
Reorganization of Microtubules in Endosperm Cells and Cell ...
Upload
independentCategory
view
0download
0
# 2008 The Authors
Journal compilation # 2008 Blackwell Publishing Ltd
doi: 10.1111/j.1600-0854.2008.00725.xTraffic 2008; 9: 833–847Blackwell Munksgaard
Single Vesicle Analysis of Endocytic Fission onMicrotubules In Vitro
John W. Murray1,2,†,*, Souvik Sarkar1,2,† and
Allan W. Wolkoff1,2
1Marion Bessin Liver Research Center and Departmentof Medicine, and Division of Hepatology, Albert EinsteinCollege of Medicine, 1300 Morris Park Avenue,Bronx, NY 10461, USA2Department of Anatomy and Structural Biology,Albert Einstein College of Medicine, 1300 MorrisPark Avenue, Bronx, NY 10461, USA*Corresponding author: John W. Murray,[email protected]†These authors contributed equally to this work.
Following endocytosis, internalized molecules are found
within intracellular vesicles and tubules that move along
the cytoskeleton and undergo fission, as demonstrated
here using primary cultured rat hepatocytes. Although
the use of depolymerizing drugs has shown that the
cytoskeleton is not required to segregate endocytic
protein, many studies suggest that the cytoskeleton
is involved in the segregation of protein in normal cells.
To investigate whether cytoskeletal-based movement
results in the segregation of protein, we tracked the
contents of vesicles during in vitro microscopy assays.
These studies showed that the addition of ATP causes
fission of endocytic contents along microtubules, result-
ing in the segregation of proteins that are targeted for
different cellular compartments. The plasma membrane
proteins, sodium (Naþ) taurocholate cotransporting
polypeptide (ntcp) and transferrin receptor, segregated
from asialoorosomucoid (ASOR), an endocytic ligand that
is targeted for degradation. Epidermal growth factor
receptor, which is degraded, and the asialoglycoprotein
receptor, which remains partially bound to ASOR, segre-
gated less efficiently from ASOR. Vesicles containing
ntcp and transferrin receptor had reduced fission in the
absence of ASOR, suggesting that fission is regulated to
allow proteins to segregate. A single round of fission
resulted in 6.5-fold purification of ntcp from ASOR, and
25% of the resulting vesicles were completely depleted of
the endocytic ligand.
Key words: asialoglycoprotein receptor, endosome,
fission, microtubules, sorting, transferrin receptor
Received 18 July 2007, revised and accepted for publica-
tion 11 February 2008, uncorrected manuscript published
online 16 February 2008, published online 18 March 2008
Proteins at the plasma membrane are continuously endo-
cytosed and transported to intracellular early (sorting)
endosomes. From early endosomes, proteins are typically
sorted either to late endosomes, for subsequent degrada-
tion, or to recycling endosomes, from which they traffic
back to the plasma membrane (1,2). Coat proteins and
adaptor proteins appear to function in protein sorting and
budding at the endosome (3,4). However, examination
of endosomal protein within mammalian cells by light
microscopy reveals dramatic cytoskeletal-based trafficking,
tubule formation and fission (5–7), while examination by
electron microscopy reveals tubular extensions corres-
ponding to sites of protein sorting (8,9). Microtubule motor
proteins are involved in the movement of endocytic
proteins from the early/sorting endosomes to the perinu-
clear recycling vesicles and late endosomes (10,11), and it
has been shown that microtubule depolymerization can
delay protein degradation and block apical recycling and
transcytosis (12–15). However, it has also been shown in
cultured cells that the depolymerization of microtubules
does not block protein recycling (16,17). Therefore, the
precise role of microtubules in protein segregation is
unclear. The studies presented here address this issue
by examining how microtubule-based movement of endo-
cytic material that is seen by time-lapse microscopy of
living cells contributes to protein sorting and how this
sorting manifests itself at the level of an individual fission
event. Few reports have quantified whether cytoskeletal-
based fission can result in specific segregation of different
proteins. In contrast, the mechanisms of endocytic vesicle
fusion and the importance of cellular SNARE protein
machinery have been well established, in part because of
reconstitution of fusion in vitro (18,19). It has been shown
that fission can be quantitatively analyzed in vitro using
time-lapse microscopy (20). This report broadens those
studies with an in-depth analysis of fission and the
behavior of multiple proteins in order to determine
whether studies of single vesicles can provide an import-
ant tool for understanding the underlying mechanisms of
protein sorting.
Four integral membrane proteins were used in these
studies: transferrin receptor, asialoglycoprotein receptor
(ASGPR), epidermal growth factor receptor (EGFR) and
sodium (Naþ) taurocholate cotransporting polypeptide
(ntcp) as well as the lumenal markers, asialoorosomucoid
(ASOR) and fluorescent dextran. The transferrin receptor
and ASGPR are well-studied integral membrane proteins
that undergo recycling back to the cell surface once
internalized (21,22). In contrast, a significant portion of
EGFR is transported to multivesicular bodies and degraded
once it is internalized (23,24). Ntcp is a hepatocyte-specific
bile acid transporter that localizes to the plasmamembrane
as well as a subplasma membrane intracellular pool
(25,26). ASOR is a ligand for ASGPR and once internalized
www.traffic.dk 833

undergoes dissociation followed by sorting to the lysosomes
where it is degraded (12). Dextran is a soluble pinocytic
marker that enters cells in the fluid phase (27). These
markers were selected in order to study the behavior of
molecules that are targeted for either degradation or
recycling during individual fission events that occur along
microtubules.
Results
Vesicle motility and fission in primary hepatocytes
Previous studies have documented that early endosomes,
late endosomes and lysosomes undergo microtubule-
based movement during the processing of endocytic
contents (7,28,29). However, it has been difficult to
demonstrate how this movement contributes to metabolic
functions such as the segregation of protein. To confirm
that hepatocytes display microtubule-based movement of
their early endosomes, hepatocytes were incubated with
fluorescent endocytic ligand, ASOR, in the presence or
absence of the microtubule-depolymerizing drug, nocoda-
zole. Time-lapse fluorescent microscopy revealed that
ASOR is present within intracellular vesicles after 5 min
of uptake and that these undergo a complicated display of
fusion, fission and back-and-forth movement (Movie S1).
This is seen as bright streaks in the time-lapse overlay in
the top panel of Figure 1A. Treatment of cells with
nocodazole suspended nearly all long-distance motility
events (Movie S2), as seen by the lack of bright streaks
in the bottom panel of Figure 1A. Some smaller move-
ments and vibrating activities were still observed, indicat-
ing that endosomes were not rigidly fixed and that cells
were still capable of movement that did not depend on
microtubules.
We next asked whether vesicles containing proteins that
are targeted to different cellular locations could be seen
undergoing fission while moving on microtubules. We also
examined whether these fission events resulted in the
segregation of one protein from the other. Hepatocytes
were incubated with Alexa-488-labeled transferrin and
Texas red-labeled ASOR and subsequently imaged in
two fluorescence channels (Figure 1B). Transferrin re-
mains bound to its receptor during cycles of endocytosis
and recycling where it moves from the cell surface into
endosomes and then back to the cell surface (21). How-
ever, ASOR is taken up by the ASGPR and released within
endocytic compartments where it subsequently traffics to
the lysosomes and is degraded (30). As seen in Movie S3
and zoomed in Movies S5 and S6 (Movie S4 contains the
corresponding bright-field images), vesicles containing
both markers were highly motile and could be seen to
undergo fission. Still images fromMovie S5 demonstrating
fission are shown in Figure 1C.
These data indicate that a single vesicle can contain two
different proteins and undergo microtubule-based move-
ment and fission. However, these findings do not imply
that these events can lead to the overall segregation of
these proteins from each other. Whereas many studies
indicate that microtubule-based trafficking contributes to
endocytic protein sorting and the recycling of protein
(10,31), other studies have demonstrated that depolymer-
ization of the microtubule cytoskeleton does not block
protein recycling (16,17). In an attempt to address these
issues, we initiated studies to quantify the overall redistri-
bution of proteins during single vesicle fission events within
living cells. However, varying background fluorescence,
photobleaching and the complex, three-dimensional motion
of the vesicles prohibited accurate quantification of protein
using live cell imaging techniques. Given these difficulties,
we instead took advantage of an in vitro assay that has
been developed to analyze the microtubule motor proteins
that are involved in endocytic vesicle traffic (32,33).
Reconstitution of vesicle fission in vitro
A standardized in vitromotility assay was used to generate
the data below. Similar assays have been used to identify
and purify motor proteins, to characterize mutations of
motor proteins and to show that organelles use specific
sets of motor proteins and regulatory factors for cellular
transport (33–36). In brief, endocytic vesicles are isolated
by homogenization of rat liver followed by sucrose step-
gradient centrifugation. Vesicles are then stored as single-
use, frozen aliquots. Small volume (3 mL) glass microscope
chambers are constructed from glass slides, and micro-
tubules are polymerized from fluorescent tubulin and
attached to the bottom of the chambers. Endocytic
vesicles are thawed and added to the chambers where
they attach to the microtubules, and the unattached
contents are washed away. Fifty micromolar ATP is added
to the chambers to initiate vesicle motility, and this is
captured by high-resolution, wide-field, multi-fluorescence
channel, time-lapse microscopy. Prior to ATP addition,
chambers containing endocytic vesicles that are attached
to microtubules are stored in buffer on ice where they
remain active in motility assays for more than 3 h.
