
BEYOND SIZE: MATRIX PROJECTION MODELS FOR POPULATIONS WHERE SIZE IS AN INCOMPLETE DESCRIPTOR
Upload
independentCategory
view
2download
0
Age-Size Structure in Populations with Quiescence
M. GYLLENBERG*
Helsinki University of Technology, Institute of Mechanics, SF-02150 Espoo, Finland
AND
G. F. WEBB
Vanderbilt University, Department of Mathematics, Nashville, Tennessee 37235
Recerved 7 September 1986; revised 1 June 1987
ABSTRACT
An analysis is given of a mathematical mode1 for the continuous age-size distribution of
a population that has both normal (growing and reproducing) and quiescent individuals.
Individuals can transit back and forth from one state to the other. The theory of positive
operator semigroups is used to show that under general assumptions about individual
behavior the age-size distribution of the population converges to a stable distribution. The
features of the model are illustrated by several examples. The examples are designed to
minimize technicalities, yet still reveal the interesting stability phenomena that occur when
structure is combined with quiescence. For these examples the extinction or asynchronous
exponential growth of the population is analyzed in terms of the parameters effecting
transition between the normal and quiescent states. The continuous mode1 is compared
with an analogous discrete model.
1. INTRODUCTION
In many populations not all individuals are actively growing and pro- liferating, but some are in a quiescent state. Under certain circumstances a normally growing individual can become quiescent, and later it may return to the normal state again. Often an individual can undergo several transi- tions back and forth from one state to the other.
As an example of a species with quiescence we can take a plant that reproduces by producing seeds. Some of the seeds germinate immediately and develop into active growing plants. Other seeds remain in the soil for years until they finally give rise to growing plants. In many cases the time of
*This work was performed while M. Gyllenberg was visiting Vanderbilt University
during the academic year 1985-1986.
MATHEMATICAL BIOSCIENCES 86:67-95 (1987)
OElsevier Science Publishing Co., Inc., 1987
52 Vanderbilt Ave., New York, NY 10017
67
00255564/87/$03.50

68 M. GYLLENBERG AND G. F. WEBB
quiescence is not fixed, but seems to obey a probability distribution. A second example is given by cell populations. In eukaryotic tissues most cells are not actively growing, but are in a resting state. The growth arrest usually happens in the Gl phase of the cell cycle, but can occur later [6, p. 221. Investigation of quiescence in mammalian cell populations is of considerable importance, since it is hypothesized that arrest states in the Gl phase of the cell cycle are closely related to an integrated control of cell proliferation and differentiation, and that cancer may result from defects that uncouple this integrated system [20].
Since the probability that a normal individual becomes quiescent and vice versa usually depends on the age and size (as well as other quantities) of individuals, modeling the growth of populations with quiescence naturally falls into the realm of structured population dynamics. One of the main goals in studying structured population models is to draw conclusions about the behavior of the population as a whole from known or assumed behavior of individuals.
Many authors have treated age and size structured populations; see e.g. [2], [9], [lo], [16], [17], and [18]. Several authors have treated structured population models with quiescence. In [13], [14], and [15] Rotenberg used maturity as a structure variable in models of proliferating cell populations with quiescence. In [7, Chapter XIII] Greenberg, Protopopescu, and Van der Mee continued this investigation. In [21] Wu et al. treated discrete models with quiescence using age and size as discrete structure variables and applied their results to forest growth with cohort size corresponding to diameter. In [3] Caswell treated discrete size structured populations with quiescence and applied the results to clonally reproducing organisms.
The model we consider identifies age and size as continuous structure variables. Individuals in the normal state advance in both age and size, whereas quiescent individuals advance only in age. Our model is also continuous in time and therefore consists of a system of partial differential equations with initial and boundary conditions. Our analysis focuses upon the extinction or exponential growth of the population. This question is answered by the value of a single constant called the intrinsic growth constant
or the Malthusian parameter. The intrinsic growth constant depends upon the various parameters within the equations, but not upon the initial states of the population.
The model we analyze possesses phenomena that are not present in models without structure variables. We illustrate this point with several examples that emphasize the importance of structure. One of these demon- strates that a population can escape extinction by transferring normal individuals into the quiescent state within a certain range of the structure variables. Critical values determine when this can be achieved. Another example shows that control of the mortality process in certain ranges of the

AGE-SIZE STRUCTURE WITH QUIESCENCE 69
structure variable is heavily influenced by the presence of quiescent popula- tion. This type of control is important in phase specific cancer therapy in which destruction of nonmalignant cells must be minimized.
This paper is organized as follows. In Section 2 we formulate the mathematical model and describe its mathematical setting. In Section 3 we discuss without proofs the main ideas of analyzing the equations. In Section 4 we present a qualitative and quantitative discussion of some examples. In Section 5 we discuss a discretized age-sire model as an analogue to the continuous one. In the Appendix we provide proofs of the results.
2. FORMULATION OF THE PROBLEM AND DESCRIPTION OF THE STATE SPACE
We assume that an individual is fully characterized by its age, size, and the state (either normal or quiescent) it is in. This means that all quantities that determine the development of an individual, such as growth and death rates and transition rates from one state to another, depend only on age, size, and state. We assume that only individuals in the normal state can grow (or possibly shrink) and reproduce. All individuals are born into the normal state. An individual in the quiescent state ages, but its size does not change and it cannot produce offspring as long as it stays in the quiescent state.
We let t denote time, a age, x size, and we denote the age-size distributions at time t of individuals in the normal and the quiescent state by n(t, a, x) and q(t, a, x), respectively. Thus for instance /.$/2n(f, a, x) dudx
is the number of individuals in the normal state who at time t have age between a, and u2 and size between x1 and x2. We can now write down the balance equations for the two states:
=-~(u,x)n(r,u,x)-~,(~,x)~(~,~,x)
+I;(a,x>q(t,a,x), (2.1)
n(t,O,x> = $$B(u,v,x)n(r,u,~)dudy, (2.2) cl
~~(r~u~x)+~~(r~u~x)~~u(u~x)~(r~u~x)~ri(u~x)~(r~u~x)
+r,(a,x)n(t,a,x), (2.3)
4(1,0,x) =o. (2.4)
Here Q is the domain of permitted (a, x) values to be specified below. ~1 and u are the death rates in the normal and quiescent state, respectively. r0 is the transition rate from normal state to quiescent state, and ri is the

