
Syllabus M. Tech.: Electrical Engineering Specialization in ...

17 (1): 37-46 (2010)
Among mammals, phyllostomid bats have undergone the largest known adaptive radiation with respect to feeding habits and habitat use (Gould, 1976). Originating from an
insectivorous ancestor (Ferrarezzi & Gimenez, 1996), mem-bers of the family Phyllostomidae exploit a wide range of food sources, including nectar, pollen, leaves, fruit, seeds, arthropods, and small vertebrates, as well as the blood of larger vertebrates (Gardner, 1976; Kunz & Diaz, 1994; Nogueira & Peracchi, 2003). Endemic to the New World tropical and subtropical zones, they represent the second largest tropical family of Chiroptera. Currently, 160 species
Specialization and omnivory in diverse mammalian assemblages1
Katja REX2, Leibniz Institute for Zoo and Wildlife Research, Alfred-Kowalke-Str. 17, 10315 Berlin,
Germany, e-mail: [email protected]
Benjamin I. CZACZKES, The Hebrew University of Jerusalem, Programming Instruction Unit,
Givat Ram, Jerusalem 91904, Israel.
Robert MICHENER & THOMAS H. KUNZ, Center for Ecology and Conservation Biology,
Department of Biology, Boston University, Boston 02215, Massachusetts, USA.
Christian C. VOIGT, Leibniz Institute for Zoo and Wildlife Research, Alfred-Kowalke-Str. 17,
10315 Berlin, Germany.
Abstract: Phyllostomid bats form some of the most speciose mammalian assemblages known, with more than 70 species estimated to co-occur on one site. These species encompass a dietary spectrum that ranges across several trophic levels, and many show morphological specializations for their dietary behaviour (e.g., long tongues in nectar-feeding bats). However, previously reported diets have varied among studies including assignments of species to feeding ensembles. In this study we present data that demonstrate that phyllostomid bats are opportunistic omnivores despite their specializations. We analyzed the diet of 67 phyllostomid bat species from the Neotropics based on both fecal analyses and nitrogen isotope ratios in wing tissue and found that most species complemented their primary diet with nutrients from many different food sources. From these new dietary data, we were unable to distinguish meaningful feeding ensembles of species based on diet. Thus, we argue that interspecific competition is not restricted to species within an ensemble but seems instead to occur across the entire assemblage. Our results suggest that phyllostomid species have specialized successively on distinct diets during their radiation without sacrificing their capability to exploit a variety of food types. The combination of morphological and behavioural specialization on the one hand and opportunistic omnivory on the other might promote the high diversity of phyllostomid bat assemblages. Keywords: adaptive radiation, bats, ensembles, fecal analysis, nitrogen isotope analysis, trophic levels.
Résumé : Les chauves-souris de la famille des Phyllostomidés forment l’un des assemblages connus les plus riches chez les mammifères avec plus de 70 espèces qui peuvent cohabiter sur un site. Ces espèces présentent un vaste spectre de régimes alimentaires couvrant plusieurs niveaux trophiques et plusieurs espèces possèdent des spécialisations morphologiques liées à leur comportement d’alimentation (par exemple, les chauves-souris se nourrissant de nectar ont de longues langues). Cependant, les diètes identifiées dans les études précédentes variaient entre elles incluant le classement des espèces dans les différents regroupements de régimes alimentaires. Dans cette étude, nous présentons des données qui démontrent que les chauves-souris phyllostomidés sont des omnivores opportunistes malgré leurs spécialisations. Nous avons analysé le régime alimentaire de 67 espèces de chauve-souris phyllostomidés néotropicales en nous basant sur des analyses fécales et des ratios d’isotopes de l’azote dans le tissu des ailes. Nous avons constaté que la plupart des espèces complètent leur régime de base avec des nutriments provenant de différentes sources alimentaires. De ces nouvelles données sur l’alimentation, nous avons été incapables de distinguer des regroupements cohérents d’espèces basés sur leur régime alimentaire. Ainsi, nous soutenons que la compétition interspécifique n’est pas limitée aux espèces au sein d’un même groupe, mais semble plutôt se produire dans tout l’assemblage d’espèces. Nos résultats suggèrent que les espèces de phyllostomidés se sont spécialisées successivement pour des régimes alimentaires distincts durant leur radiation sans sacrifier leur capacité à exploiter une variété de types de nourriture. La combinaison de spécialisations morphologiques et comportementales d’une part et une ominvorie opportuniste d’autre part favorise la grande diversité d’assemblages chez les chauves-souris phyllostomidés. Mots-clés : analyse d’isotopes de l’azote, analyse fécale, chauves-souris, niveaux trophiques, radiation adaptative, regroupements.
Nomenclature: Timm & LaVal, 1998; Albuja, 1999; Tirira, 1999; Baker, 2002; Muchhala, Mena & Albuja, 2005; Sánchez-
Hernandez, Romero-A. & Schnell, 2005.
Introduction
1Rec. 2009-06-15; acc. 2009-12-21. Associate Editor: Doug Morris.2Author for correspondence.DOI 10.2980/17-1-3294

Rex et al.: Phyllostomid diveRsity and feeding habits
3838
of phyllostomids are recognised (Simmons, 2005), making them almost as species-rich as the Old World plant-visiting bats (Pteropodidae, 186 species), which occur throughout the rainforests of Africa, Asia, Madagascar, Indonesia, New Guinea, and Australia and the deserts and savannahs of Africa and adjacent regions. Phyllostomid bats form the most species-rich assemblages of all mammalian families, with an estimate of more than 70 species co-occurring at one site (Rex et al., 2008). However, the mechanisms that have promoted and maintained this high local and regional diversity are not yet fully understood.
It has been suggested that phyllostomid species are highly specialized on specific food types, allowing them to efficiently exploit resources and to avoid dietary over-lap with other species (Dumont, 2003; 2004). As a con-sequence, phyllostomid species have been grouped into ensembles (sensu Fauth et al., 1996) according to their diet and foraging modes. This classification is based on the assumption that exploitation of different food sources is a consequence of resource partitioning within assem-blages and that species of the same ensemble exploit similar resources in a similar manner (Magurran, 2004). Previous researchers have concluded that species of the same ensem-ble have a higher potential for interspecific competition than do species of different ensembles (Moreno, Arita & Solis, 2006). However, there is no consensus as to how many and which ensembles of phyllostomid bat assem-blages exist, nor is there full agreement on diets of species or their assignments to specif ic ensembles (Fleming, Hooper & Wilson, 1972; Wilson, 1972; Gardner, 1976; Bonaccorso, 1978; Humphrey, Bonaccorso & Zinn, 1983; Norberg & Rayner, 1987; Willig, Camillo & Noble, 1993; Kalko, Handley & Handley, 1996; Herrera, Fleming & Sternberg, 1998; Kalko, 1998; Giannini & Kalko, 2004; Moreno, Arita & Solis, 2006). Previous studies have shown that most phyllostomid species include both plants and animals in their diet (Fleming, Hooper & Wilson, 1972; Gardner, 1976; Bonaccorso, 1978; Humphrey, Bonaccorso & Zinn, 1983; Ferrarezzi & Gimenez, 1996), although the proportions of these components may vary among species. By these accounts, phyllostomids appear to be omnivor-ous. Using multivariate statistics, Humphrey, Bonaccorso, and Zinn (1983) showed that 9 species of the subfamily Phyllostominae can be categorized into one ensemble, gleaning omnivores. But with respect to the differences among ensembles suggested in other studies (Fleming, Hooper & Wilson, 1972; Bonaccorso, 1978; Kalko, 1998; Matt, 2001; Giannini & Kalko, 2004), there is as yet no statistical support that ensembles do indeed differ signifi-cantly in diet, although the concept of ensembles has been used to draw conclusions about interspecific competition for food in bat assemblages.
