
Re-assessing the locations of components of the classical vesicle-mediated trafficking machinery in...
15
Re-assessing the locations of components of the classical vesicle-mediated trafficking machinery in transfected Plasmodium falciparum Akinola Adisa a,1 , Sarah Frankland a,c,1 , Melanie Rug a,d , Katherine Jackson a , Alexander G. Maier d , Peter Walsh e , Trevor Lithgow e , Nectarios Klonis a , Paul R. Gilson d , Alan F. Cowman d , Leann Tilley a,b, * a Department of Biochemistry, La Trobe University, Melbourne 3086, Vic., Australia b ARC Centre of Excellence for Coherent X-ray Science, La Trobe University, Melbourne 3086, Vic., Australia c Cooperative Research Centre for Vaccine Technology, La Trobe University, Melbourne 3086, Vic., Australia d The Walter and Eliza Hall Institute of Medical Research, 1G Royal Parade, Parkville, Melbourne 3050, Vic., Australia e Department of Biochemistry and Molecular Biology and Bio21 Molecular Science and Biotechnology Institute, University of Melbourne, Parkville 3010, Vic., Australia Received 11 January 2007; received in revised form 15 February 2007; accepted 16 February 2007 Abstract The malaria parasite, Plasmodium falciparum, exports proteins beyond the confines of its own plasma membrane, however there is debate regarding the machinery used for these trafficking events. We have generated transgenic parasites expressing chimeric proteins and used immunofluorescence studies to determine the locations of plasmodial homologues of the COPII component, Sar1p, and the Golgi-docking protein, Bet3p. The P. falciparum Sar1p (PfSar1p) chimeras bind to the endoplasmic reticulum surface and define a net- work of membranes wrapped around parasite nuclei. As the parasite matures, the endomembrane systems of individual merozoites remain interconnected until very late in schizogony. Antibodies raised against plasmodial Bet3p recognise two foci of reactivity in early parasite stages that increase in number as the parasite matures. Some of the P. falciparum Bet3p (PfBet3p) compartments are juxtaposed to compartments defined by the cis Golgi marker, PfGRASP, while others are distributed through the cytoplasm. The compartments defined by the trans Golgi marker, PfRab6, are separate, suggesting that the Golgi is dispersed. Bet3p-green fluorescent protein (GFP) is partly associated with punctate structures but a substantial population diffuses freely in the parasite cytoplasm. By contrast, yeast Bet3p is very tightly associated with immobile structures. This study challenges the view that the COPII complex and the Golgi apparatus are exported into the infected erythrocyte cytoplasm. Ó 2007 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved. Keywords: Malaria; Sar1p; Bet3p; GTP-binding protein; Membrane trafficking; Secretion 1. Introduction Plasmodium falciparum, the most virulent of the human malaria parasites, spends part of its life cycle within a parasitophorous vacuole inside the red blood cells (RBCs) of its host. From this intracellular niche the parasite alters the physical properties of its host cell to facilitate nutrient uptake and to cause adhesion to the vascular endothelium (Deitsch and Wellems, 1996; Kyes et al., 2001; Rowe and Kyes, 2004). The parasite achieves these changes by export- ing some of its proteins beyond its own plasma membrane and the parasitophorous vacuole membrane, and traffick- ing them to sites in the RBC cytoplasm and the RBC 0020-7519/$30.00 Ó 2007 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.ijpara.2007.02.009 * Corresponding author. Address: Department of Biochemistry, La Trobe University, Melbourne 3086, Vic., Australia. Tel.: +61 3 9479 1375; fax: +61 3 9479 2467. E-mail address: [email protected] (L. Tilley). 1 These authors contributed equally to this work. www.elsevier.com/locate/ijpara International Journal for Parasitology 37 (2007) 1127–1141
Transcript of Re-assessing the locations of components of the classical vesicle-mediated trafficking machinery in...

www.elsevier.com/locate/ijpara
International Journal for Parasitology 37 (2007) 1127–1141
Re-assessing the locations of components of the classicalvesicle-mediated trafficking machinery in transfected
Plasmodium falciparum
Akinola Adisa a,1, Sarah Frankland a,c,1, Melanie Rug a,d, Katherine Jackson a,Alexander G. Maier d, Peter Walsh e, Trevor Lithgow e, Nectarios Klonis a,
Paul R. Gilson d, Alan F. Cowman d, Leann Tilley a,b,*
a Department of Biochemistry, La Trobe University, Melbourne 3086, Vic., Australiab ARC Centre of Excellence for Coherent X-ray Science, La Trobe University, Melbourne 3086, Vic., Australiac Cooperative Research Centre for Vaccine Technology, La Trobe University, Melbourne 3086, Vic., Australia
d The Walter and Eliza Hall Institute of Medical Research, 1G Royal Parade, Parkville, Melbourne 3050, Vic., Australiae Department of Biochemistry and Molecular Biology and Bio21 Molecular Science and Biotechnology Institute,
University of Melbourne, Parkville 3010, Vic., Australia
Received 11 January 2007; received in revised form 15 February 2007; accepted 16 February 2007
Abstract
The malaria parasite, Plasmodium falciparum, exports proteins beyond the confines of its own plasma membrane, however there isdebate regarding the machinery used for these trafficking events. We have generated transgenic parasites expressing chimeric proteinsand used immunofluorescence studies to determine the locations of plasmodial homologues of the COPII component, Sar1p, and theGolgi-docking protein, Bet3p. The P. falciparum Sar1p (PfSar1p) chimeras bind to the endoplasmic reticulum surface and define a net-work of membranes wrapped around parasite nuclei. As the parasite matures, the endomembrane systems of individual merozoitesremain interconnected until very late in schizogony. Antibodies raised against plasmodial Bet3p recognise two foci of reactivity in earlyparasite stages that increase in number as the parasite matures. Some of the P. falciparum Bet3p (PfBet3p) compartments are juxtaposedto compartments defined by the cis Golgi marker, PfGRASP, while others are distributed through the cytoplasm. The compartmentsdefined by the trans Golgi marker, PfRab6, are separate, suggesting that the Golgi is dispersed. Bet3p-green fluorescent protein(GFP) is partly associated with punctate structures but a substantial population diffuses freely in the parasite cytoplasm. By contrast,yeast Bet3p is very tightly associated with immobile structures. This study challenges the view that the COPII complex and the Golgiapparatus are exported into the infected erythrocyte cytoplasm.� 2007 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
Keywords: Malaria; Sar1p; Bet3p; GTP-binding protein; Membrane trafficking; Secretion
1. Introduction
Plasmodium falciparum, the most virulent of the humanmalaria parasites, spends part of its life cycle within a
0020-7519/$30.00 � 2007 Australian Society for Parasitology Inc. Published b
doi:10.1016/j.ijpara.2007.02.009
* Corresponding author. Address: Department of Biochemistry, LaTrobe University, Melbourne 3086, Vic., Australia. Tel.: +61 3 9479 1375;fax: +61 3 9479 2467.
E-mail address: [email protected] (L. Tilley).1 These authors contributed equally to this work.
parasitophorous vacuole inside the red blood cells (RBCs)of its host. From this intracellular niche the parasite altersthe physical properties of its host cell to facilitate nutrientuptake and to cause adhesion to the vascular endothelium(Deitsch and Wellems, 1996; Kyes et al., 2001; Rowe andKyes, 2004). The parasite achieves these changes by export-ing some of its proteins beyond its own plasma membraneand the parasitophorous vacuole membrane, and traffick-ing them to sites in the RBC cytoplasm and the RBC
y Elsevier Ltd. All rights reserved.

1128 A. Adisa et al. / International Journal for Parasitology 37 (2007) 1127–1141
membrane (Przyborski et al., 2003; Cooke et al., 2004;Marti et al., 2005; Lanzer et al., 2006). This has led toconsiderable interest in the protein trafficking componentsof P. falciparum.
The presence of a ‘‘classical’’ secretory pathway withinthe parasite is indicated by the identification of homo-logues of a number of trafficking components. The endo-plasmic reticulum (ER) functions as the site of proteintranslocation and integration for all proteins destinedfor secretion. Components of the secretory pathway asso-ciated with the ER, such as PfERC and PfBip (Kumaret al., 1988; La Greca et al., 1997; van Dooren et al.,2005), the COPI protein, PfdCOP (Adisa et al., 2002)and the retrieval receptor, PfERD2 (Van Wye et al.,1996), have been identified in P. falciparum. Analysis ofthe genome has also revealed homologues of componentsof the signal recognition particle, the translocon, the sig-nal peptidase complex and many proteins involved in ves-icle budding, docking and fusion (Gardner et al., 2002).However, protein trafficking within the parasite is compli-cated by that fact that the parasite harbours a number ofunusual organelles such as the apicoplast, the digestivevacuole and the apical organelles (Foth et al., 2003; Kle-mba et al., 2004; Margos et al., 2004; Rug et al., 2004).Moreover, the machinery for protein glycoslyation is lar-gely lacking in P. falciparum and the Golgi appears to bequite rudimentary with the cis Golgi spatially separatedfrom the trans Golgi (Elmendorf and Haldar, 1993; deCastro et al., 1996; Van Wye et al., 1996; Adisa et al.,2002; Struck et al., 2005).
The mature human RBC has no endogenous vesicle-mediated trafficking machinery and it has been suggestedthat components of the parasite trafficking machinery areexported to the host cell cytoplasm. Indeed, antibodiesraised against plasmodial homologues of the COPII vesicleproteins, Sar1p, Sec31p and Sec23p (Albano et al., 1999;Adisa et al., 2001; Taraschi et al., 2003; Wickert et al.,2003) recognise parasite-derived structures in the host cellcytoplasm known as Maurer’s clefts. Similarly, antibodiesagainst a protein referred to as antigen 41-2 (Knappet al., 1989) which shows homology to the Golgi proteins,Bet3p and Trs31p (Gardner et al., 2002), and plasmodialN-ethylmaleimide-sensitive factor (PfNSF) (Hayashiet al., 2001), which is involved in soluble N-ethylmalei-mide-sensitive fusion protein receptors (SNARE)-mediatedmembrane fusion, also recognise structures in the RBCcytoplasm. Moreover, antibodies raised against a Maurer’scleft protein recognise mammalian Golgi (Li et al., 1995)and sphingomyelin synthase activity, which is found pre-dominantly in the Golgi of other eukaryotic cells, has beenreported to be located within the host cell (Elmendorf andHaldar, 1993). This suggests that some Golgi functionsmight be exported to the RBC cytoplasm, although thesphingomyelin synthase gene has not been identified. Thesedata have led to the proposal that the malaria parasiteexports vesicle-mediated trafficking machinery outside theboundaries of its own plasma membrane.
Vesicle-mediated transport is dependent upon thehydrolysis of guanosine triphosphate (GTP) (Tang et al.,2005), however recent data indicates that GTPase inhibi-tors do not prevent trafficking of proteins across theinfected RBC cytoplasm (Frankland et al., 2006). Thus,there is some debate as to whether classical secretory com-ponents are involved in ferrying parasite proteins to desti-nations in the host cell. Indeed, most of the evidence forexport of secretory pathway components is based onimmunolabelling experiments. However, some concernexists about the effects of fixing samples and about theselectivity of antibody-based markers for different com-partments in P. falciparum (Anders et al., 1987; Tonkinet al., 2004; Spielmann et al., 2006). With the advent oftransfection procedures for P. falciparum, it has becomepossible to use ‘‘reporter’’ proteins (eg. green fluorescentprotein, GFP) to monitor the locations and dynamics ofcomponents within living cells (Waller et al., 2000; Wick-ham et al., 2001). In this work we have generated transfec-ted parasites to follow the fates of the plasmodialhomologues of the COPII coat protein, Sar1p, andGolgi-docking protein, Bet3p.
