
Central nervous system myelination in mice with deficient expression of Notch1 receptor
Upload
independentCategory
view
0download
0
In Vitro Myelination byOligodendrocyte Precursor Cells
Transfected With theNeurotrophin-3 Gene
NAZARIO RUBIO,* RODRIGO RODRIGUEZ, AND MARIA ANGELES AREVALOInstituto Cajal, Consejo Superior de Investigaciones Cientificas (CSIC), Madrid, Spain
KEY WORDS neurotrophin-3; oligodendrocyte precursors; myelination; myelin basicprotein; regeneration
ABSTRACT Oligodendrocyte precursor cells require exogenous neurotrophin-3(NT-3) for differentiation into oligodendrocytes. We transfected precursor cells with thegene for NT-3 and looked for changes in their development into myelin-forming cells.The expression of NT-3 in transfected cells was demonstrated by reverse transcriptionfollowed by PCR as well as by Northern blots. Direct synthesis of the neurotrophinproduct and its release to the culture supernatants were also shown by specific ELISA.Transfection converts precursor cells into actively dividing cells that can incorporate3H-thymidine into DNA. In the absence of growth factors, a parallel increase in thesurvival of the transfected cultures was also demonstrated by the MTT test. The finaldemonstration of biological changes in transfected versus untreated cells was a 10-foldincrease in myelin basic protein production observed in Western blots and the directobservation by phase-contrast and electron microscopy of myelin membranes in cocul-tures with hippocampal neurons. We discuss the future use of this transfected cells inregeneration and functional recovery in experimental models of multiple sclerosis.© 2004 Wiley-Liss, Inc.
INTRODUCTION
Oligodendrocytes are nervous system cells responsi-ble for the synthesis of myelin. They arise from progen-itors or precursors that proliferate, differentiate, andmyelinate axons in postnatal stages (Pfeiffer et al.,1993; Miller, 1996). These bipotential precursor cells oroligodendrocyte precursor cells (OPCs) give rise to as-trocytes (in cultures containing 10% FCS) or to oligo-dendrocytes (in serum-free cultures) (Bogler et al.,1990; Barres et al., 1992; Kondo and Raff, 2000) whencultured in vitro. Neurotrophin-3 (NT-3) regulates theproliferation and differentiation of oligodendrocytes inculture (Barres et al., 1994; Cohen et al., 1996; Hein-rich et al., 1999). Platelet-derived growth factor(PDGF) and fibroblast growth factor (FGF) promotedivision and prevent the differentiation of OPCs (Nobleet al., 1988; Bogler et al., 1990).
There have been many reports of experimental ther-apeutic interventions in rats aimed at promoting the
regeneration of demyelinated corticospinal tracts invivo. The development of appropriate animal modelsand these successful cellular therapeutic interventionswill be crucial for the treatment of all kinds of centralnervous system disorders in human clinical practice.There has also been previous reports of the stimulationof myelinogenesis in culture by microglia (Hamiltonand Rome, 1994) or by the interleukin-6 receptor-inter-leukin-6 fusion protein (Haggiag et al., 2001). Thepresent experiments were designed to determinewhether oligodendrocyte precursor cells transfected
Grant sponsor: Direccion General de Investigacion, Ministerio de Ciencia yTecnologia, Spain; Grant number: PM99-0102.
*Correspondence to: Nazario Rubio, Instituto Cajal, CSIC, Dr. Arce Avenue 37,28002 Madrid, Spain. E-mail: [email protected]
Received 7 November 2003; Accepted 23 December 2003
DOI 10.1002/glia.20035
Published online 24 March 2004 in Wiley InterScience (www.interscience.wiley.com).
GLIA 47:78–87 (2004)
© 2004 Wiley-Liss, Inc.

with the NT-3 gene were able to stimulate in vitrosynthesis of myelin with respect to untreated precursorcells. We genetically engineered these cells to inducethe production of the proregenerative factor NT-3,which promotes myelination and axonal regeneration.Our findings demonstrated the enhanced in vitro pro-duction of myelin by NT-3-transfected cell (OPCNT-3),quantifying the myelin basic protein (MBP) producedby Western blotting and characterizing the myelinmembranes formed by electron microscopy.
Multiple sclerosis (ME) is an immune-mediated de-myelinating disorder of the human nervous system.Two different experimental animal models are com-monly used to study ME; experimental autoimmuneencephalomyelitis (EAE) and Theiler’s murine enceph-alomyelitis virus (TMEV)-induced demyelination. OurNT-3-transfected cells had promising therapeutic pros-pects due to its in vitro increasing division ability andNT-3 production. As both effects surely could promoteoligodendrogenesis and maturation of host precursorcell populations producing myelin-forming cells, weplan to graft OPCNT-3 cells in TMEV-demyelinatedmice in order to study the possible induction of recov-ery in sick animals.
MATERIALS AND METHODSOPC Cultures
The following technique for oligodendrocyte precur-sor isolation was modified from that of McCarthy andde Vellis (1980) and Molina-Holgado et al. (2001).Briefly, mixed glial cultures were prepared by mechan-ical dissociation of the cerebral cortex of newbornWistar rats for 8–10 days (Rubio et al., 1990) in Dul-becco’s minimum essential medium (MEM) with glu-tamax-I containing 10% fetal calf serum (FCS) andgentamicin (Gibco-BRL, Life Technologies, Scotland).At this time, oligodendrocyte progenitor bright cellswere present on the top of astrocyte monolayers. Con-taminating microglia was dislodged by shaking for 2 hat 37°C in a G24 environmental shaker (New Bruns-wick Scientific, Edison, NJ) at 220 rpm. Oligodendro-cyte progenitors were removed from the astrocytemonolayer by further shaking for 18 h at 280 rpm andfiltered trough a 40 �m nylon cell strainer (Falcon-Becton Dickinson, Le Pont de Claix, France) to removeany large cell aggregates. Oligodendrocyte progenitorswere further purified after centrifugation by selectiveplating on bacterial-grade petri dishes twice for 1 heach at 37°C. Under these conditions, microglia andastrocytes adhered to the plastic and oligodendrocyteprogenitors remained in suspension. After centrifuga-tion, purified cells were plated out (2 � 105 cells perwell) on poly-D-lysine (Sigma, St. Louis, MO)-coated6-well multiwell culture plates (Falcon-Becton Dickin-son) for 3 h in complete medium. Medium was thenchanged to serum-free defined medium composed ofDMEM/F-12 medium containing O.1% bovine serumalbumin fraction V, N-2 supplement (containing insu-
lin, human transferrin, progesterone, putrescin, andselenite), and gentamicin, all from Gibco-BRL. Theabove medium was supplemented with 5 ng/ml plate-let-derived growth factor (PDGF-AA) and fibroblastgrowth factor-basic (bFGF; Sigma) in order to expandthe number of progenitors and prevent their differen-tiation. These conditions allowed the formation of bi-polar bright cell cultures reaching approximately 70%confluence after 4 days.