Two approaches are used to identify specific proteins
within the endocytic vesicles: (i) the hepatic portal vein is
injected with fluorescent ligand, and hepatocytes are
allowed to take up the ligand prior to homogenization of
the liver and (ii) the endosomes are labeled by immuno-
fluorescence within the microscope chambers after they
have bound to microtubules. Unless specifically noted, all
the biochemical preparations of vesicles in the following
experiments contain fluorescent-labeled ASOR that has
been injected into the hepatic portal vein 5 min prior to liver
homogenization. The red fluorescent vesicles within the
preparation represent ‘early’ endocytic ASOR as they
contain both ligand and receptor (20). This preparation also
contains a multitude of other vesicles that copurify with
these ASOR-containing vesicles. As described previously,
these vesicles bind to microtubules and move in the
834 Traffic 2008; 9: 833–847
Murray et al.

presence of ATPwithout the addition of exogenous protein
or cytosol (20,32,37). In addition, the moving vesicles
undergo fission. As in living cells, fission is observed as
the separation of a single region, or spot, of fluorescence.
Unlike in living cells, vesicles in vitro remain within the
focal plane during movement, and the history of all move-
ments can be tracked. This facilitates the differentiation of
fission from other motile events, such as the movement of
two vesicles in opposite directions (e.g. vesicle crossing).
Also unlike in living cells, vesicle motility by these techniques
occurs in a single burst that lasts approximately 90 sec-
onds. Subsequently, vesicles detach from microtubules
Figure 1: Legend on next page.
Traffic 2008; 9: 833–847 835
Endocytic Segregation In Vitro

or stop moving, presumably as a result of ATP hydrolysis
and depletion (32) or possibly because of changes in
protein phosphorylation or other factors (26). This relatively
short burst of movement simplifies data collection and
analysis.
Figure 1C (right panels) presents an example of fission
occurring in vitro. A vesicle containing ASOR (red) and
stained with antibody to the transferrin receptor (green) is
attached to a microtubule (white arrow). ATP was added
to the chamber, and after 21 seconds, a daughter vesicle
containing only transferrin receptor emerges, moving
to the right. A portion of the transferrin receptor remains
with the mother vesicle. Note that the size of the vesicle
and duration of fission are similar for the examples in the
living cell and in vitro (Figure 1C), that is, the apparent
diameter for both vesicles is approximately 1 mm and the
separation into two vesicles takes less than 10 seconds of
real time. Movies S7 and S8 provide additional examples of
fission events between ASOR and ntcp, and ASOR, ntcp
and transferrin receptor, respectively.
Fission is not the result of the movement of
overlapped vesicles
It is possible that fission observed during in vitro microtu-
bule motility assays results from the separation of over-
lapping or aggregated vesicles. Previous studies have
shown that fission is not reduced by diluting the vesicles
prior to attachment to microtubules (20), indicating that
fission is not the result of overlapped, unconnected
vesicles. To determine whether the vesicle isolation pro-
cedure produces aggregated vesicles, livers from rats that
had endocytosed either Texas red ASOR (red) or Alexa-
labeled ASOR (green) were homogenized together and
vesicles were isolated using the standard protocol. Any
vesicle that aggregates during this process would be
revealed by overlap of red and green fluorescence. The
degree of overlap was assessed in the presence and
absence of microtubules, and the level of ATP-induced
motility and fission was also measured. Images from these
experimentsareshown inFigure 1D.Of the961microtubule-
bound vesicles scored, 671 were green and 290 were red,
and of these, 32 (3.3%) showed some overlap of red and
green fluorescence. Of the overlapped vesicles, only three
underwent separation of green and red during motility
experiments. Thus, the clustering of red and green
vesicles was rare, and those that did cluster did not show
a high level of separation upon addition of ATP.
Vesicle motility and fission varies as a function of
protein constituents
In order to measure the segregation of protein during
microtubule-based fission, we first sought to characterize
the amount of motility and fission of vesicles that con-
tained different endocytic proteins. Several groups have
demonstrated that different populations of endocytic
vesicles have different amounts of motility (6,38). The
endocytic vesicle preparation used in these studies con-
tains the endocytic marker, ASOR, as well as numerous
other vesicles (11,32). Because motor proteins, coat
proteins or other regulatory proteins have the potential to
exchange between vesicles during their purification, it was
possible that all vesicles would display an identical level of
motility and fission in vitro. Alternatively, it was possible
that the amount of fission for some vesicles would be too
low to allow quantification of protein segregation.
We performed in vitro vesicle motility assays and sub-
sequently measured the level of motility and fission for
vesicles that contained the endocytic ligand, ASOR. Motil-
ity is defined as the per cent of microtubule-bound vesicles
that undergo movement, while fission is defined as the per
cent of motile vesicles that undergo fission. Twenty-one
per cent of microtubule-bound vesicles that contained
ASOR were motile (n ¼ 67), and 14% of these underwent
fission (Figure 2). These results are very similar to those
Figure 1: Movement and fission of endocytic vesicles in hepatocytes and in vitro. A) Control or nocodazole-treated hepatocytes
were allowed to endocytose fluorescent ASOR for 5 min. Cells werewashed and imaged by time-lapsemicroscopy for 1- to 1.5-min, 8 min
following the wash. The left panels show the first frame of the movie, and the right panels shows an overlay (maximum intensity
projection) of all movie frames. Arrows point to fluorescence streaks that indicate the movement of the internalized ASOR. These streaks
were absent in nocodazole-treated cells, but short-distance movement was observed as broadening of the fluorescence spots. B) Control
hepatocytes were allowed to endocytose Texas red ASOR (red) and Alexa-488 transferrin (Tfn) (green) and washed, imaged and displayed
as described above. Both ligands showed prominent long-distance movement as indicated by the streaks (arrows). The yellow box
indicates the region of the cell that is magnified for the subsequent figure. C) On the left, fission is observed in a living hepatocyte (from
Movie S5, an additional example is seen in Movie S6), while on the right, fission is observed in vitrowithin a microscope chamber. Arrows
mark the location of ‘mother’ vesicles, while arrowheads mark the location of emerging ‘daughter’ vesicles. In the hepatocyte, a vesicle
containing both ASOR (red) and Tfn (green) moves to the right and at 30 seconds begins to undergo fission, resulting in a separate daughter
vesicle by 42 seconds. In the in vitro example, vesicles were bound to microtubules (MTs) (red filaments) and labeled for endocytic ASOR
(red) as well as the transferrin receptor (TfR) (green), and ATP was added to initiate motility and fission at 0 seconds. Fission occurred at
21 seconds, resulting in a separate daughter vesicle by 30 seconds. D) Vesicles were assessed for the degree of aggregation in the
presence or absence ofMTs. A rat liver was allowed to endocytose Alexa-488 ASOR (green), and a second liver was allowed to endocytose
Texas red ASOR (red), and the green and red livers were homogenized together and endocytic vesicleswere purified according to protocol.
Most vesicles contained either red or green fluorescence (arrows), indicating that vesicles did not aggregate, although occasional overlap
was seen (arrowhead). The lack of aggregation is better seen when vesicles are imaged in the absence of microtubule (right). Scale bars,
5 mm, except in C, 2 mm.
836 Traffic 2008; 9: 833–847
Murray et al.

obtained in previous studies that used the same procedure
to purify endocytic vesicles. In those studies, 25% of the
ASOR-containing vesicles were motile and 13% of the
motile vesicles underwent fission [(20); n ¼ 832]. A sec-
ond endocytic marker, fluorescent dextran, was injected
into the rat hepatic portal vein instead of fluorescent
ASOR, and vesicles were isolated by the same procedure.
Dextran is a pinocytotic fluid-phasemarker frequently used
to track intracellular vesicles (39,40). In vitromotility assays
were performed, and the level of motility and fission of
dextran-containing vesicles was measured. It was found
that dextran-containing vesicles had higher motility than
ASOR-containing vesicles (58.2%, n ¼ 531, p � 0.001
versus 21% for ASOR) but significantly less fission of the
dextran fluorescence (8.4%, compared with 14% for
ASOR, p � 0.001; Figure 2). This indicates that the level
of motility and fission is different for different vesicle
populations, despite the fact that the vesicles were purified
by the same procedure. Interestingly, the dextran-containing
vesicles had higher motility than the ASOR-containing
vesicles, yet fission was significantly lower. This demon-
strates that fission and motility are regulated independ-
ently. Currently, it is not clear why dextran-containing
vesicles would have such high levels of motility, and we do
not know how these activities affect endocytic processing.
As mentioned above, the endocytic vesicle preparation
contains many different vesicles (11), and, when used as
an endocytic ligand, a subset of the vesicles will contain
ASOR. We had previously determined that many of the
vesicles within the preparation contain the transmembrane
protein, transferrin receptor, as well as the bile acid trans-
porter, ntcp (26). To determine if vesicles containing either
of these proteins had similar levels of motility and fission
as that of the ASOR-containing vesicles, motility assays
were performed for vesicles that were labeled by im-
munofluorescence for either transferrin receptor or ntcp.