2. Materials and methods
2.1. Plasmid constructs and transfection of P. falciparum
We prepared transfection constructs in the pARL1a vec-tor (Marti et al., 2004). The sequence encoding full-lengthPfSar1p was PCR amplified from a cDNA library (Raw-lings and Kaslow, 1992) with or without a five glycine lin-ker. The constructs were cloned into pARL1a (Marti et al.,2004) or a modified vector, pARLmGFPmT, as previouslydescribed (Spycher et al., 2006). Full length P. falciparum
BET3 (without the stop codon) was amplified from acDNA preparation (kindly donated by Dr Sue Kyes, Insti-tute of Molecular Medicine, Oxford, UK) and ligated ontothe 5 0 end of GFP in pARLmGFPmT. All constructs wereverified by enzyme digestion and sequencing. Plasmodiumfalciparum-infected RBCs (3D7 ring stage) were transfectedwith the constructs and cultured in the presence of 10 nMWR99210 (Fidock and Wellems, 1997).
2.2. Parasites and solubility studies
Parasites (3D7 strain) were cultured as described previ-ously (Raynes et al., 1996) using RBCs and pooled serumfrom the Red Cross Transfusion Service, Melbourne. Cul-tures were synchronised and harvested by floatation on aPercoll/sorbitol cushion or by saponin-induced release.For studies of PfSar1p solubility, mature parasitised RBCs(�107 cells) were either sonicated in PBS or 50 mMNaHCO3, pH 10, at 4 �C, or agitated in 1% Triton X-100in PBS for 20 min at 4 �C. For PfBet3p solubility studies,parasites were vortexed in either 2% Triton X-100 in N-2-hydroxyehtylpiperazine-N 0-2-ethanesulfonic acid (Hepes)or 1 M NaCl in Hepes (Sacher et al., 2000). The soluble

A. Adisa et al. / International Journal for Parasitology 37 (2007) 1127–1141 1129
and particulate fractions were separated by centrifugation(100,000g, 60 min, 4 �C). The solubility of the ER lumen-located P. falciparum protein, PfERC (La Greca et al.,1997), was monitored to assess the efficiency of the extrac-tion procedures.
2.3. Expression of recombinant proteins
PfBET3 (561 bp) was ligated into pGEX-5X-3 (GEHealthcare) restricted with BamHI and EcoRI, for expres-sion in Escherichia coli (BL21 strain). The resultant recom-binant protein, fused to glutathione-S-transferase (GST),was purified on a glutathione–Sepharose column (GEHealthcare) according to the manufacturer’s protocols.The protein was analysed by Western blotting (using ananti-GST antibody, GE Healthcare) and the identity ofthe protein was confirmed by N-terminal sequencing. Thepurified protein (GST-PfBet3p) was used to generate anti-sera in rabbits with approval from the La Trobe UniversityAnimal Ethics Committee. GST-PfBet3p was cleaved withFactor Xa to release the 22 kDa PfBet3p.
2.4. Immunochemical techniques
For time-course analyses, aliquots of a synchronous cul-ture of parasite-infected RBCs (3D7 strain, 10% parasite-mia) were harvested at 8 h intervals and washed withPBS. The cells were lysed in 10 ml of 0.15% saponin inPBS and haemoglobin-depleted infected RBCs were col-lected by centrifugation at 18,000g for 10 min. The sampleswere subjected to SDS–PAGE (12% or 14% acrylamide)and Western blotting was performed using enhancedchemiluminescence (ECL) detection (La Greca et al.,1997). Antibodies recognising GFP (murine monoclonal)were obtained from Roche.
2.5. Fluorescence microscopy
Microscopy was performed using Olympus IX8 andBX50 epifluorescence microscopes or an inverted LeicaTCS-SP2 confocal microscope, using 100· oil immersionobjectives (1.4 NA). Live transfectants were mounted weton a glass slide and imaged within 20 min at 20 �C. Trans-fectants were incubated in medium containing 0.25 lM ERTracker� for 30 min. Thin smears of infected RBCs werefixed with acetone/methanol (1:1) at 0 �C, 10 min, andprobed with antibodies recognising PfSar1p (Albanoet al., 1999), KAHRP (Wickham et al., 2001), MAHRP1(Spycher et al., 2003), PfEMP1 (Frankland et al., 2006),PfERC (Albano et al., 1999), PfRab6 (kindly donated byDr Debasish Chattopadhyay, University of Alabama atBirmingham, USA and Dr Gordon Langsley, PasteurInstitute, France) and GFP (Roche), and visualised usingeither fluorescein isothiocyanate (FITC)- or Alexa-Fluor568-conjugated goat anti-rabbit or anti-mouse IgG (Sigma,Molecular Probes). The intensities of the images wereadjusted to optimise the fluorescence signal.
2.6. Fluorescence recovery after photobleaching
Details of photobleaching measurements have beendescribed elsewhere (Klonis et al., 2002; Spycher et al.,2003; Knuepfer et al., 2005). Image processing, includingbackground correction, smoothing and image analysis wereperformed using NIH ImageJ (http://rsb.info.nih.gov/ij) asdescribed previously (Adisa et al., 2003).
2.7. Yeast complementation and expression experiments with
PfBET3
A PCR amplified PfBET3 gene was cloned into pYEUra3(Clonetech) and pYX141 (centromeric, single copy vector;formerly distributed by Novagen) plasmids and transformedinto a Saccharomyces cerevisiae bet3 heterozygous diploidmutant strain (BY4743 MATa/MATa; his3D1/his3-
D1;leu2D0/leu2D0;met15D0/MET15;lys2D0/LYS2;ura3D0/
ura3D0;bet3::kanMX4/BET3) ((Brachmann et al., 1998);EUROFAN II Acc No.Y2000) kindly provided byProf Rod Devenish, Monash University, Australia. Trans-formed cells were selected on synthetic dextrose (SD)–uracilor SD–leucine medium (which selects for the presence ofpYEUra3 and pYX141 vectors, respectively). Leucine oruracil auxotrophic colonies were plated on sporulationplates and zymolase-digested tetrads were dissected using aSinger MSM dissecting microscope. Open-reading framesrepresenting PfBET3 and ScBET3 were ligated into the yeastexpression vector pGFP-C-FUS (Niedenthal et al., 1996)and transformed into diploid yeast cells (MH272 3fa/aleu2/leu2, ura3/ura3, his3/his3, rme1/rme1, trp1/trp1, ade2/
ade2) for expression as C-terminal GFP-tagged proteins.Transformed cells were selected on SD–uracil platesand mounted in 0.5% agarose and viewed by confocalmicroscopy.
3. Results
3.1. Full length PfSar1p fused to the N- or C-terminus of
GFP can be expressed in transfected P. falciparum
The PfSAR1 gene (576 bp) encodes a protein of 192amino acids with a predicted Mr of 22,020 (Albano et al.,1999). The protein has a series of motifs that are conservedin the small GTPase family (Nakano and Muramatsu,1989; Barlowe, 2002). We generated chimeras comprisingthe full-length PfSar1p coding sequence linked to the 5 0
end (PfSar1p-GFP) or the 3 0 end (GFP-PfSar1p) of thesequence encoding mut 2-enhanced GFP. The chimeraswere inserted into the transfection vector, pARL1a, fromwhich expression of the coding sequence is driven by themoderate constitutive promoter, PfCRT 5 0 (Marti et al.,2004). The resultant GFP fusion proteins are expressedfrom stably maintained episomes within transformed 3D7P. falciparum blood stage parasites, under continuousselection with the antifolate drug WR99210 (Crabb et al.,2004).
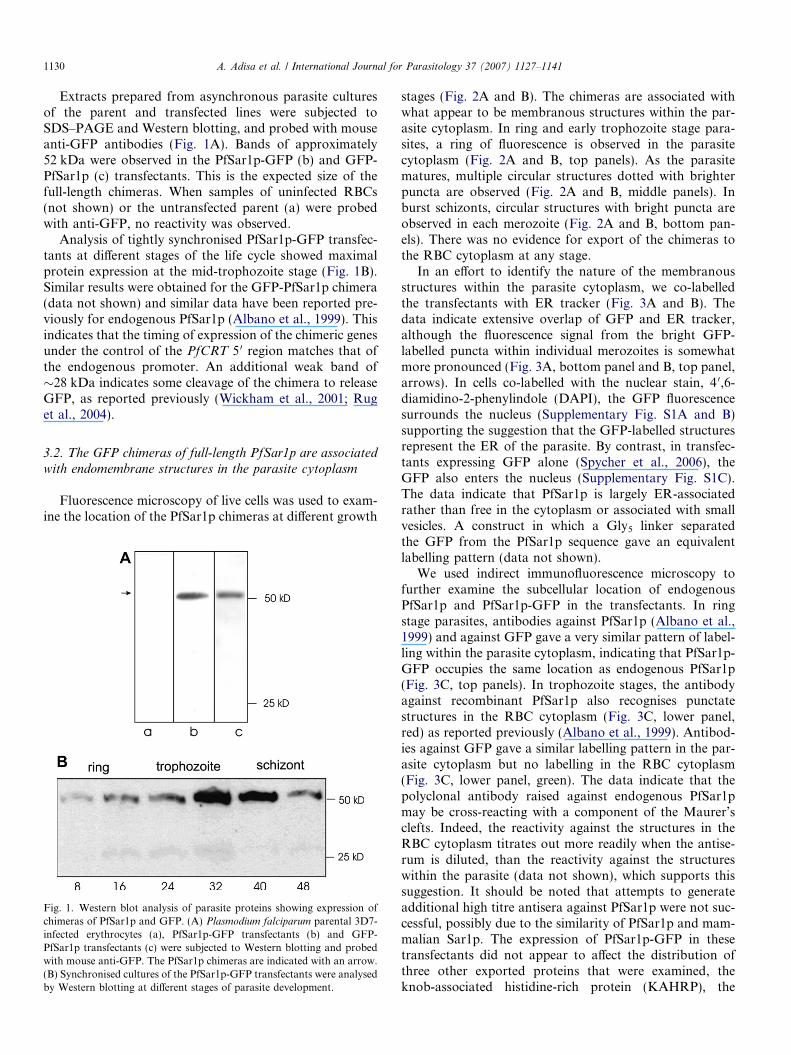
1130 A. Adisa et al. / International Journal for Parasitology 37 (2007) 1127–1141
Extracts prepared from asynchronous parasite culturesof the parent and transfected lines were subjected toSDS–PAGE and Western blotting, and probed with mouseanti-GFP antibodies (Fig. 1A). Bands of approximately52 kDa were observed in the PfSar1p-GFP (b) and GFP-PfSar1p (c) transfectants. This is the expected size of thefull-length chimeras. When samples of uninfected RBCs(not shown) or the untransfected parent (a) were probedwith anti-GFP, no reactivity was observed.
Analysis of tightly synchronised PfSar1p-GFP transfec-tants at different stages of the life cycle showed maximalprotein expression at the mid-trophozoite stage (Fig. 1B).Similar results were obtained for the GFP-PfSar1p chimera(data not shown) and similar data have been reported pre-viously for endogenous PfSar1p (Albano et al., 1999). Thisindicates that the timing of expression of the chimeric genesunder the control of the PfCRT 5 0 region matches that ofthe endogenous promoter. An additional weak band of�28 kDa indicates some cleavage of the chimera to releaseGFP, as reported previously (Wickham et al., 2001; Ruget al., 2004).
