
Cancer development induced by graded expression of Snail in mice
13
Cancer development induced by graded expression of Snail in mice Pedro Antonio Pe ´ rez-Mancera 1,{ , Marı ´a Pe ´ rez-Caro 1,{ , Ine ´s Gonza ´lez-Herrero 1 , Teresa Flores 2 , Alberto Orfao 3 , A. Garcia de Herreros 4 , Alfonso Gutie ´ rrez-Ada ´n 5 , Bele ´ n Pintado 5 , Ana Sagrera 1 , Manuel Sa ´nchez-Martı ´n 1,6 and Isidro Sa ´nchez-Garcı ´a 1, * 1 Laboratorio 13, Instituto de Biologı ´a Molecular y Celular del Ca ´ ncer (IBMCC), CSIC/Universidad de Salamanca, Campus Unamuno, 37007 Salamanca, Spain, 2 Servicio de Anatomı ´a Patolo ´gica and 3 Servicio de Citometrı ´a, University of Salamanca, Salamanca, Spain, 4 Unitat de Biologia Cellular i Molecular, Institut Municipal d’Investigacio Medica, Universitat Pompeu Fabra, Barcelona, Spain, 5 Area de Reproduccio ´ n Animal, Centro de Investigacio ´n y Tecnologı ´a, Ctra de la Corun ˜a km 5.9, 28040 Madrid, Spain and 6 Departamento de Medicina, Universidad de Salamanca, Salamanca, Spain Received August 18, 2005; Accepted September 29, 2005 The zinc-finger transcription factor Snail is believed to trigger epithelial–mesenchymal transitions (EMTs) during cancer progression. This idea is supported by analysis of Snail knockout mice, which uncovered crucial role of Snail in gastrulation, and of individuals with cancer, in whom Snail expression is frequently upregulated. However, these results have not shown a direct link between Snail and the pathogenesis of cancer. Here we show that mice carrying hypomorphic tetracycline-repressible Snail transgenes, that increase Snail expression to 20% above normal levels, exhibit no morphological alterations and develop both epithelial and mesenchymal tumours (leukaemias). Suppression of the Snail transgene did not rescue the malignant phenotype, indicating that alterations induced by Snail are irreversible. CombitTA- Snail murine embryonic fibroblasts show similar migratory ability to that of control mouse embryonic fibro- blasts (MEFs). However, CombitTA-Snail-MEFs induce tumour formation in nude mice. CombitTA-Snail expression results in increased radioprotection in vivo, although it does not affect p53 regulation in response to DNA damage. In concert with these results, Snail expression is repressed following DNA damage. This regulation of Snail by DNA damage is p53-independent. Our results connect DNA damage with the require- ment of a critical level of an EMT regulator and provide genetic evidence that Snail plays essential roles in cancer development in mammals and thereby influences cell fate in the genotoxic stress response. INTRODUCTION The Snail family of zinc-finger transcription factors occupies a central role for mesoderm formation in several organisms from flies to mammals (1). The first member of the Snail family, snail, was described in Drosophila melanogaster (2,3), where it was shown to be essential for the formation of mesoderm (4). The transfection of Snail in mammalian epithelial cells (5,6) and the phenotype of the Snail mutant mice, where it is essential for gastrulation (7), confirmed this function. The mouse phenotype is reminiscent of that shown by snail mutants in Drosophila (8). In vitro studies have shown that Snail attenuates the cell cycle and confers resistance to cell death induced by the withdrawal of survival factors (9) or by DNA damage (10). The resistance to cell death conferred by Snail provides a selective advantage to cells to separate from the primary site and migrate. Epithelial – mesenchymal transition (EMT) is the mech- anism by which epithelial cells can dissociate from the epi- thelium and migrate. As such, EMT is fundamental to both normal development and the progression of epithelial tumours (11). Thus, Snail expression is able to trigger EMT and is increasingly recognized as an alteration in cancer. Stable expression of Snail in prototypic epithelial cell # The Author 2005. Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected] { The authors wish it to be known that, in their opinion, the first two authors should be regarded as joint First Authors. * To whom correspondence should be addressed. Tel: þ34 923238403; Fax: þ34 923294813; Email: [email protected] Human Molecular Genetics, 2005, Vol. 14, No. 22 3449–3461 doi:10.1093/hmg/ddi373 Advance Access published on October 5, 2005 by guest on May 15, 2014 http://hmg.oxfordjournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Cancer development induced by graded expression of Snail in mice

Cancer development induced by gradedexpression of Snail in mice
Pedro Antonio Perez-Mancera1,{, Marıa Perez-Caro1,{, Ines Gonzalez-Herrero1, Teresa Flores2,
Alberto Orfao3, A. Garcia de Herreros4, Alfonso Gutierrez-Adan5, Belen Pintado5, Ana Sagrera1,
Manuel Sanchez-Martın1,6 and Isidro Sanchez-Garcıa1,*
1Laboratorio 13, Instituto de Biologıa Molecular y Celular del Cancer (IBMCC), CSIC/Universidad de Salamanca,
Campus Unamuno, 37007 Salamanca, Spain, 2Servicio de Anatomıa Patologica and 3Servicio de Citometrıa,
University of Salamanca, Salamanca, Spain, 4Unitat de Biologia Cellular i Molecular, Institut Municipal d’Investigacio
Medica, Universitat Pompeu Fabra, Barcelona, Spain, 5Area de Reproduccion Animal, Centro de Investigacion y
Tecnologıa, Ctra de la Coruna km 5.9, 28040 Madrid, Spain and 6Departamento de Medicina, Universidad de
Salamanca, Salamanca, Spain
Received August 18, 2005; Accepted September 29, 2005
The zinc-finger transcription factor Snail is believed to trigger epithelial–mesenchymal transitions (EMTs)during cancer progression. This idea is supported by analysis of Snail knockout mice, which uncoveredcrucial role of Snail in gastrulation, and of individuals with cancer, in whom Snail expression is frequentlyupregulated. However, these results have not shown a direct link between Snail and the pathogenesis ofcancer. Here we show that mice carrying hypomorphic tetracycline-repressible Snail transgenes, thatincrease Snail expression to 20% above normal levels, exhibit no morphological alterations and developboth epithelial and mesenchymal tumours (leukaemias). Suppression of the Snail transgene did notrescue the malignant phenotype, indicating that alterations induced by Snail are irreversible. CombitTA-Snail murine embryonic fibroblasts show similar migratory ability to that of control mouse embryonic fibro-blasts (MEFs). However, CombitTA-Snail-MEFs induce tumour formation in nude mice. CombitTA-Snailexpression results in increased radioprotection in vivo, although it does not affect p53 regulation in responseto DNA damage. In concert with these results, Snail expression is repressed following DNA damage. Thisregulation of Snail by DNA damage is p53-independent. Our results connect DNA damage with the require-ment of a critical level of an EMT regulator and provide genetic evidence that Snail plays essential roles incancer development in mammals and thereby influences cell fate in the genotoxic stress response.
INTRODUCTION
The Snail family of zinc-finger transcription factors occupies acentral role for mesoderm formation in several organismsfrom flies to mammals (1). The first member of the Snailfamily, snail, was described in Drosophila melanogaster(2,3), where it was shown to be essential for the formationof mesoderm (4). The transfection of Snail in mammalianepithelial cells (5,6) and the phenotype of the Snail mutantmice, where it is essential for gastrulation (7), confirmedthis function. The mouse phenotype is reminiscent of thatshown by snail mutants in Drosophila (8). In vitro studies
have shown that Snail attenuates the cell cycle and confersresistance to cell death induced by the withdrawal of survivalfactors (9) or by DNA damage (10). The resistance to celldeath conferred by Snail provides a selective advantage tocells to separate from the primary site and migrate.
Epithelial–mesenchymal transition (EMT) is the mech-anism by which epithelial cells can dissociate from the epi-thelium and migrate. As such, EMT is fundamental to bothnormal development and the progression of epithelialtumours (11). Thus, Snail expression is able to trigger EMTand is increasingly recognized as an alteration in cancer.Stable expression of Snail in prototypic epithelial cell
# The Author 2005. Published by Oxford University Press. All rights reserved.For Permissions, please email: [email protected]
{The authors wish it to be known that, in their opinion, the first two authors should be regarded as joint First Authors.