Plasmid Purification and Transfection
A plasmid Bluescript encoding the NT-3 gene(pNT-3) was a generous gift of Dr. Kevin R. Jones fromHoward Hughes Medical Institute Research Labora-tory (San Francisco, CA) (Jones and Reichardt, 1990).A plasmid encoding rat MBP designed pRMBP700(pMBP) was a gift from Dr. L. Wrabetz, Department ofNeurology, San Raffaele Scientific Institute (Milano,Italy). All plasmids were purified using the Miniprepprotocol of the Concert rapid purification system fromGibco-BRL and its purity was checked by electrophore-sis in 1% agarose. For the transfection of the NT-3 geneor the control MBP gene, the Effectene transfection kitfrom Qiagen (Valencia, CA) was used.
RT-PCR Analysis of NT-3 Expression
Total RNA from normal or transfected oligodendro-cyte precursor cultures was purified by using theRNeasy Mini purification kit (Qiagen). Samples werereverse-transcribed using Moloney murine leukemiavirus reverse transcriptase (RT; Promega, Madison,WI) and the 3� amplimer as template/primer. Previ-ously, RNA samples were treated with 10 units ofDNAseI RNAse-free (Boehringer Mannheim, Ger-many) for 15 min at 37°C to destroy chromosomal DNAcontaminations. Amplimers used for NT-3 amplifica-tion were 5� (5�-CAAAGGAGTTTGCCGGAAGACTCT-3�) and 3� (5�-AATGGCTGAGGACTTGTCGGTCAC-3�). PCR reactions were carried out after a 5-min cycleat 90°C for 30 cycles at an annealing temperature of65°C using a Perkin Elmer Cetus 480 DNA thermalcycler. The resulting PCR products were purified usingS-400 MicroSpin columns (Pharmacia Biotech, Upp-sala, Sweden) and electrophoresed in 2% NuSieve aga-rose gels (FMC Bio Products, Rockland, ME) in TAEbuffer, stained with ethidium bromide, and photo-graphed in a Gelstation system (TDI, Barcelona,Spain). After electrophoresis in 2% agarose, the DNAband was excised from the agarose gel with a scalpeland purified using the QIAEX II kit provided by Qia-gen. This DNA was used as a probe for Northern blotanalysis.
79NT-3-TRANSFECTED OLIGODENDROCYTE PRECURSORS

Northern Blot Analysis
Total RNA was purified using the RNeasy Mini pu-rification kit (Qiagen); 10 �g/lane were electrophoresedon a formaldehyde-agarose gel and transferred to nylonZeta-Probe membranes (Bio-Rad Laboratories, Her-cules, CA). Probes were labeled with 32P-dCTP usingthe random primed method (Boehringer). Unincorpo-rated radiolabel was removed with S-300 MicroSpincolumns (Pharmacia Biotech). The blots were exposedto Kodax X-OMAT film with an intensifying screen at�75°C. Hybridization to a human �-actin probe (Clon-tech, Palo Alto, CA) was done as an internal control forloading.
ELISA Determination of NT-3
The NT-3 Emax ELISA kit (Promega) was used forNT-3 determinations as described by the manufac-turer. Results were read at 450 nm in a Titertek Mul-tiskan Plus MK II reader (Flow Laboratories, Irvine,U.K.). The kit has less than 3% cross-reactivity to re-lated neurotrophic factors at a concentration of 10 pg/ml.
Thymidine Incorporation
OPCs in 96-well clusters (Falcon-Becton Dickinson)were incubated overnight in serum-free defined me-dium containing FGF and PDGF, washed, and trans-fected or not with the plasmid NT-3. The cultures werepulsed with 1 �Ci of [3H] thymidine per well (5 Ci/mmol; Amersham) for different periods of time, washedtwice with cold PBS, and lysed by incubating with 50 �lof 0.4% SDS for 1 h at room temperature; 150 �l ofEcoscint A scintillation cocktail (National Diagnostics,Atlanta, GA) were then added per well, and after anovernight period, the amount of [3H] thymidine incor-porated into DNA was determined in a 1450 MicroBetaTrilux liquid scintillation and luminiscence counterfrom Wallac (Turku, Finland).
MTT Test
The method of Mosmann (1983) was used. Briefly,after different periods of culture, 5 �l of a sterile 3-(4,5-dimethylthiazol-2-yl)-2.5-diphenyltetrazolium bromide(MTT; Sigma) solution (1.5 mg/ml in culture medium,stored at �20°C in aliquots and thawed immediatelybefore use) were added to each well and the platescultured for a further 24 h.
MTT-containing cultures were terminated by adding50 �l of isopropanol-0.1 N HCl. The isopropanol-acidand culture medium were mixed by up- and downaspi-rations using a 12-channel pipette set at 50 �l volume.After no longer than 1 h, plates were read on a Titertek
Multiskan reader at a test wavelength of 570 nm and areference wavelength of 630 nm.