3.2. The GFP chimeras of full-length PfSar1p are associated
with endomembrane structures in the parasite cytoplasm
Fluorescence microscopy of live cells was used to exam-ine the location of the PfSar1p chimeras at different growth
Fig. 1. Western blot analysis of parasite proteins showing expression ofchimeras of PfSar1p and GFP. (A) Plasmodium falciparum parental 3D7-infected erythrocytes (a), PfSar1p-GFP transfectants (b) and GFP-PfSar1p transfectants (c) were subjected to Western blotting and probedwith mouse anti-GFP. The PfSar1p chimeras are indicated with an arrow.(B) Synchronised cultures of the PfSar1p-GFP transfectants were analysedby Western blotting at different stages of parasite development.
stages (Fig. 2A and B). The chimeras are associated withwhat appear to be membranous structures within the par-asite cytoplasm. In ring and early trophozoite stage para-sites, a ring of fluorescence is observed in the parasitecytoplasm (Fig. 2A and B, top panels). As the parasitematures, multiple circular structures dotted with brighterpuncta are observed (Fig. 2A and B, middle panels). Inburst schizonts, circular structures with bright puncta areobserved in each merozoite (Fig. 2A and B, bottom pan-els). There was no evidence for export of the chimeras tothe RBC cytoplasm at any stage.
In an effort to identify the nature of the membranousstructures within the parasite cytoplasm, we co-labelledthe transfectants with ER tracker (Fig. 3A and B). Thedata indicate extensive overlap of GFP and ER tracker,although the fluorescence signal from the bright GFP-labelled puncta within individual merozoites is somewhatmore pronounced (Fig. 3A, bottom panel and B, top panel,arrows). In cells co-labelled with the nuclear stain, 4 0,6-diamidino-2-phenylindole (DAPI), the GFP fluorescencesurrounds the nucleus (Supplementary Fig. S1A and B)supporting the suggestion that the GFP-labelled structuresrepresent the ER of the parasite. By contrast, in transfec-tants expressing GFP alone (Spycher et al., 2006), theGFP also enters the nucleus (Supplementary Fig. S1C).The data indicate that PfSar1p is largely ER-associatedrather than free in the cytoplasm or associated with smallvesicles. A construct in which a Gly5 linker separatedthe GFP from the PfSar1p sequence gave an equivalentlabelling pattern (data not shown).
We used indirect immunofluorescence microscopy tofurther examine the subcellular location of endogenousPfSar1p and PfSar1p-GFP in the transfectants. In ringstage parasites, antibodies against PfSar1p (Albano et al.,1999) and against GFP gave a very similar pattern of label-ling within the parasite cytoplasm, indicating that PfSar1p-GFP occupies the same location as endogenous PfSar1p(Fig. 3C, top panels). In trophozoite stages, the antibodyagainst recombinant PfSar1p also recognises punctatestructures in the RBC cytoplasm (Fig. 3C, lower panel,red) as reported previously (Albano et al., 1999). Antibod-ies against GFP gave a similar labelling pattern in the par-asite cytoplasm but no labelling in the RBC cytoplasm(Fig. 3C, lower panel, green). The data indicate that thepolyclonal antibody raised against endogenous PfSar1pmay be cross-reacting with a component of the Maurer’sclefts. Indeed, the reactivity against the structures in theRBC cytoplasm titrates out more readily when the antise-rum is diluted, than the reactivity against the structureswithin the parasite (data not shown), which supports thissuggestion. It should be noted that attempts to generateadditional high titre antisera against PfSar1p were not suc-cessful, possibly due to the similarity of PfSar1p and mam-malian Sar1p. The expression of PfSar1p-GFP in thesetransfectants did not appear to affect the distribution ofthree other exported proteins that were examined, theknob-associated histidine-rich protein (KAHRP), the

Fig. 2. Expression of PfSar1p chimeras at different stages of the intraerythrocytic cycle of P. falciparum. The images represent differential interferencecontrast (DIC) micrographs and the GFP fluorescence signal for (A) PfSar1p-GFP and (B) GFP-PfSar1p. Ring and early trophozoite stage parasites (toprows) show a perinuclear ring of fluorescence in the parasite cytoplasm with bright puncta (arrows). Schizont stage parasites (fourth row) show rings offluorescence around individual nuclei (see Supplementary Fig. S1). In burst schizonts (bottom row), a focus of bright GFP fluorescence is often observed atone end of the merozoite (arrows). Bar = 5 lm.
A. Adisa et al. / International Journal for Parasitology 37 (2007) 1127–1141 1131
membrane-associated histidine-rich protein-1 (MAHRP1)and the P. falciparum RBC membrane protein-1 (PfEMP1)(data not shown).
3.3. Characterisation of the association of the PfSar1p
chimeras with the ER membranes
We examined the solubility properties of PfSar1p-GFPand compared them with those of the ER lumen proteinPfERC (Fig. 4). Vortexing parasite samples in PBS partlyreleases PfERC from the disrupted ER compartment, how-ever PfSar1p-GFP remains associated with the pellet, indi-cating that it is tightly associated with a membranecompartment as reported previously for endogenousPfSar1p (Albano et al., 1999). Sonication in a high pH,low ionic strength buffer to release both soluble and periph-
eral proteins completely released PfERC but only partlyreleased PfSar1p-GFP, indicating that hydrophobic inter-actions may contribute to the binding of PfSar1p-GFP tothe ER membranes as for endogenous PfSar1p (Albanoet al., 1999). Treatment of samples with the non-ionicdetergent, Triton X-100, released PfSar1p-GFP, suggestingthat this protein is associated with membranes by hydro-phobic interactions. Our previous data showed that endog-enous PfSar1p is not released by Triton X-100, whichsuggests that PfSar1p-GFP may be less tightly associatedwith the cytoskeletal elements than endogenous PfSar1p(Albano et al., 1999). Similar data were obtained forGFP-PfSar1p (not shown).
To further examine the physical nature of the interac-tion of the PfSar1p chimeras with the ER membranes, weperformed photobleaching studies (Klonis et al., 2002)
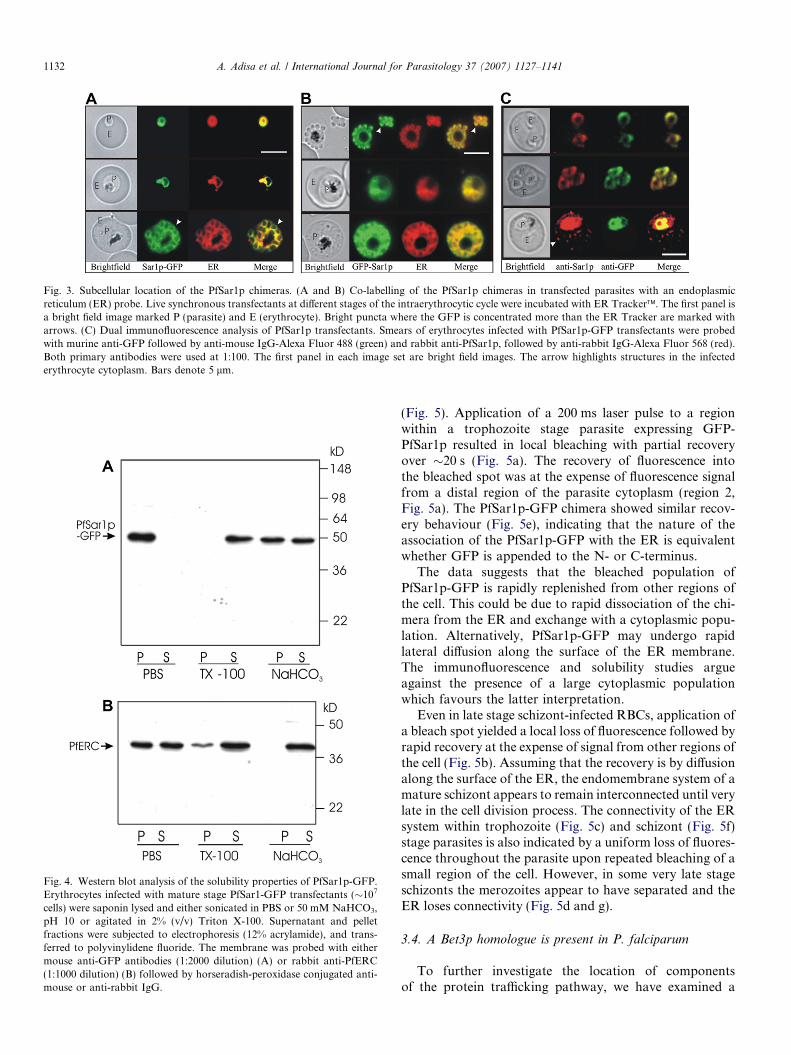
Fig. 3. Subcellular location of the PfSar1p chimeras. (A and B) Co-labelling of the PfSar1p chimeras in transfected parasites with an endoplasmicreticulum (ER) probe. Live synchronous transfectants at different stages of the intraerythrocytic cycle were incubated with ER Tracker�. The first panel isa bright field image marked P (parasite) and E (erythrocyte). Bright puncta where the GFP is concentrated more than the ER Tracker are marked witharrows. (C) Dual immunofluorescence analysis of PfSar1p transfectants. Smears of erythrocytes infected with PfSar1p-GFP transfectants were probedwith murine anti-GFP followed by anti-mouse IgG-Alexa Fluor 488 (green) and rabbit anti-PfSar1p, followed by anti-rabbit IgG-Alexa Fluor 568 (red).Both primary antibodies were used at 1:100. The first panel in each image set are bright field images. The arrow highlights structures in the infectederythrocyte cytoplasm. Bars denote 5 lm.
Fig. 4. Western blot analysis of the solubility properties of PfSar1p-GFP.Erythrocytes infected with mature stage PfSar1-GFP transfectants (�107
cells) were saponin lysed and either sonicated in PBS or 50 mM NaHCO3,pH 10 or agitated in 2% (v/v) Triton X-100. Supernatant and pelletfractions were subjected to electrophoresis (12% acrylamide), and trans-ferred to polyvinylidene fluoride. The membrane was probed with eithermouse anti-GFP antibodies (1:2000 dilution) (A) or rabbit anti-PfERC(1:1000 dilution) (B) followed by horseradish-peroxidase conjugated anti-mouse or anti-rabbit IgG.
1132 A. Adisa et al. / International Journal for Parasitology 37 (2007) 1127–1141
(Fig. 5). Application of a 200 ms laser pulse to a regionwithin a trophozoite stage parasite expressing GFP-PfSar1p resulted in local bleaching with partial recoveryover �20 s (Fig. 5a). The recovery of fluorescence intothe bleached spot was at the expense of fluorescence signalfrom a distal region of the parasite cytoplasm (region 2,Fig. 5a). The PfSar1p-GFP chimera showed similar recov-ery behaviour (Fig. 5e), indicating that the nature of theassociation of the PfSar1p-GFP with the ER is equivalentwhether GFP is appended to the N- or C-terminus.
The data suggests that the bleached population ofPfSar1p-GFP is rapidly replenished from other regions ofthe cell. This could be due to rapid dissociation of the chi-mera from the ER and exchange with a cytoplasmic popu-lation. Alternatively, PfSar1p-GFP may undergo rapidlateral diffusion along the surface of the ER membrane.The immunofluorescence and solubility studies argueagainst the presence of a large cytoplasmic populationwhich favours the latter interpretation.
Even in late stage schizont-infected RBCs, application ofa bleach spot yielded a local loss of fluorescence followed byrapid recovery at the expense of signal from other regions ofthe cell (Fig. 5b). Assuming that the recovery is by diffusionalong the surface of the ER, the endomembrane system of amature schizont appears to remain interconnected until verylate in the cell division process. The connectivity of the ERsystem within trophozoite (Fig. 5c) and schizont (Fig. 5f)stage parasites is also indicated by a uniform loss of fluores-cence throughout the parasite upon repeated bleaching of asmall region of the cell. However, in some very late stageschizonts the merozoites appear to have separated and theER loses connectivity (Fig. 5d and g).
