
Wheat on permanent beds in Punjab, India- CSIRO Technical Report
Upload
khangminh22Category
view
0download
0
OFFICIAL JOURNAL OF THE AUSTRALIAN SOCIETY FOR MICROBIOLOGY INC.OOOOOFFFFFIICCIIALLL JJJJJOURNALLL OOOOFFFF TTTTTHHHEEE AAAAUSSTTTRRRALLIIAAN SSSSOCCIIEEETTTYY FFOOORR MMMMICROBIIOLLOOGYYYY IINNNCC..OOOOOFFFFIICIIALLLL JJJJJJOOURRRRNNAALLLL OOOOFFF TTTTTTHHHEEEEE AAAAAUSTTTRRALLLIIIIAAANN SSSSOCIEETTTTYYYY FFFFOOORRR MMMMMMMMIICRROOBBBIOLLLLOOOGYYYY IINNNNNCCCCC..OFFICIAL JOURNAL OF THE AUSTRALIAN SOCIETY FOR MICROBIOLOGY INC.OFFICIAL JOURNAL OF THE AUSTRALIAN SOCIETY FOR MICROBIOLOGY INCOOOOOOFFFFFFFFFFFIIIIIICCCCCCIIIIIIAAAAAALLLLLL JJJJJOOOOOOUUUUUURRRRRRNNNNNNAAAAAALLLLLL OOOOOOFFFFFF TTTTTTHHHHHHEEEEEE AAAAAAUUUUUUSSSSSSTTTTTTRRRRRRAAAAAALLLLLLIIIIAAAAAANNNNNN SSSSSSOOOOOOCCCCCCIIIIIIEEEEEETTTTTTYYYYYY FFFFFFOOOOOORRRRRR MMMMMMMIIIIIICCCCCCRRRRRROOOOOOBBBBBBIIIIIIOOOOOOLLLLLLOOOOOOGGGGGGYYYYYY IIIINNNNNNCCCCCCOFFICIAL JOURNAL OF THE AUSTRALIAN SOCIETY FOR MICROBIOLOGY INC
Volume 36 Number 1 March 2015Volume 36 Number 1 March 2015
Mammalian microbiomes

At Charles River, we know that the safety of your manufacturing environment depends on a clear picture of your microbial risk. Our solution? Accugenix®. Unmatched for its relevance and coverage, our microbial database paired with sequencing and MALDI-TOF provide an unparalleled 98% accuracy rate. Together with our tracking and trending tools, we deliver the precision and data yo need to monitor and report your state of control with confidence.
Learn more about our Accugenix® solutions at www.criver.com/accugenix.
C O N F I D E N C E
Building C, Suite G04, Rodborough RoadFrenchs Forest, NSW, 2086 (02)94547701

OFFICIAL JOURNAL OF THE AUSTRALIAN SOCIETY FOR MICROBIOLOGY INC.
Volume 36 Number 1 March 2015
ContentsVertical Transmission 2
Jon Iredell
Guest Editorial 3Mammalian microbiomes 3
Linda L Blackall
In Focus 4Methane matters in animals and man: from beginning to end 4
Emily Hoedt, Paul Evans, Stuart Denman, Chris McSweeney, Paraic ÓCuív and Mark Morrison
The marine mammal microbiome: current knowledge
and future directions 8
Tiffanie M Nelson, Amy Apprill, Janet Mann, Tracey L Rogers and Mark V Brown
The role of the gut microbiome in host systems 14
Clarissa Febinia, Connie Ha, Chau Le and Andrew Holmes
Under the Microscope 18Modulation of the rumen microbiome 18
Rosalind Gilbert, Diane Ouwerkerk and Athol Klieve
Polymicrobial nature of chronic oral disease 22
Stuart Dashper, Helen Mitchell, Geoff Adams and Eric Reynolds
Gastrointestinal microbiota, diet and brain functioning 25
Shakuntla Gondalia and Andrew Scholey
Marsupial oral cavity microbiome 29
Philip S Bird, Wayne SJ Boardman, Darren J Trott and Linda L Blackall
Lab Report 32Relative abundance of Mycobacterium in ovine Johne’s disease 32
Andy O Leu, Paul Pavli, David M Gordon, Jeff Cave, Jacek M Gowzdz, Nick Linden, Grant Rawlin, Gwen E Allison and Claire L O’Brien
ASM Affairs 37Interactions with other microbiology societies
through Microbiology Australia 37
ASM History SIG: Microbiology Australia 37
Recent developments in virology by Australian researchers 38
Clinical Serology and Molecular SIG 39
Report from the ASM Antimicrobial Special Interest Group (ASIG) 40
MICROBIOLOGY AUSTRALIA • MARCH 2015 1
Dr Gary Lum Dr John MerlinoProf. Wieland MeyerProf. William RawlinsonDr Paul SelleckDr David SmithMs Helen SmithDr Jack WangDr Paul Young
The Australian Societyfor Microbiology Inc.9/397 Smith StreetFitzroy, Vic. 3065Tel: 1300 656 423Fax: 03 9329 1777Email: [email protected] 24 065 463 274
For Microbiology Australiacorrespondence, see address below.
Editorial teamProf. Ian Macreadie, Mrs Jo Macreadieand Mrs Hayley Macreadie
Editorial BoardDr Chris Burke (Chair)Prof. Mary BartonProf. Linda BlackallProf. Sharon ChenProf. Peter ColoeDr Narelle FeganDr Geoff HoggProf. Jonathan IredellDr I
.pek Kurtböke
Subscription ratesCurrent subscription rates are availablefrom the ASM Melbourne offi ce.
Editorial correspondenceProf. Ian Macreadie/Mrs Jo MacreadieTel: 0402 564 308 (Ian)Email: [email protected]
Published four times a year in print and open access online by
Unipark, Building 1, Level 1 195 Wellington Road, Clayton, Vic. 3168http://microbiology.publish.csiro.au
Publishing enquiriesJenny BennettEmail: [email protected]
Production enquiriesHelen PavlatosEmail: [email protected]
Advertising enquiriesDoug WaltersTel: 03 9545 8505Mobile: 0419 357 779Email: [email protected]© 2015 The Australian Society for Microbiology Inc. The ASM, through CSIRO Publishing, reserve all rights to the content, artwork and photographs in Microbiology Australia. Permission to reproduce text, photos and artwork must be sought from CSIRO Publishing.
The Australian Copyright Act 1968 and subsequent amendments permit downloading and use of an article by an individual or educational institution for non-commercial personal use or study. Multiple reproduction of any Microbiology Australia article in a study block is governed by rights agreement managed by Copyright Agency Limited and fees may apply.
Authors published in Microbiology Australia have the moral right under Australian law to be acknowledged as the creator.
ISSN 1324-4272eISSN 2201-9189
While reasonable effort has been made to ensure the accuracy of the content, the Australian Society for Microbiology, CSIRO, and CSIRO Publishing accept no responsibility for any loss or damage from the direct or indirect use of or reliance on the content. The opinions expressed in articles, letters, and advertisements in Microbiology Australia are not necessarily those of the Australian Society for Microbiology, the Editorial Board, CSIRO, and CSIRO Publishing. Cover image: Background is faecal homogenate from mouse that has been Gram stained.
Photo credit from Yi Vee Chew and Andy Holmes (University of Sydney).

Jon Iredell
President of ASM
Along with all the sciences, our discipline is evolving quickly in all
areas from pure research to applied and professional. The ASM was
formed more than half a century ago to promote the discipline of
microbiology and its role is now more important than ever. The
national leadership is conscious of the need to adapt and change
and has been moving in the past few years to do so, with gathering
momentum. One of our most important platforms is our national
meeting, developed to promote the exchange of ideas. The mem-
bership of the society is increasingly drawn to other conferences to
meet their needs and this must be recognised and accommodated.
Our approach tomeetings including our national scientificmeeting
must evolve with it, as in comparable societies.
We now see increased centralisation and automation and much
greater incorporation of molecular diagnostics and other aspects of
biotechnology in industry, environmental and diagnostic microbi-
ology. Accordingly, a national meeting under the auspices of the
ASM togive aplatform todiscuss research anddevelopment in these
fast-moving areas and to provide workshops for skill development
is being explored, initially focusing on clinical diagnostic microbi-
ology. This follows the development of an additional ASM Travel
award for clinical microbiologists (http://www.theasm.org.au/
awards/asm-clinical-microbiology-travel-award/) and theLynGilbert
Award (http://www.theasm.org.au/awards/asm-lyn-gilbert-award/)
for contributions in clinical microbiology, awarded for the first time
in 2014.
Highly specialised meetings are important and essential for career
development and networking and for exchange of the latest infor-
mation between experts in fast-moving and competitive areas of
endeavour. Deep and narrow in scope by definition, attendance at
these to the exclusion of broader conversations may not meet the
complete needs of early career microbiologists. We must therefore
not only embrace and nurture new directions in microbiology but
be open to ideas coming from outside that which we have long
regarded as conventional or traditional microbiology. We can look
to the annual scientific meeting as a venue for transdisciplinary
microbiology that is difficult to manage in highly specialised meet-
ings. This shift will be seen at the integrated symposia in Canberra
(ASM 2015), in the themed meeting in Perth (ASM 2016) and in the
planning of ASM 2017 in Hobart.
A link to state branches and the Visiting Speaker program and other
well-established infrastructure is another easy route that ASM pro-
vides for members to work through new proposals, and give early
career researchers and professional microbiologists a taste of con-
ference organising and a chance to test ideas. This ismanaged easily
through state branches and can be easily progressed to a national
meeting if the idea demands it. The relationships between states
and the utility of the Visiting Speaker Program has been enhanced
by a clearer pathway for VSP engagement, available on the website
(http://www.theasm.org.au/events/visiting-speakers-program/) and
by more formal networking between the state branches, beginning
in 2015.
A key part of our renaissance is a review of governance and
organisational structure, which also begins in 2015, and the realign-
ment of Divisional Chair responsibilities. Nominations for new
Chairs for 2017 are actively sought and will be a key part of the
repositioning of ASM. The revisedConstitution is due to be released
to all members shortly and to be voted on in the July 2015 AGM at
the Canberra meeting.
How can we participate in this rebirth and strengthenmicrobiology
as a discipline in a competitive environment? Readwidely, talk freely
with your colleagues inside and outsidemicrobiology, and promote
the community by supporting ASM: join state branches and national
council, apply formembership, including professionalmembership
andFellowship, developnew ideas formeetingswith your branchor
national office, and honour your colleagues by commending them
for awards and honorary memberships.
The discipline of microbiology is somewhat different to what it was
when theSociety began in 1959 and theSocietymust keeppacewith
it. ASM must better support progressive specialisation of the entire
membership by supporting specialised meetings, some now long-
established and completely autonomous and others that are yet be
conceived. We must at the same time enrich this with wider
engagement and bring into our community those who do not see
themselves as Microbiologists and yet work with us. The role of
the ASM is not to push back against this natural evolution but to
foster it.
Vertical Transmission
2 10.1071/MA15001 MICROBIOLOGY AUSTRALIA * MARCH 2015

Mammalian microbiomes
Linda L Blackall
Email: [email protected]
Endothermic (an organism that maintains its body at a metabolicallyfavourable temperature) amniotes (who lay their eggs on land or retainthe fertilised egg within the mother), also known asmammals, countamong their cohort the largest (whales) and the most intelligent (someprimates, cetaceans and elephants) animals on Earth. However, none ofthe 5488mammalian species live alone since they all support a complexmenagerie of microbes including prokaryotes (Bacteria and Archaea),microbial eukaryotes, and viruses. That so-called ‘microbiome’ playsmyriad roles ranging from the very well known and well studied(disease) through to provocative involvements (mood alteration andbrain activity). Indeed even the microbiome has been subdivided bysome into the bacteriome, the mycobiome and the virome. It is verytimely that this issue be devoted to mammalian microbiomes since thestudy of microbiomes is going through an unprecedented revolutiondue to current and projected capabilities to generate metagenomesequences, determine metatranscriptomic, metaproteomic and meta-metabolomic information and crucially, analyse the deluge of data andinterpret findings ecologically. The novel procedures are broadly in theeconomic realm of numerous researchers, but many do pose consid-erable technical challenges. Thepractical outcomes for host species andtheir environments are diverse.
Fundamental and applied hostmicrobiome research began very early inthe history of microbiology. Indeed, Antonie van Leeuwenhoek(1632–1723), ‘the father of microbiology’, observed and reported onwhat were large selenomonads from the human mouth in 1676. Morerecently, Robert Hungate1 (1906–2004), ‘the father of rumen micro-biology’, developed critical techniques that allowed the study of anaer-obic microbes – the roll tube technique2. Using this method that hemeticulously described, he explored methanogenesis and cellulosebiochemistry (among other metabolic functions) in ruminants and themicrobial ecology of monkey and human guts. The practical outcomesof improved milk, meat and wool production were major drivers torumen microbiology studies.
It has been nearly four decades since ‘Sanger DNA sequencing’ wasintroduced3, the first organism was sequenced4 and ribosomal RNAanalysis was used to determine the third domain of cellular life on Earth,the Archaea5. Since the late 1970s, substantial method developmentsincluding polymerase chain reaction (PCR), improved acquisition ofDNA sequences (automated DNA sequencing) and their analyses haveoccurred. The first decade of this century was part of that methodimprovement and subsequent data eruption. These fundamental
discoveries in the late 1970s were paramount in facilitating mammalianmicrobiome research.
The diversity of prokaryotes in a plethora of environments could becomprehended by applying massively parallel high throughputDNA sequencing methods (starting with 454 Life Sciences‘pyrosequencing’ in 2005) to small subunit rRNA genes. Full genomeswere determined in large numbers (currently of 31,241 prokaryotes),whole-genome phylogenies were reported, and the ‘meta-omics’ fieldsof endeavour were well and truly spawned into the general arena of theMicrobiome. The giddy rate of progress in omics is difficult to keeppace with but critical questions about microbial function and dynamics(stable and mobile) and the chemical interplay between microbes andtheir hosts should be overriding drivers of their investigation, particu-larly in mammals. Pondering the future of microbiome research, acollection of authors recently reported on their individual opinions6,and to quote from this paper:
Overall, future microbiome research regarding the mole-cules and mechanisms mediating interactions betweenmembers of microbial communities and their hosts shouldlead to discovery of excitingnewbiology and transformativetherapeutics.
The articles in thisMicrobiology Australia issue cover a broad range ofmammalian microbiome studies in Australia. The majority are onhumans (oral and gut), but marine mammals (skin, gut and respiratorytract including blowhole), ruminants and terrestrial Australian nativeanimals (oral and gut) are also explored. The motivations for the host-microbe studies in these papers cover the host species fromboth healthand well-being perspectives as well as from general ecological andanimal production improvement viewpoints. Monotreme microbialecology and potential biotechnological discoveries from the consump-tion of toxic diets (e.g. those high in essential oils like Eucalyptus spp.)should attract more attention and are of essential Australian relevance.
References1. Hungate, R.E. (1966) The Rumen and its Microbes. Academic Press, New York.
2. Hungate, R.E. and Macy, J. (1973) The roll-tube method for cultivation of strict
anaerobes. Bulletins of the Ecological Research Committee 123–126.
3. Sanger, F. et al. (1977) DNA sequencing with chain-terminating inhibitors. Proc.
Natl. Acad. Sci. USA 74, 5463–5467. doi:10.1073/pnas.74.12.5463
4. Sanger, F. et al. (1977) Nucleotide sequence of bacteriophage PHICHI174 DNA.
Nature 265, 687–695. doi:10.1038/265687a0
5. Woese, C.R. and Fox, G.E. (1977) Phylogenetic structure of prokaryotic domain –
primary kingdoms. Proc. Natl. Acad. Sci. USA 74, 5088–5090. doi:10.1073/
pnas.74.11.5088
6. Waldor, M.K. et al. (2015) Where next for microbiome research? PLoS Biol.
doi:10.1371/journal.pbio.1002050
BiographyLinda L Blackall is a microbial ecologist who has studied manydifferent complex microbial communities ranging from host associatedthrough to free living in numerous environments. Her research hascoveredmammalian microbiomes spanningmarsupials, humans, rumi-nants and horses and the methods used allow elucidation of massivemicrobial complexity and function in these diverse biomes. She is aProfessor of Biosciences at Swinburne University of Technology in theFaculty of Science, Engineering and Technology.
GuestEditorial
MICROBIOLOGY AUSTRALIA * MARCH 2015 10.1071/MA15002 3

Methane matters in animals and man: frombeginning to end
Emily HoedtA, Paul EvansB, Stuart DenmanC, Chris McSweeneyC, Paraic ÓCuívD and Mark MorrisonD,E
ASchool of Chemistry and Molecular Biosciences, University of Queensland, St Lucia, Qld 4072, Australia
BAutralian Centre for Ecogenomics, School of Chemistry and Molecular Biosciences, University of Queensland, St Lucia, Qld 4072, Australia
CCSIRO Agriculture, Queensland Bioscience Precinct, St Lucia, Qld 4067, Australia
DUniversity of Queensland Diamantina Institute, Translational Research Institute, Woolloongabba, Qld 4102, Australia
ECorresponding author. Tel: +61 7 3443 6957, Fax: +61 7 3443 6966, Email: [email protected]
Methanogenic archaea resident in themammalian gastroin-
testinal tract have long been recognised for their capacity to
participate in interspecies hydrogen transfer,with commen-
surate positive effects on plant biomass conversion. Howev-
er, there is also stillmuch to learnabout thesemethanogenic
archaea in regards to their metabolic versatility, host adap-
tation, and immunogenic properties that is of relevance to
host health and nutrition.
Methane, man and best laid plansThe methane club has an exclusive membership, principally
restricted to the Domain Archaea and more specifically, the
Euryarchaeota. Five orders of methanogens have long been recog-
nised: the Methanopyrales, Methanococcales, Methanobacteriales,
Methanomicrobiales, andMethanosarcinales1.However, themember-
ship has recently been expanded to include the Methanocellales2
as well as the provisionally named ‘Methanoplasmatales’3. Members
of the methane club are very popular, invited to join virtually all
anaerobic microbial communities and especially those where
sulphate is limiting. Popular hangouts includemoist soil biomes, fresh
water sediments and rice paddies, landfills, the gastrointestinal tracts
of invertebrate and vertebrate animals, anaerobic lagoons and waste
management facilities4–6. Indeed, the number and distribution of
these hangouts have dramatically increased in recent decades in
response to human population growth and urbanisation, as well as
the intensification of agriculture to feed a hungry world; but the
hangoverhasarrived.Wearenowbeingchallengedtoreducemethane
gas emissions, and in particular, methane emissions from livestock
production systems, which are attributed with producing ~20% of
global methane emissions7, in response to global concerns about our
impacts on the environment and climate change. Additionally, the
resurgent interest in themicrobiota we share our body with, and their
In Focus
4 10.1071/MA15003 MICROBIOLOGY AUSTRALIA * MARCH 2015

impacts on our health and well-being, extends to the methane club8.
For these reasons, there is a renewed interest in gut methanogens,
but is it more of the same or something new? We contend that there
is still much to be learned about members of the methane club
and their behaviour in the digestive tracts of animals and man, from
beginning to end.
Separating the sheep from the goats: methane
and livestockAs herbivores, ruminants rely upon their microbial communities
within the rumen-reticulum to not only deconstruct plant biomass,
but provide the schemes of anaerobic fermentation necessary to
support the formation of protein-yielding and energy-yielding
nutrients such as microbial biomass and short chain fatty acids9,10.
Methanogens have long been recognised to support these process-
es via minimising pH2, with the concept of ‘interspecies hydrogen
transfer’ (IHT) first demonstrated by Bryant and Wolin11 using
culture based experiments with rumen bacteria and methanogens.
Much of the subsequent research focused on the taxonomic and
ecological variations among methanogen communities as affected
by diet, animal breed, and production system. In general terms,
these studies have shown that while autotrophic Methanobrevi-
bacter spp. are often numerically predominant there is also a
relatively diverse population of heterotrophic archaea present in
these animals6,12. In recent years, the application of ‘omics’
approaches has provided new insights into the roles the archaea
might play in rumen function. Poulsen et al. (2013) showed that
the reduced methane emissions from dairy cows fed rapeseed
oil could be attributed to a selective suppression of the
‘Methanoplasmatales’, with coincident decreases in transcripts
encoding for methylotrophic methanogenesis from the rumen
contents of these animals13. New Zealand and US-DOE researchers
have also studied the rumen microbiota of sheep stratified with
respect to methane production, and demonstrated that the trait
is heritable14. Using a combination of metagenomic and metatran-
scriptomic methods they found no significant differences in total
methanogen numbers between the ‘low’ and ‘high’ methane
producers, although there were differences in the relative abun-
dances of the methylotrophic Methanosphaera spp. (increased in
‘low methane’ sheep) and the hydrogenotrophic Methanobrevi-
bacter gottschalkii clade (increased in ‘high methane’ sheep). The
metatranscriptomic data revealed that 7/10 genes coordinating
the hydrogenotrophic pathway were significantly increased in high
methane producing sheep. Collectively, these findings suggest
that while the inhibition of select populations of methanogens can
mitigate livestock methane emissions, it is also a heritable trait,
suggesting host-mediated effects on the rumen microbiota. In that
context, ‘high methane’ emitting animals have been postulated
to possess a longer retention of feed within the rumen as well as
alterations in the bacterial ‘ruminotype’ increasing the levels of
ruminal hydrogen, with coordinate elevated expression of
genes encoding the hydrogenotrophic pathway and greater
methane yield15,16. It seems intuitive then to further suggest that
the increased relative abundance of hydrogen-dependent
methylotrophic methanogens in ‘low methane’ animals relates to
their capacity for growthwhen the bacterial ruminotype favours less
hydrogen production during fermentation12,17.
Differences downunder: the low methane
emitting macropodidsThemacropodids (kangaroos, wallabies, pademelons and relatives)
bear some similarity to ruminants in so far as their reliance on
forestomach colonisation bymicrobes for plant biomass conversion
and nutrient provision. In contrast, the foregut microbiota resident
in these animals releases relatively low amounts of methane com-
pared to sheep18,19. Although these observations were initially
proposed to reflect the absence of methanogenic archaea within
the macropodid forestomach, several studies have now demon-
strated the presence of Methanobrevibacter, Methanosphaera,
and ‘Methanoplasmatales’ archaea, albeit at numbers substantially
less than found for ruminant livestock (~106 g.sample–1 c.f. ~108 g.
sample–1)6. Our group has now produced an axenic culture of
Methanosphaera sp. (strain WGK6) from foregut digesta collected
from a Western grey kangaroo (Macropus fuliginosus). Like
the human strain Methanosphaera stadtmanae DSM-3091, WGK6
uses methanol for methane formation, energy production and
growth. However, the annotated draft sequence of the WGK6
genome suggests the macropodid isolate possesses some unique
features that may support a greater metabolic versatility than
previously characterised from studies of the human strain. So
it seems that the adaptations to herbivory in the ‘low methane’
emitting macropodids includes the maintenance of Methano-
sphaera spp., which also seem to be present in greater abundance
in ruminant animals individually confirmed to be ‘low methane’
emitters.
Humans and methanogens: a docile partnership
or secret frenemies?Methanogens are consistently identified from human subjects
deemed healthy or suffering from disease; however the relation-
ships between the diversity of methanogen community members
and the health status of the host are still unclear. Early studies
determined that like other mammals the human large bowel
was colonised by hydrogenotrophic Methanobrevibacter spp.
(principallyMbb. smithii) and themethylotrophicMethanosphaera
spp. (principallyMsp. stadtmanae20). More recently, the analysis of
human microbiota samples from subgingival, intestinal or vaginal
mucosae has further expanded the diversity of methanogenic
archaea to include a new species of Methanobrevibacter (Mbb.
oralis), as well as two isolates of methylotrophic archaea (Candi-
datus ‘Methanomethylophilus alvus’ and Methanomassiliicoccus
luminyensis) affiliated with the newly defined orderMethanoplas-
matales21,22. Interestingly, our own unpublished studies, as well
as the findings of Poulsen et al.13, Dridi et al.21 and Borrel et al.22
show these archaea are capable of using methylated amines arising
from phosphatidylcholine metabolism to support growth. In that
context, establishment of the Methanoplasmatales in the human
large bowel might be of clinical relevance for persons known to
In Focus
MICROBIOLOGY AUSTRALIA * MARCH 2015 5