Movies resulting from these experiments were analyzed
for vesicles that contained either ntcp or transferrin
receptor but not ASOR. Forty-one per cent of vesicles that
contained ntcp were motile, yet no fissions were detected
(n ¼ 861). Thirty-two per cent of the vesicles that con-
tained transferrin receptor were motile (n ¼ 322), and only
3.8% of these underwent fission (Figure 2). These data
indicate that some vesicles within the preparation have
a low amount of fission, despite having high motility.
Approximately 20% of the vesicles that contain either ntcp
or transferrin receptor also contain ASOR (data not
shown). When these vesicles were analyzed, it was found
that 29.1% of the vesicles that contained ASOR and ntcp
were motile (n ¼ 316) and 26.1% underwent fission of the
ntcp protein (Figure 2). Twenty point eight per cent of the
vesicles that contained transferrin receptor and ASOR
were motile, and 22.2% underwent fission of the trans-
ferrin receptor (n ¼ 303; Figure 2). In both cases, the
presence of the endocytic ligand corresponded with
increased fission and slightly reduced motility. When
vesicles containing endocytic dextran instead of endocytic
ASOR were used for similar studies, it was found that
ntcp, transferrin receptor and several other proteins failed
to show significant colocalization with the fluorescent
dextran in vitro, precluding the generation of additional
data (not shown). Overall, however, these experiments
demonstrated that different vesicles within the same
preparation have different levels of motility and fission
and that vesicles containing ASOR as well as other
proteins have sufficient fission to allow an analysis of the
segregation of ASOR.
Inhibition of PI3-kinase decreases fission but not
motility of vesicles that contain ASOR and ntcp
To confirm that fission and motility can be regulated
independently, we measured these activities following
preincubation with the phosphatidylinositol 3-kinase (PI3-
kinase) inhibitor, LY294002.Many reports have shown that
PI3-kinase inhibition reduces protein recycling and sorting
Figure 2: The level of motility and fission is different for
different populations of endocytic vesicles; fission is low for
vesicles that contain ntcp or transferrin receptor (TfR) but
lack ASOR. Motility and fission were measured in the in vitro
motility assay for vesicles that contained ASOR [n (number of
vesicles) ¼ 67, e (number of experiments) ¼ 5], dextran (n ¼ 531,
e ¼ 10), ntcp but no ASOR (n ¼ 861, e ¼ 7), transferrin receptor
(TfR) but no ASOR (n ¼ 322, e ¼ 4), ntcp and ASOR (n ¼ 316,
e ¼ 22), TfR and ASOR (n ¼ 303, e ¼ 13), and ntcp and ASOR
(n ¼ 212, e ¼ 15) after pretreatment with 50 mM LY294002, as
indicated. Motility was scored as the per cent of microtubule-
bound vesicles that underwent continuous directed movement
(y-axis). Fission was scored as the per cent of moving vesicles that
underwent fission (y-axis). Assays were performed using different
aliquots of the same preparations of endocytic vesicles, which
were purified following endocytic uptake of fluorescent ASOR into
rat liver. In some cases, fluorescent dextran was substituted for
ASOR (as indicated). Different populations of vesicles within the
preparation are indicated on the x-axis. These were identified
using fluorescent antibodies and fluorescent endocytic ASOR,
which labeled subsets of the vesicles within the preparation.
Traffic 2008; 9: 833–847 837
Endocytic Segregation In Vitro

(41–43), and we therefore examined whether the fission of
endocytic vesicles, which may participate in protein sorting
and recycling, may also be affected. PI3-kinase activity has
also been shown to be important for the formation of
phospholipids that allow the attachment of motor proteins
to vesicles (10,44). However, in these studies, LY294002
was added after vesicles had attached to microtubules,
and the PI3-kinase activity might not be limiting for the
motility of pre-attached vesicles.
The motility and fission of microtubule-bound vesicles
containing ASOR and ntcp were quantified following pre-
incubation with 50 mM LY294002 in the motility chamber.
As can be seen in Figure 2 (rightmost bars), 34.4% of the
inhibitor-treated vesicles moved along microtubules (n ¼212) compared with 29.1% of untreated vesicles (n ¼ 316),
suggesting that PI3-kinase inhibition has minimal effect
on the motility of vesicles that contain ASOR and ntcp
(p ¼ 0.2). However, fission was reduced from 26.1 to 9.6%
(p ¼ 0.01), indicating that PI3-kinase regulates the level of
vesicle fission without affecting motility.
Segregation of ASOR from ntcp
Previous studies have shown that the bile acid transporter,
ntcp, resides on the plasma membrane as well as intra-
cellularly (25,45), and recently, we have shown that
a fraction of this protein colocalizes with early endocytic
ASOR but not late endocytic ASOR (26). Fission of ntcp
away from ASOR in early endocytic vesicles represents
a potential means by which ntcp is depleted from the late
endosomes. This was examined in this study bymeasuring
the degree to which ASOR and ntcp segregate during
fission along microtubules. To obtain this data, early
endocytic ASOR-containing vesicles were bound to micro-
tubules in vitro and the presence of ntcp was detected on
the vesicles by immunofluorescence. ATP was added to
the vesicles during multi-channel, time-lapse fluorescence
microscopy experiments. Subsequently, the time-lapse
movies were analyzed for the presence of vesicles that
contain both ntcp and ASOR, and this population was
analyzed for fission. The amount of ASOR and ntcp segre-
gating to each daughter vesicle was determined by mea-
suring the amount of fluorescence for each fission. These
data are tabulated (Supplementary Tables 1, 2 and 6) and
displayed as a correlation of the two proteins (see below).
Figure 3 is a cartoon that indicates how a correlation plot
can be used to view protein segregation. On the left, the
figure depicts a vesicle undergoing fission into two daugh-
ter vesicles, and on the right, the per cent of protein
recovered in each daughter vesicles is plotted as a corre-
lation. Four hypothetical examples demonstrate different
distributions of red and green proteins into daughter
vesicles, while yellow indicates overlap of red and green
in the same vesicle. In the first example, red and green
proteins divide equally between the two daughter vesicles
and the proteins do not segregate. The correlation plot
shows that each daughter vesicle contains approximately
50% of the red and green proteins. In the second example,
the proteins divide unequally into the daughter vesicles but
again there is no segregation. One vesicle contains a large
percentage of both red and green proteins, while the other
vesicle contains a small percentage of both proteins. In this
case, the daughter vesicles reside in the upper right and
lower left quadrants of the correlation plot. In the third
example, the proteins divide unequally into the daughter
vesicles and partial segregation is achieved. One daughter
vesicle contains a large percentage of red protein along
with a small percentage of green protein, while the other
daughter contains a small percentage of red protein along
with a large percentage of green protein. These vesicles
reside in the upper left and lower right quadrants of the plot.
In the final example, complete segregation is achieved, and
one vesicle contains 100% of the red protein and 0% of the
green protein, and the other vesicle contains 0% of the red
protein and 100% of the green protein. These vesicles
reside in the extreme upper left and lower right of the plot.
The correlation plot therefore indicates whether proteins
segregate from each other during fission, and all fissions
are displayed so that the scatter in the data is revealed. The
percentage of vesicles that are found in each quadrant can
be included as an additional measure of the segregation of
the two proteins as is frequently performed for flow
cytometry analysis (46). Proteins that segregate will be
negatively correlated, localizing to the upper left and lower
right quadrants, while proteins that do not segregate will
be positively correlated, localizing to the upper right and
lower left quadrants. Because both daughter vesicles are
included, the graph is rotationally symmetric (each vesicle
is related to the other daughter of the pair by the formula:
100% � co-ordinate value). Also note that the correlation,
plotted as a percentage (as in Figure 3), does not provide
information on the total amount of fluorescence.
Figure 4A presents the correlation plot for vesicle fissions
that contain ntcp and ASOR. Overall, the vesicles cluster to
the upper left and lower right quadrants of the plot,
indicating that the proteins are segregating. The degree
of segregation can be represented by the average values
of the lower left and lower right quadrants. Vesicles in
these lower quadrants contain the smaller portion of
ASOR. They also contain some proportion of ntcp. If
protein divided equally likely into any portion and if the
proteins distributed independently from one another, then
the average value of the lower quadrants would be 25%
for ASOR and 50% for ntcp. Twenty-five per cent is the
middle value for the lower half of a population, and 50%
is the middle value for a total population. However, in
the actual correlation plot, these quadrants contained
11.3 � 1.6% (mean � SEM) of the ASOR and 72.9 �3.0% of the ntcp, as indicated by the triangle and dotted
lines of Figure 4A. The scatter of the data indicates that
the per cent of protein distributing to daughter vesicles
is variable. In addition, there are many vesicles that
838 Traffic 2008; 9: 833–847
Murray et al.

completely lack ASOR and some that completely lack ntcp,
and therefore, some fission events achieve complete
segregation of the proteins. It should also be noted that
if the observed fission was actually the result of independ-
ent movement of overlapped ‘pure’ vesicles (i.e. vesicles
that contained only ASOR or only ntcp), then ‘mixed’
daughter vesicles (containing both ntcp and ASOR) should
rarely be observed. Instead, about 60% of all daughter
vesicles were mixed. In summary, these results suggest
that intracellular fission produces daughter vesicles con-
taining a broad distribution of protein, with many vesicles
(32 of 130 or 25%) completely devoid of ASOR, a marker
that is targeted for degradation within lysosomes.