Phyllostomid diets have been analyzed by fecal analy-ses, both microscopical (Bonaccorso, 1978; Humphrey, Bonaccorso & Zinn, 1983; Fleming, 1986; 1991; Matt, 2001; Giannini & Kalko, 2004) and using molecular gen-etics (Clare et al., 2009), by analysis of stomach contents (Fleming Hooper & Wilson, 1972), by examination of culled and expelled food parts (Kunz & Diaz, 1994; Dumont,
2003) and the use of nitrogen isotopes (Fleming, 1995; Herrera, Fleming & Sternberg, 1998; Herrera et al., 2001; 2002; Voigt et al., 2003). Fecal analyses of bats are very specific as to the identity of fruit or insect order consumed and also can indicate nectarivory based on the presence of pollen even though nectar itself is not traceable in feces. Thus, fecal samples are a good indicator of dietary overlap among species (Fleming, 1986). They also yield informa-tion about the dietary components used for purposes other than synthesis of body tissue, such as locomotion, that cannot be detected with isotope analyses in bulk samples. However, analyses of feces and culled parts (e.g., insect wings, expelled leaf pellets) at best represent snapshots of an animal’s diet, covering the most recently consumed items.
Nitrogen isotope analyses yield information on dietary components that were assimilated into the body tissue over the previous 1-3 months, depending on the type of tissue (Voigt etal., 2003; Voigt & Matt, 2004; Miron etal., 2006). The lighter 14N isotope is more abundant in nature than the heavier 15N isotope. Enzymatic processes in metabol-ism favour the naturally more abundant 14N when excret-ing nitrogen, which leads to an enrichment of 15N in the organism’s body tissue of about 3-5‰ in relation to the diet (DeNiro & Epstein, 1981; Voigt etal., 2006). This enrich-ment occurs with each trophic level along the food chain. Thus, the 15N/14N ratio (noted as d15N when set in relation to air) of an animal’s body tissue indicates its trophic level, allowing a classification of species into plant-eating, insec-tivorous, or carnivorous species. In contrast to fecal analyses, d15N yields information about the portion of food items assimilated into the body tissue rather than what the animal expels or excretes (DeNiro & Epstein, 1981; Bearhop etal., 2004; Voigt etal., 2006).
We analysed fecal samples and nitrogen isotope ratios in wing tissues of 67 phyllostomid species from 3 Neotropical assemblages to shed light on resource util-ization in assemblages with the highest local diversity observed for a mammalian family. Based on the large variety of food items reported for phyllostomid species, we hypothesized that phyllostomids are highly flexible in their food choices and are capable of adjusting to differ-ent resource availabilities. Resource abundances, espe-cially regarding fruit and flowers, differ between sites and also show seasonal variation. Thus, we expected to find regional as well as seasonal differences in the diet of phyllostomid bats. Because fruit and nectar are high in carbohydrates but low in protein, these dietary components are not fully reflected in the nitrogen isotope values, but they can be readily detected in feces in the form of seeds and pollen. Insects are high in protein and low in digest-ible carbohydrates: thus, insect components in the diet should be reflected in the nitrogen isotope values even if they make up a small proportion of an animal’s diet and might therefore be missed in fecal analyses. Therefore, we expected that fecal samples would contain a higher propor-tion of plant products, whereas nitrogen isotope analyses of wing tissue would indicate a more animal-biased diet. We hypothesized that phyllostomid species are opportunistic omnivores and thus cannot be categorized easily into dis-tinct feeding ensembles.

Methods
study sites
We collected data at 3 study sites: La Selva Biological Station (LS, 10° 25' 52" n, 84°' 00' 12" w) in Costa Rica, the Tiputini Biodiversity Station (TBS, 0° 38.31' s, 76° 8.92' w) in Ecuador, and the Bombuscaro River in the Podocarpus National Park (BOM, 4° 1' s, 79° 1' w) in Ecuador. LS is located in the Caribbean lowland of Costa Rica at an elevation of 30-100 m above sea level and covers 1600 ha of protected forest, including about 600 ha of old-growth forest contiguous with the Braulio Carillo National Park (46 000 ha). Habitat types include primary pre-montane forest, primary and secondary terra firme, and seasonally flooded forest (varzea), including areas of different succes-sional stages and abandoned plantations with adjacent pastures (Hartshorn & Hammel, 1994). Monthly rainfall averages 333 mm, with a drier period from February to April and rain-fall peaks (more than 400 mm per month) from June to July and from November to December (Sanford etal., 1994).
TBS is located in the Orellana Province in eastern Ecuador and is surrounded by 1.5 million ha of primary Amazonian lowland rainforest. Habitat types encompass terra firme forest, varzea forest, palm swamps, small oxbow lakes, and streams, with elevation ranging from 190 to 270 m above sea level (Kreft etal., 2004). Monthly rainfall varies between 50 and 650 mm, with heaviest rainfall from May to August and a dry period from December to April (TBS weather station).
BOM is located on the eastern slope of the Andean mountains of southern Ecuador adjacent to the Podocarpus National Park (146 200 ha). The study area is situated along the Bombuscaro River between 900 and 1200 m elevation on slopes ranging from 40° to 60°. The forest is dominated by lower montane, broad-leaf vegetation (Bussmann, 2001) consisting of old-growth forest and areas at different stages of succession after land slides and is adjacent to pasture land. Monthly rainfall varies between 150 and 200 mm without a pronounced dry season (Emck, 2005).
samPling PRotocol
Bats were captured in mist-nets from October to December 2004 and February to April 2005 at LS, September to November 2004, and April to June 2005 at TBS, and September to November 2004 and February to April 2005 at BOM. We set up at total of 36 × 2.5-m ground nets (70 denier/2 ply, 16-mm mesh, 5 shelves, R. Vohwinkel, Velbert, Germany) across trails and one 9- × 3-m canopy net (CAN-BH9 nylon mist-nets, 50 denier/2 ply, 38-mm mesh, 9 shelves, Avinet, Dryden, New York, USA) at 10 arbitrarily chosen 1-ha plots for approximately 40 full nights at each site (20 nights for each of the 2 study periods at each site). On each plot we randomly col-lected 10 mostly herbivorous insects (Caelifera, Ensifera, Hymenoptera, Lepidoptera) as well as leaf material and, where possible, fruit from at least 15 plants at each plot.
Bats were identified according to Timm and LaVal (1998) and Baker (2002) for Costa Rican species and sev-eral taxonomic keys and publications for Ecuadorian spe-cies (Albuja, 1999; Tirira, 1999; Muchhala, Mena & Albuja, 2005; Sánchez-Hernandez, Romero-A. & Schnell, 2005).