possess relatively high levels of trimethylamine-oxide in blood,
because of its association with cardiovascular disease pathogenesis
(reviewed by Morrison23 and Brugère24). However, there is also
mounting evidence from cross-sectional studies that variations in
archaeal communities at different body sites might impact human
health25–27. For instance, patients with periodontitis have been
found to harbour large numbers of methanogenic archaea, in
addition to acetogenic and sulphate-reducing bacteria within sub-
gingival periodontal pockets28. Blais Lecours et al.29 also confirmed
that both Mbb. smithii and Msp. stadtmanae can be immunosti-
mulatory in animal models of respiratory disease, with the latter
provoking a stronger immune response. Furthermore,Blais Lecours
et al.30 reported that while the total numbers of methanogenic
archaea are less in patients suffering from inflammatory bowel
disease (IBD), the prevalence of Msp. stadtmanae was greater
in these patients, and healthy human subjects produced an
antigen-specific IgG response to this archaeon. These results
suggest that Msp. stadtmanae prevalence and/or abundance may
be a biomarker of gut dysbiosis, being more prevalent in persons
with an altered ‘low hydrogen’ fermentation scheme. This hypoth-
esis warrants more detailed examination as part of well-designed
clinical studies of IBD and perhaps, other chronic inflammatory
diseases.
SummaryDespite the widespread recognition of the roles methanogenic
archaea play in gut environments, there is still much to learn about
their metabolic versatility, host adaptation, and immunomodula-
tion. Recent research of the methylotrophic archaea from three
divergent mammalian hosts suggests that methane matters in
animals and man, from beginning to end!
References1. Anderson, I. et al. (2009) Genomic characterization of methanomicrobiales
reveals three classes of methanogens. PLoS ONE 4, e5797. doi:10.1371/journal.
pone.0005797
2. Sakai, S. et al. (2008) Methanocella paludicola gen. nov., sp. nov., a methane-
producing archaeon, thefirst isolate of the lineage ‘RiceCluster I’, andproposal of
the new archaeal order Methanocellales ord. nov. Int. J. Syst. Evol. Microbiol. 58,
929–936. doi:10.1099/ijs.0.65571-0
3. Paul, K. et al. (2012) ‘Methanoplasmatales,’ Thermoplasmatales-related archaea
in termite guts and other environments, are the seventh order of methanogens.
Appl. Environ. Microbiol. 78, 8245–8253. doi:10.1128/AEM.02193-12
4. Liu, Y. andWhitman,W.B. (2008)Metabolic, phylogenetic, andecological diversity
of the methanogenic archaea. Ann. N. Y. Acad. Sci. 1125, 171–189. doi:10.1196/
annals.1419.019
5. Edwards, T. and McBride, B.C. (1975) New method for the isolation and identi-
fication of methanogenic bacteria. Appl. Microbiol. 29, 540–545.
6. Evans, P.N. et al. (2009) Community composition and density of methanogens
in the foregut of the Tammar wallaby (Macropus eugenii). Appl. Environ.
Microbiol. 75, 2598–2602. doi:10.1128/AEM.02436-08
7. Lowe, D.C. (2006) Global change: a green source of surprise. Nature 439,
148–149. doi:10.1038/439148a
8. Samuel, B.S. et al. (2007) Genomic and metabolic adaptations of Methanobre-
vibacter smithii to the human gut. Proc. Natl. Acad. Sci. USA 104, 10643–10648.
doi:10.1073/pnas.0704189104
9. Karasov, W.H. and Carey, H.V. (2009) Metabolic teamwork between gut microbes
and hosts. Microbe 4, 323–328.
10. Hobson, P.N. (1988) The RumenMicrobial Ecosystem, First edn. Elsevier Applied
Science, New York.
11. Bryant, M.P. andWolin, M.J. (1975) Proceedings of the first international congress
of the international association of the microbiological society, Developmental
Microbiology E, Science Council of Japan, Tokyo, Japan. p. 297.
12. Janssen, P.H. and Kirs, M. (2008) Structure of the archaeal community of the
rumen. Appl. Environ. Microbiol. 74, 3619–3625. doi:10.1128/AEM.02812-07
13. Poulsen, M. et al. (2013) Methylotrophic methanogenic Thermoplasmata impli-
cated in reduced methane emissions from bovine rumen. Nat Commun 4, 1428.
doi:10.1038/ncomms2432
14. Shi, W. et al. (2014) Methane yield phenotypes linked to differential gene
expression in the sheep rumen microbiome. Genome Res. 24, 1517–1525.
doi:10.1101/gr.168245.113
15. Janssen, P.H. (2010) Influence of hydrogen on rumen methane formation and
fermentation balances through microbial growth kinetics and fermentation
thermodynamics. Anim. Feed Sci. Technol. 160, 1–22. doi:10.1016/j.anifeedsci.
2010.07.002
16. Kittelmann, S. et al. (2014) Two different bacterial community types are linked
with the low-methane emission trait in sheep. PLoS ONE 9, e103171. doi:10.1371/
journal.pone.0103171
17. Attwood, G.T. et al. (2011) Exploring rumen methanogen genomes to identify
targets for methane mitigation strategies. Anim. Feed Sci. Technol. 166–167,
65–75. doi:10.1016/j.anifeedsci.2011.04.004
18. Madsen, J. and Bertelsen, M.F. (2012) Methane production by red-necked walla-
bies (Macropus rufogriseus). J. Anim. Sci. 90, 1364–1370. doi:10.2527/jas.2011-
4011
19. von Engelhardt, W. et al. (1978) Production of methane in two non-ruminant
herbivores Comp. Biochem. Physiol. Part A. Physiol. 60, 309–311. doi:10.1016/
0300-9629(78)90254-2
20. Miller, T.L. and Wolin, M.J. (1985) Methanosphaera stadtmaniae gen. nov., sp.
nov.: a species that forms methane by reducing methanol with hydrogen. Arch.
Microbiol. 141, 116–122. doi:10.1007/BF00423270
21. Dridi, B. et al. (2012) Methanomassiliicoccus luminyensis gen. nov., sp. nov., a
methanogenic archaeon isolated from human faeces. Int. J. Syst. Evol. Microbiol.
62, 1902–1907. doi:10.1099/ijs.0.033712-0
22. Borrel, G. et al. (2012)Genome sequence of ‘CandidatusMethanomethylophilus
alvus’ Mx1201, a methanogenic archaeon from the human gut belonging to a
seventh order of methanogens. J. Bacteriol. 194, 6944–6945. doi:10.1128/
JB.01867-12
23. Morrison,M. (2013) Looking large, tomakemore, out of gutmetagenomics.Curr.
Opin. Microbiol. 16, 630–635. doi:10.1016/j.mib.2013.10.003
24. Brugère, J.F. et al. (2014) Archaebiotics: proposed therapeutic use of archaea to
prevent trimethylaminuria and cardiovascular disease. Gut Microbes 5, 5–10.
doi:10.4161/gmic.26749
25. Furnari, M. et al. (2012) Reassessment of the role ofmethane production between
irritable bowel syndrome and functional constipation. J. Gastrointestin. Liver Dis.
21, 157–163.
26. Pimentel, M. et al. (2003) Methane production during lactulose breath test is
associated with gastrointestinal disease presentation. Dig. Dis. Sci. 48, 86–92.
doi:10.1023/A:1021738515885
27. Lepp, P.W. et al. (2004) Methanogenic Archaea and human periodontal disease.
Proc. Natl. Acad. Sci. USA 101, 6176–6181. doi:10.1073/pnas.0308766101
28. Vianna, M.E. et al. (2008) Quantitative analysis of three hydrogenotrophic micro-
bial groups, methanogenic archaea, sulfate-reducing bacteria, and acetogenic
bacteria, within plaque biofilms associated with human periodontal disease.
J. Bacteriol. 190, 3779–3785. doi:10.1128/JB.01861-07
29. Blais Lecours, P. et al. (2011) Immunogenic properties of archaeal species found
in bioaerosols. PLoS ONE 6, e23326. doi:10.1371/journal.pone.0023326
30. Blais Lecours, P. et al. (2014) Increased prevalence ofMethanosphaera stadtma-
nae in inflammatory bowel diseases. PLoS ONE 9, e87734. doi:10.1371/journal.
pone.0087734
BiographiesEmily Hoedt is a PhD student at The University of Queensland
SchoolofChemistry andMolecularBiosciences, and is supervisedby
In Focus
6 MICROBIOLOGY AUSTRALIA * MARCH 2015

Mark Morrison, Phil Hugenholtz and Gene Tyson. Her PhD studies
focus on functional and comparative studies of heterotrophic
methanogens from different gut environments, supported by an
Australian Postgraduate Award and a top-up scholarship from Meat
and Livestock Australia.
Paul Evans is a Postdoctoral Fellow, based at the Australia Centre
for Ecogenomics at University of Queensland, Brisbane and his
research interests are related to microbial ecology in anaerobic
environments. His research involves examinations of novel
microbes from a range of environments including ruminants, per-
mafrost soils and coal bed methane aquifers. Paul is specifically
interested in archaea that populate these anaerobic environments
and how they interact with other members of the microbial com-
munity to produce energy and make their living.
Stuart Denman is a research scientist with the CSIRO in the
Agriculture Flagship. He has been actively involved in projects that
use molecular methods to detect and monitor key microbial popu-
lations within the rumen. His current research focus is onmicrobial
metagenomics and uses advanced molecular and bioinformatic
techniques to ascertain the interactions and functional changes
that take place in the rumenmicrobiome as they pertain tomethane
abatement strategies.
Chris McSweeney is a Senior Principal Research Scientist at
CSIRO and leads research into the gut microbiology of livestock,
humans and native animals. His current research is focussed on the
molecular basis of hydrogenotropy in gut microbial ecosystems
with emphasis on the function ofmethanogenic archaea. The aim is
to eventually modify the ecosystem to reduce methane emissions
from ruminant livestock.
Dr Páraic ÓCuív is a gut microbiologist at The University of
Queensland Diamantina Institute where he has a long standing
interest in host-microbe interactions as they relate to the aetiology
of chronic gut diseases. He is an expert in gut microbiology,
microbial genetics and functional metagenomics and he currently
leads the isolation and functional characterisation of microorgan-
isms from the human gut as part of the Australian Healthy Micro-
biome Project.
Professor Mark Morrison is trained as a microbiologist with a
specific interest in the role thatmicrobes play in affecting the health
and well-being of humans and animals. After nearly 20 years within
US academia, he returned to Australia in 2006 initially as a Science
Leader within CSIRO, and now as Chair and Group Leader in
Microbial Biology and Metagenomics at The University of Queens-
land Diamantina Institute. His work since returning to Australia
has emphasised the use of ‘omics’ technologies to produce new
insights into the microbial world, and from which, improved meth-
ods for monitoring and adjustment of the gut microbiota might
be achieved; with the goal of improving host animal health and
well-being.
In Focus
MICROBIOLOGY AUSTRALIA * MARCH 2015 7

The marine mammal microbiome: currentknowledge and future directions
Marine mammals are globally significant because of their
sensitivity to environmental change and threatened status,
often serving as ‘ecosystem sentinels’1. Disease is a major
cause of marinemammal population decline and the role of
the microbiome in disease has generated considerable
interest. Recent research in humans has greatly enhanced
our understanding of how the host-associated microbial
community, the microbiome, affects host health. In this
review, we provide an overview of the extent of the marine
mammal microbiome with a focus on whole community
characterisation using genomic methods. This research
highlights the overlap in microbial communities between
geographically distinct species and populations of marine
mammals, suggesting tight links betweenmarinemammals
and their microbial symbionts over millions of years of
evolution. An understanding of these links in both healthy
and compromised hosts is essential to identifying at-risk
populations and making ecologically appropriate manage-
ment decisions. We advocate further development of
innovative sampling and analytic techniques that advance
the field of microbial ecology of marine mammals.
Recent investigations have highlighted the capacity of the micro-
biome to act strongly and significantly in maintaining host health
with a vital role in disease manifestation and immune system
function2,3. Members of the microbial community can directly
influence the progression of a disease via infection and also mod-
ulate the host’s own immune system regulation and response4.
Indeed thehost’smicrobial partners areessential to immunesystem
function. Themicrobiome has been observed to be species-specific
in a variety of vertebrate hosts5–7 and is influenced by host phylog-
eny, as a result ofmillionsof yearsof co-evolution8.Marinemammals
represent unique evolutionary lineages and investigations into their
associated microbes will provide a deeper understanding of their
ecology and evolution.
Marine mammals form a diverse group of 129 species in three
orders, and of those, 28 are considered endangered or threatened9.
Disease is one of the main causes of death in marine mammals and
somepopulationshave sufferedmassmortalities causedbybacterial
pathogens10. Bacteria exist as part of the normal, or even beneficial,
flora associated with a host, fluctuating and changing with a host’s
physiology andmetabolism11. Inmammals, disease canoccur under
Tiffanie M NelsonA,F, Amy ApprillB, Janet MannC, Tracey L RogersDand Mark V BrownD,E
ADepartment of Animal and Range Sciences, Montana State University, Bozeman, MT 59715, USA
BWoods Hole Oceanographic Institution, 266 Woods Hole Road, Mailstop #4, Woods Hole, MA 02543, USA
CGeorgetown University, Regents Hall 516, Washington, DC 20057, USA
DEvolution and Ecology Research Centre, University of New South Wales, Kensington, NSW 2052, Australia
ESchool of Biotechnology and Biomolecular Sciences, University of New South Wales, Kensington, NSW 2052,Australia
FCorresponding author. Tel: +1 406 539 6898, Email: [email protected]
In Focus
8 10.1071/MA15004 MICROBIOLOGY AUSTRALIA * MARCH 2015

a number of different circumstances, most commonly on occasions
when the host’s immune system is compromised. For marine
mammals, susceptibility to pathogens may be particularly elevated
due to anthropogenic stressors such as depleted food resources,
habitat degradation and chemical or sound exposure12–15. Addi-
tionally, succession events occurring after an initial bacterial infec-
tionmay lead to dysbiosis, and alterations in the host’s microbiome
may be a better predictor of disease progression than following the
presence of individual pathogenic agents16. Hence, we need to
establish baseline data on microorganisms commonly associated
with marine mammals in order to detect anomalies. In the last
decade genomic sequencing technologies have provided a previ-
ously unrecognised diversity of microorganisms in numerous
diverse habitats. In this brief review we highlight the current
knowledgeof themicrobial composition in associationswithmarine
mammals with a focus on whole community characterisation.
Skin microbiomeSkin, as the largest organ of mammals, serves as a thick physical
barrier that provides defense against the surrounding marine
environment. Marine mammal skin is prone to lesions and disor-
ders, however the role of microorganisms in these conditions is
still largely unresolved and knowledge is primarily founded on
cultivation-based studies17. The recent application of cultivation-
independent sequencing-survey approaches to humpback whale
(Megaptera novaeangliae) skin has demonstrated that a unique
ecosystem of microbes resides on the skin surface (Table 1), which
differs from the community present in seawater18.
Among populations of humpback whales surveyed in diverse geo-
graphic regions, two genera of bacteria (Bacteroidetes genus Tena-
cibaculum and Gammaproteobacteria genus Psychrobacter) were
found to be cosmopolitan and abundant associates on humpback
whale skin26. Scanning electron microscopy of humpback whale
skin revealed a rich layer of microbial cells on the skin surface26, but
ashumpbackwhales regularly undergo skin sloughing throughboth
behavioural27 and physiological activities28 it is possible that the
robust Tenacibaculum and Psychrobacter cells may have some
means to maintain their residence on the whale skin and could
provide benefits to their host. Sequencing survey-based data also
demonstrate differences between the skin bacterial associates of
healthy and health-compromised humpbacks18,26. Additional data
on and study of the skinmicrobiomemight potentially improve our
ability to assess health status among free-rangingmarine mammals,
in particular cetaceans.
Gut microbiomeThe gastrointestinal tract is home to an abundant community of
microorganisms. The gutmicrobiomeplays a significant role in food
breakdown and digestion, the production of essential vitamins and
minerals and regulationof the immunesystem3. In youngmammals, Table
1.Relativeabundanceofbacterialp
hyla
comparedbetw
eenknownstudiesofmarinemammalspeciesandanatomicalsitesin
healthyindividuals
Order
Cetacea
Carnivora
Sirenia
Sub-O
rder
Mysticeti
Odontoceti
Phocidae
Pinnipedia
Com
mon
name
Hum
pbackwhale
Bottlenose
dolphin
Leopard
seal
Sothern
elephant
seal
Hooded
seal
Harbour
seal
Grey
seal
Australianfurseal
Australian
sealion
Dugong
Manatee
Species
Megaptera
noveangliae
Turisops
truncatus,
T.aduncus,
hybridA
T.truncatus
Hydrurga
leptonyx
Miroungaleonina
Cystophora
cristata
Phoca
vitulina
Halichoerus
grypus
Arctocephalus
pusillusdoriferus
Neophoca
cinera
Dugong
dugong
Trichechus
manatus
latirostris
Age
group
Adult
Calf
Adultand
Sub-adult
NR
Adult
Adultand
sub-adult
Pup
NR
NR
NR
9m
pup
6m
pup
2m
pup
NR
Adult
Adult
Sub-adult
Calf
Sam
ple
Skin
Skin
Blow
Blow
Faeces
Faeces
Faeces
Colon
Colon
Colon
Faeces
Faeces
Faeces
Faeces
Faeces
Faeces
Faeces
Faeces
Bacterial
phyla(%
of
community)
Firm
icutes
1<1
15
4443
1822
5076
8387
8380
8379
7971
Bacteroidetes
4063
134
821
1468
4924
106
42
1517
1926
Proteobacteria
6036
5060
3115
59
00
2<1
48
<1<1
<1<1
Fusobacteria
<1<1
1<1
1320
621
<1<1
<1<1
<1<1
0<1
<1<1
Num
berofindividuals
516
244
1218
69
11
44
41
118
117
Methodology
PP
PCL
PP
PCL
CL
CL
PP
PM
CL
PP
P
Reference
19
19
19
20
77
721
21
21
22
22
22
23
24
25
25
25
Datasummarised
forthedominantbacterialphylaacross
speciesandanatom
icalsites.Tabledataareas
follows:notrecorded(NR);month(m);clonelibraries
(CL);pyrosequencing(P);metagenom
icsequencing
(M).
AHybridbottlenose
dolphinreferstoindividualssiredby
T.truncatustoT.aduncusfemales
born
incaptivity.
In Focus
MICROBIOLOGY AUSTRALIA * MARCH 2015 9

the gutmicrobiome is required for full development of the immune
system and maturation of the gut29,30. Studies of the complete gut
microbiome of marine mammals include leopard seals (Hydrurga
leptonyx), southern elephant seals (Mirounga leonine), grey seals
(Halichoerus grypus), hooded seals (Cystophora cristata), harbor
seals (Phoca vitulina), Australian fur seals (Arctocephalus pusillus
doriferus), Australian sea lions (Neophoca cinerea), Florida mana-
tees (Trichecus manatus latirostris) and dugongs (Dugong du-
gong). Across all these species the gut microbiome is composed
largely of Firmicutes, Bacteroidetes and Proteobacteria (Table 1).
Diet and age have been identified as factors that shape the com-
position of the gut microbiome7,25.
Amongst the seals, the gut microbiome of pinnipeds has a greater
abundance of the phylum Firmicutes compared with phocids
(Table 1). A ‘core’ group of microorganisms including the genera
Ilyobacter, Psychrilyobacter, Fusobacterium, Bacteroides, Subdo-
lingranulum, Sporobacter, Sutterella,Weisella, Anaerococcus and
Campylobacterhavebeenobservedwithinphocid seals7,21,22whilst
their herbivorous relatives, within the order Sirenia, shared mem-
bers from the order Clostridiales, including the genera Clostridium
and Ruminococcus24,25,31. The presence of shared bacterial oper-
ational taxonomic units (OTUs) in multiple hosts from different
studies highlights the strong phylogenetic influence on microbial
assembly.
Respiratory microbiomeRespiratory illnesses such as pneumonia are a major cause of
mortality in both wild and captivemarinemammals32. The cetacean
upper respiratory tract terminates in a blowhole, positioned at the
top of the head. This feature is a unique adaptation to life in the
marine environment, and allows airways to be effectively sealed off
from seawater. Upon surfacing, cetaceans forcefully exhale and in
the process eject a substance termed blow (also called condensed
respiratory vapor or exhaled breath condensate). This material has
been shown toharbour potential pathogens inwhales33 andhas also
been used to characterise the normal respiratory-associated micro-
biome residing in the upper respiratory tract of bottlenose dol-
phins19,20 (see collection methods in Figure 1). Members of the
bacterial genera Plesiomonas, Aeromonas, Escherichia, Clostridi-
um and Pseudomonas, Burkholdaria, Mycobacterium, Haemo-
phylis, Streptococcus and Staphylococcus (including multiple
resistant Staphylococcus aureus) have been detected in both
sick/dead34 and healthy, free-ranging cetaceans20,33,35.
Blow samples from both free-ranging Tursiops truncatus and
captive T. aduncus and T. truncatus were dominated by three
novel dolphin associated clades (termed DAC 1, 2 and 3) within
the Cardiobacteraceae lineage of the Gammaproteobacteria19,20.
The Cardiobacteraceae are facultative anaerobic, Gram-negative
rod-shaped cells, members of which form part of the commensal
microbiome of humans, and whose growth is enhanced by the
presence of carbon dioxide36, which occurs in high abundances at
the termination of the respiratory tract. Representatives from each
of DAC 1, 2, and 3 have been present in every bottlenose dolphin
surveyed thus far, although themajority of sequences are associated
with DAC 3, indicating this is likely a ubiquitous and critical com-
ponent of the dolphin respiratory system. Other ‘core’ taxa asso-
ciated with the dolphin respiratory microbial community appear
to include the Arcobacter, Hydrogenimonaceae, Halotalea, Aqui-
marina, Helococcus, Mycetocola, Methylococcus and Marinimi-
crobium19. Temporal analysis of captive dolphins suggests
community composition in healthy animals is quite stable
and that individual dolphins harbour consistently unique microbial
communities19.
Sampling techniquesSampling of material for microbiological analysis from marine
mammals is logistically challenging (reviewed by Hunt et al.37),
hence themajority of information onmicrobial disease comes from
captiveor strandedanimals that arenotnecessarily representativeof
(a)
(b)
Figure 1. Exhaled ‘blow’ samples provide access to respiratorymicrobiome, host DNA, hormones and associated metabolites.Bottlenose dolphins can be trained to exhale on demand allowingcollections to be made routinely as shown here by Jillian Wisse fromthe National Aquarium in Baltimore, Maryland, USA in captive dolphins(a) and Dr Ewa Krzyszczyk, collecting samples from wild bottlenosedolphins that visit a beach in Shark Bay, WA, Australia (b). Photo creditmonkeymiadolphins.org.
In Focus
10 MICROBIOLOGY AUSTRALIA * MARCH 2015

