
K+ channel KV3.1 associates with OSP/claudin-11 and regulates oligodendrocyte development
13
doi: 10.1152/ajpcell.00510.2005 291:C687-C698, 2006. First published 19 April 2006; Am J Physiol Cell Physiol Vu and Jeff Bronstein Seema Tiwari-Woodruff, Luis Beltran-Parrazal, Andrew Charles, Thomas Keck, Trung regulates oligodendrocyte development 3.1 associates with OSP/claudin-11 and V channel K + K You might find this additional info useful... 44 articles, 23 of which you can access for free at: This article cites http://ajpcell.physiology.org/content/291/4/C687.full#ref-list-1 3 other HighWire-hosted articles: This article has been cited by http://ajpcell.physiology.org/content/291/4/C687#cited-by including high resolution figures, can be found at: Updated information and services http://ajpcell.physiology.org/content/291/4/C687.full found at: can be American Journal of Physiology - Cell Physiology about Additional material and information http://www.the-aps.org/publications/ajpcell This information is current as of May 7, 2013. 0363-6143, ESSN: 1522-1563. Visit our website at http://www.the-aps.org/. Rockville Pike, Bethesda MD 20814-3991. Copyright © 2006 the American Physiological Society. ISSN: molecular physiology. It is published 12 times a year (monthly) by the American Physiological Society, 9650 is dedicated to innovative approaches to the study of cell and American Journal of Physiology - Cell Physiology at NIH-NIEHS Library on May 7, 2013 http://ajpcell.physiology.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of K+ channel KV3.1 associates with OSP/claudin-11 and regulates oligodendrocyte development

doi: 10.1152/ajpcell.00510.2005291:C687-C698, 2006. First published 19 April 2006;Am J Physiol Cell Physiol
Vu and Jeff BronsteinSeema Tiwari-Woodruff, Luis Beltran-Parrazal, Andrew Charles, Thomas Keck, Trungregulates oligodendrocyte development
3.1 associates with OSP/claudin-11 andV channel K+K
You might find this additional info useful...
44 articles, 23 of which you can access for free at: This article citeshttp://ajpcell.physiology.org/content/291/4/C687.full#ref-list-1
3 other HighWire-hosted articles: This article has been cited by http://ajpcell.physiology.org/content/291/4/C687#cited-by
including high resolution figures, can be found at: Updated information and serviceshttp://ajpcell.physiology.org/content/291/4/C687.full
found at: can beAmerican Journal of Physiology - Cell Physiology about Additional material and information
http://www.the-aps.org/publications/ajpcell
This information is current as of May 7, 2013.
0363-6143, ESSN: 1522-1563. Visit our website at http://www.the-aps.org/. Rockville Pike, Bethesda MD 20814-3991. Copyright © 2006 the American Physiological Society. ISSN:molecular physiology. It is published 12 times a year (monthly) by the American Physiological Society, 9650
is dedicated to innovative approaches to the study of cell andAmerican Journal of Physiology - Cell Physiology
at NIH
-NIE
HS
Library on May 7, 2013
http://ajpcell.physiology.org/D
ownloaded from

K� channel KV3.1 associates with OSP/claudin-11 and regulatesoligodendrocyte development
Seema Tiwari-Woodruff, Luis Beltran-Parrazal, Andrew Charles,Thomas Keck, Trung Vu, and Jeff BronsteinUCLA Multiple Sclerosis Program, Department of Neurology,David Geffen School of Medicine at UCLA, Los Angeles, California
Submitted 12 October 2005; accepted in final form 12 April 2006
Tiwari-Woodruff, Seema, Luis Beltran-Parrazal, AndrewCharles, Thomas Keck, Trung Vu, and Jeff Bronstein. K� channelKV3.1 associates with OSP/claudin-11 and regulates oligodendrocytedevelopment. Am J Physiol Cell Physiol 291: C687–C698, 2006. Firstpublished April 19, 2006; doi:10.1152/ajpcell.00510.2005.—K�
channels are differentially expressed throughout oligodendrocyte(Olg) development. KV1 family voltage-sensitive K� channels havebeen implicated in proliferation and migration of Olg progenitor cell(OPC) stage, and inward rectifier K� channels (KIR)4.1 are requiredfor OPC differentiation to myelin-forming Olg. In this report we haveidentified a Shaw family K� channel, KV3.1, that is involved inproliferation and migration of OPC and axon myelination. Applicationof anti-KV3.1 antibody or knockout of Kv3.1 gene decreased thesustained K� current component of OPC by 50% and 75%, respec-tively. In functional assays block of KV3.1-specific currents or knock-out of Kv3.1 gene inhibited proliferation and migration of OPC. AdultKv3.1 gene-knockout mice had decreased diameter of axons anddecreased thickness of myelin in optic nerves compared with age-matched wild-type littermates. Additionally, KV3.1 was identified asan associated protein of Olg-specific protein (OSP)/claudin-11 viayeast two-hybrid analysis, which was confirmed by coimmunopre-cipitation and coimmunohistochemistry. In summary, the KV3.1 K�
current accounts for a significant component of the total K� current incells of the Olg lineage and, in association with OSP/claudin-11, playsa significant role in OPC proliferation and migration and myelinationof axons.
membrane potential; tight junction; myelin; progenitor cell
OLIGODENDROCYTES (Olg), which are responsible for centralnervous system (CNS) axon myelination, arise from the pro-liferating neuroepithelial cells of the ventricular and subven-tricular zones in mammalian brain and the ventral midline anddorsal origin in the spinal cord (23, 37). During development,Olg lineage cells show stage-dependent K� channel expres-sion. Olg progenitor cells (OPC) express all six members of thedelayed rectifier Shaker family K� channels, voltage-sensitiveK� channels (KV)1.1–KV1.6, and show large voltage-depen-dent outward currents, some of which is due to KV1 channels(1, 7, 13, 18, 30), which have also been suggested to have a keyrole in OPC cell cycle progression (7, 14, 28). Olg lineage cellsalso express Ca2�-sensitive and inwardly rectifying K� (KIR)channels, and during differentiation, as the cells elaborate themyelin, the inward rectifiers are upregulated (2, 5, 8, 17, 32,34). Despite this rich population of K� channels, a largecomponent of sustained outward K� current is still unac-
counted for. Here we report that the bulk of OPC K� current isdue to KV3.1 from the Shaw family of K� channels, which areexpressed early but persist in differentiated cells and play animportant role in proper myelination.
KV3.1 has been well characterized in different neuronalsubtypes, including cerebellar granule cells, substantia nigrareticulata, thalamic nuclei, inferior colliculus, cochlear nuclei,the superior olive medial nucleus of the trapezoid body, andnodes of Ranvier (10, 19, 20, 24, 25, 27, 41). The presence ofKV3.1 has not been investigated in OPC/Olg lineage cells. Ourattention focused on this channel because it was identified byyeast two-hybrid analysis as an associated protein of Olg-specific protein (OSP)/claudin-11 (38, 39), a member of theclaudin family of tight junction proteins and a probable au-toantigen in the development of autoimmune demyelinatingdisease (6, 36). We hypothesized then that KV3.1 is expressedin significant quantities in Olg cells and plays a role in theirstructure and function. In this report we present findings insupport of our hypothesis, including 1) the presence of signif-icant KV3.1 message and protein in Olg lineage cells, 2)structural changes in myelin of Kv3.1-knockout (Kv3.1�/�)mice, 3) significant reduction of outward K� currents by KV3.1antibody, and 4) significant decrease in outward K� current inOPC of Kv3.1�/� mice. Additionally, the association ofKV3.1 with OSP/claudin-11 was further tested by coimmuno-precipitation, coimmunohistochemistry, and electrophysiologyanalysis, and the functional effects of this association wereprobed by proliferation and migration assays, functions previ-ously shown to be affected by OSP/claudin-11 knockout andimmunoblock (38, 39).
MATERIALS AND METHODS
Materials. Rabbit anti-KV1.3, -KV1.5, -2.1, -SK1, -SK2, -SK3,-KIR4.1, -KV3.2, -KV3.3, -KV3.4, and -KV3.1 antibodies (1:200 to1:500), agitoxin-2 (Alomone Labs, Israel), anti-bromodeoxyuridine(BrdU) (1:20; Oncogene, San Diego, CA), rabbit anti-KV1.2,-KV3.1b, and -KIR4.1 antibodies (1:500); mouse anti-O4 antibody,GalC, mouse anti-Caspr antibody (1:250); and rat anti-myelin basicprotein (MBP) (1:50) and rabbit anti-NG2 (1:100) antibodies werepurchased from Chemicon (Temecula, CA). Rabbit anti-KV3.1loop
was previously kindly donated by Dr. X. Y. Huang (44); later wegenerated a similar antibody, using the same peptide sequence [rabbitanti-KV3.1loop, against a 14-amino acid peptide (GAQPNDP-SASEHTH) from the external vestibule of the KV3.1 channel protein;Sigma Genesis]. The peptide sequence has high homology to othermembers of the KV3 family (KV3.2, -3.3, and -3.4). KV3.1loop anti-
Address for reprint requests and other correspondence: S. K. Tiwari-Woodruff, Glia Biology Group, UCLA Multiple Sclerosis Program, Dept. ofNeurology, NRB1, 635 Charles Young Dr. S., Los Angeles, CA 90095 (e-mail:[email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Am J Physiol Cell Physiol 291: C687–C698, 2006.First published April 19, 2006; doi:10.1152/ajpcell.00510.2005.
