
Experimental validation of 5 in-silico predicted glioma biomarkers
Upload
independentCategory
view
0download
0
2012;72:4856-4868. Published OnlineFirst August 3, 2012.Cancer Res Joseph D. Dougherty, Elena I. Fomchenko, Afua A. Akuffo, et al. of Oligodendrocyte Progenitor-like Cells in GliomaCandidate Pathways for Promoting Differentiation or Quiescence
Updated Version 10.1158/0008-5472.CAN-11-2632doi:
Access the most recent version of this article at:
MaterialSupplementary
http://cancerres.aacrjournals.org/content/suppl/2012/08/03/0008-5472.CAN-11-2632.DC1.htmlAccess the most recent supplemental material at:
Cited Articles http://cancerres.aacrjournals.org/content/72/18/4856.full.html#ref-list-1
This article cites 68 articles, 18 of which you can access for free at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
[email protected] atTo order reprints of this article or to subscribe to the journal, contact the AACR Publications
To request permission to re-use all or part of this article, contact the AACR Publications Department at
American Association for Cancer Research Copyright © 2012 on September 17, 2012cancerres.aacrjournals.orgDownloaded from
Published OnlineFirst August 3, 2012; DOI:10.1158/0008-5472.CAN-11-2632

Tumor and Stem Cell Biology
Candidate Pathways for Promoting Differentiation orQuiescence of Oligodendrocyte Progenitor-like Cells inGlioma
Joseph D. Dougherty1, Elena I. Fomchenko4, Afua A. Akuffo1, Eric Schmidt8, Karim Y. Helmy4,Elena Bazzoli5, Cameron W. Brennan2,3, Eric C. Holland2,3,4,5,6, and Ana Milosevic7
AbstractPlatelet-derived growth factor receptor alpha–positive oligodendrocyte progenitor cells (OPC) located within
the mature central nervous system may remain quiescent, proliferate, or differentiate into oligodendrocytes.Human glioblastoma multiforme tumors often contain rapidly proliferating oligodendrocyte lineage transcrip-tion factor 2 (Olig2)-positive cells that resemble OPCs. In this study, we sought to identify candidate pathwaysthat promote OPC differentiation or quiescence rather than proliferation. Gene expression profiling conducted inboth normal murine OPCs and highly proliferative Olig2-positive glioma cells identified all the transcriptsassociated with the highly proliferative state of these cells and showed that among the various cell types foundwithin the brain, Olig2-positive tumor cells are most similar to OPCs. We then subtracted OPC transcripts foundin tumor samples from those found in normal brain samples and identified 28 OPC transcripts as candidatesfor promoting differentiation or quiescence. Systematic analysis of human glioma data revealed that these geneshave similar expression profiles in human tumors and were significantly enriched in genomic deletions,suggesting an antiproliferative role. Treatment of primary murine glioblastoma cells with agonists of onecandidate gene, Gpr17, resulted in a decreased number of neurospheres. Together, our findings show thatcomparison of the molecular phenotype of progenitor cells in tumors to the equivalent cells in the normal brainrepresents a novel approach for the identification of targeted therapies. Cancer Res; 72(18); 4856–68.�2012 AACR.
IntroductionGlioma is the most prevalent type of primary brain tumor in
adults and prognosis, especially for the high-grade, stage IVglioblastoma multiforme (GBM), is dismal. Recently, work atthemRNA (1–3) andprotein signaling levels (4) have convergedupon similar classification schemes that identify distinct sub-groups of high-grade glioma. These subgroups show differen-tial gene expression profiles, signaling cascades, and responseto treatment. In particular, one subgroup, referred to asproneural, is characterized by elevated expression of plate-let-derived growth factor (PDGF), amplifications of PDGFreceptor alpha (PDGFRA; ref. 4), and expression of oligoden-drocyte lineage transcription factor 2 (Olig2; refs. 1, 2).
In the mature central nervous system (CNS), the PDGFRAand the chondroitin sulfate proteoglycan antigen (NG2) havebeen considered as markers primarily of the oligodendrocyteprogenitor cells (OPC; refs. 5, 6). OPCs are sparsely and evenlydistributed throughout white and gray matter. The adult OPCwas thought to mainly serve as a repository for the generationof new mature oligodendrocytes (6) but may also serve as aneural stem cell (7). Although they are typically quiescent, theyare more proliferative than any other population in the brain,with 1% to 5% actively cycling at any time (8). Regardless oftheir role, these cells are capable of at least 3 fates: they mayterminally differentiate into oligodendrocytes, they may pro-liferate to produce additional progenitors, or they may remainquiescent.
We have previously generated a mouse model of glioma inwhich a Nestin (Nes)-positive neural stem cell can be inducedto overproduce PDGF-B (9, 10). The PDGFRA is activated byboth homo- and heterodimers of A and B forms of PDGF (11).These mice generate aggressive gliomas resulting in a tumormass composed of the highly proliferative cells of origin andcells derived from initially normal progenitors that arerecruited to the tumor (9, 12–14). The presence of cellsexpressing Olig2 and PDGFRA in these tumors suggests thattreatment strategies in the proneural subtype may be inform-ed by knowledge of the signaling pathways that regulate thechoice between proliferation, quiescence, and differentiationin these OPCs.
Authors' Affiliations: 1Departments ofGenetics andPsychiatry,Washing-ton University, St. Louis, Missouri; 2Department of Neurosurgery, 3BrainTumor Center, 4Department of Cancer Biology and Genetics, 5Departmentof Neurology, 6Department of Surgery, Memorial Sloan Kettering CancerCenter; and 7The GENSAT project, 8Laboratory of Molecular Biology, TheRockefeller University, New York, NY
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/).
Corresponding Author: Ana Milosevic, The Rockefeller University, 1230York Ave, Box 296, NY 10065. Phone: 212-327-7286; Fax: 212-327-7888;E-mail: [email protected]
doi: 10.1158/0008-5472.CAN-11-2632
�2012 American Association for Cancer Research.
CancerResearch
Cancer Res; 72(18) September 15, 20124856
American Association for Cancer Research Copyright © 2012 on September 17, 2012cancerres.aacrjournals.orgDownloaded from
Published OnlineFirst August 3, 2012; DOI:10.1158/0008-5472.CAN-11-2632
Current glioma treatment, following resection of the tu-mor, is focused on targeting proliferating cells with temo-zolomide and radiotherapy (15). However, as previouslynoted (16), a complementary strategy would be to promotepathways for maintaining quiescence and/or driving termi-nal differentiation of the progenitors present in the tumor,as this may also serve to slow tumor growth. Prodifferentia-tion treatments may be particularly promising; although thetumor environment is likely selecting for mutations in genesthat normally suppress cell-cycle progression and maintainquiescence, there may not be as much selective pressure onall prodifferentiation pathways.We undertook a study in the PDGF-B–induced mouse
model of glioblastoma to identify transcripts that could beimportant for the regulation of quiescence and differentiationin OPC-like cells. Applying our newly developed methodologyfor in vivo cell-specific translational profiling (17, 18), weidentified all mRNAs specifically enriched in OPCs in normalmouse brain, including those likely to be important for pro-liferation, quiescence, and differentiation. We then contrastedthis to the cell-specific translational profile of Olig2-positivecells in the mouse model of a proneural glioma, in which OPC-like cells are committed to proliferation at the expense ofdifferentiation or quiescence. This permitted identification ofcandidate pathways that may serve as targets for promotingdifferentiation andquiescence inOPCs inmice. Examination ofThe Cancer Genome Atlas (TCGA) expression profiles ofhuman gliomas established that analogous pathways are sim-ilarly regulated in human proneural GBM, suggesting theirconservation as targets. Also consistent with an antiprolifera-tive role, many of these targets show deletions in human GBM.
From this combined human and mouse screening, we haveidentified several candidate pathways for promoting quies-cence and differentiation, which may serve as targets forcomplementary treatments.
Material and MethodsFull materials and methods are available online. All proto-
cols involving animals were approved by the Rockefeller Uni-versity and Memorial Sloan Kettering Cancer Center Institu-tional Animal Care and Use Committee.
New mouse linesBacterial artificial chromosomes (BAC) containing genes
PDGFRA (RP23-55P22), Cnp1 (RP23-78L12), and Snap25 (RP23-290A18)weremodifiedasdescribed (17), to insert anEGFP-L10afusion protein into the relevant translation start site.
HistologyAnatomy was carried out as described (17, 19). For immu-
nofluorescence, samples were incubated with the cell-specificantibodies, and quantification was done on�40 cortical fieldsimaged with confocal microscopy.
Profiling tumor modelTumors were generated as described (9). Cells producing
RCAS-h PDGF-B or RCAS-Cre virus were injected into theprogeny of ink4a/arf�/� mice expressing tv-a receptor forRCAS under the Nes promoter, crossed to Olig2-Egfp::L10amice. Telomeric repeat amplification protocol (TRAP) andmicroarray hybridizations were carried out as described (17,18), except tumors were processed individually.
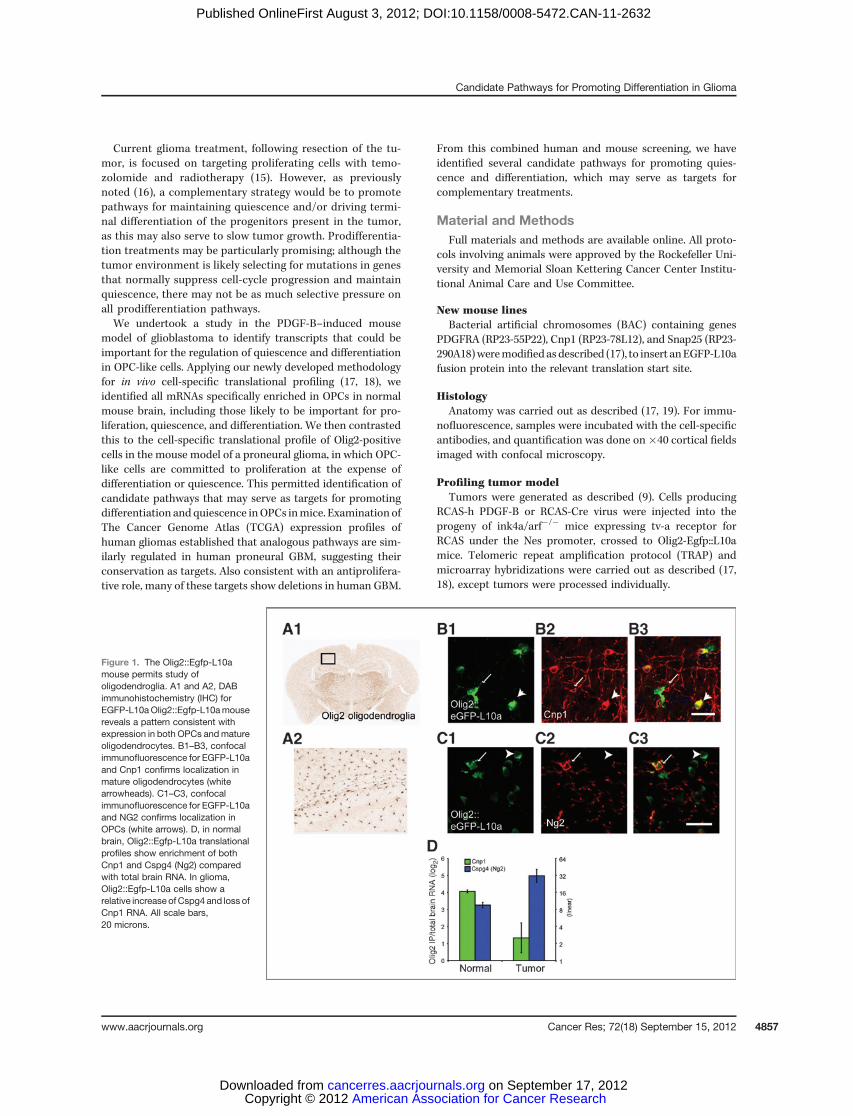
Figure 1. The Olig2::Egfp-L10amouse permits study ofoligodendroglia. A1 and A2, DABimmunohistochemistry (IHC) forEGFP-L10aOlig2::Egfp-L10amousereveals a pattern consistent withexpression in both OPCs andmatureoligodendrocytes. B1–B3, confocalimmunofluorescence for EGFP-L10aand Cnp1 confirms localization inmature oligodendrocytes (whitearrowheads). C1–C3, confocalimmunofluorescence for EGFP-L10aand NG2 confirms localization inOPCs (white arrows). D, in normalbrain, Olig2::Egfp-L10a translationalprofiles show enrichment of bothCnp1 and Cspg4 (Ng2) comparedwith total brain RNA. In glioma,Olig2::Egfp-L10a cells show arelative increase ofCspg4 and loss ofCnp1 RNA. All scale bars,20 microns.