Segregation of ASOR from transferrin receptor
Several other protein markers were selected for study
in order to determine whether segregation during fis-
sion is dependent on the identity of the protein and
whether proteins that are targeted for different cellular
destinations undergo greater segregation than those
that are targeted for the same destination. Figure 4B
presents a correlation plot for vesicles containing
transferrin receptor and ASOR. These experiments were
performed identically to the experiments containing ntcp
except that antibodies to transferrin receptor were
included to identify this protein. Figure 4B indicates
that the transferrin receptor also segregates away from
Figure 3: Graphing the distribution of protein from vesicle fission as a correlation. A cartoon indicates several examples of how two
proteins may distribute to different vesicles during fission and how this can be plotted as a correlation. A mother vesicle containing red and
green proteins is shown on the left bound to a microtubule (red filament); yellow indicates overlap of the red and green proteins. The
mother vesicle undergoes fission (arrow) into two daughter vesicles, and the fraction of red and green proteins distributing to each
daughter is plotted as a percentage as two dots. In the first example, protein distributes approximately equally to the daughter vesicles, and
the daughter vesicles appear centrally localized on the correlation plot. In the second example, protein distributes unequally to the
daughters, but red and green proteins do not segregate. These daughters localize to the lower left and upper right quadrants of the plot.
In the third example, protein distributes unequally to the daughters, and red and green proteins partially segregate. Vesicles from such
fissions localize to the upper left and lower right quadrants of the plot. In the final example, the proteins segregate completely, and
daughter vesicles align at the extreme upper left (0 and 100%) and lower right (100 and 0%) portions of the plot. Because both daughter
vesicles are included, the graph is rotationally symmetric (each vesicle is related to the other daughter of the pair by the formula:
100% � co-ordinate value).
Traffic 2008; 9: 833–847 839
Endocytic Segregation In Vitro

Figure 4: Segregation of ASOR from ntcp and transferrin receptor during microtubule-based motility in vitro. Vesicles were
purified from rat liver that contained fluorescent endocytic ASOR as well as subpopulations of unlabeled vesicles as described in Figure 2.
These were bound to microtubules and labeled for ntcp and transferrin receptor by immunofluorescence. Motility assays were performed
and subsequently analyzed for vesicles that have undergone fission and contain ASOR and ntcp or transferrin receptor. The amount of
protein present on each endocytic vesicle was measured by fluorescence intensity. The distribution of ASOR was plotted as a correlation
versus ntcp or transferrin receptor in the manner outlined in the cartoon of Figure 3. Each point represents a single daughter vesicle after it
has undergone fission. In (A) and (B), the per cent of protein segregating to daughter vesicles is presented, and the graph is rotationally
symmetric. In (C) and (D), the total amount of fluorescence is presented. Fissions showing identical percentages were offset by 0.5%
along the x-axis for visualization. A) ASOR and ntcp predominantly segregated to opposite vesicles during fission. The lower quadrants
contained an average of 11.3 � 1.6% (SEM) of the ASOR and 72.9 � 3.0% of the ntcp, as indicated by the triangle and dotted line.
B) ASOR and transferrin receptor segregated to opposite vesicles during fission, although the trend was not as strong as for ASOR and
ntcp. The lower quadrants contained an average of 10.9 � 1.7% of the ASOR and 61.0 � 3.2% of transferrin receptor as indicated by the
triangle and dotted line. C) A plot of total fluorescence (rather than per cent) of ASOR versus ntcp for the same vesicles that were used in
(A). This does not reveal a negative correlation, which indicates that the negative correlation of (A) is a result of fission rather than the
amount of protein on the vesicles per se. D) A plot of the total fluorescence of ASOR versus transferrin receptor for the same vesicles that
were used in (B). Fluorescence is in arbitrary units for all probes. UL, UR, LL and LR are abbreviations for upper left, upper right, lower left
and lower right, and the adjacent number indicates the percentage of vesicles that are found in the quadrant.
840 Traffic 2008; 9: 833–847
Murray et al.
ASOR during fission. The lower right and lower left
quadrants contained 10.9 � 1.7% of the ASOR and
61 � 3.2% of transferrin receptor, as indicated by the
triangle and dotted lines. There is also a similar degree of
scatter and a similar amount of vesicles that completely
lack ASOR (30%).
Lack of correlation between total fluorescence
in daughter vesicles
It could be argued that the degree of protein segregation
that we have attributed to fission is in reality a reflection of
the degree to which the proteins are present in vesicles
overall. That is, perhaps those vesicles that contain a low
amount of ASOR also contain a high amount of ntcp,
independent of whether or not these vesicles have under-
gone fission. We tested this hypothesis by plotting a cor-
relation for the total amount of fluorescence, rather than
the per cent, that is present on the vesicles of the previous
two data sets. Figure 4C,D shows that there is no strong
overall trend for either ntcp or transferrin receptor with
respect to ASOR, that is, vesicles containing either a high
or a low amount of ASOR can contain a high or low amount
of ntcp or transferrin receptor. It is only when the vesicles
are paired because of fission (Figure 4A,B) that we see
a low amount of ASOR distributing with a high amount of
ntcp. Mean values were used to divide the total fluores-
cence data into quadrants in the manner of Figure 4A,B,
and these did not reveal any unusual distributions
of protein. Eighty-three per cent and 68% of the vesicles
were found in the negatively correlated (upper left and
lower right) quadrants in Figure 4A,B, while 47 and 55% of
the vesicles were found in the negatively correlated
quadrants of Figure 4C,B.
Segregation of transferrin receptor from ntcp, ASOR
from ASGPR and ASOR from EGFR
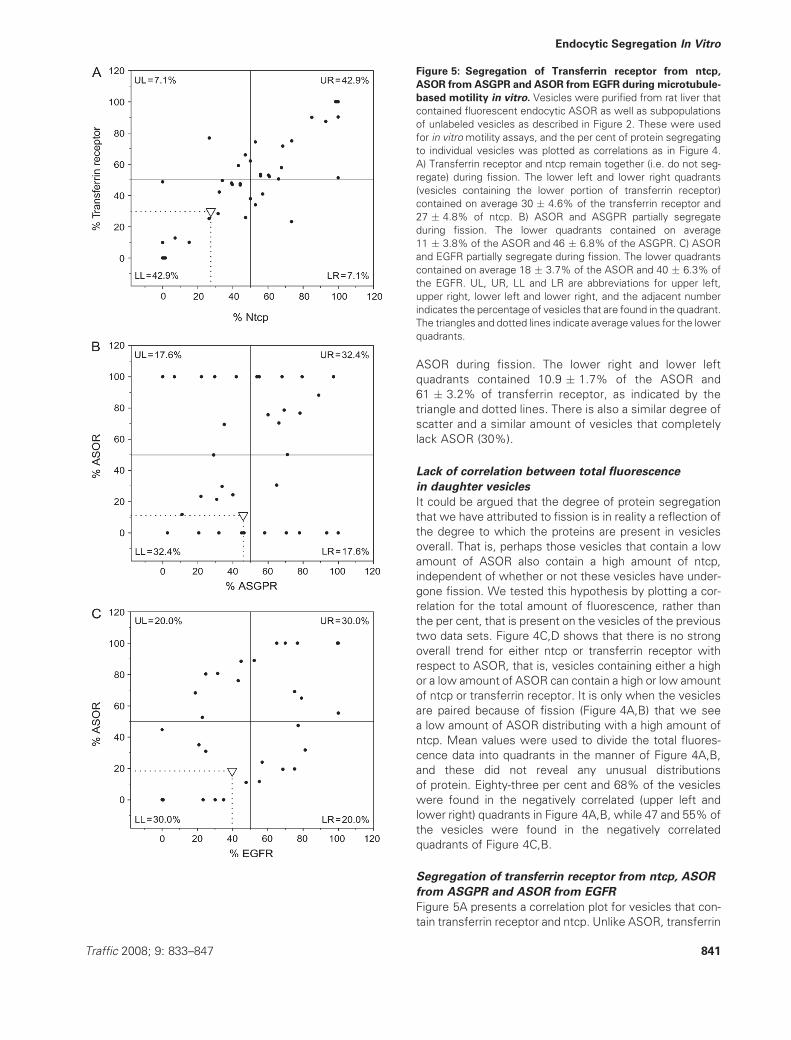
Figure 5A presents a correlation plot for vesicles that con-
tain transferrin receptor and ntcp. Unlike ASOR, transferrin
Figure 5: Segregation of Transferrin receptor from ntcp,
ASOR fromASGPR and ASOR from EGFR during microtubule-
based motility in vitro. Vesicles were purified from rat liver that
contained fluorescent endocytic ASOR as well as subpopulations
of unlabeled vesicles as described in Figure 2. These were used
for in vitromotility assays, and the per cent of protein segregating
to individual vesicles was plotted as correlations as in Figure 4.
A) Transferrin receptor and ntcp remain together (i.e. do not seg-
regate) during fission. The lower left and lower right quadrants
(vesicles containing the lower portion of transferrin receptor)
contained on average 30 � 4.6% of the transferrin receptor and
27 � 4.8% of ntcp. B) ASOR and ASGPR partially segregate
during fission. The lower quadrants contained on average
11 � 3.8% of the ASOR and 46 � 6.8% of the ASGPR. C) ASOR
and EGFR partially segregate during fission. The lower quadrants
contained on average 18 � 3.7% of the ASOR and 40 � 6.3% of
the EGFR. UL, UR, LL and LR are abbreviations for upper left,
upper right, lower left and lower right, and the adjacent number
indicates the percentage of vesicles that are found in the quadrant.
