
Effects of Dietary Cholesterol on Antibody Dependent Phagocytosis and Cell-Mediated Lysis in Guinea...
9
Effects of Dietary Cholesterol on Antibody- Dependent Phagocytosis and Cell-Mediated Lysis in Guinea Pigs1 AXEL K. DUWE,2-3 MARK FITCH AND ROSEMARIE OSTWALD Department of Nutritional Sciences, University of California, Berkeley, CA 94720 ABSTRACT The effect of dietary cholesterol on antibody-dependent phago cytosis and cell-mediated cytotoxicity (ADCC) by peritoneal cells and on the susceptibility to lysis of erythrocytes was studied in the guinea pig. We found that peritoneal cells from cholesterol-fed animals (CHOL PEC) demonstrated a decreased ability to both phagocytose and lyse antibody-coated (Ab) guinea pig erythrocytes than did those from control guinea pigs (CONT PEC). This decrease was equal in groups fed cholesterol for SVz-13 weeks, preanemic or anemic, and with normal or enlarged spleens. Dose response curves varying Ab concentration showed that CHOJ-, PEC required higher concentrations of Ab to effect phagocytosis and lysis than did CONT PEC. Dietary cholesterol, while rapidly inducing morphological changes such as spurring in guinea pig eryth rocytes, was found not to affect the susceptibility of the cells to lysis or phago cytosis in this assay system. These findings suggest that the increased incidence of infection in cholesterol-fed guinea pigs may be due to impaired phagocytic function and that the anemia observed in guinea pigs after 8-10 weeks of feeding cholesterol is not due to increased antibody-dependent removal of spurred erythrocytes by the phagocytic system. J. Nutr. Ill; 1672-1680, 1981. INDEXING KEY WORDS cholesterol phagocytosis lysis •erythrocytes Addition of 1% cholesterol to the diet of lipids, such as fatty acid esters, de- of guinea pigs results in an increase of presses activity of the reticuloendothelial cholesterol levels in both plasma and cell system (RES) and phagocytic function in membranes (1). This is followed by the vivo (4, 5). In addition, rendering animals development of a severe hemolytic ane- hypercholesterolemic by feeding choles- mia 8-10 weeks later (2). We have also terol increases the cholesterol content of observed that the animals fed cholesterol both plasma and cell membranes in the are more susceptible to infections than rat (6), rabbit (7) and guinea pig (1) and control animals—to such a degree that suppresses the non-specific phagocytosis experimental groups being fed choies- of carbon particles in vivo in the rabbit terol must be made larger than control (7) and the in vitro ingestion of latex par- groups due to attrition (unpublished data), tides and oil droplets by rat macrophages Phagocytic cells of the reticuloendoth elial system are the first line of defense against infection (3), and modulation Of Received for publication 2 February 1981. the lipid phase of phagocyte membranes 'This work wasupported byUSPHS. NIHgrant AMMSO. by in vitro or in vivo methods has been H^äJÃoeuK^^* shown to affect phagocytosis. A number »supported byNIH training grant *082AMO729u 1672 by guest on March 21, 2014 jn.nutrition.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Effects of Dietary Cholesterol on Antibody Dependent Phagocytosis and Cell-Mediated Lysis in Guinea...

Effects of Dietary Cholesterol on Antibody-Dependent Phagocytosis and Cell-Mediated Lysis inGuinea Pigs1
AXEL K. DUWE,2-3 MARK FITCH AND ROSEMARIE
OSTWALD
Department of Nutritional Sciences,University of California, Berkeley, CA 94720
ABSTRACT The effect of dietary cholesterol on antibody-dependent phagocytosis and cell-mediated cytotoxicity (ADCC) by peritoneal cells and on thesusceptibility to lysis of erythrocytes was studied in the guinea pig. We foundthat peritoneal cells from cholesterol-fed animals (CHOL PEC) demonstrateda decreased ability to both phagocytose and lyse antibody-coated (Ab) guineapig erythrocytes than did those from control guinea pigs (CONT PEC). Thisdecrease was equal in groups fed cholesterol for SVz-13 weeks, preanemic oranemic, and with normal or enlarged spleens. Dose response curves varying Abconcentration showed that CHOJ-, PEC required higher concentrations of Ab toeffect phagocytosis and lysis than did CONT PEC. Dietary cholesterol, whilerapidly inducing morphological changes such as spurring in guinea pig erythrocytes, was found not to affect the susceptibility of the cells to lysis or phagocytosis in this assay system. These findings suggest that the increased incidenceof infection in cholesterol-fed guinea pigs may be due to impaired phagocyticfunction and that the anemia observed in guinea pigs after 8-10 weeks of feedingcholesterol is not due to increased antibody-dependent removal of spurrederythrocytes by the phagocytic system. J. Nutr. Ill; 1672-1680, 1981.