Candidate Pathways for Promoting Differentiation in Glioma
www.aacrjournals.org Cancer Res; 72(18) September 15, 2012 4857
American Association for Cancer Research Copyright © 2012 on September 17, 2012cancerres.aacrjournals.orgDownloaded from
Published OnlineFirst August 3, 2012; DOI:10.1158/0008-5472.CAN-11-2632
Microarray analysisMicroarray data were analyzed with Bionconductor module
of the R statistical package, normalized as described (17, 20, 21)and deposited at GEO (GSE30626; ref. 22). To identifymessagesspecific to each cell type specificity indices (pSI) were calcu-lated as described (18). Transcripts with pSI < 0.05 wereselected for further analysis.
Heat maps and hierarchical clustering were carried outin R. All color-coded scatter plots show only top 50 tran-scripts for each cell type, but all statistics were conducted
on full lists (Supplementary Table S1). Differentiation orquiescence (DorQ) candidates were selected as those tran-scripts from the OPC list that were 2-fold higher in theaverage of the Olig2-Egfp::L10a normal cortex than theaverage of all 12 Olig2-Egfp::L10a tumor samples. Statisticalcomparisons were conducted with LIMMA module ofBioconductor.
For cross-species comparisons, human and mouse homo-logs were mapped by Gene Symbol. For DorQ candidates,mapping was confirmed by Blating mouse protein to human
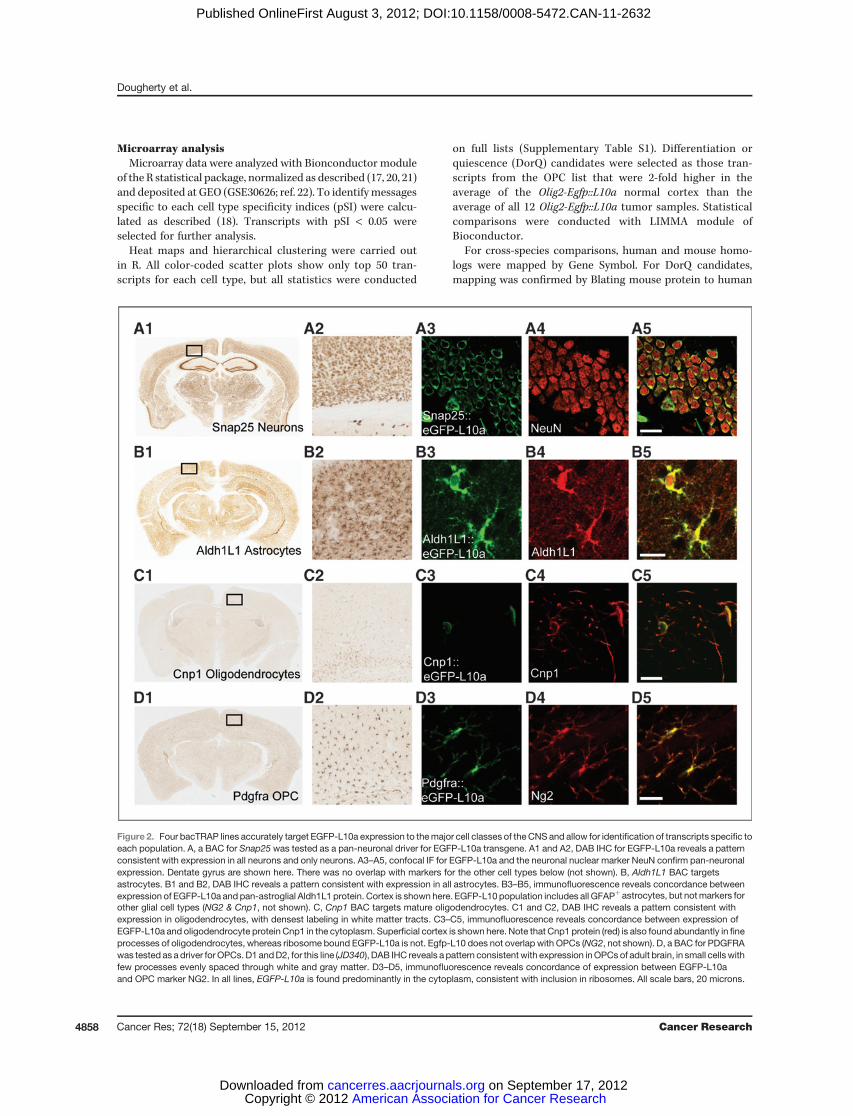
Figure 2. Four bacTRAP lines accurately target EGFP-L10a expression to themajor cell classes of the CNS and allow for identification of transcripts specific toeach population. A, a BAC for Snap25 was tested as a pan-neuronal driver for EGFP-L10a transgene. A1 and A2, DAB IHC for EGFP-L10a reveals a patternconsistent with expression in all neurons and only neurons. A3–A5, confocal IF for EGFP-L10a and the neuronal nuclear marker NeuN confirm pan-neuronalexpression. Dentate gyrus are shown here. There was no overlap with markers for the other cell types below (not shown). B, Aldh1L1 BAC targetsastrocytes. B1 and B2, DAB IHC reveals a pattern consistent with expression in all astrocytes. B3–B5, immunofluorescence reveals concordance betweenexpression of EGFP-L10a and pan-astroglial Aldh1L1 protein. Cortex is shown here. EGFP-L10 population includes all GFAPþ astrocytes, but notmarkers forother glial cell types (NG2 & Cnp1, not shown). C, Cnp1 BAC targets mature oligodendrocytes. C1 and C2, DAB IHC reveals a pattern consistent withexpression in oligodendrocytes, with densest labeling in white matter tracts. C3–C5, immunofluorescence reveals concordance between expression ofEGFP-L10a and oligodendrocyte protein Cnp1 in the cytoplasm. Superficial cortex is shown here. Note that Cnp1 protein (red) is also found abundantly in fineprocesses of oligodendrocytes, whereas ribosome bound EGFP-L10a is not. Egfp-L10 does not overlap with OPCs (NG2, not shown). D, a BAC for PDGFRAwas tested as a driver for OPCs. D1 andD2, for this line (JD340), DAB IHC reveals a pattern consistent with expression inOPCs of adult brain, in small cells withfew processes evenly spaced through white and gray matter. D3–D5, immunofluorescence reveals concordance of expression between EGFP-L10aand OPC marker NG2. In all lines, EGFP-L10a is found predominantly in the cytoplasm, consistent with inclusion in ribosomes. All scale bars, 20 microns.
Dougherty et al.
Cancer Res; 72(18) September 15, 2012 Cancer Research4858
American Association for Cancer Research Copyright © 2012 on September 17, 2012cancerres.aacrjournals.orgDownloaded from
Published OnlineFirst August 3, 2012; DOI:10.1158/0008-5472.CAN-11-2632
genome (UCSC). Human microarray data were downloadedfrom TCGA and normalized as described (4).
Gene ontology analysisFor each cell type, all gene symbols formessages with a pSI <
0.05 were analyzed with BINGO (23). Biologic Process GOcategories were evaluated to identify those with P < 0.01 usingthe hypergeometric test and Benjamini–Hochberg correction.
Neurosphere cell culturePrimary neurosphere cultures from tumors and wild-type
mice were generated as described (24). Neurosphereswere grown in 20 mg/mL EGF and 10 mg/mL bFGF, anduridine 50-diphosphate sodium salt (UDP; 10–50 mmol/L),UDP glucose (100 mmol/L), and leukotriene D4 (LTD4;100 nmol/L) were added on a daily basis. All data are averageof cultures from 4 independent mous tumors, counted intriplicate in wells of 100 to 1,000 spheres. Counts werenormalized within each tumor to the number of spheres in0 UDP condition.
ResultsWe recently developed the TRAP strategy, which allows
profiling of all mRNAs bound to ribosomes in defined cell
populations. This strategy entails using BAC transgenesis toexpress EGFP fused to the ribosomal protein L10a under thecontrol of a "driver" gene specific to certain cell types in thebrain. Thus, the cells of interest contain ribosomes with anEGFP tag enabling affinity purification of all ribosome-asso-ciated mRNA. We have generated and characterized bacTRAPmouse lines for a variety of cell types, including the Olig2-positive oligodendroglia (17).