Artibeus planirostris was treated as distinct from Artibeusjamaicensis according to Lim et al. (2004). Subfamily designation for all other species was based on Simmons (2005). Bat species were provisionally grouped into frugi-vore, nectarivore, insectivore, and carnivore ensembles fol-lowing Rex etal. (2008). Bats were banded with a coloured and numbered plastic split ring (Hughes Ltd., London, UK) on the right (males) or left (females) forearm to establish individual identity. After capture, bats were placed into individual cotton bags and left there for up to 1 h for defeca-tion. Fecal samples were taken from bags immediately and preserved in 70% ethanol. All bats were processed shortly after capture and released at the site of capture as soon as possible (typically within less than 2 h).
analyses and statistics We analyzed the volume percentage of plant material,
insect parts, bones, and other particles in fecal samples according to Whitaker, McCracken, and Siemers (2009) and Voigt, Bradley, and Ortmann (2009). The presence of pollen in feces was noted but not included in the analysis as we could not determine whether the bat had derived it from the flower itself or from pollen-carrying insects and the vol-ume percentage of pollen in the feces was extremely small. Fecal samples were analyzed using a blind protocol, and samples consisting of 100% plant material were assigned to tropic level 1, trophic level 2 consisting of 100% insect parts and trophic level 3 consisting of 100% vertebrate remains (mostly bones). Intermediate values were assigned according to the proportion of plant, insect, and vertebrate material present by volume. The presence of hair was noted but not included in the calculation of trophic level (TLfeces) because hair in feces can be attributed as much to grooming as to diet.
Wing tissue samples were collected from each bat using a sterile 3-mm biopsy punch. These tissue samples were dried at 50 °C for 3 d, weighed on a Sartorius microbalance (Sartorius AG, Göttingen, Germany), and loaded in tin capsules. Plant and insect material was dried, ground, and homogenized, and 1- to 2-mg subsamples were loaded in tin capsules. For plant species from which both leaves and fruits were available, we tested for differences in nitrogen isotope ratios between fruit and leaf tissue from the same individual plant. All samples were combusted and analyzed using a Fisons NA 1500 elemental analyzer and a Finnigan continuous flow system, coupled to a Delta-S isotope ratio mass spectrometer at Boston University’s Stable Isotope Laboratory. Atmospheric nitrogen was used as the standard for the 15N/14N ratio, and nitrogen isotope ratios are given in the d15N notation (‰).
Equation [1] was used to calculate d15N of all analyzed material.
[1]
Rsample and Rstandard represent the 15N/14N ratios of the sample and standard, respectively.
Isotope data were converted into trophic levels (TLnitrogen) according to equation [2].
[2]
Écoscience, vol. 17 (1), 2010
39
d15N = (Rsample / Rstandard – 1) 1000
TLnitrogen = (d15Nbat – d15Nplant)/Dtrophic_enrichment
Rex et al.: Phyllostomid diveRsity and feeding habits
40
where d15Nbat is the 15N/14N ratio of the bat’s wing tissue, d15Nplants is the average 15N/14N ratio of all collected plant material, and D trophic_enrichment is the enrichment of d15N between 2 trophic levels (e.g., from plant to insect).
To determine D trophic_enrichment we averaged the enrichment in d15N from plant to insect, from insects to non-phyllostomid insectivorous bats, and from plants to herbivorous bats (i.e., bats with d15N < 7‰) separately for each site and season.
We compared the trophic level of each bat species calculated from a) fecal analyses (TLfeces) and b) the d15N value of its wing tissue (TLnitrogen) using a Wilcoxon matched-pairs test.
The effects of species, site, and season on TLnitrogen were analyzed using a three-way ANOVA. We tested for differences in trophic level among ensembles (ANOVA) and ran a null-model simulation to determine if these differen-ces would also occur if species were assigned to ensembles randomly. For this simulation, the original data set was rep-licated 1000 times. For each resulting data set the various species were assigned randomly to 1 of 4 ensembles (frugiv-orous, nectarivorous, insectivorous, carnivorous [including sanguinivorous]), keeping the number of species per ensem-ble the same as in the original data. Each of the resulting data sets was analyzed using the same ANOVA procedure as for the original data set.
Species abundance data at all sites are summarized in Rex etal. (2008). Statistics were calculated using GraphPad InStat 3 (1992) and SPSS 12.0 (2003), and all tests were two-tailed with accepted significance levels of P ≤ 0.05. Results are stated as means ± 1 standard deviation.
Results
isotoPe analyses
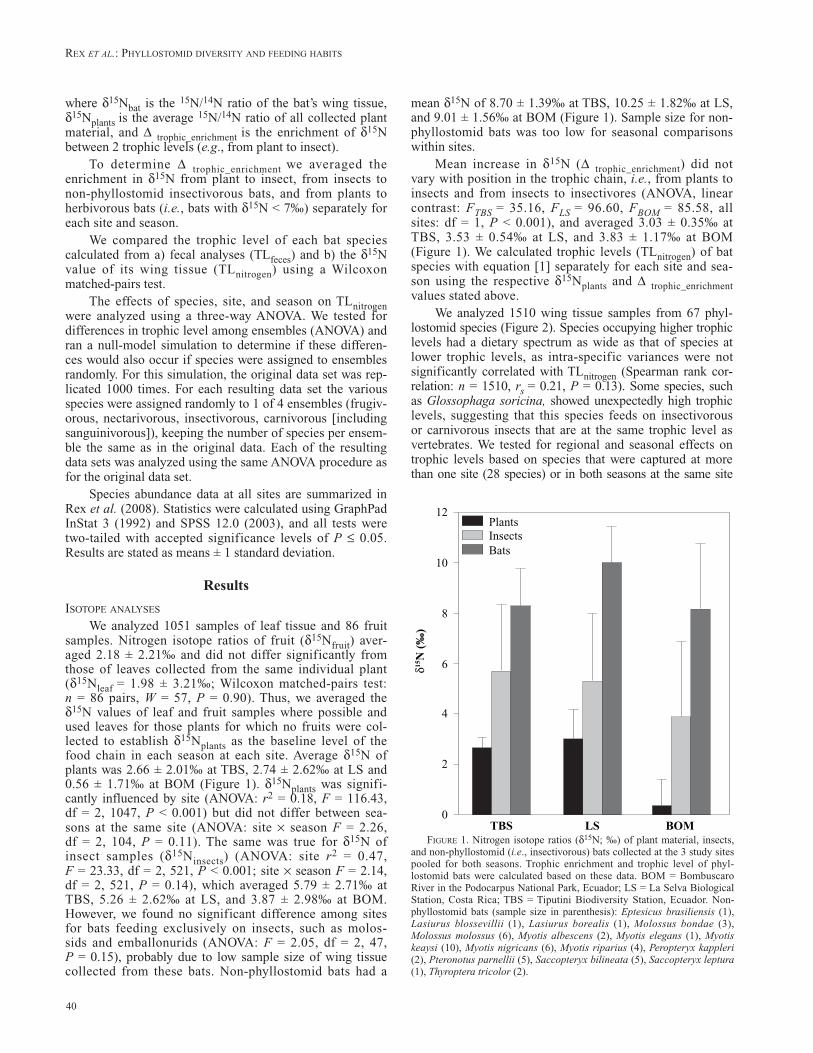
We analyzed 1051 samples of leaf tissue and 86 fruit samples. Nitrogen isotope ratios of fruit (d15Nfruit) aver-aged 2.18 ± 2.21‰ and did not differ significantly from those of leaves collected from the same individual plant (d15Nleaf = 1.98 ± 3.21‰; Wilcoxon matched-pairs test: n = 86 pairs, W = 57, P = 0.90). Thus, we averaged the d15N values of leaf and fruit samples where possible and used leaves for those plants for which no fruits were col-lected to establish d15Nplants as the baseline level of the food chain in each season at each site. Average d15N of plants was 2.66 ± 2.01‰ at TBS, 2.74 ± 2.62‰ at LS and 0.56 ± 1.71‰ at BOM (Figure 1). d15Nplants was signifi-cantly influenced by site (ANOVA: r2 = 0.18, F = 116.43, df = 2, 1047, P < 0.001) but did not differ between sea-sons at the same site (ANOVA: site × season F = 2.26, df = 2, 104, P = 0.11). The same was true for d15N of insect samples (d15Ninsects) (ANOVA: site r2 = 0.47, F = 23.33, df = 2, 521, P < 0.001; site × season F = 2.14, df = 2, 521, P = 0.14), which averaged 5.79 ± 2.71‰ at TBS, 5.26 ± 2.62‰ at LS, and 3.87 ± 2.98‰ at BOM. However, we found no significant difference among sites for bats feeding exclusively on insects, such as molos-sids and emballonurids (ANOVA: F = 2.05, df = 2, 47, P = 0.15), probably due to low sample size of wing tissue collected from these bats. Non-phyllostomid bats had a
mean d15N of 8.70 ± 1.39‰ at TBS, 10.25 ± 1.82‰ at LS, and 9.01 ± 1.56‰ at BOM (Figure 1). Sample size for non-phyllostomid bats was too low for seasonal comparisons within sites.