the greater wild population. However, current sampling methods
(see examples in Figures 1 and 2) still provide considerable insight
into the microbiome of marine mammals. Capture by sedation or
restraint has been employed on smaller species such as seals and
dolphins7,38,39 and has recently been used for some larger whales40.
However, there are few opportunities to sample using these meth-
ods. It is increasingly common to use biopsy darts for collection of
skin and blubber samples for genetic and, now, microbiological
studies18,41. Permissions for biopsy sampling can be challenging for
some species ofmarinemammals, and repeated samplings areoften
not possible for the same individuals. In order to increase existing
data on the marine mammal microbiome, logistically feasible, non-
or minimally-invasive sampling protocols that are easily reproduc-
ible and provide biological material suitable for a range of studies
are necessary. For example, respiratory blow can be used to
examine host DNA42 and hormone levels43,44 as well as respiratory
associated microorganisms19,33,37, while non-invasively collected
fecal samples can be used to study host DNA45, prey items46 and
the gut microbiome22,23.
Future researchIt appears likely that there are deep branching clades of bacteria
that are uniquely associated with marine mammals and have been
conserved throughout the evolution of their hosts. Many bacterial
sequences obtained from marine mammal studies have close rela-
tives that originate from other marine mammal species. This has
significant implications for the transmission of disease amongst
these hosts. As they are usually highly social animals, there are
numerous opportunities for the transfer of microorganisms
between individuals47. Diseases inmarinemammals have also been
shown tohave their roots inothermammals, includingdogs48,49 and
humans50. In many cases where disease has caused significant
mortality in wild marine mammals, it has been linked to viruses,
includingmorbillivirus, phocine distemper and influenza virus51–55.
Despite these links being made there is really very little known
regarding the ecological role of viruses in marine mammal hosts.
Further investigations into the factors responsible for shaping the
marine mammal microbiome need to be made. Designing studies
that control for host variation will allow us to make headway in our
understanding of disease manifestation. Studies that focus on the
functionality of themicrobiomewill reveal the interactions between
host and the microbial community23,56. In human subjects, similar
target investigations have allowed for the development of novel
metabolites to treat and prevent disease57. Unlike humans, howev-
er, to access adequatebiologicalmaterial, stridesneed tobe taken to
develop innovative andnon-invasive techniques for the collectionof
relevant samples from wild populations.
AcknowledgementsWe thank Dr Ewa Krzyszczyk and Jillian Wisse for allowing us to use
their photographs.
References1. Moore, S.E. (2008) Marine mammals as ecosystem sentinels. J. Mammal. 89,
534–540. doi:10.1644/07-MAMM-S-312R1.1
2. Hooper, L.V. et al. (2002) How host-microbial interactions shape the nutrient
environment of the mammalian intestine. Annu. Rev. Nutr. 22, 283–307.
doi:10.1146/annurev.nutr.22.011602.092259
3. Bäckhed, F. et al. (2005)Host-bacterialmutualism in thehuman intestine. Science
307, 1915–1920. doi:10.1126/science.1104816
4. Maynard, C.L. et al. (2012) Reciprocal interactions of the intestinal microbiota
and immune system. Nature 489, 231–241. doi:10.1038/nature11551
5. Yildirim,S.etal. (2010)Characterizationof the fecalmicrobiome fromnon-human
wild primates reveals species specific microbial communities. PLoS ONE 5,
e13963. doi:10.1371/journal.pone.0013963
6. McKenzie, V.J. et al. (2012) Co-habiting amphibian species harbor unique skin
bacterial communities in wild populations. ISME J. 6, 588–596. doi:10.1038/
ismej.2011.129
7. Nelson, T.M. et al. (2013) Diet and phylogeny shape the gut microbiota of
Antarctic seals: a comparison of wild and captive animals. Environ. Microbiol.
15, 1132–1145. doi:10.1111/1462-2920.12022
8. Ley, R.E. et al. (2008) Evolution ofmammals and their gutmicrobes. Science 320,
1647–1651. doi:10.1126/science.1155725
9. Pompa, S. et al. (2011) Global distribution and conservation of marine mammals.
Proc. Natl. Acad. Sci. USA 108, 13600–13605. doi:10.1073/pnas.1101525108
10. Waltzek, T.B. et al. (2012) Marine mammal zoonoses: a review of disease
manifestations. Zoonoses Public Health 59, 521–535. doi:10.1111/j.1863-2378.
2012.01492.x
11. Pamer, E.G. (2007) Immune responses to commensal and environmental
microbes. Nat. Immunol. 8, 1173–1178. doi:10.1038/ni1526
12. Mos, L. et al. (2006) Chemical and biological pollution contribute to the immu-
nological profiles of free-ranging harbor seals. Environ. Toxicol. Chem. 25,
3110–3117. doi:10.1897/06-027R.1
13. Fair, P.A. et al. (2013) Associations between perfluoroalkyl compounds and
immune and clinical chemistry parameters in highly exposed bottlenose dolphins
(Tursiops truncatus).Environ.Toxicol.Chem.32, 736–746.doi:10.1002/etc.2122
14. Kight, C.R. and Swaddle, J.P. (2011) How and why environmental noise impacts
animals: an integrative,mechanistic review.Ecol. Lett.14, 1052–1061.doi:10.1111/
j.1461-0248.2011.01664.x
Figure 2. Collection of samples fromwildmarinemammals is logisticallychallenging. The collection of quality biological material with minimalimpact on the animal requires the development of innovative samplingmethods. This photo shows petri dishes attached to a modified pole forthe collection of exhaled ‘blow’ samples from a southern humpbackwhale off the coast of northeast Australia. Photo credit Tracey Rogers.
In Focus
MICROBIOLOGY AUSTRALIA * MARCH 2015 11

15. Kannan, K. et al. (2007) A comparative analysis of polybrominated diphenyl
ethers and polychlorinated biphenyls in southern sea otters that died of infectious
diseases andnoninfectious causes. Arch. Environ. Contam. Toxicol.53, 293–302.
doi:10.1007/s00244-006-0251-8
16. Klepac-Ceraj, V. et al. (2010) Relationship between cystic fibrosis respiratory
tract bacterial communities and age, genotype, antibiotics and Pseudomonas
aeruginosa. Environ. Microbiol. 12, 1293–1303. doi:10.1111/j.1462-2920.2010.
02173.x
17. Mouton, M. and Botha, A. (2012) Cutaneous lesions in cetaceans: an indicator of
ecosystem status? in New Approaches to the Study of Marine Mammals,
A. Romero and E.O. Keith, Editors. InTech.
18. Apprill, A. et al. (2011) Humpback whales harbour a combination of specific and
variable skin bacteria. Environ. Microbiol. Rep. 3, 223–232. doi:10.1111/
j.1758-2229.2010.00213.x
19. Lima, N. et al. (2012) Temporal stability and species specificity in bacteria
associated with the bottlenose dolphins respiratory system. Environ. Microbiol.
Rep. 4, 89–96. doi:10.1111/j.1758-2229.2011.00306.x
20. Johnson, W.R. et al. (2009) Novel diversity of bacterial communities associated
with bottlenose dolphin upper respiratory tracts. Environ. Microbiol. Rep. 1,
555–562. doi:10.1111/j.1758-2229.2009.00080.x
21. Glad, T. et al. (2010) Ecological characterisation of the colonicmicrobiota in arctic
and sub-arctic seals. Microb. Ecol. 60, 320–330. doi:10.1007/s00248-010-9690-x
22. Smith, S.C. et al. (2013) Age-related differences revealed in Australian fur seal
Arctocephalus pusillus doriferus gut microbiota. FEMS Microbiol. Ecol. 86,
246–255. doi:10.1111/1574-6941.12157
23. Lavery, T.J. et al. (2012) High nutrient transport and cycling potential revealed
in the microbial metagenome of Australian sea lion (Neophoca cinerea) faeces.
PLoS ONE 7, e36478. doi:10.1371/journal.pone.0036478
24. Tsukinowa, E. et al. (2008) Fecal microbiota of a dugong (Dugong dugong) in
captivity at Toba Aquarium. J. Gen. Appl. Microbiol. 54, 25–38. doi:10.2323/
jgam.54.25
25. Merson, S.D. et al. (2014) Variation in the hindgut microbial communities of
the Florida manatee, Trichechus manatus latirostris over winter in Crystal River,
Florida. FEMS Microbiol. Ecol. 87, 601–615. doi:10.1111/1574-6941.12248
26. Apprill, A. et al. (2014) Humpback whale populations share a core skin bacterial
community: towards a health index for marine mammals? PLoS ONE 9, e90785.
doi:10.1371/journal.pone.0090785
27. Clapham, P.J. et al. (1993) High-energy behaviors in humpback whales as a source
of sloughed skin formolecular analysis.Mar.Mamm. Sci.9, 213–220. doi:10.1111/
j.1748-7692.1993.tb00448.x
28. Durban, J.W. andPitman, R.L. (2012) Antarctic killerwhalesmake rapid, round-trip
movements to subtropical waters: evidence for physiological maintenance
migrations? Biol. Lett. 8, 274–277. doi:10.1098/rsbl.2011.0875
29. Palmer, C. et al. (2007) Development of the human infant intestinal microbiota.
PLoS Biol. 5, e177. doi:10.1371/journal.pbio.0050177
30. Vael, C. and Desager, K. (2009) The importance of the development of the
intestinal microbiota in infancy. Curr. Opin. Pediatr. 21, 794–800. doi:10.1097/
MOP.0b013e328332351b
31. Eigeland, K. (2012) Bacterial community structure in the hindgut of wild and
captive dugongs (Dugong dugon). Aquat. Mamm. 38, 402–411. doi:10.1578/
AM.38.4.2012.402
32. Venn-Watson, S. et al. (2012) Thirty year retrospective evaluation of pneumonia
in a bottlenose dolphin Tursiops truncatus population. Dis. Aquat. Organ. 99,
237–242. doi:10.3354/dao02471
33. Acevedo-Whitehouse, K. et al. (2010) A novel non-invasive tool for disease
surveillance of free-ranging whales and its relevance to conservation programs.
Anim. Conserv. 13, 217–225. doi:10.1111/j.1469-1795.2009.00326.x
34. Buck, C.D. and Schroeder, J.P. (1990) Public health significance of marine
mammal disease, in Handbook of Marine Mammal Medicine, L.A. Dierauf,
Editor. CRC Press, Boca Raton, FL. pp. 163–173.
35. Morris, P.J. et al. (2011) Isolation of culturable microorganisms from free-ranging
bottlenose dolphins (Tursiops truncatus) from the southeastern United States.
Vet. Microbiol. 148, 440–447. doi:10.1016/j.vetmic.2010.08.025
36. Savage, D.D. et al. (1977) Cardiobacterium hominis endocarditis: description
of two patients and characterization of the organism. J. Clin. Microbiol. 5, 75–80.
37. Hunt, K.E. et al. (2013) Overcoming the challenges of studying conservation
physiology in large whales: a review of available methods. Conservation Physi-
ology 1, cot006. doi:10.1093/conphys/cot006
38. Ortiz, R.M. and Worthy, G.A.J. (2000) Effects of capture on adrenal steroid
and vasopressin concentrations in free-ranging bottlenose dolphins (Tursiops
truncatus). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 125, 317–324.
doi:10.1016/S1095-6433(00)00158-6
39. Fair, P.A. et al. (2014) Stress response of wild bottlenose dolphins (Tursiops
truncatus) during capture-release health assessment studies. Gen. Comp. Endo-
crinol. 206, 203–212. doi:10.1016/j.ygcen.2014.07.002
40. St Aubin, D.J. et al. (2001) Hematology and plasma chemistry as indicators of
health and ecological status in beluga whales, Delphinapterus leucas. Arctic 54,
317–331. doi:10.14430/arctic791
41. Palsbøll, P.J. et al. (1997) Genetic tagging of humpback whales. Nature 388,
767–769. doi:10.1038/42005
42. Frère, C.H. et al. (2010) Thar she blows! A novel method for DNA collection from
cetacean blow. PLoS ONE 5, e12299. doi:10.1371/journal.pone.0012299
43. Hogg, C.J. et al. (2005) Determination of testosterone in saliva and blow of
bottlenose dolphins (Tursiops truncatus) using liquid chromatography-mass
spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 814,
339–346. doi:10.1016/j.jchromb.2004.10.058
44. Hunt, K.E. et al. (2014) Detection of steroid and thyroid hormones via
immunoassay of North Atlantic right whale (Eubalaena glacialis) respiratory
vapor. Mar. Mamm. Sci. 30, 796–809. doi:10.1111/mms.12073
45. Green, M.L. et al. (2007) Noninvasive methodology for the sampling and extrac-
tion of DNA from free-ranging Atlantic spotted dolphins (Stenella frontalis).
Mol. Ecol. Notes 7, 1287–1292. doi:10.1111/j.1471-8286.2007.01858.x
46. Deagle, B.E. et al. (2009) Analysis of Australian fur seal diet by pyrosequencing
prey DNA in faeces. Mol. Ecol. 18, 2022–2038. doi:10.1111/j.1365-294X.2009.
04158.x
47. Lombardo, M. (2008) Access to mutualistic endosymbiotic microbes: an under-
appreciated benefit of group living. Behav. Ecol. Sociobiol. 62, 479–497.
doi:10.1007/s00265-007-0428-9
48. Butina, T.V. et al. (2010) Canine distemper virus diversity in Lake Baikal seal
(Phoca sibirica) population. Vet. Microbiol. 144, 192–197. doi:10.1016/j.vetmic.
2009.12.027
49. Greig, D.J. et al. (2014) Surveillance for zoonotic and selected pathogens in
harbor seals Phoca vitulina from central California. Dis. Aquat. Organ. 111(2),
93–106. doi:10.3354/dao02762
50. Osterhaus, A.D. (2000) Influenza B virus in seals. Science 288, 1051–1053.
doi:10.1126/science.288.5468.1051
51. Thompson, P.M. and Miller, D. (1992) Phocine distemper virus outbreak in the
Moray Firth common seal population: an estimate ofmortality. Sci. Total Environ.
115, 57–65. doi:10.1016/0048-9697(92)90032-N
52. Van Bressem, M.F. et al. (2014) Cetacean morbillivirus: current knowledge and
future directions. Viruses 6, 5145–5181.
53. Pollack, J.D. (2001) Caspian seal die-off is caused by canine distemper virus.
Trends Microbiol. 9, 108. doi:10.1016/S0966-842X(01)01988-6
54. Anthony, S.J. et al. (2012) Emergence of fatal avian influenza in New England
harbor seals. MBio 3, e00166–12. doi:10.1128/mBio.00166-12
55. Ramis, A.J. et al. (2012) Influenza A and B virus attachment to respiratory tract in
marine mammals. Emerg. Infect. Dis. 18, 817–820. doi:10.3201/eid1805.111828
56. Stewart, J.R. et al. (2014) Survey of antibiotic-resistant bacteria isolated from
bottlenose dolphins Tursiops truncatus in the southeastern USA. Dis. Aquat.
Organ. 108, 91–102. doi:10.3354/dao02705
57. Donia, M.S. et al. (2014) A systematic analysis of biosynthetic gene clusters in the
humanmicrobiome reveals a common family of antibiotics. Cell 158, 1402–1414.
doi:10.1016/j.cell.2014.08.032
BiographiesTiffanie Nelson is a researcher from Australia currently undertak-
ing a postdoctoral fellowship atMontana StateUniversity, Bozeman,
USA. Tiff is a microbial ecologist, who focuses on the microbiome
In Focus
12 MICROBIOLOGY AUSTRALIA * MARCH 2015

of marine mammals as well as humans and environmental
samples. Her interests are in health and disease associated with
the microbiome. Tiff’s current project is investigating the vaginal
tract microbiome of women in relation to bacterial vaginosis using
both culture-dependant and -independent methods.
Amy Apprill is a researcher at the Woods Hole Oceanographic
Institution in Massachusetts, USA. Amy is a marine microbiologist
researching questions that focus on the contributionofmicroorgan-
isms to the health and ecology of marine animals. Amy is also
interested in how animal-associatedmicrobes reflect the alterations
occurring in their surrounding marine environment. Her current
research uses a combination of field measurements and observa-
tions and laboratory experiments and reliesondiversemethodology
(cultivation, genomic, metagenomic and bioinformatic) to examine
the microbiomes of reef-building corals and marine mammals.
Janet Mann is professor of biology and psychology and vice
provost for research at Georgetown University, Washington DC,
USA. Janet has expertise in the field of animal behavior with
extensive research focusing on marine mammals. Her work has
focusedon social networks, female reproduction, calf development,
life history, conservation, tool-use, social learning and culture
among bottlenose dolphins in Shark Bay, Australia. Her long-term
study ‘The Shark Bay Dolphin Research Project’, tracks over 1600
dolphins throughout their lives and includes an international team
on three continents where each group studies different aspects of
delphinid biology.
Tracey Rogers is associate professor at the University of New
South Wales. Tracey works across a diverse range of research fields
with many years of experience working in Antarctica with marine
mammals. The common theme inTracey’s research is in attempting
to understand how mammals respond to change. Tracey uses
multidisciplinary approaches to understand the ecology of mam-
mals. Most of her work uses models and techniques with captive
populations for applications in field settings. Other techniques
include stable isotope analysis, satellite telemetry and acoustics.
Mark Brown is a senior research fellow at the University of
New South Wales, Sydney, Australia. He has extensive expertise in
research that focuses onmicrobes (Bacteria, Archaea andmicrobial
Eukaryotes), primarily from marine environments. Mark’s main
interest is in investigating how microbes interact with each other
and their environment to form communities that sustain critical
ecosystem processes. His current research couples innovative
in situ sampling methods, genetic tools, bioinformatics and eco-
logical theory to elucidate and predict the form, function and
impact of microbes in rapidly changing ecosystems.
In Focus
MICROBIOLOGY AUSTRALIA * MARCH 2015 13

The role of the gut microbiome in host systems
Clarissa FebiniaA, Connie HaA, Chau LeA and Andrew HolmesA,B
ASchool of Molecular Bioscience and Charles Perkins Centre, University of Sydney, Sydney, NSW, Australia
BCorresponding author. Email: [email protected]
The presence of microbes exerts such a profound influence
on animals that they are best considered holobionts – an
organism comprised of multiple biological partners. The
concept of dysbiosis is disease states that result from unde-
sirable interactions between the partners in a holobiont.
Many molecular mechanisms that link the gut microbiome
with host health and disease have now been established and
these are giving rise to new insights in healthcare. In essence
these studies show that our microbiome is so closely inter-
twinedwith our physiology thatmicrobiome composition is
reflective of many aspects of our health. Of special impor-
tance is recognition of the intersection between chronic diet
habits and the microbiome in driving changes in our phys-
iological state. In the foreseeable future it is likely micro-
biome profiling will be a standard diagnostic test in diverse
areas of medicine and that interventions targeting the
microbiome will be developed.
All animals are associated with microorganisms for the majority of
their life, only embryonic stages are microbe-free. However the
complexity of animal-microbe interactions and the nature of their
outcomes vary. For some animals their associations with microbes
include obligate partnerships with a specific microbe that has
obvious benefits for the animals life history (e.g. Coral:Zooxanthel-
lae, Squid:Vibrio, Aphid:Buchnera). For others, the animal may
have a specialised structure in which it receives obvious benefits
frommicrobes (e.g. the rumen), but these arise via a community of
many microbial species. In contrast microbes can also interact with
animals to cause disease. For the majority of animals such specific
pathogens have historically been the focus of scientific attention.
The remaining microbes were traditionally viewed as commensals.
The past decade has seen a dramatic, and ongoing, revision of this
view with recognition that those microbes that form communities
of stable composition at various body sites (our microbiomes)
influence many aspects of our postnatal development and physiol-
ogy. This is especially true of the gut microbiome.
The links between the gut microbiome and host physiological
properties are now known to be important to the pathophysiology
ofmetabolic and immunological diseases. Studies of germ-free (GF)
animals have demonstrated robust connections between the gut
microbiome and host development and physiology. These include
roles in vascular development1 and immune cell maturation2,3.
A consequence of such developmental effects is that emergent
aspects of animal health andphysiology such as inflammatory tone4,
energy balance5,6, feeding behaviour and even mood and gross
anatomy can differ in germ-free animals. Three key points that have
emerged from these studies are schematically represented as they
might apply to human biology in Figure 1. First, the existence of GF
animals indicates the presence of microbes is not essential for the
viable development and physiology of an animal. However, GF
animals are different, with significant constraints on their environ-
mental fitness, including susceptibility to systemic infection should
they be exposed to pathogens and having additional nutritional
demands (Figure 1a). Second, if microbes are non-essential to
normal physiological processes of animals it is arguable that their
most fundamental contribution to the animals state is alteration of
how the animal system perceives and responds to its environment,
both internal and external. Finally, both microbiome association
studies and transplant studies show that different compositions of
themicrobiome are associated with different host states. Where the
microbiome composition gives benefits to desirable host functions
In Focus
14 10.1071/MA15005 MICROBIOLOGY AUSTRALIA * MARCH 2015
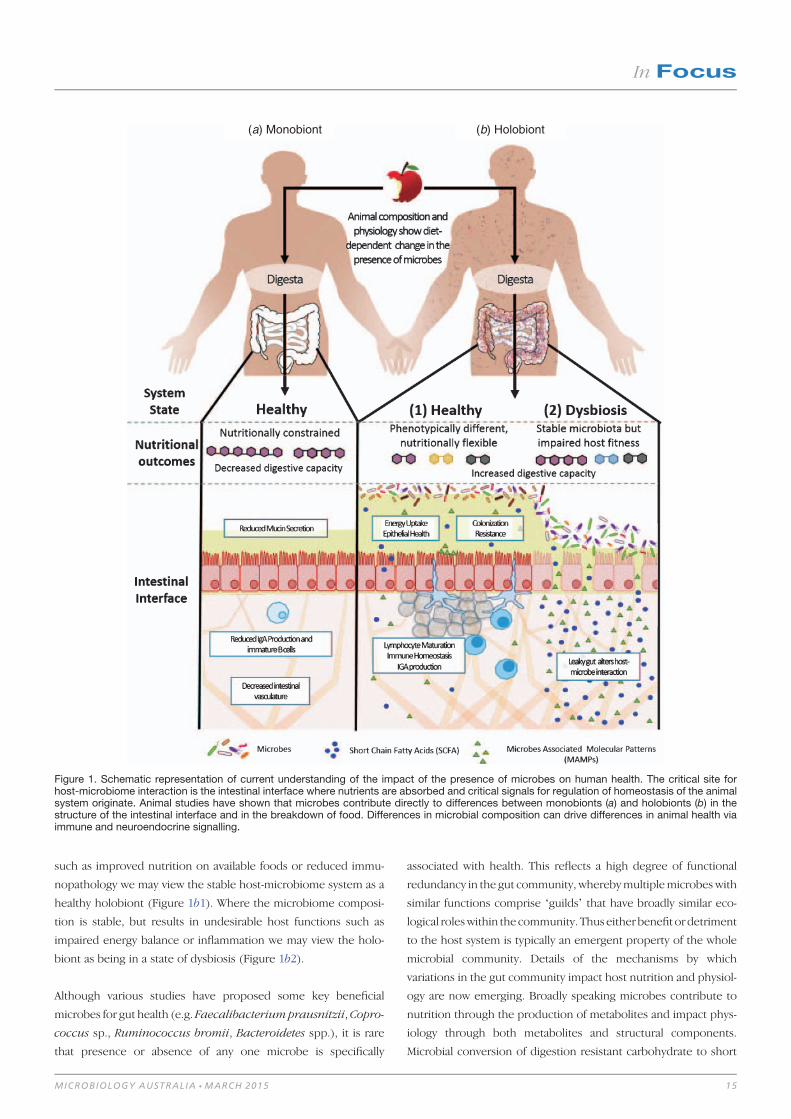
such as improved nutrition on available foods or reduced immu-
nopathology we may view the stable host-microbiome system as a
healthy holobiont (Figure 1b1). Where the microbiome composi-
tion is stable, but results in undesirable host functions such as
impaired energy balance or inflammation we may view the holo-
biont as being in a state of dysbiosis (Figure 1b2).
Although various studies have proposed some key beneficial
microbes for gut health (e.g. Faecalibacteriumprausnitzii,Copro-
coccus sp., Ruminococcus bromii, Bacteroidetes spp.), it is rare
that presence or absence of any one microbe is specifically
associated with health. This reflects a high degree of functional
redundancy in the gut community, wherebymultiplemicrobes with
similar functions comprise ‘guilds’ that have broadly similar eco-
logical roleswithin the community. Thus either benefit or detriment
to the host system is typically an emergent property of the whole
microbial community. Details of the mechanisms by which
variations in the gut community impact host nutrition and physiol-
ogy are now emerging. Broadly speaking microbes contribute to
nutrition through the production of metabolites and impact phys-
iology through both metabolites and structural components.
Microbial conversion of digestion resistant carbohydrate to short
(a) Monobiont (b) Holobiont
Figure 1. Schematic representation of current understanding of the impact of the presence of microbes on human health. The critical site forhost-microbiome interaction is the intestinal interface where nutrients are absorbed and critical signals for regulation of homeostasis of the animalsystem originate. Animal studies have shown that microbes contribute directly to differences between monobionts (a) and holobionts (b) in thestructure of the intestinal interface and in the breakdown of food. Differences in microbial composition can drive differences in animal health viaimmune and neuroendocrine signalling.
In Focus
MICROBIOLOGY AUSTRALIA * MARCH 2015 15