0363-6143/06 $8.00 Copyright © 2006 the American Physiological Societyhttp://www.ajpcell.org C687
at NIH
-NIE
HS
Library on May 7, 2013
http://ajpcell.physiology.org/D
ownloaded from

body recognized only KV3.1a and KV3.1b protein subunits in wild-type (WT) homogenates and none in Kv3.1�/� homogenates. Ratanti-platelet-derived growth factor receptor (PDGFR)-� and anti-BrdU antibodies were obtained from BD Biosciences-Pharmingen andOncogene. The second antibody step was performed by labeling withantibodies conjugated to tetramethylrhodamine isothiocyanate, FITC,and Cy5 (Vector Labs and Chemicon). All other chemicals werepurchased from Sigma-Aldrich (St. Louis, MO) or Invitrogen (Carls-bad, CA).
Animals. WT, OSP/claudin-11 heterozygous knockout (C57BL/6 � C3H; Ref. 15), and KV3.1 heterozygous knockout (C57BL/6 fromDr. Bernardo Rudy, New York University, New York, NY; Refs. 16,24) mouse breeding pairs were used to obtain postnatal day(P)0–P50animals. Littermates were genotyped by using tail DNA as thetemplate for amplification by PCR as shown previously (12, 15).Animals were housed under guidelines set by the National Institutesof Health and experiments were conducted in accordance with theUCLA Chancellor’s Animal Research Committee and the PublicHealth Service Policy on Humane Care and Use of LaboratoryAnimals.
Perfusion. Mice were deeply anesthetized with isoflurane andperfused transcardially with ice-cold 0.9% saline followed by 4%paraformaldehyde. Brains with optic nerves (ON) attached wereremoved, postfixed overnight in 4% paraformaldehyde, and cryopro-tected with 20% sucrose solution in PBS. Free-floating sections (25�m thick) were cut coronally with a sliding microtome and collectedserially in PBS.
Yeast two-hybrid screen, yeast two-hybrid bait, and library con-structions. Three OSP/claudin-11 constructs were used as bait foryeast two-hybrid screen: the entire OSP/claudin-11 open readingframe (ORF), the first extracellular domain of OSP/claudin-11 be-tween amino acids 35 and 84, and the COOH terminus of OSP/claudin-11 between amino acids 122 and 207. These cDNAs werecloned into the pGBT9 backbone (Matchmaker system; ClonetechLabs). GAL4 activation domain cDNA fusion libraries were con-structed in modified pGAD GH vector (Clonetech), using mRNAfrom mouse brain. Yeast two-hybrid screening was performed accord-ing to Tiwari-Woodruff et al. (38).
Isolation and differentiation of O2A cells. OPC were isolated fromP0–P2 mouse brains by immunopanning with anti-A2B5 (3) or withthe shaking technique (22). For differentiation to mature Olg mediumwas supplemented with T3-T4-containing Sato medium, and cellswere postfixed after 2 (O4�), 4 (GalC�), and 8 (MBP�) days tomaximize stage-specific Olg.
RT-PCR. RNA was isolated from P0 and P25 A2B5� OPC andMBP� Olg, and first-strand cDNA was synthesized with randomhexamer primers and Superscript II reverse transcriptase (Invitrogen).PCR primers were designed according to the mouse/rat KV3.1 cDNAsequence: [KV3.1a (accession no. NM008421): primer pairs to coverbp 1018–1650; KV3.1b (accession no. M68880): primer pairs to coverbp 2653–2902]. To semiquantify various samples, coamplification ofglyceraldehyde-3-phosphate dehydrogenase (GAPDH) enzyme (ac-cession no. X02231; primer pairs to cover bp 591–1042) mRNA wasperformed simultaneously.
Western blot analysis. Immunoprecipitation and Western blot anal-ysis were performed as described previously (38, 39). OPC and mousebrain were homogenized in buffer A (mM: 150 NaCl, 1 EDTA, and 10Tris �HCl and 1% Triton X-100) or radioimmunoprecipitation assay(RIPA) buffer (containing 0.1% SDS, 0.5% deoxycholate in additionto 1% Triton X-100) in the presence of protease and phosphataseinhibitors. Twenty micrograms of protein in sample loading buffercontaining 10% �-mercaptoethanol and 5 mM DTT was subjected toSDS-PAGE and transferred to nitrocellulose. Specific primary anti-bodies followed by horseradish peroxidase-conjugated secondary an-tibody were used and visualized with chemiluminesence substrate(Pierce, Rockford, IL). For immunoprecipitation 100 �g of homoge-nates was incubated with antibodies on protein A agarose beads.
Beads were washed and boiled in 40 �l of sample buffer (containing2% SDS, 10% �-mercaptoethanol, and 10 mM DTT), subjected toSDS-PAGE, and transferred to nitrocellulose. Antibody concentrationand immunoprecipitation efficiency (by Western blot) were confirmedfor each antibody used. Images were scanned in Adobe Photoshop(Adobe Systems, San Jose, CA).
Proliferation and migration assays. To identify proliferating cellsin vitro, primary OPC from WT, transgenic KV3.1, and OSP/clau-din-11 littermates were isolated and counted and then labeled withBrdU for 24 h. Cell migration was quantified by counting the numberof cells moving from agarose drops as described previously (38).Statistical significance was assessed with the Student’s paired t-test.
Histopathology and immunohistochemistry. Serial sections weremounted on slides and stained with hematoxylin and eosin. Consec-utive sections were also examined by immunohistochemistry. Brainslices and Olg lineage cells were immunostained as previously de-scribed (38, 39). Gelatin-embedded optic nerves were similarly im-munostained. IgG control experiments were performed for all primaryantibodies, and no staining was observed under these conditions.Briefly, sections and cells were fixed with 4% paraformaldehyde,permeabilized with 0.3% Triton X-100, immunostained with primaryfollowed by secondary antibodies, washed and mounted on coverslides, dried, and coverslipped with Fluoromount G. For better pen-etration of antibodies (KV3.1, OSP/claudin-11, and MBP staining) tolabel white matter structures, some spinal cord and brain sections werepretreated with 95% ethanol-5% acetic acid for 15 min, beforeproceeding through the permeabilization steps. Confocal imaging wasperformed with a Leica TCS-SP (Mannheim, Germany) microscope,and some images were also taken with a fluorescence microscope(BX51WI; Olympus, Tokyo, Japan) equipped with Plan Fluor objec-tives connected to a camera (DP70, Olympus). Digital images werecollected and analyzed with Leica confocal and DP70 camera soft-ware. Images were assembled with Adobe Photoshop.
Electrophysiology. Current recordings were obtained with thewhole cell configuration of the patch-clamp technique as previouslydescribed (28, 33). OPC were perfused with (mM) 150 NaCl, 5.4 KCl,1.5 CaCl2, 1.5 MgSO4, 10 glucose, and 10 HEPES with 0.5–1 �Mtetrodotoxin (pH 7.3). K� channel blockers 4-aminopyridine (4-AP)and tetraethylammonium chloride (TEA) and KV3.1 channel antibod-ies were directly added to the bath solution. Patch-clamp pipettes werefilled with internal solution containing (mM) 130 KCl, 2 MgCl2, 1.0CaCl2, 10 EGTA, and 10 HEPES, pH 7.3. Delayed rectifier currentswere routinely investigated by applying a series of voltage stepsranging from �80 to �80 mV for 500 ms from a holding potential(Vh) of either �80 or �40 mV. Cell sealing and breakthrough intowhole cell mode was performed in current-clamp conditions permit-ting an accurate determination of cell resting membrane potential. Todetermine the voltage dependence of steady-state activation, currentswere elicited by 50-ms voltage pulses (�80 to �80 mV, 10-mVincrements) from a Vh of �80 mV. After the steady-state peakoutward currents (IK) were converted into conductance (GK) [GK �IK/(Vh � EK), where EK is Nernst potential of K� at 21°C, i.e., �80mV], conductance (G) at various membrane potentials was normal-ized to maximal conductance (Gmax). G/Gmax was plotted vs. Vh.Curves were fitted to Boltzmann functions to determine the voltage ofhalf-maximal activation (V1/2). In some cases, the peak outwardcurrents were directly plotted vs. Vh. Patch-clamp recordings wereperformed with an Axopatch 200A amplifier; data were stored andanalyzed with Axon Instruments (Foster City, CA) pCLAMP softwareand Origin 6.0 (Microcal Software) on a Pentium PC. All data werecollected at 10 kHz and analog filtered at 5 kHz.
