
In vivo targeting of dendritic cells in lymph nodes with poly(propylene sulfide) nanoparticles
9
In vivo targeting of dendritic cells in lymph nodes with poly(propylene sulfide) nanoparticles Sai T. Reddy a , Annemie Rehor a , Hugo G. Schmoekel a,b , Jeffrey A. Hubbell a , Melody A. Swartz a, ⁎ a Institute of Bioengineering, École Polytechnique Fédérale de Lausanne (EPFL) CH-1015 Lausanne, Switzerland b Polyclinic Small Animal Medicine, FU-Berlin, D-14163 Berlin, Germany Received 8 September 2005; accepted 16 January 2006 Available online 10 March 2006 Abstract Delivery of biodegradable nanoparticles to antigen-presenting cells (APCs), specifically dendritic cells (DCs), has potential for immunotherapy. This study investigates the delivery of 20, 45, and 100nm diameter poly(ethylene glycol)-stabilized poly(propylene sulfide) (PPS) nanoparticles to DCs in the lymph nodes. These nanoparticles consist of a cross-linked rubbery core of PPS surrounded by a hydrophilic corona of poly(ethylene glycol). The PPS domain is capable of carrying hydrophobic drugs and degrades within oxidative environments. 20 nm particles were most readily taken up into lymphatics following interstitial injection, while both 20 and 45 nm nanoparticles showed significant retention in lymph nodes, displaying a consistent and strong presence at 24, 72, 96 and 120 h post-injection. Nanoparticles were internalized by up to 40–50% of lymph node DCs (and APCs) without the use of a targeting ligand, and the site of internalization was in the lymph nodes rather than at the injection site. Finally, an increase in nanoparticle-containing DCs (and other APCs) was seen at 96h vs. 24h, suggesting an infiltration of these cells to lymph nodes. Thus, PPS nanoparticles of 20–45nm have the potential for immunotherapeutic applications that specifically target DCs in lymph nodes. © 2006 Elsevier B.V. All rights reserved. Keywords: Antigen-presenting cells; Vaccines; Immunotherapy; Macrophages 1. Introduction Antigen-presenting cells (APCs) are highly efficient phago- cytic cells that utilize MHC class I, II and other co-stimulatory molecules to stimulate naive T cells and induce cell-mediated immunity. APCs, which include some macrophages and the more potent dendritic cells (DCs), are present in peripheral tissues where they act as sentinels that, following internalization and processing of foreign antigens, subsequently undergo maturation and migration to lymph nodes for the purpose of antigen presentation to T cells [1–3]. With the critical roles that APCs and DCs play in adaptive immunity, new strategies are being developed to target these cells with immunomodulating agents, such as DNA, proteins, and peptides [4–14]. Polymer- and liposome-based delivery systems have focused primarily on delivery of protein and DNA to peripheral DCs, where they first internalize the drug vehicles and then migrate to lymph nodes within ∼ 1–2 days to activate T cells [9,12,13]. Until recently, it was not clear whether immature DCs, capable of taking up antigens, were present in lymph nodes. However, recent studies have established that a substantial fraction of resident DCs in the lymph nodes are phenotypically immature and capable of internalizing antigens and particles there [15,16]; thus, resident lymph node APCs may also be utilized as targets for immunotherapeutic drugs. A potential benefit of targeting lymph node APCs or DCs instead of those in peripheral sites is that premature antigen presentation (i.e., a migrating DC that expresses antigen on its surface prior to reaching a lymph node) can often lead to immune cell tolerance [13,17,18]; therefore, delivery to lymph node APCs may potentially avert this Journal of Controlled Release 112 (2006) 26 – 34 www.elsevier.com/locate/jconrel ⁎ Corresponding author. Laboratory for Mechanobiology and Morphogenesis, Institute of Bioengineering, Station 15, École Polytechnique Fédérale de Lausanne (EPFL) CH-1015 Lausanne, Switzerland. Tel.: +41 21 693 9686; fax: +41 21 693 9685. E-mail address: [email protected] (M.A. Swartz). 0168-3659/$ - see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.jconrel.2006.01.006
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of In vivo targeting of dendritic cells in lymph nodes with poly(propylene sulfide) nanoparticles

e 112 (2006) 26–34www.elsevier.com/locate/jconrel
Journal of Controlled Releas
In vivo targeting of dendritic cells in lymph nodes withpoly(propylene sulfide) nanoparticles
Sai T. Reddy a, Annemie Rehor a, Hugo G. Schmoekel a,b,Jeffrey A. Hubbell a, Melody A. Swartz a,⁎
a Institute of Bioengineering, École Polytechnique Fédérale de Lausanne (EPFL) CH-1015 Lausanne, Switzerlandb Polyclinic Small Animal Medicine, FU-Berlin, D-14163 Berlin, Germany
Received 8 September 2005; accepted 16 January 2006Available online 10 March 2006
Abstract
Delivery of biodegradable nanoparticles to antigen-presenting cells (APCs), specifically dendritic cells (DCs), has potential forimmunotherapy. This study investigates the delivery of 20, 45, and 100nm diameter poly(ethylene glycol)-stabilized poly(propylene sulfide)(PPS) nanoparticles to DCs in the lymph nodes. These nanoparticles consist of a cross-linked rubbery core of PPS surrounded by a hydrophiliccorona of poly(ethylene glycol). The PPS domain is capable of carrying hydrophobic drugs and degrades within oxidative environments. 20 nmparticles were most readily taken up into lymphatics following interstitial injection, while both 20 and 45nm nanoparticles showed significantretention in lymph nodes, displaying a consistent and strong presence at 24, 72, 96 and 120h post-injection. Nanoparticles were internalized by upto 40–50% of lymph node DCs (and APCs) without the use of a targeting ligand, and the site of internalization was in the lymph nodes rather thanat the injection site. Finally, an increase in nanoparticle-containing DCs (and other APCs) was seen at 96h vs. 24h, suggesting an infiltration ofthese cells to lymph nodes. Thus, PPS nanoparticles of 20–45nm have the potential for immunotherapeutic applications that specifically targetDCs in lymph nodes.© 2006 Elsevier B.V. All rights reserved.