INDEXING KEY WORDS cholesterol phagocytosis lysis•erythrocytes
Addition of 1% cholesterol to the diet of lipids, such as fatty acid esters, de-of guinea pigs results in an increase of presses activity of the reticuloendothelialcholesterol levels in both plasma and cell system (RES) and phagocytic function inmembranes (1). This is followed by the vivo (4, 5). In addition, rendering animalsdevelopment of a severe hemolytic ane- hypercholesterolemic by feeding choles-mia 8-10 weeks later (2). We have also terol increases the cholesterol content ofobserved that the animals fed cholesterol both plasma and cell membranes in theare more susceptible to infections than rat (6), rabbit (7) and guinea pig (1) andcontrol animals—to such a degree that suppresses the non-specific phagocytosisexperimental groups being fed choies- of carbon particles in vivo in the rabbitterol must be made larger than control (7) and the in vitro ingestion of latex par-groups due to attrition (unpublished data), tides and oil droplets by rat macrophages
Phagocytic cells of the reticuloendothelial system are the first line of defenseagainst infection (3), and modulation Of Received for publication 2 February 1981.the lipid phase of phagocyte membranes 'ThisworkwassupportedbyUSPHS.NIHgrantAMMSO.by in vitro or in vivo methods has been H^äJÜuK^^*shown to affect phagocytosis. A number »supportedbyNIHtraininggrant*082AMO729u
1672
by guest on March 21, 2014
jn.nutrition.orgD
ownloaded from

CHOLESTEROL EFFECTS ON PHAGOCYTOSIS AND LYSIS 1673
(6). Engulfment of antibody (Ab)-coatedparticles has been shown to be dependent upon multiple interactions betweenthe Fc (fragment, crystallizable) portionof particle-bound Ab molecules and receptors for the Fc portion of the phagocyte (8). Thus it would be expected thatnot only the rate of phagocytosis but alsoa receptor-dependent ingestion dependson the physico-chemical properties of themembrane lipid phase.
The anemia in cholesterol-fed guineapigs develops 8-10 weeks after the increase of red blood cell (rbc) membranecholesterol and the appearance of a severe echinocytosis (2). Factors such asparasitic infections, vitamins C and E,folie acid and protein déficiencescan beexcluded as important causes of the anemia (9). Increased levels of membranecholesterol have been reported to decrease complement mediated lysis oferythrocytes, liposomes and tumor cells(10, 11), but do not seem to affect lysis ofmycoplasma (12).
In this study we have compared theability of phagocytes from hypercholes-terolemic and normal guinea pigs to ingest erythrocytes coated with specific Aband the susceptibility to lysis or phagocytosis of red blood cells from animals orcells from animals fed cholesterol(CHOL) and from animals or cells fromanimals fed the control diet (CONT) inan Ab-dependent assay.
MATERIALS AND METHODS
Animals and diets. Male Hartley guineapigs (Simonsen Laboratories Inc., Gilroy,CA) weighing 200 g were fed a commercial guinea pig stock diet (Ralston PurinaCo., St. Louis, MO) supplemented with5% cottonseed oil (CONT). After 3-5days the experimental animals wereswitched to the same diet supplementedwith 1% recrystallized cholesterol(CHOL). Animals were housed individually in laminar-flow cage rack units in atemperature-controlled room, weighedweekly and fed ad libitum. Blood wasobtained from an ear vein or by closedheart puncture in the presence of EDTAfrom animals fasted 16-18 hours.
