
Novel Sterol Metabolic Network of Trypanosoma brucei Procyclic and Bloodstream Forms
15
Biochem. J. (2012) 443, 267–277 (Printed in Great Britain) doi:10.1042/BJ20111849 267 Novel sterol metabolic network of Trypanosoma brucei procyclic and bloodstream forms Craigen R. NES* 1 , Ujjal K. SINGHA† 1 , Jialin LIU*, Kulothungan GANAPATHY*, Fernando VILLALTA†, Michael R. WATERMAN‡, Galina I. LEPESHEVA‡, Minu CHAUDHURI† and W. David NES* 2 *Department ofChemistry and Biochemistry, Texas Tech University, Lubbock, TX 79409, U.S.A., †Department of Microbiology and Immunology, Meharry Medical College, Nashville, TN 37208, U.S.A., and ‡Department of Biochemistry, Vanderbilt University School of Medicine, Nashville, TN 37232, U.S.A. Trypanosoma brucei is the protozoan parasite that causes African trypanosomiasis, a neglected disease of people and animals. Co-metabolite analysis, labelling studies using [methyl- 2 H 3 ]- methionine and substrate/product specificities of the cloned 24-SMT (sterol C24-methyltransferase) and 14-SDM (sterol C14- demethylase) from T. brucei afforded an uncommon sterol metabolic network that proceeds from lanosterol and 31- norlanosterol to ETO [ergosta-5,7,25(27)-trien-3β -ol], 24-DTO [dimethyl ergosta-5,7,25(27)-trienol] and ergosterol [ergosta- 5,7,22(23)-trienol]. To assess the possible carbon sources of ergo- sterol biosynthesis, specifically 13 C-labelled specimens of lanosterol, acetate, leucine and glucose were administered to T. brucei and the 13 C distributions found were in accord with the operation of the acetate–mevalonate pathway, with leucine as an alternative precursor, to ergostenols in either the insect or bloodstream form. In searching for metabolic signatures of procyclic cells, we observed that the 13 C- labelling treatments induce fluctuations between the acetyl-CoA (mitochondrial) and sterol (cytosolic) synthetic pathways detected by the progressive increase in 13 C-ergosterol production (control <[2- 13 C]leucine<[2- 13 C]acetate<[1- 13 C]glucose) and corres- ponding depletion of cholesta-5,7,24-trienol. We conclude that anabolic fluxes originating in mitochondrial metabolism constitute a flexible part of sterol synthesis that is further fluctuated in the cytosol, yielding distinct sterol profiles in relation to cell demands on growth. Key words: [1- 13 C]glucose, ergosterol biosynthesis, sterol C24-methyltransferase, sterol C14-demethylase, Trypanosoma brucei, trypanosome. INTRODUCTION Eukaryotes exhibit extensive diversity in the composition of their membrane sterols, which have a fossil record that extends back billions of years. The most valuable feature giving rise to the different sterol patterns in nature are largely due to the presence of various combinations of C24-alkyl groups in the sterol side chain, which are synthesized by a family of structurally similar 24- SMT (sterol C24-methyltransferase) enzymes [1]. Algae, fungi and protozoa typically synthesize 24β -alkyl sterols, whereas vascular plants synthesize 24α-alkyl sterols from the acetate– MVA (mevalonate) or MVA-independent pathways. In previous years, much effort has been devoted to understanding the 24-alkyl sterol metabolic networks that operate throughout Nature [2–4], in order to identify the evolutionary pathway for how these structure specificities arose [5–7] and, in the case of neglected diseases, to develop chemotherapeutic leads designed to selectively inhibit their production [8–11]. For some time our laboratories have been engaged in a study designed to establish the in vitro properties of cloned enzymes that act on sterols and in vivo sterol compositions from organisms of the protozoan order Kinetoplastida [12–17]. In the present study, a further examination of the enzyme properties of the 24- SMT and 14-SDM (sterol C14-demethylase), and the identities and amounts of sterols from one of these members responsible for sleeping sickness, Trypanosoma brucei, has been undertaken; in addition, we studied the incorporation of different 2 H- and 13 C-labelled carbon sources into the final biosynthesized products. One reason why we have pursued probing the sterol biosyn- thesis abilities of T. brucei is to establish the basis of variability in ergosterol homoeostasis as the parasite responds to adaptations in the changing environments of human bloodstream and alimentary gut of the tsetse fly, which to date remains enigmatic. Thus the bloodstream forms of T. brucei have been reported to be blocked at some unknown stage of isoprenoid metabolism, generating ergosterol auxotrophy to live [18–21]. To satisfy the sterol requirements for the bloodstream form, it is proposed that host cholesterol is absorbed by the cell via a receptor-mediated lipoprotein uptake mechanism [22]. As a result, it has long been held that their ability to scavenge cholesterol from their animal hosts, and their inability to undergo sterol biosynthesis in this adaptive stage, makes them resistant to antiparasitic drugs that target ergosterol or ergosterol biosynthesis [20–23]. On the other hand, testing sterol biosynthesis inhibitors that bind specifically to the cloned TbSMT (T. brucei 24-SMT) and TbSDM (T. brucei 14-SDM) enzymes has shown some success [12,13,16]. The existence of a functional 24-alkyl sterol pathway was demonstrated in vitro in both the procyclic and bloodstream forms [14,15]. It is noteworthy that several independent studies indicate that ergosterol (ergosta-5,7,22- trienol) is the major 24-alkyl sterol of T. brucei procyclic cells cultured on FGM (full-growth medium) [18,20,24], as reported Abbreviations used: AdoMet, S-adenosylmethionine; a.m.u., atomic mass unit; CDM, cholesterol-depleted medium; 24-DTO, dimethyl ergosta- 5,7,25(27)-trienol; ETO, ergosta-5,7,25(27)-trien-3β-ol; FBS, fetal bovine serum; FGM, full-growth medium; IPP, isopentyl diphosphate; MVA, mevalonate; RRTc, retention time relative to the retention time of cholesterol; 14-SDM, sterol C14-demethylase; 24-SMT, sterol C24-methyltransferase; TbSDM, T. brucei 14-SDM; TbSMT, T. brucei 24-SMT. 1 These authors contributed equally to this work. 2 To whom correspondence should be addressed (email [email protected]). c The Authors Journal compilation c 2012 Biochemical Society www.biochemj.org Biochemical Journal
Transcript of Novel Sterol Metabolic Network of Trypanosoma brucei Procyclic and Bloodstream Forms

Biochem. J. (2012) 443, 267–277 (Printed in Great Britain) doi:10.1042/BJ20111849 267
Novel sterol metabolic network of Trypanosoma brucei procyclic andbloodstream formsCraigen R. NES*1, Ujjal K. SINGHA†1, Jialin LIU*, Kulothungan GANAPATHY*, Fernando VILLALTA†, Michael R. WATERMAN‡,Galina I. LEPESHEVA‡, Minu CHAUDHURI† and W. David NES*2
*Department of Chemistry and Biochemistry, Texas Tech University, Lubbock, TX 79409, U.S.A., †Department of Microbiology and Immunology, Meharry Medical College, Nashville,TN 37208, U.S.A., and ‡Department of Biochemistry, Vanderbilt University School of Medicine, Nashville, TN 37232, U.S.A.
Trypanosoma brucei is the protozoan parasite that causes Africantrypanosomiasis, a neglected disease of people and animals.Co-metabolite analysis, labelling studies using [methyl-2H3]-methionine and substrate/product specificities of the cloned24-SMT (sterol C24-methyltransferase) and 14-SDM (sterol C14-demethylase) from T. brucei afforded an uncommon sterolmetabolic network that proceeds from lanosterol and 31-norlanosterol to ETO [ergosta-5,7,25(27)-trien-3β-ol], 24-DTO[dimethyl ergosta-5,7,25(27)-trienol] and ergosterol [ergosta-5,7,22(23)-trienol]. To assess the possible carbon sources of ergo-sterol biosynthesis, specifically 13C-labelled specimens oflanosterol, acetate, leucine and glucose were administered toT. brucei and the 13C distributions found were in accordwith the operation of the acetate–mevalonate pathway, withleucine as an alternative precursor, to ergostenols in either
the insect or bloodstream form. In searching for metabolicsignatures of procyclic cells, we observed that the 13C-labelling treatments induce fluctuations between the acetyl-CoA(mitochondrial) and sterol (cytosolic) synthetic pathways detectedby the progressive increase in 13C-ergosterol production (control<[2-13C]leucine<[2-13C]acetate<[1-13C]glucose) and corres-ponding depletion of cholesta-5,7,24-trienol. We concludethat anabolic fluxes originating in mitochondrial metabolismconstitute a flexible part of sterol synthesis that is furtherfluctuated in the cytosol, yielding distinct sterol profiles in relationto cell demands on growth.
Key words: [1-13C]glucose, ergosterol biosynthesis, sterolC24-methyltransferase, sterol C14-demethylase, Trypanosomabrucei, trypanosome.
INTRODUCTION
Eukaryotes exhibit extensive diversity in the composition of theirmembrane sterols, which have a fossil record that extends backbillions of years. The most valuable feature giving rise to thedifferent sterol patterns in nature are largely due to the presenceof various combinations of C24-alkyl groups in the sterol sidechain, which are synthesized by a family of structurally similar 24-SMT (sterol C24-methyltransferase) enzymes [1]. Algae, fungiand protozoa typically synthesize 24β-alkyl sterols, whereasvascular plants synthesize 24α-alkyl sterols from the acetate–MVA (mevalonate) or MVA-independent pathways. In previousyears, much effort has been devoted to understanding the 24-alkylsterol metabolic networks that operate throughout Nature [2–4], inorder to identify the evolutionary pathway for how these structurespecificities arose [5–7] and, in the case of neglected diseases,to develop chemotherapeutic leads designed to selectively inhibittheir production [8–11].