Mean increase in d15N (D trophic_enrichment) did not vary with position in the trophic chain, i.e., from plants to insects and from insects to insectivores (ANOVA, linear contrast: FTBS = 35.16, FLS = 96.60, FBOM = 85.58, all sites: df = 1, P < 0.001), and averaged 3.03 ± 0.35‰ at TBS, 3.53 ± 0.54‰ at LS, and 3.83 ± 1.17‰ at BOM (Figure 1). We calculated trophic levels (TLnitrogen) of bat species with equation [1] separately for each site and sea-son using the respective d15Nplants and D trophic_enrichment values stated above.
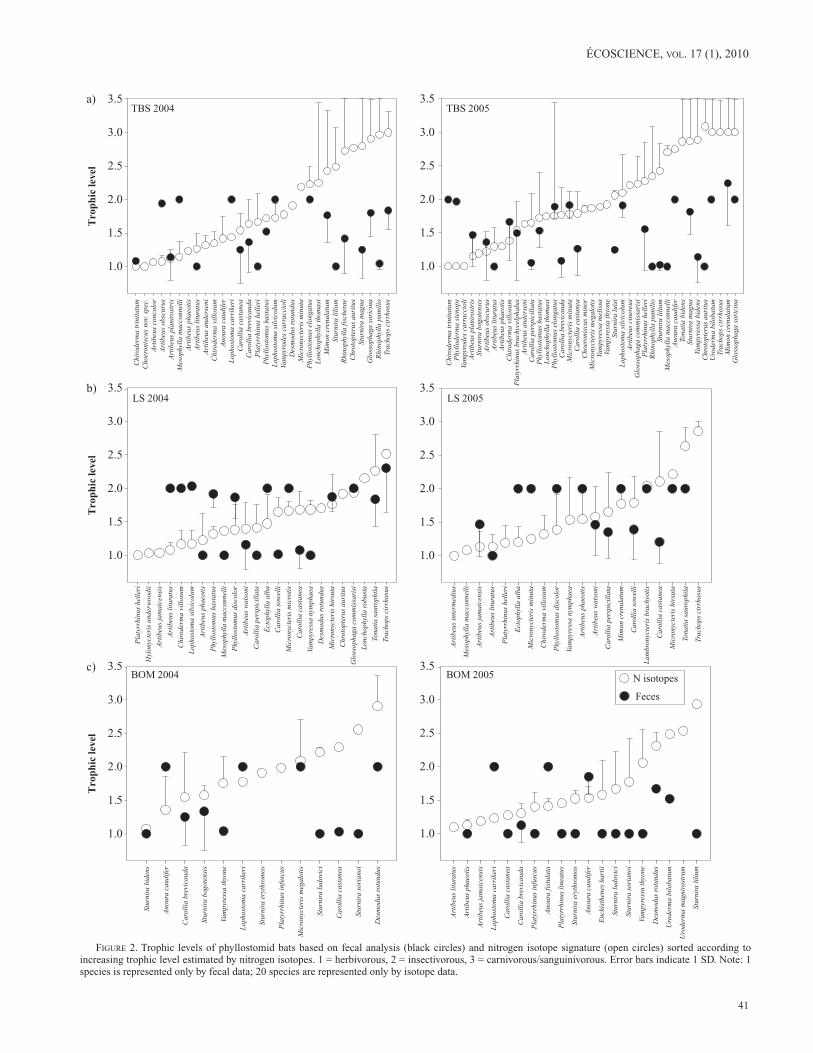
We analyzed 1510 wing tissue samples from 67 phyl-lostomid species (Figure 2). Species occupying higher trophic levels had a dietary spectrum as wide as that of species at lower trophic levels, as intra-specific variances were not significantly correlated with TLnitrogen (Spearman rank cor-relation: n = 1510, rs = 0.21, P = 0.13). Some species, such as Glossophaga soricina, showed unexpectedly high trophic levels, suggesting that this species feeds on insectivorous or carnivorous insects that are at the same trophic level as vertebrates. We tested for regional and seasonal effects on trophic levels based on species that were captured at more than one site (28 species) or in both seasons at the same site
figuRe 1. Nitrogen isotope ratios (d15N; ‰) of plant material, insects, and non-phyllostomid (i.e., insectivorous) bats collected at the 3 study sites pooled for both seasons. Trophic enrichment and trophic level of phyl-lostomid bats were calculated based on these data. BOM = Bombuscaro River in the Podocarpus National Park, Ecuador; LS = La Selva Biological Station, Costa Rica; TBS = Tiputini Biodiversity Station, Ecuador. Non-phyllostomid bats (sample size in parenthesis): Eptesicus brasiliensis (1), Lasiurus blossevillii (1), Lasiurus borealis (1), Molossus bondae (3), Molossus molossus (6), Myotis albescens (2), Myotis elegans (1), Myotiskeaysi (10), Myotisnigricans (6), Myotisriparius (4), Peropteryxkappleri (2), Pteronotusparnellii (5), Saccopteryxbilineata (5), Saccopteryxleptura (1), Thyropteratricolor (2).
Écoscience, vol. 17 (1), 2010
41
figuRe 2. Trophic levels of phyllostomid bats based on fecal analysis (black circles) and nitrogen isotope signature (open circles) sorted according to increasing trophic level estimated by nitrogen isotopes. 1 = herbivorous, 2 = insectivorous, 3 = carnivorous/sanguinivorous. Error bars indicate 1 SD. Note: 1 species is represented only by fecal data; 20 species are represented only by isotope data.