Purification of MBP and Preparation ofAntiserum
Myelin basic protein from white matter of rats wasisolated by acid extraction of defatted material accord-ing to the method of Oshiro and Eylar (1970). All buff-ers and organic solvents contained 50 �g/ml of PMSF(Sigma) as a protease inhibitor. Final purification wasaccomplished by high-performance liquid chromatogra-phy (HPLC) using a Protein-Pack 125 column (Waters,Milford, MA) and a Perkin-Elmer series 3B HPLC sys-tem (Perkin-Elmer, Norwalk, CT). Purity of MBP wasmonitored by SDS-PAGE electrophoresis (Laemmli,1970). Guinea pigs were immunized over a 2-monthperiod with four i.p. injections of 200 �g of HPLC-purified rat MBP in Freund’s complete adjuvant (DifcoLaboratories, Detroit, MI). The animals were bled bydecapitation 10 days after the last booster and anti-serum were aliquoted and stored at �20°C until used(Rubio and Cuesta, 1989).
Western Blots
Cocultures of hippocampal neurons with nontrans-fected or NT-3-transfected OPCs were solubilized with20 mM Tris, pH 7.4 buffer, containing 1 mM MgCl2, 1mM PMSF, and 10 �g/ml leupeptin. The protein con-centration was measured by the RC DC Protein Assay(Bio-Rad). After centrifugation and electrophoresis on12% polyacrylamide gels, separated proteins weretransferred to nitocellulose membranes and blockedwith 10% defatted milk in PBS. Membranes were in-cubated with 1:1,000 dilutions of the guinea pig anti-serum against MBP, washed in Tris buffer, pH 7.4,containing 0.1% Tween, followed by treatment with a1:10,000 dilution of protein A-horseradish peroxidase(Pharmacia Biotech) for 2 h at room temperature. Afterthree washes with Tris buffer-0.1% Tween, immunode-tectable bands were visualized by chemoluminescence(ECL, Amersham).
Myelinating of Hippocampal Neuron Cultures
The hippocampal neuron cultures have been previ-ously described by Dotti et al. (1988). Briefly, hip-pocampi were dissected from the brains of 18-day-oldrat fetuses, treated with 0.25% trypsin for 15 min at37°C, washed in Ca-Mg-free Hank’s Balanced Salt So-lution, and dissociated by repeated passage through aconstricted Pasteur pipette. Between 2,000 and 6,000cells were added to poly-D-lysine-coated culture dishesin MEM containing 10% horse serum. After 3–4 h at37°C, when the cells had attached to the substrate, themedia were replaced with MEM, supplemented with 10
80 RUBIO ET AL.

mM HEPES (pH 7.3), pyruvate (0.01 mg/ml), and theN-2 supplement. After that, NT-3-transfected OPCswere added on the neurons at a concentration of 2 �104/cm2 and after a 2- to 3-week period tested for my-elination by microscopy or Western blotting.
Immunochemistry, Phase-Contrast, andElectron Microscopy
Immunostaining of different antigens was done onlive cultures incubated for 30 min at room temperaturewith different dilutions of rabbit antiglial fibrillaracidic protein (anti-GFAP) antiserum (Dakopatts,Glostrup, Denmark), rabbit antigalactocerebroside(anti-GalC) antiserum (Sigma), anti-A2B5 monoclonalantibody (a kind gift of Dr. E. Molina-Holgado, Insti-tuto Cajal, Madrid, Spain), and a monoclonal anti-Mac-1 antibody (Serotec, Oxford, U.K.). After washingwith PBS-1% FCS, the cultures were incubated for 30min at room temperature with the suitable dilutions ofthe corresponding secondary fluorescein-labeled anti-bodies (purchased from Sigma), washed, and fixed with4% paraformaldehyde. The stained cells were examinesunder an Axiovert 35 (Zeiss, Germany) inverted micro-scope equipped with an MC100 camera. Morphologicalobservations and photomicrographs under phase con-trast were performed on living cultures using the Ax-iovert microscope. Myelinating cultures were preparedfor electron microscopy in polycarbonate membraneinserts of 3 �m pore size (Nunc, Roskilde, Denmark).Membranes were washed with PBS, removed from theinsert, then fixed with 2% paraformaldehyde plus 2%glutaraldehyde in 100 mM sodium cacodylate at pH 7.4for 48–72 h at 4°C. Cells were postfixed with 1% OSO4,dehydrated in ethanol, and embedded in Epon/Araldite. After thin sectioning and poststaining in leadcitrate, the specimens were observed by transmission
electron microscopy on a JEOL 100-EX electron micro-scope.
RESULTSTransfection Optimization
To achieve optimal transfection levels and minimalcell cytotoxicity, the original protocol provided by themanufacturer of the Effectene transfection kit usedwas optimized by changing several variables such asthe amount of plasmid DNA, amount of Effectene re-agent, DNA/Effectene ratio, removal of complexes, etc.We found that optimal transfection efficiency for OPCswas achieved using 1 �g of plasmid DNA and 10 �l ofEffectene (ratio 1:10) per well of a 6-well plate. Theseconditions were therefore applied to all the experi-ments.
RT-PCR Analysis
The presence of neurotrophin-3 mRNA in cultures ofmouse oligodendrocyte precursor cells, which were ei-ther untreated or transfected with the pNT-3 vector(OPCNT-3), was determined by RT-PCR. Specific am-plimers (described above) were used to amplify a 430bp fragment from the plasmid, which encodes the NT-3gene (Fig. 1, lanes �). Oligodendrocyte precursorstransfected with control plasmid encoding rat myelinbasic protein (lane �) or nontransfected (lane 0) do notexpress the NT-3 gene. This was suggested previouslyby the fact that exogenous NT-3, probably synthesizedby astrocytes, is needed for oligodendrocyte develop-ment (Barres et al., 1994). Conversely, mRNA is fullyreverse-transcribed and amplified from cells trans-fected with the plasmid pNT-3 (Fig. 1, left). The rightpanel of Figure 1 shows that NT-3 mRNA production
Fig. 1. Expression of NT-3 mRNA in transfected cells. In the left,plasmid pNT-3 amplified as a positive control by PCR with NT-3-specific amplimers produces a fragment of 430 bp (lane �). RNA fromcultures transfected with the control pMBP (lane �), mock-trans-fected (0), or transfected with 0.2, 1, and 2 �g of plasmid DNA wasreverse-transcribed and PCR-amplified. The kinetics of the reaction
was shown in the right panel. The positive control (�), and the RNAfrom cultures transfected with 1 �g of plasmid DNA for periods oftime ranging from 2 to 48 h were amplified. Lanes MW: DNA molec-ular weight markers (pBR322 DNA-MspI digest; New England Bio-labs).