For some time our laboratories have been engaged in a studydesigned to establish the in vitro properties of cloned enzymesthat act on sterols and in vivo sterol compositions from organismsof the protozoan order Kinetoplastida [12–17]. In the presentstudy, a further examination of the enzyme properties of the 24-SMT and 14-SDM (sterol C14-demethylase), and the identitiesand amounts of sterols from one of these members responsiblefor sleeping sickness, Trypanosoma brucei, has been undertaken;
in addition, we studied the incorporation of different 2H- and13C-labelled carbon sources into the final biosynthesized products.
One reason why we have pursued probing the sterol biosyn-thesis abilities of T. brucei is to establish the basis of variability inergosterol homoeostasis as the parasite responds to adaptationsin the changing environments of human bloodstream andalimentary gut of the tsetse fly, which to date remains enigmatic.Thus the bloodstream forms of T. brucei have been reported tobe blocked at some unknown stage of isoprenoid metabolism,generating ergosterol auxotrophy to live [18–21]. To satisfy thesterol requirements for the bloodstream form, it is proposed thathost cholesterol is absorbed by the cell via a receptor-mediatedlipoprotein uptake mechanism [22]. As a result, it has long beenheld that their ability to scavenge cholesterol from their animalhosts, and their inability to undergo sterol biosynthesis in thisadaptive stage, makes them resistant to antiparasitic drugs thattarget ergosterol or ergosterol biosynthesis [20–23].
On the other hand, testing sterol biosynthesis inhibitors thatbind specifically to the cloned TbSMT (T. brucei 24-SMT)and TbSDM (T. brucei 14-SDM) enzymes has shown somesuccess [12,13,16]. The existence of a functional 24-alkylsterol pathway was demonstrated in vitro in both the procyclicand bloodstream forms [14,15]. It is noteworthy that severalindependent studies indicate that ergosterol (ergosta-5,7,22-trienol) is the major 24-alkyl sterol of T. brucei procyclic cellscultured on FGM (full-growth medium) [18,20,24], as reported
Abbreviations used: AdoMet, S-adenosylmethionine; a.m.u., atomic mass unit; CDM, cholesterol-depleted medium; 24-DTO, dimethyl ergosta-5,7,25(27)-trienol; ETO, ergosta-5,7,25(27)-trien-3β-ol; FBS, fetal bovine serum; FGM, full-growth medium; IPP, isopentyl diphosphate; MVA, mevalonate;RRTc, retention time relative to the retention time of cholesterol; 14-SDM, sterol C14-demethylase; 24-SMT, sterol C24-methyltransferase; TbSDM, T. brucei14-SDM; TbSMT, T. brucei 24-SMT.
1 These authors contributed equally to this work.2 To whom correspondence should be addressed (email [email protected]).
c© The Authors Journal compilation c© 2012 Biochemical Society
www.biochemj.org
Bio
chem
ical
Jo
urn
al
268 C. R. Nes and others
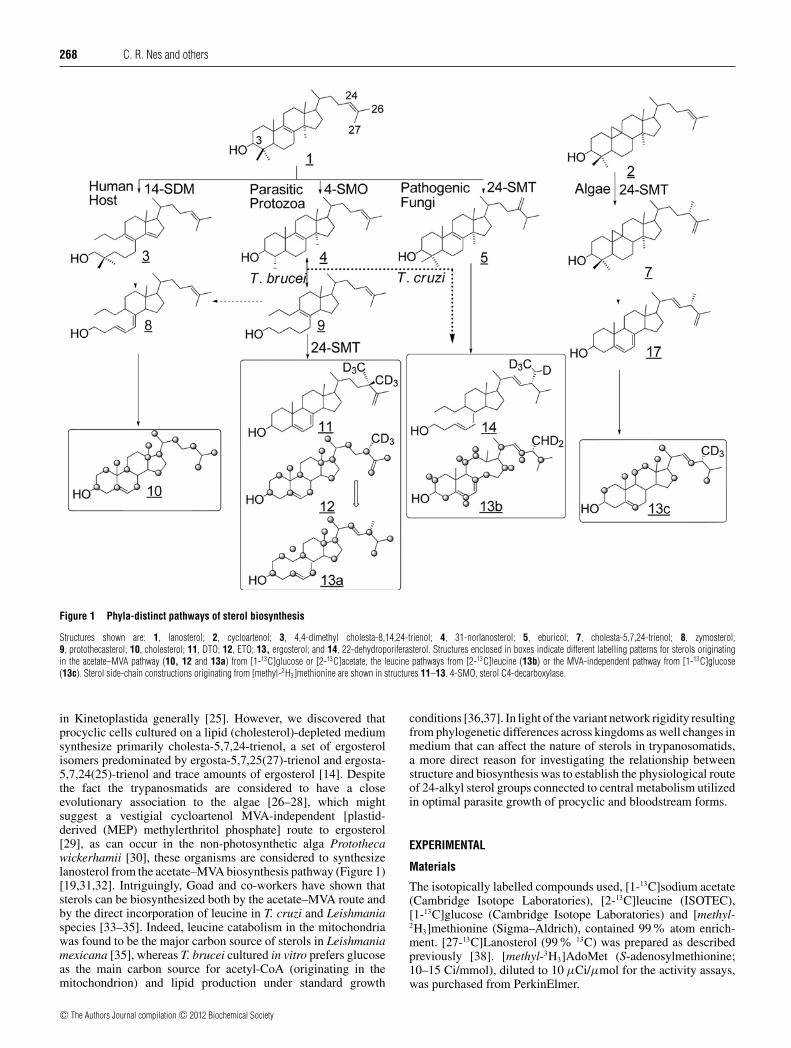
Figure 1 Phyla-distinct pathways of sterol biosynthesis
Structures shown are: 1, lanosterol; 2, cycloartenol; 3, 4,4-dimethyl cholesta-8,14,24-trienol; 4, 31-norlanosterol; 5, eburicol; 7, cholesta-5,7,24-trienol; 8, zymosterol;9, protothecasterol; 10, cholesterol; 11, DTO; 12, ETO; 13, ergosterol; and 14, 22-dehydroporiferasterol. Structures enclosed in boxes indicate different labelling patterns for sterols originatingin the acetate–MVA pathway (10, 12 and 13a) from [1-13C]glucose or [2-13C]acetate, the leucine pathways from [2-13C]leucine (13b) or the MVA-independent pathway from [1-13C]glucose(13c). Sterol side-chain constructions originating from [methyl-2H3]methionine are shown in structures 11–13. 4-SMO, sterol C4-decarboxylase.
in Kinetoplastida generally [25]. However, we discovered thatprocyclic cells cultured on a lipid (cholesterol)-depleted mediumsynthesize primarily cholesta-5,7,24-trienol, a set of ergosterolisomers predominated by ergosta-5,7,25(27)-trienol and ergosta-5,7,24(25)-trienol and trace amounts of ergosterol [14]. Despitethe fact the trypanosmatids are considered to have a closeevolutionary association to the algae [26–28], which mightsuggest a vestigial cycloartenol MVA-independent [plastid-derived (MEP) methylerthritol phosphate] route to ergosterol[29], as can occur in the non-photosynthetic alga Protothecawickerhamii [30], these organisms are considered to synthesizelanosterol from the acetate–MVA biosynthesis pathway (Figure 1)[19,31,32]. Intriguingly, Goad and co-workers have shown thatsterols can be biosynthesized both by the acetate–MVA route andby the direct incorporation of leucine in T. cruzi and Leishmaniaspecies [33–35]. Indeed, leucine catabolism in the mitochondriawas found to be the major carbon source of sterols in Leishmaniamexicana [35], whereas T. brucei cultured in vitro prefers glucoseas the main carbon source for acetyl-CoA (originating in themitochondrion) and lipid production under standard growth
conditions [36,37]. In light of the variant network rigidity resultingfrom phylogenetic differences across kingdoms as well changes inmedium that can affect the nature of sterols in trypanosomatids,a more direct reason for investigating the relationship betweenstructure and biosynthesis was to establish the physiological routeof 24-alkyl sterol groups connected to central metabolism utilizedin optimal parasite growth of procyclic and bloodstream forms.
EXPERIMENTAL
Materials
The isotopically labelled compounds used, [1-13C]sodium acetate(Cambridge Isotope Laboratories), [2-13C]leucine (ISOTEC),[1-13C]glucose (Cambridge Isotope Laboratories) and [methyl-2H3]methionine (Sigma–Aldrich), contained 99% atom enrich-ment. [27-13C]Lanosterol (99 % 13C) was prepared as describedpreviously [38]. [methyl-3H3]AdoMet (S-adenosylmethionine;10–15 Ci/mmol), diluted to 10 μCi/μmol for the activity assays,was purchased from PerkinElmer.
c© The Authors Journal compilation c© 2012 Biochemical Society

Sterol biosynthesis in Trypanosoma brucei 269
Cell cultures and growth studies
T. brucei strain 427 procyclic-form cells were grown in SDM-79 medium supplemented with 10% heat-inactivated FBS (fetalbovine serum; Atlanta Biologicals), hereafter referred to asFGM, at 27 ◦C. Bloodstream-form cells were maintained inHMI-9 medium supplemented with 10% heat-inactivated FBS(Atlanta Biologicals) and 10 % Serum Plus (SAFC Biosciences),which is also a FGM. For preparation of a CDM (cholesterol-depleted medium), procyclic forms in SDM-79 medium weresupplemented with 10% heat-inactivated lipid-free FBS (Sigma–Aldrich). For a low-cholesterol medium, the bloodstream-formcells were grown in HMI-9 medium containing 10% lipid-freeheat-inactivated FBS (Sigma–Aldrich) and 10% Serum Plus,which contains serum lipids equivalent to 2% of FBS. Cultureswere harvested by centrifugation. (5000 g for 10 min at 4 ◦C) Celldensities were determined using a Neubauer haemocytometercounter. Growth curves were performed in triplicate and thevariation in growth at each data point never exceeded 10%.
Enzyme assays
Procedures for the heterologous overexpression in E. coli and theenzyme preparations of TbSMT and TbSDM were performed asdescribed previously [12,13].