70 M. GYLLENBERG AND G. F. WEBB
transition rate in the other direction (the subscripts i and o stand for “into” and “out,” respectively). g is the growth rate of individuals in the normal state and has the following interpretation. Let X( a, a) 5‘ denote the solution of the initial value problem
(we will state later precise assumptions on g that guarantee the existence and uniqueness of solutions). Then X( a, a) 5 is the size of an individual of age a that had size 6 when it was of age a. The curve x = X( a, a),$ in the (a, x) plane is called the characteristic curve passing through (LY, 5) in the normal state.
Equation (2.2) is the birth law. It relates the flux (per unit size) n(t,O, x) of newborn individuals of size x through the boundary a = 0 of Q to the age-size distribution n( t, a, x). /3 is called the fecundity function. Roughly speaking, the larger the value of /3(u, y, x) is, the more probable it is that an individual of age a and size y produce offspring of size x. We assume that there exists a maximal age a, of reproduction, that is, the fecundity function /I is zero for a > a,. Since individuals older than ui do not contribute to the renewal of the population, we simply neglect them and consider only the population of normal and quiescent individuals of age less than or equal to
al.
The birth law (2.2) is sufficiently general to serve as a model for the reproduction pattern of higher animals and plants as well as for asymmetri- cal divisions of cells. It does not, however, include fission into two equal
parts. We now specify the individual state space, that is, the domain of per-
mitted age and size values in both states. We assume that a newborn individual can have any size between two fixed finite positive values x0 and
x1, and no other size. This is achieved by requiring /3( a, y, x) to be nonnegative for x E (x,,, xi) and zero for x $Z (x0, xi). The individual state variable (a, x) of an individual born with size 6 will then travel along the characteristic curve x = X( a ,O)[ as long as the individual stays in the normal state. If there were no quiescent state, that is, if r, = r, = 0 in (2.1), then the state variables (a, x) would be confined to the domain 8, = {(u, x) E R2 (0 < a < a,, X(u,O)x, <x Q X(u,O)x,} (see Figure 1).
It is immediately seen that Q2, is too small to serve as individual state space for the normal state, when the quiescent state is taken into account. Let us for the sake of simplicity assume that the transition rates r0 and ri are
strictly positive everywhere they are defined. This means that an individual can transit from the normal to the quiescent state and vice versa at every point (u, x), and it follows that the individual state spaces must be identical

AGE-SIZE STRUCTURE WITH QUIESCENCE 71
FIG. 1. The region a,, and the characteristic curve x = X(a,O)[.
for the normal and the quiescent state. The individual state space is therefore of the form D x a for some s2 containing 8,. Since the individual state variables travel along characteristic curves, it is clear that the individual state space D must contain these curves.
We have already described the characteristic curves of the normal state. Since individuals in the quiescent state do not grow, the characteristic curves of the quiescent state are simply straight lines parallel to the u-axis. The natural choice for individual state space is therefore the set D x 3, where D is the smallest subset of R2 with the properties
(2) if (cy, x) E 52, then (a,~) ED for all a E ](Y, a,]; (3) if (cy, x) E 8, then X(u,a)x E Q for all a E [(Y, aI].
The set fJ corresponding to the P, of Figure 1 is sketched in Figure 2. G can be characterized as follows: Q = {(a, x) E R2 10 < a < a,, fo(u) < x <
fi (a)}, where f. is the unique solution of the initial value problem
fi(0) =xi (i-0,1) (2.6)

72
X
and
M. GYLLENBERG AND G. F. WEBB
.-x= f,(a) 1
FIG. 2. The set Q. The individual state space is B x 0.
F,(a,x) = i
g(a,x) if g(a,x) GO,
0 if g(a,x) SO,
F,(a,x) = i
0 if g(U,x) GO,
g(a,x) if g(a,x)>O.
Of co&e, we have to impose conditions on g which ensure that 52 lies entirely in the positive quadrant.
As population state space we choose L’(il x a). That is, given initial distributions (p E L’(B), 4 E L’(Q), we want to find continuous functions n and q from R+ to L’(0) which satisfy (2.1)-(2.4) in an appropriate weak

