
Probing for primary functions of prohibitin in Trypanosoma brucei

EUKARYOTIC CELL, Aug. 2010, p. 1272–1282 Vol. 9, No. 81535-9778/10/$12.00 doi:10.1128/EC.00076-10Copyright © 2010, American Society for Microbiology. All Rights Reserved.
Cellular and Molecular Remodeling of the Endocytic Pathway duringDifferentiation of Trypanosoma brucei Bloodstream Forms�†
Benoit Vanhollebeke,1,2 Pierrick Uzureau,1 Daniel Monteyne,1David Perez-Morga,1 and Etienne Pays1*
Laboratory of Molecular Parasitology, Institute of Molecular Biology and Medicine, Universite Libre de Bruxelles, 12 rue desProfesseurs Jeener et Brachet, B-6041 Gosselies, Belgium,1 and Department of Biochemistry and Biophysics,
UCSF, 1550 Fourth Street, San Francisco, California 94158-23242
Received 26 March 2010/Accepted 15 June 2010
During the course of mammalian infection, African trypanosomes undergo extensive cellular differentiation,as actively dividing long slender (SL) forms progressively transform into intermediate (I) forms and finallyquiescent G1/G0-locked short stumpy (ST) forms. ST forms maintain adaptations compatible with theirsurvival in the mammalian bloodstream, such as high endocytic activity, but they already show preadaptationsto the insect midgut conditions. The nutritional requirements of ST forms must differ from those of SL formsbecause the ST forms stop multiplying. We report that the uptake of several ligands was reduced in ST formscompared with that in SL forms. In particular, the haptoglobin-hemoglobin (Hp-Hb) complex was no longertaken up due to dramatic downregulation of its cognate receptor, TbHpHbR. As this receptor also allowsuptake of trypanolytic particles from human serum, ST forms were resistant to trypanolysis by human serumlipoproteins. These observations allowed both flow cytometry analysis of SL-to-ST differentiation and thegeneration of homogeneous ST populations after positive selection upon exposure to trypanolytic particles. Inaddition, we observed that in ST forms the lysosome relocates anterior to the nucleus. Altogether, we identifiednovel morphological and molecular features that characterize SL-to-ST differentiation.
Trypanosoma brucei is a protozoan parasite of clinical, vet-erinary, and economical importance. It develops in the blood-stream of a variety of mammals that invariably die if left un-treated. Its division rate is high, with typical populationdoubling times ranging from 6 h in vitro to less than 3 h in vivo.If this process was uncontrolled, the parasite would rapidly killits host, with impaired transmission through the tsetse fly vec-tor. The parasite exhibits a surface coat made of an abundantand immunogenic antigen, termed VSG for variant surfaceglycoprotein (24). This coat triggers an efficient antitrypano-some humoral response, but this response is not sufficient tocontrol infection, as revealed from the rapid death of miceinfected with monomorphic trypanosomes in which growth isderegulated. Instead, it is the combination of parasite celldensity self-regulation and the immune response that shapesthe parasitemia in a succession of characteristic waves (26).
The process underlying the transformation of rapidly divid-ing long slender (SL) forms into G1/G0-arrested short stumpy(ST) forms is only partially understood. It has been proposedto depend on diffusible small metabolites that might triggercyclic AMP (cAMP)-dependent signaling (16). Different pro-tein kinases appear to be involved in a cascade that results instrong overall transcriptional repression with the specific acti-vation of some genes (1, 6, 8, 14, 31).
In T. brucei bloodstream forms, the endocytic rate is very
high (9). However, so far only a few surface receptors havebeen described. The transferrin (Tf) receptor is a heterodimerassociating the proteins encoded by two genes present ingenomic expression sites for VSG, termed ESAG7 and ESAG6(for expression site-associated genes 7 and 6, respectively)(25). While ESAG6 is anchored to the plasma membrane by aglycosylphosphatidylinositol (GPI) moiety, ESAG7 is not.Bloodstream forms also possess surface receptors for serumlipoproteins, because these parasites require exogenous sterol,mainly in the form of cholesterol, as a bulk membrane sterolconstituent (4, 32). Both low-density lipoprotein (LDL) andhigh-density lipoprotein (HDL), the major classes of serumlipoproteins, serve as primary sources of lipids (2, 4, 10, 32).However, the molecular identity of such receptors has re-mained elusive, and contradictory reports have been generatedregarding their substrate specificity and diversity. While somefavor a scenario in which HDL and LDL are engaged bydistinct surface molecules (2), others have provided evidencethat a single scavenger receptor might recognize both li-poprotein particles (10). Finally, a T. brucei haptoglobin-hemoglobin (Hp-Hb) surface receptor was discovered andtermed TbHpHbR (28). It is a GPI-anchored glycoproteinwith unknown structure, except for the presence of linearchains of poly-N-acetyllactosamine (pNAL), a hallmark ofendocytic proteins in T. brucei (21).
SL and ST forms are exposed to the same environmentalchallenges in terms of immune escape and antibody clearance.Therefore, in contrast with the marked differences distinguish-ing bloodstream and insect-specific procyclic forms (PFs), SLand ST forms share most endocytic characteristics (20). Forinstance, high Tf and membrane uptake rates were shown to bemaintained and even increased in ST forms (7, 20). However,
* Corresponding author. Mailing address: Laboratory of MolecularParasitology, IBMM, Universite Libre de Bruxelles, 12 rue des Pro-fesseurs Jeener et Brachet, B-6041 Gosselies, Belgium. Phone: 32 26509759. Fax: 32 2 6509750. E-mail: [email protected].
† Supplemental material for this article may be found at http://ec.asm.org/.
� Published ahead of print on 25 June 2010.
1272
on August 12, 2016 by guest
http://ec.asm.org/
Dow
nloaded from

the ability to divide clearly distinguishes these developmentalforms, and this difference might result in different require-ments for nutrients.
In order to investigate the adaptation of the endocytic sys-tem to the cellular differentiation between SL and ST forms,we undertook a systematic study of the regulation of liganduptake during this process. Our analyses revealed that while Tfand bulk phase uptake remain high in ST forms, the uptake ofHp-Hb and LDL was reduced. This was linked to reduction ofcell labeling with tomato lectin (TL), the pNAL-specific lectinthat can be used as a general marker for receptor-mediatedendocytosis in T. brucei (21). This inhibition of ligand uptakewas particularly dramatic in the case of Hp-Hb, the receptor ofwhich appeared to be absent from ST forms. Accordingly, STforms were resistant to the trypanosome lytic factor 1 (TLF-1),a potent toxic complex of human blood that enters the parasitethrough this receptor (28).
During the course of this analysis, we discovered anothercharacteristic of the endocytic apparatus of ST forms. Thelysosome was found to relocate to the anterior part of the cell,resulting in the surrounding of the nucleus by the endocyticsystem. Altogether, our results highlight important endocyticdifferences between SL and ST parasites, namely, remodelingof receptor-mediated endocytosis and relocation of the lyso-some. These observations could be useful to accurately dis-criminate and isolate ST forms, particularly in the first steps ofthe differentiation process.
MATERIALS AND METHODS
Trypanosome growth and differentiation. The AnTat 1.1E clone of Trypano-soma brucei brucei was used in all experiments. Intraperitoneal injections (100 �lof a suspension of 105 ml�1 parasites in phosphate-buffered saline glucose[PSGS]) were performed in 8-week-old female Swiss mice preinjected with cy-clophosphamide (300 mg kg of body weight�1). Parasites were purified frominfected blood by separation on DEAE cellulose. The procedure was miniatur-ized to allow the differentiation process to be followed in the same infected miceover several days. Sixty microliters of infected blood was harvested from the tailvein in capillary tubes with sodium heparin and applied to 1 ml of PSGS-equilibrated DEAE resin bed. Elution was performed in 1 ml PSGS solutionequilibrated at 4°C to minimize toxicity. Experiments were also validated afterpurification at 37°C to exclude cold shock-induced differentiation. Briefly, allmaterials and solutions were equilibrated at 37°C. Blood collection was per-formed, DEAE columns were used, and washes were performed in a 37°C heatedroom. Parasites were added to 37°C equilibrated medium for uptake experi-ments.