Data analysis and statistics. Data are reported as means � SE.Statistical significance was assessed with a two-sided t-test for pairedor unpaired samples at the significance level (P). SEs of fittedparameters were obtained by analyzing data of individual experimentsseparately. Results of electrophysiology were statistically evaluated
C688 K� CHANNEL KV3.1 REGULATES OLIGODENDROCYTE FUNCTION
AJP-Cell Physiol • VOL 291 • OCTOBER 2006 • www.ajpcell.org
at NIH
-NIE
HS
Library on May 7, 2013
http://ajpcell.physiology.org/D
ownloaded from

with the Origin 6.0 software package. Boltzmann fits were performedwith the same software.
Assaying process length and complexity. Differences in length andcomplexity of processes between Kv3.1�/� and littermate Olg weredetermined by using a modification of the Scholl method (29). Acircular grid was constructed on a plastic transparency that wassuperimposed on a �40 image of cells printed on paper. Twenty-fiveMBP-stained Olg from four different litters of Kv3.1�/� and controlmice were photographed at �40. The nucleus of the cell was posi-tioned beneath the center of the grid, and each section was assigned ascore equal to the diameter of the circle crossed by the longest processin the section. An approximate diameter score was obtained byaveraging these numbers.
Quantification of light and electron microscopy. For light micros-copy, semithin sections of ON and brain stem were cut at 2 �m,toluidine blue counterstained, and photographed. Three regions fromeach section were randomly chosen for quantification. Axon diameterwas measured by tracing the perimeter of 50 neighboring axons inseveral samples for a total of 200 axons from Kv3.1�/� (n � 5) andKv3.1�/� (n � 4) age-matched P50 littermates, and pooled dataacross individual animals within each group were used to calculatemeans � SE. For electron microscopy, age-matched littermates wereperfused with saline followed by fixative (2% glutaraldehyde � 2%paraformaldehyde). Serial ultrathin sections from fixed ON embeddedin Epon oriented to visualize myelinated axons were stained withuranyl acetate-lead citrate, and representative areas were photo-graphed. Montages (4–6) consisting of either �4,800 or �14,000were obtained from select sections. To measure myelin thickness weoverlaid a square grid on the photographs and measured the myelinwidth every time a vertical grid encountered myelin. We avoidedmeasuring myelin areas that were loosely compacted or frayed. Formost axons two encounters were measured. The ratio of axon diameterto total fiber diameter (g ratio) was measured from both Kv3.1�/�and Kv3.1�/� P25 and P50 ON and brain stem by dividing thecircumference of an axon without myelin by the circumference of thesame axon including myelin.
RESULTS
Kv3.1 mRNA and protein are expressed in cells of Olglineage. Kv3.1 gene transcription can result in alternative-spliced products that differ only in their COOH-terminal se-quence, with the last 18 amino acid sequences in KV3.1areplaced by 84 amino acid residues in KV3.1b (20). Thepresence of both alternatively spliced KV3.1 mRNAs, inroughly equal amounts, in primary cultures of OPC and Olgwas confirmed by RT-PCR (Fig. 1A). OPC obtained for cul-turing by immunopanning, which results in 99% OPC har-vested, were induced to differentiate to Olg. P45 KV3.1 WT(�/�) mouse brain homogenate (MBH) was used as positivecontrol, whereas P45 littermate Kv3.1 homozygous knockout(�/�) MBH, used as a negative control, did not show KV3.1-specific products. The relative amounts of KV3.1a and KV3.1bmRNA was estimated by coamplification of GAPDH mRNA.
To establish the expression of KV3.1 protein in brain andphenotypically defined stages of Olg development in vitro,Western blot analysis was performed. Native and heterolo-gously expressed KV3.1a (55–95 kDa) and KV3.1b (60–110kDa) protein has been shown to be of varying lengths depend-ing on phosphorylation state and cell line (21, 25, 44). KV3.1b-specific antibody recognized a single protein of 96 kDa inKv3.1�/� MBH, OPC, and Olg (Fig. 1B, left). The 96-kDaband may correspond to the glycosylated form of KV3.1bprotein because the size is larger than predicted from the aminoacid sequence of the core polypeptides (66 kDa; Ref. 24).
KV3.1loop antibody, which recognizes both KV3.1a andKV3.1b, bound three proteins of 96, 75, and 60 kDa in OPCand Olg (Fig. 1B, right). The 75-kDa and 60-kDa bands couldbe glycosylated and nonglycosylated KV3.1, as the bandsdisappear when Kv3.1�/� homogenates are used. Eventhough the peptide (GAQPNDPSASEHTH) from the externalvestibule of the KV3.1 channel protein used to make KV3.1loop
antibody is highly homologous to KV3.2 and to a lesser amountto KV3.3 and KV3.4, the antibody is specific to KV3.1a andKV3.1b, as no significant immunostaining is observed whenKv3.1�/� tissue is used. The specific bands were also notobserved when appropriate antigenic peptides were used topreadsorb the antibodies.
The presence of KV3.1 in OPC and Olg in culture was alsodetected immunohistochemically, with both KV3.1b-specificand KV3.1loop antibodies giving robust staining. Figure 1C, left,shows cultured Olg labeled with KV3.1loop antibody. Figure1C, middle, shows the same field labeled with MBP staining, amarker for late differentiating Olg. The merged images (Fig.1C, right) show that a KV3.1� Olg was also MBP�. The insetin Fig. 1C shows an A2B5 immunopanned OPC in culturelabeled with KV3.1b antibody. No staining was observed whenpeptide-adsorbed antibodies were used or when OPC and Olgfrom Kv3.1�/� mice were used. OPC also immunolabeledmoderately with antibodies to KV1.3, KV2.1, SK3-type Ca2�-activated K� channels, and KV3 subtypes KV3.2 and KV3.3and intensely with antibodies to KV1.5, SK2-type Ca2�-acti-vated K� channels, KIR4.1, and KV3.4 (Fig. 1D). No immu-nostaining difference was observed between WT andKv3.1�/� OPC and Olg for these K� channels.
Olg lineage cells also label with anti-KV3.1 in vivo. Someperfusion-fixed P4–P12 brain sections were labeled with he-matoxylin and eosin stain for orientation, and selected sectionswere coimmunostained with KV3.1loop antibody and two Olglineage markers, anti-PDGFR-� and anti-rip. Anti-PDGFR-�labels precursor cells committed to Olg lineage, and anti-ripspecifically labels rip, a protein of unknown function, found inthe cytoplasm of premyelinating and myelinating Olg. OPC inthe corpus callosum (box in hematoxylin and eosin-stainedsection in Fig. 2A) of P4–P12 brains were KV3.1loop/KV3.1b-and PDGFR-�- and KV3.1- and rip-immunopositive (Fig. 2Bi–iii). Anti-chondroitin sulfate proteoglycan NG2, which identi-fies OPC in adult brain, also colabeled with KV3.1b antibody(results not shown). P15 optic nerve Olg showed robust im-munostaining with anti-KV3.1loop (Fig. 2Bi, inset) and KV3.1b(data not shown). It is important to note that although most ofthe positively indicated OPC and Olg cells were also positivefor KV3.1, Olg and OPC markers did not label many of theKV3.1� cells in vivo. These cells are likely neurons, but we didnot analyze this directly.
Myelin thickness and average axon diameter are reduced inoptic nerve of Kv3.1�/� mice. Anti-MBP immunostaining(Fig. 2, C and D) and electron microscopy analysis (Fig. 2, Eand F) revealed five consequences of Kv3.1 knockout for Olgmorphology and axon myelination. 1) Olg showed a significant(P 0.05) decrease in diameter, 54 � 8 �m (n � 25, 4different litters) for Kv3.1�/� compared with 79 � 4 �m forKv3.1�/� littermates (n � 25). 2) There was reduced Olgmembrane branching complexity (Fig. 2C) [MBP immuno-stained 8–12 Olg per well per animal, and 6 different pairs ofKv3.1�/� and Kv3.1�/� littermates were compared]. 3) As
C689K� CHANNEL KV3.1 REGULATES OLIGODENDROCYTE FUNCTION
AJP-Cell Physiol • VOL 291 • OCTOBER 2006 • www.ajpcell.org
at NIH
-NIE
HS
Library on May 7, 2013
http://ajpcell.physiology.org/D
ownloaded from

C690 K� CHANNEL KV3.1 REGULATES OLIGODENDROCYTE FUNCTION
AJP-Cell Physiol • VOL 291 • OCTOBER 2006 • www.ajpcell.org
at NIH
-NIE
HS
Library on May 7, 2013
http://ajpcell.physiology.org/D
ownloaded from

shown in the white matter tracts near the corpus callosum (Fig.2D), there was discontinuous and punctuate MBP� staining inKv3.1�/� compared with the even staining observed withKv3.1�/� littermates [corpus callosum areas from 5Kv3.1�/� and Kv3.1�/� littermate brains were compared]. 4)There is a substantial reduction in the number of large-diameteraxons, also shown for the cross section of the ON and brainstem in Fig. 2Ei–iii. 5) The thickness of the myelin layer isreduced, as is shown for the ON in the electron microscopysections of Fig. 2Eii. Axon diameter was measured by tracingthe perimeter of 50 neighboring axons in several samples for atotal of 200 axons from Kv3.1�/� (n � 5) and Kv3.1�/�(n � 4) P50 littermates. Mean axon diameter was reduced inKv3.1�/� (0.78 � 0.04 �m compared with controls, 0.93 �0.14 �m; Fig. 2Eiv). ON myelin sheath thickness was 0.069 �0.04 �m in Kv3.1�/� compared with 0.089 � 0.03 �m inKv3.1�/� littermates (Fig. 2Evi). These differences weresignificant (P 0.05) across multiple sections of age-matchedanimals (n � 4).