AGE-SIZE STRUCTURE WITH QUIESCENCE
sense (defined precisely in Section 3) such that
73
n(O,a,x) =+(a,x), (a,x) EQ, (2.7)
4(O,a,x) =#(a,x), (a,x) EQ. (2.8)
The boundary conditions (2.2) and (2.4) describe the influx of individuals through the boundary a = 0. A glance at Figure 2 shows that characteristics can enter Q at points of the boundary as2 where a is greater than zero. This happens at those parts of the boundary x =&(a), x = fi(a) which do not coincide with characteristics. Since there is no influx of individuals through
these parts of the boundary, we add to the system (2.1)-(2.4), (2.7)-(2.8) the boundary conditions
g(a,h(a))+,u,h(a)) =o, r,‘(a) =o, O~ada,, i=O,l (2.9)
q(t,a,h(a)) =o, f,‘(a)+O, O<a<u,, i=O,l. (2.10)
Concerning the developmental rates g, /3, CL, v, r,, r, , which are all assumed to be given, we make the following assumptions:
(H,) g is continuous on [0, al]XIO, co) and has a continuous partial derivative with respect to x. The sets {a~[O,a,]~g(a,X(a,O)x~)<O},
i = O,l, have only finitely many connected components. g(a,O) > 0, a E
10, a, I. (HP) /3 is bounded and continuous on Q X(x,, x1) except possibly for
finitely many jump discontinuities in each place. There exists an a, E [0, a,)
such that &a, y, x) = 0 for a E [0, a,], B(a, y, x) > 0 for a E (a,,, al].
(HP,.) P, v E L”(Q), CL, v > 0. (H,) rO, r; E L”(O), r,, ri > 0.
Remark. If the condition r, > 0, r, > 0 is violated, it is very difficult to describe the individual state space in general. However, in certain concrete situations it is easily done. Since we will deal with examples where r, and ri are not strictly positive in Section 4, we will here briefly indicate how to choose the individual state space in one such case.
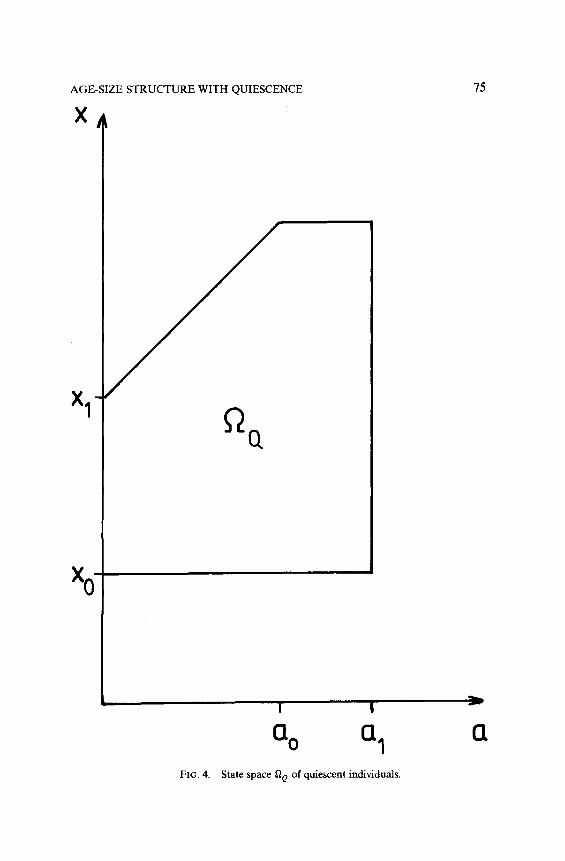
Let g(a, x) ~1, and assume that r,(a, x) > 0 for 0 6 a Q a,, r,(a, x) = 0
for a,Gada,, ri(a,x)=O for O<aga,, r,(a,x)>O for a,<a~a,. Then the state spaces Q, and Qe of normal and quiescent individuals should be chosen as in Fig. 3 and 4, respectively. The reason why there can be no normal individuals with state (a, x) in the triangle 0 G a Q a;,
x,<x<a+x,, is that the quiescent individuals in this region cannot become normal. Similarly, because normal cells cannot become quiescent for a 2 a,, there are no quiescent cells in the triangle a, Q a Q a,, x1 + a,, G x
< x1 + a. If there are initially individuals with states (a, x) in these empty

74
x
x1
xO
M. GYLLENBERG AND G. F. WEBB
I 1 w a. a
I al FIG. 3. State space a, of normal individuals.
AGE-SIZE STRUCTURE WITH QUIESCENCE 75
X
FIG. 4. State space tia of quiescent individuals

76 M. GYLLENBERG AND G. F. WEBB
regions, they will disappear from there either by dying or by reentering either tit, or 8, after finite time. In fact, for t > a, the individual states are concentrated on 8, X ii,, which is therefore a proper choice of individual state space. The population state space is L’(O, x ilQ). The reason for not allowing “holes” in the individual state space is that in that case the semigroup associated with the solutions would not be irreducible (see Appendix).
3. THE SOLUTION SEMIGROUP AND ITS PROPERTIES
Denote the state space L’(Q X iI) by L’. L’ is a Banach lattice, and its positive cone Lf+ consists of all {(p, 4) such that C$ and 4 are nonnegative almost everywhere. The solutions of the model (2.1)-(2.4), (2.7)-(2.10) form a strongly continuous semigroup of positive bounded linear operators V(t), t 2 0, in L’ according to the formula U(t){ (p, $} = (n(t, ., a), q(t, ., .)}. The operators u(t), f z 0, have the properties cI(0) = I, u(t + s) = Lr(r)Ll(s), u(t){+,+} is continuous in t for fixed ($,$} EL’, and (1(t){ $,$} E L!+ for {$,,lr/} E L:. If {@, $} is sufficiently smooth, then U(t){ (p, $} will satisfy the equations (2.1)-(2.4). For arbitrary {(p, Ir/} E L’ the function U(t){ C#I, $} may be interpreted as a solution in a weak sense.
Various properties of the solutions of the model can be proved using the general theory of positive operator semigroups in Banach lattices. The principal result we prove here is that the solutions have asynchronous exponential growth. This means that there exist a real constant X, and a projection Pa in L’ such that for all {$,$} EL’, limt,,e-Xo’Lr(t){+,~} = P, { $, 4 }. A, is called the Malthusian parameter, and P,, { C/I, 4) is called the exponential steady state. We will prove that PO is a rank one projection in L’, that is, it has one-dimensional range. This means that there exists a unique normalized exponential steady state { I#+,, $a } in L!+ such that
pa{+,+> =c(+,$~){&&,), h w ere c(+, #) is a constant depending only on
{k$}. If X,+0, th en the population extinguishes. If X, > 0, then the population grows exponentially in time, but stabilizes in age and size in the
sense that for 0’ c D
and
jjG,n( t, -, .) dxda
jja2q(t;;)dxda
The asynchronous exponential growth of the semigroup u(t), t > 0, results from two important properties it possesses. One is its ultimate compactness,

AGE-SIZE STRUCTURE WITH QUIESCENCE 77
and the other is its ultimate strict positivity. These two properties can be established by viewing U(t), t > 0, as a perturbation of the positive semi-
groups T(t), t > 0, and S(t), t > 0, in L1( 0) associated with the uncoupled problem, that is, T(t)+ =: n(t, -, .) and S(t)+ =: q(t, 1, .), where n, q is the solution of (2.1)-(2.4), (2.7)-(2.10) with the q term omitted in (2.1) and the n term omitted in (2.3). For notational convenience we define B( t, x) =
Ils2P(a,y,x)(T(t)~)(a,y)dyda. Define II,(u, b, x) = exp[- /,“(p + r, + g,)(c, X(c, a)x) dc] and
If, (a, b, x) = exp[ - /,“( Y + ri)( c, x) dc]. Then
I
$(a-t,X(u-t,u)x)rII,(u-t,u,X(u-t,u)x),
t<u,(u-t,X(u-t,u)x) Es2,
(~(t)~)(~,x) = B(t-u,X(O,u)x)rI,(O,u,X(O,u)x),
t>a,(a,x) EQ.3,
0 otherwise
(3.1)
(3.2)
The formulas (3.1) and (3.2) can be verified directly for smooth $ and $. The 0 values in (3.1) and (3.2) follow from (2.9) and (2.10), respectively. To verify (3.1) use the following formula (which may be found in [8, p. 211):
&X( a u,a)x= -g(ol,x)@(w)x
= -g(a,x)exp[~‘~~(P,X(8,~)x)dB]. (3.3)
Set
(3.4)
One now sees that U(t), t > 0, has the representation
Lqt){+,rl/} =w(r){~.~}+ldw(f-s)KU(s){~,~} ds. (3.5)
The semigroup S(t), t 2 0, is zero after time a,, and the semigroup T(t), t > 0, is compact after time a,. Further, T(t), t > 0, is strictly positive for t sufficiently large. The semigroup U(t), t > 0, inherits these two properties of

