
Optical Coherence Tomography Characteristics of Choroidal Metastasis
Upload
independentCategory
view
0download
0
Inhibition of tumor growth and metastasis of human melanoma byintracellular anti-ATF-1 single chain Fv fragment
Didier Jean1, Carmen Tellez1, Suyun Huang1, Darren W Davis1, Christiane J Bruns1,David J McConkey1, Steven H Hinrichs2 and Menashe Bar-Eli*,1
1Department of Cancer Biology, The University of Texas M.D. Anderson Cancer Center, Houston, Texas, TX 77030, USA;2Department of Pathology and Microbiology, University of Nebraska Medical Center, Omaha, Nebraska, NE 68198, USA
Activating transcription factor-1 (ATF-1) and cAMP-responsive element (CRE)-binding protein (CREB) havebeen implicated in cAMP and Ca2+-induced transcrip-tional activation. The expression of the transcriptionfactors CREB and ATF-1 is upregulated in metastaticmelanoma cells. However, how overexpression of ATF-1/CREB contributes to the acquisition of the metastaticphenotype remains unclear. Here, the e�ect of disruptingATF-1 activity was investigated using intracellularexpression of an inhibitory anti-ATF-1 single chainantibody fragment (ScFv). Intracellular expression ofScFv anti-ATF-1 in MeWo melanoma cells causedsigni®cant reduction in CRE-dependent promoter activa-tion. In addition, expression of ScFv anti-ATF-1 inmelanoma cells suppressed their tumorigenicity andmetastatic potential in nude mice. ScFv anti-ATF-1rendered the melanoma cells susceptible to thapsigargin-induced apoptosis in vitro and caused massive apoptosisin tumors transplanted subcutaneously into nude mice,suggesting that ATF-1 and its associated proteins act assurvival factor for human melanoma cells. This is the®rst report to demonstrate the potential of ScFv anti-ATF-1 as an inhibitor of tumor growth and metastasis ofsolid tumor in vivo. Oncogene (2000) 19, 2721 ± 2730.
Keywords: melanoma; metastasis; apoptosis; transcrip-tional regulation
Introduction
The molecular basis of human malignant melanomaprogression has remained largely unknown, despite thefact that the worldwide incidence of melanoma isincreasing more than any other neoplastic disease (Kopfet al., 1995). The development of malignant melanoma inhumans progresses through a multistage process. Theswitches from melanocyte to nevi, to radial growth phase(RGP), and subsequently to vertical growth phase (VGP,metastatic phenotype) are associated with decreaseddependence on growth factors, diminished anchoragedependence, and reduced contact inhibition (Fidler,1990; Lu and Kerbel, 1994).
A large body of data concerning the molecularcontrol of melanoma progression has come fromstudies using mitogens. In culture, melanocytes syner-gistically respond to a number of growth factors, whichin contribution with each other or with 12-0-tetra-
decanoylphorbol-13-acetate or cAMP stimulate notonly proliferation but also pigmentation (Halaban,1993). These growth factors include several ®broblastgrowth factors, hepatocyte growth factor, and stem cellfactor (also known as KIT ligand, MGF and steelfactor), all of which stimulate tyrosine kinase receptors.As melanocyte proliferation and di�erentiation arepositively regulated by agents that increase cAMP(Halaban et al., 1983, 1984, 1987; Huang et al., 1996),we have focused on the transcription factors ATF-1(Activating transcription factor 1) and CREB (cAMP-responsive element-binding protein), which are knownto be activated by cAMP, as possible mediators oftumor growth and metastasis of human melanoma.
ATF-1 and CREB are members of the large bZIPsuperfamily of transcription factors. Members of theCREB/ATF family bind to cAMP-responsive elements(CREs) within the promoter and enhancer sequences ofmany genes. ATF-1, CREB, and the cAMP-responsiveelement modulator protein (CREM) constitute theCREB/ATF subfamily within the bZIP superfamily,whose members are de®ned by their ability toheterodimerize with each other but not with membersof other subfamilies (Meyer and Habener, 1993). ATF-1, CREB, and CREM have similar structures and arehighly homologous at the amino acid sequence level,especially within the bZIP region. Despite thesesimilarities, members of the CREB multigene subfamilyhave distinct biological activities.
ATF-1, CREB, and CREM may act as eitherpositive or negative regulators of transcription. Alter-native mRNA splicing produces numerous isoforms ofATF-1 and CREB, that can account for the variabilityin transcriptional regulatory activities (Lee andMasson, 1993; Lemaigre et al., 1993). In addition,each protein or isoform also possesses di�eringpatterns of phosphorylation, and the speci®c patternscontribute to their ability as regulators of transcription(Meyer and Habener, 1993). CREB and CREM havebeen shown to play important roles in basal andhormone-regulated transcription and di�erentiationwhile the role of ATF-1 is less well de®ned. ATF-1homodimers appear to be weaker transcriptionalactivators than either CREB or certain forms ofCREM, since ATF-1-mediated activation is enhancedby heterodimerization with either CREB or CREM(Lee and Masson, 1993). In addition, it has beendemonstrated that CREB can e�ciently form hetero-dimers with ATF-1 rather than from the CREBhomodimer (Hurst et al., 1991; Rehfuss et al., 1991).
Previous studies have demonstrated that CREBexpression correlates directly with the metastaticpotential of murine melanoma cells (Rutberg et al.,
Oncogene (2000) 19, 2721 ± 2730ã 2000 Macmillan Publishers Ltd All rights reserved 0950 ± 9232/00 $15.00
www.nature.com/onc
*Correspondence: M Bar-EliReceived 3 January 2000; revised 29 February 2000; accepted 9March 2000

1994) and that ATF-1 is not detected in normalmelanocytes but is easily found in metastatic melano-ma cells (Bohm et al., 1995). Whether these observa-tions are causally related to tumorigenicity andmetastasis of melanoma cells was not clear. To studythe contribution of CREB and its associated proteinsto tumor growth and metastasis of human melanomacells, we have previously taken the approach of using adominant-negative form of CREB, KCREB, that hadbeen mutated in the DNA-binding domain (Walton etal., 1992). Expression of KCREB in melanoma cellsdecreased their tumorigenicity and metastatic potentialin nude mice (Xie et al., 1997b). Moreover, in recentstudies, we have demonstrated that CREB and itsassociated proteins act as survival factors for humanmelanoma cells, since quenching their activities byKCREB rendered the melanoma cells susceptible toapoptosis by agents that increase intracellular Ca2+
levels (Jean et al., 1998b). Collectively, these resultsindicate that CREB and its associated proteins (mostlikely ATF-1) play pivotal roles in the development ofmalignant melanoma.
Here, we focused our investigation on the role ofATF-1 on tumor growth and metastasis of humanmelanoma. To further investigate the cellular role ofATF-1 in melanoma progression, we hereby utilizedcurrent advances in the engineering of antibodies thathave made possible the cloning of small single-chain Fv(ScFv) fragments. ScFv fragments contain the antigen-binding variable domains of the light and heavy chainsconnected by a peptide spacer (Raag and Whitlow,1995; Winter and Milstein, 1991). When constructed inthis manner, a single RNA transcript can be expressedand translated into an active protein that has thepotential to interfere with the activity of targetedintracellular proteins. Intracellular ScFv fragmentshave been successfully employed to decrease theexpression of ErbB-2 (Grim et al., 1996; Graus-Portaet al., 1995), the a subunit of human IL-2 receptor(Richardson et al., 1995), and to restore the transcrip-tion activity of mutant p53 (de Fromentel et al., 1999).