Because Olig2 is frequently overexpressed in human glioma(25) and high levels of expressionmay particularly characterizethe proneural subtype (2). We recently examined the expres-sion of neural lineage markers, proliferation markers, andtranslational profile of Olig2::Egfp-L10a in our mouse modelof glioma (12). This work revealed a severe perturbation of thetranslational profile of Olig2-positive cells. In a normal Olig2::Egfp-L10a mouse, both NG2-positive OPCs and 20,30-cyclicnucleotide 30-phosphodiesterase1 (Cnp1)-positive mature oli-godendrocytes express EGFP-L 10a (Fig. 1A–C; ref. 17). Thetumor translational profiles for theOlig2::Egfp-L10a cells reveala strong increase in translation forCspg4 (the RNA correspond-ing to NG2) and decrease in Cnp1, relative to normal Olig2::Egfp-L10a cells (Fig. 1D). Although the data come from only 2markers, it suggests the preponderance of Olig2::Egfp-L10a cells are in an immature state within the tumor. To investigate thisthoroughly, we decided to identify the translational profile of
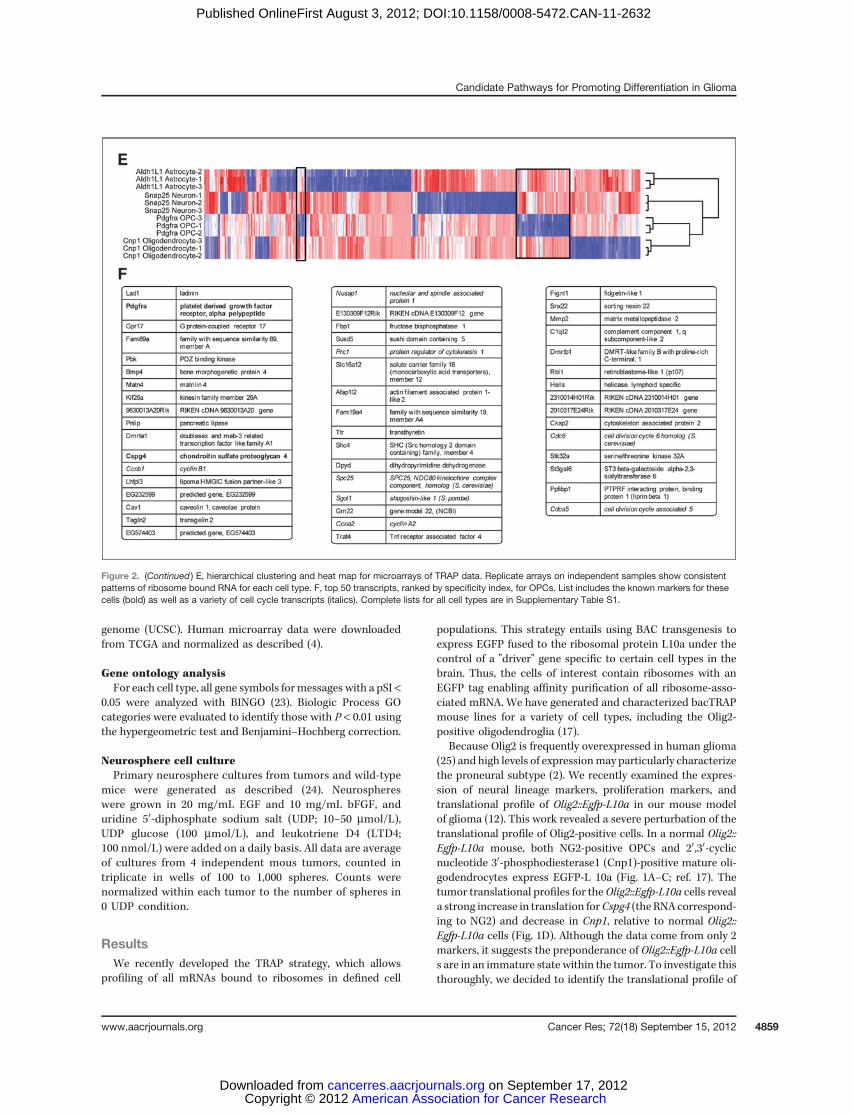
Figure 2. (Continued ) E, hierarchical clustering and heat map for microarrays of TRAP data. Replicate arrays on independent samples show consistentpatterns of ribosome bound RNA for each cell type. F, top 50 transcripts, ranked by specificity index, for OPCs. List includes the known markers for thesecells (bold) as well as a variety of cell cycle transcripts (italics). Complete lists for all cell types are in Supplementary Table S1.
Candidate Pathways for Promoting Differentiation in Glioma
www.aacrjournals.org Cancer Res; 72(18) September 15, 2012 4859
American Association for Cancer Research Copyright © 2012 on September 17, 2012cancerres.aacrjournals.orgDownloaded from
Published OnlineFirst August 3, 2012; DOI:10.1158/0008-5472.CAN-11-2632
normal adult OPCs. With this data for comparison, we wouldthen be able to determine whether Olig2::Egfp-L10a are in amore OPC-like state in the tumor, and if so, we could thenexamine how this state may differ from normal OPCs, as thesedifferences may regulate cell fate choices and represent poten-tial treatment strategies.
Identification of the translational profile of the normaloligodendrocyte progenitors
Identification of transcripts that are specific to OPCsrequires not only knowledge of their profile but also the profiles
of the other cell types. The brain contains 4 major classes ofcells: neurons, astrocytes, and 2 types of oligodendroglia;mature oligodendrocytes and OPCs. We have previouslygenerated and thoroughly characterizedmouse lines that allowtranslational profiling from astrocytes (Aldh1L1::Egfp-L10a),mature oligodendrocytes (Cnp1::Egfp-L10a), and the pan-oli-godendroglial line (Olig2::Egfp-L10a; ref. 17). However, to iden-tify those mRNAs that are enriched in each of the major cellclasses, we generated additional bacTRAP mouse lines target-ing all neurons (Snap25::Egfp-L10a), as well as Pdgfra expres-sing cells (Pdgfra::Egfp-L10a).
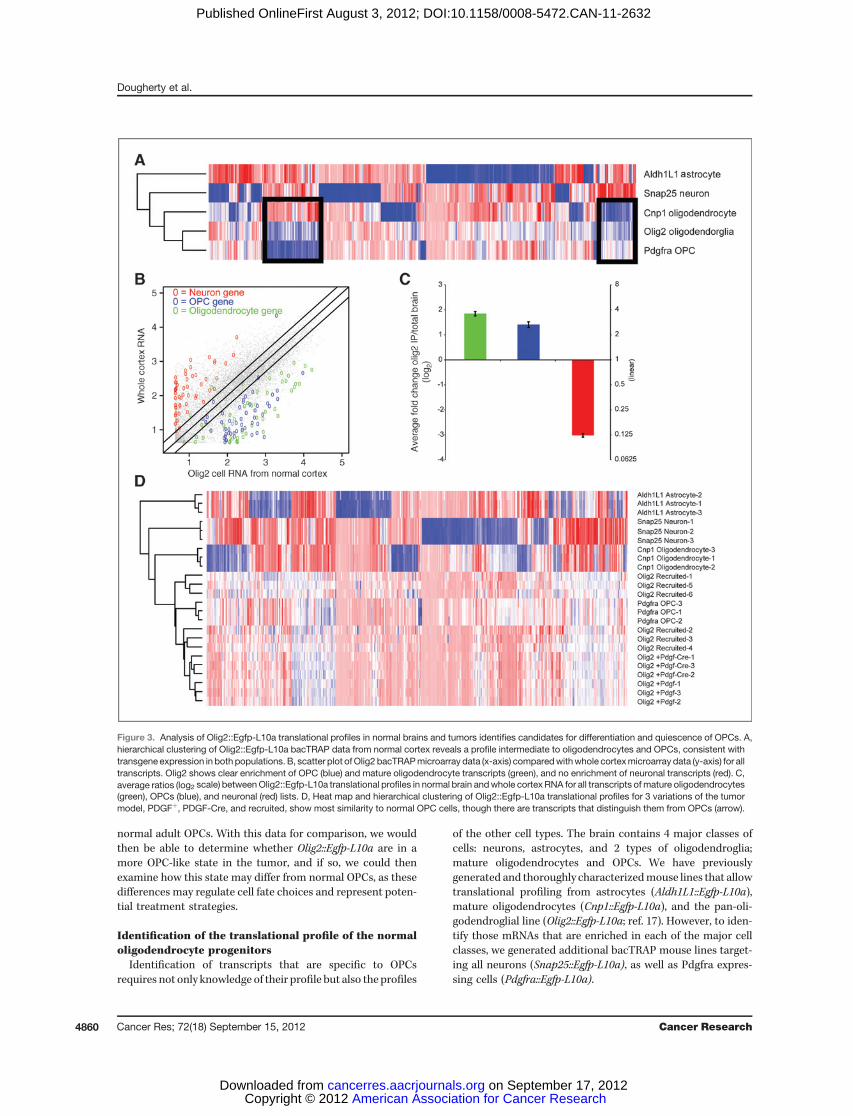
Figure 3. Analysis of Olig2::Egfp-L10a translational profiles in normal brains and tumors identifies candidates for differentiation and quiescence of OPCs. A,hierarchical clustering of Olig2::Egfp-L10a bacTRAP data from normal cortex reveals a profile intermediate to oligodendrocytes and OPCs, consistent withtransgene expression in both populations. B, scatter plot ofOlig2 bacTRAPmicroarray data (x-axis) comparedwithwhole cortexmicroarray data (y-axis) for alltranscripts. Olig2 shows clear enrichment of OPC (blue) and mature oligodendrocyte transcripts (green), and no enrichment of neuronal transcripts (red). C,average ratios (log2 scale) betweenOlig2::Egfp-L10a translational profiles in normal brain andwhole cortex RNA for all transcripts ofmature oligodendrocytes(green), OPCs (blue), and neuronal (red) lists. D, Heat map and hierarchical clustering of Olig2::Egfp-L10a translational profiles for 3 variations of the tumormodel, PDGFþ, PDGF-Cre, and recruited, show most similarity to normal OPC cells, though there are transcripts that distinguish them from OPCs (arrow).
Dougherty et al.
Cancer Res; 72(18) September 15, 2012 Cancer Research4860
American Association for Cancer Research Copyright © 2012 on September 17, 2012cancerres.aacrjournals.orgDownloaded from
Published OnlineFirst August 3, 2012; DOI:10.1158/0008-5472.CAN-11-2632
To target neurons, we selected 2 genes as putative pan-neuronal drivers, complexin 1 (Cplx1) and synaptosomal-associated protein 25 (Snap25). Examination of multipleEGFP-L10a lines for both constructs revealed that Snap25was the brighter and more reliable construct and thusselected for further analysis.To target PDGF-responsive OPCs, we tested a BAC cov-
ering the Pdgfra gene. Multiple lines showed similar but notidentical patterns of expression, targeting NG2-positive cellswith varying intensity, but also subpopulations of neurons insome lines, as well as robust expression in the choroidplexus. For this study, we selected the line that targetedcortical NG2-positive OPCs most specifically. For each of the4 cell classes, the lines used for further experiments werecarefully characterized with confocal immunofluorescenceto confirm accuracy of transgene expression and identity ofeach cell type (Fig. 2A–D).For the newly generated lines, we quantified, from randomly
collected confocal images of mouse cortex, the overlap of GFPwith 40,6-diamidino-2-phenylindole (DAPI) and with each of 2cell-specific markers. For Snap25, we labeled with pan-neuro-nal maker NeuN (Fig. 2A) or Tbr1 (not shown), a markerof deep layer neurons. In cortex, of 663 DAPI-positive cellscounted, 57.6% were positive for GFP, consistent with thelarge proportion of neurons in this tissue. A total of 96.8% ofGFP-positive cells were positive for NeuN, and 98.6% of NeuN-positive cells were positive for GFP. For Tbr1, of 592 Tbr1-positive cells counted, 99.8%were labeledwithGFP. For Pdgfra,of 3,015 DAPI-positive cells counted, 8.2% were positive forGFP, consistent with the relative rarity of the OPC cells. A totalof 88.8% of GFP-positive cells were labeled with NG2, andthough receptors are often localized on processes far from theGFP-labeled cell bodies, obfuscating counting, at least 51.4% ofGFP-positive cells were clearly labeled with Pdgfra antibodies.Most of these cells distinctly had the morphology of OPCs,though both Ng2 and Pdgfra label pericytes as well. A subsetof Pdgfra::eGFP-L10a cells also labeled with the proliferativemarker Ki-67 (<2%), consistent with expectation for thesecells (8).