3.4. A Bet3p homologue is present in P. falciparum
To further investigate the location of componentsof the protein trafficking pathway, we have examined a
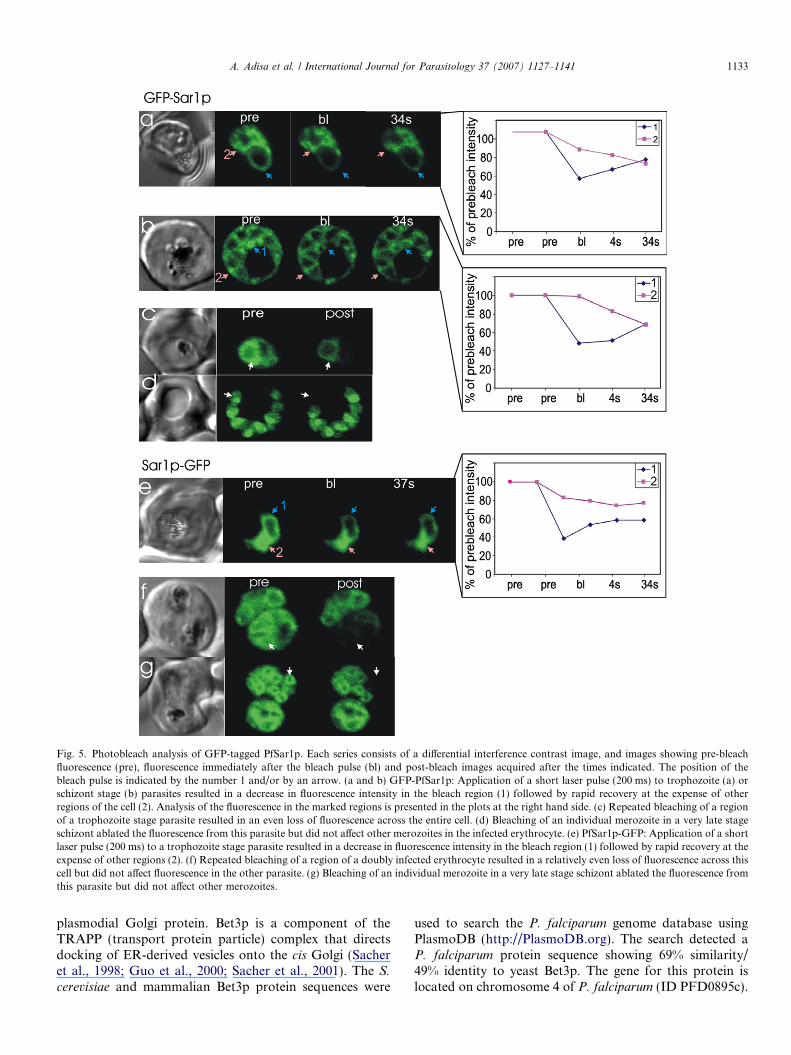
Fig. 5. Photobleach analysis of GFP-tagged PfSar1p. Each series consists of a differential interference contrast image, and images showing pre-bleachfluorescence (pre), fluorescence immediately after the bleach pulse (bl) and post-bleach images acquired after the times indicated. The position of thebleach pulse is indicated by the number 1 and/or by an arrow. (a and b) GFP-PfSar1p: Application of a short laser pulse (200 ms) to trophozoite (a) orschizont stage (b) parasites resulted in a decrease in fluorescence intensity in the bleach region (1) followed by rapid recovery at the expense of otherregions of the cell (2). Analysis of the fluorescence in the marked regions is presented in the plots at the right hand side. (c) Repeated bleaching of a regionof a trophozoite stage parasite resulted in an even loss of fluorescence across the entire cell. (d) Bleaching of an individual merozoite in a very late stageschizont ablated the fluorescence from this parasite but did not affect other merozoites in the infected erythrocyte. (e) PfSar1p-GFP: Application of a shortlaser pulse (200 ms) to a trophozoite stage parasite resulted in a decrease in fluorescence intensity in the bleach region (1) followed by rapid recovery at theexpense of other regions (2). (f) Repeated bleaching of a region of a doubly infected erythrocyte resulted in a relatively even loss of fluorescence across thiscell but did not affect fluorescence in the other parasite. (g) Bleaching of an individual merozoite in a very late stage schizont ablated the fluorescence fromthis parasite but did not affect other merozoites.
A. Adisa et al. / International Journal for Parasitology 37 (2007) 1127–1141 1133
plasmodial Golgi protein. Bet3p is a component of theTRAPP (transport protein particle) complex that directsdocking of ER-derived vesicles onto the cis Golgi (Sacheret al., 1998; Guo et al., 2000; Sacher et al., 2001). The S.
cerevisiae and mammalian Bet3p protein sequences were
used to search the P. falciparum genome database usingPlasmoDB (http://PlasmoDB.org). The search detected aP. falciparum protein sequence showing 69% similarity/49% identity to yeast Bet3p. The gene for this protein islocated on chromosome 4 of P. falciparum (ID PFD0895c).
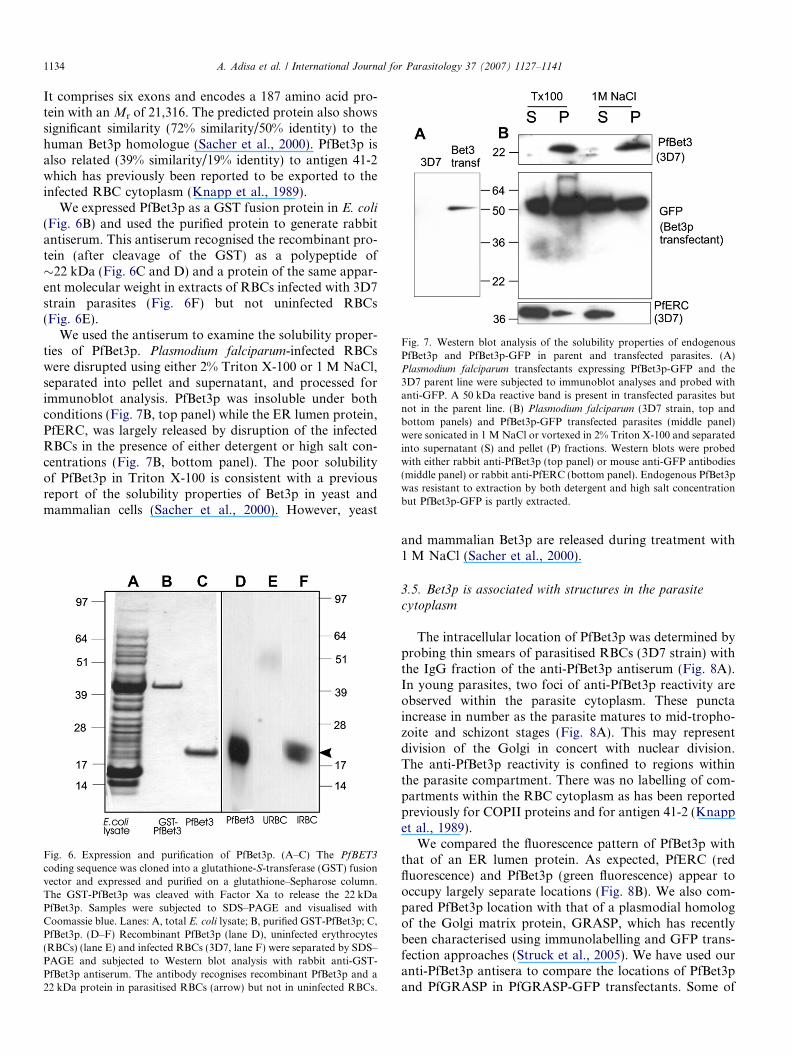
Fig. 7. Western blot analysis of the solubility properties of endogenousPfBet3p and PfBet3p-GFP in parent and transfected parasites. (A)Plasmodium falciparum transfectants expressing PfBet3p-GFP and the3D7 parent line were subjected to immunoblot analyses and probed withanti-GFP. A 50 kDa reactive band is present in transfected parasites butnot in the parent line. (B) Plasmodium falciparum (3D7 strain, top andbottom panels) and PfBet3p-GFP transfected parasites (middle panel)were sonicated in 1 M NaCl or vortexed in 2% Triton X-100 and separatedinto supernatant (S) and pellet (P) fractions. Western blots were probedwith either rabbit anti-PfBet3p (top panel) or mouse anti-GFP antibodies(middle panel) or rabbit anti-PfERC (bottom panel). Endogenous PfBet3pwas resistant to extraction by both detergent and high salt concentrationbut PfBet3p-GFP is partly extracted.
1134 A. Adisa et al. / International Journal for Parasitology 37 (2007) 1127–1141
It comprises six exons and encodes a 187 amino acid pro-tein with an Mr of 21,316. The predicted protein also showssignificant similarity (72% similarity/50% identity) to thehuman Bet3p homologue (Sacher et al., 2000). PfBet3p isalso related (39% similarity/19% identity) to antigen 41-2which has previously been reported to be exported to theinfected RBC cytoplasm (Knapp et al., 1989).
We expressed PfBet3p as a GST fusion protein in E. coli
(Fig. 6B) and used the purified protein to generate rabbitantiserum. This antiserum recognised the recombinant pro-tein (after cleavage of the GST) as a polypeptide of�22 kDa (Fig. 6C and D) and a protein of the same appar-ent molecular weight in extracts of RBCs infected with 3D7strain parasites (Fig. 6F) but not uninfected RBCs(Fig. 6E).
We used the antiserum to examine the solubility proper-ties of PfBet3p. Plasmodium falciparum-infected RBCswere disrupted using either 2% Triton X-100 or 1 M NaCl,separated into pellet and supernatant, and processed forimmunoblot analysis. PfBet3p was insoluble under bothconditions (Fig. 7B, top panel) while the ER lumen protein,PfERC, was largely released by disruption of the infectedRBCs in the presence of either detergent or high salt con-centrations (Fig. 7B, bottom panel). The poor solubilityof PfBet3p in Triton X-100 is consistent with a previousreport of the solubility properties of Bet3p in yeast andmammalian cells (Sacher et al., 2000). However, yeast
Fig. 6. Expression and purification of PfBet3p. (A–C) The PfBET3
coding sequence was cloned into a glutathione-S-transferase (GST) fusionvector and expressed and purified on a glutathione–Sepharose column.The GST-PfBet3p was cleaved with Factor Xa to release the 22 kDaPfBet3p. Samples were subjected to SDS–PAGE and visualised withCoomassie blue. Lanes: A, total E. coli lysate; B, purified GST-PfBet3p; C,PfBet3p. (D–F) Recombinant PfBet3p (lane D), uninfected erythrocytes(RBCs) (lane E) and infected RBCs (3D7, lane F) were separated by SDS–PAGE and subjected to Western blot analysis with rabbit anti-GST-PfBet3p antiserum. The antibody recognises recombinant PfBet3p and a22 kDa protein in parasitised RBCs (arrow) but not in uninfected RBCs.
and mammalian Bet3p are released during treatment with1 M NaCl (Sacher et al., 2000).
3.5. Bet3p is associated with structures in the parasite
cytoplasm
The intracellular location of PfBet3p was determined byprobing thin smears of parasitised RBCs (3D7 strain) withthe IgG fraction of the anti-PfBet3p antiserum (Fig. 8A).In young parasites, two foci of anti-PfBet3p reactivity areobserved within the parasite cytoplasm. These punctaincrease in number as the parasite matures to mid-tropho-zoite and schizont stages (Fig. 8A). This may representdivision of the Golgi in concert with nuclear division.The anti-PfBet3p reactivity is confined to regions withinthe parasite compartment. There was no labelling of com-partments within the RBC cytoplasm as has been reportedpreviously for COPII proteins and for antigen 41-2 (Knappet al., 1989).