In studies aimed at addressing the role of ATF-1 intumor growth and metastasis of human melanoma, asingle chain Fv fragment (ScFv) was derived from amonoclonal antibody (mAb) anti-ATF-1. This mAb(designated mAb 41.4) has been shown to inhibit ATF-1 binding and transcriptional activation from CRE-containing promoters in vitro (Orten et al., 1994). Inthis study we provide evidence that expression of ScFvanti-ATF-1 in MeWo melanoma cells inhibited theirtumorigenicity and metastatic potential in nude mice.Intracellular expression of ScFv anti-ATF-1 renderedthe melanoma cells susceptible to apoptosis in vivocompared to control untransfected cells. These studiesdemonstrate for the ®rst time that intracellular ScFvanti-ATF-1 can be used to quench ATF-1 activity notonly as a method to explore its function but also as amodality for cancer therapy.
Results
Expression of ScFv anti-ATF-1 in melanoma cells
MeWo melanoma cells were transfected with ScFvanti-ATF-1 expression vector. Expression of ScFv anti-
ATF-1 in MeWo melanoma cells was analysed byRT ±PCR utilizing speci®c primers corresponding tothe light and heavy chains of the ScFv. Figure 1 showsthat following transfection, we were able to isolate onepooled MeWo cells (designated MeWo-ScFv-P, lane 5)and 3 clones (designated as MeWo-ScFv-C4, C5, andC6, lanes 7, 8 and 9, respectively). No expression ofScFv anti ATF-1 was observed in control parental cells(lane 3) or in empty vector Neo-transfected cells (lanes4 and 6). GAPDH was ampli®ed simultaneously in thesame tube to verify the integrity of the DNA and equalloadings. Clone MeWo-ScFv-C4 and the pooled cellline MeWo-ScFv-P were selected for further analysesto study the e�ect of ScFv anti-ATF-1 on tumorgrowth and metastasis of the transfected cells.
Effect of intracellular ScFv anti-ATF-1 on CRE-dependent promoter transactivation
To determine whether the ScFv anti-ATF-1 in thetransfected cells was functional and capable ofquenching the ATF-1 transcriptional activity, weanalysed its ability to inhibit a CRE-dependentpromoter. To that end, we used a CRE-dependentplasmid (Somat-BglII CAT), that contains the soma-tostatin gene promoter from 771 to +53 linked to theCAT reporter gene (Montminy et al., 1986), as wepreviously described (Jean et al., 1998b). We analysedthe CAT activity driven by the Somat-BglII in MeWo-ScFv-P and in MeWo-ScFv-C4 in comparison toparental and control Neo-transfected cells (Figure 2).In the following transfection experiments, cell extractswere prepared and equivalent amounts of extractsexhibiting the same Renilla luciferase activity were
Figure 1 Expression of ScFv anti-ATF-1 by RT±PCR.GAPDH was ampli®ed simultaneously in each tube for DNAintegrity and equal loading. ScFv anti-ATF-1 expression wasobserved in one pooled cell line MeWo-ScFv-P (lane 5), and 3clones (C4, C5, and C6, lanes 7 ± 9). MeWo-ScFv-P and MeWo-ScFv-C4 were selected for further in vitro and in vivo studies.Lane 1, control naked expression vector. Lane 2, control plasmidcontaining the ScFv anti-ATF-1 DNA sequences. No expressionof ScFv anti-ATF-1 was observed in parental (lane 3) or controlNeo-transfected cells (lanes 4 and 6)
Regulation of melanoma growth and metastasis by ScFv anti-ATF-1D Jean et al
2722
Oncogene

tested for CAT activity. Figure 2 demonstrates thatCAT activity driven by Somat-BglII promoter wasinhibited by 10-fold in MeWo-ScFv-P (lane 5) ascompared with control pooled Neo-transfected cells(lane 3). CAT activity in ScFv-C4 was inhibited by 35-fold (lane 6) as compared with the Neo-transfectedclone (lane 4). The inhibition of CAT activity in ScFv-C4 was comparable to the inhibition observed in cellstransfected with the dominant-negative construct ofCREB, KCREB (35-fold, lane 7).
To make sure that the inhibition of ScFv anti-ATF-1was speci®c for CRE-dependent promoters, we choseto work with a luciferase reporter gene expressionvector driven by three AP-2 consensus responseelements from the human metallothionein gene IIAligated in front of a minimal TK promoter (3xAP-2Luc), that we described previously (Huang et al.,1998). There are no CRE's or CRE-like elementspresent, allowing for demonstration of speci®city of theScFv anti-ATF-1. Luciferase activity was not inhibitedin the ScFv anti-ATF-1 melanoma-transfected cells ascompared with parental and control Neo-transfectedcells (Figure 3). Collectively these data indicate thatexpression of ScFv anti-ATF-1 in MeWo melanomacells signi®cantly and speci®cally inhibited the CRE-driven promoter.
Effect of intracellular ScFv anti-ATF-1 on tumor growthand metastasis of human melanoma cells
To determine the tumorigenicity of the ScFv anti-ATF-1 transfected cells, we injected 16106 cells s.c. into
BALB/c nude mice and tumor growth was monitoredonce a week for a period of 55 days. Both MeWoparental and Neo-transfected control cells (MeWo-Neo-C) grew progressively in all mice injected andreached 1.1 ± 1.3 cm in mean diameter within 55 days(Figure 4). In contrast, MeWo cells expressingintracellular ScFv anti-ATF-1, both the pooled cellline (MeWo-ScFv-P) and clone MeWo-ScFv-C4 pro-duced smaller tumors (0.45 ± 0.5 cm in mean diameter).Growth of ScFv anti-ATF-1 transfected cells di�eredsigni®cantly from parental and Neo-control cells at 40,45, 50 and 55 days after injections (P50.01). Thesedata indicate that intracellular ScFv anti-ATF-1 inMeWo cells suppressed their tumor growth in vivo.
In the next set of experiments, the metastaticpotential of ScFv-transfected MeWo cells was deter-mined in an experimental lung metastasis assay(Radinsky et al., 1994; Luca et al., 1997). To thatend, BALB/c nude mice were injected i.v. with 16106
MeWo-ScFv-transfected, parental, or Neo-control cellsand 60 days later the number of lung metastases wascounted. As shown in Table 1, parental and Neo-control cells produced lung tumor colonies in allinjected mice with a median of 4200. In contrast,
Figure 2 Intracellular expression of ScFv anti-ATF-1 inhibitsCRE-dependent promoter activation. E�ect of ScFv anti-ATF-1on CAT activity driven by a CRE-dependent promoter (Somat-BglII), was analysed in MeWo-P, Neo- or ScFv-transfected cells.CAT activity was inhibited by 10-fold and 35-fold in MeWo-ScFv-P and in MeWo-ScFv-C4, respectively (lanes 5 and 6) whencompared to parental and Neo-transfected cells (lanes 2 ± 4). Theinhibition of CAT activity in ScFv-C4 was comparable to thatobserved in clone transfected with the dominant-negativeconstruct of CREB, KCREB (35-fold, lane 7). Lane 1, Basic;control plasmid without the CRE-dependent promoter. -Foldinhibition is calculated in comparison with untransfected cells(lanes 2 ± 4), which were given the reference value of 1. Figure isrepresentative of three experiments performed
Figure 3 E�ect of intracellular ScFv anti-ATF-1 on non-CRE-dependent promoter. No change in luciferase activity driven bythe 3xAP-2 Luc promoter was observed in the ScFv-transfectedcells as compared with control cells
Figure 4 Tumorigenicity of MeWo parental, neo-transfected andScFv-transfected cells, 55 days after s.c. injection into nude mice.Tumor cells (16106 in 0.2 ml of HBSS) were injected over theright scapular region. Growth of s.c. tumors was monitored byweekly examination and measurement of tumors with calipers.The data represent the mean diameter observed in ®ve animalsper group
Oncogene
Regulation of melanoma growth and metastasis by ScFv anti-ATF-1D Jean et al
2723

cells expressing intracellular ScFv anti-ATF-1 pro-duced fewer lung metastases with a median of 73 and35 for MeWo-ScFv-P and MeWo-ScFv-C4, respec-tively.