*To whom correspondence should be addressed. Tel: þ34 923238403; Fax: þ34 923294813; Email: [email protected]
Human Molecular Genetics, 2005, Vol. 14, No. 22 3449–3461doi:10.1093/hmg/ddi373Advance Access published on October 5, 2005
by guest on May 15, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

system of MDCK cells induces a complete epithelial tomesenchymal transition (5,6) and these cells overexpressingSnail exhibit tumorigenic properties when injected in nudemice (6). The involvement of Snail in tumour progression isalso supported by its expression in invasive carcinoma celllines (6), and by the expression of Snail in the invasive cellsof tumours induced in the skin of mice (6) and in biopsiesfrom patients with ductal breast carcinomas (12,13), gastriccancer (14), hepatocellular carcinomas (15) and synovial sar-comas (16). Thus, Snail expression appears to be correlatedwith invasive growth potential in human cancer.
In this study, to further investigate the function of Snailduring cancer development, we generated mice harbouring atetracycline-repressible Snail transgene. These mice did notexhibit morphological defects at birth, but did developcancers similar to those associated with SNAIL expression inhumans. These defects were not corrected by suppression ofthe Snail transgene. We found that CombitTA-Snail mouseembryonic fibroblasts (MEFs) and mice expressed Snail atlevels considerably lower than those of endogenous counter-parts. We further show here that CombitTA-Snail does not
confer a migratory advantage, although it does inducetumour formation. CombitTA-Snail expression results inincreased radioprotection in vivo. Snail expression is repressedfollowing DNA damage in a p53-independent manner. Thus, itseems likely that failure to regulate Snail expression explainswhy CombitTA-Snail mice develop cancer. These resultssuggest that tightly graded increase of Snail can induce cancer.
RESULTS
Derivation of CombitTA-Snail mice
To determine the effect of upregulation of Snail expressionin cancer development, we generated transgenic mice usingthe CombitTA system, in which the expression of Snail genecould be exogenously regulated. This system, which has thetransactivator and the tet-operator minimal promoter drivingthe expression gene unit on a single plasmid (17), ensuresthe integration of the transactivator and reporter gene unitsin equal copy numbers in a direct cis-configuration at thesame chromosomal locus and prevents genetic segregation
Figure 1. CombitTA-Snail: transgene construct, expression and effect of Snail on the survival of Ba/F3 cells deprived of growth factor. (A) Schematic represen-tation of the CombitTA-Snail vector used in this study. (B) Analysis of tetracycline-dependent Snail expression by RT–PCR in Ba/F3 cells for CombitTA-Snail(2tet, þtet in the medium). The PCR products were transferred to a nylon membrane and analysed by hybridization with a specific probe for Snail. Actin wasused to check cDNA integrity and loading. (C) Survival of Ba/F3 cells expressing Snail (Ba/F3 þ CombitTA-Snail) in the absence of IL-3. Cells growing expo-nentially in IL-3 supplemented media were adjusted to 5�105 cells/ml on day 0, and cultured after removal of IL-3. The cell number of viable cells is shown forSnail-transfected Ba/F3 cells grown in the absence of IL-3 (2tet or þtet in the medium). (D) Cell death is accompanied by nucleosome laddering after IL-3deprivation. Low-molecular-weight DNA was isolated 24 h after IL-3 deprivation from Ba/F3-CombitTA-Snail grown in the absence of IL-3 and doxycycline(2tet) (lane 1) and Ba/F3-CombitTA-Snail grown in the absence of IL-3 and with doxycycline (þtet) (lane 2). The time of treatment with doxycycline was 48 h.DNA was end-labelled, resolved by electrophoresis in a 2% agarose gel and visualized by autoradiography.
3450 Human Molecular Genetics, 2005, Vol. 14, No. 22
by guest on May 15, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

of the control elements during breeding. Insertion of themSnail gene under the control of the tetO-minimal promoteryielded the plasmid CombitTA-Snail (Fig. 1A). This was ana-lysed in a cell system, using a murine hematopoietic precursorBa/F3 cell line (18). In the absence of tetracycline, the tet-repressor protein (fused to the viral VP16 transactivatordomain) binds to an engineered tet-operator minimal promoterand activates Snail transcription (CombitTA-Snail). In thepresence of the tetracycline, binding is abolished and thepromoter silenced (Fig. 1A). CombitTA-Snail expressionwas determined in transfected Ba/F3 cells after culturing for2 days in the presence or absence of tetracycline (Fig. 1B).CombitTA-Snail was detected in Ba/F3 cells withouttetracycline but not in cells cultured with tetracycline(20 ng/ml). In vitro studies have previously shown that Snailconfers resistance to cell death induced by the withdrawalof survival factors (9). The physiological relevance of theCombitTA-Snail suppression was confirmed in vitro byassaying survival of Ba/F3 cells expressing CombitTA-Snail24 h after IL-3 withdrawal. The effects of Snail expressionon apoptosis were evaluated by analysing internucleosomalDNA cleavage leading to the formation of DNA ladders inagarose gels, which is a hallmark of apoptosis. Normally,Snail expression protects Ba/F3 cells from apoptosis followingIL-3 withdrawal (Fig. 1C and D) and the level of CombitTA-Snail expression was sufficient in Ba/F3 cells to prevent celldeath. The sensitivity to IL-3 removal was restored by theaddition of tetracycline (Fig. 1C and D). This physiologicalrelevance of the CombitTA-Snail function was further studiedby demonstrating the nuclear localization of CombitTA-Snailprotein (Fig. 2).
We generated three founder transgenic lines for CombitTA-mSnail (59A, 59B and 59C) (Fig. 3A). Only two founder lines,59A and 59B, showed germline transmission of the transgene(Table 1). In both lines, the CombitTA-Snail expression wasdetected in all tissues analysed (Fig. 3B). The CombitTA-Snail expression was the result of transactivation, as the sup-pression of expression to undetectable values was confirmedwhen mice were supplied with tetracycline in their drinkingwater (see below, Fig. 7A).
CombitTA-Snail mice show no morphologicalabnormalities
Cohorts of CombitTA-Snail mice were generated to analysethe effect of the Snail expression in vivo. A total of 63 trans-genic animals (34 mice corresponded to line 59A and 29 miceto line 59B) were analysed in detail and similar phenotypicfeatures were seen in both lines. CombitTA-Snail mice wereborn alive without overt morphological abnormalities andwere fully fertile with apparently no differences in theprogeny. Autopsy of pups, including extensive histologicalanalysis, revealed no abnormality of the kidneys, skin, liver,brain, lung or gastrointestinal tract of CombitTA-Snail mice,indicating that this level of overexpression of Snail does notperturb normal embryonic development. These results contrastwith the sustained expression of Snail in K14-Snail[HA] trans-genic animals that results in small-sized mice and in epidermalhyperproliferation and differentiation defects in mouse skin(19). A priori, these differences could be simply due to vari-ations in the Snail expression level. Although CombitTA-Snail mice show no morphological abnormalities, the
Figure 2. Subcellular localization of CombitTA-Snail. CombitTA-Snail was transiently expressed in COS7 cells. After fixation of the cells, the localization ofCombitTA-Snail protein (green) and nuclei (blue; DAPI) was examined under a fluorescent microscope. Panels before and after doxycycline treatment for Snail,nuclei alone, phase and merge images are presented (2tet or þtet in the medium). The time of treatment with doxycycline was 48 h. CombitTA-Snail wasdetected in COS7 cells without tetracycline but not in cells cultured with tetracycline (20 ng/ml).