The thickness of the myelin sheath is related to the axondiameter, and the g ratio, defined as the ratio of the innerdiameter to the outer diameter, presents the precise relationbetween the axon diameter and the myelin sheath thickness. Todetermine whether there was a variation in the amount ofmyelin around myelinated axons in the CNS of Kv3.1�/�mice, we calculated the ratio of the axon diameter to the totalfiber diameter (g ratio). Axon regions that were without com-pact myelin were not included in this analysis. The calculatedg ratio was not significantly different between Kv3.1�/� andKv3.1�/� animals in either ON (Fig. 2Ev) or brain stem. Toanalyze whether there were any major changes in myelinprotein of Kv3.1�/� animals we analyzed PLP and OSPprotein levels of age-matched Kv3.1�/� and Kv3.1�/� P50brain by Western blot but saw no significant difference bydensitometry of the gel bands (data not shown).
KV3.1 channel is major component IK in mouse OPC. Fromprevious characterizations in neurons, KV3.1-dependent cur-rents should 1) not be inactivated at modestly depolarizedpotentials, 2) be sensitive to low concentrations of TEA (rel-ative to KV1 currents, which are sensitive to high TEA), and 3)be sensitive to anti-KV3.1loop antibody. In addition, outwardcurrents should be substantially reduced in Kv3.1�/�. Thesepredictions were tested in cultured A2B5� OPC.
The resting potential of OPC was �35 � 6 mV (n � 62).K� currents activated from a Vh near the resting membranepotential (�40 mV) were slowly activating and noninactivat-ing and had a V1/2 of �10–6.5 mV, similar to the range thathas been described for KV3.1 channels (21). K� currentsactivated from a more hyperpolarized potential of �80 mVwere a combination of rapidly activating, fast inactivating A
currents (IA) and slowly activating, noninactivating currents(Fig. 3C).
KV3.1 channels are relatively sensitive to TEA (IC50 �150–200 �M) compared with KV1 channels (IC50 � 2–50mM). From Vh � �40 mV outward currents were decreased by19 � 5% by 100 �M TEA and 78 � 4% by 1 mM TEA (Fig.4, A and B). From Vh � �80 mV outward currents weredecreased by only 5 � 2% by 100 �M TEA and by only 50 �4% by 1 mM TEA (Fig. 3, C and D).
KV3.1loop antibody (250 nM), directed to an epitope in theexternal vestibule of KV3.1 channel, blocked 79% of theKV3.1 currents from KV3.1-transfected HEK cells but not otherrelated (KV1.2) K� channels (44). In OPC KV3.1loop antibodyblocked 48 � 6.4% of the outward K� conductance within7–10 min of application (n � 6; Fig. 3, E and F). Additionaluse of 1 mM TEA reduced outward current to 25% of control(Fig. 3, E and F). G/Gmax vs. voltage curves show nearly 50%block with KV3.1loop antibody present for 10 min and littleeffect on the voltage sensitivity. The antibody specific to splicevariant KV3.1b is directed to the cytoplasmic COOH termini ofKV3.1b and was used as a control. It did not show inhibition ofthe currents (data not shown).
Outward K� currents of OPC are significantly reduced inthe absence of KV3.1 protein. KV3.1 current was also investi-gated by comparing the physiological properties of Kv3.1�/�OPC to Kv3.1�/� littermates. The resting membrane potentialof Kv3.1�/� OPC was �35 � 3 mV (n � 12), not differentfrom controls; however, outward K� currents from either Vh ��40 or �80 mV were dramatically reduced in Kv3.1�/� (Fig.3G), primarily as a result of decrease in the sustained compo-nent. Kv3.1�/� OPC showed decreased current throughout therange of activating potentials (Fig. 3H), with the largest dif-ference for large depolarizations. From Vh � �40 mV, a stepto �60 mV produced 1,596 � 284 pA (means � SE; n � 42)in controls compared with only 147 � 58 pA (n � 9) inKv3.1�/�.
Much of the voltage-activated OPC currents from Vh � �80mV is fast IA, which is inactivated when voltage is held at �40mV. This current can be estimated by subtracting outward K�
currents obtained from Vh � �40 mV from those obtained at�80 mV. If this is done for Kv3.1�/� and Kv3.1�/� the“subtracted” IA components are very similar in amplitude andshape, suggesting that when Kv3.1 gene is knocked out the K�
channels that underlie the IA current are expressed normallyand are unaffected. Consistent with this finding, V1/2 of acti-vation in Kv3.1�/� obtained from Vh � �40 mV and �80mV showed a significant difference (�5.1 � 4 and �20 � 2.5mV, respectively) compared with Kv3.1�/� (8.6 � 4 and20 � 6 mV) (P 0.01). Thus the remainder of K� currents in
Fig. 1. Presence of Kv3.1 mRNA and protein in oligodendrocyte (Olg) cells. A: transcription of KV3.1a and KV3.1b mRNA in GalC� Olg and A2B5� Olgprogenitor cells (OPC). RT-PCR products of wild-type (WT) and Kv3.1�/� postnatal day (P)45 mouse brain were used as controls. GalC� Olg and A2B5� OPCwere positive for KV3.1a (570 bp) and for KV3.1b (249 bp). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH; 451 bp) controls are used for quantifyingrelative transcript levels. Mr denotes DNA standards for size calibration. MBH, mouse brain homogenate. B: Western blot analysis of KV3.1 protein in GalC�
Olg and A2B5� OPC. A band of 96 kDa was present when anti-KV3.1b antibody was used. To estimate the possible presence of both KV3.1a and KV3.1bprotein, anti-KV3.1loop antibody identified 3 bands (96, 75, and 60 kDa). Preincubation of antibody with KV3.1b-specific peptide or KV3.1loop-specific peptideeliminated specific bands, confirming antibody specificity (�Pep). Kv3.1�/� P45 and Kv3.1�/� P45 brain homogenates were used as controls. C: KV3.1 isexpressed in primary cell cultures in all stages of Olg lineage. Differentiated Olg were labeled with KV3.1loop (red) and myelin basic protein (MBP; blue)antibody. Proliferating A2B5-immunopanned OPC in culture were immunolabeled with anti-KV3.1b-specific antibody (inset). Magnification �40. D: cells of Olglineage express different K� channel subtypes. Early differentiating primary cell cultures of OPC/Olg were colabeled with KV1.5, SK2, KIR4.1, and KV3.4antibodies (red) and anti-GalC antibody (green). Magnification �20.
C691K� CHANNEL KV3.1 REGULATES OLIGODENDROCYTE FUNCTION
AJP-Cell Physiol • VOL 291 • OCTOBER 2006 • www.ajpcell.org
at NIH
-NIE
HS
Library on May 7, 2013
http://ajpcell.physiology.org/D
ownloaded from

C692 K� CHANNEL KV3.1 REGULATES OLIGODENDROCYTE FUNCTION
AJP-Cell Physiol • VOL 291 • OCTOBER 2006 • www.ajpcell.org
at NIH
-NIE
HS
Library on May 7, 2013
http://ajpcell.physiology.org/D
ownloaded from

Fig. 3. OPC outward K� currents are sensitiveto tetraethylammonium chloride (TEA), anti-KV3.1 antibody, and Kv3.1�/�. A: OPC out-ward currents when the cell was depolarized to60 mV from holding potential (Vh) of �40 mVbefore (E) and after addition of 100 �M (■ ) and1 mM (�) TEA. Calibration bars are for A, C,and E. B: currents from a Vh of �40 mV for 28OPC were converted to conductance (G), andG/Gmax was plotted against voltage before (E)and after TEA (■ , 100 �M; �, 1 mM). Errorbars are SE. C: currents from the same cellfrom a Vh of �80 mV before (E) and after 100�M (■ ) and 1 mM (�) TEA. D: as in B froma Vh of �80 mV. E: OPC K� currents evokedfrom Vh �40 mV to 60 mV before (E) and afteraddition of anti-KV3.1loop antibody (250 nM)for 3 (F), 7 (‚), and 10 (■ ) min followed by 1mM TEA (�). F: conductance-voltage curvesof the same experiment. G: outward OPC cur-rents were generated by voltage steps in 10-mVincrements from �80 mV to 80 mV from Vh
�40 mV (top) or �80 mV (bottom) inKv3.1�/� littermates (left) and Kv3.1�/�(right). Only the 20-mV increment steps areshown, for clarity. Calibration bars are for all 4sets of data. H: steady-state current-voltagevalues for WT (E; n � 42) and Kv3.1�/� (F;n � 9) OPC for Vh �40 mV. I: normalizedconductance-voltage plots for the OPC in H.