Rex et al.: Phyllostomid diveRsity and feeding habits
42
(42 species). The three-way interaction site × species × season and the main effect of season were not statistically signifi-cant (ANOVA: F = 1.11, df = 10, 1355, P > 0.35; F = 1.8, df = 1, 1355, P > 0.18, respectively). The two-way interactions site × species, site × season, and season × species were all significant (ANOVA: F = 2.599, df = 26, 1355, P < 0.001; F = 6.3, df = 2, 1355, P < 0.01; F = 2.638, df = 42, 1355, P < 0.001, respectively), as were the main effects for spe-cies (ANOVA: F = 9.427, df = 63, 1355, P< 0.001) and site (ANOVA: F = 7.7771, df = 2, 1355, P < 0.001). Body mass ranged from 5 g in the smallest species (Ectophyllaalba) to over 100 g in the largest (Phyllostomushastatus) but was not related to trophic level (TLnitrogen) (Spearman rank correlation: rs = –0.06, n = 63, P > 0.60).
fecal analyses
We obtained and analyzed 881 fecal samples. Samples consisting entirely of hair were excluded from further analy-ses, resulting in 787 fecal samples from 44 species being retained in our analysis (Figure 2). The dietary spectrum reflected by fecal analyses was narrower for species for higher trophic levels as variance within species was nega-tively correlated with TLfeces (Spearman rank correlation: n = 787, rs = –0.48, P = 0.004). Species with a trophic level of 2 consumed insects rather than equal proportions of fruit and vertebrates, as vertebrate remains were only found in species whose average trophic level is larger than 2. Sample size was not sufficient to calculate the three-way interaction species × site × season. The two-way interactions spe-cies × site and site × season were not significant (ANOVA: F = 1.397, df = 8, 711, P = 0.20 and F = 0.848, df = 2, 711, P = 0.43, respectively). Likewise the main effects of site and season were not signif icant (ANOVA: F = 1.895, df = 2, 711, P = 0.15 and F = 0.125, df = 1, 711, P = 0.72, respectively). The two-way interaction species × season and the main effect of species on TLfeces were significant (ANOVA: F = 1.722, df = 23, 711, P = 0.02 and F = 3.468, df = 44, 711, P < 0.001, respectively). Species-specific trophic levels derived from fecal analysis (TLfeces) were not related to the mean body mass of these species (Spearman rank correlation: rs = –0.06, n = 44, P = 0.70).
comPaRison of fecal and isotoPe analyses
Many species showed a high variability in trophic level as determined by nitrogen isotope ratios (mean SD of TLnitrogen within species: 0.36) and by fecal analyses (mean SD of TLfeces within species: 0.28), indicating a wide spectrum of food sources (Figure 2). Estimated values of TLfeces were significantly lower than those of TLnitrogen at TBS and BOM in both seasons (Wilcoxon test: BOM2004: n = 11, W = 48, P = 0.03; BOM2005: n = 16, W = 86, P = 0.025; TBS2004: n = 19, W = 108, P = 0.03; TBS2005: n = 26, W = 234, P = 0.002). Isotope analyses indicated 43–56% less plant material in the bat’s diet (i.e., a 0.43 to 0.56 increase in trophic level) than did fecal samples (Figures 2a and c). However, there was no significant dif-ference between TLnitrogen and TLfeces in either season at LS (Wilcoxon test: LS2004: n = 18, W = –16, P = 0.73; LS2005: n = 14, W = 5, P = 0.90; Figure 2b).
While both TLnitrogen and TLfeces were influenced sig-nificantly by the traditional assignment of species to ensem-bles (ANOVA TLnitrogen-ensembles: r2 = 0.22, P < 0.001;
TLfeces-ensembles: r2 = 0.26, P < 0.001), the same was true when species were randomly assigned to ensembles (ANOVA TLrandom-ensembles [average over 1000 runs]: r2 = 0.05, P < 0.001 for both calculations).
Discussion
comPaRing methodologies
Nitrogen isotope ratios of plants can differ substantially between sites (Figure 1). This may result from environmental variability, including different levels of nitrogen in the soil, differences in nitrogen uptake, or different mechanisms of nitrogen fixation in the ecosystem. Because plants are at the base of food webs, this base value is crucial for mak-ing regional comparisons among assemblages. Similarly, enrichment factors may vary due to regional differences in the nitrogen availability in soil. A high nitrogen abundance in the ecosystem might lead to higher enrichment factors along the trophic cascade as organisms excrete more nitro-gen, thereby increasing enzymatic fractionation. Thus, it is important to calculate trophic enrichment before drawing conclusions about trophic levels of organisms based on their d15N values.
Fecal analyses showed a higher proportion of plant material in the diet than was indicated by the nitrogen iso-tope ratio of body tissue for both of our Ecuadorian study sites (Figures 2a and c), but there was no difference at our Costa Rican study site (Figure 2b). If pollen had been included in the analysis, fecal analyses would have yielded even lower trophic levels. We assume that both methods underestimate the importance of plant-based resources in the diet of bats. Many stenodermatine species ingest only the juice of fruits, while spitting out seeds and pulp (Kunz & Diaz, 1994; Dumont, 2003), whereas glossophagine bats consume nectar as their main carbohydrate source (Helverson & Winter, 2003). These liquid plant components can be detected neither in feces nor in the nitrogen isotope ratio of tissue samples. Fruit and nectar are rich in carbohy-drates and are therefore useful to fuel locomotion and other energy-consuming behaviours in bats (Voigt & Speakman, 2007). Insects and vertebrates are good protein sources and are most likely used by bats to satisfy their protein requirements for growth, tissue synthesis, and reproduction. Apparently, many bat species complement a frugivorous diet with animal protein, as analysis of their body tissue reflected the protein intake from animal sources. In contrast, fecal samples showed a high proportion of seeds, suggesting that bats consume large quantities of carbohydrate-rich fruit, possibly in order to meet the high energy requirements of these small flying endotherms. Alternatively, herbivorous bats may over-ingest carbohydrates in order to meet protein requirements when eating a protein-poor diet. Thus, it is important to compare the results of fecal and nitrogen iso-tope analysis to get a full picture of the diet of a species.
dietaRy flexibility
Our results indicate that phyllostomid species are highly flexible in their dietary habits (Figure 2). Despite morpho-logical adaptations that suggest preferences for certain food types (Freeman, 2000; Dumont, 2003), species appear to be opportunistic feeders for at least part of the time. The trophic
Écoscience, vol. 17 (1), 2010
43
levels of phyllostomid species were influenced by capture site as well as season, indicating that these bats readily adjust their diets to local and seasonal changes in resource avail-ability. On a finer scale, similar results have been found in previous studies. Fleming (1986) hypothesized that the diet-ary spectrum of bats should be as broad as possible given the availability of local resources. His study focused on the variety of plant remains represented in the feces of 12 phyl-lostomid bat species from 4 rainforest sites and on the degree of selectivity of the respective species. Fleming concluded that phyllostomid bats are generalists that show a varying degree of selectivity depending on spatio-temporal resource availability and that they complement their preferred fruits opportunistically with fruits from other plants. Our results support his hypothesis and extend it beyond the restriction to a single trophic level. Our results indicate that phyl-lostomids are opportunistic feeders that include dietary items from several trophic levels. This was especially obvious in the genus Carollia, known as Piper specialists and consist-ently grouped in the understory frugivorous ensemble in other studies (Bonaccorso, 1978; Kalko & Handley, 2001; Bonaccorso etal., 2007). This genus showed a much higher (60–70%) level of insect-derived nitrogen in its wing tis-sue than one would expect in a strictly frugivorous species (Figures 2 and 3). Herrera etal. (2002) found that C.brevi-caudafeeds on insects only for a few months a year, whereas our data indicate that the 3 Carollia species in our study include large proportions of insects in their diet both in the rainy and the dry season, a result that is also supported by the findings of York and Billings (2009). The high proportion of insect protein in the diet of Carollia was particularly surpris-ing as the Piper fruit is much higher in protein than most other fruits consumed by phyllostomid bats (Fleming, 1986). In contrast, several stenodermatine species obtained most of their protein from plants even though they often fed on pro-tein-poor figs. Based on the findings of previous studies, we assume that some species, such as Artibeus, feed heavily on leaves for additional protein intake (Zortea & Mendes, 1993; Kunz & Diaz, 1994; Kunz & Ingalls, 1994), while others, such as Chiroderma, supplement their protein intake by feed-ing on seeds (Nogueira & Peracchi, 2003).