To determine whether the inhibition of tumorgrowth and the decreased capacity to produce lungmetastases by ScFv anti-ATF-1 in vivo was due todi�erent growth rates in vitro, we compared theirdoubling time with control cells in culture. Nodi�erence in doubling time was observed in culturebetween the transfected and control cells (data notshown).
Intracellular ScFv anti-ATF-1 renders melanoma cellssusceptible to apoptosis induction by thapsigargin
Since ATF-1 mediates both cAMP and Ca2+ transcrip-tional responses (Gonzales and Montiminy, 1989;Sheng et al., 1991) and since the induction of apoptosisby diverse exogenous signals is dependent on elevationof intracellular Ca2+ (McConkey and Orrenius, 1996),we decided to analyse if ATF-1 (and its associatedproteins) could be involved in the resistance ofmelanoma cells to apoptosis induction, hence con-tributing to tumor growth and metastasis. To study theinvolvement of ATF-1 and its associated proteins inapoptosis, we used thapsigargin (Tg), which inhibitsendoplasmic reticulum-dependent Ca2+-ATPase andthereby increases cytosolic Ca2+ (Thastrup et al.,1990). To that end, we next evaluated the ability ofTg to induce apoptosis in MeWo parental cells(MeWo-P), Neo-transfected cells (MeWo-Neo-P,MeWo-Neo-C), and ScFv anti-ATF-1-transfected clone(MeWo-ScFv-C4). As a control, we also analysed aKCREB-transfected clone (MeWo-KCREB-C8).
Apoptosis was determined by propidium iodidestaining and FACS analysis. Figure 5A shows theresults of a representative experiment after 48 htreatment with 1 mM Tg. The cells treated with Tgshowed a hypodiploid DNA content indicative ofapoptosis. The percentages of hypodiploid cells werehigher in the cells expressing intracellular ScFv anti-ATF-1 (74.5 vs 17.9, 18.3 and 19.1% in parental andNeo-transfected cells). Apoptosis was also observed inthe KCREB-transfected clone, MeWo-KCREB-C8, aswe previously described (Jean et al., 1998b). Agentsthat stimulate adenylate cyclase or cAMP analogs(such as forskolin and dibutyryl-cAMP) did not induceapoptosis in the melanoma cells in our assay (data notshown), indicating that Tg-induced apoptosis wasmediated via the Ca2+ and not the cAMP pathway.
Figure 5B is a summary of at least three independentFACS analyses of propidium iodide staining.
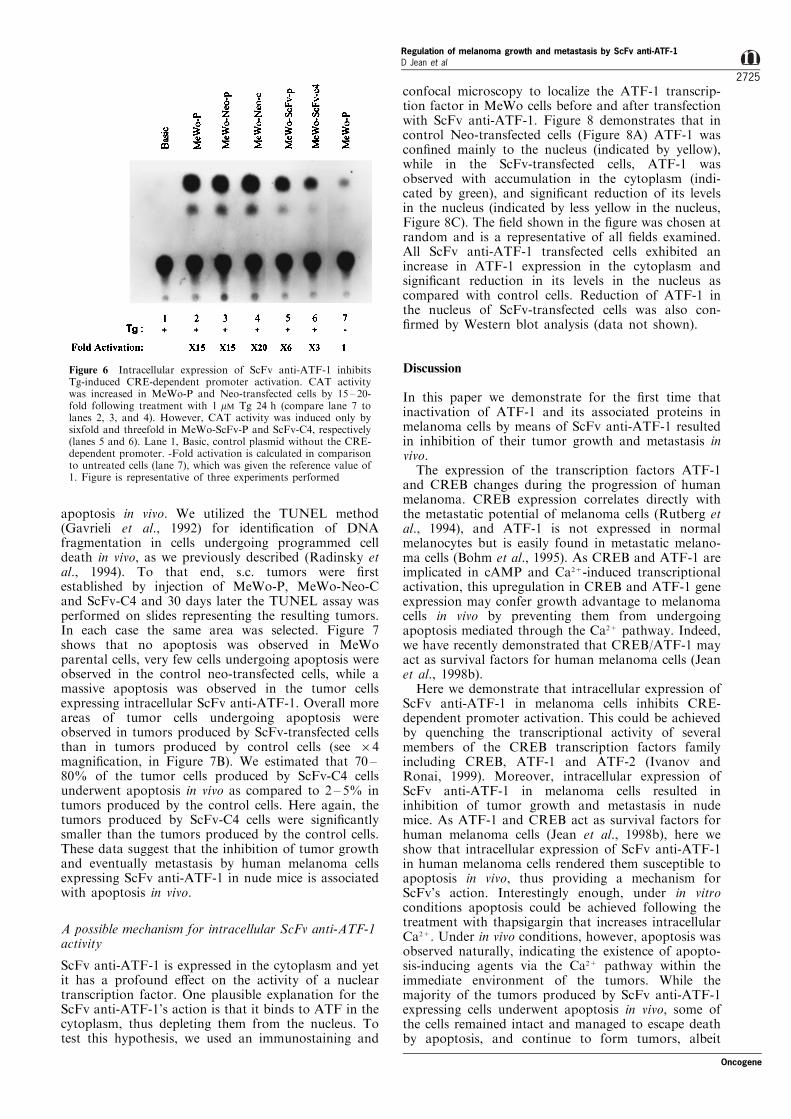
We have previously demonstrated that treatment ofmelanoma cells with thapsigargin (Tg) caused phos-phorylation of CREB and ATF-1 and activated theirCRE-dependent transcription (Jean et al., 1998b). Toverify that ScFv anti-ATF-1 expression inhibited thetransactivation of a CRE-dependent promoter inducedby Tg, we next analysed the CAT activity in thetransfected cells after treatment with 1 mM Tg for 24 h.Figure 6 shows that CAT activity was increased by15 ± 20-fold in parental and Neo-transfected cellsfollowing treatment with Tg (lanes 2 ± 4) whencompared to untreated cells (lane 7). However, CATactivity driven by the Somat-BglII promoter wasinduced only by sixfold in MeWo-ScFv-P (lane 5)and by threefold in MeWo-ScFv-C4 (lane 6) incomparison to untreated cells (lane 7). These resultsshow that ScFv anti-ATF-1 was able to suppress thefold induction of CRE-CAT activity induced by Tg.