chain fatty acids (SCFAs) or production of vitamins both result in
increased capacity for the host system to extract nutrition from
food6,7. The SCFAs also exert other effects in the host system,
particularly butyrate which is a primary energy source for colono-
cytes, and therefore important for maintaining epithelial health.
SCFAs also impact the function of other tissues and organs in
the host by acting as signalling molecules for G-coupled protein
receptors (e.g. GPR41, GPR43). Known regulatory roles of SCFAs
include: appetite regulation, epigenetic state, gut motility, energy
metabolism, endocrine functions, and immune regulation8-10. Since
these SCFAs are primarily microbial metabolites it could be argued
that the host is monitoring the activity of its microbiome via
metabolite sensors and integrating this information into homeo-
static regulation. Similarly thehost alsomonitorsmicrobial presence
viapattern recognition receptors andsignallingpathways contribute
to regulation of diverse aspects of immune andmetabolic functions.
Significantly, disruption of the key metabolite receptors (e.g.
GPR4111) or PRR receptors (e.g. TLR512) inmouseknockoutmodels
is capable of eliciting disease states, highlighting the importance
of microbial signalling for dysbiosis. Collectively these observations
show that change in the nature or strength of various microbiome
signals, rather than presence/absence of specific microbes, is the
primary determinant of health or dysbiosis. Given this, in order
to understand dysbioses we must ask what drives disturbance
to this?
Inwild type animals this signalling-based disturbance to host-micro-
biome interactions is thought to primarily arise through changes in
microbial composition or activity. Since different microbes (e.g.
Gram-positive vs Gram-negative) contain different microbe-associ-
atedmolecular patterns (MAMPs) theydrivedifferent PRR-signalling
pathways. Similarly since microbes differ in their capacities to
degrade macromolecules and which metabolites they produce,
changes in community composition will also drive changes in
metabolite-signalling pathways. Although diverse factors including
anatomical, genotypic, cultural and environmental factors can in-
fluence the gutmicrobial community13–15, it is chronic diet patterns
that are thought to be the dominant factor, since what we eat, and
the pattern of food consumption, impact the availability of nutrients
to gut bacteria for their growth and metabolism. Arguably, the key
insight is not the role of themicrobes, but rather the role of diet as a
key modulator of the interaction between microbes and the
host6,14,16. This reflects that major mechanisms of microbial influ-
ence are via small molecules that are uniquely microbial cellular
components ormetabolites. In summary, the concepts of dysbiosis,
and animals as holobionts, are changing the way we view human
biology, especially modern diseases with a lifestyle component.
In general terms there are two routes to improve health via under-
standing of the microbiome; diagnostics and interventions. In
diagnostics, microbiome signals are included in our evaluation of
the host state to inform disease prognosis or intervention
plans. In intervention the microbiome is itself the target of manip-
ulation (e.g. prebiotics or probiotics). Greater understanding of
host-microbiome interactions can inform both routes through:
(1) identification of biomarkers of health or disease in microbiome
association studies (e.g. cancer diagnostics17); (2) identification
of specific microbes or consortia of microbes that are capable of
effecting change if introduced18–20; or (3) intervention in signalling
pathways that derive from microbes21. Progress toward these
objectives could be achieved across a very wide range of diseases
and conditions if microbial community profiling were broadly
adopted as a standard test. However, standardised experimental
protocols and metadata collection (e.g. sample collection, DNA
extraction method22, diet formula) need to be implemented in
order to discern patterns that are robust across geographically and
culturally diverse populations23.
References1. Stappenbeck, T.S. et al. (2002) Developmental regulation of intestinal angiogen-
esis by indigenous microbes via Paneth cells. Proc. Natl. Acad. Sci. USA 99,
15451–15455. doi:10.1073/pnas.202604299
2. Cebra, J.J. (1999) Influences of microbiota on intestinal immune system devel-
opment. Am. J. Clin. Nutr. 69, 1046s–1051s.
3. Rhee, K.-J. et al. (2004) Role of commensal bacteria in development of gut-
associated lymphoid tissues and preimmune antibody repertoire. J. Immunol.
172, 1118–1124. doi:10.4049/jimmunol.172.2.1118
4. Lam, Y.Y. et al. (2011) Role of the gut in visceral fat inflammation and metabolic
disorders. Obesity (Silver Spring) 19, 2113–2120. doi:10.1038/oby.2011.68
5. Bäckhed, F. et al. (2004) The gut microbiota as an environmental factor that
regulates fat storage. Proc. Natl. Acad. Sci. USA 101, 15718–15723. doi:10.1073/
pnas.0407076101
6. Turnbaugh, P.J. et al. (2006) An obesity-associated gut microbiome with
increased capacity for energy harvest. Nature 444, 1027–1031. doi:10.1038/
nature05414
7. LeBlanc, J.G. et al. (2013) Bacteria as vitamin suppliers to their host: a gut
microbiota perspective. Curr. Opin. Biotechnol. 24, 160–168. doi:10.1016/
j.copbio.2012.08.005
8. Brown, A.J. et al. (2003) The orphan G protein-coupled receptors GPR41 and
GPR43 are activated by propionate and other short chain carboxylic acids. J. Biol.
Chem. 278, 11312–11319. doi:10.1074/jbc.M211609200
9. Licciardi, P.V. et al. (2011) Histone deacetylase inhibition and dietary short-chain
Fatty acids. ISRN Allergy 2011, 869647. doi:10.5402/2011/869647
10. Kumar, H. et al. (2014) Gut microbiota as an epigenetic regulator: pilot study
based on whole-genome methylation analysis. mBio 5.
11. Samuel, B.S. et al. (2008) Effects of the gut microbiota on host adiposity are
modulated by the short-chain fatty-acid binding G protein-coupled receptor,
Gpr41. Proc. Natl. Acad. Sci. USA 105, 16767–16772. doi:10.1073/pnas.08085
67105
12. Vijay-Kumar, M. et al. (2010) Metabolic syndrome and altered gut microbiota
in mice lacking Toll-like receptor 5. Science 328, 228–231. doi:10.1126/
science.1179721
13. Ley, R.E. et al. (2008) Worlds within worlds: evolution of the vertebrate gut
microbiota. Nat. Rev. Microbiol. 6, 776–788. doi:10.1038/nrmicro1978
14. Swartz, T.D. et al. (2013) Preserved adiposity in the Fischer 344 rat devoid of gut
microbiota. FASEB J. 27, 1701–1710. doi:10.1096/fj.12-221689
15. Yatsunenko, T. et al. (2012) Human gut microbiome viewed across age and
geography. Nature 486, 222–227.
16. Fleissner, C.K. et al. (2010) Absence of intestinal microbiota does not protect
mice from diet-induced obesity. Br. J. Nutr. 104, 919–929. doi:10.1017/
S0007114510001303
In Focus
16 MICROBIOLOGY AUSTRALIA * MARCH 2015

17. Zackular, J.P. et al. (2014) The human gut microbiome as a screening tool for
colorectal cancer. Cancer Prev. Res. (Phila. Pa.) 7, 1112–1121.
18. Lawley, T.D. et al. (2012) Targeted restoration of the intestinal microbiota with
a simple, defined bacteriotherapy resolves relapsing Clostridium difficile disease
in mice. PLoS Pathog. 8, e1002995. doi:10.1371/journal.ppat.1002995
19. Adamu, B.O. and Lawley, T.D. (2013) Bacteriotherapy for the treatment of
intestinal dysbiosis caused by Clostridium difficile infection. Curr. Opin. Micro-
biol. 16, 596–601. doi:10.1016/j.mib.2013.06.009
20. Atarashi, K. et al. (2013) Treg induction by a rationally selected mixture of
Clostridia strains from the human microbiota. Nature 500, 232–236.
doi:10.1038/nature12331
21. Ryan, K.K. et al. (2014) FXR is a molecular target for the effects of vertical sleeve
gastrectomy. Nature 509, 183–188. doi:10.1038/nature13135
22. Kennedy, N.A. et al. (2014) The impact of different DNA extraction kits and
laboratories upon the assessment of human gut microbiota composition by 16S
rRNA gene sequencing. PLoS ONE 9, e88982. doi:10.1371/journal.pone.0088982
23. Finucane, M.M. et al. (2014) A taxonomic signature of obesity in the microbiome?
Getting to the guts of the matter. PLoS ONE 9, e84689. doi:10.1371/journal.
pone.0084689
BiographiesThe authors are all members of the School of Molecular Bioscience
and the Microbiome node of the Charles Perkins Centre at the
University of Sydney. Their research is focussed on understanding
the dynamics of gut microbial community composition, the
mechanisms of host-microbe interaction in the gut and develop-
ment of tools to enablemanagement of the gutmicrobial ecosystem
forhealth.Aparticular focus is the relationshipbetweenournutrient
environment and its effect on host-microbiome interactions in
health. We gratefully acknowledge funding support from the ARC
and NHMRC.
Clarissa Febinia is a postgraduate student and the recipient of an
Australia Awards Scholarship with affiliations to the Eijkman Insti-
tute forMolecular Biology, Jakarta. Her project is on the intersection
between cultural, genetic and diet factors in lifestyle disease.
Connie Ha is in the final year of PhD studies on themechanisms of
diet-induced obesity and the recipient of an APRA.
Chau Le is a PhD student working on the influence of gutmicrobes
on regulation of feeding behaviour. She is the recipient of an APRA.
Andrew Holmes is currently an Associate Professor in the School
of Molecular Bioscience at the University of Sydney and leads
research programs in the Charles Perkins Centre and Marie Bashir
Institute. He was the recipient of the 2006 Fenner Prize from the
Australian Society for Microbiology. He has general interests in
microbial diversity, its evolutionary origins and ecological applica-
tions. He is a Senior Editor forMicrobiology and The ISME Journal,
and a member of the Editorial Boards of Applied and Environmen-
tal Microbiology and Environmental Microbiology.
In Focus
MICROBIOLOGY AUSTRALIA * MARCH 2015 17

Modulation of the rumen microbiome
Rosalind GilbertA,B,D, Diane OuwerkerkA,B,E and Athol KlieveB,C,F
ARumen Ecology Unit, Department of Agriculture and Fisheries, Level 2A East, EcoSciences Precinct, Dutton Park, Qld 4102, Australia
BQueensland Alliance for Agriculture and Food Innovation, University of Queensland, St Lucia, Qld 4067, Australia
CSchool of Agriculture and Food Sciences, University of Queensland, Gatton Campus, Gatton, Qld 4343, Australia
DCorresponding author. Tel: +61 7 3255 4289, Email: [email protected]
ETel: + 61 7 3255 4291, Email: [email protected]
FTel: +61 7 5460 1255, Email: [email protected]
A combination of animal genetics and the unique, enlarged
fore-stomach of ruminants (rumen) enable domesticated
ruminants to be sustained on forages and fibrous feedstuffs
that would be otherwise indigestible. Ruminants can also
utilise more easily digestible, high energy plant material
such as grain, to achieve rapid increases in weight gain,
muscle bulk and in the case of dairy cows, high milk yields.
Since the mid-1900s there has been a steady research effort
into understanding the digestive processes of ruminants,
striving to maintain animal health and nutrition whilst
maximising theproductivity andenvironmental sustainabil-
ity of livestock production systems. This article describes
strategies developed to modulate the rumen microbial eco-
system, enabling the utilisation of plant feedstuffs that may
otherwise be toxic and enhancing feed utilisation efficiency
or controlling populations of specific rumenmicrobes, such
as those contributing to lactic acidosis and enteric methane
emissions. It also traces advances in technologies that have
enabledus tounderstand theunderlyingbiologicalmechan-
isms involved in the modulation of the rumen microbiome.
The rumen microbial communityThe rumen contains a dense microbial community that actively
degrades plant material, providing the animal with energy via the
end-products of fermentation (short chain fatty acids) and protein in
the form of microbial protein, which flows from the rumen into the
lower intestine1. Rumen microbes not only adhere to and degrade
plant material they may also utilise substrates produced by other
microbes. The rumenmicrobial population includes bacteria that are
predominantly strict anaerobes with the capacity to be highly fibro-
lytic and proteolytic (generally of the phyla Bacteroidetes, Firmicutes
andProteobacteria),methanogenic archaea (phylumEuryarchaeota)
and anaerobic fungi (fungal division Neocallimastigomycota)2,3.
These rumenmicrobes are predated on by populations of anaerobic
protozoa (predominantly of the phylum Ciliophora)4,5 and viruses
(predominantly dsDNA bacteriophages of the order Caudovirales)6.
Rumen microbes friend or foe? Strategies
for reducing plant toxicity, acidosis
and enteric methaneThe modulation of rumen microbial populations has traditionally
focussedon strategies to improve feeddigestibility and consequent-
ly increase overall animal productivity, reducing the time taken for
ruminant livestock to reach market-weight specifications. Micro-
biologists and animal nutritionists have sought to determine the
impact of different diet formulations on rumen function and live-
weight gains, investigating the effects of feedstuff pre-treatment
employing either physical change (for example steam treatment,
rolling or flaking of grain7–9) or physical and chemical changes
through microbial and enzymatic pre-treatment (for example ensi-
lage of fodder crops with or without the application of silage
inoculants10–12). Research has also sought to increase the environ-
mental sustainability of livestock production by investigating the
Under theMicroscrope
18 10.1071/MA15006 MICROBIOLOGY AUSTRALIA * MARCH 2015

viability of alternative feedstuffs such as those that may also be a by-
product of food production or industry (for example cotton seed
meal13) or feeds that may be readily propagated on-farm (for
example microalgae14,15). Strategies have also been developed to
allow cattle to utilise plant feedstuffs that may otherwise be toxic to
the animal, for example the leguminous shrub Leucaena leucoce-
phala may be propagated on-farm as a high-protein fodder crop
(Figure 1). Leucaena however, contains the toxic amino acid
mimosine and normal rumen microbial degradation results in the
formation of the toxin 3-hydroxy-4(1H)-pyridone (DHP). The ac-
cumulation of these toxins leads to negative health implications
includinghair loss, reduced live-weight gain andgoitre. The solution
to preventing the development of Leucaena toxicity arose from
rumen microbes, as these toxins were first shown to be de-toxified
bybacteria of the genera Synergistes isolated from the rumenof feral
goats16. A mixed microbial drench containing Synergistes jonesii is
currentlyproduced inQueensland for the treatmentof cattlegrazing
Leucaena17.
In addition to strategies to improve feed breakdown in the rumen,
research has also been undertaken to specifically target and control
certain rumenmicrobial populations including the rumenprotozoa,
amylolytic bacteria, and more recently, the methanogenic archaea.
Rumen protozoa may positively contribute to ruminant feed break-
down18; however, as these eukaryotes actively graze on the rumen
bacterial populations, their growth and proliferation within the
rumenmay also contribute to the inefficient, intra-ruminal recycling
of microbial protein5. Strategies to reduce rumen protozoa have
included the use of diet (high grain diets tend to reduce protozoal
populations), dietary additives such as the clay bentonite, nitrates
and vaccination19,20. While these strategies have been shown to
impact onprotozoal populations, it hasprovendifficult to complete-
ly remove protozoa from the rumen.
Strategies have also been developed to control rumen populations
of the amylolytic bacterium Streptococcus bovis. Amylolytic, lactic
acid-producing bacteria such as S. bovis, may over-proliferate in the
rumen when cattle are fed high concentrate or high grain diets,
contributing to the development of a condition known as lactic
acidosis. S. bovis has been targeted through the application of
antibiotics such as monensin21 and phage therapies22. The strate-
gies developed to control S. boviswere largely undertaken between
the 1970s and1990s and the relative importanceof controlling these
organisms and their overall contribution to the development of
lactic acidosis has been cause for debate23,24. Feeding practices
avoiding sudden dietary changes to large quantities of fermentable
carbohydrates can prevent the development of acidosis25 and most
of the novel strategies to prevent rumen acidosis reported in the
literature26,27 havenot beeneither commercialised or implemented
within the livestock production industry.
In the past decade, investigations to modulate rumen microbial
populations have focused on strategies to reduce the amount of the
potent greenhouse gasmethanegeneratedby livestockproduction.
Normal rumen microbial fermentation results in the accumulation
of hydrogen. This hydrogen may be utilised by acetogenic bacteria
(for example of the genera Acetitomaculum, Eubacterium
and Blautia), however the majority of hydrogen is consumed by
populations of methanogenic archaea belonging to the genera
Methanobrevibacter,Methanobacterium,Methanococcus,Metha-
nomicrobium andMethanosaeta, with the methane produced lost
to the animal via eructation28. Strategies currently in development
to control rumen methanogen populations include specific diets
Figure 1. Cattle herd grazing the leguminous shrub Leucaena leucocephala propagated on-farm in Northern Australia.
Under theMicroscrope
MICROBIOLOGY AUSTRALIA * MARCH 2015 19

(high grain and/or oil), dietary additives (for example naturally-
occurring plant-derived compounds and synthetic anti-methano-
genic compounds), animal breeding, phage-based therapies and
vaccination29–32.While several of these novel approaches have been
shown to be effective in reducing rumenmethanogen populations,
the majority are not yet at the stage of commercial application and
adoption by the livestock production industry.
Feed supplements, probiotics and direct fed
microbialsThe Australian agricultural feed industry produces many feed sup-
plements designed and marketed to improve ruminant production
efficiency, particularly for the dairy industry. Australian law requires
that all agricultural and veterinary chemical products sold in
Australia be registered by the Australian Pesticides and Veterinary
MedicinesAuthority (APVMA,http://apvma.gov.au) andare listedon
the permits and PubCRIS database (https://portal.apvma.gov.au/
pubcris). These feed supplements include direct fed microbials
(DFM) or probiotics and may also incorporate additional enzymes
(amylases, proteases), minerals and salts (selenium, potassium).
There are approximately 30 formulations of probiotic microbes
available for use within Australia to enhance the overall digestive
efficiency of ruminants incorporating bacteria such as Bifidobacter-
ium (including the species bifidum, longum and thermophilum),
Lactobacillus (speciesacidophilus,delbrueckii subspecies bulgar-
icus, plantarum and rhamnosus) and Enterococcus faecium.
In addition, a further 11 registered products are available that are
exclusively basedon the yeast Saccharomyces cerevisiae.While the
survival and proliferation of these organisms in the rumen and the
effect of these probiotic bacteria on rumen digestive processes
has been largely under-represented in the scientific literature, the
probiotic effects of yeasts (S. cerevisiae) has beenmore extensively
assessed33,34. Results can be highly variable between studies, how-
ever several investigations have established that the provision of
these commercially available probiotics ismost useful when applied
to young ruminants to accelerate the establishment of a healthy
gastrointestinal microflora35,36. Probiotics may be used to exclude
undesirable zoonotic pathogens suchasEscherichia coliO157 from
establishing in the ruminant gastrointestinal tract and may also
impact on the ruminant host immune system and feed breakdown
efficiency36–38.
While bacterial strains of rumen origin would be anticipated to
survive and proliferate in the rumen and therefore have a selective
advantage overmicrobes of non-rumen origin36, there are currently
no commercial formulations of rumen-derived probiotic bacteria
registered for use in Australia. The mixed microbial drench for
Leucaena toxicity is the only APVMA approved rumen-derived
microbial treatment. There have however been several reports in
the scientific literature of rumen-derived bacterial isolates being
examined for application as potential probiotics, for example,
Megasphaera eldenii, Ruminococcus sp., R. flavefaciens, Prevo-
tella bryantii, Butyrivibrio fibrisolvens37,39. These strains were
selected for further testing in order to increase starch utilisation
(by the genera Ruminococcus, Prevotella and Butyrivibrio) and to
assist with the prevention of lactic acid accumulation contributing
to acidosis (M. elsdenii) in feedlot cattle.
Future directionsThe success of any strategies developed to modulate the microbial
population of the rumen will always need to be underpinned by
fundamental research efforts to understand how the strategies
impact on the baseline or ‘normal’ functioning of the rumen
microbial ecosystem40. Addressing the gaps in current understand-
ing of the rumen microbial ecosystem41 is therefore key to the
developmentof newstrategies to control themicrobial populations.
The rumen contains a very dense microbial ecosystem, end-
products of microbial digestion, salts and plant material including
partially digested fibre, carbohydrate, and phenolic compounds
and as such, samples of rumen material often present unique
technical challenges. Rumen microbiologists have therefore always
been quick to adopt new developments in technology in order to
more fully understand the complex microbial ecosystem of the
rumen. Early research efforts relied on microbial cultivation and
although the study of cultivated rumen microbes is important for
characterising microbial genera and elucidating their specific
genetic and metabolic traits, culture-independent studies for the
detection of specific microbes (real-time PCR assays) and commu-
nity analysis based on the comparative phylogeny of the prokaryote
16S rRNA gene are often employed to ascertain the extent to which
probiotic microbes survive and proliferate within the rumen31.
Rapid advances in high-throughput sequencing technologies have
also facilitated investigations into how probiotics can influence
both the community composition and functional gene capacity of
the rumen. Metagenomic studies of the rumen have progressed
current understandingof the functional genepotential of the rumen
microbial populationenabling the in silico identificationofenzymes
involved in feed breakdown42.
In the future, a greater reliance on gene sequence-based technol-
ogies or ‘omics’ will lead to an increased understanding of the
interactions occurring between probiotics and the microbial popu-
lations indigenous to the ruminant gastrointestinal tract. This is of
particular interest for the development and optimisation of new
and more effective strategies for the modulation of the rumen
microbiome. Development of new strategies, treatments and pro-
biotics to enhance rumen feed utilisation efficiency, represents an
area of great potential for the Australian livestock industries andwill
enable the production of quality products to meet global demands.
References1. Bryant, M.P. (1959) Bacterial species in the rumen. Bacteriol. Rev. 23, 125–153.
2. Hungate, R.E. et al. (1964) The rumen bacteria and protozoa. Annu. Rev.
Microbiol. 18, 131–166. doi:10.1146/annurev.mi.18.100164.001023
3. Orpin, C.G. (1975) Studies on the rumen flagellate Neocallimastix frontalis.
J. Gen. Microbiol. 91, 249–262. doi:10.1099/00221287-91-2-249
Under theMicroscrope
20 MICROBIOLOGY AUSTRALIA * MARCH 2015