Ligand preparation and labeling. Alexa Fluor 488-labeled transferrin (Tf-A488; human holotransferrin) and lysine-fixable 40,000-molecular-weight (MW)dextran-fluorescein isothiocyanate (FITC) were from Molecular Probes, and Hpand tomato lectin-FITC were purchased from Sigma-Aldrich. Human HDL andLDL were prepared by sequential flotation ultracentrifugation. The purity of thefractions was over 98% as revealed by Coomassie blue staining of ApoA-I andApoB. Haptoglobin (Sigma-Aldrich), HDL, and LDL were labeled with theAlexa Fluor 488 protein labeling kit (Molecular Probes) according to the man-ufacturer’s instructions.
Flow cytometry and microscopic examinations. Cells (106 cells at 107 ml�1) inserum-free HMI-9 growth medium (12) supplemented with 1% bovine serumalbumin, 1% glucose, and 50 �g ml�1 human hemoglobin (Sigma-Aldrich) werepretreated for 30 min with the lysosomal protease inhibitor FMK-024 (17) whenindicated and incubated in the presence of FMK-024 and the indicated concen-tration of labeled ligand (25 �g ml�1 TL-FITC; 20 �g ml�1 Hp-A488; 100 �gml�1 LDL-A488 and HDL-A488; 20 �g ml�1 LDL-A488 and HDL-A488 forcompetition assays; 5 �g ml�1 Tf-A488 or 5 mg ml�1 dextran-FITC). Afteruptake, cells were washed in 1 ml phosphate-buffered saline (PBS) and fixed for10 min at room temperature in 0.1 ml 3.7% paraformaldehyde in PBS. Fixed cellswere resuspended at a density of 2 � 106 cells ml�1 in 10 mM Tris, pH 8, 0.15M NaCl solution and either analyzed with a FACSCanto II cytometer (BD
Biosciences) or spread on poly-L-lysine-coated slides. Coated cells were perme-abilized with 0.1% (vol/vol) Triton X-100 in Tris-buffered saline for 5 min atroom temperature. DNA was stained with DAPI (4�,6-diamidino-2-phenylin-dole). Lysosomes were immunodetected with monoclonal anti-p67 antibodies(mAb280; J. Bangs and D. Russel). EP1 and PAD1 (5) immunodetection wasperformed as described previously. Primary antibodies were detected with anAlexa 488-conjugated goat anti-mouse secondary antibody (Molecular Probes).Image processing was applied to all parts of the image and identically to allimages of the same figure assembly with Adobe Photoshop software. Thedistances between the kinetoplast, the lysosome, and the nucleus were estab-lished using ImageJ software after calibration with a micrometer slide. Forlive lysis phenotype examination, cells were immobilized by spreading con-centrated suspensions of cells on a slide coated with a thin layer of HMI-9containing 0.65% (wt/vol) agar at 30°C as described in reference 29. Differ-entiation in procyclic forms was triggered by incubating freshly isolated cellsin SDM-79 for 6 h at 27°C.
Morphological and biochemical analyses of slender, intermediate, and stumpyforms. All morphological and biochemical scoring assays were performedthrough blinded examinations. For NAD diaphorase assays, cells applied as acoating on poly-L-lysine slides were fixed for 5 min with 2.5% (vol/vol) glutaral-dehyde in PBS, rinsed in distilled water, and incubated for 2 h with freshlyprepared NAD diaphorase reaction mixture (1.3 mg ml�1 of reduced �-NADHdisodium salt and 0.3 mg ml�1 nitroblue tetrazolium dissolved in PBS) beforebeing washed in PBS and processed for microscopic examination. Morphologicaldiscrimination of different forms relied on integration of the following criteria:overall cell morphology and volume, length of protruding flagellum, distance ofthe flagellum from the cell body, and position of the nucleus relative to theposterior end of the cell.
Northern blot analysis. Total RNA was isolated using the Trizol reagent(Gibco BRL) from at least 108 cells freshly isolated from mice for bloodstreamforms and from in vitro cultures for procyclic forms. Ten-microgram sampleswere electrophoresed on a 1% (wt/vol) agarose-formaldehyde gel and trans-ferred to a Hybond-C nitrocellulose membrane (Amersham Biosciences). Blotswere hybridized with 32P-radiolabeled probes (Rediprime II random prime la-beling system; Amersham Biosciences) corresponding to the 580-bp TbHpHbRopen reading frame (ORF) 5� extremity. After hybridization, the blots werewashed three times with 0.1� SSC (1� SSC is 0.15 M NaCl, 0.015 M sodiumcitrate) for 45 min at 65°C. Ethidium bromide-stained rRNA served as loadingcontrol.
Western blot analysis. Western blots (Hybond-C Extra membranes; Amer-sham Biosciences) were incubated for 2 h with a rat polyclonal anti-TbHpHbRantibody (described in reference 28) or a mouse monoclonal antitubulin antibody(gift from K. Gull, Oxford University) in 150 mM NaCl, 0.5% (wt/vol) Tween 20,20 mM Tris-HCl (pH 7.5), and 1% nonfat milk. The secondary antibodies,peroxidase-conjugated monoclonal mouse anti-rat IgG (Serotec) and peroxi-dase-conjugated sheep anti-mouse IgG (Amersham Biosciences), respectively,were diluted in the same buffer, and the bound antibodies were detected bychemiluminescence (Amersham Biosciences). For anti-tubulin staining, eachlane was loaded with 1 � 106 cells. For anti-TbHpHbR staining, each lane wasloaded with protein fractions purified by binding to Hp-Hb resins and equivalentto 5 � 108 cells, as described in reference 28.
In vitro trypanolysis assays. TLF-1 and TLF-2 were prepared by gel filtrationon Superdex 200 (GE Healthcare). Trypanosomes isolated from mice wereincubated at densities between 1 � 105 and 1 � 106 ml�1 in HMI-9 medium(with 10% fetal calf serum [FCS]) at 37°C in a CO2-equilibrated incubator.Cell densities were established by triplicate counting using a Neubauerhemocytometer.
Electron microscopy studies. Bloodstream trypanosomes were fixed overnightat 4°C in 2.5% glutaraldehyde, 0.1 M cacodylate buffer (pH 7.2), and postfixed in2% OsO4 in the same buffer. After serial dehydration in increasing ethanolconcentrations, samples were embedded in agar 100 (Agar Scientific Ltd., UnitedKingdom) and left to polymerize for 3 days at 60°C. Ultrathin sections (50 to 70nm thick) were collected in Formvar-carbon-coated copper grids by using a LeicaEM UC6 ultramicrotome and stained with uranyl acetate and lead citrate. Ob-servations were made on a Tecnai 10 electron microscope (FEI), and imageswere captured with a MegaView II camera and processed with AnalySIS andAdobe Photoshop software.