We compared the fluorescence pattern of PfBet3p withthat of an ER lumen protein. As expected, PfERC (redfluorescence) and PfBet3p (green fluorescence) appear tooccupy largely separate locations (Fig. 8B). We also com-pared PfBet3p location with that of a plasmodial homologof the Golgi matrix protein, GRASP, which has recentlybeen characterised using immunolabelling and GFP trans-fection approaches (Struck et al., 2005). We have used ouranti-PfBet3p antisera to compare the locations of PfBet3pand PfGRASP in PfGRASP-GFP transfectants. Some of

Fig. 8. Subcellular location of endogenous PfBet3p in P. falciparum-infected parasites. Smears of erythrocytes infected with 3D7 parent strain (A, B, andD) or PfGRASP-GFP transfectant (C) or PfBet3p-GFP transfectant (E) were probed with rabbit anti-PfBet3p either alone (A) or in combination withmouse anti- PfERC (B) or anti-GFP (C and E) or anti-PfRab6 (D). Binding of primary antibody was detected using fluorescein isothiocyanate (FITC)-conjugated anti-rabbit IgG and Alexa-Fluor 568-conjugated anti-mouse IgG.
A. Adisa et al. / International Journal for Parasitology 37 (2007) 1127–1141 1135
the PfBet3p foci appear to be closely adjacent to thePfGRASP foci (Fig. 8C, arrows) however additionalPfBet3p foci were also observed. We also compared thelocation of PfBet3p with the trans Golgi marker PfRab6.PfRab6 has previously been reported to be located in acompartment that is spatially separate from the cis Golgi(de Castro et al., 1996; Van Wye et al., 1996). We foundthat an anti-PfRab6 antibody recognised different struc-tures to the compartments recognised by anti-PfBet3p(Fig. 8D). These data support the suggestion that the Golgicompartments of P. falciparum are dispersed.
3.6. Bet3p fused to GFP can be expressed in transfected
P. falciparum
We generated a chimera comprising full-length PfBet3plinked to the N-terminus of GFP (PfBet3p-GFP) as suc-cessfully employed in yeast (Wooding and Pelham, 1998).Asynchronous parasite cultures of the parent and transfec-ted lines were subjected to SDS–PAGE and Western blot-ting and probed with mouse antibodies against GFP(Fig. 7A). A band of approximately 50 kDa was observed,which is equivalent to the expected size. No reactivity wasobserved in the untransfected parental line.
3.7. The PfBet3p-GFP chimera is associated with structures
in the parasite cytoplasm
Fluorescence microscopy of live cells expressingPfBet3p-GFP at different stages of growth (Fig. 9) revealeda diffuse labelling of the parasite cytoplasm with somebrighter structures. The immunolabelling data suggestendogenous PfBet3p is largely associated with punctate(presumably organellar) structures, however, there mayalso be a cytoplasmic population. Dual immunofluores-
cence analysis of PfBet3-GFP transfectants with anti-PfBet3p and anti-GFP reveals substantial overlap ofendogenous PfBet3p and the transgene product (Fig. 8E)although PfBet3p-GFP occupies additional sites whichmay represent a cytoplasmic population. Again, therewas no evidence for export of the chimera to the RBCcytoplasm.
We examined the solubility properties of PfBet3p-GFP(Fig. 7B, middle panel). PfBet3p-GFP was more readilyreleased than endogenous PfBet3p, by disruption of para-sites in the presence of either 2% Triton X-100 or 1 MNaCl. This increased solubility suggests that the bindingsites for PfBet3p on Golgi structures may be saturatedin the transfectants, resulting in an increase in thecytoplasm-located population.
To further examine the dynamics of the PfBet3p-GFPchimera, we performed photobleaching studies(Fig. 10A). Upon application of a 50 ms bleach pulse toone of the bright puncta of fluorescence in the parasitecytoplasm, the signal decreased in intensity and showedpartial recovery after about 30 s (Fig. 10A). This is consis-tent with a rapid equilibrium with a cytoplasmic pool and aGolgi-bound pool of PfBet3p-GFP. Repeated bleaching ofthe cytoplasm (Fig. 10D) caused complete loss of fluores-cence from both the cytoplasm and the bright puncta, indi-cating that the two populations are in rapid equilibrium.
3.8. PfBET3 and PfSAR1 do not complement defects of
trafficking defective yeast
Complementation of yeast SAR1 mutants with SAR1
from several other species has been successful (d’Enfertet al., 1992; Veldhuisen et al., 1997; Takeuchi et al.,1998). We therefore attempted to use the PfSAR1 gene tocomplement SAR1 mutant yeast cells, however, viable
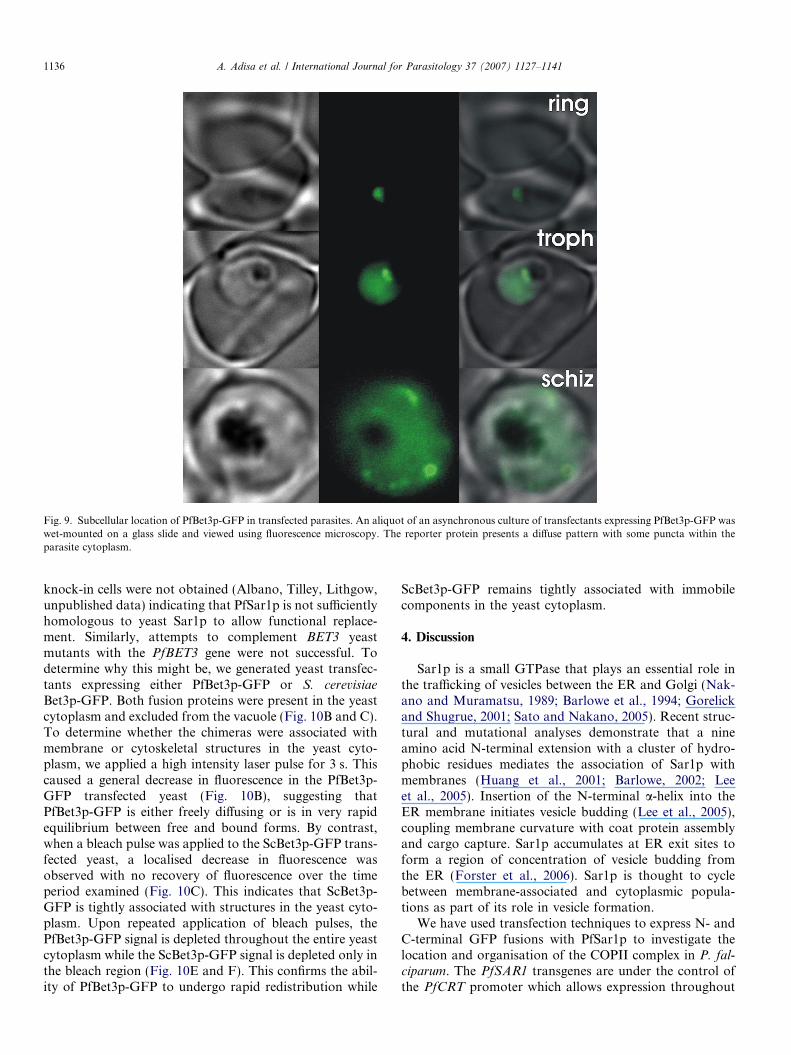
Fig. 9. Subcellular location of PfBet3p-GFP in transfected parasites. An aliquot of an asynchronous culture of transfectants expressing PfBet3p-GFP waswet-mounted on a glass slide and viewed using fluorescence microscopy. The reporter protein presents a diffuse pattern with some puncta within theparasite cytoplasm.
1136 A. Adisa et al. / International Journal for Parasitology 37 (2007) 1127–1141
knock-in cells were not obtained (Albano, Tilley, Lithgow,unpublished data) indicating that PfSar1p is not sufficientlyhomologous to yeast Sar1p to allow functional replace-ment. Similarly, attempts to complement BET3 yeastmutants with the PfBET3 gene were not successful. Todetermine why this might be, we generated yeast transfec-tants expressing either PfBet3p-GFP or S. cerevisiae
Bet3p-GFP. Both fusion proteins were present in the yeastcytoplasm and excluded from the vacuole (Fig. 10B and C).To determine whether the chimeras were associated withmembrane or cytoskeletal structures in the yeast cyto-plasm, we applied a high intensity laser pulse for 3 s. Thiscaused a general decrease in fluorescence in the PfBet3p-GFP transfected yeast (Fig. 10B), suggesting thatPfBet3p-GFP is either freely diffusing or is in very rapidequilibrium between free and bound forms. By contrast,when a bleach pulse was applied to the ScBet3p-GFP trans-fected yeast, a localised decrease in fluorescence wasobserved with no recovery of fluorescence over the timeperiod examined (Fig. 10C). This indicates that ScBet3p-GFP is tightly associated with structures in the yeast cyto-plasm. Upon repeated application of bleach pulses, thePfBet3p-GFP signal is depleted throughout the entire yeastcytoplasm while the ScBet3p-GFP signal is depleted only inthe bleach region (Fig. 10E and F). This confirms the abil-ity of PfBet3p-GFP to undergo rapid redistribution while
ScBet3p-GFP remains tightly associated with immobilecomponents in the yeast cytoplasm.
4. Discussion
Sar1p is a small GTPase that plays an essential role inthe trafficking of vesicles between the ER and Golgi (Nak-ano and Muramatsu, 1989; Barlowe et al., 1994; Gorelickand Shugrue, 2001; Sato and Nakano, 2005). Recent struc-tural and mutational analyses demonstrate that a nineamino acid N-terminal extension with a cluster of hydro-phobic residues mediates the association of Sar1p withmembranes (Huang et al., 2001; Barlowe, 2002; Leeet al., 2005). Insertion of the N-terminal a-helix into theER membrane initiates vesicle budding (Lee et al., 2005),coupling membrane curvature with coat protein assemblyand cargo capture. Sar1p accumulates at ER exit sites toform a region of concentration of vesicle budding fromthe ER (Forster et al., 2006). Sar1p is thought to cyclebetween membrane-associated and cytoplasmic popula-tions as part of its role in vesicle formation.