Human Molecular Genetics, 2005, Vol. 14, No. 22 3451
by guest on May 15, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

thymus of CombitTA-Snail mice was small. However, T-cellnumbers in peripheral blood of CombitTA-Snail mice arenormal, suggesting that this overexpression of Snail doesinterfere with the growth of primitive mouse thymocytes butnot single CD4þ and CD8þ cells. Consistent with this inter-pretation, analysis of thymus composition from 4-week-oldCombitTA-Snail mice shows reduced cell production anddifferentiation towards CD4þCD8þ cells (Fig. 4). This specificT-cell differentiation block has also been observed in thethymus of Slug2/ 2 mice (20).
Cancer development in CombitTA-Snail mice
We next analysed whether the CombitTA-Snail mice developcancer. All CombitTA-Snail mice became unwell fromapproximately 5–7 months of age onward (Table 1) withclinical manifestations that included decreased physicalactivity, tachypnea, pilo-erection, shivering and sustainedweight loss, prior to sacrifice. The cancers were from bothmesenchymal and epithelial origin (Table 1). The mesenchy-mal cancers were acute leukaemias (Fig. 5A) and lymphomas
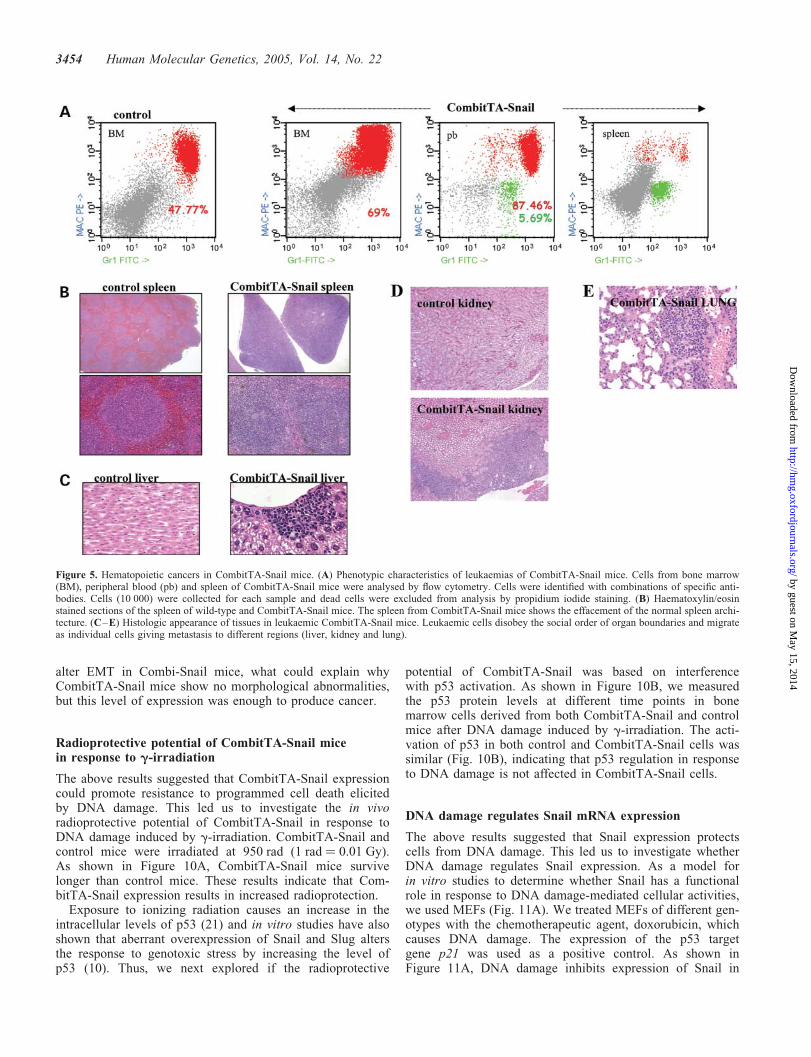
(Fig. 5B). No sarcomas were seen in any of the CombitTA-Snail mice analysed, even though with ubiquitous expres-sion of CombitTA-Snail. Detailed analysis of the epithelialtumour cells established the diagnosis as lung carcinomas(Fig. 6A), germ cell hyperplasias (Fig. 6B) and hepatocarcino-mas (Fig. 6C). We detected one type of carcinoma per animalalthough 20–25% of them also develop a hematopoieticneoplasia. The histological examination could not show dis-semination of the carcinomas. However, histological analysisrevealed marked leukaemic cell infiltration of hematopoieticand non-hematopoietic tissues. These leukaemic cells prefer-entially infiltrate kidney, liver and lung (Fig. 6C–E). Peri-pheral blood mononuclear cells from leukaemic mice wereidentified by flow cytometry using combination of specificantibodies. These studies defined the acute leukeamias asacute myeloid leukaemias (Fig. 5A).
To test the malignant potential of cells from the CombitTA-Snail mice, 1�106 peripheral blood blast cells fromCombitTA-Snail leukaemias were injected subcutaneouslyinto twelve 40-day-old nude mice. All 12 mice developed pro-gressive tumours within 4–7 weeks of transplantation. The
Figure 3. CombitTA-Snail mice: transgene expression. (A) Identification of transgenic mice by Southern analysis of tail snip DNA after EcoRI digestion.We used the cDNA for mouse Snail for detection of the transgene. (B) Expression of the transgene was demonstrated by RT–PCR. Expression ofCombitTA-Snail and endogenous Snail was analysed by RT–PCR in tissues derived of CombitTA-Snail and control mice. Actin was used to check cDNA integ-rity and loading.
Table 1. Incidence and age of tumour onset in CombitTA-Snail mice
Transgenicline
Miceautopsieca
Mice withtumour (%)b
Age in monthsat tumour onset
Tumour type (%)
IS59A 34 34 (100) 7–11 AML (40%), lymphoma (50%), lung carcinoma (12%),hepatocarcinoma (10%), germ cell hyperplasia (15%)
IS59B 29 29 (100) 5–10 AML (35%), lymphoma (40%), lung carcinoma (15%),hepatocarcinoma (15%), germ cell hyperplasia (15%)
IS59Cc 1 1 (NA) 1 Leukaemia
aNumber of mice during or after the period of cancer.bNumber of mice killed with cancer and percentage of tumour incidence.cNo lineage established
3452 Human Molecular Genetics, 2005, Vol. 14, No. 22
by guest on May 15, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

tumours in the nude mice were histologically identical to theoriginal leukaemias. Overall, these data indicate that Snail isable to induce cancer development.
In vivo suppression of Snail does not block cancerdevelopment
The above results support the view that Snail expression isenough to induce cancer development. Therefore abolitionof Snail overexpression might be expected to either halt orreduce the growth and/or spread of the Snail-expressingcells. To assess this, 40 leukaemic CombitTA-Snail micewere evaluated for disease progression by flow cytometryprior to and following administration of tetracycline (4 g/l inthe drinking water for 2 weeks, a dose sufficient tosuppress exogenous Snail expression) (Fig. 7A). None of theCombitTA-Snail mice exhibited amelioration of the leukaemicphenotype despite complete CombiTA-Snail suppression.Flow cytometry analysis identified the persistence of leukae-mic cells in the peripheral blood (Fig. 7B) with infiltrationof non-hematopoietic tissues evident on histology (Fig. 7C).Autopsy of these animals identified carcinomas (Fig. 7C).Thus, these results show that the alterations induced bySnail are irreversible.
A limited amount of Snail mRNA was expressedin CombitTA-Snail MEFs and mice
To understand the molecular basis underlying cancer develop-ment in CombitTA-Snail mice, we examined the expression oftransgene-encoded Snail in the spleen and in primary MEFsderived from CombitTA-Snail embryos, where the endogen-ous Snail is expressed (Fig. 8A). The expression level oftransgene-encoded Snail in spleen and MEFs of mice withrespect to the endogenous expression was increased to 20%
of wild-type levels (Fig. 8A). A limited amount of Snail wasexpressed in all tissues examined. In fact, the expressionof transgene-encoded Snail was present in epithelium ofCombitTA-Snail mice (Fig. 3B) and in the carcinomas appear-ing in CombitTA-Snail mice (Fig. 8B). Thus, these mice arean ideal in vivo model to study the consequences of lowlevels of Snail. In conclusion, our genetic studies point, forthe first time, to the critical role for an appropriate expres-sion level of an essential EMT regulator in cancer mousedevelopment.