RESULTS
Endocytosis rate of the known ligands of T. brucei. Welabeled the known T. brucei-specific ligands with fluorescent
VOL. 9, 2010 ENDOCYTIC REMODELING 1273
on August 12, 2016 by guest
http://ec.asm.org/
Dow
nloaded from
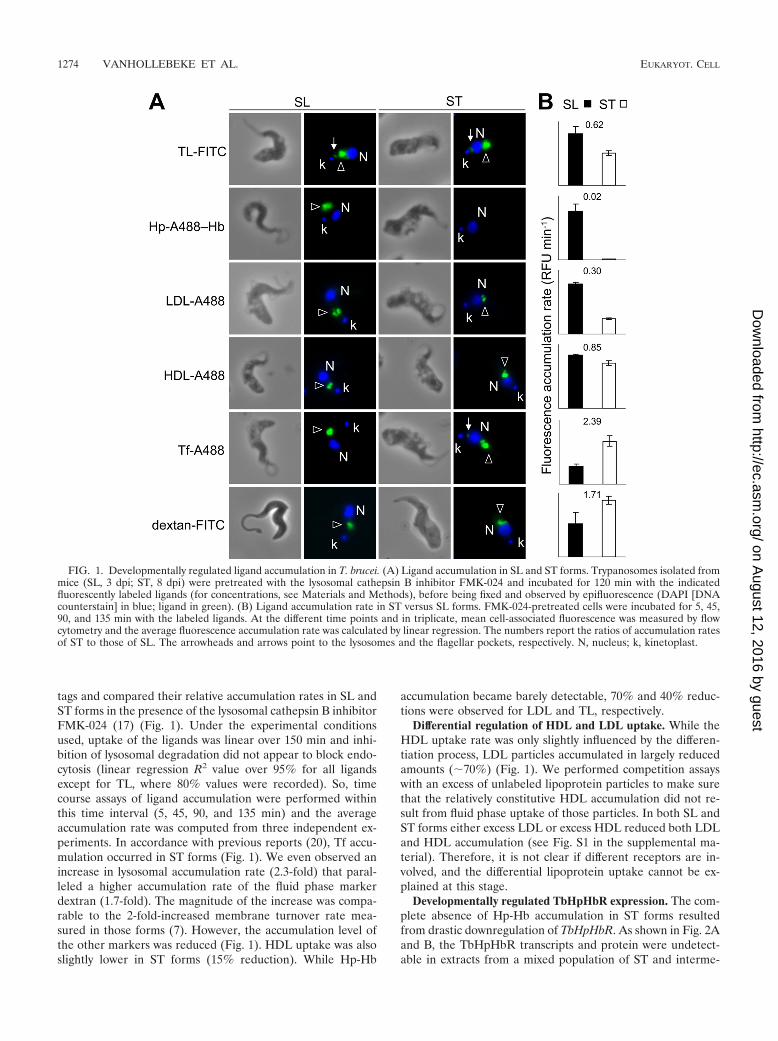
tags and compared their relative accumulation rates in SL andST forms in the presence of the lysosomal cathepsin B inhibitorFMK-024 (17) (Fig. 1). Under the experimental conditionsused, uptake of the ligands was linear over 150 min and inhi-bition of lysosomal degradation did not appear to block endo-cytosis (linear regression R2 value over 95% for all ligandsexcept for TL, where 80% values were recorded). So, timecourse assays of ligand accumulation were performed withinthis time interval (5, 45, 90, and 135 min) and the averageaccumulation rate was computed from three independent ex-periments. In accordance with previous reports (20), Tf accu-mulation occurred in ST forms (Fig. 1). We even observed anincrease in lysosomal accumulation rate (2.3-fold) that paral-leled a higher accumulation rate of the fluid phase markerdextran (1.7-fold). The magnitude of the increase was compa-rable to the 2-fold-increased membrane turnover rate mea-sured in those forms (7). However, the accumulation level ofthe other markers was reduced (Fig. 1). HDL uptake was alsoslightly lower in ST forms (15% reduction). While Hp-Hb
accumulation became barely detectable, 70% and 40% reduc-tions were observed for LDL and TL, respectively.
Differential regulation of HDL and LDL uptake. While theHDL uptake rate was only slightly influenced by the differen-tiation process, LDL particles accumulated in largely reducedamounts (�70%) (Fig. 1). We performed competition assayswith an excess of unlabeled lipoprotein particles to make surethat the relatively constitutive HDL accumulation did not re-sult from fluid phase uptake of those particles. In both SL andST forms either excess LDL or excess HDL reduced both LDLand HDL accumulation (see Fig. S1 in the supplemental ma-terial). Therefore, it is not clear if different receptors are in-volved, and the differential lipoprotein uptake cannot be ex-plained at this stage.
Developmentally regulated TbHpHbR expression. The com-plete absence of Hp-Hb accumulation in ST forms resultedfrom drastic downregulation of TbHpHbR. As shown in Fig. 2Aand B, the TbHpHbR transcripts and protein were undetect-able in extracts from a mixed population of ST and interme-
FIG. 1. Developmentally regulated ligand accumulation in T. brucei. (A) Ligand accumulation in SL and ST forms. Trypanosomes isolated frommice (SL, 3 dpi; ST, 8 dpi) were pretreated with the lysosomal cathepsin B inhibitor FMK-024 and incubated for 120 min with the indicatedfluorescently labeled ligands (for concentrations, see Materials and Methods), before being fixed and observed by epifluorescence (DAPI [DNAcounterstain] in blue; ligand in green). (B) Ligand accumulation rate in ST versus SL forms. FMK-024-pretreated cells were incubated for 5, 45,90, and 135 min with the labeled ligands. At the different time points and in triplicate, mean cell-associated fluorescence was measured by flowcytometry and the average fluorescence accumulation rate was calculated by linear regression. The numbers report the ratios of accumulation ratesof ST to those of SL. The arrowheads and arrows point to the lysosomes and the flagellar pockets, respectively. N, nucleus; k, kinetoplast.
1274 VANHOLLEBEKE ET AL. EUKARYOT. CELL
on August 12, 2016 by guest
http://ec.asm.org/
Dow
nloaded from
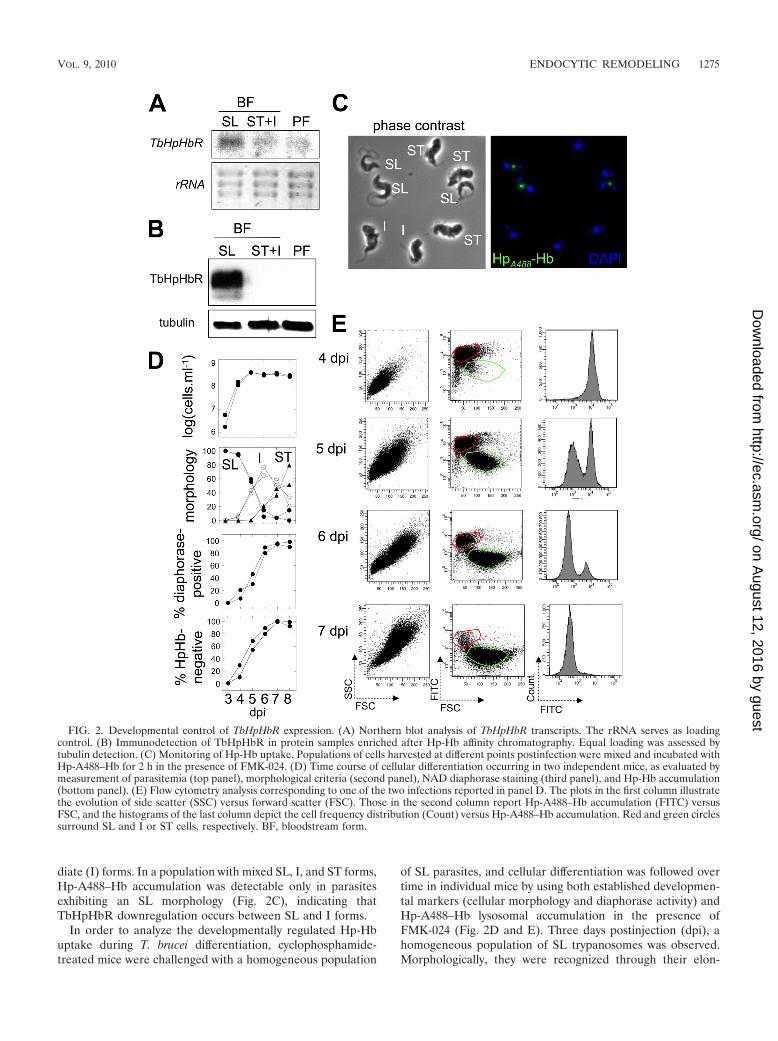
diate (I) forms. In a population with mixed SL, I, and ST forms,Hp-A488–Hb accumulation was detectable only in parasitesexhibiting an SL morphology (Fig. 2C), indicating thatTbHpHbR downregulation occurs between SL and I forms.