Intracellular ScFv anti-ATF-1 induces apoptosis in vivo
We next determined whether the inhibition of tumorgrowth and metastasis by melanoma cells expressingScFv anti-ATF-1 in nude mice was associated with
Table 1 Metastatic potential of MeWo human melanoma cellstransfected with ScFv anti-ATF-1
Experimental lungmetastasisb
Cell line Incidence Median Range Pb
MeWo-P 5/5 4200 All 4200MeWo-Neo-C 5/5 4200 1524200MeWo-ScFv-P 5/5 73 62 ± 125 50.05MeWo-ScFv-C4 5/5 35 23 ± 118 50.05
aTumor cells (16106 mouse) were injected i.v. into nude mice;experimental metastasis was determined 60 days after tumorinjection. bThe di�erences in metastatic potential between controlcells and transfected cells were analysed by the Mann±Whitney Utest
Figure 5 Intracellular ScFv anti-ATF-1 renders melanoma cellssusceptible to apoptosis by thapsigargin (Tg). Apoptosis wasassayed by propidium iodide staining and ¯ow cytometry. (A)example of a representative analysis by ¯ow cytometry of cellsstained with propidium iodide after 48 h treatment with Tg. (B) asummary of at least three independent FACS analyses ofpropidium iodide staining following treatment with 1 mM Tg for48 h. ScFv-transfected cells showed a higher rate of hypodiploidDNA contents indicative of apoptosis as compared with controlcells. Apoptosis was also observed in KCREB-transfected cells
Regulation of melanoma growth and metastasis by ScFv anti-ATF-1D Jean et al
2724
Oncogene
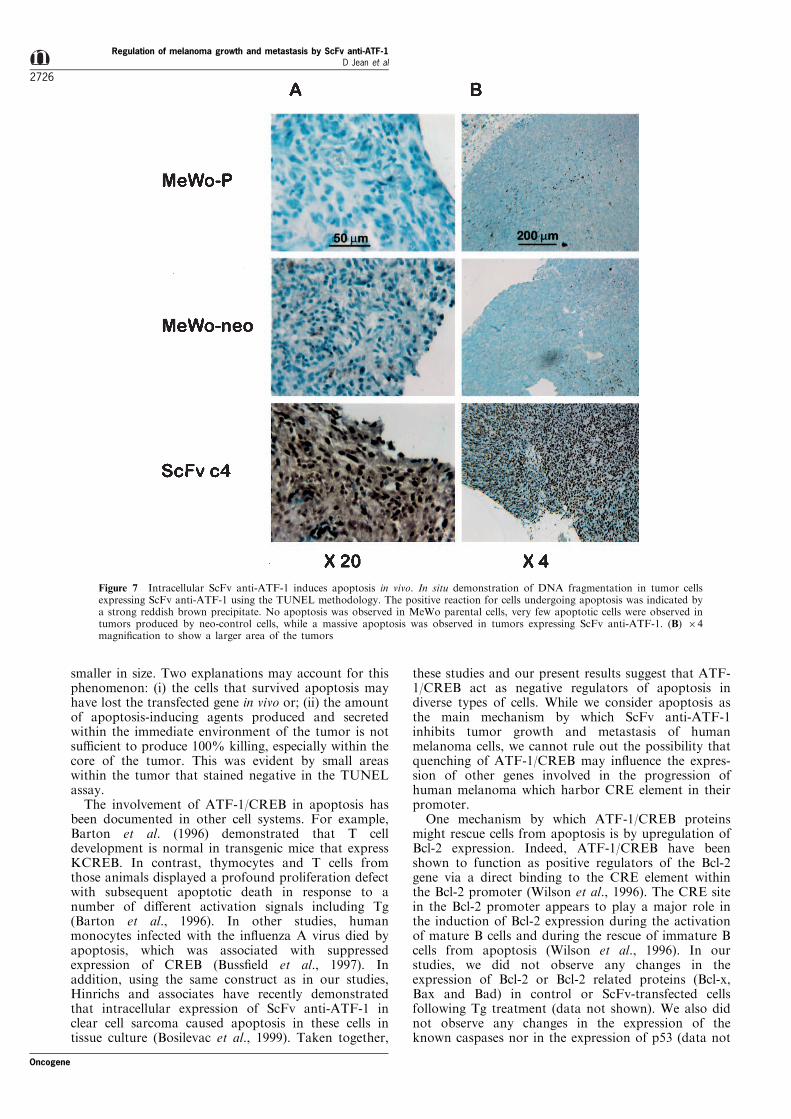
apoptosis in vivo. We utilized the TUNEL method(Gavrieli et al., 1992) for identi®cation of DNAfragmentation in cells undergoing programmed celldeath in vivo, as we previously described (Radinsky etal., 1994). To that end, s.c. tumors were ®rstestablished by injection of MeWo-P, MeWo-Neo-Cand ScFv-C4 and 30 days later the TUNEL assay wasperformed on slides representing the resulting tumors.In each case the same area was selected. Figure 7shows that no apoptosis was observed in MeWoparental cells, very few cells undergoing apoptosis wereobserved in the control neo-transfected cells, while amassive apoptosis was observed in the tumor cellsexpressing intracellular ScFv anti-ATF-1. Overall moreareas of tumor cells undergoing apoptosis wereobserved in tumors produced by ScFv-transfected cellsthan in tumors produced by control cells (see 64magni®cation, in Figure 7B). We estimated that 70 ±80% of the tumor cells produced by ScFv-C4 cellsunderwent apoptosis in vivo as compared to 2 ± 5% intumors produced by the control cells. Here again, thetumors produced by ScFv-C4 cells were signi®cantlysmaller than the tumors produced by the control cells.These data suggest that the inhibition of tumor growthand eventually metastasis by human melanoma cellsexpressing ScFv anti-ATF-1 in nude mice is associatedwith apoptosis in vivo.
A possible mechanism for intracellular ScFv anti-ATF-1activity
ScFv anti-ATF-1 is expressed in the cytoplasm and yetit has a profound e�ect on the activity of a nucleartranscription factor. One plausible explanation for theScFv anti-ATF-1's action is that it binds to ATF in thecytoplasm, thus depleting them from the nucleus. Totest this hypothesis, we used an immunostaining and
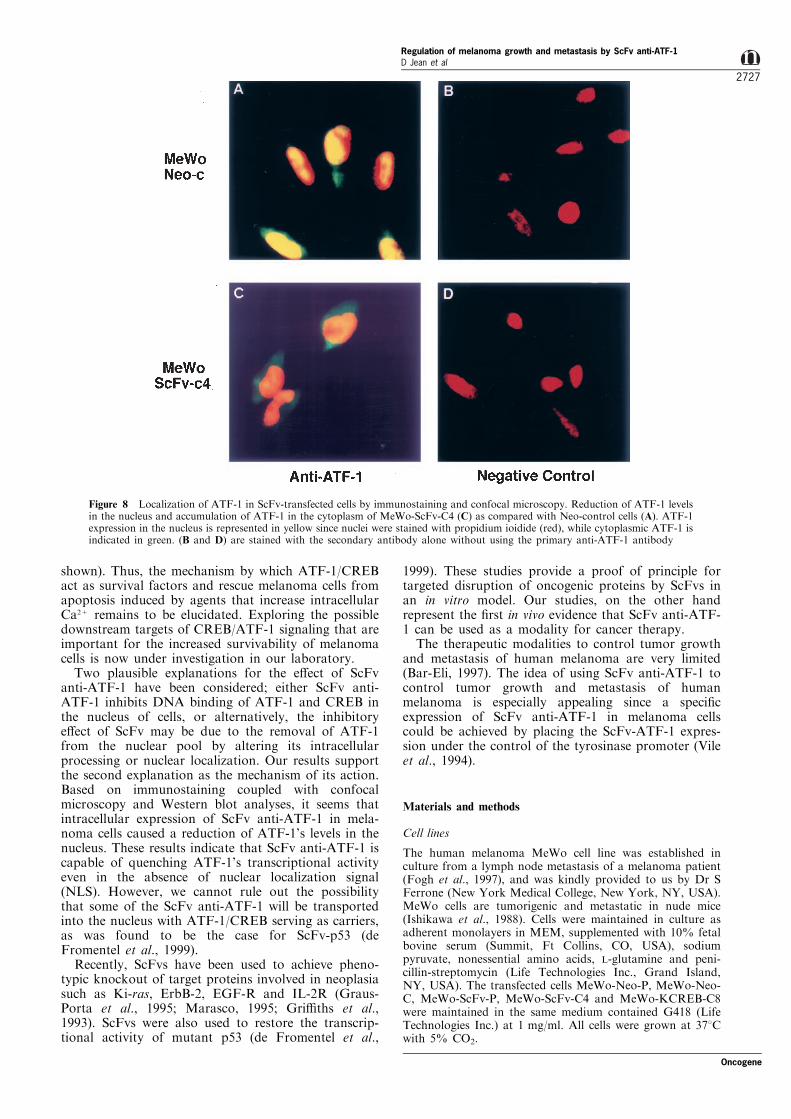
confocal microscopy to localize the ATF-1 transcrip-tion factor in MeWo cells before and after transfectionwith ScFv anti-ATF-1. Figure 8 demonstrates that incontrol Neo-transfected cells (Figure 8A) ATF-1 wascon®ned mainly to the nucleus (indicated by yellow),while in the ScFv-transfected cells, ATF-1 wasobserved with accumulation in the cytoplasm (indi-cated by green), and signi®cant reduction of its levelsin the nucleus (indicated by less yellow in the nucleus,Figure 8C). The ®eld shown in the ®gure was chosen atrandom and is a representative of all ®elds examined.All ScFv anti-ATF-1 transfected cells exhibited anincrease in ATF-1 expression in the cytoplasm andsigni®cant reduction in its levels in the nucleus ascompared with control cells. Reduction of ATF-1 inthe nucleus of ScFv-transfected cells was also con-®rmed by Western blot analysis (data not shown).