Fig. 2. Olg lineage cells express KV3.1 in vivo. A: P12 brain section near the hippocampus (hp) and corpus callosum (cc), hematoxylin and eosin stained, showingthe region immunolabeled in solid box in B. Bar, 0.5 mm. B: i: OPC labeled with platelet-derived growth factor receptor (PDGFR)-� antibody (green) alsocolabeled with anti-KV3.1loop (red), shown more clearly for the few cells (boxed area in i) with individual staining (ii). Bar, 10 �m. Inset, optic nerve OPCimmunolabeled with anti-KV3.1loop (red; magnification �40). iii: Same region of the brain labeled with anti-rip (green) and anti-KV3.1loop (red) showing separatestaining and merged images. iv: Similar region of the brain immunostained with PDGFR-� (green) and KV3.1b (red). Merged image shows costaining in 2 of3 cells. C: Olg in differentiating medium are immunolabeled with MBP (�40 magnification). Kv3.1�/� Olg in differentiating medium immunolabeled with MBPshows extensive myelin sheaths compared with Kv3.1�/� Olg. D: Kv3.1�/� MBP-immunostained white matter tracts near the outer borders of corpus callosum(�63 magnification) show bunched and discontinuous staining. E: i: 70-nm optic nerve cross sections were photographed with electron microscope at �4,800magnification. ii: Optic nerve cross sections photographed at �19,000. Insets, examples showing the thinner myelin sheath in Kv3.1�/�. iii: Cross section ofa similar area from the brain stem of Kv3.1�/� and Kv3.1�/� at �19,000. The loosely compacted myelin of a Kv3.1�/� axon is shown. iv: Axon diameterwas significantly reduced in optic nerves of Kv3.1�/� compared with Kv3.1�/� littermates. v: Ratio of axon diameter to total fiber diameter (g ratio) wasmeasured from Kv3.1�/�, Kv3.1�/�, and Kv3.1�/� animals at P50. No difference was observed between animals. vi: Myelin sheathing on individual axonswas also disrupted and significantly thinner in optic nerves of Kv3.1�/� compared with Kv3.1�/� littermates (*P 0.05).
C693K� CHANNEL KV3.1 REGULATES OLIGODENDROCYTE FUNCTION
AJP-Cell Physiol • VOL 291 • OCTOBER 2006 • www.ajpcell.org
at NIH
-NIE
HS
Library on May 7, 2013
http://ajpcell.physiology.org/D
ownloaded from
C694 K� CHANNEL KV3.1 REGULATES OLIGODENDROCYTE FUNCTION
AJP-Cell Physiol • VOL 291 • OCTOBER 2006 • www.ajpcell.org
at NIH
-NIE
HS
Library on May 7, 2013
http://ajpcell.physiology.org/D
ownloaded from

Kv3.1�/� OPC could be due to remaining KV1.3, KV1.5, andIA-producing K� channels.
OSP/claudin-11 and KV3.1 form a protein complex in Olgprogenitors. A mouse brain cDNA GAL4 activation domainlibrary was screened with OSP/claudin-11 GAL4 binding do-main baits in pGBT vector (38). Only transfection with OSP/claudin-11 C-portion bait (amino acids 122 and 207) resultedin positive clones. The two clones were called OSP/claudin-11-associated protein (OAP)-1 and OAP-2. OAP-1, found to bea novel member of the tetraspanin superfamily, forms a com-plex with OSP/claudin-11 and �1-integrin (38). OAP-2 con-tained nucleotide sequence 372–1506 of the ORF, correspond-ing to amino acids 124–502 (COOH termini) of the voltage-sensitive K� channel of the Shaw family, KV3.1. The region ofKV3.1 associated with OSP/claudin-11 is common to bothsplice variants of the channel.
Coimmunoprecipitation experiments were done to confirmthe association of KV3.1 with OSP/claudin-11. OPC homoge-nates were immunoprecipitated under relatively stringent con-ditions (1% Triton X-100; Ref. 9) with anti-OSP/claudin-11 oranti-KV3.1b specific antibodies, with anti-glial fibrillary acidicprotein (GFAP) and anti-KV1.3, proteins not expected to as-sociate with either KV3.1 or OSP/claudin-11, used as controls.Western blots were probed with anti-KV3.1b antibody (Fig. 4A,left) or anti-OSP/claudin-11 (Fig. 4A, right). There was recip-rocal immunoprecipitation of OSP/claudin-11 with anti-KV3.1and KV3.1 with anti-OSP/claudin-11. Neither KV3.1b nor OSP/claudin-11 was precipitated when the primary antibody wasomitted or when primary antibodies to unrelated proteins,GFAP or KV1.3, were used.
Immunoprecipitation was also done in a more stringentRIPA buffer, containing 0.5% deoxycholate, 0.1% SDS, and1% Triton X-100 (Fig. 4B). Homogenates were immunopre-cipitated with KV3.1b-specific, KV3.1loop, and OSP/claudin-11antibodies, and Western blots of the precipitates were probedwith KV3.1loop (Fig. 4B, top) or OSP/claudin-11 antibodies(Fig. 4B, bottom). Again there was reciprocal precipitation ofOSP/claudin-11 and KV3.1; however, probing with anti-KV3.1revealed that anti-OSP/claudin-11 could associate with bothsplice variants, KV3.1b and KV3.1a. Anti-KV3.1b antibody wasnot able to pull down KV3.1a, consistent with the specificity ofthe antibody and with the notion that OSP/claudin-11 canassociate with either KV3.1a or KV3.1b. This was expected
given that the cDNA fragment isolated from the yeast two-hybrid screen codes for a peptide common to both KV3.1a andKV3.1b. Anti-KV3.1loop antibody was able to precipitate the75-kDa species, labeled by the anti-KV3.1loop, but anti-OSP/claudin-11 antibody was not. The 75-kDa species may be aform of KV3.1 that does not associate with OSP/claudin-11.Negative control immunoprecipitation antibodies anti-GFAPand anti-KV1.3 did not pull down OSP/claudin-11, KV3.1a orKV3.1b, or the 75-kDa species.
Association between KV3.1 and OSP/claudin-11 was exam-ined in cultured OPC and brain sections with immunocyto-chemistry and confocal microscopy (Fig. 4, C and D). KV3.1band OSP/claudin-11 colocalization was observed in primarycell cultures of A2B5� progenitors (Fig. 4C, inset) and inGalC� Olg (Fig. 4C). KV3.1b staining in processes of A2B5�/GalC� cells was punctuate and discontinuous, and colocaliza-tion with OSP/claudin-11 was limited to cell body and discreteregions of processes. Similar to OSP/claudin-11, KV3.1loop
(not KV3.1b) immunostained live OPC on the cell surface, andthe two proteins showed strong colocalization in cell soma andnot processes (Fig. 4Cii). KV3.1loop also colocalized withOSP/claudin-11 in fixed mature MBP� Olg (Fig. 4Ciii). Co-localization of KV3.1 and OSP/claudin-11 in NG2�, PDGFR-��, and rip� cells was also observed in P4-P8 brain and ON(data not shown). In adult brain, KV3.1b and KV3.1loop immu-nostaining was also found in neurons and in most white mattertracts, where it partially colocalized with OSP/claudin-11 (Fig.4D, i and iv). OSP/claudin-11 colabeled with the paranodalprotein Caspr, whereas some KV3.1loop and KV3.1b (notshown) immunostaining was observed in the nodes, betweenCaspr-immunostained paranodes, consistent with previousstudies (Fig. 4D, ii and iii). KV3.1 and OSP/claudin-11 did notappear to colocalize in either the paranode or the nodal region.Magnified images in the corpus callosum and ON (not shown)indicated specific regions that had colabeling of the two pro-teins (Fig. 4Div, merged image). We have not been able toclearly define the colocalization of these proteins to the myelintight junctions.