Feeding habits across trophic levels have also been observed in some tropical bird species. Like glossophagine bats, sunbirds and hummingbirds are specialized on nec-tar as their primary food source, and they also consume large quantities of insects and spiders (Feinsinger, 1976; Roxburgh & Pinshow, 2002; Yanega & Rubega, 2004). Toucans and hornbills are some of the largest flying fru-givores, but they too feed on insects and small verte-brates, while predominantly insectivorous woodpeckers feed on fruits when they are superabundant (Bühler, 1997; Shanahan et al., 2001; Primack & Corlett, 2005). The advantage of such opportunistic omnivory is the possibility of complementing a diet rich in carbohydrates with protein-rich food sources or vice versa without the need of over-ingesting one type of nutrient.
ensembles
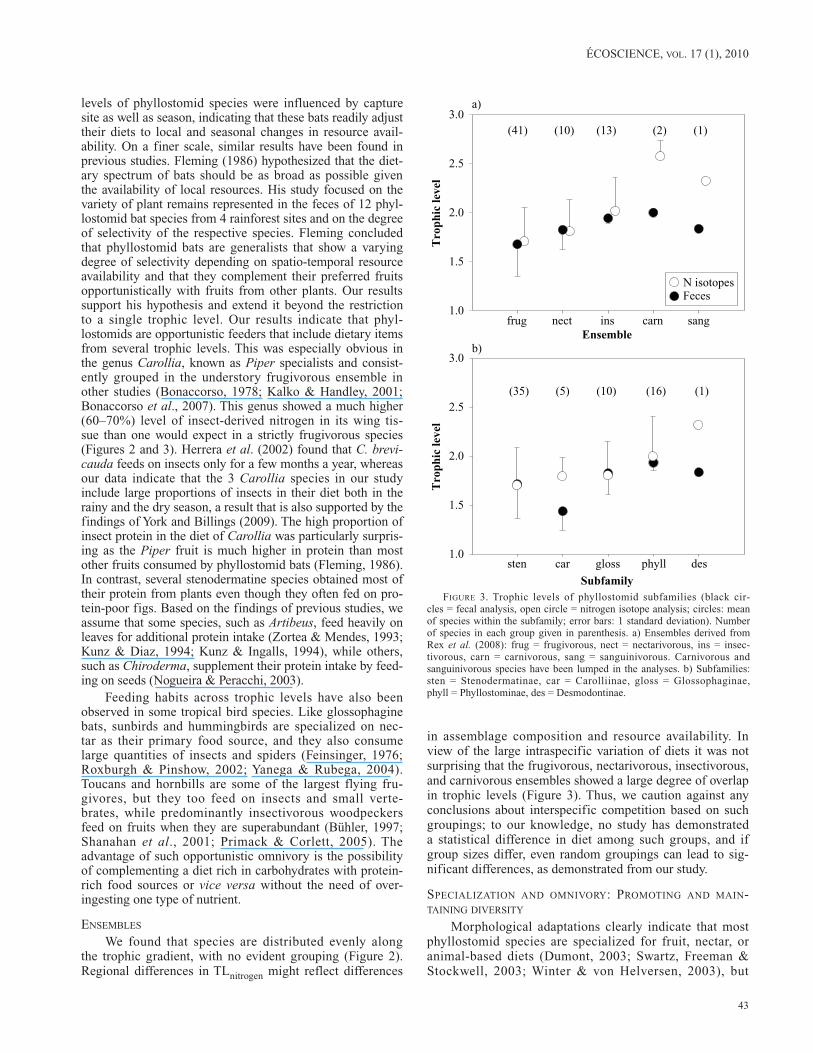
We found that species are distributed evenly along the trophic gradient, with no evident grouping (Figure 2). Regional differences in TLnitrogen might reflect differences
in assemblage composition and resource availability. In view of the large intraspecific variation of diets it was not surprising that the frugivorous, nectarivorous, insectivorous, and carnivorous ensembles showed a large degree of overlap in trophic levels (Figure 3). Thus, we caution against any conclusions about interspecific competition based on such groupings; to our knowledge, no study has demonstrated a statistical difference in diet among such groups, and if group sizes differ, even random groupings can lead to sig-nificant differences, as demonstrated from our study.
sPecialization and omnivoRy: PRomoting and main-taining diveRsity
Morphological adaptations clearly indicate that most phyllostomid species are specialized for fruit, nectar, or animal-based diets (Dumont, 2003; Swartz, Freeman & Stockwell, 2003; Winter & von Helversen, 2003), but
figuRe 3. Trophic levels of phyllostomid subfamilies (black cir-cles = fecal analysis, open circle = nitrogen isotope analysis; circles: mean of species within the subfamily; error bars: 1 standard deviation). Number of species in each group given in parenthesis. a) Ensembles derived from Rex et al. (2008): frug = frugivorous, nect = nectarivorous, ins = insec-tivorous, carn = carnivorous, sang = sanguinivorous. Carnivorous and sanguinivorous species have been lumped in the analyses. b) Subfamilies: sten = Stenodermatinae, car = Carolliinae, gloss = Glossophaginae, phyll = Phyllostominae, des = Desmodontinae.

Rex et al.: Phyllostomid diveRsity and feeding habits
44
nonetheless a generalist pattern is apparent across species. Based on our results, we argue that the high local species richness of phyllostomid bats is promoted by a combina-tion of both specialization and opportunistic generalization. Specialization allows species to exploit resources with maximal efficiency, but their ability to switch to alterna-tive food types helps them to complement their diet with additional nutrients and survive time periods when their pre-ferred food is scarce (Fleming, 1986). Thus, the 2 antipodal processes of specialization and omnivory both seem to be of importance for the observed high diversity of phyllostomid bat assemblages: specialization might have been the driving force for the rapid diversification of phyllostomids, but the generalist nature of these species might play a role in main-taining the high diversity of their assemblages.
Acknowledgements
This study would not have been possible without our field assistants (in alphabetical order): N. Bauer, B. Buchner, K. Jochum, M. Jones, D. Kelm, A. Kunz, B. Lammers, C. Neumann, F. Noriega, A. Arguero Santos, K. Sachanowizs, H. Schütt, C. Werres, K. Wiesner, and J. Zamora, to whom we are most grateful. We also wish to thank J. Guerra and the staff at Tiputini Biodiversity Station for their help in the field, D. Romo for his administrative support throughout our study at TBS, and B. Rinehart, E. Sampaio, and E. Kalko for their help with spe-cies identification. D. Fichte supported our field research and deserves special thanks. Furthermore, we wish to thank two anonymous referees for constructive comments on earlier ver-sions of this manuscript. We also wish to thank GML alfaplast GmbH Munich for providing important supplies for our field-work. This study complied with the Boston University Animal Care and Use Committee and the current laws of Costa Rica and Ecuador. We thank J. Guevarra and the Ministerio de Energía y Ambiente in Costa Rica as well as the Ecuadorian authorities for authorizing the data collection for this study. Funding for this project was provided by the Deutsche Forschungsgemeinschaft (Vo 890/7) and the Center for Ecology and Conservation Biology at Boston University.