Discussion
In this paper we demonstrate for the ®rst time thatinactivation of ATF-1 and its associated proteins inmelanoma cells by means of ScFv anti-ATF-1 resultedin inhibition of their tumor growth and metastasis invivo.
The expression of the transcription factors ATF-1and CREB changes during the progression of humanmelanoma. CREB expression correlates directly withthe metastatic potential of melanoma cells (Rutberg etal., 1994), and ATF-1 is not expressed in normalmelanocytes but is easily found in metastatic melano-ma cells (Bohm et al., 1995). As CREB and ATF-1 areimplicated in cAMP and Ca2+-induced transcriptionalactivation, this upregulation in CREB and ATF-1 geneexpression may confer growth advantage to melanomacells in vivo by preventing them from undergoingapoptosis mediated through the Ca2+ pathway. Indeed,we have recently demonstrated that CREB/ATF-1 mayact as survival factors for human melanoma cells (Jeanet al., 1998b).
Here we demonstrate that intracellular expression ofScFv anti-ATF-1 in melanoma cells inhibits CRE-dependent promoter activation. This could be achievedby quenching the transcriptional activity of severalmembers of the CREB transcription factors familyincluding CREB, ATF-1 and ATF-2 (Ivanov andRonai, 1999). Moreover, intracellular expression ofScFv anti-ATF-1 in melanoma cells resulted ininhibition of tumor growth and metastasis in nudemice. As ATF-1 and CREB act as survival factors forhuman melanoma cells (Jean et al., 1998b), here weshow that intracellular expression of ScFv anti-ATF-1in human melanoma cells rendered them susceptible toapoptosis in vivo, thus providing a mechanism forScFv's action. Interestingly enough, under in vitroconditions apoptosis could be achieved following thetreatment with thapsigargin that increases intracellularCa2+. Under in vivo conditions, however, apoptosis wasobserved naturally, indicating the existence of apopto-sis-inducing agents via the Ca2+ pathway within theimmediate environment of the tumors. While themajority of the tumors produced by ScFv anti-ATF-1expressing cells underwent apoptosis in vivo, some ofthe cells remained intact and managed to escape deathby apoptosis, and continue to form tumors, albeit
Figure 6 Intracellular expression of ScFv anti-ATF-1 inhibitsTg-induced CRE-dependent promoter activation. CAT activitywas increased in MeWo-P and Neo-transfected cells by 15 ± 20-fold following treatment with 1 mM Tg 24 h (compare lane 7 tolanes 2, 3, and 4). However, CAT activity was induced only bysixfold and threefold in MeWo-ScFv-P and ScFv-C4, respectively(lanes 5 and 6). Lane 1, Basic, control plasmid without the CRE-dependent promoter. -Fold activation is calculated in comparisonto untreated cells (lane 7), which was given the reference value of1. Figure is representative of three experiments performed
Oncogene
Regulation of melanoma growth and metastasis by ScFv anti-ATF-1D Jean et al
2725
smaller in size. Two explanations may account for thisphenomenon: (i) the cells that survived apoptosis mayhave lost the transfected gene in vivo or; (ii) the amountof apoptosis-inducing agents produced and secretedwithin the immediate environment of the tumor is notsu�cient to produce 100% killing, especially within thecore of the tumor. This was evident by small areaswithin the tumor that stained negative in the TUNELassay.
The involvement of ATF-1/CREB in apoptosis hasbeen documented in other cell systems. For example,Barton et al. (1996) demonstrated that T celldevelopment is normal in transgenic mice that expressKCREB. In contrast, thymocytes and T cells fromthose animals displayed a profound proliferation defectwith subsequent apoptotic death in response to anumber of di�erent activation signals including Tg(Barton et al., 1996). In other studies, humanmonocytes infected with the in¯uenza A virus died byapoptosis, which was associated with suppressedexpression of CREB (Buss®eld et al., 1997). Inaddition, using the same construct as in our studies,Hinrichs and associates have recently demonstratedthat intracellular expression of ScFv anti-ATF-1 inclear cell sarcoma caused apoptosis in these cells intissue culture (Bosilevac et al., 1999). Taken together,
these studies and our present results suggest that ATF-1/CREB act as negative regulators of apoptosis indiverse types of cells. While we consider apoptosis asthe main mechanism by which ScFv anti-ATF-1inhibits tumor growth and metastasis of humanmelanoma cells, we cannot rule out the possibility thatquenching of ATF-1/CREB may in¯uence the expres-sion of other genes involved in the progression ofhuman melanoma which harbor CRE element in theirpromoter.
One mechanism by which ATF-1/CREB proteinsmight rescue cells from apoptosis is by upregulation ofBcl-2 expression. Indeed, ATF-1/CREB have beenshown to function as positive regulators of the Bcl-2gene via a direct binding to the CRE element withinthe Bcl-2 promoter (Wilson et al., 1996). The CRE sitein the Bcl-2 promoter appears to play a major role inthe induction of Bcl-2 expression during the activationof mature B cells and during the rescue of immature Bcells from apoptosis (Wilson et al., 1996). In ourstudies, we did not observe any changes in theexpression of Bcl-2 or Bcl-2 related proteins (Bcl-x,Bax and Bad) in control or ScFv-transfected cellsfollowing Tg treatment (data not shown). We also didnot observe any changes in the expression of theknown caspases nor in the expression of p53 (data not
Figure 7 Intracellular ScFv anti-ATF-1 induces apoptosis in vivo. In situ demonstration of DNA fragmentation in tumor cellsexpressing ScFv anti-ATF-1 using the TUNEL methodology. The positive reaction for cells undergoing apoptosis was indicated bya strong reddish brown precipitate. No apoptosis was observed in MeWo parental cells, very few apoptotic cells were observed intumors produced by neo-control cells, while a massive apoptosis was observed in tumors expressing ScFv anti-ATF-1. (B) 64magni®cation to show a larger area of the tumors
Regulation of melanoma growth and metastasis by ScFv anti-ATF-1D Jean et al
2726
Oncogene
shown). Thus, the mechanism by which ATF-1/CREBact as survival factors and rescue melanoma cells fromapoptosis induced by agents that increase intracellularCa2+ remains to be elucidated. Exploring the possibledownstream targets of CREB/ATF-1 signaling that areimportant for the increased survivability of melanomacells is now under investigation in our laboratory.
Two plausible explanations for the e�ect of ScFvanti-ATF-1 have been considered; either ScFv anti-ATF-1 inhibits DNA binding of ATF-1 and CREB inthe nucleus of cells, or alternatively, the inhibitorye�ect of ScFv may be due to the removal of ATF-1from the nuclear pool by altering its intracellularprocessing or nuclear localization. Our results supportthe second explanation as the mechanism of its action.Based on immunostaining coupled with confocalmicroscopy and Western blot analyses, it seems thatintracellular expression of ScFv anti-ATF-1 in mela-noma cells caused a reduction of ATF-1's levels in thenucleus. These results indicate that ScFv anti-ATF-1 iscapable of quenching ATF-1's transcriptional activityeven in the absence of nuclear localization signal(NLS). However, we cannot rule out the possibilitythat some of the ScFv anti-ATF-1 will be transportedinto the nucleus with ATF-1/CREB serving as carriers,as was found to be the case for ScFv-p53 (deFromentel et al., 1999).