78 M. GYLLENBERG AND G. F. WEBB
T(t), t > 0. In the Appendix we will establish these facts and use the general theory of positive operator semigroups in Banach lattices to prove the asynchronous exponential growth of U(t), t 2 0.
4. TWO SIMPLE EXAMPLES
In this section we illustrate some of the features of the model by two examples. To keep the examples as simple as possible we assume that the vital rates ~1, v, and /I as well as the transition rates r, and r, depend on age
only, but we do not impose any such further restrictions on the growth rate g. The model then reduces to a system involving only age but not size. In fact, introducing the age distributions
and integrating the balance equations (2.1) and (2.3) over all sizes [that is, from f0 (a) to fi (a) for any given age a] and taking the boundary conditions (2.9) and (2.10) into account, one obtains
aN(t, a> at
+ aN(t,a) au =-~(~)~(~,~)-r,(~)N(~,u)+r;(~)Q<~,~)
(4.1)
aQ(f,u) + aQ(t,a) at au =-v(a)Q<t,u)-r,(u)Q(~,u)+r,(u)~(~,u)
(4.2)
for t > 0 and 0 < a G ui. To say that /_I depends on age only means in particular that the birth sizes are uniformly distributed over the permitted birth sizes [x,, xi]. By normalizing b we can assume that it has the form /3( a)/( xi - x0). Integration of the birth law (2.2) with respect to x from x,, to xi then yields
The boundary condition (2.4) becomes
Q(t,O) = 0, t>o. (4.4)
In both our examples we assume that there is no mortality in the quiescent state, that is, v = 0. We normalize the maximum age a, to equal 1. We choose step functions with a single step at a = f for the rates p, 8, r,, and I;.

AGE-SIZE STRUCTURE WITH QUIESCENCE 19
In our first example we assume that the mortality is highest in younger age groups of normal individuals while fecundity is concentrated among older individuals. The young individuals have a tendency to become quies- cent, whereas there is a high probability that an older quiescent individual becomes normal and reproducing. Specifically, we choose the rates as follows:
r,( u) = t-0 i
Oga<+,
1
0 Ogagt,
0 +<a<l, ri( u) =
‘I f<a<l.
The constants ~1 and /I are assumed positive, and r, and ri nonnegative. This choice of transition rates violates hypothesis (H,), so the proof of asynchronous exponential growth given in the Appendix cannot directly be applied, but it can easily be adapted to the present situation by a proper choice of the individual state space (see the remark at the end of Section 2).
We investigate the ultimate behavior (exponential growth, extinction) of the population from the following point of view. We consider p and j3 as fixed and r, and r, as parameters, and describe how the dominant real eigenvalue of the infinitesimal generator corresponding to the system (4.1)-(4.4) depends on r, and r,.
It turns out that when the rates have the forms chosen above it is possible to explicitly write down the characteristic equation for the system (4.1)-(4.4), the unique real root of which is the dominant eigenvalue of the generator. By formally substituting N( t, a) = &#J( a), Q( t, a) = e”JI( a) into (4.1) and (4.2) one obtains the following equations:
+‘(a) =-_(X+~+r~)+(a),
$‘(a) = -x$(a) + r,+(a)
whenO<a<t, and
+‘(a> = -G(a) + r;+(a),
+‘(a> = -(A + r,>+(a)
when ) < a < 1. The boundary conditions (4.3) and (4.4) become
(4.5)
(4.6)
(4.7)
(4.8)
$40) = BQ(4 da (4.9)
and
q(0) = 0. (4.10)

80 M. GYLLENBERG AND G. F. WEBB
To obtain the characteristic equation we proceed as follows. Equation (4.5) can be solved on 0 G a G 5. The expression for +(a), which contains the still undetermined constant G(O), is substituted into (4.6), which taking (4.10) into account can then be solved explicitly for $(a) on 0 Q a < f. At this point we know in particular the values +(f),+(i). We can therefore continue to solve (4.8) and then (4.7) on f < a ~1. The expression obtained for $(a) on f =G a ~1, still containing +(O) as a factor, is substituted into (4.9). (p(O) cancels and we arrive at the characteristic equation
p( e++‘+(h) + --+(l- e-f(P+‘e))gx( r;)] =l (4.11)
where
p(X) =Jlle-““da 2
gh( r,) = Jlle-““[l- e-+f)] da. 2
(4.12)
(4.13)
For fixed values of r0 and r, the characteristic equation (4.11) has a unique real root X -the Malthusian parameter of the population.
To get a better understanding of how the Malthusian parameter depends on the transition rates we fix X and interpret (4.11) as a relation between r,
and r,. The corresponding curve is then plotted in the r,ro plane. This is repeated for several values of X. The result is sketched in Figure 5.
From Figure 5 we first observe that if r, = 0, then the Malthusian parameter has a fixed value A, for all values of ri. This is clear, since when r, = 0, (4.11) reduces to
pe-fPp(XJ =l, (4.14)
which is independent of r,. When the transition rate from the normal to the quiescent state is zero, our model reduces to the usual age-structured Lotka-McKendrick model with only one (normal) state. The characteristic equation of the Lotka-McKendrick model with our choice of /3(u), ~(a) is precisely (4.14). Thus the unique real solution A, of (4.14) is the Malthusian parameter of the population in the absence of a quiescent state. It is very interesting to observe that the curve corresponding to h = &, has another branch bifurcating from the branch r0 = 0 at the point ( ri* ,O) [ ri* can easily
be found to equal gi0’(y/2p(1 - e-T’>)]. This means that in the presence of a quiescent state the population will ultimately behave as if there were no quiescent state, provided the transition rates are such that (I;, r,) lies on the upper branch of the curve corresponding to h = X,.