Keywords: Antigen-presenting cells; Vaccines; Immunotherapy; Macrophages
1. Introduction
Antigen-presenting cells (APCs) are highly efficient phago-cytic cells that utilize MHC class I, II and other co-stimulatorymolecules to stimulate naive T cells and induce cell-mediatedimmunity. APCs, which include some macrophages and themore potent dendritic cells (DCs), are present in peripheraltissues where they act as sentinels that, following internalizationand processing of foreign antigens, subsequently undergomaturation and migration to lymph nodes for the purpose ofantigen presentation to T cells [1–3]. With the critical roles thatAPCs and DCs play in adaptive immunity, new strategies are
⁎ Corresponding author. Laboratory for Mechanobiology and Morphogenesis,Institute of Bioengineering, Station 15, École Polytechnique Fédérale deLausanne (EPFL) CH-1015 Lausanne, Switzerland. Tel.: +41 21 693 9686; fax:+41 21 693 9685.
E-mail address: [email protected] (M.A. Swartz).
0168-3659/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.jconrel.2006.01.006
being developed to target these cells with immunomodulatingagents, such as DNA, proteins, and peptides [4–14].
Polymer- and liposome-based delivery systems have focusedprimarily on delivery of protein and DNA to peripheral DCs,where they first internalize the drug vehicles and then migrate tolymph nodes within ∼1–2 days to activate T cells [9,12,13].Until recently, it was not clear whether immature DCs, capableof taking up antigens, were present in lymph nodes. However,recent studies have established that a substantial fraction ofresident DCs in the lymph nodes are phenotypically immatureand capable of internalizing antigens and particles there [15,16];thus, resident lymph node APCs may also be utilized as targetsfor immunotherapeutic drugs. A potential benefit of targetinglymph node APCs or DCs instead of those in peripheral sites isthat premature antigen presentation (i.e., a migrating DC thatexpresses antigen on its surface prior to reaching a lymph node)can often lead to immune cell tolerance [13,17,18]; therefore,delivery to lymph node APCs may potentially avert this

27S.T. Reddy et al. / Journal of Controlled Release 112 (2006) 26–34
problem. Additionally, other DC targeting studies use conju-gated ligands such as anti-Dec-205 and anti-CD11c to increaseDC specificity [4,5,8,9,12,19]. However, since DCs are bynature highly phagocytic and present in lymph nodes at highconcentrations, it may be possible to specifically target thesecells in lymph nodes without the use of a targeting ligand.
In order to target APCs, including DCs, in lymph nodes, it iscritical to design drug delivery vehicles that can be readily takenup into lymphatic vessels and retained in draining lymph nodes.A primary role of the lymphatic system is the uptake of fluid andparticulates from the interstitial space as a small but importantcomponent of the microcirculation [20–23]. Previous in vivolymphatic targeting systems using liposomes and polymerparticles have established that particle size is the most criticalfactor for lymphatic uptake from the interstitial space [21,24–29]. Liposomes larger than 170nm generally show poorlymphatic uptake and remain at the injection site, whereasparticles in the range of 40–70nm show significant uptake intolymphatic vessels [25]. Additionally, it is likely that interactionsbetween the particle surface and the interstitium play a role inthis incomplete lymphatic uptake [30]. Steric stabilization bycoating liposomes and particles with hydrophilic layers such aspoly(ethylene glycol) (PEG) and its copolymers such asPluronics (copolymers of poly(ethylene glycol)-bl-poly(propyl-ene glycol)-bl-poly(ethylene glycol)) can reduce the non-specific interactions with proteins of the interstitium asdemonstrated by improved lymphatic uptake following subcu-taneous injections [21,27,31–35].
However, while smaller particles are taken up more readily,they also pass through the lymph node more easily; in this wayit can be difficult to achieve both efficient lymphatic uptake andlymph node retention. For example, it has been observed thatonly 1–2% of injected 70nm liposomes are retained in thelymph nodes beyond 12h post-injection [30], and that noderetention of large liposomes (>70nm) is more efficient than thatof smaller liposomes [24,29]. This apparently is due in part tothe fact that lymph node macrophages phagocytose largerparticles more efficiently. While it is assumed that coating ofliposomes with steric protectants such as PEG should reducephagocytosis by macrophages, it has been shown that suchcoating does not significantly affect lymph node retention [36].It has also been generally assumed that particles phagocytosedin the lymph nodes are done so primarily by macrophages[21,27,29,30,32,36,37]. Therefore, it will be of criticalimportance for lymph node targeting with immunomodulatingdrugs to determine whether, in addition to macrophages, thatAPCs, primarily DCs are also phagocytosing particles that reachthe lymph nodes.
As described above, both size and surface characteristics arecritical to nanoparticle uptake and retention in lymph nodes.This study employs a polymer nanoparticle system based onpoly(propylene sulfide) (PPS) and PEG that has been recentlydeveloped and characterized for systemic delivery [38,39]. Thenanoparticles consist of a hydrophobic rubbery core of cross-linked PPS surrounded by a hydrophilic corona of PEG derivedfrom a Pluronic that was used as a stabilizer during synthesis asan emulsion. PPS is an interesting polymer because it degrades
under oxidative conditions to the polysulfoxide and finally thepolysulfone [40], thus transitioning from a hydrophobic rubberto a hydrophilic, water-soluble polymer [38]. Using micro-emulsion approaches, PEG-stabilized PPS nanoparticles can beformed with diameters as low as 20nm and as large as 250nm.Furthermore, the chain termini on the PEG stabilizing thenanoparticles can be readily functionalized for specific celltargeting (unpublished data, A. Rehor, N. Tirelli, J.A. Hubbell)or protein and peptide drug delivery.
The goal of this study was to design an effective nanoparticlesystem to target the DCs and other APCs located in lymphnodes. The effects of nanoparticle size on lymphatic uptake,lymph node retention, and internalization by lymph node APCsand DCs were explored following intradermal injections in mice(which have similar size lymphatic capillaries as those inhumans–10–80μm–although these are highly variable in bothspecies [20,41]). It was found that 20nm particles are morereadily taken up into the lymphatics than 45 and 100nm, andthat they are retained in the lymph nodes for longer times (up to120h) than have been previously reported [31–34,36]. Withinthe lymph node, it was shown that the nanoparticles areinternalized exclusively by resident APCs (MHCII+ cells whichinclude DCs and some macrophages) and other non-antigenpresenting macrophages (MHCII−) without a targeting ligand.A large fraction (up to 50%) of lymph node resident DCsinternalized the 20nm particles, with the number increasingover time. PPS nanoparticles thus offer the potential for a novelstrategy to deliver immunotherapeutic agents to DCs and otherAPCs in lymph nodes.