Instrumental methods
Sterol compounds, referenced to the retention time of cholesterolin capillary GC at 13.8 min and HPLC at 22 or 37 min (analyticalcompared with semi-preparative C18-column), were quantified byintegration of the detector signal [FID (flame ionization detector)in GC and UV at 210 in HPLC], from 10 to 30 min. Productswere routinely identified by their retention times in GC andelectron-impact spectrum with those of reference specimens. Forselect products, unambiguous identification was established afterHPLC (Phenomenex C18-column linked to a diode array detectorwhich provided UV spectra) fractionation followed by GC/MS(ZB-5 capillary column of 30 m coupled to a HP LS 6500 gaschromatograph interfaced to a 5973 mass spectrometer) and NMRanalysis [spectra measured on deuterochloroform solutions usinga Varian Unity Inova 500 MHz spectrometer operating at 500MHz for protons and 125 MHz for 13C nuclei; chemical shiftswere referenced to chloroform at δ = 77.00 and reported as δ(p.p.m.)] [14,30]. For biosynthetically 13C-labelled sterols, broad-band de-coupled 13C-NMR spectra were measured as follows:32520.3 Hz spectrum width, 90
◦pulse, 1.3 s acquisition time,
70000 transients, 84618 K data points, line broadening of 0.5 Hzbefore Fourier transformation and a relaxation delay of 6 s with arepetition time of 3.3 s.
Sterol analysis, metabolic labelling and evaluation of13C/2H-enrichment in sterol
Two sets of experiments were performed to determine the sterolcomposition of T. brucei cells. For the first set of experimentsto isolate sterols for chemical characterization, procyclic cellswere cultured on FGM to yield approximately 1–2 × 109 cells.T. brucei cell pellets were saponified directly in 10 % KOHin 80% aqueous methanol at the reflux temperature for 1 h,which yielded total sterol (free plus esterified sterols). The neutrallipids obtained by dilution with water and extraction with hexane(Fisher) after the saponification were analysed by GC (3 % SE-30pack column operated isothermally at 245 ◦C). Total sterols in thenon-saponifiable lipid fraction were purified on an Agilent 1100
series HPLC system coupled to a diode array detector. The totalsterols were loaded on to a C18 reversed-phase analytical column(Phenomenex, 4 μm) eluted isocratically with methanol at a flowrate of 1 ml/min. The standard for HPLC data was cholesterol, andthe rates of movement are given relative to cholesterol (αc). HPLCfractions in the sterol region of the chromatogram (αc = 0.5–2.0)were collected and analysed by GC/MS with a Hewlett-PackardLS 6500 gas chromatograph at 70 eV. GC was performed usingan Agilent ZB-5 column (30m × 25 μm in diameter) and in somecases by 1H-NMR (500 MHz) recorded in deuterochloroformsolutions using a Varian INOVA 500 spectrometer. Cholesterolwas the standard for determination of the RRTc (retention timerelative to the retention time of cholesterol). Quantification of theamounts of unlabelled sterols was accomplished by GC with astandard curve for cholesterol. To several cell pellets undergoingthe saponification process, 5α-cholestane was added as an internalstandard. Using GC, cholesterol at 1 ng, or with HPLC, ergosterolat 1 ng, can be detected in the sample preparation [14].
For the metabolic labelling experiments, cells cultured in FGMcontaining physiological concentrations of the 13C- or 2H-labelledcompound in place of the corresponding unlabelled compoundwas added to the growth medium and the cultures left at 27 ◦C for72 h. The total sterols in the organic extract after saponificationof the cells were analysed as described above. For determin-ation of the stable IEF (isotope enrichment factors), the extent of2H and 13C incorporation were determined by GC/MS accordingto the calculations of Masse et al. [3]. The 13C-NMR spectraof the mixture of [13C]ergostenols obtained from an incorporationexperiment with [1-13C]glucose and a reference specimen of yeastergosterol were recorded at 500 MHz in deuterochloroform underidentical conditions using 16C in ergostenol as a reference. Relative13C-abundance of the individual carbon atoms was then calculatedby comparison of 13C signal integrals between 13C-labelled andunlabelled material.
Incorporation of [1-13C]glucose
Glucose-free SDM-79 medium (300 ml) containing D-[1-13C]Glucose (Cambridge Isotope Laboratories) (1.0 g/l) wereinoculated with T. brucei 427 procyclic-form cells (3 × 106/ml)and were allowed to grow in the presence or absence of lanosterol(38 μM) at 27 ◦C in a 1 litre conical flask with constant stirring.Cells were harvested after 72 h, washed twice with phosphate-buffered saline (50 ml each) and stored at − 70 ◦C.
Incorporation of L-[2-13C]leucine
SDM-79 medium was prepared using leucine-free MEM (minimalessential medium; Sigma–Aldrich) and supplemented withL-[2-13C]leucine (ISOTEC) (0.052 g/l). T. brucei 427 procycliccells were grown in this medium in the presence or absence oflanosterol (38 μM) for 72 h and harvested as described above.
Incorporation of [1-13C]sodium acetate
The SDM-79 medium (300 ml) supplemented with [1-13C]sodiumacetate (2 mM) was inoculated with 3 × 106 cells/ml and allowedto grow in the presence or absence of lanosterol (38 μM) at 27 ◦Cin a 1 litre conical flask with constant stirring. Cells were harvestedas described above.
Incorporation of [methyl-2H3]methionine
The methionine-free SDM-79 medium (300 ml) supplementedwith [methyl-2H3]methionine (0.07 g/l) was inoculated with
c© The Authors Journal compilation c© 2012 Biochemical Society

270 C. R. Nes and others
Figure 2 Gas–liquid chromatographic separation of the total sterol fractionderived from Trypanosoma brucei procyclic cells cultured on a FGM (A) or aCDM (B)
The major sterol component associated with GC peak 1 is cholesterol (M + 386), GC peak 2 ischolesta-5,7,24-trienol (M + 382), GC peak 3 is ergosta-5,7,25(27)-trienol (M + 396) and GCpeak4 is ergosta-5,7,24(25)-trienol (M + 396).
3 × 106 cells/ml and allowed to grow in the presence or absence oflanosterol (38 μM) at 27 ◦C in a 1 litre conical flask with constantstirring. Cells were harvested as described above.
RESULTS AND DISCUSSION
Sterol composition of procyclic T. brucei
To elucidate the full complement of metabolites in the sterolmetabolome from T. brucei, we conducted a thorough analysisof the sterol composition of procyclic cells cultured on FGMor CDM for 6 days (Supplementary Figure S1 at http://www.BiochemJ.org/bj/443/bj4430267add.htm). To accomplish thepresent study, a combination of chemical approaches wasemployed including: HPLC (to separate and isolate sterolsoriginating in the non-saponifiable lipid fraction) coupled to aUV detector to gain information about the position and number ofdouble bonds in the sterol molecule; GC profiling of the total sterolfraction relative to the chromatographic behaviour of authenticstandards in our steroid collection was used to provide a tentativeidentification of these compounds. Proof of the sterol identitydetected in GC was obtained by MS. In this spectral analysis,the molecular ion and fragmentation pattern of the compoundprovide relevant structural information; and finally 1H-NMR canfurnish a complete stereochemical description of protozoan sterolsgenerated in pure form by HPLC.
Procyclic cell growth was significantly inhibited in CDM. Afterday 4, cell number was decreased and upon longer incubation cellsdied. However, growth of the bloodstream form was minimallyaffected in CDM, indicating that the presence of 2% serum lipidfrom the Serum Plus is sufficient for cell growth. Bloodstreamcells did not grow in the absence of Serum Plus. The majorsterols (<95%) of the non-saponifiable lipids, identified byGC/MS, were qualitatively similar except the proportion ofcholesterol in the sterol samples varied with culture medium;FGM and CDM yielded approximately 55% and 35% cholesterolrespectively (Figure 2). The major endogenously formed sterols(several of which are shown in Figure 1) were cholesta-5,7,24-trienol (Figure 1, compound 7) with lesser amounts ofETO [ergosta-5,7,25(27)-trien-3β-ol] (Figure 1, compound 12)and ergosta-5,7,24(25)-trienol (not shown). Ergosterol (ergosta-5,7,22E-trienol) (Figure 1, compound 13) was routinely detected
only as a trace compound co-migrating in GC with cholesta-5,7,24-trienol. The two sterols are readily differentiated by theirmass spectra. Thus the molecular ion for ergosterol appears at M +
396 a.m.u. (atomic mass unit), whereas for cholesta-5,7,24-trienolit appears at M + 382 a.m.u.
A description of the distributions of 15 HPLC-purifiedsterols from procyclic cells cultured on CDM has beenpublished previously [14]; key intermediates detected in thesterol composition were lanosterol (Figure 1, compound 1),31-norlanosterol (Figure 1, compound 4) and zymosterol(Figure 1, compound 8) present at less than 2% of the totalsterols. During the course of the present study, one additional newcompound was isolated from the HPLC of the total sterol fractionfrom procyclic cells grown on FGM, 24-DTO [dimethyl ergosta-5,7,25(27)-trienol] which exhibited chromatographic behaviour(RRTc, 1.36), UV (λmax = 282 nm) and mass spectra (M + 410a.m.u.) identical with an authentic sample generated by TbSMTcatalysis of cholesta-5,7,24-trienol [39]. We also isolated ETOfrom the HPLC to prove the stereochemistry of the C24-methylgroup in the sterol side chain. The 1H-NMR spectrum of ETOafforded signals at δ H 18, 0.621 (s), H 19, 0.951 (s), H 21, 0.942(d), H 26, 1.648 (br. s), H 27, 4.672 (br. s), H 28, 1.01 (d)- C24methyl group is β-oriented, H 6, 5.577 (dd), H 7, 5.380 (m) p.p.m.,consistent with its proposed structure [40,41].
An important purpose of repeating the sterol analyses of freshcells cultured at Meharry Medical College (we previously studiedprocyclic cells grown at Rockefeller University [14]) was toshow clearly that T. brucei normally synthesizes three majoruncommon sterols, cholesta-5,7,24-trienol, ergosta-5,7,25(27)-trienol and ergosta-5,7,24-trienol, and that the proportion of themrelative to cholesterol can vary dramatically with the availabilityof lipid (cholesterol) in the growth medium. Equally importantto us was to confirm that cells can produce ergosterol, albeit attrace levels and that the usual practice of culturing T. bruceion FGM or CDM fails to affect the low abundance of thiscompound in the sterol mixture. On the basis of these results, wesurmise that changes in sterol homoeostasis associated with theprocyclic growth conditions, which are marked by high and lowconcentrations of cholesta-5,7,24-trienol (Figure 2, peak 2), maybe the result of a heretofore unrecognized regulation mechanismof post-squalene enzyme activities.