CombitTA-Snail induces a tumorigenic but notmigratory phenotype in MEFs
The above results suggested that CombitTA-Snail may not bepresent at a level sufficient to alter EMT in CombitTA-Snailmice. Thus, we next study the migratory/invasive propertiesof CombitTA-Snail MEFs. The migratory properties were ana-lysed in a wound culture assay (6), where CombitTA-SnailMEFs showed a similar migratory behaviour to controlMEFS. Approximately 80% of the wound surface wascolonized by both control and CombitTA-Snail MEFs 15 hafter the wound was made (Fig. 9A). To test the tumorigenicproperties of the CombitTA-Snail MEFs, 1 � 106 control andCombitTA-Snail cells were injected subcutaneously into40-day-old nude mice. Mice injected with control MEFs didnot develop tumours (0 out of 10). However, CombitTA-Snail MEFs gave rise to tumours within 5–9 weeks oftransplantation at the injection sites (10 out of 10) that wereclassified as fibrosarcomas according to histologic appearance.These results indicate that low levels of the transcription factorSnail induce a tumorigenic but not migratory phenotypein MEFs. In fact, we have not observed metastasis inCombitTA-Snail mice with carcinomas. Thus the transgene-encoded Snail may not be present at a level sufficient to
Figure 4. Deficient T-cell development in thymus of CombitTA-Snail mice. Representative analysis of the cells present in the thymus of these mice is shown.Cells isolated from a wild-type (control) and a CombitTA-Snail mouse were stained with the monoclonal antibodies and analysed by flow cytometry. The per-centage of cells is indicated.
Human Molecular Genetics, 2005, Vol. 14, No. 22 3453
by guest on May 15, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from
alter EMT in Combi-Snail mice, what could explain whyCombitTA-Snail mice show no morphological abnormalities,but this level of expression was enough to produce cancer.
Radioprotective potential of CombitTA-Snail micein response to g-irradiation
The above results suggested that CombitTA-Snail expressioncould promote resistance to programmed cell death elicitedby DNA damage. This led us to investigate the in vivoradioprotective potential of CombitTA-Snail in response toDNA damage induced by g-irradiation. CombitTA-Snail andcontrol mice were irradiated at 950 rad (1 rad ¼ 0.01 Gy).As shown in Figure 10A, CombitTA-Snail mice survivelonger than control mice. These results indicate that Com-bitTA-Snail expression results in increased radioprotection.
Exposure to ionizing radiation causes an increase in theintracellular levels of p53 (21) and in vitro studies have alsoshown that aberrant overexpression of Snail and Slug altersthe response to genotoxic stress by increasing the level ofp53 (10). Thus, we next explored if the radioprotective
potential of CombitTA-Snail was based on interferencewith p53 activation. As shown in Figure 10B, we measuredthe p53 protein levels at different time points in bonemarrow cells derived from both CombitTA-Snail and controlmice after DNA damage induced by g-irradiation. The acti-vation of p53 in both control and CombitTA-Snail cells wassimilar (Fig. 10B), indicating that p53 regulation in responseto DNA damage is not affected in CombitTA-Snail cells.
DNA damage regulates Snail mRNA expression
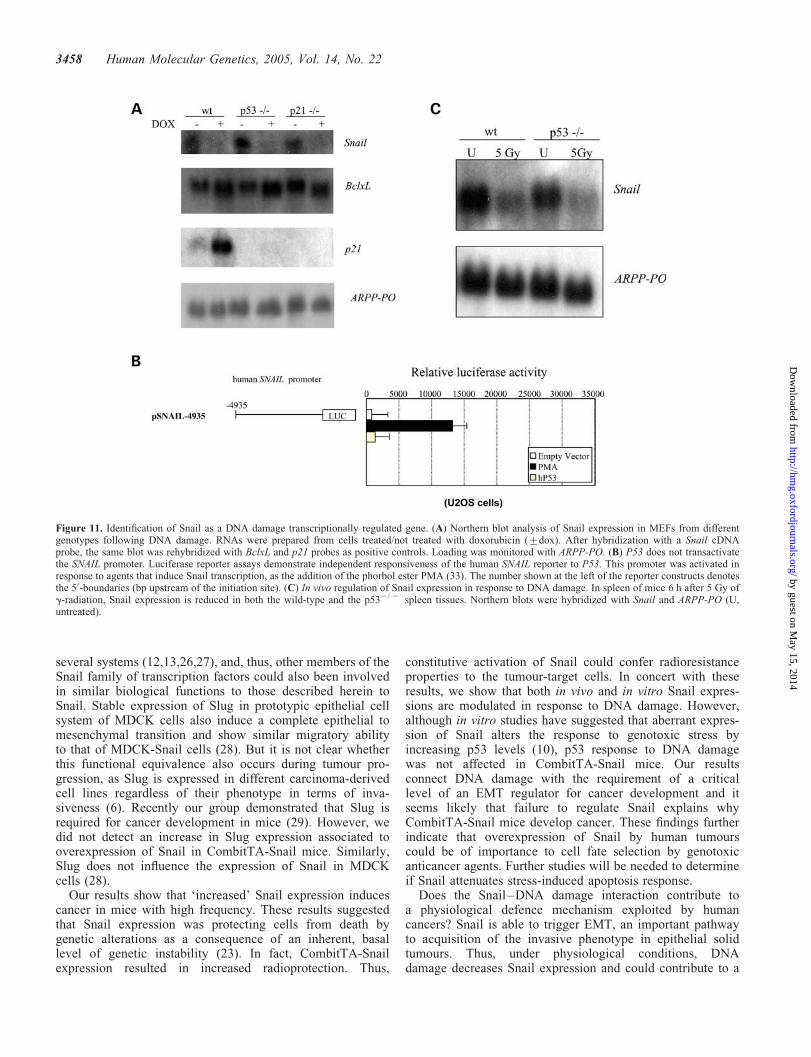
The above results suggested that Snail expression protectscells from DNA damage. This led us to investigate whetherDNA damage regulates Snail expression. As a model forin vitro studies to determine whether Snail has a functionalrole in response to DNA damage-mediated cellular activities,we used MEFs (Fig. 11A). We treated MEFs of different gen-otypes with the chemotherapeutic agent, doxorubicin, whichcauses DNA damage. The expression of the p53 targetgene p21 was used as a positive control. As shown inFigure 11A, DNA damage inhibits expression of Snail in
Figure 5. Hematopoietic cancers in CombitTA-Snail mice. (A) Phenotypic characteristics of leukaemias of CombitTA-Snail mice. Cells from bone marrow(BM), peripheral blood (pb) and spleen of CombitTA-Snail mice were analysed by flow cytometry. Cells were identified with combinations of specific anti-bodies. Cells (10 000) were collected for each sample and dead cells were excluded from analysis by propidium iodide staining. (B) Haematoxylin/eosinstained sections of the spleen of wild-type and CombitTA-Snail mice. The spleen from CombitTA-Snail mice shows the effacement of the normal spleen archi-tecture. (C–E) Histologic appearance of tissues in leukaemic CombitTA-Snail mice. Leukaemic cells disobey the social order of organ boundaries and migrateas individual cells giving metastasis to different regions (liver, kidney and lung).
3454 Human Molecular Genetics, 2005, Vol. 14, No. 22
by guest on May 15, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

MEFs in a p53-independent manner. To confirm this result,approximately 4935 bp of the promoter region of the humanSNAIL gene was cloned upstream of a luciferase reportergene (pGL3-basic). To directly assess the ability of P53 toactivate transcription from DNA sequences present in theSNAIL promoter, an expression vector containing a humanP53 cDNA (22) was co-transfected into U2OS cells alongwith the reporter vector containing the SNAIL promoter.Co-expression of P53 did not result in an increase in luciferaseactivity compared to the activity with the empty vector(Fig. 11B). These results further indicate that P53 does notregulate the SNAIL promoter.