For each new line, TRAP was conducted in triplicate onpooled cortices from 2 to 3 mice, and cell-specific polysomalmRNAs were queried with microarrays. Replicate arraysshowed high reproducibility (minimum correlation ¼ 0.978),though each cell type had a distinct profile (Fig. 2E). We usedthe specificity index statistic (21) to select for those transcriptssignificantly enriched in each cell population (Snap25, Ald1L1,Cnp1, and Pdgfra) relative to the other 3. These lists includedknown markers for each cell type (Supplementary Table S1,and Fig. 2F). The non-OPC cell types had significant over-representation of Gene Ontology (GO) categories consistentwith their known roles in synaptic transmission (Neurons,Supplementary Fig. S2A), myelination (Oligodendrocytes,Supplementary Fig. S2B), or a role in lipid metabolism(Astrocytes, Supplementary Fig. S2C; ref. 26). As the OPCpopulation is the most proliferative population in the cortex(8), the transcripts specific to OPCs showed a highly signif-icant overrepresentation of categories related to the cellcycle (P < 1E-7, hypergeometric test with Benjamini–Hoch-berg correction; Supplementary Fig. S3), as well as in cat-egories related to protein phosphorylation and biopolymermodification (P < 1E-3). For all cell types, there were also avariety of transcripts not currently annotated in knownpathways. Overall, this evidence, along with presence of allknown markers for OPCs on the list, suggests we haveidentified an OPC-specific profile, as well as generated auseful tool for assessment of these cells in vivo.
Characterization of Olig2-positive cells in mouse modelof glioblastoma
We next applied our tools to characterize the oligodendrog-lial lineage cells in the context of a mouse model of GBM.Murine PDGF-driven GBM show high expression levels ofOlig2 (Supplementary Fig. S1; refs. 12, 43). In a normal brain,the transgenic line Olig2::Egfp-L10a labels all oligodendroglialcells: both the NG2-positive OPCs and Cnp1-positive myelinat-ing oligodendrocytes (Fig. 1A–C and ref. 17). Hierarchicalclustering of TRAP microarray data positions the profile ofthese cells between that of mature oligodendrocytes and
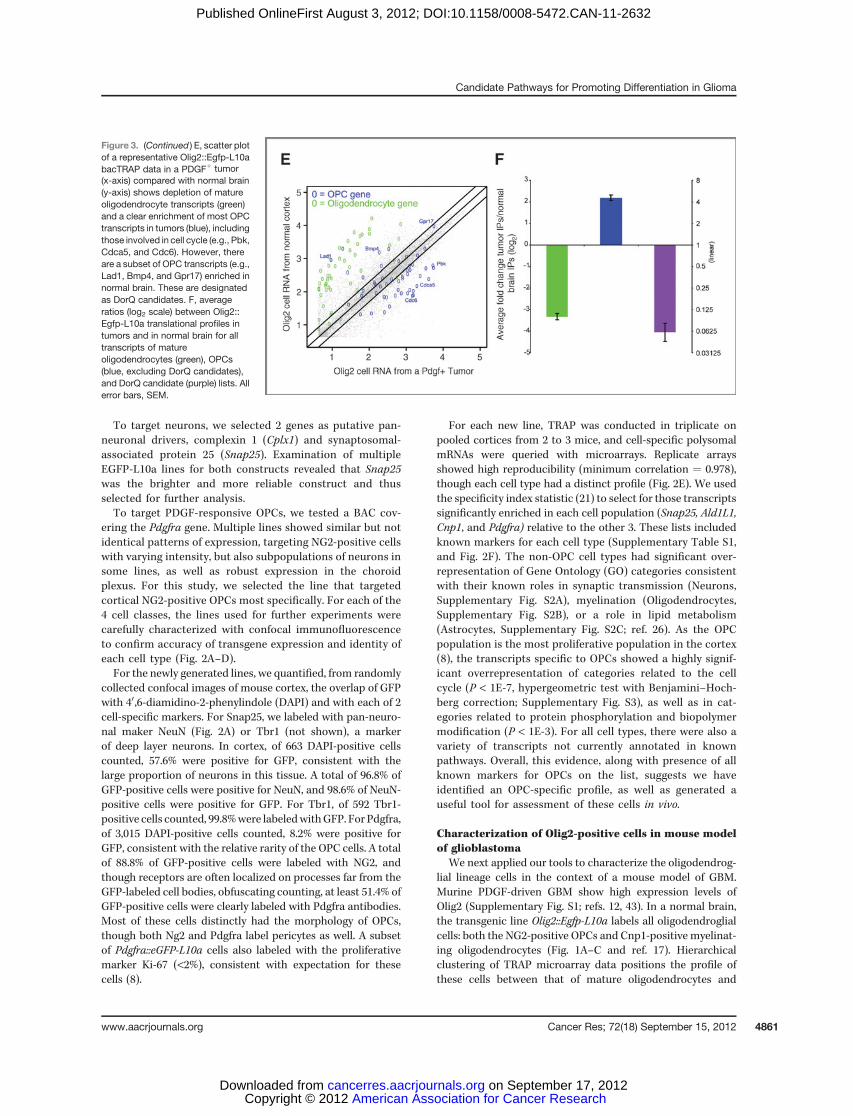
Figure 3. (Continued ) E, scatter plotof a representative Olig2::Egfp-L10abacTRAP data in a PDGFþ tumor(x-axis) compared with normal brain(y-axis) shows depletion of matureoligodendrocyte transcripts (green)and a clear enrichment of most OPCtranscripts in tumors (blue), includingthose involved in cell cycle (e.g., Pbk,Cdca5, and Cdc6). However, thereare a subset of OPC transcripts (e.g.,Lad1, Bmp4, and Gpr17) enriched innormal brain. These are designatedas DorQ candidates. F, averageratios (log2 scale) between Olig2::Egfp-L10a translational profiles intumors and in normal brain for alltranscripts of matureoligodendrocytes (green), OPCs(blue, excluding DorQ candidates),and DorQ candidate (purple) lists. Allerror bars, SEM.
Candidate Pathways for Promoting Differentiation in Glioma
www.aacrjournals.org Cancer Res; 72(18) September 15, 2012 4861
American Association for Cancer Research Copyright © 2012 on September 17, 2012cancerres.aacrjournals.orgDownloaded from
Published OnlineFirst August 3, 2012; DOI:10.1158/0008-5472.CAN-11-2632

PDGFRA-positive OPCs (Fig. 3A). Comparison of the Olig2::Egfp-L10a profile to whole cortex reveals that it is enriched inboth the OPCs and myelinating oligodendrocyte transcripts(Fig. 3B), consistent with it covering the entire lineage.
First, we translationally profiled Ntv-a/ink4a/arfþ/� Olig2::EgfpL10a cells in primary tumors from our model (PDGF-Btumors). Hierarchical clustering revealed that these cells aremost similar to OPCs of the normal brain. To test this obser-vation, we repeated the analysis in 2 additional variations of ourtumor model: primary tumors from this model with additionaldeletion of Pten (PDGF-Cre tumors), and in tumors induced bytransplanting EGFP-L10a–negative tumor cells from PDGF-driven primary murine tumors into Ntv-a/ink4a/arfþ/�Olig2::Egfp-L10amice (recruited). Although this increases the variabil-ity among the tumor samples, all samples cluster most closelywith OPCs (Fig. 3D). This provides systematic evidence thatOlig2::Egfp-L10a cells are in a more OPC-like state in glioma.
Themost parsimonious explanation of these findings is thatthe excess of PDGF-B ligand produced by the Nes-positive cellhas driven the recruitment and expansion of the proliferatingPDGFRA-positive cells, at the expense of quiescence or differ-entiation. To examine this, we plotted the translational profileof Olig2::Egfp-L10a cells in a normalmouse cortex versusOlig2::Egfp-L10a cells in tumors and labeled those transcripts that arespecific to either OPCs or mature oligodendrocytes. On aver-age, the tumor contains a clear enrichment of transcriptsspecific to OPCs and depletion of transcripts found in matureoligodendrocytes (Fig. 3E). Not surprisingly, the subset of OPCtranscripts identified by GO as involved in cell cycle/cellproliferation, such as Melk (27) and Pbk (28), are highlyenriched in the tumor samples, consistent with the Olig2:Egfp-L10a population being in a highly proliferative state insideof the tumor. However, surprisingly, there is also a subset ofOPC transcripts that seem enriched in the normal brain.
Identification of differentiation or quiescence candidatetranscripts in OPCs
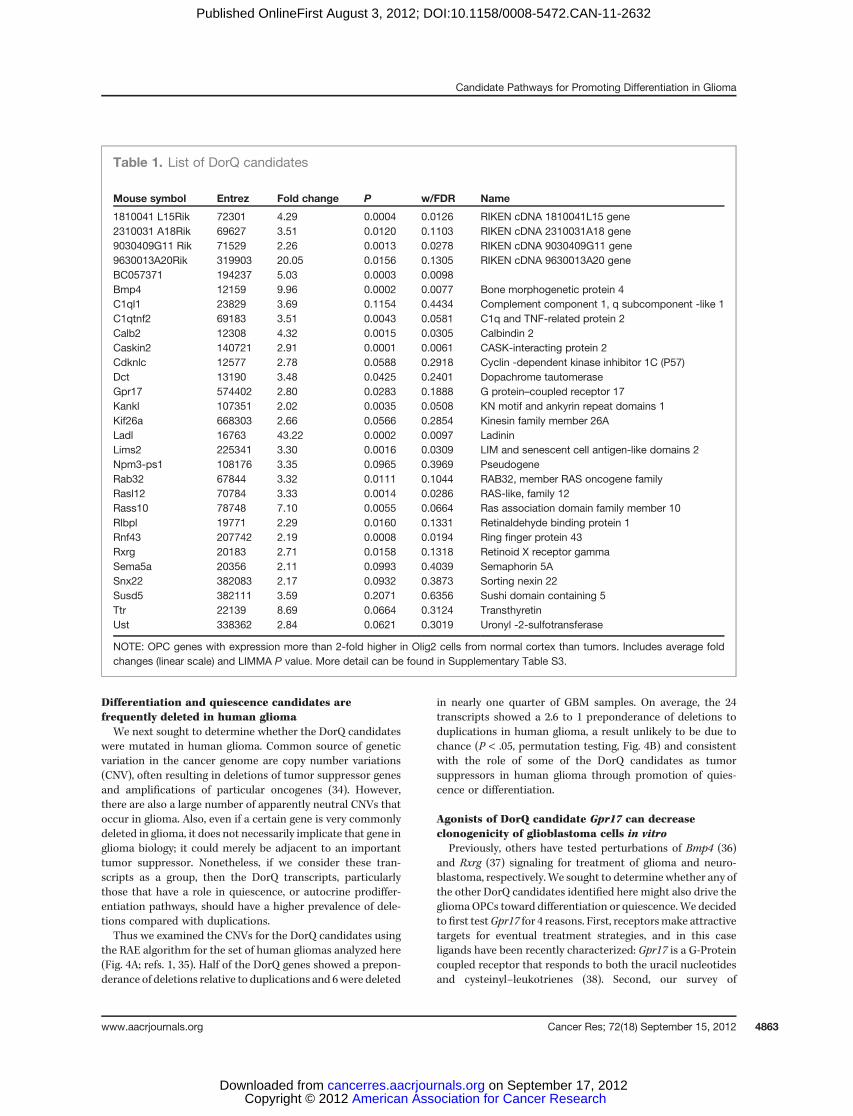
In a normal brain, OPCs may proliferate, remain quiescent,or differentiate, but in the context of a tumor, they seem toproliferate at the expense of differentiation or quiescence.Comparison of the transcriptional profiles of Olig2::Egfp-L10a cells from normal brain and tumor pointed to a subsetof 28 transcripts translated greater than 2-fold higher innormal brain (Fig. 3F, Table 1). We hypothesized that thissubset contains those transcripts used in either differentiationor quiescence in oligodendroglia.