AGE-SIZE STRUCTURE WITH QUIESCENCE 81
FIG. 5. Curves on which the dominant eigenvalue remains fixed. X- < X, < h’
If a value greater than X, is assigned to X, then the characteristic equation determines r, uniquely as a monotonically decreasing function of r,. This is the case A = h+ in Figure 5. When A = X- < X,, the correspond- ing curve again has two branches. The lower branch starts from (0,2ln[pp(X)] - p). The bifurcation point can in principle be calculated from (4.11), but the expression is very complicated. All curves have vertical asymptotes. They are found from (4.11) by letting r. tend to infinity. For a given X the vertical asymptote is given by
r;=ghl - i 1 ;. (4.15)
The curves corresponding to h = X+ > A, have horizontal asymptotes. These are obtained by solving
P pe -fb+r,)+ r
P + r. “p(A) =l (4.16)
for ro.

82 M. GYLLENBERG AND G. F. WEBB
The value of the Malthusian parameter for different values of I;, r. is not bounded below. To see this take r, = 0 in (4.11). Then letting r0 tend to infinity forces A to approach - cc. This is intuitively clear, since if r, = 0,
then quiescent individuals cannot become normal and reproduce again, and r. is essentially only an added mortality. When this mortality tends to + cc the Malthusian parameter must tend to - cc. On the other hand, X is bounded above. From Figure 5 we see that when (r, , r,) + (00, co), A
increases. In the limit the characteristic equation takes the form
BP(A) =I, (4.17)
which is the characteristic equation for the Lotka-McKendrick model with no mortality. The solution A of (4.17) is the supremum of the Malthusian parameter as r, and r, range over all nonnegative numbers. The explanation is simple. In order to m aximize X the individuals should escape death by entering immediately at birth the quiescent state. When they reach age a = f
and the danger is over [p(a) = 01, they should immediately return to the normal state and reproduce.
The curve corresponding to A = 0 is of special interest, because it divides the parameter domain {( ri, r,) 1 r, > 0, r. > 0) into a part where the popula-
FIG. 6. h, < 0. Region 1: extinction. Region 2: asynchronous exponential growth.

AGE-SIZE STRUCTURE WITH QUIESCENCE 83
1 2
FIG. 7. h, > 0. Region 1: extinction. Region 2: asynchronous exponential growth
tion goes extinct and a part where the population grows exponentially and the age distribution approaches a stable distribution. We consider two cases, one in which the population would go extinct in the absence of a quiescent state (A, < 0), and another in which the population grows exponentially without the quiescent state (A, > 0). The curves corresponding to A = 0 are plotted in each case in Figure 6 and Figure 7, respectively.
By looking at Figure 6 and Figure 7, we see that the population survives provided a certain relationship between the transition rates holds. This relationship can sometimes be rather delicate, especially in the case A, > 0, g;‘(l/@) < r, -C r,* (Figure 7). It stems from the fact that both the normal and the quiescent state have their advantages and drawbacks. The disad-

84 M. GYLLENBERG AND G. F. WEBB
vantage of the normal state is the mortality of young individuals, and the advantage is the ability to reproduce. In the quiescent state the situation is exactly the opposite: quiescent individuals neither die nor reproduce. The optimal strategy for an individual is therefore to transit back and forth between the normal and the quiescent state in such a way that it exploits the advantages of both states and avoids the drawbacks.
Let us first discuss the situation described in Figure 6, where the popula- tion goes to extinction in the absence of quiescence. We assume that j3 > 2; otherwise X = 0 is not a solution of (4.11) for any r,, ri. If the rate r, is small, then even if practically every individual escapes death by becoming quies- cent (r, 4 00) the population cannot be saved: too few individuals come back to the normal state to reproduce. On the other hand, if the rate r, is too small, then most individuals stay in the normal state and a large portion die before they reach reproductive age. A large value of ri does not help: the population is doomed to extinction. Only if both r. and ri are sufficiently large can the population avoid extinction.
The situation described in Figure 7 is quite different from the previous one. Since the population grows exponentially in the absence of quiescence (r, = 0), the mortality p is small compared with the fecundity /3. Increasing ro, which corresponds to escaping death but becoming unable to reproduce, is therefore a bad strategy for small values of r,: the drawback is greater than the advantage. If we fix r, between g;‘(l/p) and r,* and increase r,, a perhaps surprising phenomenon occurs. The parameter point (r,, ro) moves from the domain of exponential growth to the domain of extinction as it traverses the lower branch of the curve X = 0. As r, is increased further, the point traverses the upper branch of the curve and comes back to the domain of exponential growth. This shows how subtle the interplay between the relative advantages and disadvantages of the two states can be. There is still one fundamental difference between this case and the previous one. If ri is sufficiently large ( > r,* ), then the population always grows exponentially, no matter what the value of r, is.
In our second example we assume that mortality is highest in older age groups. The step functions j3, r,, and r, are as in the first example, but ~(a) is the step function
A calculation similar to the one for the first example yields the characteristic equation
/3{ ,~cc-~)p(X)i(l-e-t")gh(rj)} =l, (4.18)

AGE-SIZE STRUCTURE WITH QUIESCENCE
where
85
p(X) = jlle-(“+“)y da, 2
(4.19)
(4.20)
The unique real root X of (4.18) is the Malthusian parameter of the population. Again we analyze the dependence of the Malthusian parameter
upon the rates r, and r,. The Malthusian parameter A, for the population without quiescent state (that is, when r, = 0) is an upper bound for all possible roots of the characteristic equation. As ri --) co the left-hand side of
(4.18) approaches /3&p(X). Thus, the Malthusian parameter h + X, if we
FIG. 8. Curves on which the dominant eigenvalue remains fixed. h, < X, < A,