The triangles and dotted lines indicate average values for the lower
quadrants.
Traffic 2008; 9: 833–847 841
Endocytic Segregation In Vitro

receptor did not segregate from ntcp during fission.
Instead, 86% of the daughter vesicles were contained in
the upper right and lower left (positively correlated) quad-
rants of the plot. The lower left and lower right quadrants
(vesicles containing the lower portion of transferrin recep-
tor) contained on average 30 � 4.6% of the transferrin
receptor and 27 � 4.8% of ntcp, indicating that vesicles
with a low amount of transferrin receptor also contained
a low amount of ntcp on average. Few vesicles (4 of 42
or 10%) completely lacked transferrin receptor. Cumula-
tively, these data indicate that two basolateral, plasma
membrane proteins, ntcp and transferrin receptor, are able
to efficiently segregate from the endocytic ligand, ASOR,
during microtubule-based fission while they are unable to
segregate from each other.
ASGPR, the receptor for ASOR, was chosen as a fourth
marker for the analysis of protein segregation. ASGPR–
ASOR defines a classical pathway whereby receptor binds
to ligand at the cell surface, releases ligand intracellularly
and the ligand is targeted to lysosomes where it is
degraded, while the receptor is recycled back to the cell
surface (47,48). Further studies showed that approxi-
mately half of the ligand dissociates from the receptor
rapidly (t½ � 2.5 min) following endocytic uptake, while
the other half dissociates slowly (t½ � 50 min). Therefore,
for reasons that are still not understood, a significant
portion of ASOR remains bound to ASGPR during endo-
cytic processing and vesicular traffic (49,50).
We used the in vitro assay to address whether ASOR that
is located within endocytic vesicles after 5 min of uptake
undergoes efficient segregation from ASGPR during
microtubule-based fission. Figure 5B shows that segrega-
tion between ASOR and ASGPR was not as efficient as
the segregation of ASOR from either ntcp or transferrin
receptor. On average, 11 � 3.8% of ASOR and 46� 6.8%
of ASGPR were present in the lower quadrants of the
correlation plot. A similar level of segregation of ASGPR
from ASOR was also observed in a previous study (20).
This level of segregation was intermediate between the
segregation of ASOR from ntcp (proteins that strongly
segregated) and transferrin receptor from ntcp (proteins
that did not segregate). The percentage of vesicles in the
negatively correlated upper left and lower right quadrants
totaled 83, 35 and 14% for the segregation of ASOR from
ntcp, ASOR from ASGPR and transferrin receptor from
ntcp, respectively. Interestingly despite moderate overall
segregation, 29% of vesicles completely lacked ASOR as
a result of fission of vesicles containing ASGPR and ASOR.
These studies suggest that ASOR is not completely free to
segregate from ASGPR within the endocytic vesicles and
that the slowly dissociating [‘state 1’ (48)] receptors are
present within endocytic vesicles that can bind to and
move along microtubules.
The EGFR was chosen as a fifth marker for the study of
endocytic segregation as it is an integral membrane recep-
tor that follows a different intracellular trafficking pathway
than either transferrin receptor or ASGPR. Whereas the
transferrin receptor and ASGPR are recycled to the plasma
membrane following endocytosis, the EGFR is taken up by
clathrin-mediated endocytosis and transported to the inner
membranes of multivesicular bodies where a significant
portion of the receptormoves to lysosomes and is degraded
(23,24,51). Figure 5C shows that the EGFR does not sort
away from the degraded ligand, ASOR, during microtubule-
based fission. On average, 18 � 3.7% of the ASOR and
40 � 6.3% of the EGFR were present in the lower quad-
rants of the correlation plot, and only 40% of the vesicles
were present in the negatively correlated upper left and
lower right quadrants of Figure 5C. These studies suggest
that the different endocytic sorting pathways of EGFR and
transferrin receptor are reflected in their different sorting
behavior with respect to ASOR.
Summary of the sorting behavior of the proteins
Table 1 summarizes the sorting of the different combin-
ations of proteins that were studied in this report. Each
fission is the division of a single vesicle into two vesicles,
and therefore, the amount of protein sorting to each
vesicle was plotted as per cent of total (Figures 4A,B and
5A–C). Strong segregation would be indicated by a low
percentage of one protein sorting into the same vesicles
that contain a high percentage of the other protein. This
is shown in Table 1 as the ‘average per cent of protein
Table 1: Summary of the degree of protein segregation during
fission
Protein pair
analyzed
Average per cent
protein recovered
in one daughter
vesiclea
Fold
purificationbnc Per cent of
fissions with
protein
segregationd
ASOR 11.3 � 1.6 6.5 65 83
Ntcp 72.9 � 3.0
ASOR 10.9 � 1.7 5.6 72 68
TfR 61.0 � 3.2
ASOR 18.4 � 3.7 2.2 15 40
EGFR 39.7 � 6.3
ASOR 11.2 � 3.8 4.1 17 35
ASGPR 45.8 � 6.8
TfR 29.9 � 4.6 0.9 21 14
Ntcp 27.4 � 4.8
aThis is the average per cent protein � SEM in the lower left and
lower right quadrants of Figures 4A,B and 5A–C. These are the
daughter vesicles from each fission that contain the smaller
portion of the first listed protein.bThis is the purification of the second listed protein from the first in
these vesicles, compared with the pre-fission vesicle, obtained by
dividing the percentages in the previous column.cThe number of fissions.dThis is the per cent of vesicles located within the negatively
correlated quadrants of Figures 4A,B and 5A–C. In these fissions,
the majority of one protein was found in the vesicle containing the
minority of the other protein.
842 Traffic 2008; 9: 833–847
Murray et al.

recovered in one daughter vesicle’ and corresponds to the
average per cent of protein in the lower left and lower right
quadrants of the correlation plots. About 11.3% of the
ASOR and 72.9% of the ntcp were found in the same
vesicles, indicating that a single round of fission produced
a 6.5-fold purification of ntcp (72.9/11.3%) on average.
Table 1 ranks the proteins according to their tendency to
segregate during fission. Ntcp segregated strongly from
ASOR, transferrin receptor segregated slightly less
strongly, and EGFR and ASGPR had moderate segregation
from ASOR. Transferrin receptor, however, did not segre-
gate from ntcp. This can be seen in the column that lists
the ‘per cent fissions with protein segregation’ and corres-
ponds to the per cent of vesicles that are contained in the
negatively correlated, upper left and lower right quadrants.
The degree of segregation was significantly different for all
protein markers with p-values of 0.01, <0.005 and <0.005
when comparing the per cent of ntcp segregating from
ASOR to the per cent of transferrin receptor, EGFR and
ASGPR segregating from ASOR. These data indicate that
proteins have different degrees of segregation during
fission, and it suggests that microtubule-based fission
events are participating in the segregation of protein.
The degree of segregation does not correlate with
vesicle fluorescence intensity or size
A population of ‘highly sorted’ daughter vesicles was
analyzed for their fluorescence intensity and motility.
Daughter vesicles arising from fission were considered
highly sorted if they contained at least 80% of a single
marker protein and less than 20% of the ‘opposing’ marker
proteins. For ASOR, both ntcp and transferrin receptor
were considered opposing marker proteins, while for ntcp
and transferrin receptor, ASOR was considered the oppos-
ing marker protein (Table 2). The highly sorted vesicles
were brighter on average than the remaining vesicles,
indicating that protein segregation did not occur into dim
vesicles. The highly sorted vesicles also had equal or
greater standard deviation of fluorescence, indicating that
protein did not segregate into vesicles of uniform intensity.
Overall, this analysis did not reveal any correlation
between fluorescence intensity and the degree of protein
sorting. Furthermore, the total fluorescence correlation
plots of Figure 4C,D showed that fluorescence intensities
were dispersed. These results are consistent with the
concept that protein segregates to vesicles that have
a broad distribution in size and intensity (52).
The transmembrane proteins, ntcp and transferrin
receptor, were highly motile during fission
Fissions were observed as the division of a single vesicle
into two daughter vesicles. Typically, one of the daughter
vesicles was motile, while the other remained stationary,
although in some fissions (6%) both daughters were
motile (Supplementary Tables). A review of the data
shows that some proteins were less abundant in the
moving vesicles than others. For instance, on average,
66% of the daughter vesicles in the lower left and lower
right quadrants of Figures 4A,B and 5A–C were motile.
These are the daughter vesicles that contained the smaller
amount of ASOR from the fission. The trend is more
apparent when looking at fissions that strongly segregated
protein. Seventy-two per cent of daughter vesicles that
contained either ntcp or transferrin receptor but were
completely depleted of ASOR after fission were motile
(n ¼ 60, culled from Tables S1–S6). However, only 38% of
the vesicles that contained ASOR but were completely
depleted of ntcp were motile (n ¼ 16, p ¼ 0.01), and 20%
of vesicles that contained ASOR but were completely
depleted of transferrin receptor were motile (n ¼ 11,
p < 0.001). This is also seen in the analysis of ‘highly
sorted’ vesicles of Table 2. Seventy per cent and 86% of
the highly sorted ntcp- and transferrin receptor-containing
vesicles, respectively, were motile compared with 41%
of the highly sorted ASOR-containing vesicles (n ¼ 27,
p ¼ 0.03 and p < 0.001, respectively, compared with highly
sorted ASOR-containing vesicles). These data show that
fissions were frequently the result of the transmembrane
proteins, ntcp and transferrin receptor, moving away from the
ligand, ASOR, that is located on the lumenal side of the
vesicle. This is consistent with the concept of ‘geometric’
protein sorting inwhich transmembrane proteins are believed
to be concentrated into tubules that bud or pull away from
the endocytic lumen, resulting in protein segregation (8,53).