Recently, ScFvs have been used to achieve pheno-typic knockout of target proteins involved in neoplasiasuch as Ki-ras, ErbB-2, EGF-R and IL-2R (Graus-Porta et al., 1995; Marasco, 1995; Gri�ths et al.,1993). ScFvs were also used to restore the transcrip-tional activity of mutant p53 (de Fromentel et al.,
1999). These studies provide a proof of principle fortargeted disruption of oncogenic proteins by ScFvs inan in vitro model. Our studies, on the other handrepresent the ®rst in vivo evidence that ScFv anti-ATF-1 can be used as a modality for cancer therapy.
The therapeutic modalities to control tumor growthand metastasis of human melanoma are very limited(Bar-Eli, 1997). The idea of using ScFv anti-ATF-1 tocontrol tumor growth and metastasis of humanmelanoma is especially appealing since a speci®cexpression of ScFv anti-ATF-1 in melanoma cellscould be achieved by placing the ScFv-ATF-1 expres-sion under the control of the tyrosinase promoter (Vileet al., 1994).
Materials and methods
Cell lines
The human melanoma MeWo cell line was established inculture from a lymph node metastasis of a melanoma patient(Fogh et al., 1997), and was kindly provided to us by Dr SFerrone (New York Medical College, New York, NY, USA).MeWo cells are tumorigenic and metastatic in nude mice(Ishikawa et al., 1988). Cells were maintained in culture asadherent monolayers in MEM, supplemented with 10% fetalbovine serum (Summit, Ft Collins, CO, USA), sodiumpyruvate, nonessential amino acids, L-glutamine and peni-cillin-streptomycin (Life Technologies Inc., Grand Island,NY, USA). The transfected cells MeWo-Neo-P, MeWo-Neo-C, MeWo-ScFv-P, MeWo-ScFv-C4 and MeWo-KCREB-C8were maintained in the same medium contained G418 (LifeTechnologies Inc.) at 1 mg/ml. All cells were grown at 378Cwith 5% CO2.
Figure 8 Localization of ATF-1 in ScFv-transfected cells by immunostaining and confocal microscopy. Reduction of ATF-1 levelsin the nucleus and accumulation of ATF-1 in the cytoplasm of MeWo-ScFv-C4 (C) as compared with Neo-control cells (A). ATF-1expression in the nucleus is represented in yellow since nuclei were stained with propidium ioidide (red), while cytoplasmic ATF-1 isindicated in green. (B and D) are stained with the secondary antibody alone without using the primary anti-ATF-1 antibody
Oncogene
Regulation of melanoma growth and metastasis by ScFv anti-ATF-1D Jean et al
2727

Animals and tumor cell injection
Male athymic nude mice (BALB/c background) wereobtained from the Animal Production Area of the NCI-Frederick Cancer Research and Development Center (Fre-derick, MD, USA). The mice were housed in laminar ¯owcabinets under speci®c pathogen-free conditions and used at 8weeks of age. Animals were maintained in facilities approvedby the American Association for Accreditation of LaboratoryAnimal Care and in accordance with current regulations andstandards of the US Department of Agriculture, Departmentof Health and Human Services, and the National Institutes ofHealth. Their use in these experiments was approved by theinstitutional Animal Care and Use Committee.
To prepare tumor cells for inoculation, cells in exponentialgrowth phase were harvested by brief exposure to 0.25%trypsin, 0.2% EDTA solution (w/v). The ¯ask was sharplytapped to dislodge the cells, and supplemented medium wasadded. The cell suspension was pipetted to produce a single-cell suspension. The cells were washed and resuspended inCa2+- and Mg2+-free HBSS to the desired cell concentration.Cell viability was determined by Trypan blue exclusion, andonly single-cell suspensions of more than 90% viability wereused. Subcutaneous tumors were produced by injecting16106 tumor cells in 0.2 ml of HBSS over the right scapularregion as we previously described (Jean et al., 1998a; Luca etal., 1995). Growth of subcutaneous tumors was monitoredfor 55 days. For experimental lung metastasis, 16106 tumorcells in 0.2 ml of HBSS were injected into the lateral tail veinof nude mice. The mice were killed after 60 days, and thelungs were removed, washed in water, and ®xed with Bouin'ssolution for 24 h to facilitate counting of tumor nodules aswe described previously (Radinsky et al., 1994; Singh et al.,1995; Luca et al., 1997). The number of surface tumornodules was counted under a dissecting microscope. Sectionsof the lungs were stained with hematoxylin and eosin tocon®rm that the nodules were melanoma and to identifymicrometastasis.
RNA preparation and RT±PCR
Total cellular RNA was prepared from 60 mm culture dishesof 70 ± 80% con¯uent cells using TRIzol reagent (LifeTechnologies Inc.) according to manufacturer's instructions.1 mg of total RNA was reverse-primed with an oligo poly-dTprimer and extended with MMLV reverse transcriptase(Clontech, Palo Alto, CA, USA). Using Advantage cDNAPCR kit (Clontech), cDNA was used for PCR ampli®cationwith primers speci®c of the ScFv-ATF-1 fragment (5'-GGAGTGGGTCGCATACATTA-3' and 5'-AAGATGAG-GAGTTTGGGTGG-3') and primers of GAPDH (5'-GAGC-CACATCGCTCAGAC-3') and 5'-CTTCTCATGGTTCAC-ACCC-3'). The PCR reaction was carried out in a PerkinElmer thermal cycler (Foster City, CA, USA). The reactionwas submitted to an initial denaturation step (2 min at 968C)followed by 27 cycles of denaturation (1 min 948C), annealing(1 min 578C), and extension (1 min 728C) then followed byan extension time of 5 min 728C. PCR products wereanalysed on a 3% agarose gel containing ethidium bromide.
Plasmid constructs
The pCMV-ScFv-ATF-1 that contains the cDNA coding forthe ScFv fragment speci®c for the transcription factor ATF-1(ScFv-ATF-1) has been previously described (Bosilevac et al.,1998). ScFv-ATF-1 cDNA was ®rst subcloned in pBluescriptII S+ (Stratagene, San Diego, CA, USA) using BglII/BamHIand NotI. PRSV-ScFv-ATF-1 was generated by placing thecDNA of ScFv-ATF-1 into the multiple cloning site of pRc/RSV (Invitrogen, San Diego, CA, USA) using HindIII andNotI. pRSV-KCREB was kindly provided by Dr Richard HGoodman (Oregon Health Sciences, Portland, OR, USA).PRSV-KCREB contains a full length CREB cDNA with a
single base pair substitution in the DNA-binding domain thatcauses a change from Arg287 to Leu (Walton et al., 1992). Incontrol experiments, a pRc/RSV (Invitrogen) construct thatlacks ScFv-ATF-1 or KCREB cDNA was used. The CRE-dependent plasmid (Somat-BglII) was obtained from Dr.Marc R. Montminy (Harvard Medical School, Boston, MA,USA). The Somat-BglII CAT construct contains thesomatostatin gene promoter from 771 to +53 linked tothe chloramphenicol acetyltransferase (CAT) reporter gene(Montminy et al., 1986). The pRL-b-actin vector contains theRenilla luciferase reporter gene (Promega) which is driven bythe b-actin promoter. The b-actin promoter does not containCRE element. The AP-2 binding site-luciferase reporter(3xAP-2Luc) construct contains an AP-2-dependent promoter(without CRE) linked to the Fire¯y luciferase reporter gene(Huang et al., 1998; Jean et al., 1998a).