81NT-3-TRANSFECTED OLIGODENDROCYTE PRECURSORS
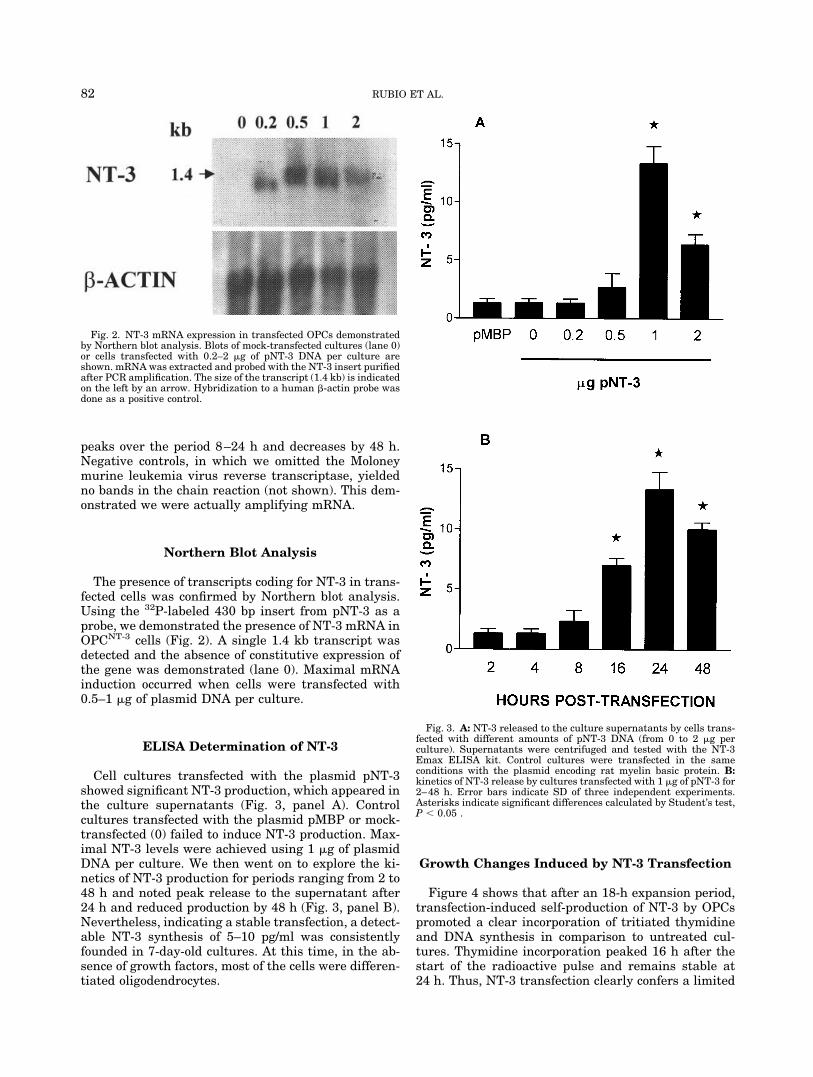
peaks over the period 8–24 h and decreases by 48 h.Negative controls, in which we omitted the Moloneymurine leukemia virus reverse transcriptase, yieldedno bands in the chain reaction (not shown). This dem-onstrated we were actually amplifying mRNA.
Northern Blot Analysis
The presence of transcripts coding for NT-3 in trans-fected cells was confirmed by Northern blot analysis.Using the 32P-labeled 430 bp insert from pNT-3 as aprobe, we demonstrated the presence of NT-3 mRNA inOPCNT-3 cells (Fig. 2). A single 1.4 kb transcript wasdetected and the absence of constitutive expression ofthe gene was demonstrated (lane 0). Maximal mRNAinduction occurred when cells were transfected with0.5–1 �g of plasmid DNA per culture.
ELISA Determination of NT-3
Cell cultures transfected with the plasmid pNT-3showed significant NT-3 production, which appeared inthe culture supernatants (Fig. 3, panel A). Controlcultures transfected with the plasmid pMBP or mock-transfected (0) failed to induce NT-3 production. Max-imal NT-3 levels were achieved using 1 �g of plasmidDNA per culture. We then went on to explore the ki-netics of NT-3 production for periods ranging from 2 to48 h and noted peak release to the supernatant after24 h and reduced production by 48 h (Fig. 3, panel B).Nevertheless, indicating a stable transfection, a detect-able NT-3 synthesis of 5–10 pg/ml was consistentlyfounded in 7-day-old cultures. At this time, in the ab-sence of growth factors, most of the cells were differen-tiated oligodendrocytes.
Growth Changes Induced by NT-3 Transfection
Figure 4 shows that after an 18-h expansion period,transfection-induced self-production of NT-3 by OPCspromoted a clear incorporation of tritiated thymidineand DNA synthesis in comparison to untreated cul-tures. Thymidine incorporation peaked 16 h after thestart of the radioactive pulse and remains stable at24 h. Thus, NT-3 transfection clearly confers a limited
Fig. 2. NT-3 mRNA expression in transfected OPCs demonstratedby Northern blot analysis. Blots of mock-transfected cultures (lane 0)or cells transfected with 0.2–2 �g of pNT-3 DNA per culture areshown. mRNA was extracted and probed with the NT-3 insert purifiedafter PCR amplification. The size of the transcript (1.4 kb) is indicatedon the left by an arrow. Hybridization to a human �-actin probe wasdone as a positive control.
Fig. 3. A: NT-3 released to the culture supernatants by cells trans-fected with different amounts of pNT-3 DNA (from 0 to 2 �g perculture). Supernatants were centrifuged and tested with the NT-3Emax ELISA kit. Control cultures were transfected in the sameconditions with the plasmid encoding rat myelin basic protein. B:kinetics of NT-3 release by cultures transfected with 1 �g of pNT-3 for2–48 h. Error bars indicate SD of three independent experiments.Asterisks indicate significant differences calculated by Student’s test,P � 0.05 .