Discussion
This study demonstrates that microtubule-based motility
results in segregation of endocytic protein and that the
Table 2: Properties of the highly sorted vesiclesa
Protein Highly sorted
vesicles
Remaining
vesicles
ASOR
Mean intensity 337.7 244.5
SD 305.8 355.3
Per cent moving 41 52
n 27 172
Ntcp
Mean intensity 399.5 301.3
SD 335.3 347.3
Per cent moving 70 54
n 20 90
Transferrin receptor
Mean intensity 717.9 477.8
SD 585.9 478.0
Per cent moving 86 50
n 14 112
aThese are a subset of fissions where proteins strongly sorted
from each other. They contained at least 80% of the listed protein
and less than 20% of the ‘opposite’ protein. For ASOR, the
opposite protein was considered as both ntcp and TfR, and for
ntcp and TfR, the opposite protein was considered as ASOR.
Traffic 2008; 9: 833–847 843
Endocytic Segregation In Vitro

degree of segregation is specific to the particular protein.
In living cells, internalized ligands were seen to undergo
multiple microtubule-based vesicle fissions, resulting in
apparent segregation of protein (Figure 1). However, vary-
ing background fluorescence, photobleaching and the
complex, three-dimensional motion of the vesicles pre-
vented accurate quantification of protein fluorescence.
Instead, we used endocytic vesicles prepared from rat
liver to recapitulate microtubule-based traffic within in vitro
microscopy assays. This allowed direct quantification of
protein with fluorescent antibodies and fluorescent-tagged
ligands. Previous reports have shown that movement of
the vesicles is dependent on specific sets of motor
proteins and that the inhibition of motor proteins blocks
fission (11,20,26,33). Fission resulted from splitting and
movement of at least one daughter vesicle along the
microtubule in every case (Supplementary Tables).
It is possible that fission under these conditions is depend-
ent only on the activity of motor proteins. However, we
found that fission is regulated differently than vesicle
motility. For instance, vesicles containing transferrin recep-
tor and ASOR had decreased motility but increased fission
compared with vesicles containing transferrin receptor but
no ASOR. Also, PI3-kinase inhibition reduced fission but
not motility (Figure 2). This suggests that fission on micro-
tubules results from the co-ordinated activity of motor pro-
teins with other cellular machineries. These may include
proteins that have been shown to be involved in vesicle
budding and protein sorting such as coatomers, adaptors
and ADP ribosylation factor 1 (54). It should be emphasized
that our studies were performed without addition of exo-
genous proteins and that soluble material is removed
during the vesicle purification and microscopy assays.
The vesicles themselves had all the machinery required
to produce motility, fission and protein segregation.
Analyses of protein levels on vesicles undergoing fission
suggest that proteins segregate during fission only when
they are destined for different cellular compartments. Ntcp
strongly segregated away from ASOR (Table 1), and this
was achieved because ASOR sorted asymmetrically, leav-
ing many ntcp vesicles completely devoid of ASOR (Fig-
ure 4A). The transferrin receptor showed a similar but
somewhat weaker segregation from ASOR (Figure 4B;
Table 1), whereas ntcp and transferrin receptor, proteins
that are both recycled to the plasma membrane, remained
together during fission (Figure 5A). ASOR showed moder-
ate segregation from its receptor, ASGPR. Although the
receptor is recycled and the ligand is degraded, a significant
portion of ASOR remains bound to receptor following
endocytosis and only dissociates slowly (49). Similar slowly
dissociating pools of ligand are seen for many if not most
receptor–ligand pairs, and a full understanding of this
phenomenon remains to be elucidated. EGFR, an integral
membrane protein that is targeted for degradation (23), also
failed to segregate efficiently from ASOR. It is interesting to
consider that neither ASGPR nor EGFR segregates effi-
ciently from ASOR in vitro. However, within cells, ASGPR is
recycled to the plasmamembrane, while EGFR is degraded.
Analysis of the data indicates that, although their overall
segregation from ASOR is similar, more ‘pure’ ASGPR
vesicles are generated than pure EGFR vesicles (Figure
5B,C). This suggests that the generation of pure vesicles is
an important step for protein segregation, and we speculate
that these vesicles may be directly targeted to the recycling
compartment and plasma membrane.
The geometric model of protein segregation proposes that
vesicles enriched in transmembrane receptors are physi-
cally pulled away from lumenal protein (53). Agreeing with
this, we found that fissions frequently produced a motile
daughter vesicle that was enriched in transmembrane
protein (Table 2). A recent study that tracked the move-
ment of endocytic vesicles within cells showed that
ligands can pre-sort to dynamic and static endosome
populations and that the dynamic population is more
rapidly processed for degradation (6). We also observed
populations of vesicles with different rates of motility.
However, in our studies, vesicles containing proteins that
are destined for recycling (ntcp and transferrin receptor)
had similar or higher motility than vesicles containing
proteins or molecules that are destined for degradation
(ASOR and dextran; Figure 2). Many factors can regulate
motility (5,26,55), and additional studies will be needed to
understand this regulation. The recycling of membrane
transporters such as GLUT4 or Naþ/Hþ exchanger (42,43)
has been found to be dependent on PI3-kinase activity, and
in this study, it is shown that PI3-kinase inhibition reduces
fissions by more than half without affecting vesicle motil-
ity. Studies from our laboratory have found that inhibition
of the microtubule-based minus-ended motor KIFC1 de-
creased fission (33) and that rab4 may have a role (55)
in regulation of the fission machinery. Activation of the
molecular motors by protein kinases (56,57) and concom-
itant involvement of the Rab proteins represent potential
mediators of membrane fission and vesicle processing.
Materials and Methods
Chemicals and reagentsCommercial antibodies used were transferrin receptor (number 13-6800;
Zymed laboratories) and EGFR (number sc-03; Santa Cruz Biotechnology).
In-house-generated antibodies used were ASGPR, to the C-terminal
cytoplasmic tail (20), ntcp, to amino acids 337–350 at the cytoplasmic
C-terminus (26). Primary antibodies were used at 0.33–20 mg/mL. Fluores-
cent secondary antibodies were from Jackson ImmunoResearch. ASOR
was prepared from human orosomucoid (Sigma-Aldrich) by acid hydrolysis
(58) and labeled with Texas red sulfonyl chloride or Alexa-488 (Invitrogen
Corporation) as described (32). Some ASOR was labeled with 125I by
a chloramine-T method to a specific activity of 10 000–15 000 c.p.m./ng
(59). Amino dextran (Cat. no. D1861; Invitrogen Corporation) was labeled
with Texas red sulfonyl chloride. Labeled and unlabeled tubulin was from
Cytoskeleton. All other reagents were from Sigma-Aldrich.
Fluorescent ASOR-containing endocytic vesicles were prepared from rat
liver as previously described (11,20,26,32,60,61). For some experiments,
the portal veins of two livers were separately injected with either Texas red
844 Traffic 2008; 9: 833–847
Murray et al.

or Alexa-488 ASOR, and these were homogenized together and vesicles
were isolated according to protocol. Taxol-stabilized fluorescent micro-
tubules were polymerized from rhodamine–tubulin as previously described
(11). Assay of vesicle motility on microtubules was as described (20,32,60).
Antibodies were screened for their appropriateness in immunofluores-
cence assays by Western blot and whole cell immunofluorescence (37).
For a given day’s experiment, 10–15 chambers with microtubule-bound,
immunofluorescence-stained endosomes were assembled and stored on
ice, and the chambers were sequentially placed on a microscope stage
maintained at 378C. Time-lapse multi-fluorescence channel microscopy
was then initiated, and 50 mM ATP was added to induce movement and
fission of the endosomes.
Immunofluorescence analysis in vitroDetection of protein on unfixed endocytic vesicles bymonoclonal or affinity-
purified polyclonal antibody followed by fluorescence secondary antibody
was performed as described (60). Wide-field imaging was performed at
378C using a 60�, 1.4 numerical aperture Olympus objective on an
Olympus 1X71 inverted microscope equipped for multi-channel fluores-
cence with a Lambda DG-4 excitation source and Lambda 10-2 emission
filter wheel (Sutter Instruments). Data were collected through a CoolSnap
HQ cooled charge-coupled device camera (Photometrics, Roper Scientific)
using METAMORPH software (Molecular Devices). Time-lapse movies were
captured in 1–3 channels (Cy2, Cy3 and Cy5). Some images were captured
on a BioRad Radiance confocal microscope in the Analytical Imaging Facility
(Albert Einstein College of Medicine). Movies were analyzed using IMAGEJ
software (National Institutes of Health public domain; http://rsb.info.nih.-
gov/ij/) and ADOBE PHOTOSHOP v 6.0 (Adobe Systems Inc.). Protein signal was
measured as the signal intensity minus background multiplied by the area.