Next, we examined the p53-independent regulation of Snailexpression following DNA damage in vivo. We treated wild-type and p532/2 mice with 5 Gy of g-radiation and analysedthe expression of Snail in spleens by northern blot (Fig. 11C).
Six hours after irradiation, the expression of Snail was down-regulated in both control and p532/2 mice. Therefore, weconclude that Snail expression is similarly modulated in vivofollowing DNA damage. Overall, our results demonstrate therequirement of a critical level of Snail for cancer developmentand it seems likely that failure to regulate Snail explains whyCombitTA-Snail mice develop cancer.
DISCUSSION
During the last years, different studies indicated the involvementof Snail in mesenchymal phenotype in cancer (5,6,11–16).Nevertheless, the molecular mechanisms by which Snailparticipates in these biological processes are not yet clear.We have utilized the single-plasmid system containing the
Figure 6. Carcinoma development in CombitTA-Snail mice. Histological analysis of lung (A), testis (B) and liver (C) of wild-type and CombitTA-Snail mice.Representative matched tissue sections from wild-type and CombitTA-Snail mice were stained with haematoxylin/eosin. The histological sections of CombitTA-Snail lung show the presence of an adenocarcinoma (A). The histological section of CombitTA-Snail testis shows the presence of a hyperplasia of germ cells (B).The histological sections of CombitTA-Snail liver show the presence of a hepatocarcinoma (C).
Human Molecular Genetics, 2005, Vol. 14, No. 22 3455
by guest on May 15, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from
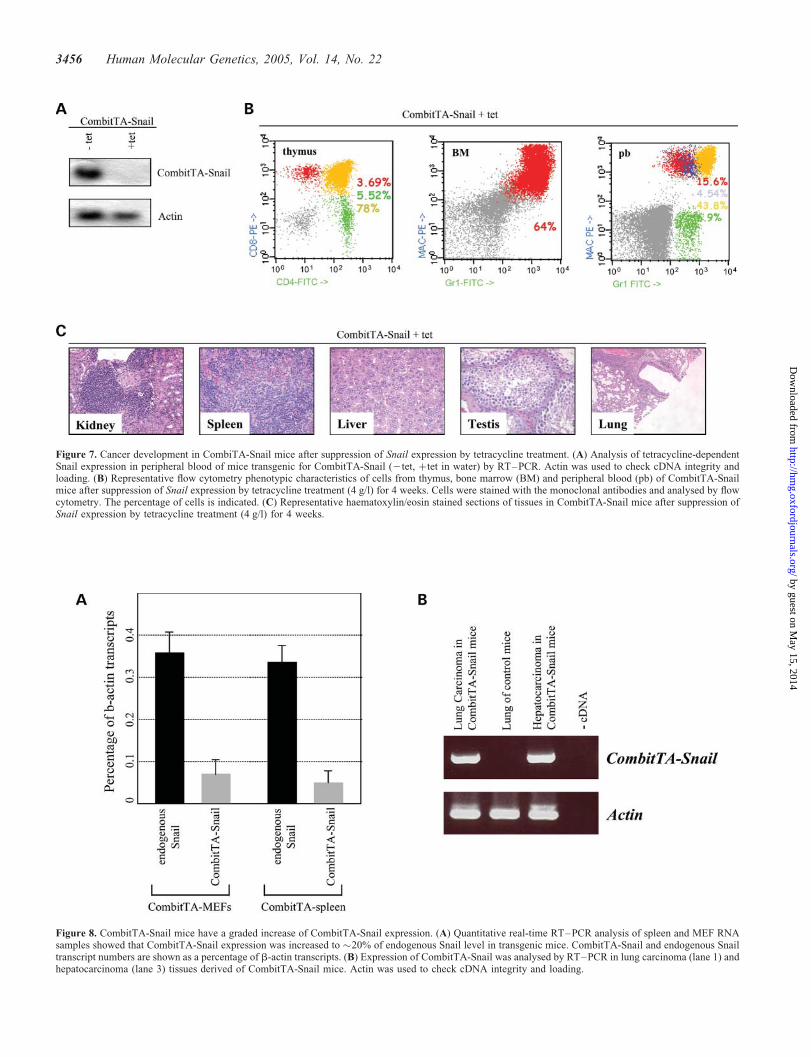
Figure 7. Cancer development in CombiTA-Snail mice after suppression of Snail expression by tetracycline treatment. (A) Analysis of tetracycline-dependentSnail expression in peripheral blood of mice transgenic for CombitTA-Snail (2tet, þtet in water) by RT–PCR. Actin was used to check cDNA integrity andloading. (B) Representative flow cytometry phenotypic characteristics of cells from thymus, bone marrow (BM) and peripheral blood (pb) of CombitTA-Snailmice after suppression of Snail expression by tetracycline treatment (4 g/l) for 4 weeks. Cells were stained with the monoclonal antibodies and analysed by flowcytometry. The percentage of cells is indicated. (C) Representative haematoxylin/eosin stained sections of tissues in CombitTA-Snail mice after suppression ofSnail expression by tetracycline treatment (4 g/l) for 4 weeks.
Figure 8. CombitTA-Snail mice have a graded increase of CombitTA-Snail expression. (A) Quantitative real-time RT–PCR analysis of spleen and MEF RNAsamples showed that CombitTA-Snail expression was increased to �20% of endogenous Snail level in transgenic mice. CombitTA-Snail and endogenous Snailtranscript numbers are shown as a percentage of b-actin transcripts. (B) Expression of CombitTA-Snail was analysed by RT–PCR in lung carcinoma (lane 1) andhepatocarcinoma (lane 3) tissues derived of CombitTA-Snail mice. Actin was used to check cDNA integrity and loading.
3456 Human Molecular Genetics, 2005, Vol. 14, No. 22
by guest on May 15, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

regulating and expression elements of the original binary tetra-cycline system to allow induction and tight control of geneexpression by tetracycline in mice (17) to try to understandthe relevance of Snail to human cancer development.In vitro studies have shown that Snail confers resistance tocell death induced by the withdrawal of survival factors (9).
The physiological relevance of the CombitTA-Snail suppres-sion was confirmed in vitro by assaying survival of Ba/F3cells expressing CombitTA-Snail after IL-3 withdrawal. Theanalysis of the Snail-expressing mice identified that thesemice develop cancer, mainly hematopoietic tumours. It isbelieved that the resistance to cell death conferred by Snailprovides a selective advantage to cell migration important tocancer development (9). Thus, the hematopoietic cancersobserved in the CombitTA-Snail mice represent an in vivodemonstration of the idea that transformation depends upongenetic changes that allow undifferentiated cells to growoutside their normal environment (23). Thus, these resultsprovide evidence that Snail expression facilitates cell migra-tion. However, the survival conferred by Snail, while reversiblein vitro (Fig. 1), can escape such control in vivo.
In the mouse, the Snail gene has been shown to triggerEMT, an important pathway to acquisition of the invasive phe-notype in epithelial solid tumours (5,6). Our data did notsupport this observation, with both no-epithelial alterationsand non-invasive carcinoma development in CombitTA-Snail mice. However, CombitTA-Snail mice expressed alimited amount of Snail, although level sufficient to promoteresistance to cell death elicited by growth factor withdrawal(Fig. 1). Thus, the transgene-encoded Snail may not bepresent at a level sufficient to alter EMT in our mice, butthis level of expression was enough to induce cancer. Itappears that Snail must be kept above a certain thresholdlevel to achieve normal development. Consistent with thisinterpretation, CombitTA-Snail induced a tumorigenic butnot migratory phenotype in MEFs. These findings indicateSnail does not require tumour formation before disseminationcan place. However, these results cannot exclude a role forSnail in carcinoma development in a context where epithelialcells show or accumulate previous tumour alterations.
Slug expression confers resistance to programmed celldeath, a function shared by Snail (9), elicited either bygrowth factor (20) or by DNA damage (10,24,25). In thissense, Slug has been shown to play similar roles to Snail in
Figure 9. CombitTA-Snail expression in MEFs does not induce a migratory phenotype. The motility/migratory behaviour of control-MEFs (a–c) and Com-bitTA-Snail-MEFs (d–f) was analysed in an in vitro wound model. Confluent cultures were gently scratched with a pipette tip to produce a wound. Photographsof the cultures were taken immediately after the incision (a, d) and after 9 h (b, e) and 15 h (c, f) in culture.