Literature cited
Albuja, L., 1999. Murciélagos del Ecuador. Circetrónic Companía Limitada Offset, Quito.
Baker, R. J., 2002. A new Central American species from the Carollia brevicauda complex. Occasional Papers, Museum of Texas Tech University, 217: 1–12.
Bearhop, S., C. E. Adams, S. Waldron, R. A. Fuller & H. Macleod, 2004. Determining trophic niche width: A novel approach using stable isotope analysis. Journal of Animal Ecology, 73: 1007–1012.
Bonaccorso, F. J., 1978. Foraging and reproductive ecology in a Panamanian bat community. Bulletin of the Florida State Museum. Biological Sciences, 24: 359–408.
Bonaccorso, F. J., J. R. Winkelmann, D. Shin, C. I. Agrawal, N. Aslami, C. Bonney, A. Hsu, P. E. Jekielek, A. K. Knox, S. J. Kopack, T. D. Jennings, J. R. Lasky, S. A. Menesale, J. H. Richards, J. A. Rutland, A. K. Sessa, L. Zhaurova & T. H. Kunz, 2007. Evidence for exploitative competition: Comparative foraging behavior and roosting ecology of short-tailed fruit bats (Phyllostomidae). Biotropica, 39: 249–256.
Bühler, P., 1997. The visual peculiarities of the toucan’s bill and their principal biological role (Ramphastidae, Aves). Pages 305–310 in H. Ulrich (ed.). Tropical Biodiversity and Systematics. Zoologisches Forschungsinstitut und Museum Alexander Koenig, Bonn.
Bussmann, R. W., 2001. The montane forests of Reserva Biologica San Francisco (Zamora-Chinchipe, Ecuador). Die Erde, 132: 9–25.
Clare, E. L., E. E. Fraser, H. E. Braid, M. B. Fenton & P. D. N. Hebert, 2009. Species on the menu of a generalist predator, the eastern red bat (Lasiurusborealis): Using a molecular approach to detect arthropod prey. Molecular Ecology, 18: 2532–2542.
DeNiro, M. J. & S. Epstein, 1981. Influence of diet on the dis-tribution of nitrogen isotopes in animals. Geochimica et Cosmochimica Acta, 45: 341–351.
Dumont, E., 2003. Bats and fruit: An ecomorphological approach. Pages 398–429 in T. H. Kunz & M. B. Fenton (eds). Bat Ecology. University of Chicago Press, Chicago, Illinois.
Dumont, E., 2004. Patterns of diversity in cranial shape among plant-visiting bats. Acta Chiropterologica, 6: 59–74.
Emck, P., 2005. Klimadifferenzierung in Suedecuador. Ph.D. thesis. Friedrich-Alexander-Universität, Erlangen-Nürnberg.
Fauth, J. E., J. Bernardo, M. Camara, W. J. Resetarits, J. van Buskirk & S. A. McCollim, 1996. Simplifying the jargon of community ecology: A conceptual approach. American Naturalist, 147: 282–286.
Feinsinger, P., 1976. Organization of a tropical guild of nectarivor-ous birds. Ecological Monographs, 46: 257–291.
Ferrarezzi, H. & E. A. Gimenez, 1996. Systematic patterns and the evolution of feeding habits in Chiroptera (Archonta: Mammalia). Journal of Comparative Biology, 1: 75–94.
Fleming, T. H., 1986. Opportunism versus specialization: The evo-lution of feeding strategies in frugivorous bats. Pages 105–118 in A. Estrada & T. H. Fleming (eds). Frugivores and Seed Dispersal. Dr. W. Junk Publishers, Dordrecht.
Fleming, T. H., 1991. The relationship between body size, diet, and habitat use in frugivorous bats, genus Carollia (Phyllostomidae). Journal of Mammalogy, 72: 493–501.
Fleming, T. H., 1995. The use of stable isotopes to study the diets of plant-visiting bats. Symposium of the Zoological Society of London, 67: 99–110.
Fleming, T. H., E. T. Hooper & D. E. Wilson, 1972. Three Central American bat communities: Structure, reproductive cycles, and movement patterns. Ecology, 53: 555–569.
Freeman, P., 2000. Macroevolution and microchiroptera: Recoupling morphology and ecology with phylogeny. Evolutionary Ecology Research, 2: 317–335.
Gardner, A. L., 1976. Feeding habits. Pages 293–350 in R. J. Baker, J. K. Jones Jr. & D. C. Carter (eds). Biology of the New World Family Phyllostomidae, Part II. Special Publications, Museum Texas Tech University, Lubbock, Texas.
Giannini, N. P. & E. K. V. Kalko, 2004. Trophic structure in a large assemblage of phyllostomid bats in Panama. Oikos, 105: 209–220.
Gould, E., 1976. Echolocation and communication. Pages 247–292 in R. J. Baker, J. K. Jones Jr. & D. C. Carter (eds). Biology of the New World Family Phyllostomidae, Part II. Special Publications, Museum Texas Tech University, Lubbock, Texas.
GraphPad Software, 1992. GraphPad Software Inc., San Diego, California.
Hartshorn, G. S. & B. E. Hammel, 1994. Vegetation types and floristic patterns. Pages 73–89 in L. McDade, K. Bawa, H. Hespenheide & G. Hartshorn (eds). La Selva: Ecology and Natural History of a Neotropical Rain Forest. University of Chicago Press, Chicago Illinois.

Écoscience, vol. 17 (1), 2010
45
Herrera, L. G., T. H. Fleming & L. S. L. Sternberg, 1998. Trophic relationships in a neotropical bat community: A preliminary study using carbon and nitrogen isotopic signatures. Tropical Ecology, 39: 23–29.
Herrera, L. G., K. A. Hobson, L. Mirón-M, M. Ramírez-P, G. Méndez-C & V. Sánchez-Cordero, 2001. Sources of protein in two species of phytophagous bats in a seasonal dry forest: Evidence from stable isotope analysis. Journal of Mammalogy, 82: 352–361.
Herrera, L. G., E. Gutierrez, K. A. Hobson, B. Altube, W. G. Díaz & V. Sánchez-Cordero, 2002. Sources of assimilated protein in five species of New World frugivorous bats. Oecologia, 133: 280–287.
Humphrey, S. R., F. J. Bonaccorso & T. L. Zinn, 1983. Guild struc-ture of surface-gleaning bats in Panama. Ecology, 64: 284–294.
Kalko, E. K. V., 1998. Organisation and diversity of tropical bat communities through space and time. Zoology, 101: 281–297.
Kalko, E. K. V. & C. O. Handley, 2001. Neotropical bats in the canopy: Diversity, community structure, and implications for conservation. Plant Ecology, 153: 319–333.
Kalko, E. K. V., C. O. Handley & D. Handley, 1996. Organization, diversity, and long-term dynamics of a Neotropical bat com-munity. Pages 503–533 in M. Cody & J. Smallwood (eds). Long Term Studies in Vertebrate Communities. Academic Press, Los Angeles, California.
Kreft, H., N. Köster, W. Küper, J. Nieder & W. Barthlott, 2004. Diversity and biogeography of vascular epiphytes in Western Amazonia, Yasuní, Ecuador. Journal of Biogeography, 31: 1463–1476.
Kunz, T. H. & C. A. Diaz, 1994. Folivory in fruit-eating bats, with new evidence from Artibeus jamaicensis (Chiroptera: Phyllostomidae). Biotropica, 72: 106–120.