In order to analyze the developmentally regulated Hp-Hbuptake during T. brucei differentiation, cyclophosphamide-treated mice were challenged with a homogeneous population
of SL parasites, and cellular differentiation was followed overtime in individual mice by using both established developmen-tal markers (cellular morphology and diaphorase activity) andHp-A488–Hb lysosomal accumulation in the presence ofFMK-024 (Fig. 2D and E). Three days postinjection (dpi), ahomogeneous population of SL trypanosomes was observed.Morphologically, they were recognized through their elon-
FIG. 2. Developmental control of TbHpHbR expression. (A) Northern blot analysis of TbHpHbR transcripts. The rRNA serves as loadingcontrol. (B) Immunodetection of TbHpHbR in protein samples enriched after Hp-Hb affinity chromatography. Equal loading was assessed bytubulin detection. (C) Monitoring of Hp-Hb uptake. Populations of cells harvested at different points postinfection were mixed and incubated withHp-A488–Hb for 2 h in the presence of FMK-024. (D) Time course of cellular differentiation occurring in two independent mice, as evaluated bymeasurement of parasitemia (top panel), morphological criteria (second panel), NAD diaphorase staining (third panel), and Hp-Hb accumulation(bottom panel). (E) Flow cytometry analysis corresponding to one of the two infections reported in panel D. The plots in the first column illustratethe evolution of side scatter (SSC) versus forward scatter (FSC). Those in the second column report Hp-A488–Hb accumulation (FITC) versusFSC, and the histograms of the last column depict the cell frequency distribution (Count) versus Hp-A488–Hb accumulation. Red and green circlessurround SL and I or ST cells, respectively. BF, bloodstream form.
VOL. 9, 2010 ENDOCYTIC REMODELING 1275
on August 12, 2016 by guest
http://ec.asm.org/
Dow
nloaded from
gated cell shape, with a flagellum tightly apposed to the cellbody and protruding largely from the cell anterior part. Bio-chemically, they were negative for diaphorase staining. As re-ported earlier (28), those parasites accumulated high levels ofHp-Hb (Fig. 2E). As infection proceeded, the proportion of SLparasites gradually decreased in favor of I forms (peak on 6dpi) that transformed into fully differentiated ST forms by 7and 8 dpi. Diaphorase staining became detectable in I forms(Fig. 2D). As shown in Fig. 2C to E, I forms were negative forHp-Hb staining. Accumulation of Hp-Hb allowed the earlysteps of cellular differentiation to be followed quantitatively byflow cytometry. At 4 dpi a homogeneous population of stronglylabeled cells was observed. At 5 dpi (thus, 24 h before theappearance of fully differentiated ST forms), two resolved clus-ters could be recognized when cell size and fluorescence wereprobed on the flow cytometer (Fig. 2E). The percentage ofunlabeled cells expanded in the following days, to representalmost 100% of the population by 7 dpi. This observation pavesthe way toward the purification of homogeneous populationsof I forms by fluorescence-activated cell sorting (FACS). In-deed, while the side scatter-forward scatter (SSC-FSC) scatterplot allows the detection of the gradual increase in cell com-plexity and cell size along the differentiation process (the meanSSC and FSC values of SL forms are lower than those of STforms), it does not allow discrimination between the two pop-ulations (Fig. 2E, first column). When the sole Hp-A488–Hblysosomal accumulation is considered, two overlapping peakscan be observed (Fig. 2E, last column). But when both cell size(FSC channel) and ligand accumulation (FITC channel) areplotted, then long SL forms (red circles) can be safely discrim-
inated from either I or ST forms (green circles) (Fig. 2E,middle column).
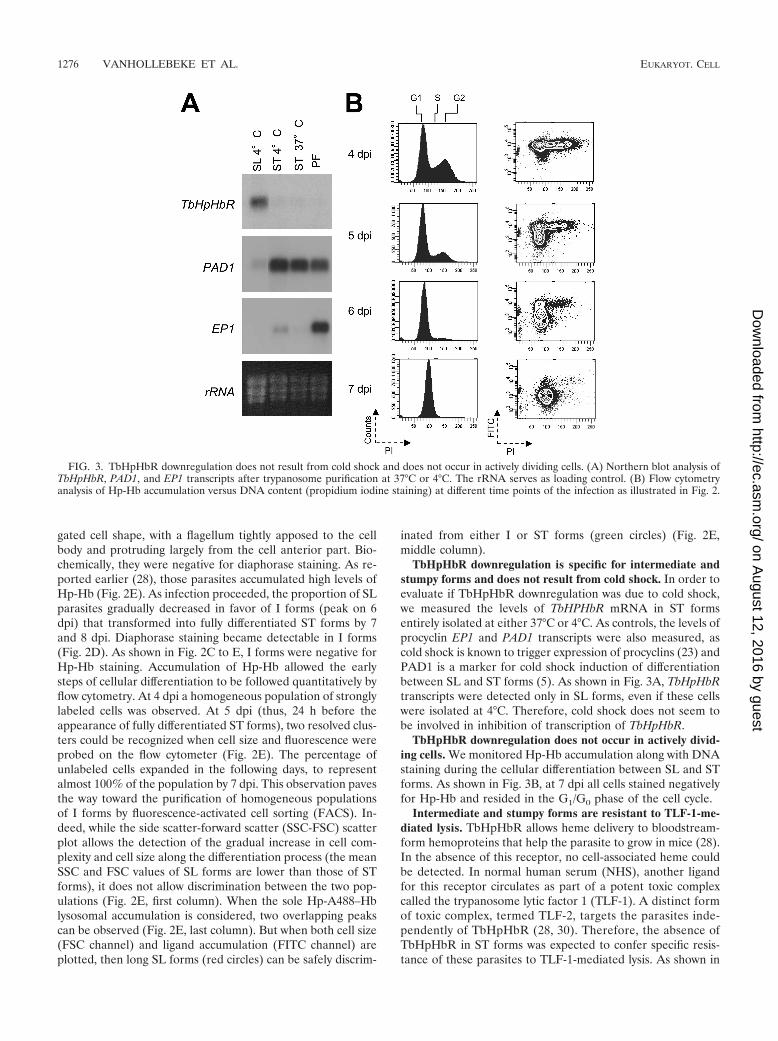
TbHpHbR downregulation is specific for intermediate andstumpy forms and does not result from cold shock. In order toevaluate if TbHpHbR downregulation was due to cold shock,we measured the levels of TbHPHbR mRNA in ST formsentirely isolated at either 37°C or 4°C. As controls, the levels ofprocyclin EP1 and PAD1 transcripts were also measured, ascold shock is known to trigger expression of procyclins (23) andPAD1 is a marker for cold shock induction of differentiationbetween SL and ST forms (5). As shown in Fig. 3A, TbHpHbRtranscripts were detected only in SL forms, even if these cellswere isolated at 4°C. Therefore, cold shock does not seem tobe involved in inhibition of transcription of TbHpHbR.
TbHpHbR downregulation does not occur in actively divid-ing cells. We monitored Hp-Hb accumulation along with DNAstaining during the cellular differentiation between SL and STforms. As shown in Fig. 3B, at 7 dpi all cells stained negativelyfor Hp-Hb and resided in the G1/G0 phase of the cell cycle.