82 RUBIO ET AL.
proliferative capacity upon OPCs, as reported for theaddition of recombinant mouse NT-3 to cultures ofthose cells isolated from the optic nerves of rats (Barreset al., 1994). NT-3 was also found to increase the num-ber of cells in cultures of oligodendrocyte precursorsabout 20% (Heinrich et al., 1999).
We also tested the capacity of NT-3 transfection tostimulate the proliferation of OPC in combinationwith its know mitogens FGF and PDGF. Both factorsbehave similarly, promoting division and self-renewal, as previously demonstrated (Noble et al.,1988; Bogler et al., 1990; Barres et al., 1992; Cohenet al., 1996). Figure 5 shows 3H-thymidine incorpo-ration in cultures treated with both added factors orin combination with the transfection-induced NT-3.Clearly, transfection-induced NT-3 prevented the de-cay of DNA synthesis starting after 8 h, probably dueto the consumption of the factors by the cultures, andmaintains them in a proliferative state for a longerperiod of time. Similar results were reported by Co-hen et al. (1996), who demonstrated that added re-combinant NT-3 behaved in the same way as endog-enous transfection-induced NT-3.
Survival of Untreated and pNT-3-TransfectedCultures
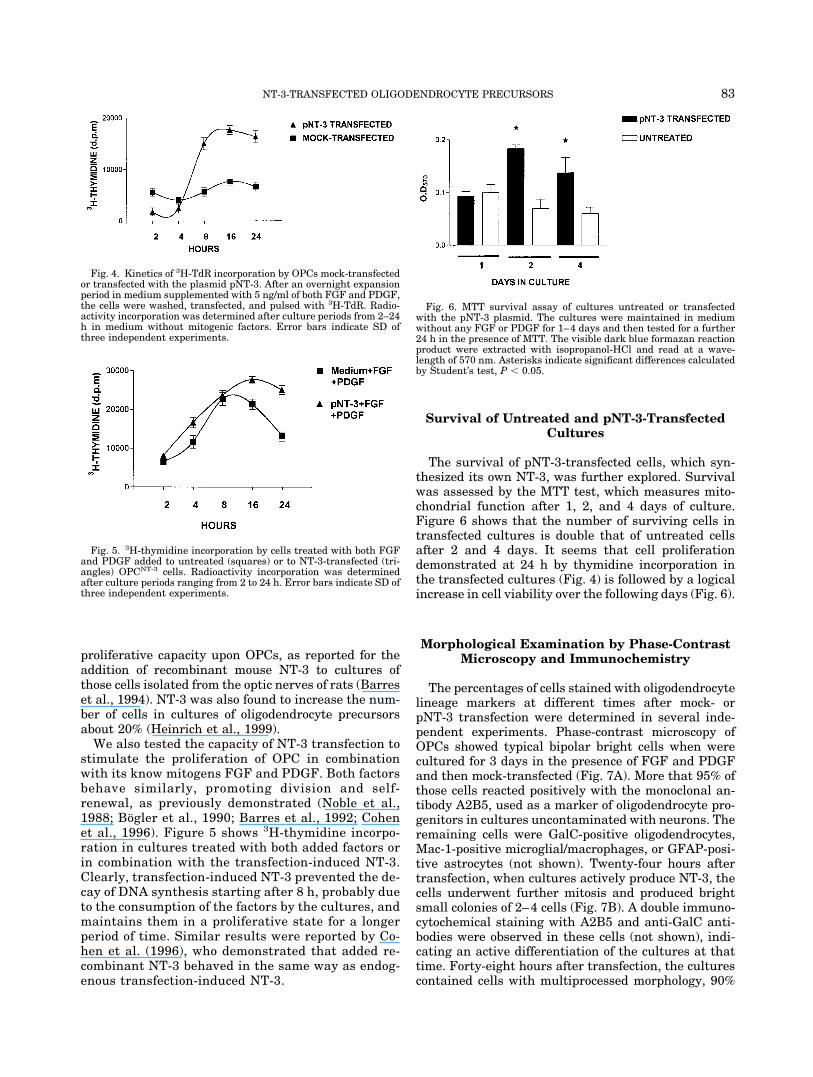
The survival of pNT-3-transfected cells, which syn-thesized its own NT-3, was further explored. Survivalwas assessed by the MTT test, which measures mito-chondrial function after 1, 2, and 4 days of culture.Figure 6 shows that the number of surviving cells intransfected cultures is double that of untreated cellsafter 2 and 4 days. It seems that cell proliferationdemonstrated at 24 h by thymidine incorporation inthe transfected cultures (Fig. 4) is followed by a logicalincrease in cell viability over the following days (Fig. 6).
Morphological Examination by Phase-ContrastMicroscopy and Immunochemistry
The percentages of cells stained with oligodendrocytelineage markers at different times after mock- orpNT-3 transfection were determined in several inde-pendent experiments. Phase-contrast microscopy ofOPCs showed typical bipolar bright cells when werecultured for 3 days in the presence of FGF and PDGFand then mock-transfected (Fig. 7A). More that 95% ofthose cells reacted positively with the monoclonal an-tibody A2B5, used as a marker of oligodendrocyte pro-genitors in cultures uncontaminated with neurons. Theremaining cells were GalC-positive oligodendrocytes,Mac-1-positive microglial/macrophages, or GFAP-posi-tive astrocytes (not shown). Twenty-four hours aftertransfection, when cultures actively produce NT-3, thecells underwent further mitosis and produced brightsmall colonies of 2–4 cells (Fig. 7B). A double immuno-cytochemical staining with A2B5 and anti-GalC anti-bodies were observed in these cells (not shown), indi-cating an active differentiation of the cultures at thattime. Forty-eight hours after transfection, the culturescontained cells with multiprocessed morphology, 90%
Fig. 4. Kinetics of 3H-TdR incorporation by OPCs mock-transfectedor transfected with the plasmid pNT-3. After an overnight expansionperiod in medium supplemented with 5 ng/ml of both FGF and PDGF,the cells were washed, transfected, and pulsed with 3H-TdR. Radio-activity incorporation was determined after culture periods from 2–24h in medium without mitogenic factors. Error bars indicate SD ofthree independent experiments.