To understand the functional significance of KV3.1 andOSP/claudin-11 interaction in OPCs, we analyzed K� currentsin the absence of OSP/claudin-11. Resting membrane potentialof OSP/claudin-11�/� OPC were �37 � 3 mV (n � 14) andwere similar to OSP/claudin-11�/� OPC. Outward K� cur-
Fig. 4. KV3.1b associates with OSP/claudin-11. A: primary OPC homogenates (80–100 �g) were immunoprecipitated (IP) (43) with anti-OSP/claudin-11,-KV3.1b, -glial fibrillary acidic protein (GFAP), or -KV1.3 antibodies, and Western blots were probed with either anti-KV3.1b (left) or anti-OSP/claudin-11(right). The IgG band is visible near 56 kDa. The anti-KV3.1b-labeled band was 96 kDa, and OSP/claudin-11 was at 23 kDa. B: IP of OPC homogenatesperformed with more stringent radioimmunoprecipitation assay (RIPA) buffer conditions. Top: probe with anti-KV3.1loop antibody. Bottom: probe withanti-OSP/claudin-11. The anti-KV3.1b-labeled band was 96 kDa; the OSP/claudin-11 band was 23 kDa; and the KV3.1a band was 60 kDa. The protein betweenKV3.1a and KV3.1b is 75 kDa. See text for details. C: i: primary cultures of GalC� Olg and A2B5� OPC (inset) were immunolabeled with anti-KV3.1b (green)followed by anti-OSP/claudin-11 (red). Merged images show regions of colocalization in yellow. ii: Live A2B5� OPC were immunolabeled with anti-KV3.1loop
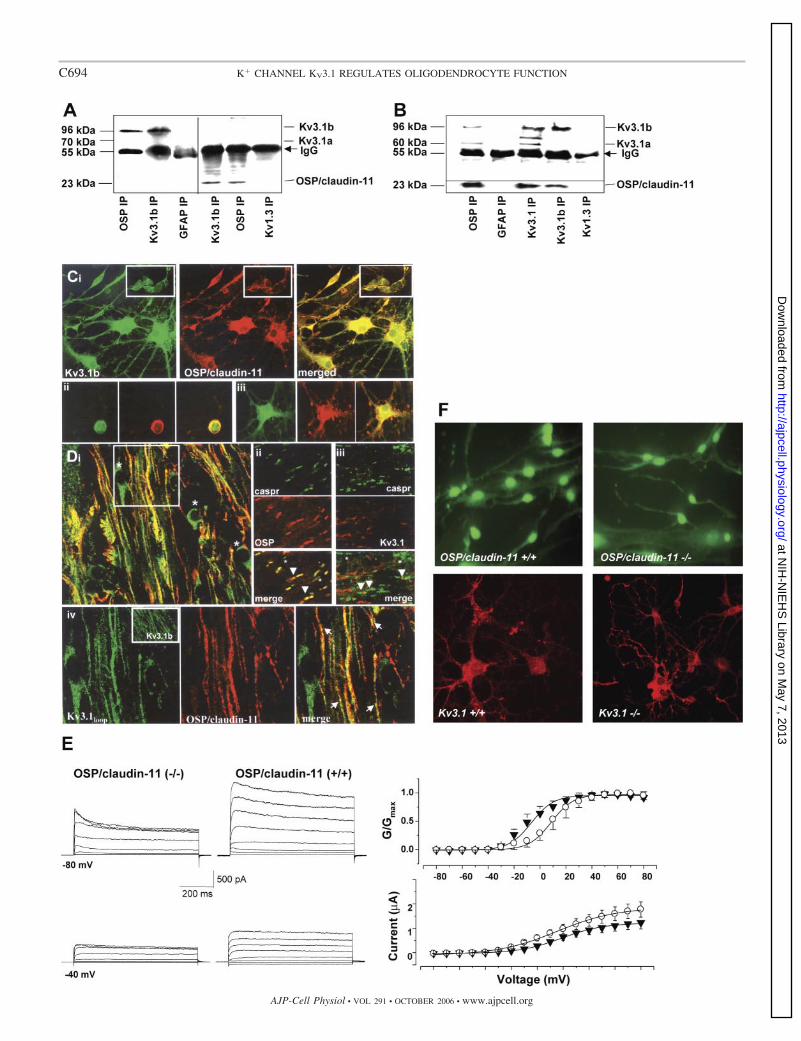
and OSP/claudin-11 and then fixed. Colocalization is observed more on the membrane surface borders. iii: MBP� Olg immunostained with KV3.1loop andOSP/claudin-11 showing maximal colocalization in the cell body and minimal in the processes. D: i: region of P45 brain including corpus callosum and corteximmunostained with KV3.1loop (green) and OSP/claudin-11 (red) in merged confocal image. Only KV3.1 immunostaining is in neuronal cells (stars), whereas insome parts of the white matter it colabels with OSP/claudin-11, which is exclusively present in the white matter. A higher magnification of the area marked inwhite square is shown in iv (arrows denote colabeling of OSP/claudin-11 and KV3.1loop). ii: P25 optic nerve immunostained with anti-Caspr antibody labels theparanode regions and OSP/claudin-11 labels most (arrowheads), but not all (stars) paranodes. iii: Anti-KV3.1loop immunolabels a few nodes (arrowhead betweenCaspr� paranodes; stars denote no KV3.1 immunostaining). Inset in iv shows anti-KV3.1b in P45 corpus callosum white matter. E: OPC K� currents weregenerated by voltage steps in 20-mV increments from �80 mV to 80 mV from Vh �40 mV (top) or �80 mV (bottom) in OSP/claudin-11�/� (left) andOSP/claudin-11�/� (right) littermates. Steady-state current-voltage values for WT (E; n � 42) and OSP/claudin-11�/� (�; n � 14) OPC for Vh �40 mV andnormalized conductance-voltage plots are shown. F: pre-Olg from OSP/claudin-11�/� and OSP/claudin-11�/� cultured for 2 days in differentiating mediumwere immunostained with anti-KV3.1loop antibody (green). Anti-A2B5 immunopanned Kv3.1�/� and Kv3.1�/� littermate OPC were cultured for 4 days andimmunostained with anti-OSP/claudin-11 antibody (red). No significant difference in either KV3.1 or OSP/claudin-1 localization was observed.
C695K� CHANNEL KV3.1 REGULATES OLIGODENDROCYTE FUNCTION
AJP-Cell Physiol • VOL 291 • OCTOBER 2006 • www.ajpcell.org
at NIH
-NIE
HS
Library on May 7, 2013
http://ajpcell.physiology.org/D
ownloaded from

rents of OSP/claudin-11�/� OPC were significantly reduced(1,105 � 221 pA, 30%) compared with OSP/claudin-11�/�OPC (Fig. 4E). Similar to Kv3.1�/� currents the reductionwas primarily due to reduction in IK. V1/2 of OSP/claudin-11�/� was similar to Kv3.1�/�, �8.6 � 3 mV at Vh ��40mV and �10.6 � 3 mV at Vh ��80 mV. The altered K�
currents in the OSP/claudin-11�/� OPC might be due toaltered membrane trafficking of the K� channels or alteredchannel kinetics. To estimate whether OSP/claudin-11 absencewas causing a decrease in overall KV3.1 expression, we per-formed Western blotting and saw no differences in eitherKV3.1a or KV3.1b bands between OSP/claudin-11�/� OPCand OSP/claudin-11�/� OPC homogenates (data not shown).Immunostaining with anti-KV3.1loop antibody of OSP/claudin-11�/� OPC and OSP/claudin-11�/� OPC or with anti-OSP/claudin-11 antibody of Kv3.1�/� pre-Olg and Kv3.1�/�pre-Olg showed no significant difference (Fig. 4F). Theseresults indicate the importance and complexity of OSP/clau-din-11 interaction with KV3.1 that is tailored for specific K�
currents required for normal OPC function.KV3.1 and OSP/claudin-11 are involved in OPC prolifera-
tion and migration. Proliferation assays were performed inOPC cultured for 24 h in the presence of mitogenic factors(PDGF and bFGF; Fig. 5A). K� channel blockers TEA and4-AP inhibited OPC proliferation in a concentration-dependentmanner, and to an extent the inhibition was comparable toanti-KV3.1loop antibody addition. Boiled anti-KV3.1loop anti-body had no effect. Anti-KV1.3 antibody that targets an extra-
cellular domain of KV1.3 inhibited proliferation slightly. Anti-�1-integrin and anti-OSP/claudin-11 caused significant reduc-tion in proliferation (38), and KV3.1loop antibody did not blockproliferation of Kv3.1�/� OPC (Fig. 5B). Kv3.1�/� OPC hada small (20%) but significant reduction in proliferation com-pared with Kv3.1�/� OPCs (Fig. 5A, right).
In vitro OPC migration, which depends on fibronectin sub-strate (Fig. 5B), was inhibited in the presence of K� channelblockers TEA and 4-AP and anti-KV3.1loop antibody. Antibodyto the COOH terminus of KV3.1b (anti-KV3.1b) and anti-GFAP antibody, both added as controls, did not affect migra-tion. Kv3.1�/� OPC showed a significant reduction in migra-tion compared with Kv3.1�/� (Fig. 5D).