Intermediate and stumpy forms are resistant to TLF-1-me-diated lysis. TbHpHbR allows heme delivery to bloodstream-form hemoproteins that help the parasite to grow in mice (28).In the absence of this receptor, no cell-associated heme couldbe detected. In normal human serum (NHS), another ligandfor this receptor circulates as part of a potent toxic complexcalled the trypanosome lytic factor 1 (TLF-1). A distinct formof toxic complex, termed TLF-2, targets the parasites inde-pendently of TbHpHbR (28, 30). Therefore, the absence ofTbHpHbR in ST forms was expected to confer specific resis-tance of these parasites to TLF-1-mediated lysis. As shown in
FIG. 3. TbHpHbR downregulation does not result from cold shock and does not occur in actively dividing cells. (A) Northern blot analysis ofTbHpHbR, PAD1, and EP1 transcripts after trypanosome purification at 37°C or 4°C. The rRNA serves as loading control. (B) Flow cytometryanalysis of Hp-Hb accumulation versus DNA content (propidium iodine staining) at different time points of the infection as illustrated in Fig. 2.
1276 VANHOLLEBEKE ET AL. EUKARYOT. CELL
on August 12, 2016 by guest
http://ec.asm.org/
Dow
nloaded from
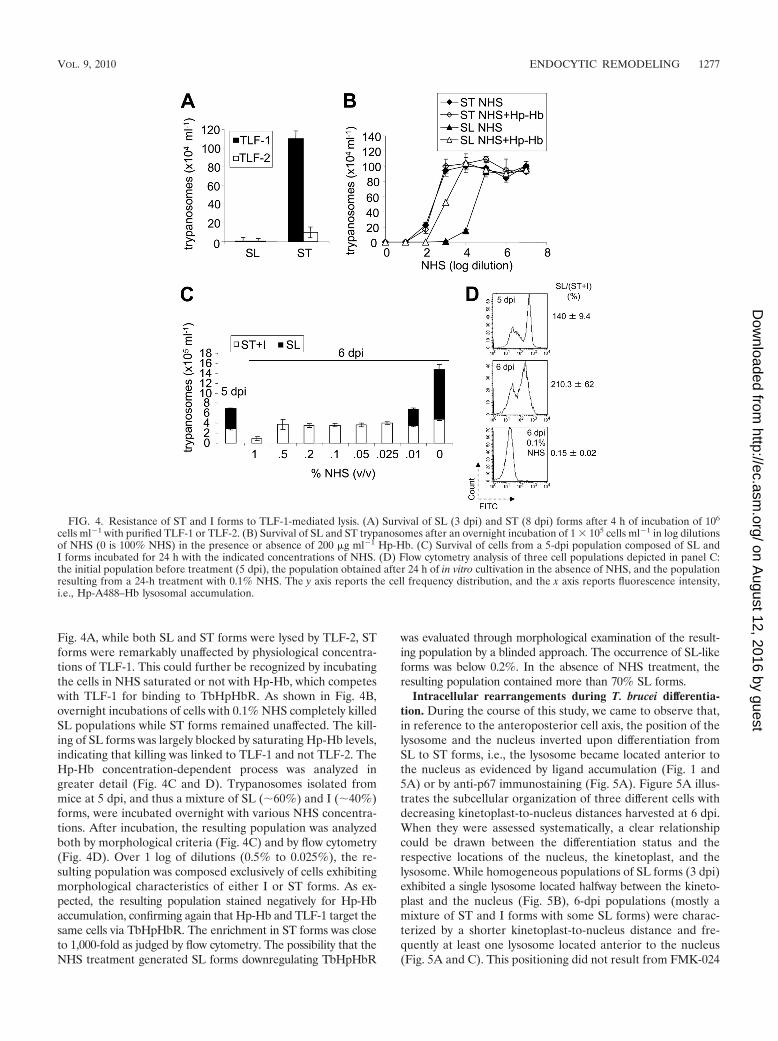
Fig. 4A, while both SL and ST forms were lysed by TLF-2, STforms were remarkably unaffected by physiological concentra-tions of TLF-1. This could further be recognized by incubatingthe cells in NHS saturated or not with Hp-Hb, which competeswith TLF-1 for binding to TbHpHbR. As shown in Fig. 4B,overnight incubations of cells with 0.1% NHS completely killedSL populations while ST forms remained unaffected. The kill-ing of SL forms was largely blocked by saturating Hp-Hb levels,indicating that killing was linked to TLF-1 and not TLF-2. TheHp-Hb concentration-dependent process was analyzed ingreater detail (Fig. 4C and D). Trypanosomes isolated frommice at 5 dpi, and thus a mixture of SL (�60%) and I (�40%)forms, were incubated overnight with various NHS concentra-tions. After incubation, the resulting population was analyzedboth by morphological criteria (Fig. 4C) and by flow cytometry(Fig. 4D). Over 1 log of dilutions (0.5% to 0.025%), the re-sulting population was composed exclusively of cells exhibitingmorphological characteristics of either I or ST forms. As ex-pected, the resulting population stained negatively for Hp-Hbaccumulation, confirming again that Hp-Hb and TLF-1 target thesame cells via TbHpHbR. The enrichment in ST forms was closeto 1,000-fold as judged by flow cytometry. The possibility that theNHS treatment generated SL forms downregulating TbHpHbR
was evaluated through morphological examination of the result-ing population by a blinded approach. The occurrence of SL-likeforms was below 0.2%. In the absence of NHS treatment, theresulting population contained more than 70% SL forms.
Intracellular rearrangements during T. brucei differentia-tion. During the course of this study, we came to observe that,in reference to the anteroposterior cell axis, the position of thelysosome and the nucleus inverted upon differentiation fromSL to ST forms, i.e., the lysosome became located anterior tothe nucleus as evidenced by ligand accumulation (Fig. 1 and5A) or by anti-p67 immunostaining (Fig. 5A). Figure 5A illus-trates the subcellular organization of three different cells withdecreasing kinetoplast-to-nucleus distances harvested at 6 dpi.When they were assessed systematically, a clear relationshipcould be drawn between the differentiation status and therespective locations of the nucleus, the kinetoplast, and thelysosome. While homogeneous populations of SL forms (3 dpi)exhibited a single lysosome located halfway between the kineto-plast and the nucleus (Fig. 5B), 6-dpi populations (mostly amixture of ST and I forms with some SL forms) were charac-terized by a shorter kinetoplast-to-nucleus distance and fre-quently at least one lysosome located anterior to the nucleus(Fig. 5A and C). This positioning did not result from FMK-024
FIG. 4. Resistance of ST and I forms to TLF-1-mediated lysis. (A) Survival of SL (3 dpi) and ST (8 dpi) forms after 4 h of incubation of 106
cells ml�1 with purified TLF-1 or TLF-2. (B) Survival of SL and ST trypanosomes after an overnight incubation of 1 � 105 cells ml�1 in log dilutionsof NHS (0 is 100% NHS) in the presence or absence of 200 �g ml�1 Hp-Hb. (C) Survival of cells from a 5-dpi population composed of SL andI forms incubated for 24 h with the indicated concentrations of NHS. (D) Flow cytometry analysis of three cell populations depicted in panel C:the initial population before treatment (5 dpi), the population obtained after 24 h of in vitro cultivation in the absence of NHS, and the populationresulting from a 24-h treatment with 0.1% NHS. The y axis reports the cell frequency distribution, and the x axis reports fluorescence intensity,i.e., Hp-A488–Hb lysosomal accumulation.
VOL. 9, 2010 ENDOCYTIC REMODELING 1277
on August 12, 2016 by guest
http://ec.asm.org/
Dow
nloaded from

treatment, as was confirmed by anti-p67 staining on untreatedcells (data not shown). In some developmental forms wherethe kinetoplast-to-nucleus distance was intermediate (around2.5 �m), two vesicles were decorated with anti-p67 antibodiesand accumulated Tf; thus, two apparent lysosomes were ob-served (Fig. 5A). The possibility that the observed repositioningof the lysosome resulted from fixation artifacts was ruled outthrough the observation of the swelling lysosome of live but im-mobilized ST forms treated with TLF-2 (Fig. 5D). Under thisexperimental setting, the swollen lysosome appeared anterior tothe nucleus, in contrast with observations of SL forms (29).