Sterol biosynthesis in procyclic cells
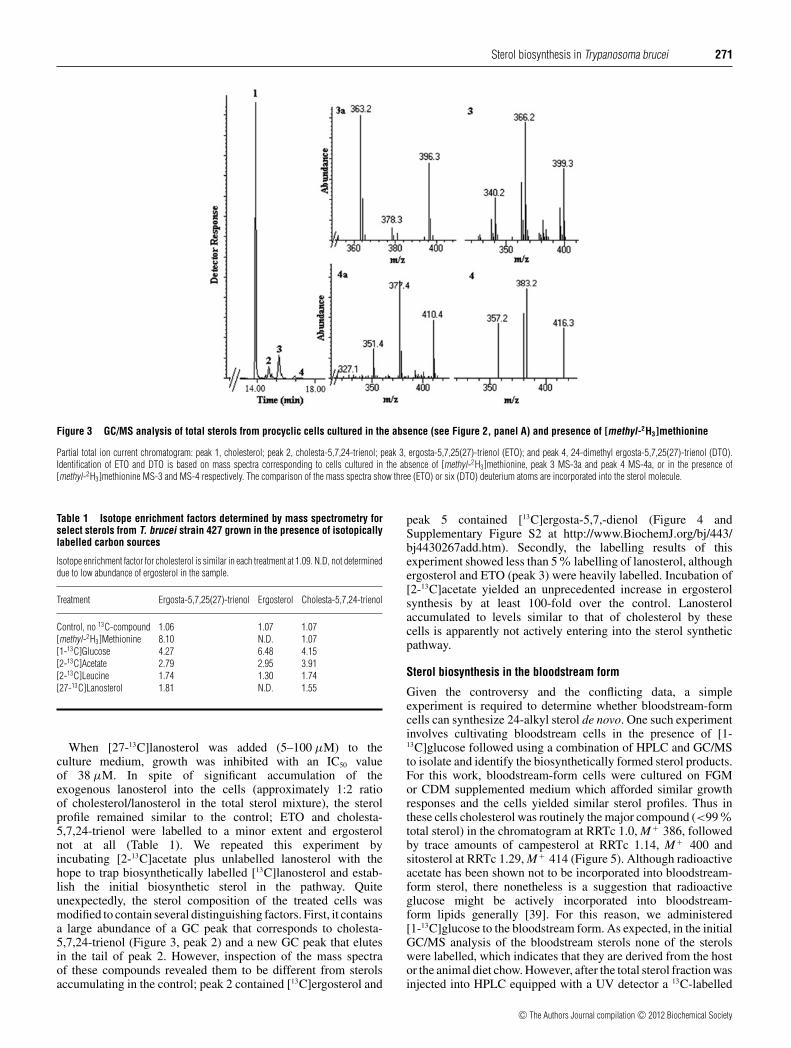
In the present study, GC/MS was used to determine the numberof deuterium atoms from [methyl-2H3]AdoMet (synthesizedin vivo from [methyl-2H3]methionine) incorporated into the sterolside chain at carbon-28. Thus procyclic cells were cultured inthe presence of [methyl-2H3]methionine and neither the growthresponse nor total sterol content of the cells changed comparedwith the control. GC/MS analysis of total sterols from [methyl-2H3]methionine-treated cells revealed significant incorporation ofdeuterium into the C24-methyl sterols ETO and ergosta-5,7-24-trienol showing the incorporation of none and three deuteriumatoms (M + 396 and 399 a.m.u.) and in the 24-dimethyl sterol DTOshowing the addition of six deuterium atoms in the mass spectrum(M + at 410 shifted to M + 416 a.m.u.). (Figure 3); as expected, nodeuterium was detected in cholesterol (Table 1). These resultsprovide convincing evidence for the intermediacy of ergosta-8,25(27)-dienol and ergosta-8,24(25)-dienol in the biosynthesis ofETO and DTO respectively, and rule out ergosta-8,24(28)-dienol(fecosterol) as an intermediate to the major 24-alkyl sterols thataccumulate in these cells (Figure 1). Ergosterol, detected as atrace compound in the sterol mixture, was not labelled from theseincubations.
c© The Authors Journal compilation c© 2012 Biochemical Society
Sterol biosynthesis in Trypanosoma brucei 271
Figure 3 GC/MS analysis of total sterols from procyclic cells cultured in the absence (see Figure 2, panel A) and presence of [methyl-2H3]methionine
Partial total ion current chromatogram: peak 1, cholesterol; peak 2, cholesta-5,7,24-trienol; peak 3, ergosta-5,7,25(27)-trienol (ETO); and peak 4, 24-dimethyl ergosta-5,7,25(27)-trienol (DTO).Identification of ETO and DTO is based on mass spectra corresponding to cells cultured in the absence of [methyl-2H3]methionine, peak 3 MS-3a and peak 4 MS-4a, or in the presence of[methyl-2H3]methionine MS-3 and MS-4 respectively. The comparison of the mass spectra show three (ETO) or six (DTO) deuterium atoms are incorporated into the sterol molecule.
Table 1 Isotope enrichment factors determined by mass spectrometry forselect sterols from T. brucei strain 427 grown in the presence of isotopicallylabelled carbon sources
Isotope enrichment factor for cholesterol is similar in each treatment at 1.09. N.D, not determineddue to low abundance of ergosterol in the sample.
Treatment Ergosta-5,7,25(27)-trienol Ergosterol Cholesta-5,7,24-trienol
Control, no 13C-compound 1.06 1.07 1.07[methyl-2H3]Methionine 8.10 N.D. 1.07[1-13C]Glucose 4.27 6.48 4.15[2-13C]Acetate 2.79 2.95 3.91[2-13C]Leucine 1.74 1.30 1.74[27-13C]Lanosterol 1.81 N.D. 1.55
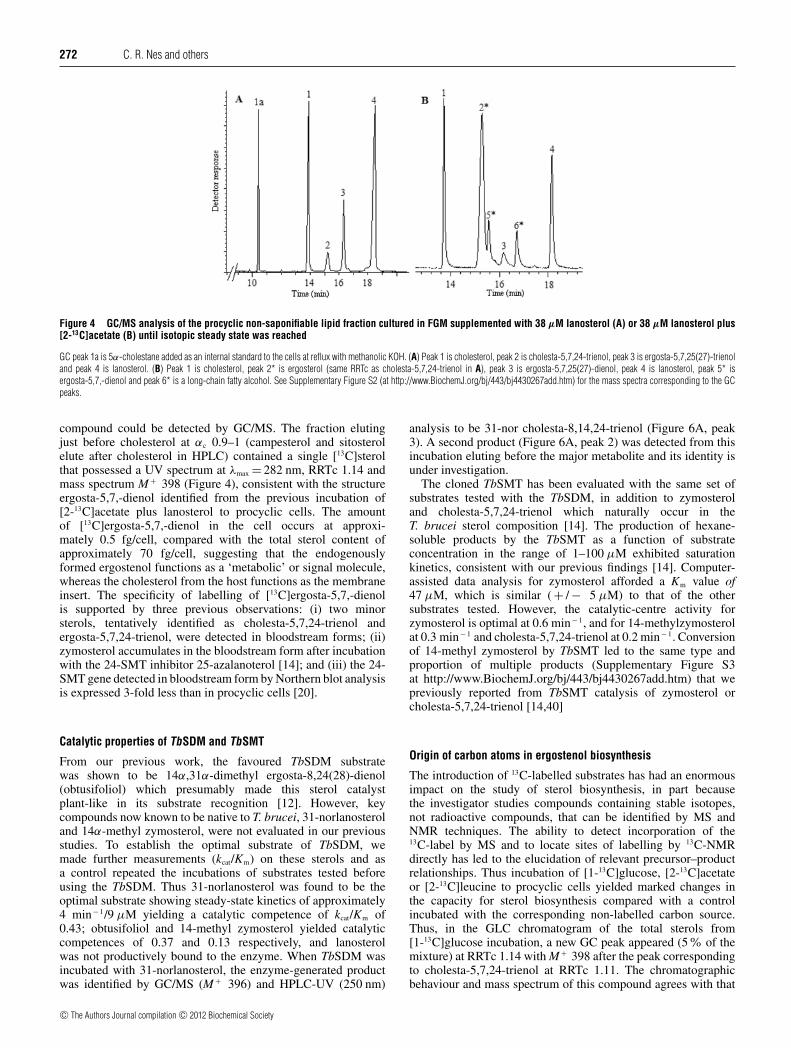
When [27-13C]lanosterol was added (5–100 μM) to theculture medium, growth was inhibited with an IC50 valueof 38 μM. In spite of significant accumulation of theexogenous lanosterol into the cells (approximately 1:2 ratioof cholesterol/lanosterol in the total sterol mixture), the sterolprofile remained similar to the control; ETO and cholesta-5,7,24-trienol were labelled to a minor extent and ergosterolnot at all (Table 1). We repeated this experiment byincubating [2-13C]acetate plus unlabelled lanosterol with thehope to trap biosynthetically labelled [13C]lanosterol and estab-lish the initial biosynthetic sterol in the pathway. Quiteunexpectedly, the sterol composition of the treated cells wasmodified to contain several distinguishing factors. First, it containsa large abundance of a GC peak that corresponds to cholesta-5,7,24-trienol (Figure 3, peak 2) and a new GC peak that elutesin the tail of peak 2. However, inspection of the mass spectraof these compounds revealed them to be different from sterolsaccumulating in the control; peak 2 contained [13C]ergosterol and
peak 5 contained [13C]ergosta-5,7,-dienol (Figure 4 andSupplementary Figure S2 at http://www.BiochemJ.org/bj/443/bj4430267add.htm). Secondly, the labelling results of thisexperiment showed less than 5% labelling of lanosterol, althoughergosterol and ETO (peak 3) were heavily labelled. Incubation of[2-13C]acetate yielded an unprecedented increase in ergosterolsynthesis by at least 100-fold over the control. Lanosterolaccumulated to levels similar to that of cholesterol by thesecells is apparently not actively entering into the sterol syntheticpathway.