Peritoneal exúdate cells (PEC). Guineapigs were injected intraperitoneally with20 ml sterile 5% sodium caseinate in saline. Approximately 18 hours later, animals were bled by cardiac puncture andkilled by anesthesia. A large patch of skinwas removed from the abdomen and 50ml saline poured into the peritoneal cavity through a small slit. The abdomen wasthen lightly massaged and the saline containing the peritoneal exúdate pouredinto a beaker. Cells were washed 3 timesin saline and tissue culture medium(RPMI 1640, Grand Island BiologicalCo., Grand Island, NY). This procedureresulted in a peritoneal exúdateconsisting of 70-80% granulocytes, as shown bycounting slides treated with Wright's stain
as has been reported by others (13).Target cells. Erythrocytes were la
beled with 51Crby adding 20 /¿Iof packedguinea pig erythrocytes (GPrbc) to 10 /¿Ifecal calf serum (FCS), SOptlmedium and30 ¿tlNa2 51CrO4(1 mCi/ml in physiological saline, Amersham Searle Corp., Arlington Heights, IL). This mixture wasincubated for 45 minutes at 37°in 10%CO2-90% air and then cells were washed3 times in saline.
Phagocytosis and antibody-dependentcell-mediated cytotoxicity (ADCC) assays. These assays are a modification ofthe method of Walker and Demus (14).Briefly, 5 x IO551Cr-labeled GPrbc andthe appropriate number of peritoneal exúdatecells (PEC) with or without rabbitanti-GPrbc serum (Cappell, Cochran-ville, PA) were suspended in 200 /u.1tissue culture medium. This suspensionwas added to 12 x 75 mm plastic tubes(No. 2054, Falcon Plastics Co., Oxnard,CA) and incubated at 37°in an atmosphere containing 10% Co2-90% air for3 hours unless indicated otherwise. Toeach culture was then added 1.8 ml saline, the tubes were centrifuged at 500 xg for 10 minutes, and 1 ml supernatantwas removed for the determination of radioactivity (A). The remaining 1 ml wasaspirated, the cell pellet was resus-pended in 2 ml water to lyse extracellularrbc and centrifuged at 500 x g for 10 minutes. The supernatant was aspirated and
by guest on March 21, 2014
jn.nutrition.orgD
ownloaded from

1674 DUWE, FITCH AND OSTWALD
the pellet counted (B). Total radioactivitywas measured in a measured aliquot ofrbc (C). Each assay was performed in triplicate and experiments were repeated 3or more times. Phagocytosis was calculated according to the formula:
% phagocytosis
cpm in water—lysis pellet (B)
total cpm (C)
ADCC was calculated according to theformula:
cpm in supernatant of culture -I-Ab (A)- cpm in supernatant of culture - Ab
= ; —total cpm (C)
Statistics. A two-tailed Student's f-test
was used to determine the significance ofdifferences between experimental groups.Differences with P > 0.05 were not considered significant.
RESULTS
Incubation of guinea pig PEC with au-tologous erythrocytes plus rabbit antiGPrbc serum resulted in both erythro-phagocytosis and extracellular lysis(ADCC) of rbc. Figures 1A and 1C showthat the degree of lysis and phagocytosisincreased as the ratio of PEC to erythrocytes, i.e. effector to target cell (E:T) ratio, was increased. Higher antiserum concentrations also mediated higher levelsof lysis and phagocytosis (figs. IB, ID).Little or no measurable activity by PECwas detected in the absence of anti GPrbcserum (figs. 1A, 1C), and incubation ofrbc plus antiserum or control serum without PEC gave only background 5lCr re
lease values. Identical results were obtained when PEC and rbc came from thesame or different animals.