Fig. 5. 3H-thymidine incorporation by cells treated with both FGFand PDGF added to untreated (squares) or to NT-3-transfected (tri-angles) OPCNT-3 cells. Radioactivity incorporation was determinedafter culture periods ranging from 2 to 24 h. Error bars indicate SD ofthree independent experiments.
Fig. 6. MTT survival assay of cultures untreated or transfectedwith the pNT-3 plasmid. The cultures were maintained in mediumwithout any FGF or PDGF for 1–4 days and then tested for a further24 h in the presence of MTT. The visible dark blue formazan reactionproduct were extracted with isopropanol-HCl and read at a wave-length of 570 nm. Asterisks indicate significant differences calculatedby Student’s test, P � 0.05.
83NT-3-TRANSFECTED OLIGODENDROCYTE PRECURSORS

of which were stained with anti-GalC antibodies (dif-ferentiated oligodendrocytes, Fig. 7C).
Control mock-transfected cultures takes a further 5-to 7-day period to reach this stage of differentiation.Hence, transfection with the NT-3 gene induces fasterdifferentiation into oligodendrocytes. The same effecthas been described in cultures treated with recombi-nant NT-3 (Heinrich et al., 1999).
Western Blot Detection of MBP Production
Next, we investigated whether self-production ofNT-3 by transfected cultures elicits further func-tional responses involved in myelination, in additionto 3H-thymidine incorporation or changes in the cul-tures survival. To this end, we analyzed cell homog-enates from cocultures using a guinea pig antiserumthat recognizes four major forms of MBP with molec-ular weight 14, 17, 18.5, and 21.5 kd in Westernblots. These four forms are produced by alternatesplicing of MBP mRNA (De Ferra et al., 1985; Taka-hashi et al., 1985). Cocultures of hyppocampal neu-rons and OPCs untreated or transfected were set upand then assayed for MBP biosynthesis by Westernblotting after several periods of time. All lanes wereloaded with the same amount of protein (25 �g) totake into consideration the substantial increase incell number promoted in transfected cultures. Asshown in Figure 8, lane E, OPCNT-3 cells were muchmore efficient synthesizing MBP than untreatedOPC (lane D). According to densitometry scans,transfected cells were 10 times better as producers ofMBP than untreated cells. As positive control, weused a rat brain extract (lane C), and cultures ofOPCs alone (lane A) or hippocampal neurons alone(lane B) served as negative controls. One-week-oldcocultures synthesized only negligible levels of MBP(Fig. 8, lane 1); maximal production occurred after 2weeks (Fig. 8, lane 2). At 3 weeks, when hyppocam-pal neurons start to die, cultures showed no furtherincrease (Fig. 8, lane 3).
Microscopic Detection of Myelination inCocultures
Using phase-contrast and electron microscopy, weobserved that transfected OPCs were consistentlyable to myelinate axons of hippocampal neurons inculture. Individual oligodendrocytic cells alignedalong and simultaneously enveloping axons or bun-dles of axons could be easily identified by phase-contrast microscopy in cocultures containing trans-fected cells (Fig. 9A).
Electron microscopy confirmed that the increasedMBP synthesis detected by Western blotting wasindeed indicative of myelination. Mature, highlywrapped, compact myelin membranes (composed of amean of 10 � 2 layers) could already be seen in3-week-old cocultures of neurons with OPCNT-3 (Fig.9C). Cocultures with nontransfected OPCs also pro-duced myelin membranes but these membranes hadless layers (mean 4 � 1) that were less compacted(Fig. 9B). Although a precise quantitation of myelinprofiles is hard to carry out, their abundance wasclearly increased in cultures containing OPCNT-3 (amean of 13 � 3 per field) when compared with cul-tures containing OPCs (mean 5 � 2). No myelinmembranes were observed in cultures of hippocam-pal neurons alone.
DISCUSSION
Using our transfection protocol, transfected cellswere able to express NT-3 mRNA, as demonstratedby RT-PCR analysis and Northern blots (Figs. 1 and2). This mRNA was transcribed into the NT-3 proteinproduct, which was detected in culture supernatantsby ELISA. Kinetic studies revealed that 24 – 48 hposttransfection is the optimal time for neurotrophinproduction and secretion (Fig. 3). One of the effects oftransfection is inducing the cells to divide, as dem-
Fig. 7. Phase-contrast microscopy of oligodendrocyte precursor cells. After 3-day expansion period inthe presence of FGF and PDGF, the cultures were washed, mock-transfected (A), or transfected with thepNT-3 plasmid (B and C). In A and B, cells were observed 24 h later; in C, 48 h later.