2. Materials and methods
2.1. Nanoparticles
PPS nanoparticles with diameters of 20, 45, and 100nm weresynthesized by emulsion polymerization as described elsewhere[38]. Briefly, an emulsion was created by adding the PEG blockcopolymer emulsifier, Pluronic F-127 (Sigma-Aldrich, Buchs,Switzerland) and the monomer propylene sulfide to ultrapuremilliQ water under constant stirring. The protected initiatorpentaerythritol tetrathioester was synthesized as describedelsewhere [38] and, in a separate flask, was deprotected bymixing it with 0.20ml of 0.5M sodiummethylate solution understirring for 10min. Following deprotection, the initiator wasthen added to the monomer emulsion and 5min later 60μl of thebase diaza[5.4.0]bicycycloundec-7-ene (DBU) was added to thereaction and it was allowed to stir continuously for 24h under aninert atmosphere. The nanoparticles were then exposed to air inorder to produce disulfide cross-linking.
The nanoparticles were purified from remaining monomers,base, or free Pluronic by 2 days of repeated dialysis with a 12–14kDa MWCO membrane (Spectrum Laboratories, RanchoDominguez, CA) against ultrapure milliQ water. The nanopar-ticle size distributions were determined by the use of a dynamiclight scattering instrument (Malvern, Worcestershire, UnitedKingdom). Fluorescent labeling was accomplished by adding 6-iodoacetamido-fluorescein or Alexa Fluor 488 malemeide
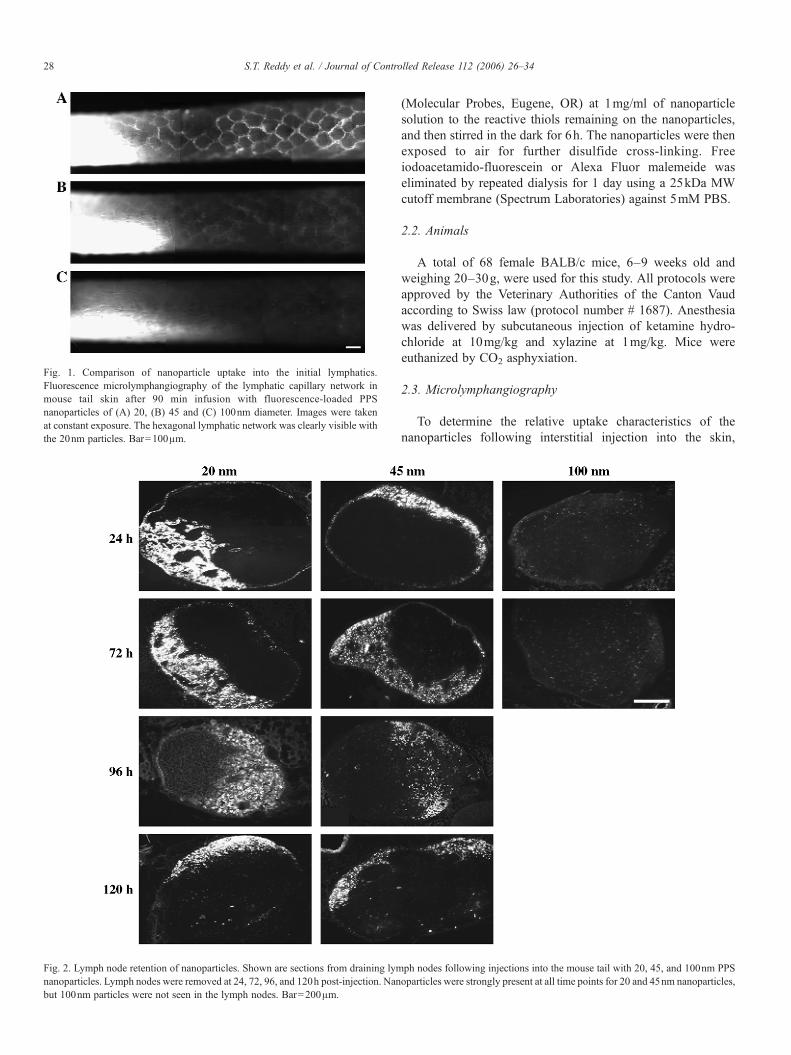
Fig. 1. Comparison of nanoparticle uptake into the initial lymphatics.Fluorescence microlymphangiography of the lymphatic capillary network inmouse tail skin after 90 min infusion with fluorescence-loaded PPSnanoparticles of (A) 20, (B) 45 and (C) 100nm diameter. Images were takenat constant exposure. The hexagonal lymphatic network was clearly visible withthe 20nm particles. Bar=100μm.
Fig. 2. Lymph node retention of nanoparticles. Shown are sections from draining lymnanoparticles. Lymph nodes were removed at 24, 72, 96, and 120h post-injection. Nanbut 100nm particles were not seen in the lymph nodes. Bar=200μm.
28 S.T. Reddy et al. / Journal of Controlled Release 112 (2006) 26–34
(Molecular Probes, Eugene, OR) at 1mg/ml of nanoparticlesolution to the reactive thiols remaining on the nanoparticles,and then stirred in the dark for 6h. The nanoparticles were thenexposed to air for further disulfide cross-linking. Freeiodoacetamido-fluorescein or Alexa Fluor malemeide waseliminated by repeated dialysis for 1 day using a 25kDa MWcutoff membrane (Spectrum Laboratories) against 5mM PBS.
2.2. Animals
A total of 68 female BALB/c mice, 6–9 weeks old andweighing 20–30g, were used for this study. All protocols wereapproved by the Veterinary Authorities of the Canton Vaudaccording to Swiss law (protocol number # 1687). Anesthesiawas delivered by subcutaneous injection of ketamine hydro-chloride at 10mg/kg and xylazine at 1mg/kg. Mice wereeuthanized by CO2 asphyxiation.