Background intensity was measured and averaged from three neighboring
regions. To adjust for different imaging conditions (e.g. exposure time and
excitation intensity), fluorescence was normalized so that each set of
experiments gave equal median vesicle fluorescence for a given protein.
For total fluorescence correlation plots, 10 units (in arbitrary units) were
added to each vesicle to allow plotting on a logarithmic scale. The p-values
for correlation coefficients were from Student’s t-test. The Supplementary
Tables present all the data from different sets of experiments that were
performed either with different combinations of antibodies or after pre-
incubation with 50 mM LY294002. In the correlation plots, fissions were
culled from all experimental sets where the listed proteins were used,
regardless of the presence of other proteins.
ASOR uptake in cultured rat hepatocytesRat hepatocytes were isolated by collagenase perfusion as described (55),
attached to coverslip-bottomed dishes (MatTek Corporation) that were
coated with 0.5 mg/mLMatrigel (BD Biosciences) and cultured overnight in
Hepatozyme media (Invitrogen Corporation). Dishes were treated with
10 mg/mL Texas red-labeled ASOR and, in some experiments, 20 mg/mL
Alexa-488 transferrin (Invitrogen Corporation) for 5 min, washed and
imaged at 378C with 200-millisecond exposure times. Images were pro-
cessed with a SpotEnhancing filter (Daniel Sage, Biomedical Imaging Group).
For some experiments, cells were cooled to 48C in the presence or absence
of 30 mM nocodazole for 30 min, followed by warming to 378C and exposure
to ASOR as described above in continuous exposure to nocodazole.
Acknowledgments
This study was supported by National Institutes of Health grants DK41918
and DK23026.
Supplementary Materials
Table S1: Per cent of protein, motility and other data for daughter vesicles
resulting from fissions of vesicles containing ASOR and ntcp
Table S2: Data from fissioned vesicles containing ASOR, ntcp and trans-
ferrin receptor
Table S3: Data from fissioned vesicles containing ASOR and transferrin
receptor
Table S4: Data from fissioned vesicles containing ASOR, ASGPR and
transferrin receptor
Table S5: Data from fissioned vesicles containing ASOR, EGFR and
transferrin receptor
Table S6: Data from fissioned vesicles containing ntcp and ASOR, and
experiments performed after incubation with the PI3-kinase inhibitor,
LY294002
Movie S1: Endocytic vesicle movement in a control cell (Figure 1A).
Hepatocytes were allowed to take up fluorescent ASOR for 5 min, washed
and after additional 8 min, time-lapse movies of the endocytic vesicles
were captured at 1 frame per 1.2 seconds for a total of 72 seconds. A
hepatocyte is shown with multiple endocytic vesicles traversing the cell
and undergoing varied morphological changes, such as fusion, fission and
tubulation. Scale bar, 5 mm.
Movie S2: Endocytic vesicle movement in a nocodazole-treated cell
(Figure 1A). Hepatocytes were treated with 30 mM nocodazole to depoly-
merize microtubules and then allowed to take up ASOR, followed by
washing as in Movie S1. Time-lapse images were captured at 1 frame per
3 seconds for a total of 90 seconds. Note the lack of long-distance vesicle
movement. Short-distance, oscillating behavior is still observed. Scale bar,
5 mm.
Movie S3: ASOR and transferrin endocytic vesicle movement in control
cells (Figure 1B). Hepatocytes were allowed to take up both Texas red
ASOR (red) and Alexa-488 transferrin (green) for 5 min, washed and imaged
and displayed as described above but with two fluorescence channels.
Time-lapse images were captured at 1 frame per 1.5 seconds for a total of
90 seconds. Both transferrin and ASOR undergo long-distance movement
with morphological changes. Scale bar, 5 mm.
Movie S4: The corresponding bright-field images of Movie S3. Scale
bar, 5 mm.
Movie S5: Endocytic fission of ASOR and transferrin in a living cell (also
presented in Figure 1C) magnified fromMovie S3. A vesicle containing both
transferrin and ASOR moves from lower left toward the upper right. At
frame 26, fluorescence divides and the resulting daughter vesicle moves to
the left. Scale bar, 2 mm.
Movie S6: Second example of endocytic fission of ASOR and transferrin
magnified fromMovie S3. A vesicle containing ASOR and transferrin moves
from central left toward the lower right and then back again. At frame 21
a green daughter emerges. Scale bar, 2 mm.
Movie S7: Endocytic fission of ASOR and transferrin receptor in vitro (see
Figure 1C for an additional example). Endocytic vesicles containing fluor-
escent ASOR (red dots) were isolated from rat liver and subsequently
bound to microtubules (red) and immunostained for transferrin receptor
(blue dots) or ntcp (green dots) within microscope chambers. ATP was
added to initiate motility. Time-lapse images were captured at 1 frame per
1.5 seconds for a total of 24.5 seconds. A vesicle containing ntcp and
ASOR moves to the lower left and at the same time undergoes a fission
event. Scale bar, 2 mm.
Traffic 2008; 9: 833–847 845
Endocytic Segregation In Vitro

Movie S8: Second example of endocytic fission captured as described in
Movie S7. A vesicle containing ntcp, transferrin receptor and ASOR
undergoes a fission event, and the resulting daughter vesicle moves to
the right. Total time was 19.5 seconds. Scale bar, 5 mm.
Supplemental materials are available as part of the online article at http://
www.blackwell-synergy.com
References
1. Gruenberg J. The endocytic pathway: a mosaic of domains. Nat Rev
Mol Cell Biol 2001;2:721–730.
2. Maxfield FR, McGraw TE. Endocytic recycling. Nat Rev Mol Cell Biol
2004;5:121–132.
3. Wessels E, Simpson JC. Impact of live cell imaging on coated vesicle
research. Semin Cell Dev Biol 2007;18:412–423.
4. Bonifacino JS, Traub LM. Signals for sorting of transmembrane
proteins to endosomes and lysosomes. Annu Rev Biochem 2003;72:
395–447.
5. Rink J, Ghigo E, Kalaidzidis Y, Zerial M. Rab conversion as a mech-
anism of progression from early to late endosomes. Cell 2005;122:
735–749.
6. Lakadamyali M, Rust MJ, Zhuang X. Ligands for clathrin-mediated
endocytosis are differentially sorted into distinct populations of early
endosomes. Cell 2006;124:997–1009.
7. Matteoni R, Kreis TE. Translocation and clustering of endosomes and
lysosomes depends on microtubules. J Cell Biol 1987;105:1253–1265.
8. Geuze HJ, Slot JW, Strous GJ, Lodish HF, Schwartz AL. Intracellular
site of asialoglycoprotein receptor-ligand uncoupling: double-label
immunoelectron microscopy during receptor-mediated endocytosis.
Cell 1983;32:277–287.
9. Harding C, Levy MA, Stahl P. Morphological analysis of ligand uptake
and processing: the role of multivesicular endosomes and CURL in
receptor-ligand processing. Eur J Cell Biol 1985;36:230–238.
10. Hoepfner S, Severin F, Cabezas A, Habermann B, Runge A, Gillooly D,
Stenmark H, Zerial M. Modulation of receptor recycling and degrada-
tion by the endosomal kinesin KIF16B. Cell 2005;121:437–450.
11. Bananis E, Nath S, Gordon K, Satir P, Stockert RJ, Murray JW, Wolkoff
AW.Microtubule-dependentmovement of late endocytic vesicles in vitro:
requirements for dynein and kinesin. Mol Biol Cell 2004;15:3688–3697.
12. Wolkoff AW, Klausner RD, Ashwell G, Harford J. Intracellular segrega-
tion of asialoglycoproteins and their receptor: a prelysosomal event
subsequent to dissociation of the ligand-receptor complex. J Cell Biol
1984;98:375–381.
13. Breitfeld PP, McKinnon WC, Mostov KE. Effect of nocodazole on
vesicular traffic to the apical and basolateral surfaces of polarized
MDCK cells. J Cell Biol 1990;111:2365–2373.
14. Hunziker W, Male P, Mellman I. Differential microtubule requirements
for transcytosis in MDCK cells. EMBO J 1990;9:3515–3525.
15. Maples CJ, Ruiz WG, Apodaca G. Both microtubules and actin
filaments are required for efficient postendocytotic traffic of the poly-
meric immunoglobulin receptor in polarized Madin-Darby canine kidney
cells. J Biol Chem 1997;272:6741–6751.
16. Koval M, Pagano RE. Lipid recycling between the plasma membrane
and intracellular compartments: transport and metabolism of fluores-
cent sphingomyelin analogues in cultured fibroblasts. J Cell Biol 1989;
108:2169–2181.
17. McGraw TE, Dunn KW, Maxfield FR. Isolation of a temperature-
sensitive variant Chinese hamster ovary cell line with a morphologically
altered endocytic recycling compartment. J Cell Physiol 1993;155:
579–594.
18. Block MR, Glick BS, Wilcox CA, Wieland FT, Rothman JE. Purification
of an N-ethylmaleimide-sensitive protein catalyzing vesicular transport.
Proc Natl Acad Sci U S A 1988;85:7852–7856.
19. Spang A, Schekman R. Reconstitution of retrograde transport from the
Golgi to the ER in vitro. J Cell Biol 1998;143:589–599.