We have used transfection techniques to express N- andC-terminal GFP fusions with PfSar1p to investigate thelocation and organisation of the COPII complex in P. fal-ciparum. The PfSAR1 transgenes are under the control ofthe PfCRT promoter which allows expression throughout
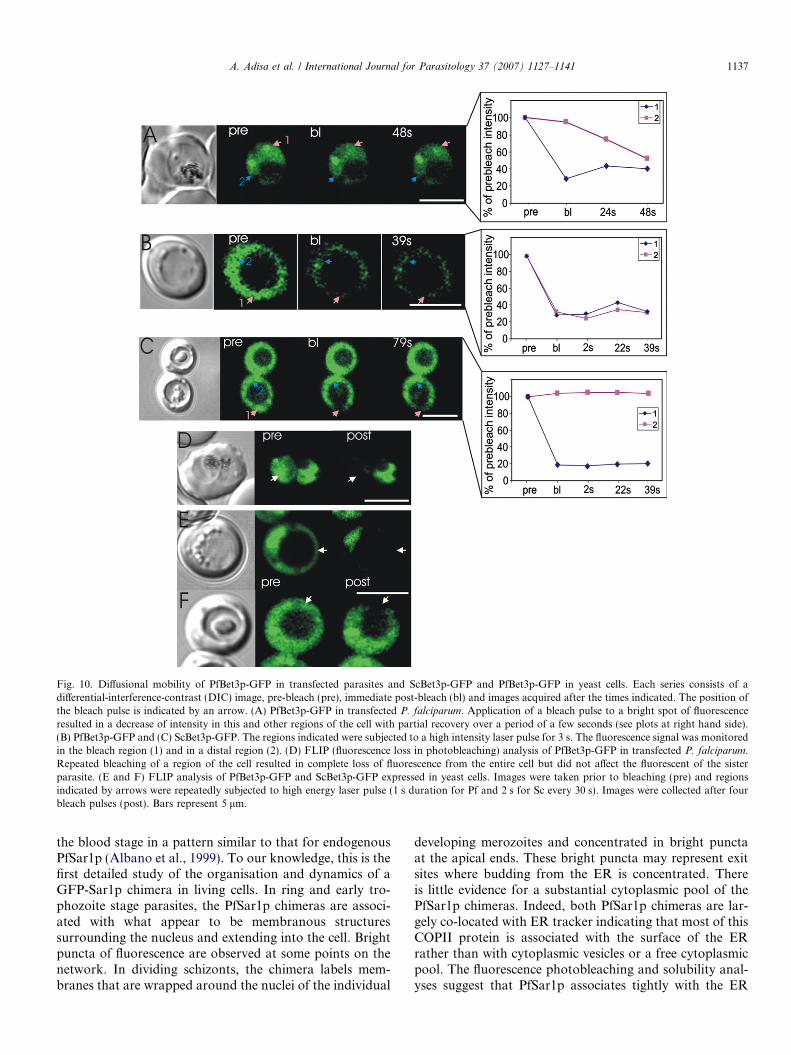
Fig. 10. Diffusional mobility of PfBet3p-GFP in transfected parasites and ScBet3p-GFP and PfBet3p-GFP in yeast cells. Each series consists of adifferential-interference-contrast (DIC) image, pre-bleach (pre), immediate post-bleach (bl) and images acquired after the times indicated. The position ofthe bleach pulse is indicated by an arrow. (A) PfBet3p-GFP in transfected P. falciparum. Application of a bleach pulse to a bright spot of fluorescenceresulted in a decrease of intensity in this and other regions of the cell with partial recovery over a period of a few seconds (see plots at right hand side).(B) PfBet3p-GFP and (C) ScBet3p-GFP. The regions indicated were subjected to a high intensity laser pulse for 3 s. The fluorescence signal was monitoredin the bleach region (1) and in a distal region (2). (D) FLIP (fluorescence loss in photobleaching) analysis of PfBet3p-GFP in transfected P. falciparum.Repeated bleaching of a region of the cell resulted in complete loss of fluorescence from the entire cell but did not affect the fluorescent of the sisterparasite. (E and F) FLIP analysis of PfBet3p-GFP and ScBet3p-GFP expressed in yeast cells. Images were taken prior to bleaching (pre) and regionsindicated by arrows were repeatedly subjected to high energy laser pulse (1 s duration for Pf and 2 s for Sc every 30 s). Images were collected after fourbleach pulses (post). Bars represent 5 lm.
A. Adisa et al. / International Journal for Parasitology 37 (2007) 1127–1141 1137
the blood stage in a pattern similar to that for endogenousPfSar1p (Albano et al., 1999). To our knowledge, this is thefirst detailed study of the organisation and dynamics of aGFP-Sar1p chimera in living cells. In ring and early tro-phozoite stage parasites, the PfSar1p chimeras are associ-ated with what appear to be membranous structuressurrounding the nucleus and extending into the cell. Brightpuncta of fluorescence are observed at some points on thenetwork. In dividing schizonts, the chimera labels mem-branes that are wrapped around the nuclei of the individual
developing merozoites and concentrated in bright punctaat the apical ends. These bright puncta may represent exitsites where budding from the ER is concentrated. Thereis little evidence for a substantial cytoplasmic pool of thePfSar1p chimeras. Indeed, both PfSar1p chimeras are lar-gely co-located with ER tracker indicating that most of thisCOPII protein is associated with the surface of the ERrather than with cytoplasmic vesicles or a free cytoplasmicpool. The fluorescence photobleaching and solubility anal-yses suggest that PfSar1p associates tightly with the ER

1138 A. Adisa et al. / International Journal for Parasitology 37 (2007) 1127–1141
surface but undergoes rapid diffusion within the plane ofthe membrane. Previous studies of Sar1p in yeast also showcompartmentalisation to the ER and nuclear membranes(Nishikawa and Nakano, 1991; Bednarek et al., 1995).
Recently transfected parasites were used to examine thecellular location of a GFP chimera of an ER lumen pro-tein, PfBiP (van Dooren et al., 2005). Using PfBiP-GFP,the ER was visualised as a ring around the nucleus withtwo small protrusions that develop into an extended retic-ular network as the parasite enlarges. These data are con-sistent with our images of the PfSar1p chimeras. Ourdata are also consistent with EM evidence for a concentra-tion of secretory activity at the anterior surface of thenuclear membrane in segmenting schizonts (Ward et al.,1997; Bannister et al., 2000b,a).
The PfSar1p chimeras examined were not trafficked tothe host cell cytoplasm. This is in contrast to previousimmunofluorescence data for COPII proteins (Albanoet al., 1999; Adisa et al., 2001; Taraschi et al., 2003; Wick-ert et al., 2003). We considered the possibility that the GFPfusion protein might interfere with export of the PfSar1pchimeras, however, the fact that the chimeras clearly asso-ciate with the ER and the fact that appending GFP ateither the N- or C-terminus or including an Gly5 linkerregion between PfSar1p and the GFP reporter did not alterthe cellular distribution argues against this possibility.While it remains possible that the presence of the GFPtag prevents export to the host cell cytoplasm, the data sug-gests that the previous reports of export of COPII proteinsmay be due to cross-reactivity of the antibodies againstcomponents of the Maurer’s clefts. Indeed there are manyreports in the literature of cross-reactivity of antibodies toP. falciparum proteins (Mattei et al., 1989; Saul et al., 1989;Mercereau-Puijalon et al., 1992; Gruner et al., 2001; Spiel-mann et al., 2006). This is thought to be due to the highlyrepetitive and immunogenic nature of many parasite pro-teins which enhances cross-reactive binding of antibodies(Anders et al., 1987).
We have also determined the intracellular location ofanother component of the protein trafficking machineryof P. falciparum. Bet3p is a 22 kDa component of theTRAPP complex. TRAPP comprises two highly relatedmultiprotein complexes, TRAPP I and TRAPP II (Barrow-man et al., 2000; Guo et al., 2000; Sacher et al., 2000;Sacher et al., 2001; Turnbull et al., 2005). The TRAPP Icomplex has seven subunits (including Bet3p and therelated Trs31p and Trs33p) and directs docking of ER-derived vesicles onto the cis Golgi. The TRAPP II complexcontains three extra subunits and functions after transportto the Golgi (Sacher et al., 1998, 2001; Guo et al., 2000).
Recent crystallographic structures of mammalian Bet3pshow that the protein forms a homodimer (or a heterodi-mer with Trs33p) and presents a flat, basic surface that isthought to dock on to negatively charged lipids on theGolgi surface (Kim et al., 2005; Turnbull et al., 2005).Bet3p has a distinct hydrophobic channel that is thoughtto provide specificity by interacting with a Golgi-located
hydrophobic moiety. A protein referred to as antigen 41-2, which is related to plasmodial PfBet3p, has beenreported (using an antibody-based analysis) to be exportedto the host cell cytoplasm (Knapp et al., 1989). In view ofour findings for PfSar1p, we analysed the location ofPfBet3p using immunolabelling and transfectionapproaches.
Antiserum against PfBet3p labelled two punctate struc-tures in ring stage parasites that increased in number as theparasites matured. Yeast Bet3p is associated with the cis
Golgi complex (Sacher et al., 1998; Barrowman et al.,2000), but TRAPP can stimulate nucleotide exchange ona GTPase pair required for exit from the trans Golgi, sug-gesting that it may also associate with the trans Golgi(Jones et al., 2000). Moreover, as Bet3p is a member of aperipheral membrane protein complex, the bound popula-tion is expected to be in equilibrium with a cytoplasmicpopulation. In yeast, the bound population of Bet3pappears to predominate (Sacher et al., 2000) while in mam-malian cells the majority of Bet3p is present in the cyto-plasm (Loh et al., 2005). Our immunofluorescence andsolubility data suggest that PfBet3p is largely present as amembrane- or cytoskeleton-bound pool.
Transgenic parasites expressing a GFP chimera of theGolgi matrix protein, PfGRASP, have recently been gener-ated (Struck et al., 2005). PfGRASP colocalises withERD2, suggesting that it is a cis Golgi marker (Strucket al., 2005). PfBet3p and PfGRASP-GFP are both foundin punctate compartments that increase in number as theparasite matures. Some of the PfBet3p-containing com-partments are closely adjacent to the PfGRASP compart-ments, indicating that PfBet3p marks a compartment thatis closely associated with the cis Golgi. However, PfBet3pis also present in additional compartments which mayrepresent cytoplasmic TRAPP complexes. There was noevidence for export of PfBet3p beyond the confines of theparasite plasma membrane. Preliminary studies with trans-fectants expressing a GFP fusion of another TRAPP com-plex protein, PfTrs31p, indicate that this chimera is alsorestricted to the parasite cytoplasm (Frankland, Maier,Cowman, Tilley, unpublished data). Taken together, thesedata argue against the export of a cannonical traffickingpathway to the host cell cytoplasm.
To confirm our antibody-based observations, we gener-ated transfected malaria parasites expressing PfBet3p-GFP. Again, PfBet3p-GFP was confined to regions withinthe parasite cytoplasm. The chimera was associated withpunctate structures in the parasite cytoplasm that are pre-sumably the same structures occupied by endogenousPfBet3p. However, a substantial amount of the chimeraalso appeared to be present free in the parasite cytoplasm.
Photobleaching analysis showed that the Golgi-boundand cytoplasmic pools of PfBet3p-GFP are in rapid equi-librium. For comparison, we have examined the dynamicsof PfBet3p-GFP and ScBet3p-GFP in transfected yeast. Inboth transfectants, the chimera was expressed in the yeastcytoplasm. ScBet3p-GFP appeared to be tightly associated

A. Adisa et al. / International Journal for Parasitology 37 (2007) 1127–1141 1139
with immobilised structures. These may represent cytoskel-etal elements as ScBet3p has previously been shown to beresistant to extraction by Triton X-100 (Sacher et al.,2000). By contrast, PfBet3p-GFP appeared to be rapidlymobile within the yeast cytoplasm, which is consistent withthe failure of PfBet3p-GFP to complement the loss ofScBet3p.
Taken together, the data provide information about theorganisation of the endomembrane system of P. falcipa-
rum. As the ring stage parasite develops, the ER expandsfrom the nuclear membrane to form a reticulum through-out the cell. Puncta of the PfSar1p chimeras along this net-work likely represent ER exit sites. As the parasite divides,the ER forms around individual dividing nuclei butremains connected to the ER within other developingmerozoites until very late in schizogony. Eventually, themerozoites undergo cytokinesis and the ER systems areseparated. The Golgi of P. falciparum is initially presentas one or two foci that multiply as the parasite matures.The data suggest that different Golgi compartments arespatially dispersed. However, given the problems usingantibodies in P. falciparum, dual labelling of different Golgisub-compartments with different fluorescent protein tags isneeded to confirm this suggestion. It is interesting to notethat a conventional Golgi and a single ER exit site associ-ated with the nuclear envelope are present in the closelyrelated parasite, Toxoplasma gondii (He, 2007).