Sterol biosynthesis in the bloodstream form
Given the controversy and the conflicting data, a simpleexperiment is required to determine whether bloodstream-formcells can synthesize 24-alkyl sterol de novo. One such experimentinvolves cultivating bloodstream cells in the presence of [1-13C]glucose followed using a combination of HPLC and GC/MSto isolate and identify the biosynthetically formed sterol products.For this work, bloodstream-form cells were cultured on FGMor CDM supplemented medium which afforded similar growthresponses and the cells yielded similar sterol profiles. Thus inthese cells cholesterol was routinely the major compound (<99%total sterol) in the chromatogram at RRTc 1.0, M + 386, followedby trace amounts of campesterol at RRTc 1.14, M + 400 andsitosterol at RRTc 1.29, M + 414 (Figure 5). Although radioactiveacetate has been shown not to be incorporated into bloodstream-form sterol, there nonetheless is a suggestion that radioactiveglucose might be actively incorporated into bloodstream-form lipids generally [39]. For this reason, we administered[1-13C]glucose to the bloodstream form. As expected, in the initialGC/MS analysis of the bloodstream sterols none of the sterolswere labelled, which indicates that they are derived from the hostor the animal diet chow. However, after the total sterol fraction wasinjected into HPLC equipped with a UV detector a 13C-labelled
c© The Authors Journal compilation c© 2012 Biochemical Society
272 C. R. Nes and others
Figure 4 GC/MS analysis of the procyclic non-saponifiable lipid fraction cultured in FGM supplemented with 38 μM lanosterol (A) or 38 μM lanosterol plus[2-13C]acetate (B) until isotopic steady state was reached
GC peak 1a is 5α-cholestane added as an internal standard to the cells at reflux with methanolic KOH. (A) Peak 1 is cholesterol, peak 2 is cholesta-5,7,24-trienol, peak 3 is ergosta-5,7,25(27)-trienoland peak 4 is lanosterol. (B) Peak 1 is cholesterol, peak 2* is ergosterol (same RRTc as cholesta-5,7,24-trienol in A), peak 3 is ergosta-5,7,25(27)-dienol, peak 4 is lanosterol, peak 5* isergosta-5,7,-dienol and peak 6* is a long-chain fatty alcohol. See Supplementary Figure S2 (at http://www.BiochemJ.org/bj/443/bj4430267add.htm) for the mass spectra corresponding to the GCpeaks.
compound could be detected by GC/MS. The fraction elutingjust before cholesterol at αc 0.9–1 (campesterol and sitosterolelute after cholesterol in HPLC) contained a single [13C]sterolthat possessed a UV spectrum at λmax = 282 nm, RRTc 1.14 andmass spectrum M + 398 (Figure 4), consistent with the structureergosta-5,7,-dienol identified from the previous incubation of[2-13C]acetate plus lanosterol to procyclic cells. The amountof [13C]ergosta-5,7,-dienol in the cell occurs at approxi-mately 0.5 fg/cell, compared with the total sterol content ofapproximately 70 fg/cell, suggesting that the endogenouslyformed ergostenol functions as a ‘metabolic’ or signal molecule,whereas the cholesterol from the host functions as the membraneinsert. The specificity of labelling of [13C]ergosta-5,7,-dienolis supported by three previous observations: (i) two minorsterols, tentatively identified as cholesta-5,7,24-trienol andergosta-5,7,24-trienol, were detected in bloodstream forms; (ii)zymosterol accumulates in the bloodstream form after incubationwith the 24-SMT inhibitor 25-azalanoterol [14]; and (iii) the 24-SMT gene detected in bloodstream form by Northern blot analysisis expressed 3-fold less than in procyclic cells [20].
Catalytic properties of TbSDM and TbSMT
From our previous work, the favoured TbSDM substratewas shown to be 14α,31α-dimethyl ergosta-8,24(28)-dienol(obtusifoliol) which presumably made this sterol catalystplant-like in its substrate recognition [12]. However, keycompounds now known to be native to T. brucei, 31-norlanosteroland 14α-methyl zymosterol, were not evaluated in our previousstudies. To establish the optimal substrate of TbSDM, wemade further measurements (kcat/Km) on these sterols and asa control repeated the incubations of substrates tested beforeusing the TbSDM. Thus 31-norlanosterol was found to be theoptimal substrate showing steady-state kinetics of approximately4 min− 1/9 μM yielding a catalytic competence of kcat/Km of0.43; obtusifoliol and 14-methyl zymosterol yielded catalyticcompetences of 0.37 and 0.13 respectively, and lanosterolwas not productively bound to the enzyme. When TbSDM wasincubated with 31-norlanosterol, the enzyme-generated productwas identified by GC/MS (M + 396) and HPLC-UV (250 nm)
analysis to be 31-nor cholesta-8,14,24-trienol (Figure 6A, peak3). A second product (Figure 6A, peak 2) was detected from thisincubation eluting before the major metabolite and its identity isunder investigation.
The cloned TbSMT has been evaluated with the same set ofsubstrates tested with the TbSDM, in addition to zymosteroland cholesta-5,7,24-trienol which naturally occur in theT. brucei sterol composition [14]. The production of hexane-soluble products by the TbSMT as a function of substrateconcentration in the range of 1–100 μM exhibited saturationkinetics, consistent with our previous findings [14]. Computer-assisted data analysis for zymosterol afforded a Km value of47 μM, which is similar ( + / − 5 μM) to that of the othersubstrates tested. However, the catalytic-centre activity forzymosterol is optimal at 0.6 min− 1, and for 14-methylzymosterolat 0.3 min− 1 and cholesta-5,7,24-trienol at 0.2 min− 1. Conversionof 14-methyl zymosterol by TbSMT led to the same type andproportion of multiple products (Supplementary Figure S3at http://www.BiochemJ.org/bj/443/bj4430267add.htm) that wepreviously reported from TbSMT catalysis of zymosterol orcholesta-5,7,24-trienol [14,40]
Origin of carbon atoms in ergostenol biosynthesis
The introduction of 13C-labelled substrates has had an enormousimpact on the study of sterol biosynthesis, in part becausethe investigator studies compounds containing stable isotopes,not radioactive compounds, that can be identified by MS andNMR techniques. The ability to detect incorporation of the13C-label by MS and to locate sites of labelling by 13C-NMRdirectly has led to the elucidation of relevant precursor–productrelationships. Thus incubation of [1-13C]glucose, [2-13C]acetateor [2-13C]leucine to procyclic cells yielded marked changes inthe capacity for sterol biosynthesis compared with a controlincubated with the corresponding non-labelled carbon source.Thus, in the GLC chromatogram of the total sterols from[1-13C]glucose incubation, a new GC peak appeared (5% of themixture) at RRTc 1.14 with M + 398 after the peak correspondingto cholesta-5,7,24-trienol at RRTc 1.11. The chromatographicbehaviour and mass spectrum of this compound agrees with that
c© The Authors Journal compilation c© 2012 Biochemical Society

Sterol biosynthesis in Trypanosoma brucei 273
Figure 5 GC/MS analysis of the total sterols isolated from bloodstream-form cells cultured on FGM
Left-hand panel: identification of peaks based on mass spectra are cholesterol (peak 1, A1), campesterol (peak 2, A2) and sitosterol (peak 3, A3). Note that ergosta-5,7-dienol isolated from13C-incorporation experiments co-migrates with campesterol in this GC system. Right-hand panel: mass and UV (inset) spectral analysis of the HPLC fraction corresponding to [13C]ergosta-5,7-dienol(αc≈0.9–1) from bloodstream forms from incubation with [1-13C]glucose. mAU, milli-absorption units.
of an authentic specimen of ergosta-5,7-dienol available fromour sterol collection. In these studies, we also observed thatthe GC peak corresponding to cholesta-5,7,24-trienol broadenedand progressively increased in height from incubation oflabelled leucine, acetate and glucose respectively. Careful GC/MS
analyses of this GC peak, revealed that the change in peak shapewas due to increases in ergosterol (M + 396) which co-migrateswith cholesta-5,7,24-trienol (M + 382). The ratio of cholesta-5,7,24-trienol to ergosterol in the control and 13C-incorporationexperiments varied from: (i) trace to 99 % from control; to (ii)
c© The Authors Journal compilation c© 2012 Biochemical Society

274 C. R. Nes and others
Figure 6 GC/MS analysis of the TbSDM enzyme-generated fraction from incubation with 31-norlanosterol
Total ion current chromatogram of the substrate 31-norlanosterol (A) and total sterol fraction from the enzyme extract (B). According to GC/MS analysis (right-hand panels), there are two products ofthe reaction; one of them, peak 3, corresponds in retention time and mass spectrum to the expected product (inset is the UV associated with the HPLC fraction corresponding to peak 3). The identityof sterol in peak 2 is unknown.
1 to 99 from [13C]leucine; to (iii) 1 to 1 from [13C]acetate; and(iv) 1 to 10 from [13C]glucose. The 13C-label was incorporatedvariably into ergosterol, ETO and cholesta-5,7,24-trienol showingisotope enrichment factors for each labelled compound thatvaried from 1.30 to 4.27% (Table 1 and Supplementary FigureS4 at http://www.BiochemJ.org/bj/443/bj4430267add.htm). Inthe [13C]leucine experiment, ETO, cholesta-5,7,24-trienol andergosterol were weakly labelled (<5%). The sterol products fromlabelled glucose and acetate possessed the same isotopomericcomposition, suggesting a common origin of their biosynthesis(Supplementary Figure S3). In contrast, cholesterol wasnot labelled from these incubations. These sterol-dependentdifferences from incubation of different 13C-labelled carbonsources are probably explained by mitochondrial and cytosolicinvolvement in metabolite processing affected by two pools ofacetyl-CoA exchanging rapidly, i.e. they are in equilibrium. Takentogether, these results signify that glucose is a better carbon sourcethan acetate and that leucine is not a relevant physiological carbonsource in sterol biosynthesis of procyclic cells.