Careful examination of current literature supports thishypothesis. For example, Cdkn1c is required for the regulationof cell cycle and specification of OPCs (29), whereas Sema5a,an oligodendrocyte-specific semaphorin, plays a role in axongrowth inhibition (30). G-coupled protein receptor Gpr17has the role in myelination during development (31). OtherDorQ candidate transcripts are known to be expressed inoligodendroglia, although their role is unclear. They includeRlbp (32) and C1qtnf2, whose mRNA expression pattern isconsistent in part with the oligodendrocyte expression (33).Additional screening of the expression patterns of DorQcandidate transcripts in Allen Brain Atlas andGene Expression
Nervous System Atlas (GENSAT) confirmed that Gpr17, Bmp4,Rlbp1, Rnf43, 90304409g11Rik, Rxrg, Caskin2, Rasl12, and2310031a18Rik expression patterns are consistent with OPClocalization. The presence of these transcripts in OPCs sug-gests that the DorQ candidates may have some relevance ascomplementary targets for treatment, if they are conserved inhumans.
Differentiation and quiescence candidate transcriptsshow similar expression pattern in human glioma
As human proneural tumors are characterized by anabundance of PDGF-B and activation of PDGFRA signalingpathways, we sought to determine whether they containeda preponderance of OPC transcripts. The results, summa-rized in Supplementary Table S2, Supplementary Fig. S2,and Supplementary Fig. S4, showed that human proneuralspecific gene list has a highly significant enrichment ofOPC transcripts. We carried out hierarchical clusteringof the human gliomas using the OPC transcripts (excludingthose categorized by GO as involved in cell cycle or DNAreplication as enhanced cell cycle is common to all glio-blastoma subtypes). Using just these transcripts, most of theproneural tumors clustered into a single large branch,suggesting that these OPC transcripts are sufficient todistinguish the proneural subtype (Supplementary Fig.S4B). On average, the non–cell cycle OPC transcripts areexpressed at a higher relative level in proneural tumors(Supplementary Fig. S4C), though they are present at detect-able levels in all subtypes. This suggests that part of whatdistinguishes the proneural subtype from the other typesmay be the relative abundance of OPC-like cells.
Although the analysis above showed that OPC transcripts ingeneral have potential relevance to human glioma, we nextexamined the DorQ candidate subset of OPC transcripts spe-cifically. We first analyzed the human TCGA dataset to deter-mine whether these transcripts show the same pattern ofexpression that is present in the mouse tumors. Because theDorQ candidates were identified by using translational profilingdata specifically for Olig2::Egfp-L10a cells, which is unavailablein the human samples, to generate an analysis analogous tohuman samples, we examined the expression of the DorQcandidates in total RNA profiles from mouse gliomas andnormal mice (Supplementary Fig. S5A). With this method allOPC transcripts, including the DorQ candidates, are somewhatenriched in tumors compared with normal brain. However,the DorQ candidate transcripts are significantly less increasedin tumors than the other OPC transcripts are (SupplementaryFig. S5D, t test P < 1E-6.). We repeated this same analysis inhumanproneural tumors comparedwithnormal human cortex,using the human homologs of these DorQ transcripts (Supple-mentary Fig. S5B). We find that, as with the mouse, the DorQcandidate transcripts are enriched, but significantly less so thanthe other OPC transcripts (P < 0.05; Supplementary Fig. S5D).Overall, the log2 fold change valuesofglioma/normalbrain for allOPC transcripts is correlated at 0.62 (Pearsons correlation)across mouse and human comparisons, suggesting that thegeneral pattern of expression of these pathways is conservedacross mouse and human glioma (Supplementary Fig. S5C).
Dougherty et al.
Cancer Res; 72(18) September 15, 2012 Cancer Research4862
American Association for Cancer Research Copyright © 2012 on September 17, 2012cancerres.aacrjournals.orgDownloaded from
Published OnlineFirst August 3, 2012; DOI:10.1158/0008-5472.CAN-11-2632
Differentiation and quiescence candidates arefrequently deleted in human gliomaWe next sought to determine whether the DorQ candidates
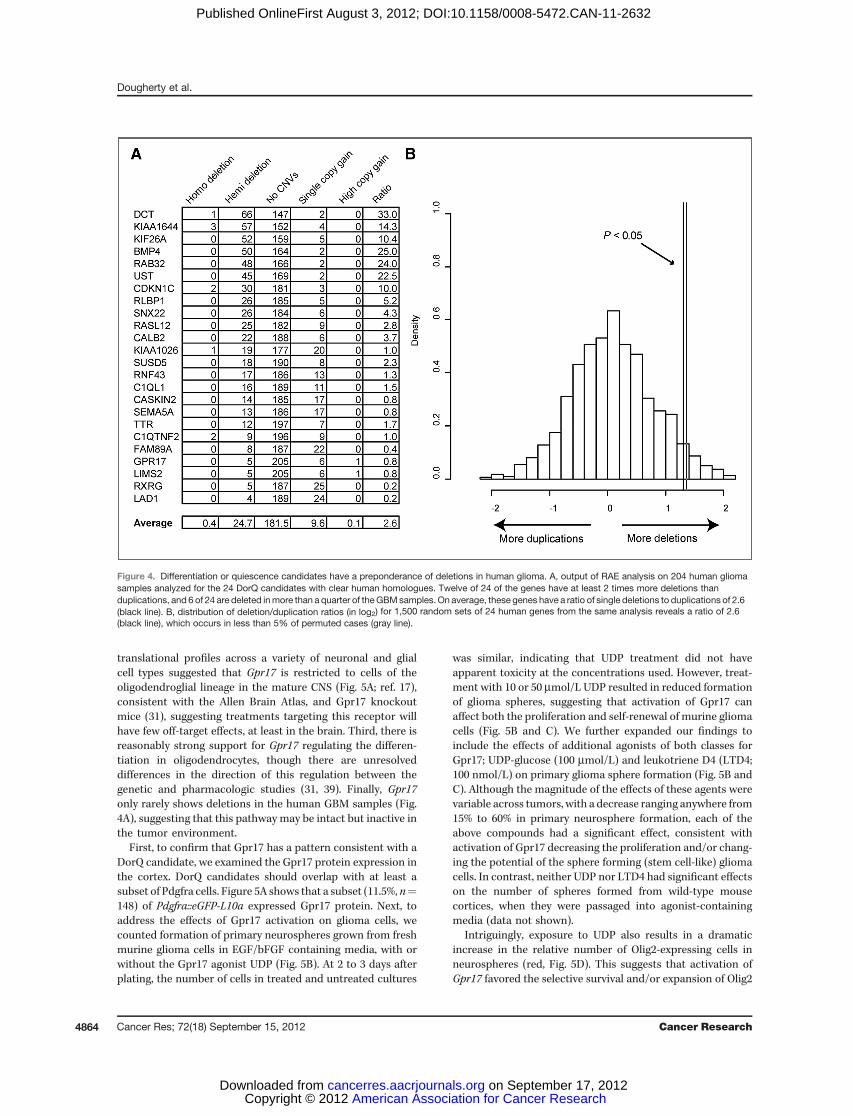
were mutated in human glioma. Common source of geneticvariation in the cancer genome are copy number variations(CNV), often resulting in deletions of tumor suppressor genesand amplifications of particular oncogenes (34). However,there are also a large number of apparently neutral CNVs thatoccur in glioma. Also, even if a certain gene is very commonlydeleted in glioma, it does not necessarily implicate that gene inglioma biology; it could merely be adjacent to an importanttumor suppressor. Nonetheless, if we consider these tran-scripts as a group, then the DorQ transcripts, particularlythose that have a role in quiescence, or autocrine prodiffer-entiation pathways, should have a higher prevalence of dele-tions compared with duplications.Thus we examined the CNVs for the DorQ candidates using
the RAE algorithm for the set of human gliomas analyzed here(Fig. 4A; refs. 1, 35). Half of the DorQ genes showed a prepon-derance of deletions relative to duplications and 6were deleted
in nearly one quarter of GBM samples. On average, the 24transcripts showed a 2.6 to 1 preponderance of deletions toduplications in human glioma, a result unlikely to be due tochance (P < .05, permutation testing, Fig. 4B) and consistentwith the role of some of the DorQ candidates as tumorsuppressors in human glioma through promotion of quies-cence or differentiation.
Agonists of DorQ candidate Gpr17 can decreaseclonogenicity of glioblastoma cells in vitro
Previously, others have tested perturbations of Bmp4 (36)and Rxrg (37) signaling for treatment of glioma and neuro-blastoma, respectively.We sought to determine whether any ofthe other DorQ candidates identified here might also drive thegliomaOPCs toward differentiation or quiescence.We decidedto first testGpr17 for 4 reasons. First, receptorsmake attractivetargets for eventual treatment strategies, and in this caseligands have been recently characterized: Gpr17 is a G-Proteincoupled receptor that responds to both the uracil nucleotidesand cysteinyl–leukotrienes (38). Second, our survey of
Table 1. List of DorQ candidates
Mouse symbol Entrez Fold change P w/FDR Name
1810041 L15Rik 72301 4.29 0.0004 0.0126 RIKEN cDNA 1810041L15 gene2310031 A18Rik 69627 3.51 0.0120 0.1103 RIKEN cDNA 2310031A18 gene9030409G11 Rik 71529 2.26 0.0013 0.0278 RIKEN cDNA 9030409G11 gene9630013A20Rik 319903 20.05 0.0156 0.1305 RIKEN cDNA 9630013A20 geneBC057371 194237 5.03 0.0003 0.0098Bmp4 12159 9.96 0.0002 0.0077 Bone morphogenetic protein 4C1ql1 23829 3.69 0.1154 0.4434 Complement component 1, q subcomponent -like 1C1qtnf2 69183 3.51 0.0043 0.0581 C1q and TNF-related protein 2Calb2 12308 4.32 0.0015 0.0305 Calbindin 2Caskin2 140721 2.91 0.0001 0.0061 CASK-interacting protein 2Cdknlc 12577 2.78 0.0588 0.2918 Cyclin -dependent kinase inhibitor 1C (P57)Dct 13190 3.48 0.0425 0.2401 Dopachrome tautomeraseGpr17 574402 2.80 0.0283 0.1888 G protein–coupled receptor 17Kankl 107351 2.02 0.0035 0.0508 KN motif and ankyrin repeat domains 1Kif26a 668303 2.66 0.0566 0.2854 Kinesin family member 26ALadl 16763 43.22 0.0002 0.0097 LadininLims2 225341 3.30 0.0016 0.0309 LIM and senescent cell antigen-like domains 2Npm3-ps1 108176 3.35 0.0965 0.3969 PseudogeneRab32 67844 3.32 0.0111 0.1044 RAB32, member RAS oncogene familyRasl12 70784 3.33 0.0014 0.0286 RAS-like, family 12Rass10 78748 7.10 0.0055 0.0664 Ras association domain family member 10Rlbpl 19771 2.29 0.0160 0.1331 Retinaldehyde binding protein 1Rnf43 207742 2.19 0.0008 0.0194 Ring finger protein 43Rxrg 20183 2.71 0.0158 0.1318 Retinoid X receptor gammaSema5a 20356 2.11 0.0993 0.4039 Semaphorin 5ASnx22 382083 2.17 0.0932 0.3873 Sorting nexin 22Susd5 382111 3.59 0.2071 0.6356 Sushi domain containing 5Ttr 22139 8.69 0.0664 0.3124 TransthyretinUst 338362 2.84 0.0621 0.3019 Uronyl -2-sulfotransferase
NOTE: OPC genes with expression more than 2-fold higher in Olig2 cells from normal cortex than tumors. Includes average foldchanges (linear scale) and LIMMA P value. More detail can be found in Supplementary Table S3.