4. Moon-van der Staay, S.Y. et al. (2014) The symbiotic intestinal ciliates and the
evolution of their hosts. Eur. J. Protistol. 50, 166–173. doi:10.1016/j.ejop.2014.
01.004
5. Williams, A.G. (1986) Rumenholotrich ciliate protozoa.Microbiol. Rev.50, 25–49.
6. Klieve, A.V. and Bauchop, T. (1988) Morphological diversity or ruminal bacter-
iophages from sheep and cattle. Appl. Environ. Microbiol. 54, 1637–1641.
7. Owens, F.N. et al. (1997) The effect of grain source and grain processing on
performance of feedlot cattle: A review. J. Anim. Sci. 75, 868–879.
8. Yang,W.Z. et al. (2001) Effects of grainprocessing, forage to concentrate ratio, and
forage particle size on rumen pH and digestion by dairy cows. J. Dairy Sci. 84,
2203–2216. doi:10.3168/jds.S0022-0302(01)74667-X
9. Yang, W.Z. et al. (2013) Quality and precision processing of barley grain affected
intake and digestibility of dry matter in feedlot steers. Can. J. Anim. Sci. 93,
251–260. doi:10.4141/cjas2012-132
10. Beauchemin, K.A. et al. (2003) Effects of particle size of alfalfa-based dairy cow
diets on chewing activity, ruminal fermentation, andmilk production. J. Dairy Sci.
86, 630–643. doi:10.3168/jds.S0022-0302(03)73641-8
11. Koenig, K. and Beauchemin, K. (2011)Optimum extent of barley grain processing
and barley silage proportion in feedlot cattle diets: growth, feed efficiency, and
fecal characteristics. Can. J. Anim. Sci. 91, 411–422. doi:10.4141/cjas2010-039
12. Zhao, Y.L. et al. (2015) Effects of volume weight, processing method and proces-
sing index of barley grain on in situ digestibility of dry matter and starch in beef
heifers. Anim. Feed Sci. Technol. 199, 93–103. doi:10.1016/j.anifeedsci.2014.
11.005
13. Rogers, G.M. et al. (2002) Feeding cotton products to cattle. Vet. Clin. North Am.
Food Anim. Pract. 18, 267–294. doi:10.1016/S0749-0720(02)00020-8
14. Holman, B.W.B. and Malau-Aduli, A.E.O. (2013) Spirulina as a livestock supple-
ment and animal feed. J. Anim. Physiol. Anim. Nutr. (Berl.) 97, 615–623.
doi:10.1111/j.1439-0396.2012.01328.x
15. Panjaitan, T. et al. (2010) Effect of the concentration of Spirulina (Spirulina
platensis) algae in the drinking water on water intake by cattle and the proportion
of algae bypassing the rumen.Anim.Prod. Sci.50, 405–409. doi:10.1071/AN09194
16. Allison,M.J. et al. (1992) Synergistes jonesii, gen. nov., sp. nov.: a rumenbacterium
that degrades toxic pyridinediols. Syst. Appl. Microbiol. 15, 522–529. doi:10.1016/
S0723-2020(11)80111-6
17. Klieve, A.V. et al. (2002) The production and storage of a fermentor-grown
bacterial culture containing Synergistes jonesii, for protecting cattle against
mimosine and 3-hydroxy-4(1H)-pyridone toxicity from feeding on Leucaena
leucocephala. Aust. J. Agric. Res. 53, 1–5. doi:10.1071/AR00121
18. Takenaka, A. et al. (2004) Fiber digestion by rumen ciliate protozoa. Microbes
Environ. 19, 203–210. doi:10.1264/jsme2.19.203
19. Hristov, A.N. et al. (2003) Evaluation of several potential bioactive agents for
reducing protozoal activity in vitro. Anim. Feed Sci. Technol. 105, 163–184.
doi:10.1016/S0377-8401(03)00060-9
20. Williams, Y.J. et al. (2014) Technical note: Protozoa-specific antibodies raised in
sheep plasma bind to their target protozoa in the rumen. J. Anim. Sci. 92,
5757–5761. doi:10.2527/jas.2014-7873
21. Newbold, C.J. and Wallace, R.J. (1988) Effects of the ionophores monensin and
tetronasin on simulated development of ruminal lactic acidosis in vitro. Appl.
Environ. Microbiol. 54, 2981–2985.
22. Tarakanov, B.V. (1994) Regulation of microbial processes in the rumen by
bacteriophages of Streptococcus bovis. Microbiol. 63, 373–378. (translated from
Mikrobiologiya 363, 657–667).
23. Calsamiglia, S. et al. (2012) Is subacute ruminal acidosis a pH related problem?
Causes and tools for its control.Anim. Feed Sci. Technol.172, 42–50. doi:10.1016/
j.anifeedsci.2011.12.007
24. Newbold, C.J. and Hillman, K. (2004) Feed Supplements: Enzymes, probiotics,
yeasts. In Encyclopedia of Animal Science (Pond, W.G., ed), pp. 376–378, CRC
Press.
25. Krause, K.M. and Oetzel, G.R. (2006) Understanding and preventing subacute
ruminal acidosis in dairy herds: a review. Anim. Feed Sci. Technol. 126, 215–236.
doi:10.1016/j.anifeedsci.2005.08.004
26. Klieve, A.V. et al. (2012) Persistence of orally administeredMegasphaera elsdenii
and Ruminococcus bromii in the rumen of beef cattle fed a high grain (barley)
diet. Anim. Prod. Sci. 52, 297–304. doi:10.1071/AN11111
27. Styriak, I. et al. (1994) Isolation and characterisation of a new ruminal bacterio-
phage lytic to Streptococcus bovis. Curr. Microbiol. 28, 355–358. doi:10.1007/
BF01570201
28. St-Pierre, B. and Wright, A.D.G. (2013) Diversity of gut methanogens in herbiv-
orous animals. Animal 7, 49–56. doi:10.1017/S1751731112000912
29. Buddle, B.M. et al. (2011) Strategies to reduce methane emissions from
farmed ruminants grazing on pasture. Vet. J. 188, 11–17. doi:10.1016/j.tvjl.2010.
02.019
30. Cottle, D.J. et al. (2011) Ruminant enteric methane mitigation: a review. Anim.
Prod. Sci. 51, 491–514. doi:10.1071/AN10163
31. Krause, D.O. et al. (2013) Board-invited review: rumen microbiology: leading
the way inmicrobial ecology. J. Anim. Sci. 91, 331–341. doi:10.2527/jas.2012-5567
32. Williams, Y.J. et al. (2009) A vaccine against rumen methanogens can alter the
composition of archaeal populations. Appl. Environ. Microbiol. 75, 1860–1866.
doi:10.1128/AEM.02453-08
33. Callaway, T. et al. (2012)Current status of practical applications: probiotics in dairy
cattle. In Direct-Fed Microbials and Prebiotics for Animals (Callaway, T.R. and
Ricke, S.C., eds), pp. 121–135, Springer, New York.
34. Pinloche, E. et al. (2013) The effects of a probiotic yeast on the bacterial diversity
and population structure in the rumen of cattle. PLoSONE8, e67824. doi:10.1371/
journal.pone.0067824
35. AlZahal, O. et al. (2014) Use of a direct-fed microbial product as a supplement
during the transition period in dairy cattle. J. Dairy Sci. 97, 7102–7114.
doi:10.3168/jds.2014-8248
36. Yeoman, C.J. and White, B.A. (2014) Gastrointestinal tract microbiota and pro-
biotics in production animals. Ann. Rev. Anim. Biosci. 2, 469–486. doi:10.1146/
annurev-animal-022513-114149
37. McAllister, T.A. et al. (2011) Review: the use of direct fed microbials to mitigate
pathogens and enhance production in cattle. Can. J. Anim. Sci. 91, 193–211.
doi:10.4141/cjas10047
38. Newbold, C.J. and Hillman, K. (2004) Feed Supplements: enzymes, probiotics,
yeasts. In Encyclopedia of Animal Science (Pond, W.G., ed), pp. 376–378, CRC
Press.
39. Klieve, A.V. et al. (2003) Establishing populations ofMegasphaera elsdenii YE 34
and Butyrivibrio fibrisolvens YE 44 in the rumen of cattle fed high grain diets.
J. Appl. Microbiol. 95, 621–630. doi:10.1046/j.1365-2672.2003.02024.x
40. Russell, J.B. and Rychlik, J.L. (2001) Factors that alter rumen microbial ecology.
Science 292, 1119–1122. doi:10.1126/science.1058830
41. Lima, F.S. et al. (2015) Prepartum and postpartum rumen fluid microbiomes:
characterization and correlation with production traits in dairy cows. Appl.
Environ. Microbiol. 81, 1327–1337. doi:10.1128/AEM.03138-14
42. Hess, M. et al. (2011) Metagenomic discovery of biomass-degrading genes
and genomes from cow rumen. Science 331, 463–467. doi:10.1126/science.
1200387
BiographiesDr Rosalind Gilbert is a scientist with the Queensland
Department of Agriculture, Fisheries and Forestry, Rumen Ecology
Unit. Her research interests include rumen microbiology and the
role of phages in controlling rumen microbial populations.
Ms Diane Ouwerkerk is a Senior Molecular Biologist within the
Rumen Ecology Unit of the Department of Agriculture, Fisheries
and Forestry based at the Ecosciences Precinct, Dutton Park,
Queensland. Her research interests include the use of molecular
techniques to investigate gut microbial ecosystems, particularly in
ruminants.
Athol Klieve is the Associate Professor in Agricultural Microbiology
at the University of Queensland. He has worked in rumen micro-
biology for 30 years and leads the collaborative UQ/DAFF Rumen
Ecology Unit.
Under theMicroscrope
MICROBIOLOGY AUSTRALIA * MARCH 2015 21

Polymicrobial nature of chronic oral disease
Stuart DashperA, Helen MitchellA, Geoff AdamsA and Eric ReynoldsA,B
AOral Health Cooperative Research Centre, Melbourne Dental School, The University of Melbourne, Parkville, Vic. 3052, Australia
BCorresponding author. Tel: +61 3 9341 1547, Fax: +61 3 9341 1596, Email: [email protected]
Recentmicrobiome studies have shown that the human oral
microbiome is composed of over 260 abundant bacterial
species that predominantly live as polymicrobial biofilms
accreted to the non-shedding hard surfaces of the teeth. In
addition representatives of both Archaea and Fungi are
found in the oral cavity and there is considerable colonisa-
tionof the soft tissues of themouth.Most of these species are
commensal and form complex biofilm communities that
restrict the colonisation of the oral cavity by exogenous
bacteria. Changes in the polymicrobial biofilm microenvi-
ronment such as those resulting from the effects of chronic
inflammation for subgingival plaque, can lead to the emer-
gence of opportunistic pathogens resulting in dysbiosis and
the development of chronic diseases such as periodontitis in
a susceptiblehost. Theapplicationofmicrobiomic studies to
the analysis of these complex and dynamic communities in
rigorously designed human clinical studies will provide
valuable mechanistic insight into the bacterial succession
and complex interactions involved in the development of
dysbiosis and disease.
The human oral cavity is the entry point of the gastrointestinal tract
and offers a number of microenvironments that enable the prolif-
eration of a wide range of largely commensal bacteria, the vast
majority of which are endemic to the human oral cavity. Consider-
able effort has been expended to identify the approximately 700
prokaryote species that compose the total human oral microbiome.
Over one-third of these species remain uncultivated and less than
half are officially named; however, draft genomes for approximately
half of these taxa are now available from the Human Oral Microbial
Database (www.homd.org/)1.
16S rRNA gene sequence surveys are providing a cost effective
means of studying microbiomes, identifying and enumerating a
relatively unbiased set of the prokaryotic species present, including
uncultivable species. This technique has been adopted for studying
the oral microbiota; however, the results produced have not yet
been definitive, with some studies finding huge variation across
individuals and limited or no differences between healthy and
diseased states. Many factors of the design and analysis of these
experiments can make it difficult to compare results between the
different studies, including pooling of samples, DNA extraction
method, marker gene or region used, primer sets, PCR conditions,
sequencing platform, choice of taxonomic classifier and level of
classification, clustering of read data into microbial groups, and the
statistical methods used for diversity analysis2. The composition of
the healthy oral microbiota can certainly vary considerably across
sites within the mouth, at the same site over time, and from person
to person3,4. Although the 16S rRNA survey techniques are available
most clinical studies to date have been cross-sectional and have
investigated a limited number of bacterial species using either real
time PCR, checkerboard DNA-DNA hybridisation, or more recently,
the Human Oral Microbe Identification Microarray (HOMIM)
techniques. To compound these limitations samples taken from a
limited number of sites within the mouth are often pooled which
can obscure comparative results and is not recommended for
Under theMicroscope
22 10.1071/MA15007 MICROBIOLOGY AUSTRALIA * MARCH 2015

diverse communities such as the oral microbiome. Generating
less information from a larger number of samples is much more
informative than generatingmore information from a small number
of samples, particularly for the classification of diseased and healthy
states5. Furthermore, clinical sampling techniquesmay result inonly
part of the polymicrobial biofilm being collected; with the surface
of the biofilm most closely associated with the host and disease
process poorly represented in the sample.
Chronic periodontitis is an inflammatory disease of the supporting
tissues of the teeth with an endogenous polymicrobial aetiology.
Over 47% of Americans over 30 (64.7million adults) have chronic
periodontitis, distributed as 8.7% mild, 30.0% moderate and 8.5%
severe6. The prevalence and severity of periodontitis increases with
age, with more than 64% of adults aged 65 and over likely to have
moderate or severe periodontitis. Numerous cross-sectional and
longitudinal epidemiological studies have shown associations be-
tween periodontal diseases and a greater risk of certain systemic
diseases and disorders, such as cardiovascular diseases, diabetes,
chronic kidney disease, metabolic syndrome, obesity, rheumatoid
arthritis, Alzheimer’s disease, pre-term andunderweight births, and
some cancers, particularly pancreatic, head and neck and oesopha-
geal cancers. These associations remain even after adjustment for
medical and socio-economic confounding factors.
The concepts of the roles of particular oral bacterial species in
chronic periodontitis have changed over the past two decades but
there is wide consensus that anaerobic, proteolytic, amino acid
fermenting species including Porphyromonas gingivalis, Trepone-
ma denticola and Tannerella forsythia play a crucial role in
initiation and/or progression of disease. A process of bacterial
succession in subgingival plaque has been described where mild
inflammationof thegingival tissue (gingivitis) and theestablishment
of an appropriate microenvironment by early colonising bacteria
allows late colonisers to emerge as opportunistic pathogens7,8.
More recently P. gingivalis has been proposed to be a keystone
pathogen that perturbs the ecological balance enabling the prolif-
eration of other oral bacterial species resulting in the formation of a
dysbiotic polymicrobial plaque, whilst remaining at very low levels
itself9. The keystone pathogen concept was adapted from conser-
vation biology’s keystone species defining a low abundance species
which has a disproportionately large effect on its environment.
While this theory does help explain the major role of a few species
in a complex polymicrobial biofilm, it is not entirely consistent
with prospective clinical trial data demonstrating that this
bacterium, amongst others, proliferates during disease and can
represent a significant proportion of subgingival plaque bacteria at
diseased sites. Furthermore the imminent progression of chronic
periodontitis at a site has been predicted by increases in the relative
proportions of P. gingivalis and/or T. denticola in subgingival
plaque at that site above threshold levels of 10–15%10. This and
more recent research demonstrating mutualism and synergistic
virulence in animal models of periodontitis of multiple late colonis-
ing species found closely associated with disease progression in
humans has led to the more generally supported view of the
emergence of an opportunistic pathogenic polymicrobial biofilm
as the trigger for disease progression in susceptible individuals
(Figure 1)11. A large number of disease-associated species have
been identified, consistent with the dysbiosis-hypothesis12, al-
though further studies are required to differentiate commensal
species that benefit from the disease process as opposed to those
that actually cause the disease.
In addition to the composition of the subgingival plaque polymi-
crobial biofilm, which will be revealed by microbiomic analyses, its
architecture is important as it has been shown that the late colonis-
ing opportunistic periodontal pathogens are found as microcolo-
nies in the outer layer of the biofilm adjacent to the epitheliumof an
inflamed periodontal pocket13. These findings indicate that the
disproportionately large impact of the late colonising opportunistic
pathogens in disease may be explained by their close proximity to
the inflamed tissue and resorbing alveolar bone.
Chronic periodontitis is episodic in nature with acute exacerbations
of destruction followed by periods of dormancy. Currently, diag-
nosis of periodontitis is achieved retrospectively by clinical assess-
ment of attachment loss. This loss of attachment is a result of
pathogenic events that have already occurred at the diseased site,
and any sampling at the time of diagnosis, may fail to identify those
species involved in active destruction. In addition different teeth
within the same patient, as well as different sites around the same
Dysbiosis
Homeostasis
Stable
Oral Healthcrc
Commensal Biofilm Reduced Inflammation
Inflammation
Progressive
Chronic Periodontitis
Pathogenic Biofilm
Figure 1. The changes in the subgingival plaque microbiome and theintimate association with the host inflammatory response that results ina shift from a stable site to one that is undergoing disease progression.
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2015 23

tooth can display varying degrees of disease severity, all undergoing
periodontal disease progression at different rates. Therefore to
conclusively determine the polymicrobial aetiology of chronic peri-
odontitis and how opportunistic pathogens emerge and proliferate
rigorously designed prospective human clinical trials coupled with
microbiomic analyses are essential, followed by the testing of the
bacterial species and communities identified in appropriate in vitro
and animal models to determine their potential as polymicrobial
biofilms to induce dysbiosis and disease.
References1. Dewhirst, F.E. et al. (2010) The human oral microbiome. J. Bacteriol. 192,
5002–5017. doi:10.1128/JB.00542-10
2. Goodrich, J.K. et al. (2014) Conducting a microbiome study. Cell 158, 250–262.
doi:10.1016/j.cell.2014.06.037
3. Simón-Soro, Á. et al. (2013) Microbial geography of the oral cavity. J. Dent. Res.
92, 616–621. doi:10.1177/0022034513488119
4. Ge, X. et al. (2013) Oral microbiome of deep and shallow dental pockets in
chronic periodontitis. PLoS ONE 8, e65520. doi:10.1371/journal.pone.0065520
5. Hamady, M. and Knight, R. (2009) Microbial community profiling for human
microbiome projects: tools, techniques, and challenges. Genome Res. 19,
1141–1152. doi:10.1101/gr.085464.108
6. Eke, P.I. et al. (2012) Prevalence of periodontitis in adults in the United States:
2009 and 2010. J. Dent. Res. 91, 914–920. doi:10.1177/0022034512457373
7. Socransky, S.S. et al. (1998) Microbial complexes in subgingival plaque. J. Clin.
Periodontol. 25, 134–144. doi:10.1111/j.1600-051X.1998.tb02419.x
8. Kolenbrander, P.E. et al. (2010) Oral multispecies biofilm development and the
key role of cell–cell distance. Nat. Rev. Microbiol. 8, 471–480. doi:10.1038/
nrmicro2381
9. Hajishengallis, G. et al. (2011) Low-abundance biofilm species orchestrates
inflammatory periodontal disease through the commensal microbiota and com-
plement. Cell Host Microbe 10, 497–506. doi:10.1016/j.chom.2011.10.006
10. Byrne, S.J. et al. (2009) Progression of chronic periodontitis can be predicted
by the levels of Porphyromonas gingivalis and Treponema denticola in sub-
gingival plaque.OralMicrobiol. Immunol.24, 469–477. doi:10.1111/j.1399-302X.
2009.00544.x
11. Tan, K.H. et al. (2014) Porphyromonas gingivalis and Treponema denticola
exhibit metabolic symbioses. PLoS Pathog. 10, e1003955. doi:10.1371/journal.
ppat.1003955
12. Curtis, M.A. (2014) Periodontal microbiology—The lid’s off the box again. J. Dent.
Res. 93, 840–842. doi:10.1177/0022034514542469
13. Zijnge, V. et al. (2010) Oral biofilm architecture on natural teeth. PLoS ONE 5,
e9321. doi:10.1371/journal.pone.0009321
Biographies
Stuart Dashper is a Professor in the Oral Health Cooperative
Research Centre and TheMelbourne Dental School, The University
of Melbourne. Over the past 15 years he has developed a systems
biology approach to the study of chronic oral diseases that incor-
porates the identification and characterisation of bacterial patho-
bionts, the composition and structures of the polymicrobial biofilm
communities in which they dwell, themolecular characterisation of
virulence-related traits and their interactionswithothermembers of
the bacterial community and the host.
Helen Mitchell is a researcher with the Oral Health CRC and
Masters of Science student in Bioinformatics at The University of
Melbourne. She is undertaking comparative genomics of the peri-
odontal pathobiont Porphyromonas gingivalis to determine viru-
lence characteristics and assist in vaccine development. She is using
Next-Generation Sequencing techniques to determine bacterial
biomarkers of early childhood caries in saliva.
Geoff Adams has over 30 years’ experience as a biostatistician
and epidemiologist involved in consulting, teaching, and research.
He has been employed by the Melbourne Dental School and the
Oral Health CRC as a biostatistician and epidemiologist since 1999.
Geoff manages the Oral and Systemic Disease program in the
Oral Health CRC, which is investigating associations between peri-
odontal disease and various systemic conditions.
Eric Reynolds AO PhD FICD FTSE FRACDS is a Melbourne
Laureate Professor and CEO and Director of Research of the Oral
Health CRC. He is also Head of the Oral Biology Section of the
Melbourne Dental School. He has been researching and teaching
for over 30 years on the aetiology and prevention of the two major
oral diseases, dental caries and periodontal diseases, which are
associated with polymicrobial biofilms.
Under theMicroscope
24 MICROBIOLOGY AUSTRALIA * MARCH 2015