84 RUBIO ET AL.
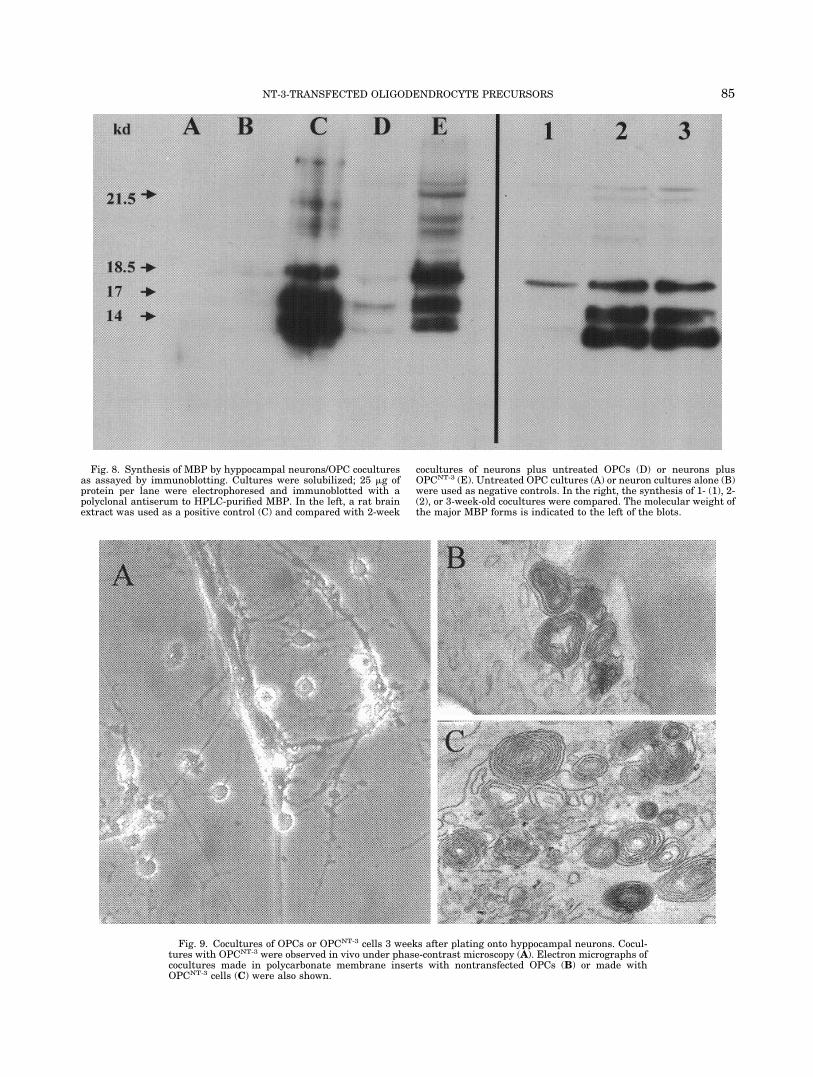
Fig. 8. Synthesis of MBP by hyppocampal neurons/OPC coculturesas assayed by immunoblotting. Cultures were solubilized; 25 �g ofprotein per lane were electrophoresed and immunoblotted with apolyclonal antiserum to HPLC-purified MBP. In the left, a rat brainextract was used as a positive control (C) and compared with 2-week
cocultures of neurons plus untreated OPCs (D) or neurons plusOPCNT-3 (E). Untreated OPC cultures (A) or neuron cultures alone (B)were used as negative controls. In the right, the synthesis of 1- (1), 2-(2), or 3-week-old cocultures were compared. The molecular weight ofthe major MBP forms is indicated to the left of the blots.
Fig. 9. Cocultures of OPCs or OPCNT-3 cells 3 weeks after plating onto hyppocampal neurons. Cocul-tures with OPCNT-3 were observed in vivo under phase-contrast microscopy (A). Electron micrographs ofcocultures made in polycarbonate membrane inserts with nontransfected OPCs (B) or made withOPCNT-3 cells (C) were also shown.
85NT-3-TRANSFECTED OLIGODENDROCYTE PRECURSORS

onstrated by enhanced 3H-thymidine incorporationinto DNA (Fig. 4). This increase in thymidine incor-poration was also detected over that produced by thejoint presence of the known mitogens FGF and PDGF(Fig. 5). In the absence of mitogens, cell survival wasclearly improved by transfection, as detected by theMTT test when compared to untreated cell cultures(Fig. 6). The morphology of the transfected culturesalso changes from their bipolar precursors tobrighter dividing cells that become GalC-positive dif-ferentiated oligodendrocytes after 48 h of culture(Fig. 7). Therefore, transfection induces in OPCs lim-ited proliferation accompanied of enhanced differen-tiation. We then evaluated if cocultures of hippo-campal neurons with OPCNT-3 cells could producemore myelin than untreated OPC. MBP is the majorstructural protein of myelin and constitutes as muchas 30% of its total protein. Western blots of thecocultures containing OPCNT-3 cells showed 10-foldincrease in MBP synthesis when compared to cocul-tures with nontransfected oligodendrocyte precur-sors (Fig. 8, lanes E and D). It was shown by phase-contrast microscopy that OPCNT-3 cells becameattached to, and aligned along, the axons in cocul-tures (Fig. 9A). When the cocultures were processedfor electron microscopy, it was possible to observecomplete myelin-like membranes. After 2 weeks ofcoculture, these membranes were composed of sev-eral layers (Fig. 9C). Untreated OPC also producemyelin-like membranes, as previously reported(Rome et al., 1986), but these membranes were com-posed of less layers that were loosely compacted(Fig. 9B).
There are many recent reports of experimentaltherapeutic interventions in rodents aimed at pro-moting regeneration of demyelinated corticospinaltracts. Grafting of embryonic stem cells (Liu et al.,2000), adult neurospheres (Pluchino et al., 2003),fibroblast engineered to produce NT-3 (Grill et al.,1997; McTigue et al., 1998), OPCs (Groves et al.,1993), or cografting of oligodendrocyte precursorswith PDGF-secreting neuroblastoma cells (Milwardet al., 2000) has been reported to induce partialrecovery after rat spinal cord injury. The potentialclinical use of neural cell transplants will depend onthe ability of transplanted cells to create areas ofrepair in demyelinated zones. The grafting of ourOPCNT-3 could combine two conventional regenera-tion strategies: cell replacement and neurotrophicfactor delivery (Hormer and Gage, 2000; Kondo andRaff, 2000). The recovery of sick animals in the EAEexperimental model, involving the intracerebroven-tricular injection of cultured adult neural stem cell,has recently been described (Pluchino et al., 2003) .We are transplanting OPCNT-3 to promote functionalrecovery in the TMEV-induced ME animal model.Positive results could provide new hope for the futuretreatment of this human disease.
ACKNOWLEDGMENTS
The technical assistance of Concepcion Torres in thetissue culture is gratefully acknowledged.
REFERENCES
Barres BA, Hart IK, Coles HSR, Burne JF, Voyvodic JT, RichardsonWD, Raff MC. 1992. Cell death and control of cell survival in theoligodendrocyte lineage. Cell 70:31–46.
Barres BA, Raff MC, Gaese F, Bartke I, Dechant G, Barde Y-A. 1994.A crucial role for neurotrophin-3 in oligodendrocyte development.Nature 367:371–375.