Candidate Pathways for Promoting Differentiation in Glioma
www.aacrjournals.org Cancer Res; 72(18) September 15, 2012 4863
American Association for Cancer Research Copyright © 2012 on September 17, 2012cancerres.aacrjournals.orgDownloaded from
Published OnlineFirst August 3, 2012; DOI:10.1158/0008-5472.CAN-11-2632
translational profiles across a variety of neuronal and glialcell types suggested that Gpr17 is restricted to cells of theoligodendroglial lineage in the mature CNS (Fig. 5A; ref. 17),consistent with the Allen Brain Atlas, and Gpr17 knockoutmice (31), suggesting treatments targeting this receptor willhave few off-target effects, at least in the brain. Third, there isreasonably strong support for Gpr17 regulating the differen-tiation in oligodendrocytes, though there are unresolveddifferences in the direction of this regulation between thegenetic and pharmacologic studies (31, 39). Finally, Gpr17only rarely shows deletions in the human GBM samples (Fig.4A), suggesting that this pathway may be intact but inactive inthe tumor environment.
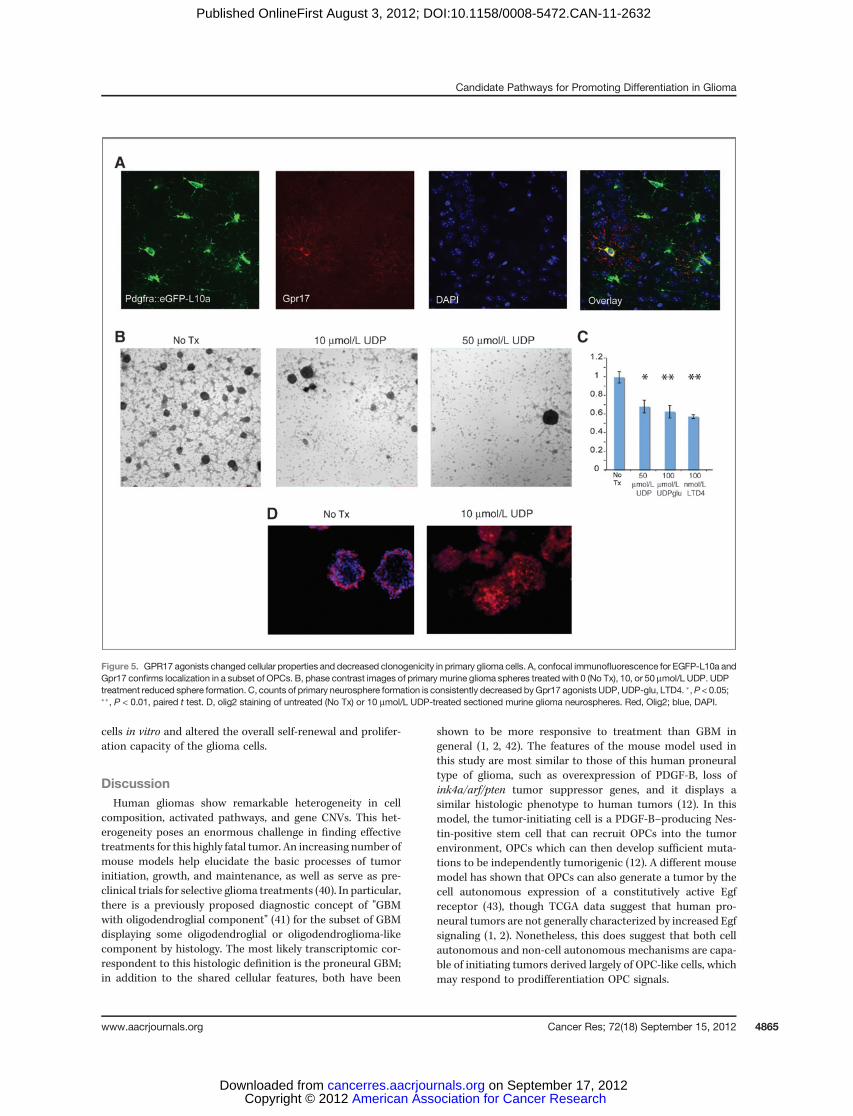
First, to confirm that Gpr17 has a pattern consistent with aDorQ candidate, we examined the Gpr17 protein expression inthe cortex. DorQ candidates should overlap with at least asubset of Pdgfra cells. Figure 5A shows that a subset (11.5%, n¼148) of Pdgfra::eGFP-L10a expressed Gpr17 protein. Next, toaddress the effects of Gpr17 activation on glioma cells, wecounted formation of primary neurospheres grown from freshmurine glioma cells in EGF/bFGF containing media, with orwithout the Gpr17 agonist UDP (Fig. 5B). At 2 to 3 days afterplating, the number of cells in treated and untreated cultures
was similar, indicating that UDP treatment did not haveapparent toxicity at the concentrations used. However, treat-ment with 10 or 50 mmol/L UDP resulted in reduced formationof glioma spheres, suggesting that activation of Gpr17 canaffect both the proliferation and self-renewal of murine gliomacells (Fig. 5B and C). We further expanded our findings toinclude the effects of additional agonists of both classes forGpr17; UDP-glucose (100 mmol/L) and leukotriene D4 (LTD4;100 nmol/L) on primary glioma sphere formation (Fig. 5B andC). Although the magnitude of the effects of these agents werevariable across tumors, with a decrease ranging anywhere from15% to 60% in primary neurosphere formation, each of theabove compounds had a significant effect, consistent withactivation of Gpr17 decreasing the proliferation and/or chang-ing the potential of the sphere forming (stem cell-like) gliomacells. In contrast, neither UDP nor LTD4 had significant effectson the number of spheres formed from wild-type mousecortices, when they were passaged into agonist-containingmedia (data not shown).
Intriguingly, exposure to UDP also results in a dramaticincrease in the relative number of Olig2-expressing cells inneurospheres (red, Fig. 5D). This suggests that activation ofGpr17 favored the selective survival and/or expansion of Olig2
Figure 4. Differentiation or quiescence candidates have a preponderance of deletions in human glioma. A, output of RAE analysis on 204 human gliomasamples analyzed for the 24 DorQ candidates with clear human homologues. Twelve of 24 of the genes have at least 2 times more deletions thanduplications, and 6 of 24 are deleted inmore than a quarter of theGBMsamples. On average, these genes have a ratio of single deletions to duplications of 2.6(black line). B, distribution of deletion/duplication ratios (in log2) for 1,500 random sets of 24 human genes from the same analysis reveals a ratio of 2.6(black line), which occurs in less than 5% of permuted cases (gray line).
Dougherty et al.
Cancer Res; 72(18) September 15, 2012 Cancer Research4864
American Association for Cancer Research Copyright © 2012 on September 17, 2012cancerres.aacrjournals.orgDownloaded from
Published OnlineFirst August 3, 2012; DOI:10.1158/0008-5472.CAN-11-2632
cells in vitro and altered the overall self-renewal and prolifer-ation capacity of the glioma cells.
DiscussionHuman gliomas show remarkable heterogeneity in cell
composition, activated pathways, and gene CNVs. This het-erogeneity poses an enormous challenge in finding effectivetreatments for this highly fatal tumor. An increasing number ofmouse models help elucidate the basic processes of tumorinitiation, growth, and maintenance, as well as serve as pre-clinical trials for selective glioma treatments (40). In particular,there is a previously proposed diagnostic concept of "GBMwith oligodendroglial component" (41) for the subset of GBMdisplaying some oligodendroglial or oligodendroglioma-likecomponent by histology. The most likely transcriptomic cor-respondent to this histologic definition is the proneural GBM;in addition to the shared cellular features, both have been
shown to be more responsive to treatment than GBM ingeneral (1, 2, 42). The features of the mouse model used inthis study are most similar to those of this human proneuraltype of glioma, such as overexpression of PDGF-B, loss ofink4a/arf/pten tumor suppressor genes, and it displays asimilar histologic phenotype to human tumors (12). In thismodel, the tumor-initiating cell is a PDGF-B–producing Nes-tin-positive stem cell that can recruit OPCs into the tumorenvironment, OPCs which can then develop sufficient muta-tions to be independently tumorigenic (12). A different mousemodel has shown that OPCs can also generate a tumor by thecell autonomous expression of a constitutively active Egfreceptor (43), though TCGA data suggest that human pro-neural tumors are not generally characterized by increased Egfsignaling (1, 2). Nonetheless, this does suggest that both cellautonomous and non-cell autonomous mechanisms are capa-ble of initiating tumors derived largely of OPC-like cells, whichmay respond to prodifferentiation OPC signals.
Figure 5. GPR17 agonists changed cellular properties and decreased clonogenicity in primary glioma cells. A, confocal immunofluorescence for EGFP-L10a andGpr17 confirms localization in a subset of OPCs. B, phase contrast images of primary murine glioma spheres treated with 0 (No Tx), 10, or 50 mmol/L UDP. UDPtreatment reduced sphere formation. C, counts of primary neurosphere formation is consistently decreased byGpr17 agonists UDP, UDP-glu, LTD4. �,P < 0.05;��, P < 0.01, paired t test. D, olig2 staining of untreated (No Tx) or 10 mmol/L UDP-treated sectioned murine glioma neurospheres. Red, Olig2; blue, DAPI.