Previous work has shown the 13C-label in [1-13C]glucose isfound at C2 of acetate [1]. Subsequently, �3-IPP (isopentyldiphosphate), which is formed via the acetate–MVA pathway,becomes labelled in positions at C2, C4 and C5, ultimately givingrise to ergosterol with 15 carbons labelled in the sterol frameat the specific locations shown in Figure 1 for ergosterol 13a.However, for [1-13C]glucose conversion via the plastid-derivedMVA-independent route, the pathway starts with pyruvate andglyceraldehyde 3-phosphate from glycolysis and leads to �3-IPP
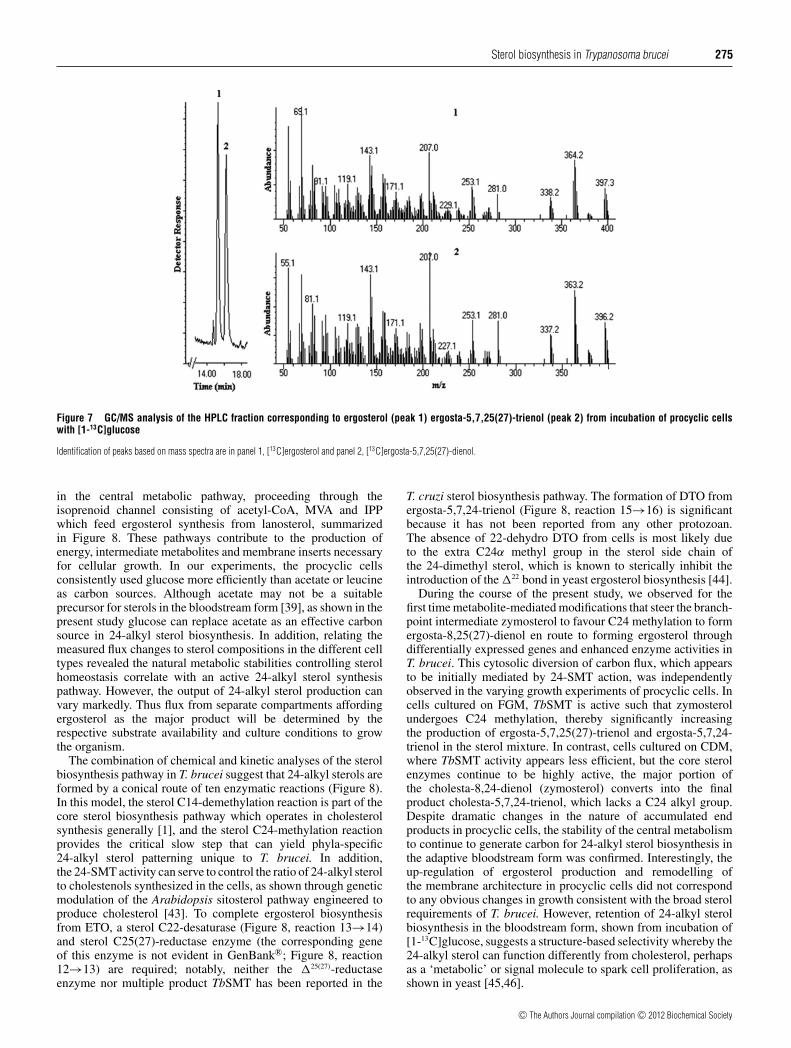
labelled at positions C1 and C5. The resulting ergosterol moleculewill possess 10 labelled carbon atoms (Figure 1, ergosterol13c). 13C incorporation into ergosterol from incubation with [2-13C]leucine can also generate six or 11 labelled carbon atoms(Figure 1, ergosterol 13b), depending on the route of leucinecatabolism. In the present study, labelled ergosterol and ETOproduced by procyclic cells was purified as an approximate 50:50mixture by HPLC. GC/MS analysis of the labelled compoundsshowed similar levels of 13C incorporation (Figure 7). 13C-NMRof the sterol mixture showed site-specific labelling of 15 13C-enriched carbon atoms (carbon number, normalized peak area toC16) at: δ 38.39 (C1, 7.25), 70.45 (C3, 5.88), 139.79 (C5, 1.75),116.31 (C7, 4.00), 46.27 (C9, 4.75), 42.84 (C13 and C24, 4.75),23.00 (C15, 4.5), 55.75 (C17, 4.5), 12.06 (C18, 4.00), 16.29 (C19,6.25), 21.12 (C21, 5.50), 135.7 (C22, 2.75), 33.10 (C25, 1.38),19.66 (C26, 5.25), 19.96 (C27, 5.00)/1.09.37 (C27, 1.27) p.p.m.The enriched signal corresponding to C27 for ergosterol resonatedat 19.96, and for ETO it resonated at 1.09.37, whereas there is noenhanced signal associated with C28 in these 24-alkyl sterols forbiosynthetic reasons [1].
Sterol flux analysis for procyclic and bloodstream forms
The results of the present study from labelling data of sinkmetabolites known to be synthesized in different compartmentsdemonstrate a sterol metabolic network that consists of threedistinct, but coordinated, sets of pathways starting with glucose
c© The Authors Journal compilation c© 2012 Biochemical Society
Sterol biosynthesis in Trypanosoma brucei 275
Figure 7 GC/MS analysis of the HPLC fraction corresponding to ergosterol (peak 1) ergosta-5,7,25(27)-trienol (peak 2) from incubation of procyclic cellswith [1-13C]glucose
Identification of peaks based on mass spectra are in panel 1, [13C]ergosterol and panel 2, [13C]ergosta-5,7,25(27)-dienol.
in the central metabolic pathway, proceeding through theisoprenoid channel consisting of acetyl-CoA, MVA and IPPwhich feed ergosterol synthesis from lanosterol, summarizedin Figure 8. These pathways contribute to the production ofenergy, intermediate metabolites and membrane inserts necessaryfor cellular growth. In our experiments, the procyclic cellsconsistently used glucose more efficiently than acetate or leucineas carbon sources. Although acetate may not be a suitableprecursor for sterols in the bloodstream form [39], as shown in thepresent study glucose can replace acetate as an effective carbonsource in 24-alkyl sterol biosynthesis. In addition, relating themeasured flux changes to sterol compositions in the different celltypes revealed the natural metabolic stabilities controlling sterolhomeostasis correlate with an active 24-alkyl sterol synthesispathway. However, the output of 24-alkyl sterol production canvary markedly. Thus flux from separate compartments affordingergosterol as the major product will be determined by therespective substrate availability and culture conditions to growthe organism.
The combination of chemical and kinetic analyses of the sterolbiosynthesis pathway in T. brucei suggest that 24-alkyl sterols areformed by a conical route of ten enzymatic reactions (Figure 8).In this model, the sterol C14-demethylation reaction is part of thecore sterol biosynthesis pathway which operates in cholesterolsynthesis generally [1], and the sterol C24-methylation reactionprovides the critical slow step that can yield phyla-specific24-alkyl sterol patterning unique to T. brucei. In addition,the 24-SMT activity can serve to control the ratio of 24-alkyl sterolto cholestenols synthesized in the cells, as shown through geneticmodulation of the Arabidopsis sitosterol pathway engineered toproduce cholesterol [43]. To complete ergosterol biosynthesisfrom ETO, a sterol C22-desaturase (Figure 8, reaction 13→14)and sterol C25(27)-reductase enzyme (the corresponding geneof this enzyme is not evident in GenBank®; Figure 8, reaction12→13) are required; notably, neither the �25(27)-reductaseenzyme nor multiple product TbSMT has been reported in the
T. cruzi sterol biosynthesis pathway. The formation of DTO fromergosta-5,7,24-trienol (Figure 8, reaction 15→16) is significantbecause it has not been reported from any other protozoan.The absence of 22-dehydro DTO from cells is most likely dueto the extra C24α methyl group in the sterol side chain ofthe 24-dimethyl sterol, which is known to sterically inhibit theintroduction of the �22 bond in yeast ergosterol biosynthesis [44].
During the course of the present study, we observed for thefirst time metabolite-mediated modifications that steer the branch-point intermediate zymosterol to favour C24 methylation to formergosta-8,25(27)-dienol en route to forming ergosterol throughdifferentially expressed genes and enhanced enzyme activities inT. brucei. This cytosolic diversion of carbon flux, which appearsto be initially mediated by 24-SMT action, was independentlyobserved in the varying growth experiments of procyclic cells. Incells cultured on FGM, TbSMT is active such that zymosterolundergoes C24 methylation, thereby significantly increasingthe production of ergosta-5,7,25(27)-trienol and ergosta-5,7,24-trienol in the sterol mixture. In contrast, cells cultured on CDM,where TbSMT activity appears less efficient, but the core sterolenzymes continue to be highly active, the major portion ofthe cholesta-8,24-dienol (zymosterol) converts into the finalproduct cholesta-5,7,24-trienol, which lacks a C24 alkyl group.Despite dramatic changes in the nature of accumulated endproducts in procyclic cells, the stability of the central metabolismto continue to generate carbon for 24-alkyl sterol biosynthesis inthe adaptive bloodstream form was confirmed. Interestingly, theup-regulation of ergosterol production and remodelling ofthe membrane architecture in procyclic cells did not correspondto any obvious changes in growth consistent with the broad sterolrequirements of T. brucei. However, retention of 24-alkyl sterolbiosynthesis in the bloodstream form, shown from incubation of[1-13C]glucose, suggests a structure-based selectivity whereby the24-alkyl sterol can function differently from cholesterol, perhapsas a ‘metabolic’ or signal molecule to spark cell proliferation, asshown in yeast [45,46].
c© The Authors Journal compilation c© 2012 Biochemical Society

276 C. R. Nes and others
Figure 8 Compartmented metabolic network and fluxes in Trypanosoma brucei procyclic cells yielding membrane sterol from glucose and acetyl-CoA
The scheme illustrates probable entrance points of the 13C-labelled carbon sources used in the present study into the mainframe biosynthesis pathways. The sterol biosynthesis pathway is definedby enzymes that catalyse the sterol structure, and the relevant enzymatic steps are shown associated with the endoplasmic reticulum. The ten enzymatic steps involved in conversion of lanosterolinto ergosterol are shown by the solid arrows. Several reaction steps are repeated by four of these enzymes, sterol C4-methyl oxidase [1→2], sterol C4-decarboxylase [2→3], sterol C3-reductase[3→4] and 24-SMT [7→8,9,10], using different substrates. G3P, glucose 3-phosphate; DHAP, dihydroxy acetonephosphate; PEP, phosphoenolpyruvate; Sq, squalene; SO, squalene oxide.