M. GYLLENBERG AND G. F. WEBB
3
FIG. 9. Region 1: extinction in both examples. Region 2: asynchronous exponen-
tial growth in first example, extinction in second example. Region 3: asynchronous expo-
nential growth in both examples. Region 4: extinction in first example, asynchronous
exponential growth in second example.
fix r, and let r, + cc. On the other hand, if we fix r, and let r, + cc, then the Malthusian parameter X approaches the solution of the equation flgx( r;) = 1. The behavior of X in the riro plane for two values A, and A, with A, < A, -C A, is illustrated in Figure 8. The vertical asymptotes give critical values of r, for which the Malthusian parameter is bounded below no matter how large the rate r, becomes.
It is interesting to compare the first example with mortality concentrated in the younger ages and the second example with mortality concentrated in the older ages. Without quiescence the Malthusian parameter for the first
example (the solution of (p/X) e-- ~@+‘)(l- e-i’) =l) is always less than
the Malthusian parameter for the second example (the solution of [fie-i’/(A + p)](l- e-fc ‘+p)) = 1). In the presence of quiescent population
this relationship can be reversed. For example, take j3 = 3 and p -1. For the first example the Malthusian parameter without quiescence is positive, which implies asynchronous exponential growth. In Figure 9 we have plotted the curves X = 0 as a function of ri and r, for both examples. The solid curve is for the first example and the dashed curve is for the second.

AGE-SIZE STRUCTURE WITH QUIESCENCE 87
5. AN ANALOGOUS DISCRETE MODEL
The well-known Leslie-Lewis-Benardelli theory of discrete age structured populations (see [12]) can be carried over to age-size structured populations. In this section we discuss a discretized time-age-size population model that also incorporates quiescence. We assume that the population is divided into p age classes and q size classes with q < p. For simplicity we assume that all individuals are born into the first age and first size class. Let no,s( t) be the number of individuals in age class a and size class s at time t with s < a. Let b,_,s be the number of newborns from age class a and size class s in the time interval At. Let a,,, be the fraction in age class a and size class s that transits to age class a + 1 and size class s + 1 in the time interval At. Let y,., be the fraction in age class a and sire class s that transits to age class a + 1 and size class s in the time interval At. The movement of individuals to various classes is illustrated in Figure 10, where normal individuals advance
diagonally and quiescent individuals advance horizontally.
FIG. 10. Transition scheme between discrete age-size classes

88 M. GYLLENBERG AND G. F. WEBB
Unlike our continuous model, which describes normal and quiescent individuals with different dependent variables, this discrete model combines the two types into one dependent variable, but distinguishes their propor- tions. The change in the age-size classes from time t to time t + At is given by the formulas
u=2s=2
n2Jt + AtI = Yllnl,lW
n2,20 + W = ~llnl,lW
n3.A t + Ad = Y2ln2.d d
n3,2U+A4 =a21n2,,(r)+Y22n2,2(r)
n3,3@ + At) = ~2,n2,2W
For an example we take p = q = 4, b,, = b,, = bd2 = 0, bj3 = bu = bd3 = 2, d 11=(1-Poh, ~22=(1-Po)c,, a,,= (I- P,) c3 9 Yll = POClT Y22 = P&2 9 Y33 = p0c3, u21 = u3, = p,. The values ci, c2, c3 are given constants in (0,l). The parameter p0 (0 Q p, d 1) measures the movement of individuals our of normal growth to quiescence. If p0 = 0, then no individual becomes quies- cent, and if p, = 1, then all do. The parameter pi (0 G pi G 1) measures the movement of individuals into normal growth from quiescence. If pi = 0, then all quiescent individuals remain quiescent, and if pi = 1, then all quiescent individuals reenter normal growth.
In term: of the classes_contributing to reproduction this example has the equation N( r + A r) = AN(r), where
G(r) =
nl,d d n2,1w n2.2( d n3,2( 0 n3.3( 0 %,3W
nd4.
7 A=
0 0 0 0 b,, bd3 b4
yll 0 0 0 0 0 0
(Jll 0 0 0 0 0 0
0 021 Y22 0 0 0 0
0 0 022 0 0 0 0
o o o u32 Y33 o o
0 0 0 0 a33 0 0
The matrix A is a square matrix with only nonnegative terms. If 0 < p0 < 1, 0 -C pi G 1, then the terms of A” are strictly positive for sufficiently large m. By the Perron-Frobenius theorem (see [12, p. 391) A has a dominant real positive algebraically simple eigenvalue A which possesses a strictly positive

AGE-SIZE STRUCTURE WITH QUIESCENCE 89
eigenvector. The dominant eigenvahte X is the unique positive solution of the characteristic equation
det( A - XI) = - A’ + A4b,,a,,u,,
+ A,( b43( %~22Y3, + Q-Y2242 + YllU21U32)
+ hJ,*~2*%3 > = 0 (5.1)
The terms inside the braces in (5.1) can be easily interpreted in terms of possible pathways in Figure 10. If X > 1, then the population has asynchro- nous exponential growth, and if X < 1, then the population becomes extinct
(see [12, p. 471). The behavior of the population in terms of the parameters p. and pi can
be obtained by setting X = 1 in (5.1) and solving for p, and pi. The behavior
PO 1.0 -
FIG. 11. Region 1: extinction. Region 2: asynchronous exponential growth.