Candidate Pathways for Promoting Differentiation in Glioma
www.aacrjournals.org Cancer Res; 72(18) September 15, 2012 4865
American Association for Cancer Research Copyright © 2012 on September 17, 2012cancerres.aacrjournals.orgDownloaded from
Published OnlineFirst August 3, 2012; DOI:10.1158/0008-5472.CAN-11-2632

Here we showed that translational profile of Olig2-positivecells in our model closely resembles the profile of OPC cells,and that OPC transcripts are sufficient to distinguish thehuman proneural glioma subtype. This confirmed the resultsfrom a study that used similar glioblastoma model with thepten/p53 deletion (44). Our data also confirmed the expressionof stem and progenitor cell markers, as well as high expressionof genes involved in cell cycle, in Olig2-positive cells in glio-blastoma, consistent with numerous studies of human tumorsand mouse models (25, 45–50). Our study went a step furthergenerating the list of candidate transcripts, deleted often inhuman gliomas that may serve to promote the differentiationor quiescence of Olig2 cells in the tumor. Of course, futureworkwill need to be done to establish which of these candidates arebona fide prodifferentiation or quiescence factors.
Gene expression and transcriptome analysis could providevaluable information about potential cell-specific therapytargets, and the expression profile of glioma cells has beenextensively studied (reviewed in ref. 51). However, these anal-yses were carried out either on glioma cell lines or using thetotal mRNA from the tumor mass. The pitfall of this approachis that glioma cell lines do not possess the physiologic char-acteristics of the tumor cells in vivo and analysis based on thetotal mRNA extracted from the tumor does not reflect thedifferences in expression profile of specific cell subpopulations.In addition, this approach does not identify the actively trans-lated pool of mRNAs. It has been shown that translationalregulation plays an important role in tumorigenesis (52, 53).Thus, this study is the first that used the bacTRAP method-ology to analyze the actively translatedmRNAs in a cell-specificmanner within the tumor.
The prevalence of hemideletions for DorQ genes in manyhuman glioma samples suggested that they might serve astargets for glioma therapy. Bone morphogenetic factor 4(Bmp4) and 4 other DorQ genes (Kank1, Lims2, Rab32, andRassf10) are frequently epigenetically silenced or deleted inother cancer types, and/or are suggested to function as tumorsuppressors (54–57). Several DorQ candidates were alreadytested either in vivo or in vitro (36, 58, 59). When we tested theactivation of Gpr17 signaling in the murine GBM cells in vitro,our results suggested that Gpr17 agonists might drive thedifferentiation of highly proliferative, uncommitted tumorcells toward oligodendoglial fates. Similarly, Bmp4 has beentested in animalmodel of glioma and human glioblastoma cells(36) as well as in meduloblastoma (60). Cells isolated fromhuman glioblastomas treated with Bmp4 showed decreasedproliferation and increased differentiation in vitro, whereas invivo treatment with Bmp4 blocked the tumor growth. Finally,agonists of the retinoid receptor gamma Rxrg promote differ-entiation in human neuroblastoma (37) and glioblastoma (58)cell lines and have already shown some efficacy as part ofcombination treatments for thyroid cancer (59). In addition,
many of the candidate DorQ transcripts have been detected inother types of tumors (54–57, 60–66), suggesting these candi-dates may have some relevance beyond GBM.
Human glioma tumors contain an abundance of cellsexpressingmarkers of glial progenitors and stem cells, suggest-ing that tumorigenesis involves expansion of a glial progeni-tor–like cell (67). The comparison of the translational profilesof tumors to OPCs and normal mature cells of the neuronal,oligodendroglial, and astrocyte lineage supports this hypoth-esis. The analysis confirmed the similarities of tumor cells withthe immature PDGFRA-positive cells from the normal brain. Itwas also suggested that OPCs might serve as source of gliomas(43, 44, 67–69). Although this study could not provide theinformation about cell-of-origin for gliomas, the approachwould be useful if normal immature glial and stem-like cellprofiles are compared with glioma cells at different time pointsduring tumorigenesis. The mouse line generated for this study(Pdgfra::Egfp-L10a) will provide reproducible access to OPCsfor future translational profiling. However, beyond applica-tions to glioma, this line should permit reproducible in vivoaccess for researchers interested in studies of the basic biologyof these cells both in the normalmouse brain and experimentalmodels of OPC-relevant diseases, such as multiple sclerosis orwhite matter stroke.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: J.D. Dougherty, E.I. Fomchenko, A. MilosevicDevelopment of methodology: E.I. Fomchenko, E. BazzoliAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): J.D. Dougherty, E.I. Fomchenko, E. Schmidt, K.Y.Helmy, E. Bazzoli, E.C. Holland, A. Milosevic, A.A. AkuffoAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): J.D. Dougherty, E.I. Fomchenko, C.W. Brennan, E.C.Holland, A.A. AkuffoWriting, review, and/or revision of the manuscript: J.D. Dougherty, E.I.Fomchenko, A. Milosevic, A.A. AkuffoStudy supervision: A. Milosevic
AcknowledgmentsThe authors thank Victoria Isakova and Daviana Martinez Osorio for their
technical assistance, ElizabethGriggs for image processing, and N. Ramanan andmembers of the N. Heintz laboratory for their advice and support and also thankthe RU Genomics Resource Center and the MSKCC Flow Cytometry ResourceCenter.
Grant SupportThiswork was supported by the Tri-Institutional StemCell Initiative award to
A. Milosevic and E.C. Holland, the NIH (4R00NS067239-03 to J.D. Dougherty) andthe Adelson Medical Research Foundation.
The costs of publication of this article were defrayed in part by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received September 15, 2011; revised July 2, 2012; accepted July 8, 2012;published OnlineFirst August 3, 2012.
References1. TCGARN. Comprehensive genomic characterization defines human
glioblastoma genes and core pathways. Nature 2008;455:1061–8.2. VerhaakRG,HoadleyKA, PurdomE,WangV,QiY,WilkersonMD, et al.
Integrated genomic analysis identifies clinically relevant subtypes of
Dougherty et al.
Cancer Res; 72(18) September 15, 2012 Cancer Research4866
American Association for Cancer Research Copyright © 2012 on September 17, 2012cancerres.aacrjournals.orgDownloaded from
Published OnlineFirst August 3, 2012; DOI:10.1158/0008-5472.CAN-11-2632

glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR,and NF1. Cancer Cell 2010;17:98–110.
3. Phillips HS, Kharbanda S, Chen R, Forrest WF, Soriano RH, Wu TD,et al. Molecular subclasses of high-grade glioma predict prognosis,delineate a pattern of disease progression, and resemble stages inneurogenesis. Cancer Cell 2006;9:157–73.
4. Brennan C, Momota H, Hambardzumyan D, Ozawa T, Tandon A,Pedraza A, et al. Glioblastoma subclasses can be defined by activityamong signal transduction pathways and associated genomic altera-tions. PLoS One 2009;4:e7752.
5. Nishiyama A, Chang A, Trapp BD. NG2þ glial cells: a novel glialcell population in the adult brain. J Neuropathol Exp Neurol 1999;58:1113–24.
6. Dawson MR, Levine JM, Reynolds R. NG2-expressing cells in thecentral nervous system: are they oligodendroglial progenitors?J Neurosci Res 2000;61:471–9.
7. Jackson EL, Garcia-Verdugo JM, Gil-Perotin S, Roy M, Quinones-Hinojosa A, VandenBerg S, et al. PDGFR alpha-positive B cellsare neural stem cells in the adult SVZ that form glioma-like growthsin response to increased PDGF signaling. Neuron 2006;51:187–99.
8. Nishiyama A, Watanabe M, Yang Z, Bu J. Identity, distribution, anddevelopment of polydendrocytes: NG2-expressing glial cells. J Neu-rocytol 2002;31:437–55.
9. Shih AH, Dai C, Hu X, RosenblumMK, Koutcher JA, Holland EC. Dose-dependent effects of platelet-derived growth factor-B on glial tumor-igenesis. Cancer Res 2004;64:4783–9.
10. UhrbomL,Dai C,Celestino JC, RosenblumMK, Fuller GN,Holland EC.Ink4a-Arf loss cooperates with KRas activation in astrocytes andneural progenitors to generate glioblastomas of various morphologiesdepending on activated Akt. Cancer Res 2002;62:5551–8.
11. Tallquist M, Kazlauskas A. PDGF signaling in cells and mice. CytokineGrowth Factor Rev 2004;15:205–13.
12. Fomchenko EI, Dougherty JD, Helmy KY, Katz AM, Pietras A, BrennanC, et al. Recruited cells can become transformed and overtake PDGF-induced murine gliomas in vivo during tumor progression. PLoS One2011;6:e20605.
13. AssanahM, Lochhead R, OgdenA, Bruce J, Goldman J, Canoll P. Glialprogenitors in adult white matter are driven to form malignant gliomasby platelet-derived growth factor-expressing retroviruses. J Neurosci2006;26:6781–90.
14. Dai C, Celestino JC, Okada Y, Louis DN, Fuller GN, Holland EC. PDGFautocrine stimulation dedifferentiates cultured astrocytes and inducesoligodendrogliomas and oligoastrocytomas from neural progenitorsand astrocytes in vivo. Genes Dev 2001;15:1913–25.
15. Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, TaphoornMJ, et al. Radiotherapy plus concomitant and adjuvant temozolomidefor glioblastoma. N Engl J Med 2005;352:987–96.
16. Pardal R, Clarke MF, Morrison SJ. Applying the principles of stem-cellbiology to cancer. Nat Rev Cancer 2003;3:895–902.
17. Doyle JP, Dougherty JD, Heiman M, Schmidt EF, Stevens TR, Ma G,et al. Application of a translational profiling approach for the compar-ative analysis of CNS cell types. Cell 2008;135:749–62.
18. HeimanM, Schaefer A, GongS, Peterson JD, DayM, RamseyKE, et al.A translational profiling approach for the molecular characterization ofCNS cell types. Cell 2008;135:738–48.
19. Bleau AM, Howard BM, Taylor LA, Gursel D, Greenfield JP, Lim TungHY, et al. New strategy for the analysis of phenotypic marker antigensin brain tumor-derived neurospheres in mice and humans. NeurosurgFocus 2008;24:E28.
20. Dougherty JD, Schmidt EF, Nakajima M, Heintz N. Analyticalapproaches to RNA profiling data for the identification of genesenriched in specific cells. Nucleic Acids Res 2010;38:4218–30.
21. Dai M,Wang P, Boyd AD, Kostov G, Athey B, Jones EG, et al. Evolvinggene/transcript definitions significantly alter the interpretation of Gen-eChip data. Nucleic Acids Res 2005;33:e175.
22. Available from: http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc¼.23. Maere S, Heymans K, KuiperM. BiNGO: a Cytoscape plugin to assess
overrepresentation of gene ontology categories in biological networks.Bioinformatics 2005;21:3448–9.
24. Charles N, Ozawa T, Squatrito M, Bleau AM, Brennan CW, Hambard-zumyan D, et al. Perivascular nitric oxide activates notch signaling andpromotes stem-like character inPDGF-inducedgliomacells. Cell StemCell 2010;6:141–52.