Bogler O, Wren D, Barnett SC, Land H, Noble M. 1990. Cooperationbetween two growth factors promotes extended self-renewal andinhibits differentiation of oligodendrocyte-type-2 astrocyte (0-2A)progenitor cells. Proc Natl Acad Sci USA 87:6368–6372.
Cohen RI, Marmur R, Norton W, Mehler MF, Kessler JA. 1996. Nervegrowth factor and neurotrophin-3 differentially regulate the prolif-eration and survival of developing rat brain oligodendrocytes.J Neurosci 16:6433–6442.
De Ferra F, Engh H, Hundson L, Kamholz J, Puckett C, Molineaux S,Lazzarini RA. 1985. Alternative splicing accounts for the four formsof myelin basic protein. Cell 43:1287–1301.
Dotti CG, Sullivan CA, Banker GA. 1988. The establishment ofpolarity by hippocampal neurons in culture. J Neurosci 8:1454 –1468.
Grill R, Murai K, Blesch A, Gage FH, Tuszynski MH. 1997. Cellulardelivery of neurotrophin-3 promotes corticospinal axonal growthand partial functional recovery after spinal cord injury. J Neurosci17:5560–5572.
Groves AK, Barnett SC, Franklin RJ, Crang AJ, Mayer M, BlakemoreWF, Noble M. 1993. Repair of demyelinate lesions by transplanta-tion of O-2A progenitor cells. Nature 362:453–455.
Haggiag S, Zhang P-L, Slutzky G, Shinder V, Kumar A, Chebath J,Revel M. 2001. Stimulation of myelin gene expression in vitro andof sciatic nerve remyelination by interleukin-6 receptor-interleu-kin-6 chimera. J Neurosci Res 64:564–574.
Hamilton SP, Rome LH. 1994. Stimulation of in vitro myelin synthe-sis by microglia. Glia 11:326–335.
Heinrich M, Gorath M, Richter-Landsberg C. 1999. Neurotrophin-3(NT-3) modulates early differentiation of oligodendrocytes in ratbrain cortical cultures. Glia 28:244–255.
Hormer PJ, Gage FH. 2000. Regenerating the damaged central ner-vous system. Nature 407:963–970.
Jones KR, Reichardt LF. 1990. Molecular cloning of a human genethat is a member of the nerve growth factor family. Proc Natl AcadSci USA 87:8060–8064.
Kondo T, Raff M. 2000. Oligodendrocyte precursor cells repro-grammed to become multipotential CNS stem cells. Science 289:1754–1757.
Laemmli UK. 1970. Cleavage of structural proteins during the assem-bly of the head of bacteriophage T4. Nature 227:680–685.
Liu S, Qu Y, Steward TJ, Howard MJ, Chakrabortty S, Holekamp TF,McDonald JW. 2000. Embryonic stem cells differentiate into oligo-dendrocytes and myelinate in culture after spinal cord transplan-tation. Proc Natl Acad Sci USA 97:6126–6131.
McCarthy KD, de Vellis J. 1980. Preparation of separate astroglialand oligodendroglial cell cultures fron rat cerebral tissue. J CellBiol 85:890–902.
McTigue DM, Horner PH, Stokes BT, Gage FH. 1998. Neurotrophin-3and brain-derived neurotrophic factor induce oligodendrocyte pro-liferation and myelination of regenerating axons in the contusedadult rat spinal cord. J Neurosci 18:5354–5365.
Miller RH. 1996. Oligodendrocyte origins. Trends Neurosci 19:92–96.
Milward EA, Zhang S-C, Zhao M, Lundberg C, Ge B, Goetz BD,Duncan ID. 2000. Enhanced proliferation and directed migration ofoligodendroglial progenitors co-grafted with growth factor-secretingcells. Glia 32:264–270.
Molina-Holgado E, Vela JM, Arevalo-Martın A, Guaza C. 2001. LPS/IFN- cytotoxicity in oligodendroglial cells: role of nitric oxide andprotection by the anti-inflammatory cytokine IL-10. Eur J Neurosci13:493–502.
Mosmann T. 1983. Rapid colorimetric assay for cellular growth andsurvival: application to proliferation and cytotoxicity. J ImmunolMeth 65:55–63.
86 RUBIO ET AL.

Noble M, Murray K, Stroobant P, Waterfield MD, Riddle P. 1988.Platelet-derived growth factor promotes division and motility andinhibits premature differentiation of the oligodendrocyte/type-2 as-trocyte progenitor cell. Nature 333:560–562.
Oshiro Y, Eylar EH. 1970. Allegic encephalitogenic basic protein frombovine brain. Archs Biochem Biophys 138:392–396.
Pfeiffer SE, Warrington AE, Bansal R. 1993. The oligodendrocyte andits many cellular processes. Trends Cell Biol 3:191–197.
Pluchino S, Quattrini A, Brambilla E, Gritti A, Salani G, Dina G, GalliR, Del Carro U, Amadio ST, Bergami A, Furlan R, Comi G, VescoviAL, Martino G. 2003. Injection of adult neurospheres induces re-covery in a chronic model of multiple sclerosis. Nature 422:688–694.
Rome LH, Bullock PN, Chiappelli F, Cardwell M, Adinolfi AM, Swan-son D. 1986. Synthesis of a myelin-like membrane by oligodendro-cytes in culture. J Neurosci Res 15:49–65.
Rubio N, Cuesta A. 1989. Lack of cross-reaction between myelin basicproteins and putative demyelinating virus envelope proteins. MolImmunol 26:663–668.
Rubio N, De Felipe C, Torres C. 1990. Theiler’s murine encephalomy-elitis virus-binding activity on neural and non-neural cell lines andtissues. J Gen Virol 71:2867–2872.
Takahashi N, Roach A, Teplow DB, Prusiner SB, Hood L. 1985.Cloning and characterization of the myelin basic protein gene frommouse: one gene can encode both 14 kd and 18.5 kd MBPs byalternate use of exons. Cell 42:139–148.
87NT-3-TRANSFECTED OLIGODENDROCYTE PRECURSORS
Copyright © 2022 FDOKUMEN




