If parasite-derived classical trafficking machinery is notexported to the host cell cytoplasm, how does the parasitetransport proteins across the host RBC cytoplasm? It ispossible that the parasite exports a completely novel reper-toire of proteins for mediating vesicle budding and fusion.Alternatively, trafficking of proteins across the infectedRBC cytoplasm may occur via alternative pathways.Indeed, we have recently provided evidence that Maurer’sclefts appear to bud from the parasitophorous vacuolemembrane as pre-formed structures and then take up resi-dence at the cell periphery (Spycher et al., 2006). MoreoverPfEMP1, an integral membrane protein destined for theinfected RBC membrane, may transit the infected RBCcytoplasm as a soluble chaperoned complex (Knuepferet al., 2005; Papakrivos et al., 2005). Similarly, we haveshown that inhibition of GTP-dependent events in theinfected RBC cytoplasm does not prevent trafficking ofPfEMP1 across this compartment (Frankland et al.,2006). Clearly further studies are needed to dissect the com-ponents of this unusual trafficking machinery and to deter-mine if these pathways are related to the alternativetrafficking pathways employed in other organisms (Nickel,2003). The novelty of the extracellular protein traffickingpathways of P. falciparum might provide suitable targetsfor the development of new antimalarial strategies.
Acknowledgments
This work was supported by the National Health andMedical Research Council, Australia. S. Frankland was
also supported by the Michael J. Osborne Centre, Institutefor Advanced Study Postgraduate Writing-up Award, LaTrobe University. We thank Sam Deed, Frank Albanoand Dr Cornelia Spycher, La Trobe University, for techni-cal support, for performing the Sar1p complementationstudies, and for donating the transfectant expressing GFPalone. PfGRASP-GFP transfectants were kindly donatedby Dr Nicole Struck and Dr Tim Gliberger, Berhard NochtInstitute, Hamburg.
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at doi:10.1016/j.ijpara.2007.02.009.
References
Adisa, A., Albano, F.R., Reeder, J., Foley, M., Tilley, L., 2001. Evidencefor a role for a Plasmodium falciparum homologue of Sec31p in theexport of proteins to the surface of malaria parasite-infected erythro-cytes. J. Cell Sci. 114, 3377–3386.
Adisa, A., Rug, M., Foley, M., Tilley, L., 2002. Characterisation of delta-COP homologue in the malaria parasite, Plasmodium falciparum. Mol.Biochem. Parasitol. 123, 11–21.
Adisa, A., Rug, M., Klonis, N., Foley, M., Cowman, A.F., Tilley, L.,2003. The signal sequence of exported protein-1 directs the greenfluorescent protein to the parasitophorous vacuole of transfectedmalaria parasites. J. Biol. Chem. 278, 6532–6542.
Albano, F.R., Berman, A., La Greca, N., Hibbs, A.R., Wickham, M.,Foley, M., Tilley, L., 1999. A homologue of Sar1p localises to a noveltrafficking pathway in malaria-infected erythrocytes. Eur. J. Cell Biol.78, 453–462.
Anders, R.F., Barzaga, N., Shi, P.T., Scanlon, D.B., Brown, L.E.,Thomas, L.M., Brown, G.V., Stahl, H.D., Coppel, R.L., Kemp, D.J.,1987. Repetitive sequences in malaria antigens, Molecular Strategies ofParasitic Invasion, Alan R. Liss, Inc., pp. 333–342.
Bannister, L.H., Hopkins, J.M., Fowler, R.E., Krishna, S., Mitchell,G.H., 2000a. A brief illustrated guide to the ultrastructure ofPlasmodium falciparum asexual blood stages. Parasitol. Today 16,427–433.
Bannister, L.H., Hopkins, J.M., Fowler, R.E., Krishna, S., Mitchell,G.H., 2000b. Ultrastructure of rhoptry development in Plasmodium
falciparum erythrocytic schizonts. Parasitology 121 (Pt. 3), 273–287.Barlowe, C., 2002. COPII-dependent transport from the endoplasmic
reticulum. Curr. Opin. Cell Biol. 14, 417–422.Barlowe, C., Orci, L., Yeung, T., Hosobuchi, M., Hamamoto, S., Salama,
N., Rexach, M.F., Ravazzola, M., Amherdt, M., Schekman, R., 1994.COPII: a membrane coat formed by Sec proteins that drive vesiclebudding from the endoplasmic reticulum. Cell 77, 895–907.
Barrowman, J., Sacher, M., Ferro-Novick, S., 2000. TRAPP stablyassociates with the Golgi and is required for vesicle docking. EMBO J.19, 862–869.
Bednarek, S.Y., Ravazzola, M., Hosobuchi, M., Amherdt, M., Perrelet,A., Schekman, R., Orci, L., 1995. COPI- and COPII-coated vesiclesbud directly from the endoplasmic reticulum in yeast. Cell 83,1183–1196.
Brachmann, C.B., Davies, A., Cost, G.J., Caputo, E., Li, J., Hieter, P.,Boeke, J.D., 1998. Designer deletion strains derived from Saccharo-
myces cerevisiae S288C: a useful set of strains and plasmids for PCR-mediated gene disruption and other applications. Yeast 14, 115–132.
Cooke, B.M., Lingelbach, K., Bannister, L., Tilley, L., 2004. Proteintrafficking in Plasmodium falciparum-infected red blood cells. TrendsParasitol. 20, 581–589.

1140 A. Adisa et al. / International Journal for Parasitology 37 (2007) 1127–1141
Crabb, B.S., Rug, M., Gilberger, T.W., Thompson, J.K., Triglia, T.,Maier, A.G., Cowman, A.F., 2004. Transfection of the human malariaparasite Plasmodium falciparum. Methods Mol. Biol. 270, 263–276.
de Castro, F.A., Ward, G.E., Jambou, R., Attal, G., Mayau, V.,Jaureguiberry, G., Braun-Breton, C., Chakrabarti, D., Langsley, G.,1996. Identification of a family of Rab G-proteins in Plasmodium
falciparum and a detailed characterisation of pfrab6. Mol. Biochem.Parasitol. 80, 77–88.
Deitsch, K.W., Wellems, T.E., 1996. Membrane modifications in eryth-rocytes parasitized by Plasmodium falciparum. Mol. Biochem. Paras-itol. 76, 1–10.
d’Enfert, C., Gensse, M., Gaillardin, C., 1992. Fission yeast and aplant have functional homologues of the Sar1 and Sec12 proteinsinvolved in ER to Golgi traffic in budding yeast. EMBO J. 11,4205–4211.
Elmendorf, H.G., Haldar, K., 1993. Identification and localization ofErd2 in the malaria parasite Plasmodium falciparum – separation fromsites of sphingomyelin synthesis and implications for organization ofthe Golgi. EMBO J. 12, 4763–4773.
Fidock, D.A., Wellems, T.E., 1997. Transformation with human dihy-drofolate reductase renders malaria parasites insensitive to WR99210but does not affect the intrinsic activity of proguanil. Proc. Natl. Acad.Sci. USA 94, 10931–10936.
Forster, R., Weiss, M., Zimmermann, T., Reynaud, E.G.,Verissimo, F., Stephens, D.J., Pepperkok, R., 2006. Secretory cargoregulates the turnover of COPII subunits at single ER exit sites. Curr.Biol. 16, 173–179.
Foth, B.J., Ralph, S.A., Tonkin, C.J., Struck, N.S., Fraunholz, M., Roos,D.S., Cowman, A.F., McFadden, G.I., 2003. Dissecting apicoplasttargeting in the malaria parasite Plasmodium falciparum. Science 299,705–708.
Frankland, S., Adisa, A., Horrocks, P., Taraschi, T.F., Schneider, T.,Elliott, S.R., Rogerson, S.J., Knuepfer, E., Cowman, A.F., Newbold,C.I., Tilley, L., 2006. Delivery of the malaria virulence proteinPfEMP1 to the erythrocyte surface requires cholesterol-rich domains.Eukaryot. Cell 5, 849–860.
Gardner, M.J., Hall, N., Fung, E., White, O., Berriman, M., Hyman,R.W., Carlton, J.M., Pain, A., Nelson, K.E., Bowman, S., Paulsen,I.T., James, K., Eisen, J.A., Rutherford, K., Salzberg, S.L., Craig, A.,Kyes, S., Chan, M.S., Nene, V., Shallom, S.J., Suh, B., Peterson, J.,Angiuoli, S., Pertea, M., Allen, J., Selengut, J., Haft, D., Mather,M.W., Vaidya, A.B., Martin, D.M., Fairlamb, A.H., Fraunholz, M.J.,Roos, D.S., Ralph, S.A., McFadden, G.I., Cummings, L.M., Subra-manian, G.M., Mungall, C., Venter, J.C., Carucci, D.J., Hoffman,S.L., Newbold, C., Davis, R.W., Fraser, C.M., Barrell, B., 2002.Genome sequence of the human malaria parasite Plasmodium falcipa-
rum. Nature 419, 498–511.Gorelick, F.S., Shugrue, C., 2001. Exiting the endoplasmic reticulum.
Mol. Cell. Endocrinol. 177, 13–18.Gruner, A.C., Brahimi, K., Eling, W., Konings, R., Meis, J., Aikawa, M.,
Daubersies, P., Guerin-Marchand, C., Mellouk, S., Snounou, G.,Druilhe, P., 2001. The Plasmodium falciparum knob-associatedPfEMP3 antigen is also expressed at pre-erythrocytic stages andinduces antibodies which inhibit sporozoite invasion. Mol. Biochem.Parasitol. 112, 253–261.
Guo, W., Sacher, M., Barrowman, J., Ferro-Novick, S., Novick, P., 2000.Protein complexes in transport vesicle targeting. Trends Cell Biol. 10,251–255.
Hayashi, M., Taniguchi, S., Ishizuka, Y., Kim, H.S., Wataya, Y.,Yamamoto, A., Moriyama, Y., 2001. A homologue of N-ethylmalei-mide-sensitive factor in the malaria parasite Plasmodium falciparum isexported and localized in vesicular structures in the cytoplasm ofinfected erythrocytes in the brefeldin A-sensitive pathway. J. Biol.Chem. 276, 15249–15255.
He, C.Y., 2007. Golgi biogenesis in simple eukaryotes. Cell Microbiol. 9,566–572.
Huang, M., Weissman, J.T., Beraud-Dufour, S., Luan, P., Wang, C.,Chen, W., Aridor, M., Wilson, I.A., Balch, W.E., 2001. Crystal
structure of Sar1-GDP at 1.7 A resolution and the role of the NH2terminus in ER export. J. Cell Biol. 155, 937–948.
Jones, S., Newman, C., Liu, F., Segev, N., 2000. The TRAPP complex is anucleotide exchanger for Ypt1 and Ypt31/32. Mol. Biol. Cell 11, 4403–4411.
Kim, Y.G., Sohn, E.J., Seo, J., Lee, K.J., Lee, H.S., Hwang, I., Whiteway,M., Sacher, M., Oh, B.H., 2005. Crystal structure of bet3 reveals anovel mechanism for Golgi localization of tethering factor TRAPP.Nat. Struct. Mol. Biol. 12, 38–45.
Klemba, M., Beatty, W., Gluzman, I., Goldberg, D.E., 2004. Traffickingof plasmepsin II to the food vacuole of the malaria parasitePlasmodium falciparum. J. Cell Biol. 164, 47–56.
Klonis, N., Rug, M., Harper, I., Wickham, M., Cowman, A., Tilley, L.,2002. Fluorescence photobleaching analysis for the study of cellulardynamics. Eur. Biophys. J. 31, 36–51.
Knapp, B., Hundt, E., Kupper, H.A., 1989. A new blood stage antigen ofPlasmodium falciparum transported to the erythrocyte surface. Mol.Biochem. Parasitol. 37, 47–56.
Knuepfer, E., Rug, M., Klonis, N., Tilley, L., Cowman, A.F., 2005.Trafficking of the major virulence factor to the surface of transfectedP. falciparum-infected erythrocytes. Blood 105, 4078–4087.