Stable transfection of melanoma cells with ScFv-ATF-1
56105 MeWo cells were transfected with 5 mg of pRSV-ScFv-ATF-1 expression vector, pRSV-KCREB expressionvector or control pRc/RSV vector using 20 ml of Lipofectinreagent (Life Technologies Inc.). Transfections were carriedout according to the manufacturer's instructions. At 5 hfollowing transfection, the medium was changed to normalgrowth medium, and the cells were then further incubated for48 h at 378C. Cells were selected after 48 h with standardmedium containing G418 at 1 mg/ml (Life Technologies,Inc.). Two-to-three weeks later, G418-resistant colonies wereisolated by trypsinization using a penicylinder and establishedin culture to obtain clones. Pools of G418-resistant cells werealso prepared as we previously described (Xie et al., 1997a,b).
CAT assay
Melanoma cells (26105) were transfected with 3 mg of thebasic CAT expression vector with no promoter/enhancersequence (pCAT-Basic) from Promega or the Somat-BglIICAT construct using 20 ml of lipofectin according to themanufacturer instructions (Life Technologies, Inc.). Fiftynanograms of pRL-b-actin vector was introduced in eachexperiment to normalize for variations in transfectione�ciency. After 5 h, the transfection medium was replacedby normal growth medium, and the cells were furtherincubated for 48 h at 378C. For Thapsigargin treatment,Thapsigargin (1 mM) was added to the culture medium 24 hafter transfection and the cells were incubated for anadditional 24 h. The transfected cells were washed twice withPBS and lysed in the Reporter Lysis Bu�er (Promega). Afterremoval of cell debris by centrifugation, cell extracts were®rst assayed for Renilla luciferase activity using the Dual-Luciferase Reporter Assay System (Promega). Samples werenormalized as the same relative luciferase activity andassayed for CAT activity (Gorman et al., 1982) as wepreviously described (Hudson et al., 1995). Brie¯y, cellextracts were incubated with 0.025 mCi of [14C]chloramphe-nicol and 0.35 mM acetyl coenzyme A in 25 mM Tris-HCl pH7.9 for 2 h at 378C. The nonacetylated and acetylatedchloramphenicol were extracted in ethyl acetate andseparated by thin-layer chromatography with chloroform/methanol (95 : 5). The chromatograph was exposed to X-ray®lm. The conversion of chloramphenicol to the acetylatedform was quanti®ed by scanning densitometry of anautoradiograph with personal densitometer (MolecularDynamics, Sunnyvale, CA, USA). Each assay was repeatedat least three times; there was less than 10% variation amongindividual transfections.
Luciferase assay
Melanoma cells (26105) were transfected with 3 mg of the3xAP-2Luc and 50 ng of pRL-b-actin vector to normalize for
Regulation of melanoma growth and metastasis by ScFv anti-ATF-1D Jean et al
2728
Oncogene

transfection e�ciency by using 20 ml of Lipofectin reagent(Life Technologies Inc.). At 5 h following transfection, themedium was changed to normal growth medium, and thecells were further incubated for 48 h at 378C. The cells werethen washed with PBS and harvested in Passive Lysis Bu�er(Promega). Fire¯y and Renilla luciferase activities weremeasured using the Dual-Luciferase Reporter Assay System(Promega). Fire¯y luciferase activities were normalized forRenilla luciferase activities. Results are reported as theaverage of at least three independent experiments.
Detection of apoptosis by propidium iodide-staining and flowcytometry
Cells were plated overnight in culture medium describedabove without G418 and incubated in the presence of 1 mMThapsigargin (Tg) for 48 h. All cells (including those whichhad detached and those remaining adherent) were collected,and adherent cells were harvested in PBS containing 0.1%EDTA. Cells were washed twice with PBS, pelleted bycentrifugation, and resuspended in PBS containing 3 mM
Sodium Citrate, 0.1% Triton X-100 and 50 mg/ml propidiumiodide (PI) and incubated 2 ± 4 h. The propidium iodide-stained cells were subjected to ¯ow cytometric analysis on anEPICS Pro®le ¯ow cytometer (Coulter Corp., Miami, FL,USA). Results are reported as the average of at least threeindependent experiments as reported previously (Jean et al.,1998b).
Detection of apoptosis by TUNEL
Sections of subcutaneous tumors, characterized by the samesize, were prepared and stained for apoptosis by in situlabeling of DNA breaks using terminal deoxynucleotidetransferase (TdT) mediated dUTP-biotin nick end-labeling(TUNEL) method (Gavrieli et al., 1992). Following depar-a�nization and hydration, tumor sections were incubatedwith proteinase K (20 mg/ml) for 15 min at room temperature(RT), and washed. Endogenous peroxydase was inactivatedwith 2% H2O2 for 5 min at RT. Tumor sections were rinsedwith water and immersed in TdT bu�er (30 mM Trizma base,pH 7.2, 140 mM sodium cacodylate, 1 mM cobalt chloride).Tumor sections were then covered with TdT (0.3 units/ml)and dUTP-biotin in TdT bu�er and incubated in ahumidi®ed chamber at 378C for 60 min. The reaction wasterminated by transferring the slides to TB bu�er (300 mMsodium chloride, 30 mM sodium citrate) for 15 min at RT.Tumor sections were rinsed with water, covered with a 2%BSA solution for 10 min, rinsed in water, immersed in PBSfor 5 min, covered with Strep-Avidin Peroxydase (DakoCorp., Carpinteria, CA, USA), diluted 1 : 400 in bu�er, andincubated for 30 min at 378C. The sections were washed,immersed in PBS for 5 min and stained with 3-amino, 9-
ethylcarbazole (AEC) for 30 min at 378C. For positivecontrol, each tumor section was treated with DNase at1 ng/ml (Life Technologies Inc.) for 10 min in bu�er (30 mM
Trizm base, pH 7.2, 104 mM sodium cacodylate, 4 mM
MgCl2, 0.1 mM DTT). For negative control, TdT anddUTP-biotin were omitted in the procedure.
Immunofluorescence staining and confocal microscopy
Cells were plated (16105) in each chamber of a two-chamberFalcon glass slide (Becton Dickinson Labware, FranklinLakes, NJ, USA). Cells were grown to 60% con¯uence inMEM+10% FBS and prepared for immuno¯uorescentstaining. Brie¯y, the medium was decanted and cells were®xed with 4% paraformaldehyde for 20 min at 48C andwashed with PBS two times for 5 min. Cells werepermeabilized with 0.2% Triton X-100 for 10 min at 48Cand washed with PBS for 5 min. Cells were incubated withblocking bu�er (10% goat serum and 5% normal horseserum in PBS) for 10 min followed by incubation with A100anti-ATF-1 in blocking bu�er (1 : 100) overnight at 48C.Antibody to ATF-1 (A100) was a gift from Dr Michael RGreen (University of Massachusetts Medical Center, Worce-ster, MA, USA) (Liu et al., 1993). Slides were washed withPBS two times for 5 min and incubated with FITC-labeledsecondary antibody (anti-rabbit Ig) in blocking bu�er (1 : 200)for 2 h at room temperature. Imaging was done using a Zeissconfocal laser scanning microscope (upright version) using anargon laser (488 nm, power 10 mW). Signals were collectedby photomultipliers with 590 nm (PI) long pass ®lter and520 ± 560 nm (FITC) band pass ®lter, respectively. Digitizedimages were transmitted to a Macintosh-based image analysissystem through a GPIB interface using BDS-LSM software(Biological Detection Systems, Pittsburgh, PA, USA).Composite images were assembled using Adobe Photoshop(Adobe Systems, Inc., Mountain View, CA, USA).
Statistics
The signi®cance of the in vitro results was determined by theStudent's t-test (two-tailed); the signi®cance of the in vivometastasis results were determined by the Mann ±Whitney Utest.