Gastrointestinal microbiota, diet and brainfunctioning
Shakuntla Gondalia
Centre for HumanPsychopharmacology, School ofHealth Science, SwinburneUniversity of Technology, Hawthorn,Vic. 3122, AustraliaTel: +61 3 9214 5100Email: [email protected]
Andrew Scholey
Centre for HumanPsychopharmacology, School ofHealth Science, SwinburneUniversity of Technology, Hawthorn,Vic. 3122, Australia
A growing interest for research in the relationship between
the gastrointestine (GI), GImicrobiota, health and disease is
due to the potential for research identifying intervention
strategies.Preclinical andclinical studieshave indicated that
initial colonisation of bacteria in the GI tract can affect the
individual’s health condition in later life. Diet is an influen-
tial factor in modulating this complex ecosystem and con-
sequently can help to modulate physiological conditions.
The broader role of the GI microbiota in modulation of
pathology and physiology of various diseases has pointed
to the importance of bidirectional communication between
thebrain and theGImicrobiota inmaintaininghomeostasis.
An association of diet withmetabolic diseases is well known
andtherearedietary supplements reported to improvebrain
function and cognitive decline. In addition to the plausible
mechanisms of inflammation and oxidative stress for psy-
chological conditions, more research into the role of the GI
microbiota in combination with dietary factors as a compo-
nent in psychological condition is warranted. From this
work, targeted interventions could result.
Although a link between GI function and health or disease in an
individual hasbeendetermined, theunderlyingmechanismsarenot
clearly understood. TheGI tract of a newborn is rapidly colonised by
microbiota during the birth process through maternal contact and
from the surrounding environment. This microbial ecosystem sta-
bilises in first 2–3 years of life and reaches maturity in the human
adult1. By adulthood, the intestine contains approximately 1012
bacteria per gram of colonic content2, which is 10 times the number
of cells in the human body. Based on 16S rRNA gene analyses, it was
estimated that an adult GI tract harbours between 500 to 1000
different species3 from three major bacterial phyla, Bacteroidetes
(Gram-negative), Firmicutes (Gram-positive) and Actinobacteria
(Gram-positive). The proportion of these phyla in any individual
depends upon that individual’s genetic makeup, dietary habits and
surrounding environment.
The initial inoculation then colonisation of the GI impacts the
GI microbiota throughout life and the dynamic microbial
ecosystem is highly influenced by the surrounding environment
and dietary factors. Any modifications in this highly organised and
complex ecosystem have the potential to influence the normal
physiological functions and are suspected to play a role in obesity,
fatty liver disease, inflammation, diabetes and also psychological
conditions4–8.
Diet that impacts GI microbiota: from infancy
to elderly
Upon ageing, weakening of dentition, salivary function, digestion
and intestinal transit time may affect the intestinal microbiota9,10.
One manageable environmental factor is diet, which has been
shown to significantly impact the GI microbial composition. The
types of bacteria present are dependent on the type of substrates
available, with some bacteria prospering on a specific substrate
while others are unable to utilise that compound11. This selective
substrate usage therefore enablesmodulationof the compositionof
GI microbiota via the diet, which can occur at any stage of life.
However, diet in infancy can greatly affect thematuration process of
the microbial profile, gastrointestinal health and immune system.
Human milk is normally the first dietary exposure to an infant. This
is fermented in the colon, stimulating the growth of specific bacteria
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2015 10.1071/MA15008 25

(including Gram positive Bifidobacterium spp.). Infants fed with
breast milk and those on milk formula developed and harboured
different and diverse bacterial populations in their GI tract12. More-
over, the composition of different milk formula facilitates different
types of bacterial colonisation. Infants fed with oligosaccharide
enriched formula harbour higher counts of Bifidobacterium
spp. and Lactobacillus spp. compared to infants fed on unsupple-
mented formulae13. This prebiotic effect of infant food could be of a
major concern as it plays a role in shaping the GI microbiota that is
believed to influence an individual’s health for their entire life. A
significant shift in the GI microbial ecology occurs when infants
switch to a more solid and varied diet, including substantial reduc-
tion in the percentage of Bifidobacterium spp. and Lactobacillus
spp. in the totalmicrobiota14. Throughout adulthoodGImicrobiota
appears to becomemore stable15 as withmore stable dietary habits.
The final shift in composition and function of gutmicrobiota occurs
during theolder age of lifespan. In general, aging is associatedwith a
decline in physiological function including in the immune system
and metabolism that consequently affects the microbial composi-
tion, or vice versa. Human age-related changes reported in the GI
microbial composition include a numerical decrease in Bifidobac-
teruim spp. and Lactobacillus spp. and an increase in Enterobac-
teriaceae and obligate anaerobes such as Clostridium spp.16
Numerous dietary components have been identified as having
positive or negative effect on brain function and behaviour, this
effect can be improved byGImicrobiota. Animal studies andhuman
clinical trials have well demonstrated the role of omega-3 polyun-
saturated fatty acid in normal brain functioning. Administration
of Bifidobacterium spp. in combination with a substrate for eico-
sapentaenoic acid (EPA), alpha-linolenic acid results in increased
concentration of EPA in liver tissue and docosahexaenoic acid
in brain tissues17,18. In wild mice model, probiotics such as
L. helveticus prevented high fat diet-induced anxiety-like behav-
iour19. Diet can modulate the GI microbial composition by provid-
ing a favourable environment while the contrary is also true that
GI microbiota can modulate the effect of diet and in turn host
physiological and psychological function.
Activity of GI microbiota
The GI microbiota can produce a vast range of metabolites and/or
structural components whose generation depends on the availabil-
ity of nutrients and the luminal environment. Thesemetabolites are
subsequently taken up by GI tissues, potentially reach circulation
and other distant tissues and can be excreted in urine and breath. GI
microbiotahavebeen linked tovery importanthealth functions such
as development and role of the immune system7, resistance to
infection by preventing pathogen colonisation20, bioactivation of
beneficial constituents such as polyphenols21, detoxification of
xenobiotics and metabolism of luminal components leading to
formation of a variety of metabolites such as short chain fatty acids
(SCFA), vitamins and several gases11,22,23.
SCFAs such as n-butyrate, acetate and propionate act as key sources
of energy for tissues andpromote cellularmechanisms thatmaintain
tissue integrity. When SCFAs reach the circulatory system, they
impact immune function and inflammation8. SCFAs are also in-
volved in host-microbe signalling and control of colonic pH with
subsequent effects onmicrobial composition, intestinalmotility and
epithelial cell proliferation24. Microbes have also been involved in
enzymatic degradation of complex substrates particularly many
forms of polysaccharides from ingested food25. For example Bac-
teroides thetaiotamicron, produces an array of enzymes for carbo-
hydrate breakdown26. During metabolism some gases such as
methane, hydrogen, hydrogen sulfide and carbon dioxide are
produced within the GI tract. Excess production of these gases may
causeGI problems such as bloating and pain. These gasesmay serve
useful purposes however, it is debatable whether hydrogen sulfide
for example, is largely beneficial or harmful!27.
Furthermore, GI microbiota can influence behaviour and brain
function by influencing the expression of certain body chemicals
such as hormons, neurotransmitters and neurotrophic factors.
Commensal bacteria such as Bifidobacteria infantis can modulate
tryptophan metabolism, this suggest that GI microbiota may influ-
ence the precursor pool for serotonin (5-HT)28. Animal studies have
demonstrated thatGImicrobioa can alsomodulates brain chemistry
such as Brain-DerivedNeurotropic Factor (BDNF) expression in the
hypothalamus and the brainstem29. BDNF is crucially involved in
neurogenesis, brain development and neural circuit formation.
BDNF has also been recognised as an important antiobesity factor.
GI microbiota maintain communication with the host using meta-
bolic, neural, immune and endocrine pathways. For health, homeo-
stasis between the GI microbiota and the host system is essential,
since any imbalance in this arrangement may result in a disease
condition4,5,7.
Interaction between GI microbiota and brain
functioning
The evidence is increasing for a bidirectional route of communica-
tion between brain, gut and GI microbiota which use immune,
neural and endocrine pathways and by this means influence gut-
brain communication, brain function and even humanbehaviour4,5.
The top-down mechanism of the effect of stress and psychological
condition on the GI functions is known from extensive research, in
Under theMicroscope
26 MICROBIOLOGY AUSTRALIA * MARCH 2015

particular with Inflammatory Bowel Disease, Irritable Bowel Syn-
drome and Crohn’s disease. However, the role of GI microbiota in
brain functions such as stress, cognition and mood need to be
explored more comprehensively.
The GI microbiota influences the release of the major neurotrans-
mitters tryptophan, serotonin, endocannabinoid ligands, ghrelin
and cholecystokinin, which can influence food intake, energy
balance and some brain tasks such as emotion, cognition andmotor
functions28,30. Changes in microbial composition and metabolism
correlate with the concept of ‘inflamm-ageing’, a low-grade chronic
pro-inflammatory status as a common basis for a broad spectrum of
age associated pathologies including cognitive decline and immu-
nosenescence31. Age-associated changes may increase intestinal
permeability and ease the passage of bacterial lipopolysaccharides
(LPS) into circulation, resulting in an elevated systemic LPS level.
When LPS binds to pattern recognition receptors such as toll-like
receptor 4 on immune cells, there is induction of inflammation by
production and release of cytokines, leukotrienes and prostaglan-
din32. Animal studieswithprobiotic supplementationdemonstrated
that probiotics can normalise immune responses, reverse beha-
vioural deficits and restore basal noradrenaline level in response to
stress6. The probiotics also normalise central nervous system (CNS)
biochemistry and improve behaviour in a mouse model of colitis,
through vagal nerve pathways for gut-brain communication33. Psy-
chological responses to the GI microbiome composition may be an
important factor in understanding the increasing prevalence of
psychological conditions in the community and research into this
topic should be promoted.
Conclusion
Clinically, a psychological conditiondoesnot standalone since there
are frequently immune system and GI comorbidities. Antidepres-
sants are the commonest treatment, and as such are focused on a
top-down approach. However, growing evidence from increasing
numbers of animal studies and human trials suggest that the GI
microbiota composition can be correlated with the incidence of
complex conditions such as cognitive decline, anxiety and depres-
sion. Detailed clinical interpretation is warranted so that novel
interventions in neuropsychological conditions can be employed.
Preclinical research combining detailed exploration of theGImicro-
biota in well-designed human cohort studies investigating the
impact of antibiotics, probiotics and diet on the brain and CNS
functions will inform us of the importance of bottom-up mechan-
isms in neuropsychological conditions. Further research in this
emerging area will provide novel targets for interventions in psy-
chological disorders.
References1. Scholtens, P.A.M.J. et al. (2012) The early settlers: intestinal microbiology in early
life.Annu.Rev. Food. Sci. Technol.3, 425–447.doi:10.1146/annurev-food-022811-
101120
2. Dominguez-Bello, M.G. et al. (2011) Development of the human gastrointestinal
microbiota and insights from high-throughput sequencing. Gastroenterology
140, 1713–1719. doi:10.1053/j.gastro.2011.02.011
3. Hooper, L.V. and Gordon, J.I. (2001) Commensal host-bacterial relationships in
the gut. Science 292, 1115–1118. doi:10.1126/science.1058709
4. Clemente, J.C. et al. (2012) The impact of the gut microbiota on human health:
an integrative view. Cell 148, 1258–1270. doi:10.1016/j.cell.2012.01.035
5. Heijtz, R.D. et al. (2011) Normal gut microbiota modulates brain development
and behavior. Proc. Natl. Acad. Sci. USA 108, 3047–3052. doi:10.1073/pnas.
1010529108
6. Desbonnet, L. et al. (2010) Effects of the probiotic Bifidobacterium infantis in
the maternal separation model of depression. Neuroscience 170, 1179–1188.
doi:10.1016/j.neuroscience.2010.08.005
7. Souza, D.G. et al. (2004) The essential role of the intestinal microbiota
in facilitating acute inflammatory responses. J. Immunol. 173, 4137–4146.
doi:10.4049/jimmunol.173.6.4137
8. Samuel, B.S. et al. (2008) Effects of the gut microbiota on host adiposity are
modulated by the short-chain fatty-acid binding G protein-coupled receptor,
Gpr41. Proc. Natl. Acad. Sci. USA 105, 16 767–16 772. doi:10.1073/pnas.08085
67105
9. Lovat, L.B. (1996) Age related changes in gut physiology and nutritional status.
Gut 38, 306–309. doi:10.1136/gut.38.3.306
10. Claesson, M.J. et al. (2012) Gut microbiota composition correlates with diet and
health in the elderly. Nature 488, 178–184. doi:10.1038/nature11319
11. Cummings, J.H. and Macfarlane, G.T. (1991) The control and consequences of
bacterial fermentation in the human colon. J. Appl. Bacteriol. 70, 443–459.
doi:10.1111/j.1365-2672.1991.tb02739.x
12. Harmsen,H.J.M. et al. (2000) Analysis of intestinal flora development in breast-fed
and formula-fed infants by using molecular identification and detection methods.
J. Pediatr. Gastroenterol. Nutr. 30, 61–67. doi:10.1097/00005176-200001000-
00019
13. Penders, J. et al. (2006) Factors influencing the composition of the
intestinal microbiota in early infancy. Pediatrics 118, 511–521. doi:10.1542/
peds.2005-2824
14. Sghir, A. et al. (2000)Quantificationof bacterial groupswithinhuman fecalflora by
oligonucleotide probe hybridization. Appl. Environ. Microbiol. 66, 2263–2266.
doi:10.1128/AEM.66.5.2263-2266.2000
15. Agans, R. et al. (2011) Distal gut microbiota of adolescent children is different
from that of adults. FEMS Microbiol. Ecol. 77, 404–412. doi:10.1111/j.1574-
6941.2011.01120.x
16. Lakshminarayanan, B. et al. (2013) Prevalence and characterization ofClostridium
perfringens from the faecal microbiota of elderly irish subjects. J. Med. Microbiol.
62, 457–466. doi:10.1099/jmm.0.052258-0
17. Wall, R. et al. (2012) Contrasting effects of Bifidobacterium breve NCIMB 702258
and Bifidobacterium breve DPC 6330 on the composition of murine brain fatty
acids and gut microbiota. Am. J. Clin. Nutr. 95, 1278–1287. doi:10.3945/ajcn.
111.026435
18. Wall, R. et al. (2010) Impact of administered Bifidobacterium onmurine host fatty
acid composition. Lipids 45, 429–436. doi:10.1007/s11745-010-3410-7
19. Ohland, C.L. et al. (2013) Effects of Lactobacillus helveticus on murine behavior
are dependent on diet and genotype and correlate with alterations in the
gut microbiome. Psychoneuroendocrinology 38, 1738–1747. doi:10.1016/
j.psyneuen.2013.02.008
20. Wells, C.L. et al. (1988) Role of intestinal anaerobic bacteria in colonization
resistance. Eur. J. Clin. Microbiol. Infect. Dis. 7, 107–113. doi:10.1007/
BF01962194
21. Miene, C. et al. (2011) Impact of polyphenol metabolites produced by colonic
microbiota on expression of COX-2 andGSTT2 in human colon cells (LT97).Nutr.
Cancer 63, 653–662. doi:10.1080/01635581.2011.552157
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2015 27

22. Donohoe, D.R. et al. (2011) The microbiome and butyrate regulate energy
metabolism and autophagy in the mammalian colon. Cell Metab. 13, 517–526.
doi:10.1016/j.cmet.2011.02.018
23. Topping, D.L. and Clifton, P.M. (2001) Short-chain fatty acids and human colonic
function: roles of resistant starch and nonstarch polysaccharides. Physiol. Rev.81,
1031–1064.
24. Musso, G. et al. (2011) Interactions between gut microbiota and host metabolism
predisposing to obesity and diabetes. Annu. Rev. Med. 62, 361–380. doi:10.1146/
annurev-med-012510-175505
25. Cantarel, B.L. et al. (2012)Complex carbohydrate utilization by thehealthy human
microbiome. PLoS ONE 7, e28742. doi:10.1371/journal.pone.0028742
26. Xu, J. et al. (2003) A genomic view of the human-Bacteroides thetaiotaomicron
symbiosis. Science 299, 2074–2076. doi:10.1126/science.1080029
27. Carbonero, F. et al. (2012) Microbial pathways in colonic sulfur metabolism
and links with health and disease. Front. Physiol. 3, 448. doi:10.3389/fphys.
2012.00448
28. Desbonnet, L. et al. (2008) The probiotic Bifidobacteria infantis: an assessment
of potential antidepressant properties in the rat. J. Psychiatr. Res. 43, 164–174.
doi:10.1016/j.jpsychires.2008.03.009
29. Schéle, E. et al. (2013) The gut microbiota reduces leptin sensitivity and the
expression of the obesity-suppressing neuropeptides proglucagon (GCG) and
brain-derived neurotrophic factor (BDNF) in the central nervous system.
Endocrinology 154, 3643–3651. doi:10.1210/en.2012-2151
30. Gruninger, T.R. et al. (2007) Molecular signaling involved in regulating feeding
and other mitivated behaviors. Mol. Neurobiol. 35, 1–19. doi:10.1007/BF0270
0621
31. Franceschi, C. et al. (2007) Inflammaging and anti-inflammaging: a systemic
perspective on aging and longevity emerged from studies in humans. Mech.
Ageing Dev. 128, 92–105. doi:10.1016/j.mad.2006.11.016
32. Piya, M.K. et al. (2013) Metabolic endotoxaemia: is it more than just a gut feeling?
Curr. Opin. Lipidol. 24, 78–85. doi:10.1097/MOL.0b013e32835b4431
33. Bercik, P. et al. (2011) The anxiolytic effect of Bifidobacterium longum ncc3001
involves vagal pathways for gut–brain communication.Neurogastroenterol.Motil.
23, 1132–1139. doi:10.1111/j.1365-2982.2011.01796.x
BiographiesShakuntla Gondalia is as Postdoctoral Fellow at Swinburne Uni-
versity of Technology, Victoria and her research interest incorpo-
rates gastrointestinal microbial ecology and nutritional intervention
with thepotential to improve thehealth and cognitive performance.
Her research aims to better understand the effectivemechanisms of
GI microbiota, nutritional and dietary intervention on individual’s
physiological and psychological health condition.
Professor Andrew Scholey is director of the Centre for Human
Psychopharmacology at Swinburne University, Melbourne. He is a
leading international researcher into the neurocognitive effects of
natural products, supplements and food components, having pub-
lished over 160 peer-reviewed journal articles, books and book
chapters. Scholey has been lead investigator in a series of studies
into the humanbiobehavioural effects of natural products, and their
neurocognition-enhancing and anti-stress/anxiolytic properties
(including first-into human neurocognitive assessment of Ginseng,
Sage and Lemon balm among others). His current research focuses
on neuroimaging and biomarker techniques to better understand
the mechanisms of cognitive enhancement.
Under theMicroscope
28 MICROBIOLOGY AUSTRALIA * MARCH 2015

Marsupial oral cavity microbiome
Philip S BirdA,D, Wayne SJ BoardmanB, Darren J TrottB and Linda L BlackallC
AThe University of Queensland, School of Veterinary Science, Faculty of Science, Gatton, Qld 4343, Australia
BThe University of Adelaide, School of Animal and Veterinary Sciences, Roseworthy, SA 5371, Australia
CSwinburne University of Technology, School of Science, Faculty of Science, Engineering and Technology, Hawthorn, Vic. 3122, Australia
DCorresponding author. Tel: +61 7 5460 1834, Fax: +61 7 5460 1922, Email: [email protected]
The oral microbiome of humans and animals will cause oral
disease within their lifetimes and include a large number of
endogenous cariogenic, periodontal and other opportunis-
tic pathogens. Studies over many decades have attempted
to determine which bacteria are involved in oral diseases.
Earlier studiesusedexclusively culture-basedmethods.Now
culture-independent methods are being used to determine
the composition of the microbiome in health and disease.
Therehavebeen limitednumbersof studies of themarsupial
microbiome and this report covers some of the research of
those studies.
Dental plaque, a natural oral biofilm is composed of many diverse
bacterial species, some of which are involved in the aetiology of
periodontal diseases (gingivitis and periodontitis)1. Factors such as
indigenous bacteria, host immune system, diet, host susceptibility
and time, interplay in thesediseases2. Therehavebeenmany studies
determining which of the causative agent(s) initiate oral diseases in
humans and domesticated animals. Marsupials also have oral dis-
eases3,4 and culture-dependent studies have shown a range of
bacteria can be isolated from the marsupial oral cavity5. However
culture-based studies, while useful to enable precise characterisa-
tion of putative periodontopathogens, generally underestimate
microbial community diversity. Culture-independent methods,
such as high-throughput DNA sequencing reveal a rich and diverse
bacterial community in the oral cavities of humans and companion
animals6-8, and the marsupial microbiome should also be more
thoroughly studied too.
The first bacteria ever to be seen under a microscope were the
plaque bacteria taken from Antonie van Leeuwenhoek’s own teeth
and reported in a letter to the Royal Society on 17 September 1683.
These first recorded observations of living bacteria showed ‘an
unbelievable great company of living animalcules and of enormous
number of a variety of shapes and sizes’. Since then many oral
prokaryotic species have been described. The Human Oral Micro-
biome Database (www.homd.org/) has to-date >200 bacterial spe-
cies described fromculture-dependentmethods and approximately
1,000 phylotypes detected by 16S rRNA gene sequencing of oral
samples from the human oral cavity using culture-independent
methods9. A search of the International Journal of Systematic and
EvolutionaryMicrobiologyover thepast 10 years for ‘oral’ revealed
a host of novel bacterial species isolated from the oral cavity of
humans and animals, including domesticated andwild (free-ranging
and captive) animals. In marsupials, novel microbial species were
revealed in the oral cavity of macropods associated with gingivitis
and oral necrobacillosis10.
In domesticated and wild animals, using culture-dependent meth-
ods, associations with disease and specific bacterial species have
been reported5. More recently, researchers have used high-
throughput DNA sequencing to study the oral microbiota of
healthy cats8 and dogs7. In adult horses 67% of 203 operational
taxonomic units (OTUs) were recovered, with the most frequent
genera being Prevotella and Porphyromonas (T Chinkangsadarn,
GJWilson, SWCorley and PS Bird, 2014, unpublished). In each case,
the results revealed a rich and diverse bacterial community in
much higher numbers than identified using culture- and cloning-
based studies. Therefore, in studies of animals with known oral
health status, health and disease could be correlated with the oral
microbiota detected using these new technologies.
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2015 10.1071/MA15009 29