2.3. Microlymphangiography
To determine the relative uptake characteristics of thenanoparticles following interstitial injection into the skin,
ph nodes following injections into the mouse tail with 20, 45, and 100nm PPSoparticles were strongly present at all time points for 20 and 45nm nanoparticles,

29S.T. Reddy et al. / Journal of Controlled Release 112 (2006) 26–34
fluorescence microlymphangiography was performed by con-stant pressure injection into the tip of the tail as previouslydescribed [42–44]. Hair was depilated from the mouse tail, andthe mouse was positioned on the microscope stage (Axiovert200M, Zeiss) with a heating pad to maintain 37°C bodytemperature. A 20mg/ml solution of fluorescent PPS nanopar-ticles (20, 45, or 100nm diameter) in sterile phosphate-bufferedsaline (PBS) was drawn into a catheter; a 30-gauge needleattached to the catheter was inserted intradermally at ∼1mmfrom the tail tip and a stopcock was opened that initiated flow ata constant pressure of 40mm Hg. The flow rate of thenanoparticle solution (monitored with a bubble far upstreamin the tubing) averaged 0.1μl/min into roughly 20mm3 of tissue(approximated from the volume visible from the injection
Fig. 3. Localization of nanoparticles within the lymph node. Shown are serial lympImmune cells were identified as indicated (red) with antibodies against (A) CD3e (Tcells (DCs)); nuclei are shown in blue. Nanoparticles are distinctly absent in T cBar=100μm. (D) The endothelial marker CD31 (red) demonstrates nanoparticle (gr
depot), or approximately 5μl/g/min. No visible swelling wasobserved. Sequential images along the length of the tail werecollected at a constant exposure time; 3 experiments wererepeated for each nanoparticle size.
2.4. Evaluation of nanoparticle distribution in lymph nodes
To evaluate lymph node retention, 20μl of 20mg/mlfluorescent PPS nanoparticles (20, 45, and 100nm diameter)was injected as a bolus into the tip of the mouse tail or frontfootpad through a 30-gauge needle; controls were performedwith 20μl injections of PBS. No inflammation was observed atthe sites of injection. At 24, 72, 96, or 120h, mice weresacrificed by CO2 asphyxiation. The sacral and lumbar lymph
h node sections at 96h following injection of 20nm PPS nanoparticles (green).cells), (B) CD45R (B cells), and (C) CD68 (macrophages (Macs) and dendriticell and B cell zones, but strongly co-localized with macrophages and DCs.een) distribution relative to lymph node sinus architecture. Bar=100μm.

30 S.T. Reddy et al. / Journal of Controlled Release 112 (2006) 26–34
nodes, which drain the tail and leg lymphatics, brachial andaxillary lymph nodes, which drain the front footpad arealymphatics were removed, flash frozen, cryosectioned into10-μm sections, and immunostained with antibodies againstmouse CD3e (Pharmingen, San Diego, CA), CD45R (Caltag,Burlingame, CA), CD68 (Serotec, Dusseldorf, Germany), Dec-205 (Serotec), and CD31 (Pharmingen) to label T cells, B cells,macrophages/DCs, DCs, and endothelial cells, respectively.Secondary detection was performed with Alexa Flour 594nm(Molecular Probes) antibodies. Lymph node sections wereimaged with fluorescence (Axiovert 200M, Zeiss) and confocallaser scanning microscopy (LSM 510 Meta, Zeiss).
2.5. Lymph node cell isolation and staining
Lymph node cells were isolated following a previouslydescribed protocol [45]. Briefly, following injections offluorescent nanoparticles or PBS as described earlier, lymphnodes were removed, teased with 26-gauge needles anddigested in Collagenase D (Roche, Basel, Switzerland) for25min at 37°C. Tissue was then passed through a 70-μm cellstrainer (BD, Basel, Switzerland) to recover a cell suspension.With the lymph node cell suspension, APCs were stained forwith anti-MHC Class II-(I-A)-R-PE (Chemicon, Temecula, CA)and DCs with anti-CD11c-allophycocyanin (Pharmingen).
2.6. In vitro nanoparticle internalization
Following lymph node cell isolation, cells were plated inRPMI (5% FBS) at ∼500,000cells/ml. Cells were then pulsed
Fig. 4. Internalization of nanoparticles by macrophages and dendritic cells (DCs). Shonm PPS nanoparticles. (A) Staining for CD68, expressed by both macrophages and D(B) Staining for Dec-205, which is found exclusively on DCs, demonstrates that DC
with 20μl of 20mg/ml of fluorescent nanoparticles andincubated for 24h. Cells were then washed twice with HBSSand stained for APCs and DCs as mentioned earlier.
2.7. Flow cytometry and analysis
Following staining, lymph node cell suspensions wereanalyzed by flow cytometry (CyAn ADP, Dako, Glostrup,Denmark). Further analysis was performed using FlowJosoftware (TreeStar, Ashland, OR). APCs and DCs that hadinternalized fluorescent nanoparticles were determined to beMHCII+FITC+ and CD11c+FITC+, respectively.
2.8. Statistics
Statistical significance was determined by performing a two-tailed Student's t test. Results indicate mean±SD and 3–8 experiments were performed for each condition.
3. Results and discussion
This study investigated the potential for PPS nanoparticles asdrug delivery vehicles to target APCs, specifically DCs, inlymph nodes. Nanoparticle lymphatic uptake, lymph noderetention, and localization within lymph nodes and among cellpopulations there were evaluated. The ability to deliver drugs toDCs in lymph nodes offers a promising approach toimmunotherapy. One challenge is obtaining both efficientlymphatic uptake and lymph node retention, since nanoparticleproperties, such as size and surface characteristics, have
wn are confocal images of lymph node sections at 96 h following injection of 20Cs, reveals internalization of nanoparticles by these cells (nuclei shown in blue).s also internalize the nanoparticles. Bar=20μm.

Fig. 5. Quantification of cell uptake of nanoparticles (NPs). Flow cytometryanalysis was used to determine the fraction of lymph node APCs (MHCII+) andDCs (CD11c+) that internalized NPs (FITC+). (A) At 24h, following injections,more than 38% of APCs and 50% of DCs in lymph nodes had internalized 20nmNPs. There was reduced uptake in both cell populations with 45nm NPs, andonly∼10% of all APCs took up 100nm NPs. (B) After in vitro pulsing of APCsand DCs with NPs for 24h, nearly all APCs and DCs had internalized NPs of all3 sizes. Thus since all 3 NP sizes are equally taken up in vitro, the differencesseen following in vivo injection are most likely due to differences in NP uptakeinto lymphatics following injection. These results also indicate that NP uptakeoccurs in the lymph nodes rather than by cells in peripheral sites, which thenmigrate to lymph nodes.
31S.T. Reddy et al. / Journal of Controlled Release 112 (2006) 26–34
different advantages. In general, smaller particles have betterlymphatic uptake than larger particles but lower lymph noderetention. Additionally, steric stabilization with hydrophilicpolymers such as PEG can improve uptake from the interstitiumby reducing non-specific interactions; however, their increasedstealth nature could also reduce internalization by phagocyticcells in lymph nodes. Therefore, this study aimed to evaluateand optimize PPS nanoparticle properties for lymphatic deliveryand furthermore establish and substantiate the strategy ofdelivery to DCs and other APCs in lymph nodes.