DISCUSSION
KV3.1 channel expression is widespread in the nervoussystem, with KV3.1b mRNA far more abundant than theKV3.1a transcript in adult brain (26, 42) but with both tran-scripts strongly expressed in neuronal subpopulations of theolfactory bulb, neocortex, hippocampus, basal nuclei, thala-mus, brain stem, and cerebellum. It is therefore surprising thatthe knockout mouse has a mild phenotype (16). We were ableto determine that a structural effect of Kv3.1 knockout is areduction in myelin thickness and a decrease in the number oflarge-diameter axons (shown for ON in Fig. 2E). The expectedfunctional consequence of this would be reduced conductionvelocity and slowed response times, a nonlethal but suboptimal
Fig. 5. KV3.1 blockers and Kv3.1�/� inhibitOPC proliferation and migration. A: prolifera-tion was assayed by counting bromodeoxyuri-dine (BrdU)-positive OPC and normalizingthem to control after 24 h of incubation in thepresence of KV3.1loop, boiled (b) anti-KV3.1loop, KV1.3 K� channel extracellular(ec), and OSP/claudin-11 antibodies andblockers TEA and 4-aminopyridine (4-AP).KV3.1loop antibody did not block proliferationof Kv3.1�/� OPC (n � 3 separate experi-ments, right of dashed line). Values are ex-pressed as % of control. **P 0.005. B: OPCswere isolated from Kv3.1�/�, Kv3.1�/�, andKv3.1�/� littermates. Difference in Kv3.1�/�vs. Kv3.1�/� OPC proliferation as indicated bynumber of BrdU-positive cells was significant(*P 0.05). C: OPC migration was assayed inthe absence (control) and presence of KV3.1loop,OPS/claudin-11, KV3.1b COOH termini, andGFAP antibodies and blockers TEA and 4-AP.Except for �fibronectin control, all conditionscontained fibronectin. Values are expressed as %of control. **P 0.005. D: migration assayswere performed with OPC isolated fromKv3.1�/� (n � 10), Kv3.1�/� (n � 6), andKv3.1�/� (n � 6) littermates in the presence offibronectin. Values are expressed as % ofKv3.1�/�. Difference in Kv3.1�/� vs.Kv3.1�/� OPC migration was significant (*P 0.05).
C696 K� CHANNEL KV3.1 REGULATES OLIGODENDROCYTE FUNCTION
AJP-Cell Physiol • VOL 291 • OCTOBER 2006 • www.ajpcell.org
at NIH
-NIE
HS
Library on May 7, 2013
http://ajpcell.physiology.org/D
ownloaded from

situation for the organism, and could explain the motor skilldeficit in these mice (16). The cause of these structural changescould reside in the neurons; because the g ratio is unchangedthe decrease in axonal width could be intrinsic to Kv3.1�/�neurons. However, because Olg and OPC also express KV3.1,the principal finding of this report, the pathology could residepartially in the Olg.
In this report we present evidence that 1) KV3.1 transcriptand protein for both alternative sliced species are found in OPCand Olg [an earlier study using RNase protection assay toidentify KV3.1 mRNA in C6 glioma cell line and mixedpopulation of ON cultures was unsuccessful (26)]; 2) 50% ofthe voltage-dependent outward K� current in OPC is due toKV3.1; 3) KV3.1 forms a complex with a major, Olg-specificprotein, OSP/claudin-11; 4) specific block of KV3.1 or Kv3.1knockout inhibits proliferation and migration of OPC; and 5)Kv3.1 knockout affects normal myelination.
KV3.1 may have functions not shared with other K� chan-nels, possibly through its association with OSP/claudin-11 andthe other proteins shown to be members of the complex,OAP-1 and �1-integrin (38). The complex is important ulti-mately as a mediator of cell-cell contact and must have a rolein myelination. The existence of a K� channel as part of thecomplex raises the possibility that voltage-dependent localincreases in extracellular K� may affect tight junction forma-tion. Also, having a voltage-dependent protein as a partner inthe complex may confer voltage sensitivity to it, which maythen allow voltage changes to influence tight junction forma-tion. Aside from specialized functions, activation of the chan-nel must also affect repolarization after depolarization stimuli,as would any of the voltage-dependent K� channels. A directway in which this could influence Olg is by affecting stimulus-dependent intracellular changes in Ca2�, an important secondmessenger that controls or modifies every cell behavior andfunction (4, 31). Restricting K� conductance with anti-KV3.1or Kv3.1 knockout should increase stimulus-dependent Ca2�
increases in the cytoplasm of Olg and OPC, and we arepresently examining this prediction.
The slowing of OPC proliferation and migration, shown herewith KV3.1 block or Kv3.1�/�, might be involved in theKv3.1�/�-dependent thinning of the ON myelin sheath; how-ever, there is likely to be slowing of other Olg behaviors aswell, for example, transitions in developmental stages and afterdifferentiation in the rate of myelination once it has begun. Ifthere is a critical period for myelination and Olg are slow torespond, the window of opportunity may close before themyelin sheath can obtain normal thickness. If this were thecase it would imply that there is an independent mechanism formyelination termination, and this has important implicationsnot only for developmental myelination but also for remyeli-nation after injury and disease.
An interesting point made by the proliferation/migrationexperiments is that acute block of KV3.1 by anti-KV3.1 is moreeffective than Kv3.1 knockout. This suggests that Kv3.1�/�Olg have a mechanism of compensation, perhaps the upregu-lation of an alternative K� channel. Another member of theKV3 subtype, KV3.3, is widely coexpressed with KV3.1 indistinct neuronal populations in the CNS. Mice lacking bothKV3.1 and KV3.3 K� channel alleles display severe motordeficits (11, 12) compared with either single mutation. Immu-nohistochemical analysis indicates that other voltage-depen-
dent K� channels including KV3.3 are probably not upregu-lated in the Kv3.1�/� Olg, but a Ca2�-dependent K� channelwould be a good candidate for compensation, especially if theprincipal effect of Kv3.1�/� is loss of feedback control overstimulus-dependent Ca2� increases. Upregulation of Ca2�-dependent K� channels would act as a substitute negative-feedback mechanism. If compensation does occur, and themechanism of it is common to other cells, it would help explainthe relatively mild phenotypic effects of Kv3.1 knockout.
Here we have demonstrated that KV3.1 associates withOSP/claudin-11, contributes significantly to OPC K� currents,and has a role in OPC development and axon myelination.Because distinct changes in the membrane K� channel pheno-type of OPC occur during lineage progression (3, 35, 40) andKV3.1 along with KV1 type channels and KIR4.1 has a majorrole in the regulation of OPC development and axon myelina-tion, a complete understanding of the mechanism by whichKV3.1 and related proteins exert their influence on OPC de-velopment would be beneficial in developing therapies fordemyelinating diseases.
ACKNOWLEDGMENTS
We thank Michael Woodruff for helpful discussion and comments on thismanuscript. We acknowledge the Carol Moss Spivak Cell Imaging Facility atUCLA.
GRANTS
This work was supported by grants from the NMSS (RG355A1, S. Tiwari-Woodruff), the National Institute of Neurological Disorders and Stroke (NS-01596, J. Bronstein), the Department of Veterans Affairs, and the SW PA-DRECC (J. Bronstein).
REFERENCES
1. Bacia A, Wollmann R, and Soliven B. K� channel blockade impairsremyelination in the cuprizone model. Glia 48: 156–165, 2004.
2. Barres BA, Chun LL, and Corey DP. Calcium current in corticalastrocytes: induction by cAMP and neurotransmitters and permissiveeffect of serum factors. J Neurosci 9: 3169–3175, 1989.
3. Barres BA, Koroshetz WJ, Swartz KJ, Chun LL, and Corey DP. Ionchannel expression by white matter glia: the O-2A glial progenitor cell.Neuron 4: 507–524, 1990.
4. Berridge MJ, Bootman MD, and Roderick HL. Calcium signalling:dynamics, homeostasis and remodelling. Nat Rev Mol Cell Biol 4: 517–529, 2003.
5. Borges K, Ohlemeyer C, Trotter J, and Kettenmann H. AMPA/kainatereceptor activation in murine oligodendrocyte precursor cells leads toactivation of a cation conductance, calcium influx and blockade of delayedrectifying K� channels. Neuroscience 63: 135–149, 1994.
6. Bronstein JM, Lallone RL, Seitz RS, Ellison GW, and Myers LW. Ahumoral response to oligodendrocyte-specific protein in MS: a potentialmolecular mimic. Neurology 53: 154–161, 1999.
7. Chittajallu R, Chen Y, Wang H, Yuan X, Ghiani CA, Heckman T,McBain CJ, and Gallo V. Regulation of Kv1 subunit expression inoligodendrocyte progenitor cells and their role in G1/S phase progressionof the cell cycle. Proc Natl Acad Sci USA 99: 2350–2355, 2002.
8. Chvatal A, Pastor A, Mauch M, Sykova E, and Kettenmann H.Distinct populations of identified glial cells in the developing rat spinalcord slice: ion channel properties and cell morphology. Eur J Neurosci 7:129–142, 1995.
9. Claas C, Stipp CS, and Hemler ME. Evaluation of prototype transmem-brane 4 superfamily protein complexes and their relation to lipid rafts.J Biol Chem 276: 7974–7984, 2001.