We have shown that oral diseases such as gingivitis and periodontitis
can be found in a range of native Australian animals including
macropods, koalas (Phascolarctos cinereus), brushtail possums
(Trichosurus vulpecular) and bandicoots (Isoodon macrourus),
and that black-pigmented, anaerobic bacteria, belonging to the
genera Porphyromonas and Prevotella, are part of themicrobiota5.
Earlier studiesusingculturedependentmethods, showed that in the
normal oral microbiota of macropods, Gram-negative anaerobes
were poorly represented11. In contrast, Dent and co-workers12
reported that macropod oral cavities had a noticeable predomi-
nance of Gram-negative andGram-positive rods with the facultative
anaerobic Gram-negative rods comprising 40% of the cultivable
organisms although no Bacteroides spp. were isolated. In a study of
10 species of kangaroos and wallabies, black-pigmented anaerobic
bacteria comprised 21% of their normal oral microbiota13. Thus the
early culture-based studies have shown variability in the microbes
isolated and suggest that this approach has limitations.
Koalas do present with severe periodontal disease4 and with severe
loss of alveolar boneassociatedwith age andconditions suchas food
impaction3 (Figure 1). In koalas <7 years old with good oral health,
there was an absence of black-pigmented bacteria, compared to
koalas >7 years of age, where 50% harboured black-pigmented
bacteria, the majority of which were identified as Porphyromonas
gingivalis-like14. Currentwork characterising this bacterium, shows
the organism to be novel and that it may be associated with
periodontal disease in marsupials [Bird et al, 2015, submitted].
Another intriguingquestion relates to thekoala’sdietwhichconsists
of Eucalyptus leaves. How have the oral bacteria evolved in the
presence of such a toxic diet (e.g. high in essential oils) and how
does this affect the koala’s oral microbiome?
Oral necrobacillosis or lumpy jaw as it is commonly known, is a
leading cause of mortality in captive macropods and has been
reported in free-rangingmacropods15. The disease progresses from
plaque formation, gingivitis and periodontal disease to a necrotis-
ing, fatal osteomyelitis,16 (Figure 2) with all macropods susceptible,
particularly Eastern grey kangaroos17. Early studies of macropods
with lumpy jaw showed that Fusobacterium necrophorumwas the
most frequent isolate from lesions (81% prevalence) as well as the
most abundant organism in mixed cultures. It also was isolated in
high abundance from gingival margin samples taken from sites
remote from the lumpy jaw lesions in 61% of the animals with
disease18. While the principle infective agent in Australia appears to
be F. necrophorum, other organisms appear to play a role in this
disease. Recent work has shown that F. necrophorum sub-species
necrophorum is associated with organisms resembling Porphyro-
monas gulae in lumpy jaw inmacropods in South Australian zoos19.
Porphyromonas organisms distinct from both P. gingivalis and
P. gulae have been proposed as a novel species14 and were isolated
with increasing frequency from the oral cavity of macropods in our
studies, whichwarrants further evaluation into the role of this newly
described organism in jaw disease.
A study of the tammar wallaby pouch young suggested factors that
protect young animals against potentially pathogenic microbial
infections could include themicrobiome from thematernal saliva20.
The microbiomes of the pouch and saliva from the mother were
compared with the gastrointestinal tract (GIT) microbiome of the
pouch young using 16S rRNA gene comparative methods. Each
study site had a unique microbiome20. The maternal pouch har-
boured 41 unique Actinobacteria phylotypes, while in the saliva
there were 48 unique Proteobacteria phylotypes. The GIT of the
pouch young had a complex microbiome of 53 unique phylotypes
andof these,only nineweredetectedat eithermaternal site.Overall,
themajority of bacteria detected were novel species and each study
site possessed its own unique microbiome20.
In conclusion, a number of culture-based studies on the oral
microbiota of marsupials were conducted some 20–30 years ago.
Nowwith the deep sequencing culture-independentmethods it will
be possible to detect novel bacterial species in their oral cavities
and these are likely to have unique properties. In addition, new
studies will undoubtedly lead to insights into their evolution,
diversity and ecological role. We can speculate that co-evolution
of the marsupial oral microbiome has occurred with its host and
organisms such as the newly identified porphyromonad unique
to marsupials may represent an ancestral lineage distinct from
P. gulae and P. gingivalis. The oral microbiome of our marsupials
has received little attention and therefore definitely warrants more
Figure 1. Oral disease in the koala presented with an old mandibular fracture with compaction vegetation resulting in bone loss (credit: Ms Lyndall MPettett).
Under theMicroscope
30 MICROBIOLOGY AUSTRALIA * MARCH 2015

thorough exploration by keen and dedicated microbiologists for
novel bacteria and associated oral diseases.
References1. Tsang, K.L. et al. (2005) Caries and periodontal disease: two diseases, one biofilm.
Microbiol. Aust. 23, 110–112.
2. Marsh, P.D. (2003) Are dental diseases examples of ecological catastrophes?
Microbiology 149, 279–294. doi:10.1099/mic.0.26082-0
3. Lee, E.F. et al. (2011) Loss of tooth-supporting bone in the koala (Phascolarctos
cinereus) with age. Aust. J. Zool. 59, 49–53. doi:10.1071/ZO10047
4. Pettett, L.M. et al. (2012) The development of an oral health charting system for
koalas (Phascolarctos cinereus). J. Vet. Dent. 29, 232–241.
5. Bird, P.S. et al. (2002)Oral disease in animals: the Australian perspective. Isolation
and characterisation of black-pigmented bacteria from the oral cavity of marsu-
pials. Anaerobe 8, 79–87. doi:10.1006/anae.2002.0412
6. Pérez-Chaparro, P.J. et al. (2014) Newly identified pathogens associated with
periodontitis: A systematic review. J. Dent. Res. 93, 846–858. doi:10.1177/
0022034514542468
7. Sturgeon, A. et al. (2014) Characterization of the oral microbiota of healthy
cats using next-generation sequencing. Vet. J. 201, 223–229. doi:10.1016/
j.tvjl.2014.01.024
8. Sturgeon, A. et al. (2013) Metagenomic analysis of the canine oral cavity as
revealed by high-throughput pyrosequencing of the 16S rRNA gene. Vet. Micro-
biol. 162, 891–898. doi:10.1016/j.vetmic.2012.11.018
9. Dewhirst, F.E. et al. (2010) The human oral microbiome. J. Bacteriol. 192,
5002–5017. doi:10.1128/JB.00542-10
10. Antiabong, J.F. et al. (2013) The oral microbial community of gingivitis and
lumpy jaw in captive macropods. Res. Vet. Sci. 95, 996–1005. doi:10.1016/
j.rvsc.2013.08.010
11. Beighton, D. and Miller, W.A. (1977) A microbiological study of normal flora
of macropod dental plaque. J. Dent. Res. 56, 995–1000. doi:10.1177/002203
45770560083101
12. Dent, V.E. (1979) The bacteriology of dental plaque from a variety of zoo-
maintained mammalian species. Arch. Oral Biol. 24, 277–282. doi:10.1016/
0003-9969(79)90089-X
13. Samuel, J.L. (1982) The normal flora of the mouths of macropods (Marsupialia:
macropodidae).Arch. Oral Biol.27, 141–146. doi:10.1016/0003-9969(82)90134-0
14. Mikkelsen, D. et al. (2008) Phylogenetic analysis of Porphyromonas species
isolated from the oral cavity of Australian marsupials. Environ. Microbiol. 10,
2425–2432. doi:10.1111/j.1462-2920.2008.01668.x
15. Borland, D. et al. (2012) Oral necrobacillosis (‘lumpy jaw’) in a free-ranging
population of eastern grey kangaroos (Macropus giganteus) in Victoria. Aust.
Mammal. 34, 29–35. doi:10.1071/AM10031
16. Bakal-Weiss, M. et al. (2010) Use of a sustained release chlorhexidine varnish
as treatment of oral necrobacillosis in Macropus spp. J. Zoo Wildl. Med. 41,
371–373. doi:10.1638/2010-0004.1
17. Butler, R. (1981) Epidemiology and management of ‘lumpy jaw’ in macropods.
In Fourth International Conference on the Wildlife Diseases Association.
Sydney, Australia.
18. Samuel, J.L. (1983) Jaw disease in macropod marsupials: bacterial flora isolated
from lesions and from themouths of affected animals. Vet. Microbiol. 8, 373–387.
doi:10.1016/0378-1135(83)90050-0
19. Antiabong, J.F. et al. (2013) Amolecular survey of a captive wallaby population for
periodontopathogens and the co-incidence of Fusobacterium necrophorum
subspecies necrophorum with periodontal diseases. Vet. Microbiol. 163,
335–343. doi:10.1016/j.vetmic.2013.01.012
20. Chhour, K.L. et al. (2010) An observational study of the microbiome of the
maternal pouch and saliva of the tammar wallaby, Macropus eugenii, and of
the gastrointestinal tract of the pouch young. Microbiology 156, 798–808.
doi:10.1099/mic.0.031997-0
BiographiesDr Philip Bird is an Adjunct Associate Professor at the School of
Veterinary Science. His research interests are the oral microbiology
of human and animals, especially that of native Australian animals.
Dr Wayne Boardman is Senior Lecturer, at the School of Animal
and Veterinary Sciences, University of Adelaide. An experienced
wildlife veterinarian, he is a diplomate of the European College of
Zoological Medicine in Wildlife Population Health.
Dr Darren Trott is Associate Professor of Veterinary Microbiology
at The University of Adelaide. His research interests cover compar-
ative aspects of antimicrobial resistance in animals andhumans, new
drug development and gastrointestinal microbial ecology.
Linda L Blackall is a microbial ecologist who has studied many
different complex microbial communities ranging from host asso-
ciated through to free living in numerous environments. Her
research has covered mammalian microbiomes spanning marsu-
pials, humans, ruminants and horses and the methods used allow
elucidation of massive microbial complexity and function in these
diverse biomes. She is a Professor of Biosciences at Swinburne
University of Technology in the Faculty of Science, Engineering and
Technology.
Figure 2. Wallaby skull showing swollen lower jaw due to oral necrobacillosis or lumpy jaw.
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2015 31

Relative abundance of Mycobacterium in ovineJohne’s disease
Nostudyhasdeterminedwhat proportion of the totalmicro-
biota comprises the genusMycobacterium in ovine Johne’s
disease (OJD) tissues. We aimed to assess the relative abun-
dance of Mycobacterium in the ileocaecal lymph node,
involved and uninvolved ileal mucosa from sheep with and
without OJD, using three extraction methods. Eight sheep,
fourwithand fourwithoutOJD,were recruited.Pyrosequen-
cingof the16SrRNAgene amplicons for all samples revealed
that Mycobacterium represented between 0-92% (average
38%) of the total microbiota of samples from sheep with
OJD, and 0-85% (average 13%) of sheep without OJD. Only
sheepwithOJDhad samples that were positive for the IS900
(MAP) element. Mycobacterial strains other than MAP may
provide competitive exclusion ofMAP and should be further
investigated.
Mycobacterium avium subspecies paratuberculosis (MAP) is
the etiologic agent of ovine Johne’s disease (OJD), a chronic,
Andy O LeuA, Paul PavliB,C, David M GordonA, Jeff CaveD, Jacek M GowzdzE, Nick LindenD,Grant Rawlin
E, Gwen E Allison
A,Cand Claire L O’Brien
A,B,C,F
AResearch School of Biology, Australian National University, Acton, ACT 0200, Australia
BGastroenterology and Hepatology Unit, Canberra Hospital, Garran, ACT 2605, Australia
CMedical School, Australian National University, Acton, ACT 0200, Australia
DDepartment of Environment and Primary Industries, Wodonga, Vic. 3690, Australia
EAgriBio, Bundoora, Vic. 3083, Australia
FCorresponding author. Tel: +61 2 6244 4023, Email: [email protected]
LabReport
32 10.1071/MA15010 MICROBIOLOGY AUSTRALIA * MARCH 2015

contagious granulomatous enteritis of ruminants. The disease
causes significant morbidity and mortality worldwide, resulting in
significant economic losses1.
To our knowledge, no study has determined what fraction of the
microbial community of non-OJD and OJD affected tissue com-
prisesMycobacterium. Cheung et al. (2013)2 found thatMycobac-
terium tuberculosis represented less than 1% of the total microbial
community of sputum samples of infected patients. Does Myco-
bacterium comprise such a small fraction of the microbial commu-
nity in all infectious states andspecimens?MAP is aputative triggerof
Crohn’s disease in humans, and is detected in some studies but not
others, including a recent study of ours3. Is this because it is not
being detected?
While other studies have quantified Mycobacterium or MAP in
clinical and subclinical specimens4,5, no study has determined what
fraction of the microbial community comprises Mycobacterium in
mucosa from sheep with and without OJD. The primary aim is to
assess the relative abundance of Mycobacterium in the microbial
communities of macroscopically normal bowel mucosa obtained
>50 cm downstream the ileocaecal valve (sheep without OJD)
or involved mucosa (sheep with OJD), involved (or normal for
sheep without OJD) ileal mucosa adjacent the ileocaecal lymph
node, and the ileocaecal lymph node of sheep with and without
OJD.
Samples of involved and uninvolved ileal mucosa, and ileocaecal
lymph nodes were collected from eight sheep, four with and four
without OJD, from farms around Rutherglen, Australia. Sterile
gloves and implements were used to extract the ileocaecal node
before the bowel was opened, so that it did not come into contact
with the skin or bowel microbiota. Total DNA was extracted from
each sample using three different extraction methods. The first
protocol followed the Qiagen DNeasy kit protocol and included an
enzymatic lysis step and bead-beating step. The second and third
extractions were the same as the first, except the second protocol
included a freeze-thaw step, and the third protocol a boiling step.
Technical replicates were included for the first protocol. All samples
were amplified using barcoded primers targeting the universal
bacterial16S rRNA gene, and pyrosequenced using a 454 Genome
Sequencer FLX-Titanium platform, as previously described3.
Sequences were analysed using Mothur6. IS900 PCR was used to
determine whether or not Mycobacterium detected in a given
sample was MAP, or not.
Four sheep (1, 2, 4 and 5; for example, see Figure 1) were diagnosed
withOJDbyqualified veterinarians andpathologists, usingmethods
previously described7,8. Briefly, sheep were initially assessed as
having OJD, or not, based on physical signs, including condition,
scouring, ill-thrift, and lethargy during mobility. Diagnosis was
confirmed by a NATA-accredited pathology laboratory (AgriBio,
Victoria), and included MAP culture and DNA analysis, as well as
microscopic visualization of acid-fast bacteria using ZN staining .
Sheep 1 was initially suspected to have OJD, however negative
culture and staining results excluded it from a diagnosis of OJD, and
it was found to have metastatic cancer.
Pyrosequencing showed thatMycobacterium representedbetween
0–92% (average 38%) of the total microbiota of samples from sheep
withOJD,and0-85%(average13%)of sheepwithoutOJD(Figure2).
Mucosal samples from sheep 7were excluded due to the inability to
amplify products from them. Sheep 4 and 5 each had a positive
lymph node sample for one extraction, comprising 6% and 68%
Mycobacterium reads, respectively.These levelswere similar to that
observed in the mucosal samples from the same animal.
The lymph node and mucosa samples of sheep 1 were negative
when tested with theMAP-specific primers. Sheep 1may have had a
paucibacillary form of OJD. Samples from all other sheep with
OJD were positive, except the normal mucosa sample of sheep 4
(Table 1). All samples fromsheepwithoutOJDwere negative for the
IS900 element.
No significant effect of extraction method was detected for either
Simpson or Shannon diversity estimates (Simpson Diversity, AOV:
P > F=0.384; ShannonDiversity, AOV: P > F=0.443), nor technical
replicates (Simpson Diversity, AOV: P > F=0.398; Shannon Diver-
sity, AOV: P > F=0.566).
Each sample was covered by an average of 2, 644 quality sequences.
Six phyla (Firmicutes, Bacteroidetes, Actinobacteria, Proteobac-
teria, Synergistetes, and TM7) represented ~98% of all sequences
Figure 1. Ovine Johne’s disease affected sheep #2, showing physicalsigns of wasting.
LabReport
MICROBIOLOGY AUSTRALIA * MARCH 2015 33

(Figure 3). Actinobacteria, of which Mycobacterium is a member,
was overrepresented in sheep with OJD, except sheep 1. Microbial
community diversity declined as Mycobacterium increased
(R2=0.705, p=0.001) (Figure 4).
To our knowledge, this is the first study to describe the full
microbial community of mucosa from sheep with and without
OJD. Although studies have enumeratedMAP in the feces of sheep
using RT-PCR5,9, no study has determined what fraction of the
100
90
80
70
60
50
Myc
obac
teriu
m (
%)
40
30
20
10
0
Figure 2. Proportion of involved and normal mucosa and lymph node tissues (sheep 4 and 5) containing Mycobacterium. Sheep with Johne’sdisease are coloured orange, without blue. Technical replicates (normal mucosa only) have diagonal texturing. The first numeral in the labelindicates the animal number; NM, normal mucosa; IM, involved mucosa; LN, lymph node; the second numeral indicates the DNA extraction: ‘1’, ‘2’and ‘3’ are replicates, ‘12’ is the technical replicate for normal mucosa (DNA extraction 1) only. Sheep 1, 2, 4 and 5 had OJD, sheep 3, 6 and 8did not.
Table 1. Sheep characteristics, pathology and MAP-specific PCR results
Sheep 1 2 3 4 5 6 7 8
Diagnosis OJD and MIC OJD Non-OJD OJD OJD Non-OJD Non-OJD Non-OJD
Sex F F F F F F F F
Age 7 4 7 7 4 6 3 6
Vaccination N Y N N Y N Y N
Breed Merino Merino Merino X
Leicester
Merino X
Leicester
Merino Merino Merino Merino
Intestinal
lesions
Severe Moderate None Moderate Mild None None None
MAP culture + + – + + – – –
AFB in
Involved
Mucosa
Absent Occasional Absent Numerous Numerous Absent Absent Absent
AFB in
Lymph node
Absent Absent Absent Moderate Numerous Absent Absent Absent
MAP-specific IS900 PCR
NM – + – – + – – –
IM – + – + + – – –
LN – + – + + – – –
OJD, ovine Johne’s disease; MIC, metastatic intestinal cancer;MAP,Mycobacterium avium subspecies paratuberculosis; Y, yes; N, no; AFB, acid-fast bacteria;NM, normal mucosa; IM, involved mucosa. All samples were positive for bacterial DNA using the 16S rRNA gene primers, except sheep 7 samples.
LabReport
34 MICROBIOLOGY AUSTRALIA * MARCH 2015
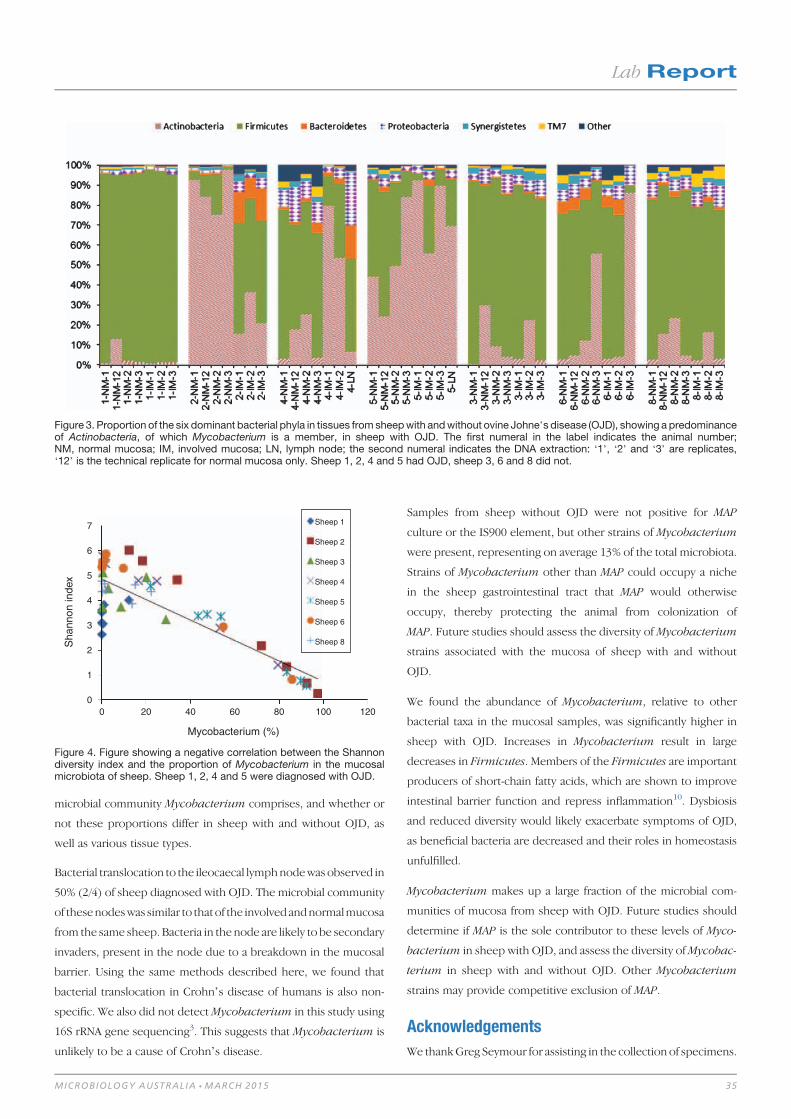
microbial community Mycobacterium comprises, and whether or
not these proportions differ in sheep with and without OJD, as
well as various tissue types.
Bacterial translocation to the ileocaecal lymphnodewas observed in
50% (2/4) of sheep diagnosed with OJD. The microbial community
of thesenodeswas similar to thatof the involvedandnormalmucosa
from the same sheep. Bacteria in the node are likely to be secondary
invaders, present in the node due to a breakdown in the mucosal
barrier. Using the same methods described here, we found that
bacterial translocation in Crohn’s disease of humans is also non-
specific. We also did not detectMycobacterium in this study using
16S rRNA gene sequencing3. This suggests that Mycobacterium is
unlikely to be a cause of Crohn’s disease.
Samples from sheep without OJD were not positive for MAP
culture or the IS900 element, but other strains of Mycobacterium
were present, representing on average 13% of the total microbiota.
Strains of Mycobacterium other than MAP could occupy a niche
in the sheep gastrointestinal tract that MAP would otherwise
occupy, thereby protecting the animal from colonization of
MAP. Future studies should assess the diversity of Mycobacterium
strains associated with the mucosa of sheep with and without
OJD.
We found the abundance of Mycobacterium, relative to other
bacterial taxa in the mucosal samples, was significantly higher in
sheep with OJD. Increases in Mycobacterium result in large
decreases in Firmicutes. Members of the Firmicutes are important
producers of short-chain fatty acids, which are shown to improve
intestinal barrier function and repress inflammation10. Dysbiosis
and reduced diversity would likely exacerbate symptoms of OJD,
as beneficial bacteria are decreased and their roles in homeostasis
unfulfilled.
Mycobacterium makes up a large fraction of the microbial com-
munities of mucosa from sheep with OJD. Future studies should
determine if MAP is the sole contributor to these levels of Myco-
bacterium in sheep with OJD, and assess the diversity ofMycobac-
terium in sheep with and without OJD. Other Mycobacterium
strains may provide competitive exclusion of MAP.
Acknowledgements
We thankGreg Seymour for assisting in the collection of specimens.
Figure 3. Proportion of the six dominant bacterial phyla in tissues from sheepwith andwithout ovine Johne’s disease (OJD), showing a predominanceof Actinobacteria, of which Mycobacterium is a member, in sheep with OJD. The first numeral in the label indicates the animal number;NM, normal mucosa; IM, involved mucosa; LN, lymph node; the second numeral indicates the DNA extraction: ‘1’, ‘2’ and ‘3’ are replicates,‘12’ is the technical replicate for normal mucosa only. Sheep 1, 2, 4 and 5 had OJD, sheep 3, 6 and 8 did not.
7
6
5
4
3
Sha
nnon
inde
x
Mycobacterium (%)
2
1
00 20 40 60 80 100
Sheep 1
Sheep 2
Sheep 3
Sheep 4
Sheep 5
Sheep 6
Sheep 8
120
Figure 4. Figure showing a negative correlation between the Shannondiversity index and the proportion of Mycobacterium in the mucosalmicrobiota of sheep. Sheep 1, 2, 4 and 5 were diagnosed with OJD.
LabReport
MICROBIOLOGY AUSTRALIA * MARCH 2015 35