Kumar, N., Syin, C.A., Carter, R., Quakyi, I., Miller, L.H., 1988.Plasmodium falciparum gene encoding a protein similar to the 78-kDarat glucose-regulated stress protein. Proc. Natl. Acad. Sci. USA 85,6277–6281.
Kyes, S., Horrocks, P., Newbold, C., 2001. Antigenic variation atthe infected red cell surface in malaria. Annu. Rev. Microbiol. 55,673–707.
La Greca, N., Hibbs, A.R., Riffkin, C., Foley, M., Tilley, L., 1997.Identification of an endoplasmic reticulum resident protein withmultiple EF-hand motifs in asexual stage of Plasmodium falciparum.Mol. Biochem. Parasitol. 89, 283–293.
Lanzer, M., Wickert, H., Krohne, G., Vincensini, L., Braun Breton, C.,2006. Maurer’s clefts: a novel multi-functional organelle in thecytoplasm of Plasmodium falciparum-infected erythrocytes. Int. J.Parasitol. 36, 23–36.
Lee, M.C., Orci, L., Hamamoto, S., Futai, E., Ravazzola, M., Schekman,R., 2005. Sar1p N-terminal helix initiates membrane curvature andcompletes the fission of a COPII vesicle. Cell 122, 605–617.
Li, W., Keller, G.A., Haldar, K., 1995. Recognition of a 170 kDa proteinin mammalian Golgi complexes by an antibody against malarialintraerythrocytic lamellae. Tissue Cell 27, 355–367.
Loh, E., Peter, F., Subramaniam, V.N., Hong, W., 2005. Mammalian Bet3functions as a cytosolic factor participating in transport from the ERto the Golgi apparatus. J. Cell Sci. 118, 1209–1222.
Margos, G., Bannister, L.H., Dluzewski, A.R., Hopkins, J., Wil-liams, I.T., Mitchell, G.H., 2004. Correlation of structuraldevelopment and differential expression of invasion-related mole-cules in schizonts of Plasmodium falciparum. Parasitology 129,273–287.
Marti, M., Good, R.T., Rug, M., Knuepfer, E., Cowman, A.F., 2004.Targeting malaria virulence and remodeling proteins to the hosterythrocyte. Science 306, 1930–1933.
Marti, M., Rug, M., Baum, J., Tilley, L., Cowman, A.F., 2005. Signalmediated export of proteins from the malaria parasite to the hosterythrocyte. J. Cell Biol. 171, 587–592.
Mattei, D., Berzins, K., Wahlgren, M., Udomsangpetch, R., Perlmann, P.,Griesser, H.W., Scherf, A., Muller-Hill, B., Bonnefoy, S., Guillotte,M., et al., 1989. Cross-reactive antigenic determinants present ondifferent Plasmodium falciparum blood-stage antigens. Parasite Immu-nol. 11, 15–29.
Mercereau-Puijalon, O., Guillotte, M., Doyen, N., 1992. Cross-reaction ofantibodies to the nine-amino acid repeats of Plasmodium falciparum
antigen 11.1 with human serum albumin. Eur. J. Immunol. 22,381–385.
Nakano, A., Muramatsu, M., 1989. A novel GTP-binding protein, Sar1p,is involved in transport from the endoplasmic reticulum to the Golgiapparatus. J. Cell Biol. 109, 2677–2691.

A. Adisa et al. / International Journal for Parasitology 37 (2007) 1127–1141 1141
Nickel, W., 2003. The mystery of nonclassical protein secretion. A currentview on cargo proteins and potential export routes. Eur. J. Biochem.270, 2109–2119.
Niedenthal, R.K., Riles, L., Johnston, M., Hegemann, J.H., 1996. Greenfluorescent protein as a marker for gene expression and subcellularlocalization in budding yeast. Yeast 12, 773–786.
Nishikawa, S., Nakano, A., 1991. The GTP-binding Sar1 protein islocalized to the early compartment of the yeast secretory pathway.Biochim. Biophys. Acta 1093, 135–143.
Papakrivos, J., Newbold, C.I., Lingelbach, K., 2005. A potential novelmechanism for the insertion of a membrane protein revealed by abiochemical analysis of the Plasmodium falciparum cytoadherencemolecule PfEMP-1. Mol. Microbiol. 55, 1272–1284.
Przyborski, J.M., Wickert, H., Krohne, G., Lanzer, M., 2003. Maurer’sclefts – a novel secretory organelle? Mol. Biochem. Parasitol. 132,17–26.
Rawlings, D.J., Kaslow, D.C., 1992. A novel 40-kDa membrane-associ-ated EF-hand calcium-binding protein in Plasmodium falciparum.J. Biol. Chem. 267, 3976–3982.
Raynes, K., Foley, M., Tilley, L., Deady, L.W., 1996. Novel bisquinolineantimalarials. Synthesis, antimalarial activity, and inhibition of haempolymerisation. Biochem. Pharmacol. 52, 551–559.
Rowe, J.A., Kyes, S.A., 2004. The role of Plasmodium falciparum vargenes in malaria in pregnancy. Mol. Microbiol. 53, 1011–1019.
Rug, M., Wickham, M.E., Foley, M., Cowman, A.F., Tilley, L., 2004.Correct promoter control is needed for trafficking of the ring-infectederythrocyte surface antigen to the host cytosol in transfected malariaparasites. Infect. Immun. 72, 6095–6105.
Sacher, M., Jiang, Y., Barrowman, J., Scarpa, A., Burston, J., Zhang, L.,Schieltz, D., Yates, J.R., Abeliovich, H., Ferronovick, S., 1998. Trapp,a highly conserved novel complex on the cis-golgi that mediates vesicledocking and fusion. EMBO J. 17, 2494–2503.
Sacher, M., Barrowman, J., Schieltz, D., Yates 3rd, J.R., Ferro-Novick,S., 2000. Identification and characterization of five new subunits ofTRAPP. Eur. J. Cell Biol. 79, 71–80.
Sacher, M., Barrowman, J., Wang, W., Horecka, J., Zhang, Y.Y.,Pypaert, M., Ferro-Novick, S., 2001. TRAPP I implicated in thespecificity of tethering in ER-to-Golgi transport. Mol. Cell 7,433–442.
Sato, K., Nakano, A., 2005. Dissection of COPII subunit-cargo assemblyand disassembly kinetics during Sar1p-GTP hydrolysis. Nat. Struct.Mol. Biol. 12, 167–174.
Saul, A., Lord, R., Jones, G., Geysen, H.M., Gale, J., Mollard, R., 1989.Cross-reactivity of antibody against an epitope of the Plasmodium
falciparum second merozoite surface antigen. Parasite Immunol. 11,593–601.
Spielmann, T., Hawthorne, P.L., Dixon, M.W., Hannemann, M., Klotz,K., Kemp, D.J., Klonis, N., Tilley, L., Trenholme, K.R., Gardiner,D.L., 2006. A cluster of ring stage-specific genes linked to a locusImplicated in cytoadherence in Plasmodium falciparum codes forPEXEL-negative and PEXEL-positive proteins exported into the hostcell. Mol. Biol. Cell.
Spycher, C., Klonis, N., Spielmann, T., Kump, E., Steiger, S., Tilley, L.,Beck, H.P., 2003. MAHRP-1, a novel Plasmodium falciparum histi-dine-rich protein, binds ferriprotoporphyrin IX and localizes to theMaurer’s clefts. J. Biol. Chem. 278, 35373–35383.
Spycher, C., Rug, M., Klonis, N., Ferguson, D.J., Cowman, A.F., Beck,H.P., Tilley, L., 2006. Genesis of and trafficking to the Maurer’s cleftsof Plasmodium falciparum-infected erythrocytes. Mol. Cell. Biol. 26,4074–4085.
Struck, N.S., de Souza Dias, S., Langer, C., Marti, M., Pearce, J.A.,Cowman, A.F., Gilberger, T.W., 2005. Re-defining the Golgi complexin Plasmodium falciparum using the novel Golgi marker PfGRASP. J.Cell Sci. 118, 5603–5613.
Takeuchi, M., Tada, M., Saito, C., Yashiroda, H., Nakano, A., 1998.Isolation of a tobacco cDNA encoding Sar1 GTPase and analysis of itsdominant mutations in vesicular traffic using a yeast complementationsystem. Plant Cell Physiol. 39, 590–599.
Tang, B.L., Wang, Y., Ong, Y.S., Hong, W., 2005. COPII and exit fromthe endoplasmic reticulum. Biochim. Biophys. Acta 1744, 293–303.
Taraschi, T.F., O’Donnell, M.E., Martinez, S., Schneider, T., Trelka, D.,Fowler, V.M., Tilley, L., Moriyama, Y., 2003. Generation of anerythrocyte vesicle transport system by Plasmodium falciparum malariaparasites. Blood 102, 3420–3426.
Tonkin, C.J., van Dooren, G.G., Spurck, T.P., Struck, N.S., Good, R.T.,Handman, E., Cowman, A.F., McFadden, G.I., 2004. Localization oforganellar proteins in Plasmodium falciparum using a novel set oftransfection vectors and a new immunofluorescence fixation method.Mol. Biochem. Parasitol. 137, 13–21.
Turnbull, A.P., Kummel, D., Prinz, B., Holz, C., Schultchen, J., Lang, C.,Niesen, F.H., Hofmann, K.P., Delbruck, H., Behlke, J., Muller, E.C.,Jarosch, E., Sommer, T., Heinemann, U., 2005. Structure of palmi-toylated BET3: insights into TRAPP complex assembly and membranelocalization. EMBO J. 24, 875–884.
van Dooren, G.G., Marti, M., Tonkin, C.J., Stimmler, L.M., Cowman,A.F., McFadden, G.I., 2005. Development of the endoplasmicreticulum, mitochondrion and apicoplast during the asexual life cycleof Plasmodium falciparum. Mol. Microbiol. 57, 405–419.
Van Wye, J., Ghori, N., Webster, P., Mitschler, R.R., Elmendorf, H.G.,Haldar, K., 1996. Identification and localization of Rab6, separationof Rab6 from Erd2 and implications for an unstacked Golgi, inPlasmodium falciparum. Mol. Biochem. Parasitol. 83, 107–120.
Veldhuisen, G., Saloheimo, M., Fiers, M.A., Punt, P.J., Contreras, R.,Penttila, M., van den Hondel, C.A., 1997. Isolation and analysis offunctional homologues of the secretion-related SAR1 gene of Saccha-
romyces cerevisiae from Aspergillus niger and Trichoderma reesei. Mol.Gen. Genet 256, 446–455.
Waller, R.F., Reed, M.B., Cowman, A.F., McFadden, G.I., 2000. Proteintrafficking to the plastid of Plasmodium falciparum is via the secretorypathway. EMBO J. 19, 1794–1802.
Ward, G.E., Tilney, L.G., Langsley, G., 1997. Rab GTPases and theunusual secretory pathway of Plasmodium. Parasitol. Today 3, 57–62.
Wickert, H., Rohrbach, P., Scherer, S.J., Krohne, G., Lanzer, M., 2003. Aputative Sec23 homologue of Plasmodium falciparum is located inMaurer’s clefts. Mol. Biochem. Parasitol. 129, 209–213.
Wickham, M.E., Rug, M., Ralph, S.A., Klonis, N., McFadden, G.I.,Tilley, L., Cowman, A.F., 2001. Trafficking and assembly of thecytoadherence complex in Plasmodium falciparum-infected humanerythrocytes. EMBO J. 20, 5636–5649.
Wooding, S., Pelham, H.R.B., 1998. The dynamics of golgi proteintraffic visualized in living yeast cells. Mol. Biol. Cell 9, 2667–2680.




