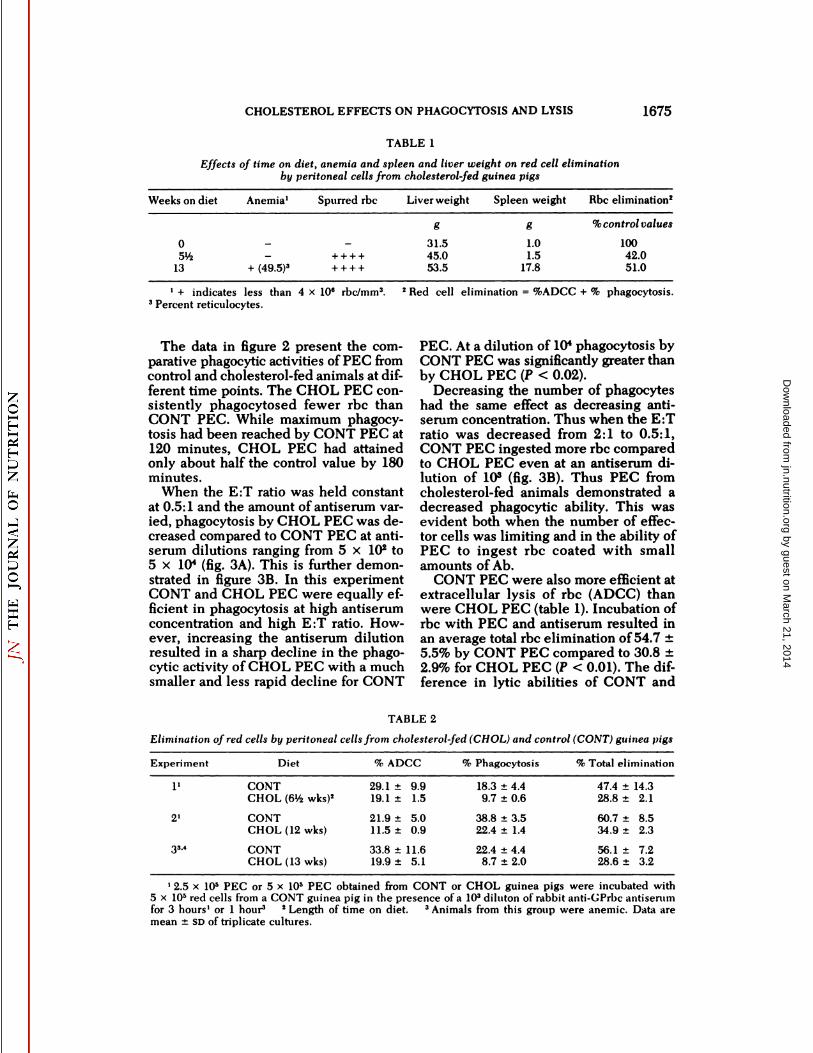
The ability of PEC from cholesterol-fed guinea pigs to eliminate rbc was significantly suppressed when compared tocontrol animals (rbc elimination is thesum of ADCC and phagocytosis). Presence or absence of anemia, increasedspleen weight or time on cholesterol diet(after 5Vi weeks) had no effect on the degree of rbc elimination (table 1). This isfurther demonstrated in table 2. BothADCC and phagocytosis were depressedin the CHOL groups (P < 0.01). In addition, the degree of rbc elimination wasapproximately equal in groups fed cholesterol for 6Yzweeks (experiment 1) and
12 weeks (experiment 2) but showing noanemia, and for animals fed cholesterolfor 13 weeks which were anemic (experiment 3). All CHOL guinea pigs werehypercholesterolemic as routinely assessed by rbc morphology. Our experience in several hundred guinea pigs hasshown that: a) If there are spurred rbc,they are also hypercholesterolemic; andb) this is invariably present by about 3weeks after initiation of cholesterol feed-ing (15).
50-
O.ZS'I 0.5.1 10" 4110* SlIO*
E'T ANTISERUM DILUTION
Fig. 1 ADCC and phagocytic activity of PECfrom control animals. 5 x IO5CONT GPrbc targetswere incubated with a 10s dilution of rabbit anti-GPrbc antiserum and varying numbers of PEC (Aand C) or with 5 x IO5 PEC and varying dilutionsof antiserum (B and D) and assayed for ADCC (Aand B) or phagocytosis (C and D). Data representthe means ±SD of triplicate cultures in a representative experiment.
by guest on March 21, 2014
jn.nutrition.orgD
ownloaded from
CHOLESTEROL EFFECTS ON PHAGOCYTOSIS AND LYSIS 1675
TABLE 1
Effects of time on diet, anemia and spleen and liver weight on red cell eliminationby peritoneal cells from cholesterol-fed guinea pigs
Weeks on diet Anemia1 Spurred rbc Liver weight Spleen weight Rbc elimination2
0S'A
13 + (49.5)"
g
31.545.053.5
1.01.5
17.8
% control values
10042.051.0
1 + indicates less than 4 x 10" rbc/mm3. 2Red cell elimination = %ADCC + % phagocytosis.3Percent reticulocytes.
The data in figure 2 present the comparative phagocytic activities of PEC fromcontrol and cholesterol-fed animals at different time points. The CHOL PEC consistently phagocytosed fewer rbc thanCONT PEC. While maximum phagocytosis had been reached by CONT PEC at120 minutes, CHOL PEC had attainedonly about half the control value by 180minutes.