25. Ligon KL, Alberta JA, Kho AT, Weiss J, Kwaan MR, Nutt CL, et al. Theoligodendroglial lineage marker OLIG2 is universally expressed indiffuse gliomas. J Neuropathol Exp Neurol 2004;63:499–509.
26. Cahoy JD, Emery B, Kaushal A, Foo LC, Zamanian JL, ChristophersonKS, et al. A transcriptome database for astrocytes, neurons, andoligodendrocytes: a new resource for understanding brain develop-ment and function. J Neurosci 2008;28:264–78.
27. Nakano I, Paucar AA, Bajpai R, Dougherty JD, Zewail A, Kelly TK,et al. Maternal embryonic leucine zipper kinase (MELK) regulatesmultipotent neural progenitor proliferation. J Cell Biol 2005;170:413–27.
28. Dougherty JD, Garcia AD, Nakano I, Livingstone M, Norris B, Pola-kiewicz R, et al. PBK/TOPK, a proliferating neural progenitor-specificmitogen-activated protein kinase kinase. J Neurosci 2005;25:10773–85.
29. Dugas JC, Ibrahim A, Barres BA. A crucial role for p57(Kip2) in theintracellular timer that controls oligodendrocyte differentiation. J Neu-rosci 2007;27:6185–96.
30. Goldberg JL, Vargas ME, Wang JT, Mandemakers W, Oster SF,Sretavan DW, et al. An oligodendrocyte lineage-specific semaphorin,Sema5A, inhibits axon growth by retinal ganglion cells. J Neurosci2004;24:4989–99.
31. Chen Y, Wu H,Wang S, Koito H, Li J, Ye F, et al. The oligodendrocyte-specific G protein-coupled receptor GPR17 is a cell-intrinsic timer ofmyelination. Nat Neurosci 2009;12:1398–406.
32. Saari JC, Huang J, Possin DE, Fariss RN, Leonard J, Garwin GG, et al.Cellular retinaldehyde-binding protein is expressed by oligodendro-cytes in optic nerve and brain. Glia 1997;21:259–68.
33. Iijima T, Miura E, Watanabe M, Yuzaki M. Distinct expression of C1q-like family mRNAs in mouse brain and biochemical characterization oftheir encoded proteins. Eur J Neurosci 2010;31:1606–15.
34. Beroukhim R, Getz G, Nghiemphu L, Barretina J, Hsueh T, Linhart D,et al. Assessing the significance of chromosomal aberrations in can-cer: methodology and application to glioma. Proc Natl Acad Sci U S A2007;104:20007–12.
35. Taylor BS, Barretina J, Socci ND, Decarolis P, LadanyiM,MeyersonM,et al. Functional copy-number alterations in cancer. PLoSOne 2008;3:e3179.
36. Piccirillo SG, Reynolds BA, Zanetti N, Lamorte G, Binda E, Broggi G,et al. Bone morphogenetic proteins inhibit the tumorigenic potential ofhuman brain tumour-initiating cells. Nature 2006;444:761–5.
37. Das A, Banik NL, Ray SK. Retinoids induce differentiation and down-regulate telomerase activity and N-Myc to increase sensitivity toflavonoids for apoptosis in humanmalignant neuroblastomaSH-SY5Ycells. Int J Oncol 2009;34:757–65.
38. CianaP, FumagalliM, TrincavelliML, VerderioC,RosaP, LeccaD, et al.Theorphan receptorGPR17 identifiedasanewdual uracil nucleotides/cysteinyl-leukotrienes receptor. EMBO J 2006;25:4615–27.
39. Lecca D, Trincavelli ML, Gelosa P, Sironi L, Ciana P, Fumagalli M,et al. The recently identified P2Y-like receptor GPR17 is a sensor ofbrain damage and a new target for brain repair. PLoS One 2008;3:e3579.
40. Huse JT, Holland EC. Genetically engineered mouse models of braincancer and the promise of preclinical testing. Brain Pathol 2009;19:132–43.
41. Fuller GN, Scheithauer BW. The 2007 revised World Health Organi-zation (WHO) classification of tumours of the central nervous system:newly codified entities. Brain Pathol 2007;17:304–7.
42. Vordermark D, Ruprecht K, Rieckmann P, Roggendorf W, Vince GH,Warmuth-MetzM, et al. Glioblastomamultiformewith oligodendroglialcomponent (GBMO): favorable outcome after post-operative radio-therapy and chemotherapy with nimustine (ACNU) and teniposide(VM26). BMC Cancer 2006;6:247.
43. Persson AI, Petritsch C, Swartling FJ, Itsara M, Sim FJ, Auvergne R,et al. Non-stem cell origin for oligodendroglioma. Cancer Cell 2010;18:669–82.
Candidate Pathways for Promoting Differentiation in Glioma
www.aacrjournals.org Cancer Res; 72(18) September 15, 2012 4867
American Association for Cancer Research Copyright © 2012 on September 17, 2012cancerres.aacrjournals.orgDownloaded from
Published OnlineFirst August 3, 2012; DOI:10.1158/0008-5472.CAN-11-2632

44. Lei L, SonabendAM,Guarnieri P, Soderquist C, LudwigT,RosenfeldS,et al. Glioblastoma models reveal the connection between adult glialprogenitors and the proneural phenotype. PLoS One 2011;6:e20041.
45. Lu QR, Park JK, Noll E, Chan JA, Alberta J, Yuk D, et al. Oligoden-drocyte lineage genes (OLIG) as molecular markers for human glialbrain tumors. Proc Natl Acad Sci U S A 2001;98:10851–6.
46. Rhee W, Ray S, Yokoo H, Hoane ME, Lee CC, Mikheev AM, et al.Quantitative analysis of mitotic Olig2 cells in adult human brain andgliomas: implications for glioma histogenesis and biology. Glia2009;57:510–23.
47. Zheng H, Ying H, Yan H, Kimmelman AC, Hiller DJ, Chen AJ, et al. p53and Pten control neural and glioma stem/progenitor cell renewal anddifferentiation. Nature 2008;455:1129–33.
48. Wang Y, Yang J, Zheng H, Tomasek GJ, Zhang P, McKeever PE, et al.Expression of mutant p53 proteins implicates a lineage relationshipbetween neural stem cells andmalignant astrocytic glioma in amurinemodel. Cancer Cell 2009;15:514–26.
49. Ellis JA,Castelli M, Bruce JN,Canoll P,OgdenAT.Retroviral delivery ofplatelet-derived growth factor to spinal cord progenitor cells drives theformation of intramedullary gliomas. Neurosurgery 2012;70:198–204.
50. Ivkovic S, Canoll P, Goldman JE. Constitutive EGFR signaling inoligodendrocyte progenitors leads to diffuse hyperplasia in postnatalwhite matter. J Neurosci 2008;28:914–22.
51. Mischel PS, Cloughesy TF, Nelson SF. DNA-microarray analysis ofbrain cancer: molecular classification for therapy. Nat Rev Neurosci2004;5:782–92.
52. Bilanges B, Stokoe D. Mechanisms of translational deregulation inhuman tumors and therapeutic intervention strategies. Oncogene2007;26:5973–90.
53. Parsa AT, Holland EC. Cooperative translational control of gene expres-sion by Ras and Akt in cancer. Trends Mol Med 2004;10:607–13.
54. Hesson LB, Dunwell TL, Cooper WN, Catchpoole D, Brini AT, Chiar-amonte R, et al. The novel RASSF6 and RASSF10 candidate tumoursuppressor genes are frequently epigenetically inactivated in child-hood leukaemias. Mol Cancer 2009;8:42.
55. Shibata D, Mori Y, Cai K, Zhang L, Yin J, Elahi A, et al. RAB32hypermethylation and microsatellite instability in gastric and endome-trial adenocarcinomas. Int J Cancer 2006;119:801–6.
56. Kim SK, Jang HR, Kim JH, Noh SM, Song KS, Kim MR, et al. Theepigenetic silencing of LIMS2 in gastric cancer and its inhibitory effecton cell migration. Biochem Biophys Res Commun 2006;349:1032–40.
57. Sarkar S, Roy BC, Hatano N, Aoyagi T, Gohji K, Kiyama R. A novelankyrin repeat-containing gene (Kank) located at 9p24 is a growthsuppressor of renal cell carcinoma. J Biol Chem 2002;277:36585–91.
58. Das A, Banik NL, Ray SK. Retinoids induced astrocytic differentiationwith down regulation of telomerase activity and enhanced sensitivity totaxol for apoptosis in human glioblastoma T98G and U87MG cells.J Neurooncol 2008;87:9–22.
59. Zhang Y, Jia S, Liu Y, Li B, Wang Z, Lu H, et al. A clinical study of all-trans-retinoid-induced differentiation therapy of advanced thyroidcancer. Nucl Med Commun 2007;28:251–5.
60. Zhao H, Ayrault O, Zindy F, Kim JH, Roussel MF. Post-transcrip-tional down-regulation of Atoh1/Math1 by bone morphogenic pro-teins suppresses medulloblastoma development. Genes Dev2008;22:722–7.
61. Lubbe SJ, Pittman AM, Matijssen C, Twiss P, Olver B, Lloyd A, et al.Evaluation of germline BMP4mutation as a cause of colorectal cancer.Hum Mutat 2011;32:E1928–38.
62. Braig S, Mueller DW, Rothhammer T, Bosserhoff AK. MicroRNA miR-196a is a central regulator of HOX-B7 and BMP4 expression inmalignant melanoma. Cell Mol Life Sci 2010;67:3535–48.
63. Miyamoto K, Sakurai H, Sugiura T. Proteomic identification of a PSF/p54nrb heterodimer as RNF43 oncoprotein-interacting proteins. Pro-teomics 2008;8:2907–10.
64. Sendoel A, Kohler I, Fellmann C, Lowe SW, Hengartner MO. HIF-1antagonizes p53-mediated apoptosis through a secreted neuronaltyrosinase. Nature 2010;465:577–83.
65. Vicent S, Luis-Ravelo D, Anton I, Garcia-Tunon I, Borras-Cuesta F,Dotor J, et al. A novel lung cancer signature mediates meta-static bone colonization by a dual mechanism. Cancer Res 2008;68:2275–85.
66. Aalto Y, El-Rifa W, Vilpo L, Ollila J, Nagy B, Vihinen M, et al. Distinctgene expression profiling in chronic lymphocytic leukemia with 11q23deletion. Leukemia 2001;15:1721–8.
67. Fomchenko EI, Holland EC. Stem cells and brain cancer. Exp Cell Res2005;306:323–9.
68. Stiles CD, Rowitch DH. Glioma stem cells: a midterm exam. Neuron2008;58:832–46.
69. Lindberg N, Kastemar M, Olofsson T, Smits A, Uhrbom L. Oligoden-drocyte progenitor cells can act as cell of origin for experimentalglioma. Oncogene 2009;28:2266–75.
Dougherty et al.
Cancer Res; 72(18) September 15, 2012 Cancer Research4868
American Association for Cancer Research Copyright © 2012 on September 17, 2012cancerres.aacrjournals.orgDownloaded from
Published OnlineFirst August 3, 2012; DOI:10.1158/0008-5472.CAN-11-2632
Copyright © 2022 FDOKUMEN




