Fluorescence microlymphangiography in the mouse tail wasused to evaluate the lymphatic uptake of PPS nanoparticles of20, 45, and 100nm diameter. Following infusion with 20nmparticles, the hexagonal lymphatic capillary network was clearlyvisible after 90min and uniformly filled from the injection site(Fig. 1A). In contrast, only a very faint lymphatic network wasobserved following injection with the 45nm particles (Fig. 1B),and almost no network could be seen with the 100nm particles(Fig. 1C), indicating poor uptake. The upper limit of size formacromolecule/protein/particle leakage into blood capillaries iswell known to be∼3.5nm [41], therefore the leakage into bloodvasculature of our smallest 20nm particles is effectively zero.This method thus qualitatively confirms that 20 nm particles aremore readily taken up into the lymphatic capillaries from theinterstitial space compared to 45nm and especially 100nmparticles.
Lymph node retention time of nanoparticles and liposomeshas been investigated by several other researchers and typicallyhas focused on time points in the range of 6–52h post-injection[31–34,36]. This study examined lymph node retention ofnanoparticles up to 120h, and showed that 20nm particles weresignificantly present at qualitatively consistent levels in thelymph node at 24, 72, 96, 120h following a bolus injection of20μl intradermally (Fig. 2). The 45nm nanoparticles were alsopresent, although in lower amounts, in lymph nodes at all timepoints, while the 100nm particles were not visibly present in thelymph nodes at any time point (Fig. 2). Thus, together with theresults in Fig. 1, these data show that 20–45nm is a good PPSnanoparticle size range for both lymphatic uptake and lymphnode retention, with 20nm being optimal, while 100nmparticles are too large for efficient lymphatic uptake from theinterstitium following a constant pressure injection. This isconsistent with previous studies that have shown liposomes>70nm remain mostly at the site of injection [24,30].
The specific locations, with respect to the various immunecells, in which PPS nanoparticles accumulated within lymphnodes, were evaluated. Staining results were consistent withknown lymph node architecture, where specific zones for T andB lymphocytes can be readily seen [2]. T cells tend to aggregatein the center regions of the node while B cells are often found ingerminal centers located towards the outer membrane. The othermajor cell types present in lymph nodes are APCs or MHCII+
cells, namely DCs and some macrophages, and their location isdispersed in the subcapsular space. Fig. 3 shows serial sectionsof the same lymph node following an intradermal injection of20nm particles. The nanoparticles were not present in the T cellor B cell zones (Fig. 3A, B). However, there was significant co-
localization of the nanoparticles with macrophages and DCs;i.e., CD68+ cells (Fig. 3C).
It has generally been assumed that liposomes and nanopar-ticles delivered to lymph nodes are primarily phagocytosed bymacrophages there [21,27,29,30,32,36,37]. However, it hasrecently been shown that immature DCs capable of taking upantigens are also present in the lymph nodes [15,16]. With PPSnanoparticle delivery, immunostaining for CD68 (which,although a transmembrane protein, is also expressed intra-cellulary [46–49]) verified that macrophages and DCs hadinternalized PPS nanoparticles (Fig. 4A). To further determinewhether the CD68+ cells were macrophages, DCs, or both,lymph nodes were immunostained for the highly specific DCreceptor Dec-205 [4,50–56]. The Dec-205+ cells and their co-localization with nanoparticles (Fig. 4B) demonstrate that asignificant fraction of the cells in the lymph node thatphagocytose the nanoparticles were, indeed, DCs. This willbe critical for future work that looks to deliver antigensto lymph nodes in order to stimulate the most potent APCtype—DCs.
32 S.T. Reddy et al. / Journal of Controlled Release 112 (2006) 26–34
Flow cytometry analysis was performed to quantify thefraction of APCs and DCs in lymph nodes that wereinternalizing nanoparticles. Fig. 5A shows that up to ∼40%of APCs (MHCII+) and specifically ∼50% of DCs (CD11c+) inlymph nodes have taken up 20nm particles after 24h post-injection. Also a significant fraction of APCs and DCsphagocytose 45nm particles while very little uptake of100nm particles was observed. APCs, including the DCs,could have either endocytosed the particles after they arrived inthe lymph node, or internalized them at the injection site beforetrafficking to lymph nodes. If the latter were the case, 100nmparticles would be seen in the lymph node, since larger particles(1–10μm) can be endocytosed by APCs just as efficiently assmaller ones [13,57]. It was verified that nanoparticle size didnot affect APC or DC internalization by in vitro experiments;
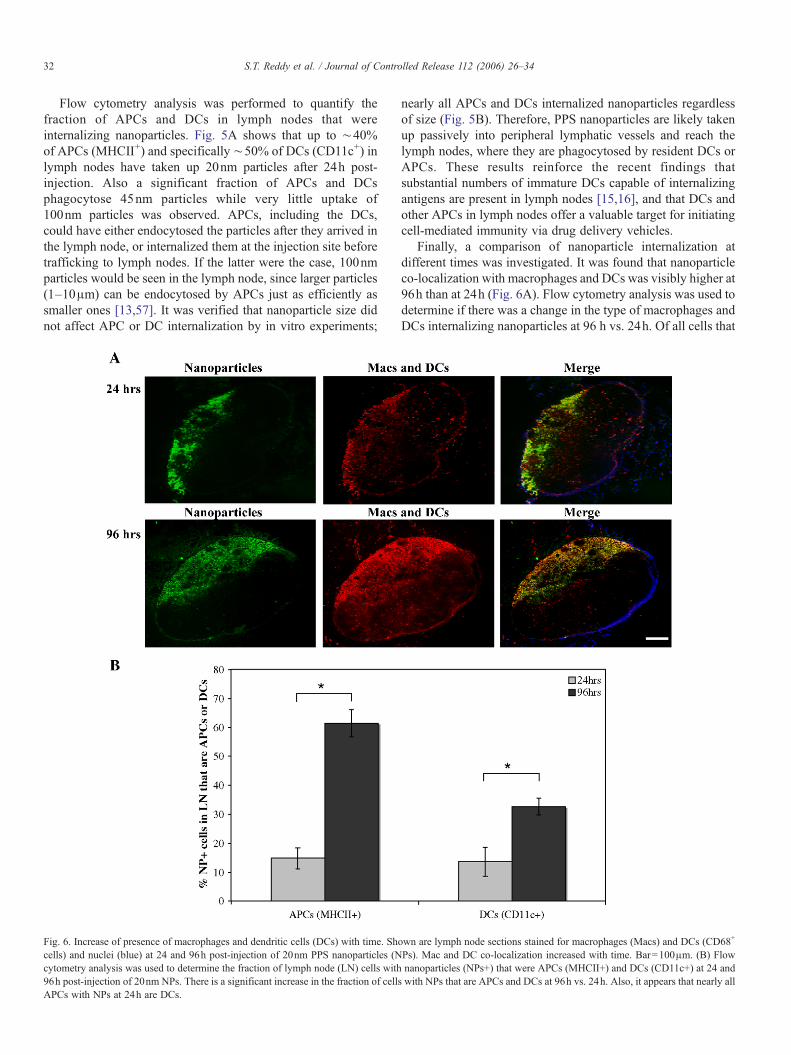
Fig. 6. Increase of presence of macrophages and dendritic cells (DCs) with time. Shcells) and nuclei (blue) at 24 and 96h post-injection of 20nm PPS nanoparticles (Ncytometry analysis was used to determine the fraction of lymph node (LN) cells with96h post-injection of 20nm NPs. There is a significant increase in the fraction of cellAPCs with NPs at 24h are DCs.