M. GYLLENBERG AND G. F. WEBB
1,
FIG. 12. Region 1: extinction. Region 2: asynchronous exponential growth.
for the specific values c, = 0.9, c2 = 0.3, and cs = 0.7 is given in Figure 11, and for c1 = 0.9, c2 = 0.4, c3 = 0.7 in Figure 12. In the first case there is higher mortality in the middle ages than in the second case. For the first case the population becomes extinct if there is no quiescence (p, = 0), but escapes extinction if p0 and pi are sufficiently large. For the second case the population is growing exponentially if there is no quiescence (p, = 0), but becomes extinct if p0 and pi are sufficiently large.
If p, = 0, then the entries of Am are not ultimately strictly positive, but there still exists a dominant simple eigenvalue.
APPENDIX
We provide here the proof of the asynchronous exponential growth of the solutions of the model. We first state some results from positive operator

AGE-SIZE STRUCTURE WITH QUIESCENCE 91
theory. Let X be a Banach lattice, and let X* be its dual space. Let X, and XT denote the positive cones of X and X*, respectively. A bounded linear operator L in X is positive if and only if it leaves X+ invariant. L is irreducible if and only if for all x E X+ , x # 0, x* E X:, x* # 0, there exists a positive integer n such that (L”x, x*) > 0. The following theorem is proved in [ll]:
THEOREM I (de Pagter)
If L is a positive irreducible compact bounded linear operator in the Banach lattice X with dim X > 1, then L has spectral radius r(L) > 0.
Let T(t), t > 0, be a strongly continuous semigroup of positive bounded linear operators in X with infinitesimal generator A. The spectral bound of r(t), t 3 0, is X, := sup(Re X: X is in the spectrum of A}. T(t), t z 0, is irreducible if and only if for all x E X, , x f 0, x* E X,*, x* # 0, there exists t > 0 such that (T( t)x, x*) > 0. The following theorem is proved in [l, Part C]:
THEOREM 2 (Greiner)
Let T(t), t > 0, be an irreducible strongly continuous semigroup of positive
bounded linear operators in the Banach lattice X such that for some t, > 0, T( to) is compact and has spectral radius r( T( to)) > 0. Then there exists a
rank one projection PO in X and constants 6 > 0, M 2 1 such that T(t) PO =
POT(t) = e -xo’PO and le- ‘o’T( t) - PO I< Me-“‘, t > 0, where A, is the spec- tral bound of T(t), t > 0.
We now apply the general theory of semigroups to the semigroup defined in Section 3.
THEOREM 3
The semigroup U(t), t 2 0, is compact for t > a,.
Proof. The semigroup T(t), t 2 0, in (3.1) may be decomposed as T(t) =
T,(t)+ G(t), where (T,(t)+)(a, x) = B(t - a, X(0, a)x)n,(O, a, X(0, a)x) for a< t, (a,x)E&, (T,(t)$)(a,x)=O otherwise, and T2(t)=T(t)- TI (t). From the definition of B we see that TI( t) is a compact operator in L’(0) for t 2 0 (see [5, p. 2981). Further, T2(t) = 0 for t > a,. From (3.2) we see also that S(t) = 0 for t > a,. Define
Define the zeroth generation at time t as U,(t) := V,(t) + w,(t), and define
the (n + 1)st generation at time t as
u,+,(t) :=~vo(t-s)Ku”(s)dF+pqt-s)KUj,(s)ds.

92 M. GYLLENBERG AND G. F. WEBB
From (3.4) and (3.5) we see that U(t) =E~_,,U,(t). Observe that
Since I%(t) is compact for t > 0, the last two terms are compact for f 2 0.
From the formula for V,(t) we see that the first term is 0 for t 2 a,. Next observe that
U,(r)=Id~(f-s)~~~~(s-r)KV,(r)drdF
+ terms which are compact for t > 0.
The formula for I/,(r) again assures that the first term is 0 for t > a,. A similar argument applies to each U,(t), n >, 1. Since the compact operators are closed in B( L’), U(t) is compact for t >, a,.
THEOREM 4
Thereexistst*>Osuchthatforall {$,$}EL$, {+,~}+{O,O}, (11,~)
EL:*, {v,P} + {O,O}, andt>t*, oneha (W~){~,#),{CP})>O.
Proof. We first show that there exists t, > 0 such that for all $I E Li (a), + # 0, and t > t,, one has T(t)+ > 0 a.e. in 9,. From (3.2) and (3.7) we see
that
B(t,x) =jb”i”(*~““~x:bq~~~(a,y,x)~(~-a,X(O,a)y)
x&@,a,X(O,a)y) &da
+I;,(, ~~)~~8(a,Y~x)9(a-f~x(u-r,u)Y) 0
xl-I,,,(a-tt,a,X(a-rt,a)y)dyda. (A.11
We claim that the hypothesis (Hg) implies there exists t, Q a,, such that B(t,,x)>O a.e. x~(xa,xi). If supp(p~{(a,x)~O(a~a,}#0, choose t,=O. If suppcp~{(a,x)~Q~aaa~}=0, choose t,<a, such that supp$(a-~,,X(U-t,,a)x)n((a,x)EDla~a,}#0. We next claim that B( t, x) > 0 a.e. x E (x,, x1) and t E (a,, + t,, a, + to), since for these t we can choose UE(U,,U,) such that t-a=tt, and /3(a,y,x)B(t- a,X(O,a)y)=/?(a,y,x)B(t,,X(O,a)y)>O a.e. In a similar fashion we can show that B(t, x) > 0 a.e. x E (x0, xi) and t E (ka, + t,, ka, + to), k =1,2,... . Now choose t, = ka,, where k is the first integer such that