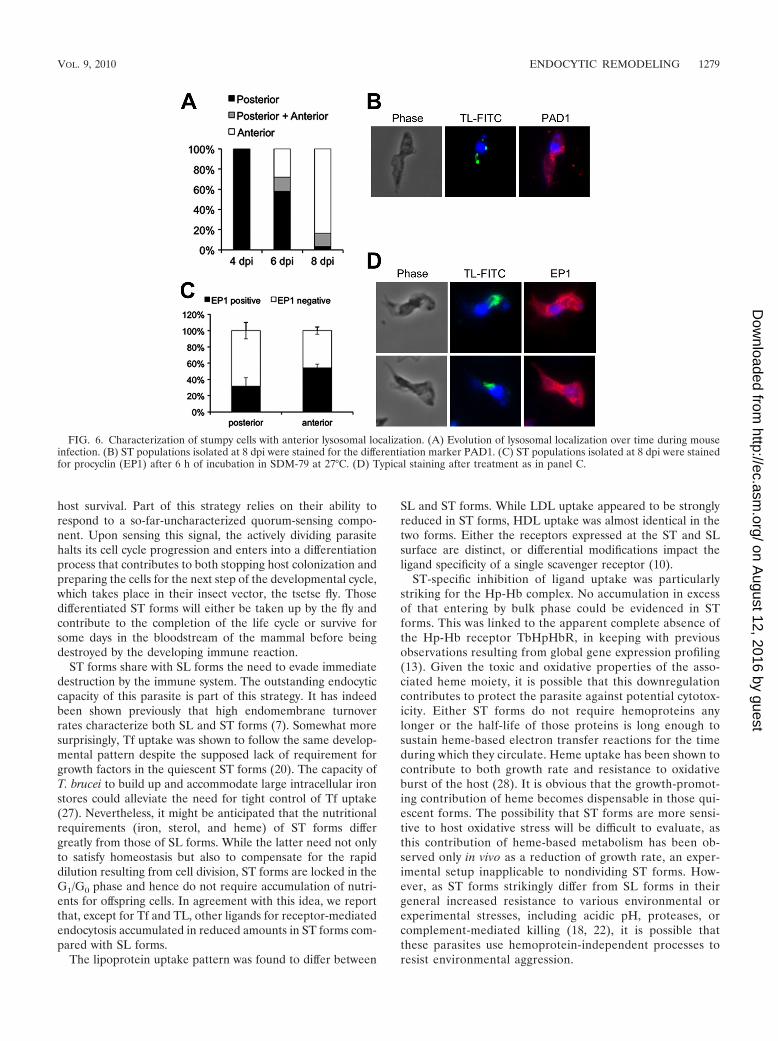
The evolution of lysosomal localization was followed overtime during mouse infection. As shown in Fig. 6A, cells withanterior lysosome localization clearly accumulated along withthe differentiation process, finally representing the majority ofthe differentiated population. Such cells (�95%) acquired ST-specific PAD1 expression (Fig. 6B). Moreover, upon triggeringof cellular differentiation into PFs, cells with anterior lysosomelocalization exhibited even better expression of EP1 procyclin
than did cells with posterior localization (Fig. 6C and D).Altogether, these results indicated that the movement of thelysosome toward the anterior compartment does not resultfrom experimental artifact and is a genuine feature of thecellular differentiation into ST forms.
The lysosome reorganization was detailed using transmis-sion electron microscopy analysis (15). On the way from itsoriginal location in SL forms to the cell anterior compartmentin ST forms, the lysosome traveled all around the nucleus (Fig.7). Interestingly, during this movement the lysosome mem-brane seemed to closely associate with that of the expandingmitochondrion (Fig. 7B). This analysis also confirmed the dif-ferentiation-linked increase of the lysosome size, which wasalready noted (3).
DISCUSSION
In order to remain durably in their mammalian hosts, try-panosomes keep their densities within limits acceptable for
FIG. 5. Subcellular reorganization during T. brucei differentiation. (A) Tf accumulation (5 �g ml�1, 120 min) and p67 (lysosomal marker)immunolocalization in a 6-dpi FMK-024-treated trypanosome population. “d(k-N)” represents the distance between the kinetoplast and thenucleus, along the virtual curve depicted by the white dotted line. (B) Distribution of the distance between the kinetoplast and the lysosome[d(k-L); y axis] and the distance between the kinetoplast and the nucleus (x axis) of a population of SL cells harvested at 3 dpi. Each cross representsan individual cell. (C) Same as in panel B, with a population harvested at 6 dpi. Each cross represents an individual cell. The colored circles referto the cells displaying two lysosomes as revealed by anti-p67 staining and Tf accumulation [two d(k-L) values of identical color for a single d(k-N)value]. (D) Phase-contrast imaging of immobilized live ST forms incubated or not with TLF-2. Arrows and arrowheads point to the swellinglysosome (L) and nucleus (N), respectively.
1278 VANHOLLEBEKE ET AL. EUKARYOT. CELL
on August 12, 2016 by guest
http://ec.asm.org/
Dow
nloaded from
host survival. Part of this strategy relies on their ability torespond to a so-far-uncharacterized quorum-sensing compo-nent. Upon sensing this signal, the actively dividing parasitehalts its cell cycle progression and enters into a differentiationprocess that contributes to both stopping host colonization andpreparing the cells for the next step of the developmental cycle,which takes place in their insect vector, the tsetse fly. Thosedifferentiated ST forms will either be taken up by the fly andcontribute to the completion of the life cycle or survive forsome days in the bloodstream of the mammal before beingdestroyed by the developing immune reaction.
ST forms share with SL forms the need to evade immediatedestruction by the immune system. The outstanding endocyticcapacity of this parasite is part of this strategy. It has indeedbeen shown previously that high endomembrane turnoverrates characterize both SL and ST forms (7). Somewhat moresurprisingly, Tf uptake was shown to follow the same develop-mental pattern despite the supposed lack of requirement forgrowth factors in the quiescent ST forms (20). The capacity ofT. brucei to build up and accommodate large intracellular ironstores could alleviate the need for tight control of Tf uptake(27). Nevertheless, it might be anticipated that the nutritionalrequirements (iron, sterol, and heme) of ST forms differgreatly from those of SL forms. While the latter need not onlyto satisfy homeostasis but also to compensate for the rapiddilution resulting from cell division, ST forms are locked in theG1/G0 phase and hence do not require accumulation of nutri-ents for offspring cells. In agreement with this idea, we reportthat, except for Tf and TL, other ligands for receptor-mediatedendocytosis accumulated in reduced amounts in ST forms com-pared with SL forms.
The lipoprotein uptake pattern was found to differ between
SL and ST forms. While LDL uptake appeared to be stronglyreduced in ST forms, HDL uptake was almost identical in thetwo forms. Either the receptors expressed at the ST and SLsurface are distinct, or differential modifications impact theligand specificity of a single scavenger receptor (10).
ST-specific inhibition of ligand uptake was particularlystriking for the Hp-Hb complex. No accumulation in excessof that entering by bulk phase could be evidenced in STforms. This was linked to the apparent complete absence ofthe Hp-Hb receptor TbHpHbR, in keeping with previousobservations resulting from global gene expression profiling(13). Given the toxic and oxidative properties of the asso-ciated heme moiety, it is possible that this downregulationcontributes to protect the parasite against potential cytotox-icity. Either ST forms do not require hemoproteins anylonger or the half-life of those proteins is long enough tosustain heme-based electron transfer reactions for the timeduring which they circulate. Heme uptake has been shown tocontribute to both growth rate and resistance to oxidativeburst of the host (28). It is obvious that the growth-promot-ing contribution of heme becomes dispensable in those qui-escent forms. The possibility that ST forms are more sensi-tive to host oxidative stress will be difficult to evaluate, asthis contribution of heme-based metabolism has been ob-served only in vivo as a reduction of growth rate, an exper-imental setup inapplicable to nondividing ST forms. How-ever, as ST forms strikingly differ from SL forms in theirgeneral increased resistance to various environmental orexperimental stresses, including acidic pH, proteases, orcomplement-mediated killing (18, 22), it is possible thatthese parasites use hemoprotein-independent processes toresist environmental aggression.