References1. Salem, M. et al. (2013) Mycobacterium avium subspecies paratuberculosis:
an insidious problem for the ruminant industry. Trop. Anim. Health Prod. 45,
351–366. doi:10.1007/s11250-012-0274-2
2. Cheung, M.K. et al. (2013) Sputummicrobiota in tuberculosis as revealed by 16S
rRNA pyrosequencing. PLoS ONE 8, e54574. doi:10.1371/journal.pone.0054574
3. O’Brien, C.L. et al. (2014) Detection of bacterial DNA in lymph nodes of Crohn’s
disease patients using high throughput sequencing. Gut 63, 1596–1606.
doi:10.1136/gutjnl-2013-305320
4. Park, K.T. et al. (2014) Development of a novel DNA extraction method for
identification and quantification of Mycobacterium avium subsp. paratubercu-
losis from tissue samples by real-time PCR. J. Microbiol. Methods 99, 58–65.
doi:10.1016/j.mimet.2014.02.003
5. Logar, K. et al. (2012) Evaluation of combined high-efficiency DNA extraction and
real-time PCR for detection ofMycobacteriumavium subsp. paratuberculosis in
subclinically infected dairy cattle: comparison with faecal culture, milk real-time
PCR and milk ELISA. BMC Vet. Res. 8, 49. doi:10.1186/1746-6148-8-49
6. Schloss, P.D. et al. (2009) Introducing mothur: open-source, platform-indepen-
dent, community-supported software for describing and comparing microbial
communities. Appl. Environ. Microbiol. 75, 7537–7541. doi:10.1128/
AEM.01541-09
7. Gwozdz, J.M. et al. (2000) Vaccination against paratuberculosis of lambs already
infected experimentally with Mycobacterium avium subspecies paratubercu-
losis. Aust. Vet. J. 78, 560–566. doi:10.1111/j.1751-0813.2000.tb11902.x
8. Whittington, R.J. et al. (2013) Development and validation of a liquid medium
(M7H9C) for routine culture ofMycobacterium avium subsp. paratuberculosis
to replace modified Bactec 12B medium. J. Clin. Microbiol. 51, 3993–4000.
doi:10.1128/JCM.01373-13
9. Sting, R. et al. (2014) Detection of Mycobacterium avium subsp. paratubercu-
losis in faeces using different procedures of pre-treatment for real-time PCR in
comparison to culture. Vet. J. 199, 138–142. doi:10.1016/j.tvjl.2013.08.033
10. Pryde, S.E. et al. (2002) The microbiology of butyrate formation in the human
colon. FEMS Microbiol. Lett. 217, 133–139. doi:10.1111/j.1574-6968.2002.
tb11467.x
Biographies
Andy Leu graduated from the Australian National University (ANU)
with aBachelor ofMedical Science (Honours I) in2013.HisHonours
project examined the relative abundanceofMycobacteriumavium.
paratuberculosis (MAP) in Johne’s disease tissues. Andy was re-
cently awarded an Australian Postgraduate Award scholarship to
pursue a PhD under the supervision of Dr Gene Tyson.
Paul Pavli is a gastroenterologist with clinical and basic scientific
research interests in the inflammatory bowel diseases. He was on
the steering committee of the multicentre Australian study using
long-term anti-mycobacterial agents in Crohn’s disease (Selby
et al., Gastroenterology, 2007). Current research interests are in
the interaction between the innate inflammatory response and the
intestinal microbiome.
David Gordon received his BSc from King’s College London and
his PhD fromMcGill University,Montreal.His research concerns the
ecology, population genetics and evolution of the Enterobacter-
iaceae, especially E. coli.
Jeff Cave graduated from Murdoch University in 1988 and spent
twoyears inmixedprivatepractice inSouthAustralia beforeworking
in the Pacific for much of the 1990s. For the past 16 years he has
worked as the District Veterinary Officer in Wodonga, Victoria.
Jacek Gwozdz is a Veterinary Pathologist at the State veterinary
laboratory in Victoria. He received his PhD in Veterinary Science
fromtheMasseyUniversity inNewZealandandworkedover thepast
16 years in research and diagnostic microbiology and pathology.
NickLinden is a research scientistwith theVictorianDepartmentof
Primary Industries and Environment, he has a particular interest in
the lamb supply chain and is passionate about linking on farm
production through to carcass appraisal and consumer acceptance.
His recent projects have been focused on lamb feed conversion
efficiency as well as on-line grading of lamb carcasses.
Grant Rawlin is Research Leader of Veterinary Pathobiology at the
State veterinary laboratory in Victoria. His career has spanned
veterinary diagnostics as a pathologist, research in veterinary and
humanmicrobiologyandresearch intocontrolof infectiousdiseases
of animals at a population level.
Dr Gwen Allison received her PhD in Microbiology from North
Carolina State University. Her research interests have included
molecular biology of lactic acid bacteria and microbial ecology of
the gastrointestinal tract and environment. The current research
was conducted when Dr Allison was a Senior Lecturer at the ANU.
Dr Claire O’Brien received her PhD from the Australian National
University Medical School (ANUMS) in 2012. Her main research
interest is thegutmicrobiome,particularly inCrohn’sdisease.Claire
is an NHMRC Peter Doherty Fellow at the ANUMS and Canberra
Hospital.
LabReport
36 MICROBIOLOGY AUSTRALIA * MARCH 2015

Interactions with other microbiology societiesthrough Microbiology AustraliaThe September 2014 issue ofMicrobiology Australia was a special
issue that reflected on the ‘Microbial diseases and products that
shaped world history’. A major focus involved microbes of WWI
and their effects on ANZACs and Turkish soldiers. The timing of
the issue is particularly fitting as we reflect on the 100th anniversary
of the terrible losses suffered by both sides involved in the Gallipoli
conflict.
The idea for the issue came from Ipek Kurtböke who Guest
Edited the joint effort between the Australian, New Zealand and
Turkish Microbiology Societies. The issue contains excellent
articles on the major diseases of world history, and early efforts to
control them.
TheTurkish articleswerehighly informative about their experiences
with smallpox, tuberculosis and typhus as well as the huge losses
the Turkish troops suffered at the hands of infectious diseases.
In addition, the articles remind us that much is owed to the Turks
for the early lessons they learned through their contributions to
infection control. This is particularly so for their early adoption of
variolation for the prevention of smallpox, an experience that led
directly to Jenner’s seminalworkwith cowpox and thedevelopment
of modern vaccination.
At the invitation of the TurkishMicrobiology Society, Professor Paul
Young, President of ASM, Ian Macreadie, Editor of Microbiology
Australia and Ipek Kurtböke attended the annual Turkish
Microbiology Society meeting in Antalya, Turkey in November
2014. They participated in the opening ceremony and gave scientific
presentations during the meeting. At the opening ceremony,
Professor Young reminded the audience of the strong bonds that
were forged between our three countries after WWI and which
remain to this day. This connection was most notably encapsulated
in the words of Ataturk in 1934. Professor Young’s quotation of this
very moving tribute gained very warm applause from the audience:
Those heroes that shed their blood and lost their lives. . .Youare now lying in the soil of a friendly country. Therefore restin peace. There is nodifference between the Johnnies and theMehmets to us where they lie side by side here in this countryof ours. . .You, themothers,who sent their sons from farawaycountries wipe away your tears; your sons are now lying inour bosom and are in peace, after having lost their lives onthis land they have become our sons as well.
Wehave learnedmuch fromour interactions. It was a privilege to be
hosted by such gracious hosts and we look forward to welcoming a
delegation of Turkish microbiologists to Canberra for ASM2015.
The success of our interactions with other microbiology societies
is something ASM would like to foster and it is hoped that every
few years a similar joint issue of Microbiology Australia can be
produced.
ASM History SIG: Microbiology AustraliaThe ASM History SIG was launched at ASM Adelaide 2013 at a
meeting on 12 noon Tuesday 9th July 2013 and Dr Diane Lightfoot
was elected convener for the first 2 years of this SIG.
The purpose of this SIG is to record the history of Microbiology in
Australia, in the ASM archives, including details about certain
microbiologists, microbiological institutions, and microorganisms,
as well as disseminate this information to ASM members.
Regularly notes on topics of interest will be contributed to the ASM
Victorian Branch Newsletter from the History SIG (e.g. prominent
microbiologists, significantmicrobiological discoveries by Victorian
microbiologists, microbiological institutions/major events at these
institutions). In future it is intended to have a History of Australian
Microbiology Symposium at the ASM annual scientific meeting.
The AGM of the History SIG will be held at the ASM scientific
meeting in Canberra July 2015.
If ASM members have that significant ASM memorabilia that they
would like to donate to the ASM archives or suggestions of topics
suitable for possible symposia at the ASM Annual Meetings. Please
send details of memorabilia or suggestions/topics of interest or for
possible symposia to:
HistorySIGconvener, c/oAustralianSociety forMicrobiologyOffice,
9/397 Smith Street, Fitzroy, Vic. 3068, Australia.
Nezahat Gurler, Paul Young, Ian Macreadie, _Ipek Kurtboke, PhilippeDesmeth and Bulent Gurler.
ASMAffairs
MICROBIOLOGY AUSTRALIA * MARCH 2015 10.1071/MA15011 37

Recent developments in virology by AustralianresearchersJoseph R Freitas and Suresh Mahalingam
Institute for Glycomics, Griffith University, Gold Coast, Queensland
Virology is a growing field within Australia. Increased funding is
being allocated to discovering how viruses interact with their
host/vectors(s) and to the development of better treatments
and vaccines. There have been many recent exciting new develop-
ments in Australian virology. Space limitations mean that we can
only highlight a small number of these achievements in this brief
overview of the current virology landscape in Australia.
The increasing prevalence or re-emergence of certain alphaviruses,
such as Ross River virus (RRV), Sindbis virus, and Chikungunya
virus, is a cause for concern and has attracted increased research
interest in recent years. These viruses are known as arthritogenic
alphaviruses because they cause arthritis-like symptoms. The ability
of these viruses to directly induce bone pathology has remained
poorly defined to date. Chen et al.1 recently shed some light on this
issue by revealing that RRV can infect human osteoblasts and that
osteoblast infection leads to IL-6-dependent bone loss in a mouse
model. This discovery of the interaction between osteoblasts,
inflammatory factors and alphaviruses is a plausible explanation for
how viruses cause chronic joint pain.
Infections with certain viruses, such as West Nile virus (WNV) and
dengue virus, can lead to encephalitis, which is associated with
high mortality. Inflammatory cells play a central in the progression
of these diseases. Currently there are no specific targeted treat-
ments to modulate the action of inflammatory cells. Getts et al.2
recently demonstrated that infusion of immune-modifying micro-
particles (IMPs) into WNV-infected mice significantly reduced
the symptoms of central nervous system infection. The continued
injection of IMPs to mice over several days led to an improved
survival rate. Similar therapeutic effects were shown for other
inflammation-mediated diseases. The therapeutic potential of
such IMPs for a variety of immune-related disorders looks to be
very promising.
Dengue (DENV) is a mosquito-borne virus that infects hundreds
of millions of people annually. It has a widespread geographical
prevalence that continues to expand. There is an urgent need for an
effective vaccine against DENV. However, this has proven difficult
due to the virus having four different serotypes, with a general
consensus that a successful vaccine will need to induce immunity to
each specific serotype. The identification of antibodies with broad
cross-reactivity to all serotypes by Dejnirattisai et al.3 led to the
discovery of a novel DENV epitope, which has high potential for use
as a successful vaccine antigen. Although this study was performed
outside of Australia, the threat of DENV in Australia and the strong
collaborative input fromUniversity ofMelbourne researchersmerits
its mention here.
Viruses with the ability to pass from animals to humans (‘zoonotic
viruses’) are very unpredictable because they have the potential to
mutate and become deadly in new hosts. The most notable local
example of this is theHendra virus outbreaks that occurred over the
past two decades. Research into this virus and its natural hosts led to
the successful development and deployment of a vaccine against
Hendra in 20124. A newly identified virus found in Australian bats,
dubbed Cedar virus, by Marsh et al.5 in 2012 was found to be very
similar to the Hendra and Nipah viruses. The one key difference is
that it does not cause disease in several mammals that are suscep-
tible to Hendra and Nipah. This fascinating discovery will no doubt
assist scientists to understand what exactly it is that makes certain
viruses deadly and others less so.
Norovirus (NoV) is the leading cause of gastroenteritis worldwide
and is known to cause thousands of deaths in developing countries.
Most outbreaks occur within institutional settings such as hospitals
and aged-care facilities. One particular genotype of the virus is
responsible for the majority of infections, the genogroup II geno-
type 46. Since current treatment for NoV infections is largely
preventative, there is an urgent need for an effective vaccine or
antiviral. By using high throughput screening methods against the
RNA polymerase of NoV, Eltahla et al.7 identified a very promising
target for development of effective antivirals against this disease.
With these and the many other researchers around Australia cur-
rently active in thefield,we canexpect continuingmajor advances in
the discipline of virology in 2015 and in the years to come.
References1. Chen, W. et al. (2014) Arthritogenic alphaviral infection perturbs osteoblast
function and triggers pathologic bone loss. Proc. Natl. Acad. Sci. USA 111,
6040–6045. doi:10.1073/pnas.1318859111
2. Getts, DR et al. (2014) Therapeutic inflammatory monocyte modulation using
immune-modifying microparticles. Sci. Transl. Med. 6, 219ra7.
3. Dejnirattisai, W. et al. (2015) A new class of highly potent, broadly neutralizing
antibodies isolated from viremic patients infected with dengue virus. Nat. Immu-
nol. 16, 170–177. doi:10.1038/ni.3058
4. Broder, C.C. et al. (2013) A treatment for and vaccine against the deadlyHendra and
Nipah viruses. Antiviral Res. 100, 8–13. doi:10.1016/j.antiviral.2013.06.012
5. Marsh, G.A. et al. (2012) Cedar virus: a novel Henipavirus isolated from Australian
bats. PLoS Pathog. 8, e1002836. doi:10.1371/journal.ppat.1002836
6. Eden, J.S. et al. (2013) Recombinationwithin thepandemicNorovirusGII.4 lineage.
J. Virol. 87, 6270–6282. doi:10.1128/JVI.03464-12
7. Eltahla, A.A. et al. (2014) Nonnucleoside inhibitors of Norovirus RNA
polymerase: scaffolds for rational drug design. Antimicrob. Agents Chemother.
58, 3115–3123. doi:10.1128/AAC.02799-13
ASMAffairs
38 10.1071/MA15013 MICROBIOLOGY AUSTRALIA * MARCH 2015

Clinical Serology and Molecular SIGFrom the retiring National Convenor, David Dickeson
The Clinical Serology Special Interest Group was established after a
meeting in Adelaide in 1989with Peter Robertson (now retired from
PrinceofWalesHospital, Randwick) the inaugural national convenor
andonly a fewdozenmembers. Itwas established toprovide a forum
for the discussion of serological techniques with the objective to
develop a strategy to monitor and control the quality of diagnostic
reagents. This was published in 1994 (Backhouse et al., Australian
Microbiologist,1994;15:37–45).BranchesofthisSIGwereformedin
most states from 1989 onwards. I became involved as NSW branch
treasurer in 1993with LeonHeron as convenor and I have remained
as secretary/treasurer ever since. The group was expanded in 2003
to include molecular techniques in the medical diagnostic industry
and the combined SIG convenor was Robyn Wood (formerly from
Queensland Medical Laboratory, then TGA). In 2004 I took on the
position as convenor at the ASM annualmeeting in Sydney and have
continued in that role until now.
The CS&M SIG Working Group was formed in 2009 after several
ASM meetings and NRL workshops with the great assistance of
Sheena Adamson (Communicable Diseases Branch, NSW Health)
who remains as the working group secretary. This group is
overseen by the ASM Clinical Microbiology Standing Committee
and aims to be a proactive source of education and information
and able to give advice on serology and related molecular testing.
Teleconferences held every 3 months have kept an expert group of
scientists and medical microbiologists in contact. Dissemination
of information has been achieved through ASM, NRL, RCPA QAP,
various direct emails and the national email routing ‘SERSIG’,
which links all members with requests for information or help in
resolving problems. Now SERSIG has 419 members.
At the ASM annual scientific meeting each year, we organise a
workshop with topics including QAP, case studies and issues with
diagnostic kit failures or problems. Previous workshops connected
with ASM annualmeetings in Adelaide,Melbourne and Sydneywere
organised by Ros Escott (RCPA QAP now retired). In 2014 a molec-
ular diagnosis workshop was held on the Saturday afternoon at the
Peter Doherty Institute prior to the national meeting. The meeting
was chaired by Wayne Dimech and Darren Jardine with over 80
attending. It included topics on molecular testing, validation and
standards, choosing equipment and new technologies.
An Annual General Meeting of the SIG is organised by the convenor
at each yearlymeeting and it includes branch reports, industry input
anddiscussionof current topics. InMelbourne last year 17members
attended, reports were tabled for NSW, QLD, VIC, SA andWA, and a
call for nominations for new national convenor, secretary and
workshop organiser was made.
After over 10 years of service as national convenor and serving as
secretary/treasurer of the NSW branch since 1993, I will be handing
over the duties and responsibilities of the ASM CS&M SIG to the
new convenor. Coordinating, participating and helping with all the
events and discussions of this SIG has been a pleasure for over
22 years. I would like to thank Wayne for his help and advice as
secretary and administrator of the SERSIG email routing and espe-
cially thank Sheena for being the driving force behind the working
group, organising teleconferences, emails to members and letters
to governing bodies. I trust the Clinical Serology and Molecular
SIG will continue to meet the needs of members by providing
useful and timely scientific information about diagnostic serology
and molecular techniques and related matters. I am sure the SIG
will be left in good hands with the help of the new executive.
New office bearers for the SIG are:
Convenor: Linda Hueston, Pathology West – ICPMR, Westmead,
NSW.
Email: [email protected]
Secretary: Megan Wieringa, Monash Health, Clayton, Victoria.
Email: [email protected]
Workshoporganiser:BruceWong,PaLMS,RNSH,St Leonards,NSW.
Email: [email protected]
State Branch Convenors 2014
NSW: Deane Byers, RCPA.
Email: [email protected]
Victoria: Wayne Dimech (retiring), NRL.
Email: [email protected]
Queensland: Cheryl Bletchley and Theo Sloots (retiring).
Email: [email protected]
South Australia: Trish Hahesy, IMVS.
Email: [email protected]
Western Australia: Justin Morgan, PathWest Laboratory
Medicine WA.
Email: [email protected]
Tasmania: Louise Cooley, RHH.
Email: [email protected]
ASMAffairs
MICROBIOLOGY AUSTRALIA * MARCH 2015 10.1071/MA15014 39

Report from the ASM Antimicrobial SpecialInterest Group (ASIG)
John Merlino, on behalfof ASIG
ASM ASIG Convenor/ChairEmail:[email protected]
The ASM Antimicrobial Special Interest Group (ASIG) is made up
of colleagues and ASM members you have an interest in antimicro-
bials. Members of the ASIG committee include medical, veterinary,
CDS Users, mycology and parasitology consultants. The Antimicro-
bial SIG is one of the largest Special Interest Groups in the ASM.
In 2012 we had over 923 ASIG members. In 2013 we had 721
members listed under ASIG. Some of our members have retired.
Numbers are yet to be finalised in 2014.
In 2014 ASM ASIG members were actively involved with local and
national ASM Branch meetings in various states – this is usual in the
form of communication and newsletters. Committee actively com-
municates via emails and website www.asig.org.au on emerging
issues – methodology on susceptibility testing and standards on
antimicrobials and antimicrobial resistance. Members of the com-
mittee actively take part in planning workshops titles and speakers,
e.g. Annual Scientific ASM Meetings – this has been the focus for
many years. Discussions on antimicrobial issues take place at these
meetings.
In Melbourne in 2014, the Antimicrobial Special Interest Group
Workshop on Antimicrobial Resistance focused onMALDI-TOF and
Susceptibility Testing with both CLSI and EUCAST methodology.
This was a DRY Workshop with practical slide demonstrations with
discussions and a QUIZ with equipment such as buzzers provide
via the RCPA. Speakers’ presentations are available on (powerpoint
in PDF) on the ASIG website www.asig.org.au, http://asig.org.
au/australian-society-for-microbiology-asm-2014-annual-scientific-
meeting-melbourne-australia-workshop-presentations/.
TheBioMerieux Identifying Resistance Award in 2014went to A/Prof
Denis Spelman for his long continued support for antimicrobials
and infectious control practices; this was presented in Melbourne
annual scientific meeting.
ASIG became affiliated with the International Society of Chemo-
therapy (ISC) after a formal invitation. Some of our members who
specialise in specific antimicrobial areashavebeen invited and taken
part in symposiums andother events at their own costs.Members of
ASIG committee actively communicate with other local and inter-
national antimicrobial groups on emerging issues in antimicrobial
resistance for example with the MRSA International Committee.
This remains ongoing in 2014–15. ASIG Members have also been
involved in EuropeanCongress Clinical Microbiology and Infectious
Diseases (ECCMID)meetings overseas – expensesmet by their own
trust fund not ASM. Members have been invited and involved in
writing articles on antimicrobials for Microbiology Australia and
overseas journals. When requested ASIG members who specialise
in specific areas of antimicrobials interests review articles for local
and international journals.
The ASIG convenor/committee and members communicates with
Division1Chairs, informally invitingoverseas speakers inorganising
symposiums and workshops at ASM Annual Scientific Meetings.
ASIG continues to review submissions and comments from other
Medical andAntimicrobialGroups in regards to antimicrobial issues,
e.g. Royal College of Pathologists Australasia Advisory Committee,
AIMS, NATA, NPAAC, with the ASM Clinical Standing Committee
when required. ASIG continues support of teaching and research
activities where possible by seeking speakers for events. ASIG
promotes theadvancementofASMSIGantimicrobials bypresenting
lectures/visits to Universities and Colleges nationally and interna-
tionally as required.
ASIG is involved with the Continuing Education Program, involved
with the AIMS APACE Official Certificate of Attendance is given out
at all ASIG workshops at National meetings. These certificates are
important for professional developments and career advancements
in industry. As the convenor I thank all our members and sponsors
for their continued support.
ASMAffairs
40 10.1071/MA15015 MICROBIOLOGY AUSTRALIA * MARCH 2015

Annual Scientific Meeting and Trade Exhibition
[email protected]://asmmeeting.theasm.org.au/
12-15 July 2015QT CANBERRA1 London Circuit, Canberra
Rubbo Oration Janet Jansson, Pacific Northwest National Laboratory, USABazeley Oration Yoshihiro Kawaoka, University Wisconsin -Madison, USA, University Tokyo, JapanPlenary Speakers Chantal Abergel, CNRS-AMU, France Judith Berman, Tel Aviv University, Israel Ed DeLong, MIT, USA Jorge Galan, Yale University, USA Stefan Schwarz, Friedrich-Loeffler-Institut, Germany
Public Lecture: From Guts to Great Oceans Janet Jansson, Mike Manefield, Ed DeLong
Workshops Antimicrobials Bioinformatics for microbial ecology Imaging in microbiology Methods in microbial proteomics and metabolomics Women in leadership
EduCon: Microbiology Educators’ Conference preceding asm2015.
Joint Mycology Meeting with the Australasian Mycological Society, July 15-16.

© 2015 Hologic, Inc. All rights reserved. Hologic, Science of Sure, Aptima, Panther and associated logos are trademarks and/or registered trademarks of Hologic, Inc. and/or its subsidiaries in the United States and/or other countries. This information is intended for medical professionals and other markets and is not intended as a product solicitation or promotion where such activities are prohibited. Because Hologic materials are distributed through websites, eBroadcasts and tradeshows, it is not always possible to control where such materials appear. For specific information on what products are available for sale in a particular country, please contact your local Hologic representative or write to [email protected] *Pending listing on the Australian Register of Therapeutic Goods (ARTG)
Gen-Probe Australia Pty Ltd. ABN 91 150 687 773 . Suite 402, Level 4, 2-4 Lyon Park Road, MacquariePark, NSW, Australia, 2113 Phone: +61 2 9888 8000 Fax: (02) 9870 7555
hologic.com | [email protected] | 1800 264 073 (Australia) | [email protected] | 0800 804 904 (New Zealand)
Aptima Combo 2® assay | Aptima CT assay | Aptima GC assay | Aptima Trichomonas vaginalis assay
Aptima HPV assay | Aptima HPV 16 18/45 Genotyping assay
Aptima HIV-1 Quant Dx Assay*
The highly evolvedsenses of a hunter
With continuous, random access
and no-batch processing the
Panther® system, along with our
highly sensitive and specific
Aptima® assays, provide
on-demand performance for a
wide range of infectious diseases.
aasa say | Aptima CT assay | Aptima GC assay | Aptima Trichomonas vaginalis assay
Aptima HPV assay | Aptima HPV 16 18/45 Genotyping assayAptima HPV assay | Aptima HPV 16 18/45 Genotyping assay
Aptima HIV-1 Quant Dx Assay*
Copyright © 2022 FDOKUMEN




