Conclusions
The current study documents a novel 24-alkyl sterol biosynthesispathway in which all but one enzymes [sterol C25(27)-reductase]have been detected by protein–protein BLAST searches of thecatalogue of predicted T. brucei sterol biosynthetic enzymes(results not shown). In both the procyclic and bloodstreamform, the energy-expensive C24-methylation pathway continuesto play an essential role in sterol biosynthesis so long as glucosemetabolism is not limiting. The reason why T. brucei synthesizesmultiple 24-alkyl sterol products is not clear; if some of themare found to be functionally redundant, such as fecosterol andDTO, then we have a glimpse of sterol evolution at play inwhich separate routes are available for sterol biosynthesis butonly one of them yields physiologically relevant end products. Wesurmise that the major factors affecting changes in the proportionof ergostenols to cholestenols in procyclic and bloodstream formsmay operate at the level of compartmented metabolic fluxes linkedto gene expression regulation through a sterol-sensing mechanismof the sort utilized by opportunistic pathogens [47]. More detailedstudies on the factors that modulate biosynthesis of the unusualergostenols in trypanosomatids, and the selective pressures thatdrive this process, are warranted.
AUTHOR CONTRIBUTION
Fernando Villata, Michael Waterman, Galina Lepesheva, Minu Chaudhuri and W. DavidNes designed the experiments and analysed data; Craigen Nes, Ujjal Singha, JialinLiu, Kulothungan Ganapathy, Galina Lepesheva and Minu Chaudhuri performed theexperiments; and W. David Ness wrote the paper.
FUNDING
This work was supported, in whole or in part, by the National Science Foundation [grantnumber MCB-0929212 (to W.D.N.)], and the National Institutes of Health [grant numbersGM 067871 (to M.R.W. and G.I.L.), AI 080580 (to F.V.) and GM 081146 (to M.C.)].
REFERENCES
1 Nes, W. D. (2011) Biosynthesis of cholesterol and other sterols. Chem. Rev. 111,6423–6451
2 Ginger, M. L., McFadden, G. I. and Michels, P. A. M. (2010) Rewiring and regulation ofcross-compartmentalized metabolism in protists. Philos. Trans. R. Soc. London Ser. B365, 831–845
3 Masse, G., Belt, S., Rowland, S. J. and Rohmer, M. (2004) Isoprenoid biosynthesis in thediatoms Rhizosolenia setigera (Brightwell) and Haslea ostrearia (Simonsen). Proc. Natl.Acad. Sci. U.S.A. 101, 4413–4418
c© The Authors Journal compilation c© 2012 Biochemical Society

Sterol biosynthesis in Trypanosoma brucei 277
4 Zhou, W., Nguyen, H. T. and Nes, W. D. (2008) Plant sterol methyltransferase:phytosterolomic analysis, enzymology and bioengineering strategies. In Advances in PlantBiochemistry and Molecular Biology (Lewis, N. G., ed.), pp. 242–281, Elsevier, Oxford
5 Kodner, R. B., Summons, R. E., Pearson, A., King, N. and Knoll, A. W. (2008) Sterols in aunicellular relative of the metazoans. Proc. Natl. Acad. Sci U.S.A. 105, 9897–9902
6 Nes, W. D. (2005) Enzyme redesign and interactions of substrate analogues with sterolmethyltransferase to understand phytosterol diversity, reaction mechanism and the natureof the active site. Biochem. Soc. Trans. 33, 1189–1196
7 Waldbauer, J. R., Newman, D. K. and Summons, R. E. (2011) Microaerobic steroidbiosynthesis and the molecular fossil record of Archean life. Proc. Natl. Acad. Sci. U.S.A.108, 13409–13414
8 Croft, S. L., Barrett, M. P. and Urbina, J. O. (2005) Chemotherapy of trypanosomiases andleishmaniasis. Trends Parasitol. 21, 508–512
9 Roberts, C. W., McLeod, R, Rice, D. W., Ginger, M., Chance, M. L. and Goad, L. J. (2003)Fatty acid and sterol metabolism: potential antimicrobial targets in apicomplexan andtrypanosomatid parasitic protozoa. Mol. Biochem. Parasitol. 126, 129–142
10 de Souza, W. D and Rodrigues, J. C. F. (2009) Sterol biosynthesis pathway as target foranti-trypanosomatid drugs. Interdiscip. Perspect. Infect. Dis. 2009, 1–19
11 Buckner, F. S. (2008) Sterol 14-demethylase inhibitors for Trypanosoma cruzi infections.Adv. Exp. Med. Biol. 625, 61–80
12 Lepesheva, G. I., Nes, W. D., Zhou, W., Hill, G. C. and Waterman, M. R. (2004) CYP51from Trypanosoma brucei is obtusifoliol specific. Biochemistry 43, 10789–10799
13 Zhou, W., Lepesheva, G. I., Waterman, M. R. and Nes, W. D. (2006) Mechanistic analysisof a multiple product sterol methyltransferase from Trypanosoma brucei implicated inergosterol biosynthesis. J. Biol. Chem. 281, 6290–6296
14 Zhou, W., Cross, G. A. M. and Nes, W. D. (2007) Cholesterol import fails to preventcatalyst-based inhibition of ergosterol synthesis and cell proliferation in Trypanosomabrucei. J. Lipid Res. 48, 665–673
15 Lepesheva, G. I., Ott, R. D., Hargrove, T. Y., Kleshchenko, Y. Y., Schuster, I., Nes, W. D.,Hill, G. C., Villalta, F. and Waterman, M. R. (2007) Sterol 14α-demethylase inhibitors aspotential target for antitrypanosomal therapy: enzyme inhibition and parasite growth.Chem. Biol. 14, 1–11
16 Lepesheva, G. I., Park, H.-W., Hargrove, T. Y., Vanhollebeke, B., Zdzislaw, W., Harp, J. M.,Sundaramoorthy, M., Nes, W. D., Pays, E., Villalta, F. and Waterman, M. R. (2010) Crystalstructures of ligand-free and inhibitor-bound sterol 14α-demethylase from Trypansomabrucei 99. J. Biol. Chem. 285, 1773–1780
17 Hargrove, T. Y., Liu, J., Nes, W. D., Waterman, M. R. and Lepesheva, G. I. (2011) Substratepreferences and catalytic parameters determined by structural characteristics of sterol14α-demethylase CYP51 from Leishmania infantum. J. Biol. Chem. 286, 26838–26848
18 Dixon, H., Ginger, C. D. and Williamson, J. (1972) Trypanosome sterols and theirmetabolic origin. Comp. Biochem. Physiol. 41B, 1–18
19 Coppens, I. and Courtoy, P. J. (1995) Exogenous and endogenous sources of sterols inthe cultured-adapted procyclic trypomastigotes of Trypanosoma brucei. Mol. Biochem.Parasitol. 73, 179–188
20 Gros, L., Castillo-Acosta, V. M., Jimenez, C. J., Sealey-Cardona, M., Vargas, S., Estevez,A. M., Yardley, V., Rattray, L., Croft, S. L., Ruiz-Perez, L. M. et al. (2006) New azasterolsagainst Trypanosoma brucei: role of 24-sterol methyl transferase in inhibitor action.Antimicrob. Agents Chemother. 50, 2595–2601
21 Alsford, S., Turner, D. J., Obado, S. O., Shanchez-Flores, Z., Glover, L., Berriman, M.,Hertz-Fowler, C. and Horn, D. (2010) High-throughput phenotyping using parallelsequencing of RNA interference targets in the African trypanosome. Genome Res. 21,915–924
22 Coopens, I and Courtoy, J. C.F. (2000) The adaptive mechanisms of Trypanosoma bruceifor sterol homeostasis in its different life-cycle environments. Annu. Rev. Microbiol. 54,129–156
23 Hinshaw, J. C., Suh, D. M., Garrier, P., Buckner, F. S., Eastman, R. T., Matsuda, S. P.,Joubert, B. M., Coopens, I., Joiner, K. A., Merali, S. et al. (2003) Oxidocyclase inhibitorsas antimicrobial agents. J. Med. Chem. 46, 4240–4243
24 Furlong, S. T. (1989) Sterols of parasitic protozoa and helminths. Exp. Parasitol. 68,482–485
25 Patterson, G. W. (1994) Phylogenetic distribution of sterols. ACS Symp. Ser. 562, 90–10826 Halvey, S. (1991) Sterols, toxins and parasitism. Med. Hypotheses 36, 204–20527 Martin, W. and Borst, P. (2003) Secondary loss of chloroplasts in trypanosomes. Proc.
Natl. Acad. Sci. U.S.A. 100, 765–76728 Cavalier-Smith, T. (2003) Protist phylogeny and the high-level classification of protozoa.
Eur. J. Protistol. 39, 338–34829 Hunter, W. N. (2007) The non-mevalonate pathway of isoprenoid biosynthesis. J. Biol.