nearly all APCs and DCs internalized nanoparticles regardlessof size (Fig. 5B). Therefore, PPS nanoparticles are likely takenup passively into peripheral lymphatic vessels and reach thelymph nodes, where they are phagocytosed by resident DCs orAPCs. These results reinforce the recent findings thatsubstantial numbers of immature DCs capable of internalizingantigens are present in lymph nodes [15,16], and that DCs andother APCs in lymph nodes offer a valuable target for initiatingcell-mediated immunity via drug delivery vehicles.
Finally, a comparison of nanoparticle internalization atdifferent times was investigated. It was found that nanoparticleco-localization with macrophages and DCs was visibly higher at96h than at 24h (Fig. 6A). Flow cytometry analysis was used todetermine if there was a change in the type of macrophages andDCs internalizing nanoparticles at 96 h vs. 24h. Of all cells that
own are lymph node sections stained for macrophages (Macs) and DCs (CD68+
Ps). Mac and DC co-localization increased with time. Bar=100μm. (B) Flownanoparticles (NPs+) that were APCs (MHCII+) and DCs (CD11c+) at 24 and
s with NPs that are APCs and DCs at 96h vs. 24h. Also, it appears that nearly all

33S.T. Reddy et al. / Journal of Controlled Release 112 (2006) 26–34
had internalized nanoparticles at 24h, ∼15% were APCs(MHCII+) and ∼13% were DCs (CD11c+), suggesting thatmost of the APCs were DCs, and that the remaining ∼85% ofnanoparticle containing cells were non-antigen presentingmacrophages (MHCII−). At 96h, the fraction of cells withnanoparticles that were APCs or DCs was 61±5% and 33±3%,respectively (note that this does not reflect an increase in thefraction of lymph node APCs and DCs that contain nanoparti-cles, which remains constant at levels shown in Fig. 5A). Theincrease in nanoparticle-containing MHCII+ and CD11c+ cellsmay be due to an infiltration of APCs and DCs into lymph nodesthat then pick up free nanoparticles still remaining in the nodaltissue. It is also possible that the increase in MHCII+ cells withnanoparticles is due to the activation ofmacrophages between 24and 96h (i.e., MHCII−macrophages that become activated andthus MHCII+ following nanoparticle internalization).
In conclusion, this study demonstrates the strong potentialfor PPS nanoparticles to be used for targeted drug delivery toAPCs, specifically DCs, in lymph nodes. The simplicity ofthis approach is that by controlling size, nanoparticles can beeffectively taken up into lymphatics as well as retained inlymph nodes for at least 5 days, and without using anyspecific targeting ligand; they are internalized exclusively bynodal resident DCs and other APCs. Additionally, it isdemonstrated that up to 40–50% of resident lymph nodeDCs internalize nanoparticles, further supporting the feasi-bility of this delivery strategy. Future studies will focus onthe ability for PPS nanoparticles to induce DC maturation,antigen presentation, T cell proliferation, and cell-mediatedimmunity.
Acknowledgments
The authors are extremely grateful to Veronique Angeli,Gwendalyn Randolph, and Dimitris Missirils for helpful adviceand Miriella Pasquier, Anissa Sidibe, and Veronique Garea forinvaluable technical assistance. This work was funded by theÉcole Polytechnique Fédérale de Lausanne and a grant from theNIH / NCI (P50 CA89018-01).
References
[1] J. Banchereau, R.M. Steinman, Dendritic cells and the control ofimmunity, Nature 392 (6673) (1998) 245–252.
[2] J.G. Cyster, Chemokines—chemokines and cell migration in secondarylymphoid organs, Science 286 (5447) (1999) 2098–2102.
[3] G.J. Randolph, V. Angeli, M.A. Swartz, Dendritic-cell trafficking to lymphnodes through lymphatic vessels, Nat. Rev. Immunol. 5 (2005) 617–628.
[4] L. Bonifaz, D. Bonnyay, K. Mahnke, M. Rivera, M.C. Nussenzweig, R.M.Steinman, Efficient targeting of protein antigen to the dendritic cellreceptor DEC-205 in the steady state leads to antigen presentation on majorhistocompatibility complex class I products and peripheral CD8(+) T celltolerance, J. Exp. Med. 196 (12) (2002) 1627–1638.
[5] L.C. Bonifaz, D.P. Bonnyay, A. Charalambous, D.I. Darguste, S. Fujii, H.Soares, M.K. Brimnes, B. Moltedo, T.M. Moran, R.M. Steinman, In vivotargeting of antigens to maturing dendritic cells via the DEC-205 receptorimproves T cell vaccination, J. Exp. Med. 199 (6) (2004) 815–824.
[6] S. Jilek, H.P. Merckle, E. Walter, DNA-loaded biodegradable micro-particles as vaccine delivery systems and their interaction with dendritecells, Adv. Drug Deliv. Res. 57 (3) (2005) 377–390.
[7] P. Elamanchili, M. Diwan, M. Cao, J. Samuel, Characterization of poly(D,L-lactic-co-glycolic acid) based nanoparticulate system for enhanceddelivery of antigens to dendritic cells, Vaccine 22 (19) (2004) 2406–2412.