20. Bananis E, Murray JW, Stockert RJ, Satir P, Wolkoff AW. Microtubule
and motor-dependent endocytic vesicle sorting in vitro. J Cell Biol
2000;151:179–186.
21. Dunn KW, McGraw TE, Maxfield FR. Iterative fractionation of recycling
receptors from lysosomally destined ligands in an early sorting endo-
some. J Cell Biol 1989;109:3303–3314.
22. Huang T, Wolkoff AW, Stockert RJ. Adaptor heat shock protein
complex formation regulates trafficking of the asialoglycoprotein re-
ceptor. Am J Physiol Gastrointest Liver Physiol 2006;290:G369–G376.
23. Dikic I. Mechanisms controlling EGF receptor endocytosis and degra-
dation. Biochem Soc Trans 2003;31:1178–1181.
24. Renfrew CA, Hubbard AL. Degradation of epidermal growth factor
receptor in rat liver. Membrane topology through the lysosomal
pathway. J Biol Chem 1991;266:21265–21273.
25. Mukhopadhayay S, Ananthanarayanan M, Stieger B, Meier PJ, Suchy
FJ, Anwer MS. cAMP increases liver Naþ-taurocholate cotransport by
translocating transporter to plasma membranes. Am J Physiol 1997;
273:G842–G848.
26. Sarkar S, Bananis E, Nath S, Anwer MS, Wolkoff AW, Murray JW.
PKCzeta is required for microtubule-based motility of vesicles contain-
ing the ntcp transporter. Traffic 2006;7:1078–1091.
27. Marbet P, Rahner C, Stieger B, Landmann L. Quantitative microscopy
reveals 3D organization and kinetics of endocytosis in rat hepatocytes.
Microsc Res Tech 2006;69:693–707.
28. Herman B, Albertini DF. A time-lapse video image intensification
analysis of cytoplasmic organelle movements during endosome trans-
location. J Cell Biol 1984;98:565–576.
29. Goltz JS, Wolkoff AW, Novikoff PM, Stockert RJ, Satir P. A role for
microtubules in sorting endocytic vesicles in rat hepatocytes. Proc Natl
Acad Sci U S A 1992;89:7026–7030.
30. Stockert RJ. The asialoglycoprotein receptor: relationships be-
tween structure, function, and expression. Physiol Rev 1995;75:
591–609.
31. Driskell OJ, Mironov A, Allan VJ, Woodman PG. Dynein is required for
receptor sorting and the morphogenesis of early endosomes. Nat Cell
Biol 2007;9:113–120.
32. Murray JW, Bananis E, Wolkoff AW. Reconstitution of ATP-dependent
movement of endocytic vesicles along microtubules in vitro: an
oscillatory bidirectional process. Mol Biol Cell 2000;11:419–433.
33. Nath S, Bananis E, Sarkar S, Stockert RJ, Sperry AO, Murray JW,
Wolkoff AW. Kif5B and Kifc1 interact and are required for motility and
fission of early endocytic vesicles in mouse liver. Mol Biol Cell 2007;18:
1839–1849.
34. Vale RD, Reese TS, Sheetz MP. Identification of a novel force-
generating protein, kinesin, involved in microtubule-based motility. Cell
1985;42:39–50.
35. Endow SA, Higuchi H. A mutant of the motor protein kinesin that
moves in both directions on microtubules. Nature 2000;406:913–916.
36. Gill SR, Schroer TA, Szilak I, Steuer ER, Sheetz MP, Cleveland DW.
Dynactin, a conserved, ubiquitously expressed component of an
activator of vesicle motility mediated by cytoplasmic dynein. J Cell
Biol 1991;115:1639–1650.
37. Murray JW, Wolkoff AW. Assay of Rab4-dependent trafficking on
microtubules. Methods Enzymol 2005;403:92–107.
38. Ichikawa T, Yamada M, Homma D, Cherry RJ, Morrison IE, Kawato S.
Digital fluorescence imaging of trafficking of endosomes containing
low-density lipoprotein in brain astroglial cells. Biochem Biophys Res
Commun 2000;269:25–30.
846 Traffic 2008; 9: 833–847
Murray et al.

39. Petiot A, Faure J, Stenmark H, Gruenberg J. PI3P signaling regulates
receptor sorting but not transport in the endosomal pathway. J Cell Biol
2003;162:971–979.
40. Murphy RF. Analysis and isolation of endocytic vesicles by flow
cytometry and sorting: demonstration of three kinetically distinct
compartments involved in fluid-phase endocytosis. Proc Natl Acad
Sci U S A 1985;82:8523–8526.
41. van Dam EM, Ten Broeke T, Jansen K, Spijkers P, Stoorvogel W.
Endocytosed transferrin receptors recycle via distinct dynamin and
phosphatidylinositol 3-kinase-dependent pathways. J Biol Chem 2002;
277:48876–48883.
42. Malide D, Cushman SW. Morphological effects of wortmannin on the
endosomal system and GLUT4-containing compartments in rat adipose
cells. J Cell Sci 1997;110:2795–2806.
43. Kurashima K, Szabo EZ, Lukacs G, Orlowski J, Grinstein S. Endosomal
recycling of the Naþ/Hþ exchanger NHE3 isoform is regulated by the
phosphatidylinositol 3-kinase pathway. J Biol Chem 1998;273:20828–
20836.
44. Klopfenstein DR, Tomishige M, Stuurman N, Vale RD. Role of
phosphatidylinositol(4,5)bisphosphate organization in membrane trans-
port by the Unc104 kinesin motor. Cell 2002;109:347–358.
45. Dranoff JA, McClure M, Burgstahler AD, Denson LA, Crawford AR,
Crawford JM, Karpen SJ, Nathanson MH. Short-term regulation of bile
acid uptake by microfilament-dependent translocation of rat ntcp to the
plasma membrane. Hepatology 1999;30:223–229.
46. Krutzik PO, Clutter MR, Nolan GP. Coordinate analysis of murine
immune cell surface markers and intracellular phosphoproteins by flow
cytometry. J Immunol 2005;175:2357–2365.
47. Morell AG, Irvine RA, Sternlieb I, Scheinberg IH, Ashwell G.
Physical and chemical studies on ceruloplasmin. V. Metabolic stu-
dies on sialic acid-free ceruloplasmin in vivo. J Biol Chem 1968;243:
155–159.
48. Weigel PH, Yik JH. Glycans as endocytosis signals: the cases of the
asialoglycoprotein and hyaluronan/chondroitin sulfate receptors. Bio-
chim Biophys Acta 2002;1572:341–363.
49. Oka JA, Weigel PH. Recycling of the asialoglycoprotein receptor in
isolated rat hepatocytes. Dissociation of internalized ligand from
receptor occurs in two kinetically and thermally distinguishable com-
partments. J Biol Chem 1983;258:10253–10262.
50. Stockert RJ, Potvin B, Tao L, Stanley P, Wolkoff AW. Human hepatoma
cell mutant defective in cell surface protein trafficking. J Biol Chem
1995;270:16107–16113.
51. WatermanH, YardenY.Molecularmechanisms underlying endocytosis and
sorting of ErbB receptor tyrosine kinases. FEBS Lett 2001;490:142–152.
52. Luini A, Ragnini-Wilson A, Polishchuck RS, De Matteis MA. Large
pleiomorphic traffic intermediates in the secretory pathway. Curr Opin
Cell Biol 2005;17:353–361.
53. Mukherjee S, Ghosh RN, Maxfield FR. Endocytosis. Physiol Rev 1997;
77:759–803.
54. Cai H, Reinisch K, Ferro-Novick S. Coats, tethers, Rabs, and SNAREs
work together to mediate the intracellular destination of a transport
vesicle. Dev Cell 2007;12:671–682.
55. Bananis E, Murray JW, Stockert RJ, Satir P, Wolkoff AW. Regulation of
early endocytic vesicle motility and fission in a reconstituted system.
J Cell Sci 2003;116:2749–2761.
56. Dillman JF III, Pfister KK. Differential phosphorylation in vivo of
cytoplasmic dynein associated with anterogradely moving organelles.
J Cell Biol 1994;127:1671–1681.
57. Okada Y, Sato-Yoshitake R, Hirokawa N. The activation of protein
kinase A pathway selectively inhibits anterograde axonal transport of
vesicles but not mitochondria transport or retrograde transport in vivo.
J Neurosci 1995;15:3053–3064.
58. Stockert RJ, Haimes HB, Morell AG, Novikoff PM, Novikoff AB,
Quintana N, Sternlieb I. Endocytosis of asialoglycoprotein-enzyme
conjugates by hepatocytes. Lab Invest 1980;43:556–563.
59. Hudgin RL, Pricer WE Jr, Ashwell G, Stockert RJ, Morell AG. The
isolation and properties of a rabbit liver binding protein specific for
asialoglycoproteins. J Biol Chem 1974;249:5536–5543.
60. Murray JW, Bananis E, Wolkoff AW. Immunofluorescence micro-
chamber technique for characterizing isolated organelles. Anal Bio-
chem 2002;305:55–67.
61. Murray JW, Wolkoff AW. In vitro motility system to study the role of
motor proteins in receptor-ligand sorting. Methods Mol Biol 2007;392:
143–158.
Traffic 2008; 9: 833–847 847
Endocytic Segregation In Vitro
Copyright © 2022 FDOKUMEN




