Kunz, T. H. & K. A. Ingalls, 1994. Folivory in bats: An adaptation derived from frugivory. Functional Ecology, 8: 665–668.
Lim, B. K., M. D. Engstrom, T. E. Lee, J. C. Patton & J. W. Bichkam, 2004. Molecular differentiation of large species of fruit-eating bats (Artibeus) and phylogenetic relationships based on the cyto-chrome b gene. Acta Chiropterologica, 6: 1–12.
Magurran, A. E., 2004. Measuring Biological Diversity. Blackwell Publishing, Oxford.
Matt, F., 2001. Pflanzenbesuchende Fledermäuse im tropischen Bergregenwald: Diversität, Einnischung und Gildenstruktur. Ph.D. thesis. Friedrich-Alexander-Universität, Erlangen-Nürnberg.
Miron, L. L., L. G. Herrera, N. Ramírez & K. A. Hobson, 2006. Effect of diet quality on carbon and nitrogen turnover and iso-topic discrimination in blood of a New World nectarivorous bat. Journal of Experimental Biology, 209: 541–548.
Moreno, C. M., H. T. Arita & L. Solis, 2006. Morphological assembly mechanisms in Neotropical bat assemblages and ensembles within a landscape. Oecologia, 149: 133–140.
Muchhala, N., P. Mena & L. Albuja, 2005. A new species of Anoura (Chiroptera: Phyllostomidae) from the Ecuadorian Andes. Journal of Mammalogy, 86: 457–461.
Nogueira, M. R. & A. L. Peracchi, 2003. Fig-seed predation by 2 species of Chiroderma: Discovery of a new feeding strategy in bats. Journal of Mammalogy, 84: 225–233.
Norberg, U. M. & J. M. V. Rayner, 1987. Ecological morphol-ogy and flight in bats (Mammalia; Chiroptera): Wing adapta-tions, flight performance, foraging strategy and echolocation. Philosophical Transactions of the Royal Society of London B, 316: 335–427.
Primack, R. & R. Corlett, 2005. Tropical Rain Forests: An Ecological and Biogeographical Comparison. Blackwell Publishing, Oxford.
Rex, K., D. Kelm, K. Wiesner, T. H. Kunz & C. C. Voigt, 2008. Species richness and structure of three Neotropical bat assem-blages. Biological Journal of the Linnean Society, 94: 617–629.
Roxburgh, L. & B. Pinshow, 2002. Digestion of nectar and insects by Palestine sunbirds. Physiological and Biochemical Zoology, 75: 583–589.
Sánchez-Hernandez, C., M. L. Romero-A. & G. D. Schnell, 2005. A new species of Sturnira (Chiroptera: Phyllostomidae) from northern South America. Journal of Mammalogy, 86: 866–872.
Sanford, R. L., Jr., P. Paaby, J. C. Luvall & E. Phillips, 1994. Climate, geomorphology and aquatic systems. Pages 19–33 in L. McDade, K. Bawa, H. Hespenheide & G. Hartshorn (eds). La Selva: Ecology and Natural History of a Neotropical Rain Forest. University of Chicago Press, Chicago, Illinois.
Shanahan, M., S. So, S. G. Compton & R. T. Corlett, 2001. Fig-eating by vertebrate frugivores: A global review. Biological Reviews, 76: 529–572.
Simmons, N. B., 2005. Chiroptera. Pages 312–529 in D. E. Wilson & D. M. Reeder (eds). Mammal Species of the World. John Hopkins University Press, Baltimore, Maryland.
SPSS, 2003. Version 12.0. SPSS Inc., Chicago, Illinois.
Swartz, S. M., P. W. Freeman & E. F. Stockwell, 2003. Ecomorphology of bats: Comparative and experimental approaches relating structural design to ecology. Pages 257–300 in T. H. Kunz & M. B. Fenton (eds). Bat Ecology. University of Chicago Press, Chicago, Illinois.
Timm, R. M. & R. K. LaVal, 1998. A Field Key to the Bats of Costa Rica. Occasional Publication 22. Center of Latin American Studies, University of Kansas, Lawrence, Kansas.
Tirira, D., 1999. Mamíferos del Ecuador. Museo de Zoología, Publicatión Especial 2, Quito.
Voigt, C. C., B. J. Bradley & S. Ortmann, 2009. Dietary analysis of plant-visiting bats. Pages 593–609 in T. H. Kunz & S. Parsons (eds). Ecological and Behavioral Methods for the Study of Bats. Johns Hopkins University Press, Baltimore, Maryland.
Voigt, C. C. & F. Matt, 2004. Nitrogen stress causes unpredictable enrichments of 15N in two nectar-feeding bat species. Journal of Experimental Biology, 207: 1741–1748.
Voigt, C. C. & J. R. Speakman, 2007. Nectar feeding bats fuel their high metabolism directly with exogenous carbohydrates. Functional Ecology, 21: 913–921.
Voigt, C. C., F. Matt, R. Michener & T. H. Kunz, 2003. Low rates of carbon isotope turnover in tissues of two nectar-feed-ing bat species (Glossophaginae, Phyllostomidae). Journal of Experimental Biology, 206: 1419–1427.
Voigt, C. C., G. U. G. Lehmann, R. Michener & M. M. Joachimski, 2006. Nuptial feeding is reflected in tissue nitro-gen isotope ratios of female katydids. Functional Ecology, 20: 656–661.
von Helversen, O. & Y. Winter, 2003. Glossophagine bats and their flowers: Costs and benefits. Pages 346–397 in T. H. Kunz & M. B. Fenton (eds). Bat Ecology. University of Chicago Press, Chicago, Illinois.
Whitaker, J. O., Jr., G. F. McCracken & B. M. Siemers, 2009. Food habits analysis of insectivorous bats. Pages 564–592 in T. H. Kunz & S. Parsons (eds). Ecological and Behavioral Methods for the Study of Bats. Johns Hopkins University Press, Baltimore, Maryland.

Rex et al.: Phyllostomid diveRsity and feeding habits
46
Willig, M. R., G. R. Camilo & S. J. Noble, 1993. Dietary overlap in frugivorous and insectivorous bats from edaphic cerrado habitats of Brazil. Journal of Mammalogy, 74: 117–128.
Wilson, D. E., 1972. Bat faunas: A trophic comparison. Systematic Zoology, 22: 14–29.
Winter, Y. & O. von Helversen, 2003. Operational tongue length in phyllostomid nectar-feeding bats. Journal of Mammalogy, 84: 886–896.
Yanega, G. M. & M. A. Rubega, 2004. Feeding mechanisms: Hummingbird jaw bends to aid insect capture. Nature, 428: 625.
York, H. A. & S. A. Billings, 2009. Stable-isotope analysis of diets of short-tailed fruit bats (Chiroptera: Phyllostomidae: Carollia). Journal of Mammalogy, 90: 1469–1477.
Zortea, M. & S. L. Mendes, 1993. Folivory in the big fruit-eating bat, Artibeuslituratus (Chiroptera: Phyllostomidae). Journal of Tropical Ecology, 9: 117–120.

Copyright of Ecoscience is the property of Ecoscience and its content may not be copied or emailed to multiple
sites or posted to a listserv without the copyright holder's express written permission. However, users may print,
download, or email articles for individual use.
Copyright © 2022 FDOKUMEN




