[8] S. Faraasen, J. Voros, G. Csucs, M. Textor, H.P. Merkle, E. Walter, Ligand-specific targeting of microspheres to phagocytes by surface modificationwith poly(L-lysine)-grafted poly(ethylene glycol) conjugate, Pharm. Res.20 (2) (2003) 237–246.
[9] Y.J. Kwon, E. James, N. Shastri, J.M. Frechet, In vivo targeting ofdendritic cells for activation of cellular immunity using vaccine carriersbased on pH-responsive microparticles, Proc. Natl. Acad. Sci. U. S. A. 102(51) (2005) 18264–18268.
[10] Y.J. Kwon, S.M. Standley, S.L. Goh, J.M. Frechet, Enhanced antigenpresentation and immunostimulation of dendritic cells using acid-degradable cationic nanoparticles, J. Control. Release 105 (3) (2005)199–212.
[11] Y.J. Kwon, S.M. Standley, A.P. Goodwin, E.R. Gillies, J.M. Frechet,Directed antigen presentation using polymeric microparticulate carriersdegradable at lysosomal pH for controlled immune responses, Mol.Pharmacol. 2 (1) (2005) 83–91.
[12] C.L. van Broekhoven, C.R. Parish, C. Demangel, W.J. Britton, J.G. Altin,Targeting dendritic cells with antigen-containing liposomes: a highlyeffective procedure for induction of antitumor immunity and for tumorimmunotherapy, Cancer Res. 64 (12) (2004) 4357–4365.
[13] C. Wang, Q. Ge, D. Ting, D. Nguyen, H.R. Shen, J.Z. Chen, H.N. Eisen, J.Heller, R. Langer, D. Putnam, Molecularly engineered poly(ortho ester)microspheres for enhanced delivery of DNA vaccines, Nat. Mater 3 (3)(2004) 190–196.
[14] R.F. Wang, H.Y. Wang, Enhancement of antitumor immunity byprolonging antigen presentation on dendritic cells, Nat. Biotechnol. 20(2) (2002) 149–154.
[15] N.S. Wilson, D. El-Sukkari, G.T. Belz, C.M. Smith, R.J. Steptoe, W.R.Heath, K. Shortman, J.A. Villadangos, Most lymphoid organ dendritic celltypes are phenotypically and functionally immature, Blood 102 (6) (2003)2187–2194.
[16] M. Sixt, N. Kanazawa, M. Seig, T. Samson, G. Roos, D.P. Reinhardt, R.Pabst, M.B. Lutz, L. Sorokin, The conduit system transports solubleantigens from the afferent lymph to resident dendritic cells in the Tcell areaof the lymph node, Immunity 22 (1) (2005) 19–29.
[17] D.W. Pack, Timing is everything, Nat. Mater. 3 (3) (2004) 133–134.[18] H.C. Probst, J. Lagnel, G. Kollias, M. van den Broek, Inducible transgenic
mice reveal resting dendritic cells as potent inducers of CD8+ T celltolerance, Immunity 18 (5) (2003) 713–720.
[19] M.J. Copland, M.A. Baird, T. Rades, J.L. McKenzie, B. Becker, F. Reck, P.C. Tyler, N.M. Davies, Liposomal delivery of antigen to human dendriticcells, Vaccine 21 (9–10) (2003) 883–890.
[20] M.A. Swartz, The physiology of the lymphatic system, Adv. Drug Deliv.Rev. 50 (1–2) (2001) 3–20.
[21] C.J.H. Porter, Drug delivery to the lymphatic system, Crit. Rev. Ther. DrugCarr. Syst. 14 (4) (1997) 333–393.
[22] C.J.H. Porter, S.A. Charman, Lymphatic transport of proteins aftersubcutaneous administration, J. Pharm. Sci. 89 (3) (2000) 297–310.
[23] M. Papisov, R. Weissleder, Drug delivery to lymphatic tissue, Crit. Rev.Ther. Drug Carr. Syst. 13 (1–2) (1996) 57–84.
[24] C. Oussoren, J. Zuidema, D.J.A. Crommelin, G. Storm, Lymphatic uptakeand biodistribution of liposomes after subcutaneous injection: 2. Influenceof liposomal size, lipid composition and lipid dose, Biochim. Biophys.Acta, Biomembr. 1328 (2) (1997) 261–272.
[25] A.E. Hawley, S.S. Davis, L. Illum, Targeting of colloids to lymph-nodes—influence of lymphatic physiology and colloidal characteristics, Adv. DrugDeliv. Rev. 17 (1) (1995) 129–148.
[26] T.M. Allen, C.B. Hansen, L.S.S. Guo, Subcutaneous administration ofliposomes—a comparison with the intravenous and intraperitoneal routesof injection, Biochim. Biophys. Acta 1150 (1) (1993) 9–16.
[27] S.M. Moghimi, Modulation of lymphatic distribution of subcutaneouslyinjected poloxamer 407-coated nanospheres: the effect of the ethyleneoxide chain configuration, FEBS Lett. 545 (2–3) (2003) 241–244.
[28] S.M. Moghimi, B. Bonnemain, Subcutaneous and intravenous deliveryof diagnostic agents to the lymphatic system: applications in lymphos-

34 S.T. Reddy et al. / Journal of Controlled Release 112 (2006) 26–34
cintigraphy and indirect lymphography, Adv. Drug Deliv. Rev. 37 (1–3)(1999) 295–312.
[29] Y. Nishioka, H. Yoshino, Lymphatic targeting with nanoparticulate system,Adv. Drug Deliv. Rev. 47 (1) (2001) 55–64.
[30] C. Oussoren, G. Storm, Liposomes to target the lymphatics bysubcutaneous administration, Adv. Drug Deliv. Rev. 50 (2001) 143–156.
[31] C. Oussoren, G. Storm, Lymphatic uptake and biodistribution of liposomesafter subcutaneous injection: 3. Influence of surface modification with poly(ethyleneglycol), Pharm. Res. 14 (10) (1997) 1479–1484.
[32] S.M. Moghimi, A.E. Hawley, N.M. Christy, T. Gray, L. Illum, S.S. Davis,Surface engineered nanospheres with enhanced drainage into lymphaticsand uptake by macrophages of the regional lymph-nodes, FEBS Lett. 344(1) (1994) 25–30.
[33] A.E. Hawley, L. Illum, S.S. Davis, Preparation of biodegradable, surfaceengineered PLGA nanospheres with enhanced lymphatic drainage andlymph node uptake, Pharm. Res. 14 (5) (1997) 657–661.