FIG. 6. Characterization of stumpy cells with anterior lysosomal localization. (A) Evolution of lysosomal localization over time during mouseinfection. (B) ST populations isolated at 8 dpi were stained for the differentiation marker PAD1. (C) ST populations isolated at 8 dpi were stainedfor procyclin (EP1) after 6 h of incubation in SDM-79 at 27°C. (D) Typical staining after treatment as in panel C.
VOL. 9, 2010 ENDOCYTIC REMODELING 1279
on August 12, 2016 by guest
http://ec.asm.org/
Dow
nloaded from

The absence of TbHpHbR explains the observed resistanceof early and late ST forms to the action of trypanolytic HDLsfrom NHS, since it is known that these particles enter theparasite through TbHpHbR (28). The documented sensitivityof ST forms to NHS appears to be entirely due to the TLF-2component, which enters trypanosomes independently fromTbHpHbR. Previous studies have reported on the increasedresistance of ST forms to TLF-1-mediated lysis (19). In thatreport, while in some assays ST forms were resistant to TLF-1-mediated lysis at 37°C as reported here, in others, ST formswere found to remain sensitive, unless forced to differentiatefurther by a mild cold shock treatment. In our analysis we did
not observe this instability of the TLF-1 phenotype, as in allexperiments ST forms were found to be fully resistant evenwhen the whole experiment from cell isolation from mice tocell fixation was entirely performed at 37°C, with preheatedsolutions and in thermostatic rooms (data not shown). Wespeculate that for some strains like TREU 667, used in refer-ence 19, a mild cold shock might sometimes be required toachieve full downregulation of TbHpHbR, while in otherstrains like AnTat 1.1, used in this study, the downregulationcould be more robust. The downregulation is likely to occur invivo, as TLF-1 injection in mice killed SL but not ST forms(data not shown).
FIG. 7. Transmission electron microscope analysis of SL-to-ST differentiation. Trypanosomes at 8 dpi were processed for transmission electronmicroscopy analysis. The posterior end of the cell is to the right of the pictures. (A) Representative micrographs of cells with an anterior lysosome,the dominant cellular organization (80.6% of the population, n � 31). (B) Less frequent cellular organizations. (Left) Posterior localization(probably relapsing SL cell; 3.2% of the population). (Center) Nuclear proximal posterior lysosome (3.2% of the population). (Right) Cellharboring both an anterior and a posterior lysosome (12.9% of the population). Arrows point to the mitochondrion, closely associated with thelysosome. L, lysosome; N, nucleus; FP, flagellar pocket. Scale bar, 1 �m.
1280 VANHOLLEBEKE ET AL. EUKARYOT. CELL
on August 12, 2016 by guest
http://ec.asm.org/
Dow
nloaded from

The downregulation of TbHpHbR was shown to result fromdecreased mRNA levels. This paralleled the transition be-tween dividing SL forms and the initiation of differentiationinto ST forms. In particular, I forms did not seem to expressthe receptor any longer. This dramatic downregulation mightbe linked to the location of the TbHpHbR gene at the very endof a transcriptional unit that immediately precedes a transcrip-tion unit for the procyclic form-specific procyclin genes (28).Indeed, strong transcriptional changes are linked to SL-to-STdifferentiation, including partial activation of procyclin loci(1, 23). Therefore, it is possible that enhanced recruitment ofRNA polymerase at the procyclin promoter could hinderproper transcription elongation in the TbHpHbR gene locatedin the close upstream environment. However, although in STcold shock triggered the synthesis of procyclin mRNAs, thedownregulation of TbHpHbR did not require cold shock. Asthis downregulation was not observed in dividing cells, it couldbe linked to the cell cycle blockade typical of early SL-to-STdifferentiation.
The absence of TbHpHbR in early ST forms offers newavenues to study the events triggering T. brucei differentiationin the bloodstream. So far, this is poorly characterized due tothe impossibility of isolating and profiling homogeneous pop-ulations of cells at the initiation of this process. Currentlyavailable techniques require plating SL forms at high densitiesin vitro, a situation that might not fully reproduce the endog-enous conditions. In vivo, the differentiation is not fully syn-chronous, as can be appreciated from Fig. 2D (except for 3 dpi,the population is always composed of at least two differentforms). This leaves the investigator with two types of relativelyhomogeneous differentiated populations: either a populationrich in I forms but containing both SL and ST forms (6 dpi) ora population of ST forms (8 dpi) sometimes contaminated withrelapsing SL forms. In both cases, cell analysis at the onset ofthe differentiation process is intractable for a population.mRNA profiling of homogeneous trypanosome populationsbetween ST and procyclic forms has allowed major advances inour understanding of the differentiation between bloodstreamand insect-specific forms (5, 14). As we have shown that ahomogeneous population of differentiated forms (most inter-estingly, early I forms) can be obtained either by exposure tocarefully titrated amounts of NHS or through FACS, the toolsdeveloped in this work should allow a similar approach to beundertaken for the SL-to-ST differentiation. Two options areavailable, either the FACS of the cells after Hp-A488–Hbuptake or the counterselection of SL forms with TLF-1. Al-though ST and I forms seemed unaffected by the overnighttreatment with TLF-1, we favor the first option because it isnot known whether mild TLF-1 treatment or overnight culturemight affect cell development, and conversely the Hp-Hb ac-cumulation can be completed in less than 2 h. More generally,Hp-Hb uptake represents a new and easy developmentalmarker, which together with diaphorase activity, mitochondrialelaboration, and morphological modifications could be used toassess early steps of the differentiation process.
Finally, we observed a so-far-undisclosed feature of the SL-to-ST differentiation process. Reciprocal translocation of thelysosome and the nucleus coincided with the appearance oftypical features of ST forms, such as characteristic flagellumlength, PAD1 immunoreactivity, and diaphorase staining in-
tensity. Therefore, we propose that this inversion is a hallmarkof the terminally differentiated ST forms. Curiously, the lyso-some movement seemed to be in concert with that of theexpanding mitochondrion, as the two organelles appeared tobe closely associated. Presumably concerted, rather than inde-pendent, segregation is more efficient and energetically morefavorable. This coupled translocation might result from theincreased size of the two organelles, which might not fit anylonger the narrowing space between the kinetoplast and thenucleus. The significance of this cytological reorganization isnot obvious, but one possibility is that this positioning couldincrease the rate of intracellular organelle autophagy uponinitiation of the differentiation into procyclic forms (11).
ACKNOWLEDGMENTS
This work was supported by the Belgian Fonds National de la Re-cherche Scientifique (FNRS) and the Interuniversity Attraction PolesProgramme-Belgian Science Policy. During the course of this workB.V. was a research fellow of the FNRS.
We thank K. Matthews (University of Edinburgh) for anti-PAD1antibodies and the Brachet Fund for help.
REFERENCES
1. Amiguet-Vercher, A., D. Perez-Morga, A. Pays, P. Poelvoorde, H. V. Xong, P.Tebabi, L. Vanhamme, and E. Pays. 2004. Loss of the mono-allelic control ofthe VSG expression sites during the development of Trypanosoma brucei inthe bloodstream. Mol. Microbiol. 51:1577–1588.