10. Devaux J, Alcaraz G, Grinspan J, Bennett V, Joho R, Crest M, andScherer SS. Kv3.1b is a novel component of CNS nodes. J Neurosci 23:4509–4518, 2003.
11. Espinosa F, Marks G, Heintz N, and Joho RH. Increased motor driveand sleep loss in mice lacking Kv3-type potassium channels. Genes BrainBehav 3: 90–100, 2004.
C697K� CHANNEL KV3.1 REGULATES OLIGODENDROCYTE FUNCTION
AJP-Cell Physiol • VOL 291 • OCTOBER 2006 • www.ajpcell.org
at NIH
-NIE
HS
Library on May 7, 2013
http://ajpcell.physiology.org/D
ownloaded from

12. Espinosa F, McMahon A, Chan E, Wang S, Ho CS, Heintz N, andJoho RH. Alcohol hypersensitivity, increased locomotion, and spontane-ous myoclonus in mice lacking the potassium channels Kv3.1 and Kv33.J Neurosci 21: 6657–6665, 2001.
13. Gallo V, Zhou JM, McBain CJ, Wright P, Knutson PL, and Arm-strong RC. Oligodendrocyte progenitor cell proliferation and lineageprogression are regulated by glutamate receptor-mediated K� channelblock. J Neurosci 16: 2659–2670, 1996.
14. Ghiani CA, Eisen AM, Yuan X, DePinho RA, McBain CJ, and GalloV. Neurotransmitter receptor activation triggers p27(Kip1)and p21(CIP1)accumulation and G1 cell cycle arrest in oligodendrocyte progenitors.Development 126: 1077–1090, 1999.
15. Gow A, Southwood CM, Li JS, Pariali M, Riordan GP, Brodie SE,Danias J, Bronstein JM, Kachar B, and Lazzarini RA. CNS myelin andsertoli cell tight junction strands are absent in Osp/claudin-11 null mice.Cell 99: 649–659, 1999.
16. Ho CS, Grange RW, and Joho RH. Pleiotropic effects of a disrupted K�
channel gene: reduced body weight, impaired motor skill and musclecontraction, but no seizures. Proc Natl Acad Sci USA 94: 1533–1538,1997.
17. Kettenmann H, Sonnhof U, and Schachner M. Exclusive potassiumdependence of the membrane potential in cultured mouse oligodendro-cytes. J Neurosci 3: 500–505, 1983.
18. Knutson P, Ghiani CA, Zhou JM, Gallo V, and McBain CJ. K�
channel expression and cell proliferation are regulated by intracellularsodium and membrane depolarization in oligodendrocyte progenitor cells.J Neurosci 17: 2669–2682, 1997.
19. Luneau C, Wiedmann R, Smith JS, and Williams JB. Shaw-like ratbrain potassium channel cDNA’s with divergent 3� ends. FEBS Lett 288:163–167, 1991.
20. Luneau CJ, Williams JB, Marshall J, Levitan ES, Oliva C, Smith JS,Antanavage J, Folander K, Stein RB, Swanson R, Kaczmarek LK,and Buhrow SA. Alternative splicing contributes to K� channel diversityin the mammalian central nervous system. Proc Natl Acad Sci USA 88:3932–3936, 1991.
21. Macica CM and Kaczmarek LK. Casein kinase 2 determines the voltagedependence of the Kv3.1 channel in auditory neurons and transfected cells.J Neurosci 21: 1160–1168, 2001.
22. McCarthy KD and de Vellis J. Preparation of separate astroglial andoligodendroglial cell cultures from rat cerebral tissue. J Cell Biol 85:890–902, 1980.
23. Miller RH. Dorsally derived oligodendrocytes come of age. Neuron 45:1–3, 2005.
24. Ozaita A, Martone ME, Ellisman MH, and Rudy B. Differentialsubcellular localization of the two alternatively spliced isoforms of theKv3.1 potassium channel subunit in brain. J Neurophysiol 88: 394–408,2002.
25. Perney TM and Kaczmarek LK. Localization of a high thresholdpotassium channel in the rat cochlear nucleus. J Comp Neurol 386:178–202, 1997.
26. Perney TM, Marshall J, Martin KA, Hockfield S, and Kaczmarek LK.Expression of the mRNAs for the Kv3.1 potassium channel gene in theadult and developing rat brain. J Neurophysiol 68: 756–766, 1992.
27. Rudy B, Kentros C, Weiser M, Fruhling D, Serodio P, Vega-Saenz deMiera E, Ellisman MH, Pollock JA, and Baker H. Region-specificexpression of a K� channel gene in brain. Proc Natl Acad Sci USA 89:4603–4607, 1992.
28. Schmidt K, Eulitz D, Veh RW, Kettenmann H, and Kirchhoff F.Heterogeneous expression of voltage-gated potassium channels of theshaker family (Kv1) in oligodendrocyte progenitors. Brain Res 843:145–160, 1999.
29. See J, Zhang X, Eraydin N, Mun SB, Mamontov P, Golden JA, andGrinspan JB. Oligodendrocyte maturation is inhibited by bone morpho-genetic protein. Mol Cell Neurosci 26: 481–492, 2004.
30. Shrager P and Novakovic SD. Control of myelination, axonal growth,and synapse formation in spinal cord explants by ion channels andelectrical activity. Brain Res Dev Brain Res 88: 68–78, 1995.
31. Soliven B. Calcium signalling in cells of oligodendroglial lineage. Mi-crosc Res Tech 52: 672–679, 2001.
32. Soliven B, Ma L, Bae H, Attali B, Sobko A, and Iwase T. PDGFupregulates delayed rectifier via Src family kinases and sphingosine kinasein oligodendroglial progenitors. Am J Physiol Cell Physiol 284: C85–C93,2003.
33. Soliven B, Szuchet S, Arnason BG, and Nelson DJ. Expression andmodulation of K� currents in oligodendrocytes: possible role in myelino-genesis. Dev Neurosci 11: 118–131, 1989.
34. Sontheimer H and Kettenmann H. Heterogeneity of potassium currentsin cultured oligodendrocytes. Glia 1: 415–420, 1988.
35. Sontheimer H, Perouansky M, Hoppe D, Lux HD, Grantyn R, andKettenmann H. Glial cells of the oligodendrocyte lineage express proton-activated Na� channels. J Neurosci Res 24: 496–500, 1989.
36. Stevens DB, Chen K, Seitz RS, Sercarz EE, and Bronstein JM.Oligodendrocyte-specific protein peptides induce experimental autoim-mune encephalomyelitis in SJL/J mice. J Immunol 162: 7501–7509, 1999.
37. Thomas JL, Spassky N, Perez Villegas EM, Olivier C, Cobos I,Goujet-Zalc C, Martinez S, and Zalc B. Spatiotemporal development ofoligodendrocytes in the embryonic brain. J Neurosci Res 59: 471–476,2000.
38. Tiwari-Woodruff SK, Buznikov AG, Vu TQ, Micevych PE, Chen K,Kornblum HI, and Bronstein JM. OSP/claudin-11 forms a complex witha novel member of the tetraspanin super family and �1 integrin andregulates proliferation and migration of oligodendrocytes. J Cell Biol 153:295–305, 2001.
39. Tiwari-Woodruff SK, Kaplan R, Kornblum HI, and Bronstein JM.Developmental expression of OAP-1/Tspan-3, a member of the tet-raspanin superfamily. J Neurosci Res 77: 166–173, 2004.
40. von Blankenfeld G and Kettenmann H. Glutamate and GABA receptorsin vertebrate glial cells. Mol Neurobiol 5: 31–43, 1991.
41. Weiser M, Bueno E, Sekirnjak C, Martone ME, Baker H, Hillman D,Chen S, Thornhill W, Ellisman M, and Rudy B. The potassium channelsubunit KV3.1b is localized to somatic and axonal membranes of specificpopulations of CNS neurons. J Neurosci 15: 4298–4314, 1995.
42. Weiser M, Vega-Saenz de Miera E, Kentros C, Moreno H, Franzen L,Hillman D, Baker H, and Rudy B. Differential expression of Shaw-related K� channels in the rat central nervous system. J Neurosci 14:949–972, 1994.
43. Zangani D, Darcy KM, Masso-Welch PA, Bellamy ES, Desole MS,and Ip MM. Multiple differentiation pathways of rat mammary stromalcells in vitro: acquisition of a fibroblast, adipocyte or endothelial pheno-type is dependent on hormonal and extracellular matrix stimulation.Differentiation 64: 91–101, 1999.
44. Zhou BY, Ma W, and Huang XY. Specific antibodies to the externalvestibule of voltage-gated potassium channels block current. J GenPhysiol 111: 555–563, 1998.
C698 K� CHANNEL KV3.1 REGULATES OLIGODENDROCYTE FUNCTION
AJP-Cell Physiol • VOL 291 • OCTOBER 2006 • www.ajpcell.org
at NIH
-NIE
HS
Library on May 7, 2013
http://ajpcell.physiology.org/D
ownloaded from




