Figure 10. Effect of irradiation on survival of CombitTA-Snail mice. (A)CombitTA-Snail (30 animals) and control mice (30 animals) were irradiatedat 950 rad to determine their survival after DNA damage. The radiationdose was given as a split dose of equal intensity, 4 h apart. (B) Levels ofp53 protein in CombitTA-Snail and control BM cells after g-irradiation. p53protein was detected by western blotting. Actin was used as a loadingcontrol. The time points are in hours.
Human Molecular Genetics, 2005, Vol. 14, No. 22 3457
by guest on May 15, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from
several systems (12,13,26,27), and, thus, other members of theSnail family of transcription factors could also been involvedin similar biological functions to those described herein toSnail. Stable expression of Slug in prototypic epithelial cellsystem of MDCK cells also induce a complete epithelial tomesenchymal transition and show similar migratory abilityto that of MDCK-Snail cells (28). But it is not clear whetherthis functional equivalence also occurs during tumour pro-gression, as Slug is expressed in different carcinoma-derivedcell lines regardless of their phenotype in terms of inva-siveness (6). Recently our group demonstrated that Slug isrequired for cancer development in mice (29). However, wedid not detect an increase in Slug expression associated tooverexpression of Snail in CombitTA-Snail mice. Similarly,Slug does not influence the expression of Snail in MDCKcells (28).
Our results show that ‘increased’ Snail expression inducescancer in mice with high frequency. These results suggestedthat Snail expression was protecting cells from death bygenetic alterations as a consequence of an inherent, basallevel of genetic instability (23). In fact, CombitTA-Snailexpression resulted in increased radioprotection. Thus,
constitutive activation of Snail could confer radioresistanceproperties to the tumour-target cells. In concert with theseresults, we show that both in vivo and in vitro Snail expres-sions are modulated in response to DNA damage. However,although in vitro studies have suggested that aberrant expres-sion of Snail alters the response to genotoxic stress byincreasing p53 levels (10), p53 response to DNA damagewas not affected in CombitTA-Snail mice. Our resultsconnect DNA damage with the requirement of a criticallevel of an EMT regulator for cancer development and itseems likely that failure to regulate Snail explains whyCombitTA-Snail mice develop cancer. These findings furtherindicate that overexpression of Snail by human tumourscould be of importance to cell fate selection by genotoxicanticancer agents. Further studies will be needed to determineif Snail attenuates stress-induced apoptosis response.
Does the Snail–DNA damage interaction contribute toa physiological defence mechanism exploited by humancancers? Snail is able to trigger EMT, an important pathwayto acquisition of the invasive phenotype in epithelial solidtumours. Thus, under physiological conditions, DNAdamage decreases Snail expression and could contribute to a
Figure 11. Identification of Snail as a DNA damage transcriptionally regulated gene. (A) Northern blot analysis of Snail expression in MEFs from differentgenotypes following DNA damage. RNAs were prepared from cells treated/not treated with doxorubicin (+dox). After hybridization with a Snail cDNAprobe, the same blot was rehybridized with BclxL and p21 probes as positive controls. Loading was monitored with ARPP-PO. (B) P53 does not transactivatethe SNAIL promoter. Luciferase reporter assays demonstrate independent responsiveness of the human SNAIL reporter to P53. This promoter was activated inresponse to agents that induce Snail transcription, as the addition of the phorbol ester PMA (33). The number shown at the left of the reporter constructs denotesthe 50-boundaries (bp upstream of the initiation site). (C) In vivo regulation of Snail expression in response to DNA damage. In spleen of mice 6 h after 5 Gy ofg-radiation, Snail expression is reduced in both the wild-type and the p532/ 2 spleen tissues. Northern blots were hybridized with Snail and ARPP-PO (U,untreated).
3458 Human Molecular Genetics, 2005, Vol. 14, No. 22
by guest on May 15, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

transient inhibition of migratory capacity of tumour-targetcell. With constitutive expression of Snail during transform-ation, this control is lost. Thus human cancers that overexpressSnail may have a survival advantage to genotoxic andpotentially other forms of stress by exploiting physiologicalmechanisms that evolved for the EMT, raising the possibilityof strategies based on Snail for the treatment of human cancer.
MATERIALS AND METHODS
Generation of transgenic mice and treatments
The cDNA for mouse Snail was cloned into the Combi-tTAvector (17). Linear DNA fragments for microinjection wereobtained by NotI digestion and injected into CBAxC57BL/6J-fertilized eggs. We identified transgenic mice by Southernanalysis of tail snip DNA after EcoRI digestion as described(30). We used the cDNA for mouse Snail for detection ofthe transgene. Founder mice were crossed to the C57BL6mice for five generations to establish co-isogenic transgenicmice. Similar phenotypic features were seen in all assays forboth the CombitTA-Snail transgenic lines generated. Miceaged 5–6 weeks were irradiated using a caesium source andmaintained in microisolator cages on sterilized food andacidified sterile water.
Histological analysis
Mice included in this study were subjected to standardnecropsy. All major organs were closely examined under thedissecting microscope, and samples of each organ were pro-cessed into paraffin, sectioned and examined histologically.All tissue samples were taken from homogenous and viableportions of the resected sample by the pathologist andfixed within 2–5 min of excision. Haematoxylin- and eosin-stained sections of each tissue were reviewed by a singlepathologist (T.F.). For comparative studies, age-matched micewere used (wild-type or Combi-Slug mice in the continuouspresence of tetracycline).
Cell culture
Cell lines used include Ba/F3 cells (18) and COS7 cells. Cellswere maintained in Dulbecco’s modified Eagle’s medium(DMEM) supplemented with 10% fetal calf serum (FCS).When required, 10% WEHI-3B-conditioned medium wasadded as a source of IL-3.
Subcellular localization of CombitTA-Snail
CombitTA-Snail was transiently expressed in COS7 cells.After fixation of the cells, immunostaining was carried outby using anti-Snail antibody (173EC3F9) (Franci et al. inpreparation)—as described (31). The localization of theCombiTA-Snail protein was examined under a fluorescentmicroscope.
Cell transfection and cell survival assay
Ba/F3 cells were transfected by electroporation (960 mF,220 V) with 20 mg of either CombitTA-Snail. The neomy-cin-resistant pool of cells (Ba/F3 þ CombitTA-Snail) wereanalysed by RT–PCR for CombitTA-Snail expression inthe presence and in the absence of tetracycline (20 ng/ml).These cells were resistant to IL-3 withdrawal when grow inthe absence of tetracycline. Cells were screened for resistanceto IL-3 withdrawal and cell viability was determined byTrypan blue exclusion.
Culture of MEFs
Heterozygous p53þ/2 (Jackson Laboratories) and p21þ/2 (agift of M. Serrano) mice were crossed to obtain wild-typeand null p532/2 and p212/2 embryos, respectively. Primaryembryonic fibroblasts were harvested from 13.5 d.p.c.embryos. Head and organs of day 13.5 embryos were dis-sected; fetal tissue was rinsed in PBS, minced and rinsedtwice in PBS. Fetal tissue was treated with trypsin/EDTAand incubated for 30 min at 378C and subsequently dissociatedin medium. After removal of large tissue clamps, the remain-ing cells were plated out in a 175-cm2 flask. After 48 h,confluent cultures were frozen down. These cells were con-sidered as being passage one MEFs. For continuous culturing,MEF cultures were split into 1:3. MEFs were grown at 378C inDMEM (Boehringer Ingelheim) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Boehringer Ingelheim).All the cells were negative for mycoplasma (MycoAlertTM
Mycoplasma Detection Kit, Cambrex).
DNA damage experiments
Cells were plated at 106 cells per 10 cm dish, and the day after,they were treated with 0.2 mg/ml of doxorubicin (Sigma) (32).After 12 h, cells were collected for RNA preparation.