2. Bastin, P., A. Stephan, J. Raper, J. M. Saint-Remy, F. R. Opperdoes, andP. J. Courtoy. 1996. An M(r) 145,000 low-density lipoprotein (LDL)-bindingprotein is conserved throughout the Kinetoplastida order. Mol. Biochem.Parasitol. 76:43–56.
3. Brown, R. C., D. A. Evans, and K. Vickerman. 1972. Developmental changesin ultrastructure and physiology of Trypanosoma brucei. Trans. R. Soc. Trop.Med. Hyg. 66:336–337.
4. Coppens, I., P. Baudhuin, F. R. Opperdoes, and P. J. Courtoy. 1988. Re-ceptors for the host low density lipoproteins on the hemoflagellate Trypano-soma brucei: purification and involvement in the growth of the parasite. Proc.Natl. Acad. Sci. U. S. A. 85:6753–6757.
5. Dean, S., R. Marchetti, K. Kirk, and K. R. Matthews. 2009. A surfacetransporter family conveys the trypanosome differentiation signal. Nature459:213–217.
6. Domenicali Pfister, D., G. Burkard, S. Morand, C. K. Renggli, I. Roditi, andE. Vassella. 2006. A mitogen-activated protein kinase controls differentiationof bloodstream forms of Trypanosoma brucei. Eukaryot. Cell 5:1126–1135.
7. Engstler, M., T. Pfohl, S. Herminghaus, M. Boshart, G. Wiegertjes, N.Heddergott, and P. Overath. 2007. Hydrodynamic flow-mediated proteinsorting on the cell surface of trypanosomes. Cell 131:505–515.
8. Fenn, K., and K. R. Matthews. 2007. The cell biology of Trypanosoma bruceidifferentiation. Curr. Opin. Microbiol. 10:539–546.
9. Field, M. C., J. H. Lumb, V. O. Adung’a, N. G. Jones, and M. Engstler. 2009.Macromolecular trafficking and immune evasion in African trypanosomes.Int. Rev. Cell Mol. Biol. 278:1–67.
10. Green, H. P., M. Del Pilar Molina Portela, E. N. St. Jean, E. B. Lugli, andJ. Raper. 2003. Evidence for a Trypanosoma brucei lipoprotein scavengerreceptor. J. Biol. Chem. 278:422–427.
11. Herman, M., D. Perez-Morga, N. Schtickzelle, and P. A. Michels. 2008.Turnover of glycosomes during life-cycle differentiation of Trypanosomabrucei. Autophagy 4:294–308.
12. Hirumi, H., and K. Hirumi. 1989. Continuous cultivation of Trypanosomabrucei blood stream forms in a medium containing a low concentration ofserum protein without feeder cell layers. J. Parasitol. 75:985–989.
13. Jensen, B. C., D. Sivam, C. T. Kifer, P. J. Myler, and M. Parsons. 2009.Widespread variation in transcript abundance within and across develop-mental stages of Trypanosoma brucei. BMC Genomics 10:482.
14. Kabani, S., K. Fenn, A. Ross, A. Ivens, T. K. Smith, P. Ghazal, and K.Matthews. 2009. Genome-wide expression profiling of in vivo-derived blood-stream parasite stages and dynamic analysis of mRNA alterations duringsynchronous differentiation in Trypanosoma brucei. BMC Genomics 10:427.
15. Langreth, S. G., and A. E. Balber. 1975. Protein uptake and digestion inbloodstream and culture forms of Trypanosoma brucei. J. Protozool. 22:40–53.
16. Laxman, S., A. Riechers, M. Sadilek, F. Schwede, and J. A. Beavo. 2006.Hydrolysis products of cAMP analogs cause transformation of Trypanosomabrucei from slender to stumpy-like forms. Proc. Natl. Acad. Sci. U. S. A.103:19194–19199.
VOL. 9, 2010 ENDOCYTIC REMODELING 1281
on August 12, 2016 by guest
http://ec.asm.org/
Dow
nloaded from

17. Lonsdale-Eccles, J. D., and D. J. Grab. 1987. Lysosomal and non-lysosomalpeptidyl hydrolases of the bloodstream forms of Trypanosoma brucei brucei.Eur. J. Biochem. 169:467–475.
18. McLintock, L. M., C. M. Turner, and K. Vickerman. 1993. Comparison ofthe effects of immune killing mechanisms on Trypanosoma brucei parasites ofslender and stumpy morphology. Parasite Immunol. 15:475–480.
19. Moore, D. R., A. Smith, K. M. Hager, R. Waldon, J. D. Esko, and S. L.Hajduk. 1995. Developmentally regulated sensitivity of Trypanosoma bruceito the cytotoxic effects of human high-density lipoprotein. Exp. Parasitol.81:216–226.
20. Natesan, S. K., L. Peacock, K. Matthews, W. Gibson, and M. C. Field. 2007.Activation of endocytosis as an adaptation to the mammalian host by try-panosomes. Eukaryot. Cell 6:2029–2037.
21. Nolan, D. P., M. Geuskens, and E. Pays. 1999. N-linked glycans containinglinear poly-N-acetyllactosamine as sorting signals in endocytosis in Trypano-soma brucei. Curr. Biol. 9:1169–1172.
22. Nolan, D. P., S. Rolin, J. Rueda Rodriguez, J. Van Den Abbeele, and E. Pays.2000. Slender and stumpy bloodstream forms of Trypanosoma brucei displaya differential response to extracellular acidic and proteolytic stress. Eur.J. Biochem. 267:18–27.
23. Pays, E. 2005. Regulation of antigen gene expression in Trypanosoma brucei.Trends Parasitol. 21:517–520.
24. Pays, E., L. Vanhamme, and D. Perez-Morga. 2004. Antigenic variation inTrypanosoma brucei. Facts, challenges and mysteries. Curr. Opin. Microbiol.7:369–374.
25. Salmon, D., M. Geuskens, F. Hanocq, J. Hanocq-Quertier, D. Nolan, L.Ruben, and E. Pays. 1994. A novel heterodimeric transferrin receptor en-coded by a pair of VSG expression site-associated genes in Trypanosomabrucei. Cell 78:75–86.
26. Savill, N. J., and J. R. Seed. 2004. Mathematical and statistical analysis of theTrypanosoma brucei slender to stumpy transition. Parasitology 128:53–67.
27. Steverding, D. 1998. Bloodstream forms of Trypanosoma brucei require onlysmall amounts of iron for growth. Parasitol. Res. 84:59–62.
28. Vanhollebeke, B., G. De Muylder, M. J. Nielsen, A. Pays, P. Tebabi, M. Dieu,M. Raes, S. K. Moestrup, and E. Pays. 2008. A haptoglobin-hemoglobinreceptor conveys innate immunity to Trypanosoma brucei in humans. Science320:677–681.
29. Vanhollebeke, B., L. Lecordier, D. Perez-Morga, A. Amiguet-Vercher, and E.Pays. 2007. Human serum lyses Trypanosoma brucei by triggering uncon-trolled swelling of the parasite lysosome. J. Eukaryot. Microbiol. 54:448–451.
30. Vanhollebeke, B., and E. Pays. 2010. The trypanolytic factor of humanserum: many ways to enter the parasite, a single way to kill. Mol. Microbiol.76:806–814.
31. Vassella, E., R. Kramer, C. M. Turner, M. Wankell, C. Modes, M. van denBogaard, and M. Boshart. 2001. Deletion of a novel protein kinase with PXand FYVE-related domains increases the rate of differentiation of Trypano-soma brucei. Mol. Microbiol. 41:33–46.
32. Zhou, W., G. A. Cross, and W. D. Nes. 2007. Cholesterol import fails toprevent catalyst-based inhibition of ergosterol synthesis and cell proliferationof Trypanosoma brucei. J. Lipid Res. 48:665–673.
1282 VANHOLLEBEKE ET AL. EUKARYOT. CELL
on August 12, 2016 by guest
http://ec.asm.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