AcknowledgmentsWe thank Dr MR Montiminy for the gift of the CRE-dependent promoter-CAT vector, Dr MR Green for theantibody against ATF-1, Dr RH Goodman for theKCREB expression vector, and Patherine Greenwood forexpert preparation of this manuscript. Supported in partby NIH grant CA76098 (to M Bar-Eli).
References
Bar-Eli M. (1997). J. Cell Physiol., 173, 275 ± 278.Barton K, Muthusamy N, Chanyangam M, Fischer C,
Clendenin C and Leiden JM. (1996). Nature, 379, 81 ± 85.Bohm M, Moellmann G, Cheng E, Alvarez-Franco M,
Wagner S, Sassone-Corsi P and Halaban R. (1995). CellGrowth Di�er., 6, 291 ± 302.
Bosilevac JM, Gilchrist CA, Jankowski PE, Paul S, Rees ARand Hinrichs SH. (1998). J. Biol. Chem., 273, 16874 ±16879.
Bosilevac JM, Olsen RJ, Bridge JA and Hinrichs SH. (1999).J. Biol. Chem., 274, 34811 ± 34818.
Buss®eld D, Bacher M, Moritz A, Gemsa D and Sprenger H.(1997). Immunobiol., 198, 291 ± 298.
de Fromentel CC, Gruel N, Venot C, Debussche L,Consieller E, Dureuil C, Teillaud J-L, Tocque B andBracco L. (1999). Oncogene, 18, 551 ± 557.
Fidler IJ. (1990). Cancer Res., 50, 6130 ± 6138.Fogh J, Wright WC and Loveless JD. (1997). J. Natl. Cancer
Inst., 58, 209 ± 214.Gavrieli Y, Sherman Y and Ben-Sasson SA. (1992). J. Cell
Biol., 119, 493 ± 501.Gonzalez GA andMontminy MR. (1989). Cell, 59, 675 ± 680.Gorman CM, Mo�att LF and Howard BH. (1982).Mol. Cell
Biol., 2, 1044 ± 1051.Graus-Porta D, Beerli R and Hyns N. (1995).Mol. Cell Biol.,
15, 1182 ± 1191.
Oncogene
Regulation of melanoma growth and metastasis by ScFv anti-ATF-1D Jean et al
2729

Gri�ths A, Malmquist M, Marks J, Bye J, McCa�erty J,Hollinger K, Gorick B, Hughes N and Winter G. (1993).EMBO J., 12, 725 ± 734.
Grim J, Deshane J, Feng M, Lieber A, Kay M and CurielDT. (1996). Am. J. Respir. Cell Mol. Biol., 15, 348 ± 354.
Halaban R. (1993). Semin. Cancer Biol., 4, 171 ± 181.Halaban R, Ghosh S and Baird A. (1987). In Vitro Cell
Develop. Biol., 23, 47 ± 52.Halaban R, Pomerantz SH, Marshall S and Lerner AB.
(1984). Arch. Biochem. Biophys., 230, 383 ± 387.Halaban R, Pomerantz SH, Marshall S, Lambert DT and
Lerner AB. (1983). J. Cell Biol., 97, 480 ± 488.Huang S, Jean D, Luca M, Tainsky M and Bar-Eli M. (1998).
EMBO J., 17, 4358 ± 4369.Huang S, Luca M, Gutman M, McConkey DJ, Langley KE,
Lyman SD and Bar-Eli M. (1996). Oncogene, 13, 2339 ±2347.
Hudson JM, Frade R and Bar-Eli M. (1995). DNA Cell Biol.,14, 759 ± 766.
Hurst HC, Totty NF and Jones NC. (1991). Nucl. Acids Res.,19, 4601 ± 4609.
Ishikawa M, Dennis JW, Man S and Kerbel RS. (1988).Cancer Res., 48, 665 ± 670.
Ivanov VN and Ronai Z. (1999). J. Biol. Chem., 274, 14079 ±14089.
Jean D, Gershenwald JE, Huang S, Luca M, Hudson MJ,Tainsky AM and Bar-Eli M. (1998a). J. Biol. Chem., 273,16501 ± 16508.
Jean D, Harbison M, McConkey DJ, Ronai Z and Bar-EliM. (1998b). J. Biol. Chem., 273, 24884 ± 24890.
Kopf AW, Salopek TG, Slade J, Marghoob AA and Bart RS.(1995). Cancer, 75, 684 ± 690.
Lee KA and Masson N. (1993). Biochim. Biophys. Acta.,1174, 221 ± 233.
Lemaigre FP, Ace CI and Green MR. (1993). Nucl. AcidsRes., 21, 2907 ± 2911.
Liu F, Thompson MA, Wagner S, Greenberg ME and GreenMR. (1993). J. Biol. Chem., 268, 6714 ± 6720.
Luca M, Xie S, Gutman M, Huang S and Bar-Eli M. (1995).Oncogene, 11, 1399 ± 1402.
Lu C and Kerbel RS. (1994). Curr. Opin. Oncol., 6, 212 ± 220.
Luca M, Huang S, Gershenwald JE, Singh RK, Reich R andBar-Eli M. (1997). Am. J. Pathol., 151, 1105 ± 1113.
Marasco W. (1995). Immunotech., 1, 1 ± 19.McConkey DJ and Orrenius S. (1996). J. Leukocyte Biol., 59,
775 ± 783.Meyer TE and Habener JF. (1993). Endocr. Rev., 14, 269 ±
290.Montminy MR, Sevarino KA, Wagner JA, Mandel G and
Goodman RH. (1986). Proc. Natl. Acad. Sci. USA, 83,6682 ± 6686.
Orten DJ, Strawhecker JM, Sanderson SD, Huang D,Prystowsky MB and Hinrichs SH. (1994). J. Biol. Chem.,269, 32254 ± 32263.
Raag R and Whitlow M. (1995). FASEB, 9, 73 ± 80.Radinsky R, Fidler IJ, Price JE, Esumi N, Tsan R, Petty CM,
Bucana CD and Bar-Eli M. (1994). Oncogene, 9, 1877 ±1883.
Rehfuss RP, Walton KM, Loriaux MM and Goodman RH.(1991). J. Biol. Chem., 266, 18431 ± 18434.
Richardson JH, Sodroski JG, Waldmann TA and MarascoWA. (1995). Proc. Natl. Acad. Sci. USA, 92, 3137 ± 3141.
Rutberg SE, Goldstein IM, Yang YM, Stackpole CE andRonai Z. (1994). Mol. Carcinogen., 10, 82 ± 87.
Sheng M, Thompson MA and Greenberg ME. (1991).Science, 252, 1427 ± 1430.
Singh RK, Gutman M, Reich R and Bar-Eli M. (1995).Cancer Res., 55, 3669 ± 3674.
Thastrup O, Cullen PJ, Drobak BK, Hanley MR andDawson AP. (1990). Proc. Natl. Acad. Sci. USA, 87,2466 ± 2470.
Vile RG, Nelson JA, Castleden S, Chong H and Hart IR.(1994). Cancer Res., 54, 6228 ± 6234.
Walton KM, Rehfuss RP, Chrivia JC, Lochner JE andGoodman RH. (1992). Mol. Endocrinol., 6, 647 ± 655.
Wilson BE, Mochon E and Boxer LM. (1996). Mol. Cell.Biol., 16, 5546 ± 5556.
Winter G and Milstein C. (1991). Nature, 349, 293 ± 299.Xie S, Huang S, Luca M, Gutman M, Reich R, Johnson JP
and Bar-Eli M. (1997a). Cancer Res., 57, 2295 ± 2303.Xie S, Price JE, Luca M, Jean D, Ronai Z and Bar-Eli M.
(1997b). Oncogene, 15, 2069 ± 2075.
Regulation of melanoma growth and metastasis by ScFv anti-ATF-1D Jean et al
2730
Oncogene
Copyright © 2022 FDOKUMEN




