Chem. 282, 21573–2157730 Zhou, W.-X. and Nes, W. D. (2000) Stereochemistry of hydrogen introduction at C-25 in
ergosterol synthesized by the mevalonate-independent pathway. Tetrahedron Lett. 41,2791–2795
31 Buckner, F. S., Nguyen, L. N., Joubert, B. M. and Matsuda, S. P. (2000) Cloning andheterologous expression of the Trypanosoma brucei lanosterol synthase gene. Mol.Biochem. Parasitol. 110, 399–403
32 Smith, T. K. and Butikofer, P. (2010) Lipid metabolism in Trypanosoma brucei. Mol.Biochem. Parasitol. 172, 66–79
33 Ginger, M. L., Chance, M. L. and Goad, L. J. (1999) Elucidation of carbon sources usedfor the biosynthesis of fatty acids and sterols in the trypanosomatid Leishmaniamexicana. Biochem. J. 342, 397–405
34 Ginger, M. L., Prescott, M. C., Reynolds, D. G., Chance, M. L. and Goad, L J. (2000)Utilization of leucine and acetate as carbon sources for sterol and fatty acid biosynthesisby Old and New World Leishmania species, Endotrypanum monterogeii and Trypanosomacruzi. Eur. J. Biochem 267, 2555–2566
35 Ginger, M. L., Chance, M. L., Sadler, I. H. and Goad, L. J. (2001) The biosyntheticincorporation of the intact leucine skeleton into sterol by the trypanosomatid Leishmaniamexicana. J. Biol. Chem. 276, 11674–11682
36 Riviere, L., Moreau, P., Allmann, S., Hahn, M., Biran, M., Plazolles, N., Franconi, J.-M.,Boshart, M. and Bringaud, F. (2009) Acetate produced in the mitochondrion is theessential precursor for lipid biosynthesis in procylic cell trypanosomes. Proc. Natl. Acad.U.S.A. 106, 12694–12699
37 Coustou, V., Biran, M., Breton, M., Guegan, F., Riveiere, L., Plazolles, N., Nolan, D.,Barrett, M. P., Franconi, J.-M. and Bringaud, F. (2008) Glucose-induced remodeling ofintermediary and energy metabolism in procyclic Trypanosoma brucei. J. Biol. Chem.283, 16342–16354
38 Zhou, W., Guo, D. and Nes, W. D. (1996) Stereochemistry of hydrogen migration fromC-24 to C-25 during biomethylation in ergosterol biosynthesis. Tetrahedron Lett. 37,1339–1342
39 Liu, J., Ganapathy, K., Wywial, E., Bujnicki, J. M., Nwogwugwu, C. A. and Nes, W. D.(2011) Effect of substrate features and mutagenesis of active site tyrosine residues on thereaction course catalyzed by Trypanosoma brucei sterol C24-methyl transferase.Biochem. J. 439, 413–422
40 Norton, R. A. and Nes, W. D. (1991) Identification of ergosta-6(8),8(14),25(27)-trien-3β-ol and ergosta-5(6),7(8),25(27)-trien-3β-ol: two new steroidal trienessynthesized by Prototheca wickerhamii. Lipids 26, 247–249
41 Goad, L. J. and Akihsa, T. (1997) Analysis of sterols, Blackie Academic, London42 Dixon, H., Ginger, C. D. and Williamson, J. (1971) The lipid metabolism of blood and
culture forms of Trypanosoma lewis and Trypanosoma rhodesiense. Comp. Biochem.Physiol. 39B, 247–266
43 Diener, A. C., Li, H., Zhou, W.-X., Whoriskey, W. J., Nes, W. D. and Fink, G. R. (2000)Sterol methyl transferase I (SMTI) controls the level of cholesterol in plants. Plant Cell12, 853–870
44 Li, Y. (1996) Stereochemical studies on the metabolism of sterols by Saccharomycescerevisiae stain GL 7. Masters Thesis, Texas Tech University, Lubbock, Texas, U.S.A.
45 Nes, W. R. (1987) Structure–function relationships for sterols in Saccharomycescerivisiae. ACS Symp. Ser. 325, 252–267
46 Rodriquez, R. J., Low, C., Bottema, C. D. K. and Parks, L. W. (1985) Multiple functions forsterols in Saccharomyces cerevisiae Biochim. Biophys. Acta 837, 336–343
47 Bien, C. M. and Espenshade, P. J. (2010) Sterol regulatory element binding proteins infungi: hypotoxic transcription factors linked to pathogenesis. Eukaryotic Cell 9,352–359
Received 17 October 2011/5 December 2011; accepted 19 December 2011Published as BJ Immediate Publication 19 December 2011, doi:10.1042/BJ20111849
c© The Authors Journal compilation c© 2012 Biochemical Society
Biochem. J. (2012) 443, 267–277 (Printed in Great Britain) doi:10.1042/BJ20111849
SUPPLEMENTARY ONLINE DATANovel sterol metabolic network of Trypanosoma brucei procyclic andbloodstream formsCraigen R. NES*1, Ujjal K. SINGHA†1, Jialin LIU*, Kulothungan GANAPATHY*, Fernando VILLALTA†, Michael R. WATERMAN‡,Galina I. LEPESHEVA‡, Minu CHAUDHURI† and W. David NES*2
*Department of Chemistry and Biochemistry, Texas Tech University, Lubbock, TX 79409, U.S.A., †Department of Microbiology and Immunology, Meharry Medical College, Nashville,TN 37208, U.S.A., and ‡Department of Biochemistry, Vanderbilt University School of Medicine, Nashville, TN 37232, U.S.A.
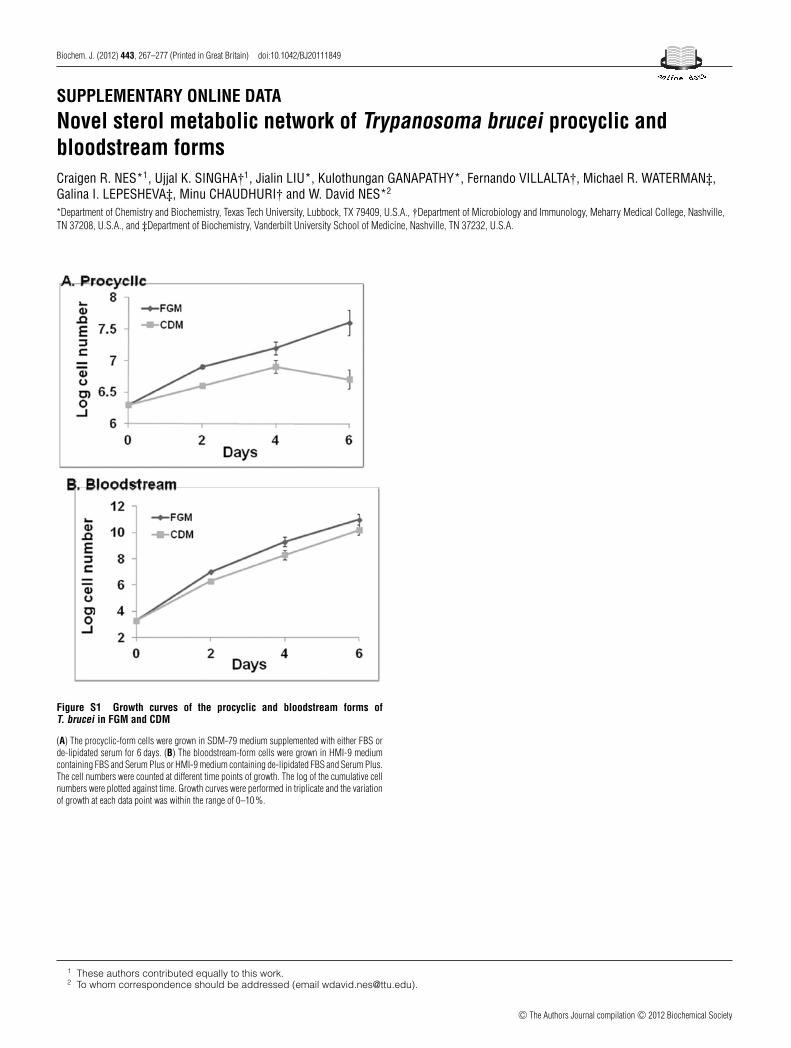
Figure S1 Growth curves of the procyclic and bloodstream forms ofT. brucei in FGM and CDM
(A) The procyclic-form cells were grown in SDM-79 medium supplemented with either FBS orde-lipidated serum for 6 days. (B) The bloodstream-form cells were grown in HMI-9 mediumcontaining FBS and Serum Plus or HMI-9 medium containing de-lipidated FBS and Serum Plus.The cell numbers were counted at different time points of growth. The log of the cumulative cellnumbers were plotted against time. Growth curves were performed in triplicate and the variationof growth at each data point was within the range of 0–10 %.
1 These authors contributed equally to this work.2 To whom correspondence should be addressed (email [email protected]).
c© The Authors Journal compilation c© 2012 Biochemical Society
C. R. Nes and others
Figure S2 Mass spectra (EI, 70 eV) of the peaks corresponding to the GC/MS analyses shown in Figure 4 of the main text
c© The Authors Journal compilation c© 2012 Biochemical Society

Sterol biosynthesis in Trypanosoma brucei
Figure S3 GC/MS analysis of the total TbSMT reaction product from incubation of 14-methylzymosterol
Mass spectra of the peaks 1 (substrate) to 2–5 (products) are shown along with their proposed structures.
c© The Authors Journal compilation c© 2012 Biochemical Society

C. R. Nes and others
Figure S4 Mass spectra (EI, 70 eV) at the high mass end (abundance is generally normalized to M + 386 in cholesterol the spectrum) of the 13C- and2H-biosynthetically formed sterols, ergosterol-5,7,25(27)-trienol, ETO, ergosterol, ERG and cholesta-5,7,24-treinol from the corresponding incubations ofprocyclic cells
The spectra are taken from GC peaks (which for some treatments contained mixtures of ergosterol and cholesta-5,7,24-trienol): (A), control; (B), [2-13C]acetate; (C), [1-13C]glucose; (D), [2-13C]leucine;(E), [methyl-2H3]methionine; and (F), [27-13C]lanosterol. In none of the incubations was cholesterol labelled, and in the incubation with [methyl-2H3]methionine ergosterol was not detected in thetotal sterol mixture by GC/MS analysis.
Received 17 October 2011/5 December 2011; accepted 19 December 2011Published as BJ Immediate Publication 19 December 2011, doi:10.1042/BJ20111849
c© The Authors Journal compilation c© 2012 Biochemical Society




