Low-molecular-weight DNA analysis
Low-molecular-weight DNA was isolated as follows. Cellswere collected into 1.5 ml of culture medium and micro-centrifuged for 1 min at 1500 rpm (400g), and the pellet wassuspended in 300 ml of proteinase K buffer. After overnightincubation at 558C, DNA was ethanol-precipitated, suspendedin 200 ml of TE buffer containing 50 mg/ml of RNase A, andincubated at 378C for 2 h. DNA was extracted with phenol andchloroform, and precipitated with ethanol. Aliquots of DNA(2 mg) were end-labelled with a32-dCTP and electrophoresedon 2% agarose gels. After electrophoresis, the gel was blottedonto Hybond-N (Amersham) and autoradiographed for 2 hat 2708C.
Reverse transcription–PCR (RT–PCR) and real-timePCR quantification
To analyse expression of CombitTA-Snail and endogenousSnail in mouse cell lines and mice, RT was performed accord-ing to the manufacturer’s protocol in a 20-ml reaction contain-ing 50 ng of random hexamers, 3 mg of total RNA and 200 U
Human Molecular Genetics, 2005, Vol. 14, No. 22 3459
by guest on May 15, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

of Superscript II RNase H2 reverse transcriptase (GIBCO/BRL). The sequences of the specific primers were asfollows: Combi-polyA-B1: 50-TTGAGTGCATTCTAGTTGTG-30; mSnailF: 50-CAGCTGGCCAGGCTCTCGGT-30;mSnailB: 50-GCGAGGGCCTCCGGAGCA-30. EndogenousSnail expression was analysed with mSnailF and mSnailBprimers and exogenous Snail expression with mSnailF andCombi-polyA-B1 primers. Amplification of b-actin RNAserved as a control to assess the quality of each RNAsample. The PCR conditions used to amplify CombitTA-Snail and endogenous Snail were as follows: 948C for1 min, 568C for 1 min and 728C for 2 min for 40 cycles forCombitTA-Snail and 30 cycles for endogenous Snail, respect-ively. The PCR products were confirmed by hybridizationwith specific internal probes. Real-time quantitative PCRwas carried out for the quantitation of both CombitTA-Snailand endogenous Snail. Fluorogenic PCRs were set up in areaction volume of 50 ml using the TaqMan PCR CoreReagent kit (PE Biosystems). cDNA amplifications werecarried out using the same primers in a 96-well reactionplate format in a PE Applied Biosystems 5700 SequenceDetector. Thermal cycling was initiated with a first denatura-tion step of 10 min at 958C. The subsequent thermal profilewas 40 cycles of 958C for 15 s, 568C for 30 s and 728C for1 min. Multiple negative water blanks were tested and a cali-bration curve was determined in parallel with each analysis.The b-actin endogenous control (PE Biosystems) wasincluded to relate both CombitTA-Snail and endogenousSnail to total cDNA in each sample.
Phenotype analysis
The following anti-mouse monoclonal antibodies fromPharmingen were used for cytometry staining: CD45R/B220(B-cell-specific), IgM (B-cell-specific), Mac1 (myeloid-cell-specific), Gr-1 (myeloid-cell-specific), CD4 (T-cell-specific)and CD8 (T-cell-specific). Single cell suspensions fromdifferent tissue samples obtained by routine techniques wereincubated with purified anti-mouse CD32/CD16 (Pharmingen)to block binding via Fc receptors and with an appropriatedilution of the different antibodies at room temperature or48C. The samples were washed twice with PBS and resus-pended in PBS. Dead cells in samples were excluded bypropidium iodide staining. The samples and the data wereanalysed in a FACScan using CellQuest software (BectonDickinson).
Tumorigenicity assay
To test the tumorigenicity of the various CombitTA-Snailcancers and MEFs, 4–6-week-old athymic (nude) male micewere injected subcutaneously on both flanks with 106 cellsresuspended in 200 ml of phosphate-buffered saline (PBS).The animals were examined for tumour formation every week.
Luciferase assays
Approximately 4935-bp upstream promoter sequence ofSNAIL was isolated from a P1 clone containing the SNAILgene (Genome Systems) and cloned into the luciferase reporter
plasmid pGL3-basic (Promega) and termed PSNAIL-4935.For reporter assays, U2OS cells were transfected usingDual-Luciferase (Promega) with normalization to Renillaluciferase, and mean + standard error was determined fromat least three data points. U2OS cells were maintained inDMEM supplemented with 10% FBS. When indicated, cellswere supplemented with phorbol 12-myristate 13-acetate(PMA) (from Sigma).
Northern blot analysis
Total cytoplasmic RNA of different MEFs and spleen tissuesfrom both untreated and 5 Gy-irradiated wild-type and thep532/2 mice was glyoxylated and fractionated in 1.4%agarose gels in 10 mM Na2HPO4 buffer (pH 7.0). After elec-trophoresis, the gel was blotted onto Hybond-N (Amersham),UV-cross-linked and hybridized to 32P-labelled mouse SlugcDNA probe. Loading was monitored by reprobing the filterwith a ARPP-P0 probe.
Western blot analysis
Bone marrow cells were collected by flushing the marrowcavity of femurs. Western blot assays were done using extractsfrom 1�107 BM cells per lane. Extracts were normalized forprotein content by Bradford analysis (Bio-Rad Laboratories,Inc., Melville, NY, USA) and Coommasie blue gel staining.Lysates were run on a 10% SDS–PAGE gel and transferredto a PVDF membrane. After blocking, the membrane wasprobed with the following primary antibodies: mouse p53was detected using the antibody FL-393 (Santa Cruz) andthe polyclonal antibody C-11 (Santa Cruz) was used todetect actin. Reactive bands were detected with an ECLsystem (Amersham).
Migration assays
The migratory/motility behaviour of transfectant cells wasanalysed by the wound assay. Monolayers of confluent cul-tures were lightly scratched with a Gilson pipette tip and,after washing to remove detached cells, the cultures wereobserved at timely intervals as previously described (6).
ACKNOWLEDGEMENTS
We thank all members of lab 13 for their helpful commentsand constructive discussions on the manuscript. We are grate-ful to Dr Pedro Soria for continuous and generous help withthe mice irradiation, to Dr Tyler Jacks for various usefulreagents, to Dr Manuel Serrano for the p21 mice, toDr Martin Haas for the human p53 expression vector and toDr H. Bluethmann for the Combi-tTA vector. This work wassupported by MEyC (BIO2000-0453-P4-02, SAF2003-01103, FIT-010000-2004-157 and PETRI no. 95-0913.OP),Junta de Castilla y Leon (CSI03A05), ADE de Castillay Leon (04/04/SA/0001), FIS (PI020138, G03/179 and G03/136), Fundacion de Investigacion MMA and USAL-CIBASA project. M.S.M. was supported by FIS grant no.PI041271 and M.P.C. is a recipient of a MCyT fellowship.
3460 Human Molecular Genetics, 2005, Vol. 14, No. 22
by guest on May 15, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

Conflict of Interest statement. None declared.
REFERENCES
1. Nieto, M.A. (2002) The Snail superfamily of zinc finger transcriptionfactors. Nat. Rev. Mol. Cell Biol., 3, 155–166.
2. Grau, Y., Carteret, C. and Simpson, P. (1984) Mutations andchromosomal rearrangements affecting the expression of snail, a geneinvolved in embryonic patterning in Drosophila melanogaster. Genetics.,108, 347–360.
3. Nusslein-Volhard, C., Weischaus, E. and Kluding, H. (1984) Mutationsaffecting the pattern of the larval cuticle in Drosophila malanogaster.I. Zygotic loci on the second chromosome. Wilheim Roux’s Arch. Dev.Biol., 193, 267–282.
4. Alberga, A., Boulay, J.L., Kempe, E., Dennefeld, C. and Haenlin,M. (1991) The snail gene required for mesoderm formation in Drosophilais expressed dynamically in derivatives of all three germ layers.Development., 111, 983–992.