AGE-SIZE STRUCTURE WITH QUIESCENCE 93
q < k(a, - a,), to obtain B(r, x) > 0 a.e. x E (x,, xi), r > t,. From (3.1) we obtain T(t)+ > 0 a.e. in O,, t > t,.
To prove the theorem let { $, I/J} E Lc, (4, I/J} # {O,O}. From (3.5) we obtain
n(t)=T(t)OfldT(t-s)r;q(s)ds, (A.2)
4(r)=S(t)~+lbS(t-S)r~n(s)ds. (A.3)
If I$ # 0, then, as shown above, T(t) $J > 0 a.e. in Sz, for t > t, , and hence by (A.2) n(t) > 0 a.e. in 8,) r > 1,. If $J # 0, then T(t)r,q(O) = T(t)ri$ > 0 a.e. in 9, for t > t, [recall that by (H,), r; > 01. It now follows from (A.2) that n(t) > 0 a.e. in 52, for t > t,.
Let 0, be the smallest set such that
(2) if ((Y, x) E Q2,, then (a,~) E fit for all a E [cY,u,].
Let (u,x)~S&. Then there exists an LYE[O,U,] such that (a-a,x)~&,.
Since S(t -~)r,n(s)(u,x)=r,(u-(t-s),~)n(s)(u-(t-s)),x)~~(u- (t-s),u,x), we have s(t-s)r,n(s)>O a.e. in 8, for all t>t,+u, and some s > t,. (A.3) now implies that q(r) > 0 a.e. in Q2, for t > t, + a,, and from (A.2) we can then deduce that n(t) > 0 a.e. in Q2, for t > t, + ul.
Let Q2, be the smallest set such that
(1) 4 = a*, (2) if (OL, x) E Cl,, then X(u, a)x E Q2, for all a E [a, ai].
Let (a, x) E Qt,. There exists an (Y E [0, aI] such that (u - (Y, X( a - cq a)~) E 0,. Since, as shown above, q(s) > 0 a.e. in 8,, s > t, + ul, we have
T(t - s)r,q(s)(u, x) = );(a -(t - s), X(a -(t - s), u)x)q(s)(u -(t - s),X(u-(t-s),u)x)II,(u-(t-s),u,X(u-(t-s),u)x)>O a.e. in 8, for all t > t, + 2u, and some s > t, + aI (set cy = t - s < a < al). Using (A.2) and (A.3), we find exactly as above that n(t) and q(r) are positive a.e. in 9,.
Continuing in this way and extending the domain Qj in turn along the characteristics of the quiescent and normal states, we obtain a larger and larger domain on which n(t) and q(t) are positive a.e. By the definition of G and our assumption in (H,) that the boundaries of D can decrease only on finitely many intervals, this process terminates after a finite number of steps: Q, = fJ for some finite k. Thus both n(t) and q(t) are positive almost everywhere in the entire domain Q provided t > t* =: t, + ku,. This proves the theorem.

94 M. GYLLENBERG AND G. F. WEBB
THEOREM 5
The semigroup U(t), t > 0, has the property of asynchronous exponential growth.
Proof. The proof follows immediately from Theorems 1, 2, 3, and 4.
REFERENCES
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
W. Arendt, A. Grabosch, G. Greiner, U. Groh, H. P. Lotz, U. Moustakas, R. Nagel, F.
Neubrander, and U. Schlotterbeck, in One-Parameter Semigroups of Positive Operators
(R. Nagel, Ed.), Lecture Notes in Mathematics 1184, Springer, New York, 1986.
G. I. Bell and E. C. Anderson, Cell growth and division. I. A mathematical model with
applications to cell volume distributions in mammalian suspension cultures, Biophys.
J. 7:329-351 (1967).
H. Caswell, The evolutionary demography of clonal reproduction, in Popularion
Biology und Evolution of Clonal Organisms (J. B. C. Jackson, L. W. Buss, and R. E.
Cook, Eds.), Yale U.P., New Haven, 1985.
0. Diekmann, H. J. A. M. Heijmans, and H. Thieme, On the stability of the cell size
distribution, J. Math. Biol. 19(2):227-248 (1984).
N. Dunford and J. Schwartz, Linear Operators, Part I: Generul Theory, Interscience,
1958.
M. Eisen, Mathematical models in cell biology, in Lecture Notes in Biomathematics,
Vol. 30, Springer, New York, 1979.
W. Greenberg, V. Protopopescu, and C. V. M. Van der Mee, Boundary Value Problems
in Abstract Kinetic Theory, Birtiauser, to appear.
J. K. Hale, Ordinary Differential Equations, Interscience Series on Pure and Applied
Mathematics, Vol. 21, Wiley-Interscience, New York, 1969.
N. R. Hartmann, The continuity equation, a fundamental in modeling and analysis in
cell kinetics, in Modeling and Analysis in Biomedicine (C. Nicolini, Ed.), World
Scientific Publishing Company, Singapore, 1984.
H. J. A. M. Heijmans, The dynamical behaviour of the age-size distribution of a cell
population, in Dynamics of Physiologically Structured Populations (J. A. J. Metz and 0.
Diekmann, Eds.), Lecture Notes in Biomathematics, Vol. 68, Springer, 1986, pp.
185-202.
B. de Pagter, Irreducible compact operators, Math. 2. 192:149-154 (1986). J. H. Pollard, Mathematical Models for the Growth of Human Populations, Cambridge
U.P., Cambridge, 1973.
M. Rotenberg, Correlations, asymptotic stability and the G,, theory of the cell cycle, in
Biomathematics and Cell Kinefics, Developments in Cell Biology, Vol. 2 (A.-J. Valleron
and P. D. M. Macdonald, Eds.), Elsevier North Holland Biomedical Press, 1978, pp.
59-67.
M. Rotenberg, Theory of distributed quiescent state in the cell cycle, J. Theoret. Biol. 96:495-509 (1982).
M. Rotenberg, Transport theory for growing cell populations, J. Theoret. Biol.
103:181-199 (1983).
E. Sinestrari and G. F. Webb, Nonlinear hyperbolic systems with nonlocal boundary
conditions, J. Math. Anal. Appl. 121:449-464 (1987).

AGE-SIZE STRUCTURE WITH QUIESCENCE 95
17 J. W. Sink0 and W. Streifer, A new model for age-sire structure of a population,
Ecologv 48:910-918 (1967).
18 G. F. Webb, Dynamics of populations structured by internal variables, Math. Z.
189:319-335 (1985).
19 R. White, A review of some mathematical models in cell kinetics, in Biomarhematics
und Cell Kinetics, Developments in Cell Biology, Vol. 8 (M. Rotenberg, Ed.), Elsevier
North-Holland Biomedical Press, 1981, pp. 243-261.
20 J. J. Wille, Jr., and R. E. Scott, Cell cycle-dependent integrated control of cell
proliferation and differentiation in normal and neoplastic mammalian cells, in Cell
Cycle Clocks (L. N. Edmunds, Jr., Ed.), Marcel Dekker, New York, 1984.
21 H. Wu, P. J. H. Sharpe, and E. J. Rykiel, Jr., Age-sire population dynamics: derivation
of a general matrix methodology, to appear.
Copyright © 2022 FDOKUMEN



![GÉOPOLITIQUE ET POPULATIONS AU TCHAD [Geopolitics and populations in Chad]](https://static.fdokumen.com/doc/165x107/631378e5fc260b71020f1c3f/geopolitique-et-populations-au-tchad-geopolitics-and-populations-in-chad.jpg)
