[34] A.E. Hawley, L. Illum, S.S. Davis, Lymph node localisation ofbiodegradable nanospheres surface modified with poloxamer andpoloxamine block co-polymers, FEBS Lett. 400 (3) (1997) 319–323.
[35] L. Illum, A.E. Church, M.D. Butterworth, A. Arien, J. Whetstone, S.S.Davis, Development of systems for targeting the regional lymph nodes fordiagnostic imaging: in vivo behaviour of colloidal PEG-coated magnetitenanospheres in the rat following interstitial administration, Pharm. Res. 18(5) (2001) 640–645.
[36] C. Oussoren, M. Velinova, G. Scherphof, J.J. van der Want, N. vanRooijen, G. Storm, Lymphatic uptake and biodistribution of liposomesafter subcutaneous injection: IV. Fate of liposomes in regional lymphnodes, Biochim. Biophys. Acta, Biomembr. 1370 (2) (1998) 259–272.
[37] C. Oussoren, G. Storm, Role of macrophages in the localisation ofliposomes in lymph nodes after subcutaneous administration, Int. J. Pharm.183 (1) (1999) 37–41.
[38] A. Rehor, J.A. Hubbell, N. Tirelli, Oxidation-sensitive polymericnanoparticles, Langmuir 21 (1) (2005) 411–417.
[39] A. Rehor, N. Tirelli, J.A. Hubbell, A new living emulsion polymerizationmechanism: episulfide anionic polymerization, Macromolecules 35 (23)(2002) 8688–8693.
[40] A. Napoli, M. Valentini, N. Tirelli, M. Muller, J.A. Hubbell, Oxidation-responsive polymeric vesicles, Nat. Mater. 3 (3) (2004) 183–189.
[41] W.L. Olszewski, Lymph Stasis Pathophysiology, Diagnosis and Treatment,CRC Press, Boca Raton, 1991.
[42] D.A. Berk, M.A. Swartz, A.J. Leu, R.K. Jain, Transport in lymphaticcapillaries: 2. Microscopic velocity measurement with fluorescencephotobleaching, Am. J. Physiol.: Heart Circ. Physiol. 39 (1) (1996)H330–H337.
[43] M.A. Swartz, D.A. Berk, R.K. Jain, Transport in lymphatic capillaries: 1.Macroscopic measurements using residence time distribution theory, Am.J. Physiol.: Heart Circ. Physiol. 39 (1) (1996) H324–H329.
[44] M.A. Swartz, A. Kaipainen, P.A. Netti, C. Brekken, Y. Boucher, A.J.Grodzinsky, R.K. Jain, Mechanics of interstitial-lymphatic fluid transport:
theoretical foundation and experimental validation, J. Biomech. 32 (12)(1999) 1297–1307.
[45] V. Angeli, J. Llodra, J.X. Rong, K. Satoh, S. Ishii, T. Shimizu, E.A. Fisher,G.J. Randolph, Dyslipidemia associated with atherosclerotic diseasesystemically alters dendritic cell mobilization, Immunity 21 (4) (2004)561–574.
[46] R.P. da Silva, S. Gordon, Phagocytosis stimulates alternative glycosylationof macrosialin (mouse CD68), a macrophage-specific endosomal protein,Biochem. J. 338 (Pt 3) (1999) 687–694.
[47] C.L. Holness, R.P. da Silva, J. Fawcett, S. Gordon, D.L. Simmons,Macrosialin, a mouse macrophage-restricted glycoprotein, is a member ofthe lamp/lgp family, J. Biol. Chem. 268 (13) (1993) 9661–9666.
[48] S. Rabinowitz, H. Horstmann, S. Gordon, G. Griffiths, Immunocytochem-ical characterization of the endocytic and phagolysosomal compartmentsin peritoneal macrophages, J. Cell Biol. 116 (1) (1992) 95–112.
[49] S.S. Rabinowitz, S. Gordon, Macrosialin, a macrophage-restrictedmembrane sialoprotein differentially glycosylated in response to inflam-matory stimuli, J. Exp. Med. 174 (4) (1991) 827–836.
[50] M. Breel, R.E. Mebius, G. Kraal, Dendritic cells of the mouse recognizedby 2 monoclonal-antibodies, Eur. J. Immunol. 17 (11) (1987) 1555–1559.
[51] D. Hawiger, K. Inaba, Y. Dorsett, M. Guo, K. Mahnke, M. Rivera, J.V.Ravetch, R.M. Steinman, M.C. Nussenzweig, Dendritic cells induceperipheral T cell unresponsiveness under steady state conditions in vivo,J. Exp. Med. 194 (6) (2001) 769–779.
[52] K. Inaba, W.J. Swiggard, M. Inaba, J. Meltzer, A. Mirza, T. Sasagawa, M.C. Nussenzweig, R.M. Steinman, Tissue distribution of the Dec-205protein that is detected by the monoclonal-antibody Nldc-145: 1.Expression on dendritic cells and other subsets of mouse leukocytes,Cell. Immunol. 163 (1) (1995) 148–156.
[53] W.P. Jiang, W.J. Swiggard, C. Heufler, M. Peng, A. Mirza, R.M. Steinman,M.C. Nussenzweig, The Receptor Dec-205 expressed by dendritic cellsand thymic epithelial-cells is involved in antigen-processing, Nature 375(6527) (1995) 151–155.
[54] W.P. Jiang, W.J. Swiggard, A. Mirza, M. Peng, R.M. Steinman, M.C.Nussenzweig, Molecular characterization of a 205-kD protein that isabundant on dendritic cells and identified with the Nldc-145 monoclonal-antibody, J. Cell. Biochem. (1995) 20.
[55] G. Kraal, M. Breel, M. Janse, G. Bruin, Langerhans cells, veiled cells, andinterdigitating cells in the mouse recognized by a monoclonal-antibody,J. Exp. Med. 163 (4) (1986) 981–997.
[56] K. Mahnke, M. Guo, S. Lee, H. Sepulveda, S.L. Swain, M. Nussenzweig,R.M. Steinman, The dendritic cell receptor for endocytosis, DEC-205, canrecycle and enhance antigen presentation via major histocompatibilitycomplex class II-positive lysosomal compartments, J. Cell Biol. 151 (3)(2000) 673–683.
[57] Y. Tabata, Y. Ikada, Phagocytosis of polymer microspheres by macro-phages, Adv. Polym. Sci. 94 (1990) 107–141.




