5. Battle, E., Sancho, E., Franci, C., Dominguez, D., Monfar, M., Baulida,J. and de Herreros, A.G. (2000) The transcription factor Snail is arepressor of E-cadherin gene expression in epithelial tumour cells. Nat.Cell Biol., 2, 84–89.
6. Cano, A., Perez-Moreno, M.A., Rodrigo, I., Locascio, A., Blanco, M.J.,del Barrio, M.G., Portillo, F. and Nieto, M.A. (2000) The transcriptionfactor Snail controls epithelial–mesenchymal transitions by repressingE-cadherin expression. Nat. Cell Biol., 2, 76–83.
7. Carver, E.A., Jiang, R., Lan, Y., Oram, K.F. and Gridley, T. (2001) Themouse snail gene encodes a key regulator of the epithelial–mesenchymaltransition. Mol. Cell Biol., 21, 8184–8188.
8. Oda, H., Tsukita, S. and Takeichi, M. (1998) Dynamic behavior of thecadherin-based cell–cell adhesion system during Drosophila gastrulation.Dev. Biol., 203, 435–450.
9. Vega, S., Morales, A.V., Ocana, O.H., Valdes, F., Fabregat, I. and Nieto,M.A. (2004) Snail blocks the cell cycle and confers resistance to celldeath. Genes Dev., 18, 1131–1143.
10. Kajita, M., McClinic, K.N. and Wade, P.A. (2004) Aberrant expression ofthe transcription factors snail and slug alters the response to genotoxicstress. Mol. Cell Biol., 24, 7559–7566.
11. Hay, E.D. (1995) An overview of epithelio-mesenchymal transformation.Acta Anat. (Basel), 154, 8–20.
12. Cheng, C.W., Wu, P.E., Yu, J.C., Huang, C.S., Yue, C.T., Wu, C.W. andShen, C.Y. (2001) Mechanisms of inactivation of E-cadherin in breastcarcinoma: modification of the two-hit hypothesis of tumor suppressorgene. Oncogene, 20, 3814–3823.
13. Blanco, M.J., Moreno-Bueno, G., Sarrio, D., Locascio, A., Cano, A.,Palacios, J. and Nieto, M.A. (2002) Correlation of Snail expression withhistological grade and lymph node status in breast carcinomas. Oncogene,21, 3241–3246.
14. Rosivatz, E., Becker, I., Specht, K., Fricke, E., Luber, B., Busch, R.,Hofler, H. and Becker, K.F. (2002) Differential expression of theepithelial–mesenchymal transition regulators snail, SIP1, and twist ingastric cancer. Am. J. Pathol., 161, 1881–1891.
15. Sugimachi, K., Tanaka, S., Kameyama, T., Taguchi, K., Aishima, S.,Shimada, M., Sugimachi, K. and Tsuneyoshi, M. (2003) Transcriptionalrepressor snail and progression of human hepatocellular carcinoma. Clin.Cancer Res., 9, 2657–2664.
16. Saito, T., Oda, Y., Kawaguchi, K., Sugimachi, K., Yamamoto, H.,Tateishi, N., Tanaka, K., Matsuda, S., Iwamoto, Y., Ladanyi M. et al.(2004) E-cadherin mutation and Snail overexpression as alternativemechanisms of E-cadherin inactivation in synovial sarcoma. Oncogene,23, 8629–8638.
17. Schultze, N., Burki, Y., Lang, Y., Certa, U. and Bluethmann, H. (1996)
Efficient control of gene expression by single step integration of the
tetracycline system in transgenic mice. Nat. Biotechnol., 14, 499–503.
18. Palacios, R. and Steinmetz, M. (1985) IL-3 dependent mouse clones that
express B-220 surface antigen, contain Ig genes in germ-line
configuration, and generate B-lymphocytes in vivo. Cell, 41, 727–734.
19. Jamora, C., Lee, P., Kocieniewski, P., Azhar, M., Hosokawa, R., Chai,
Y. and Fuchs E. (2005) A signaling pathway involving TGF-beta2 and
snail in hair follicle morphogenesis. PLoS Biol., 3, e11.
20. Perez-Losada, J., Sanchez-Martin, M., Rodriguez-Garcia, A., Sanchez,M.L., Orfao, A., Flores, T. and Sanchez-Garcıa, I. (2002) Zinc-finger
transcription factor Slug contributes to the function of the stem cell factorc-kit signaling pathway. Blood, 100, 1274–1286.
21. Kastan, M.B., Onyekwere, O., Sidransky, D., Vogelstein, B. and Craig,
R.W. (1991) Participation of p53 protein in the cellular response to DNAdamage. Cancer Res., 51, 6304–6311.
22. Norris, P.S. and Haas, M. (1997) A fluorescent p53GFP fusion protein
facilitates its detection in mammalian cells while retaining the propertiesof wild-type p53. Oncogene, 15, 2241–2247.
23. Sanchez-Garcıa, I. (1997) Consequences of chromosomal abnormalities in
tumour development. Annu. Rev. Genet., 31, 429–453.
24. Inoue, A., Seidel, M.G., Wu, W., Kamizono, S., Ferrando, A.A., Bronson,R.T., Iwasaki, H., Akashi, K., Morimoto, A., Hitzler, J.K. et al. (2002)
Slug, a highly conserved zinc finger transcriptional repressor, protectshematopoietic progenitor cells from radiation-induced apoptosis in vivo.
Cancer Cell, 2, 279–288.
25. Perez-Losada, J., Sanchez-Martin, M., Perez-Caro, M., Perez-Mancera,P.A. and Sanchez-Garcıa, I. (2003) The radioresistance biological
function of the SCF/kit signaling pathway is mediated by the zinc-fingertranscription factor Slug. Oncogene, 22, 4205–4211.
26. del Barrio, M.G. and Nieto, M.A. (2002) Overexpression of Snail family
members highlights their ability to promote chick neural crest formation.
Development, 129, 1583–1593.
27. Hajra, K.M., Chen, D.Y. and Fearon, E.R. (2002) The SLUG zinc-finger
protein represses E-cadherin in breast cancer. Cancer Res., 62,
1613–1618.
28. Bolos, V., Peinado, H., Perez-Moreno, M.A., Fraga, M.F., Esteller, M. and
Cano, A. (2003) The transcription factor Slug represses E-cadherin
expression and induces epithelial to mesenchymal transitions: acomparison with Snail and E47 repressors. J. Cell Sci., 116, 499–511.
29. Perez-Mancera, P.A., Gonzalez-Herrero, I., Perez-Caro, M., Gutierrez-
Cianca, N., Flores, T., Gutierrez-Adan, A., Pintado, B., Sanchez-Martın,M. and Sanchez-Garcıa, I. (2005) SLUG (SNAI2) in cancer development.
Oncogene, 24, 3073–3082.
30. Garcia-Hernandez, B., Castellanos, A., Lopez, A., Orfao, A. and Sanchez-Garcıa, I. (1997) Murine hematopoietic reconstitution after tagging and
selection of retrovirally transduced bone marrow cells. Proc. Natl Acad.Sci. USA, 94, 13239–13244.
31. Garcia-Hernandez, B. and Sanchez-Garcıa, I. (1996) Retroviral vector
design for gene therapy of cancer: specific inhibition and tagging of BCR-ABLp190 cells. Mol. Med., 2, 124–133.
32. Attardi, L.D., Reczek, E.E., Cosmas, C., Demicco, E.G., McCurrach,
M.E., Lowe, S.W. and Jacks, T. (2000) PERP, an apoptosis-associatedtarget of p53, is a novel member of the PMP-22/gas3 family. Genes Dev.,
14, 704–718.
33. Barbera, M.J., Puig, I., Dominguez, D., Julien-Grille, S., Guaita-Esteruelas, S., Peiro, S., Baulida, J., Franci, C., Dedhar, S., Larue, L. and
Garcia de Herreros, A. (2004) Regulation of Snail transcription during
epithelial to mesenchymal transition of tumor cells. Oncogene, 23,7345–7354.
Human Molecular Genetics, 2005, Vol. 14, No. 22 3461
by guest on May 15, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from




