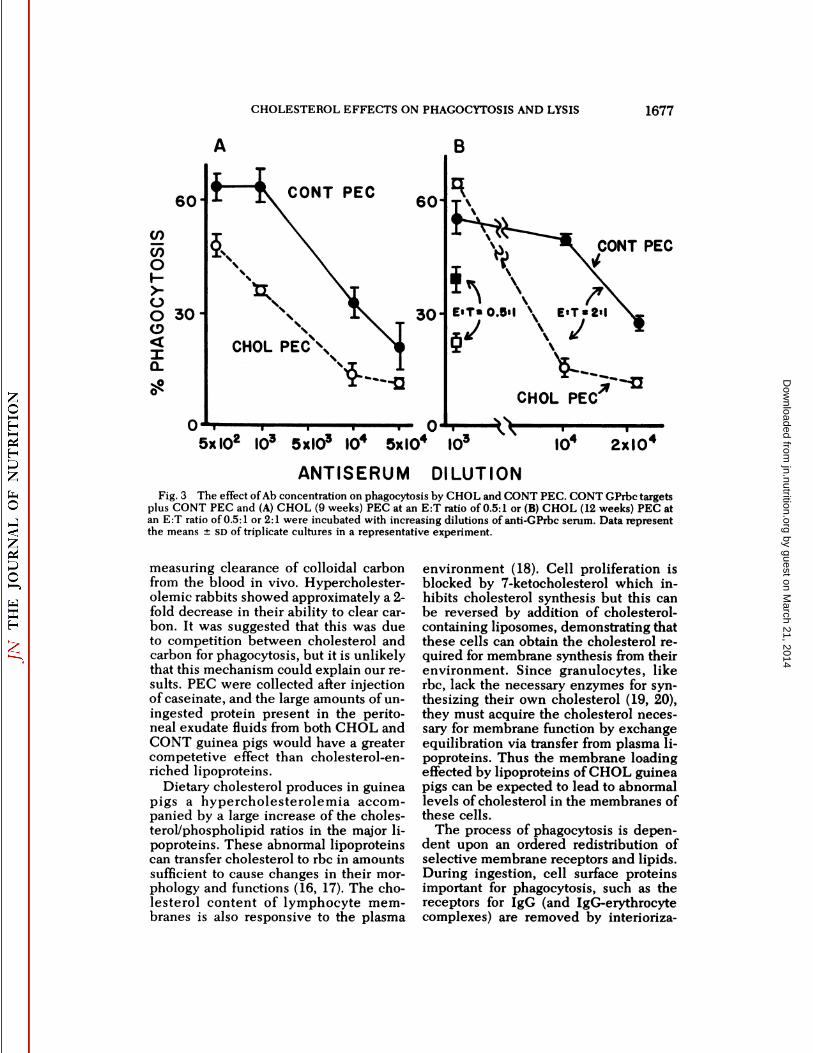
When the E:T ratio was held constantat 0.5:1 and the amount of antiserum varied, phagocytosis by CHOL PEC was decreased compared to CONT PEC at anti-serum dilutions ranging from 5 x IO2to5 x IO4(fig. 3A). This is further demonstrated in figure 3B. In this experimentCONT and CHOL PEC were equally efficient in phagocytosis at high antiserumconcentration and high E:T ratio. However, increasing the antiserum dilutionresulted in a sharp decline in the phagocytic activity of CHOL PEC with a muchsmaller and less rapid decline for CONT
PEC. At a dilution of IO4phagocytosis byCONT PEC was significantly greater thanby CHOL PEC (P < 0.02).
Decreasing the number of phagocyteshad the same effect as decreasing anti-serum concentration. Thus when the E:Tratio was decreased from 2:1 to 0.5:1,CONT PEC ingested more rbc comparedto CHOL PEC even at an antiserum dilution of IO3 (fig. 3B). Thus PEC fromcholesterol-fed animals demonstrated adecreased phagocytic ability. This wasevident both when the number of effector cells was limiting and in the ability ofPEC to ingest rbc coated with smallamounts of Ab.
CONT PEC were also more efficient atextracellular lysis of rbc (ADCC) thanwere CHOL PEC (table 1). Incubation ofrbc with PEC and antiserum resulted inan average total rbc elimination of 54.7 ±5.5% by CONT PEC compared to 30.8 ±2.9% for CHOL PEC (P < 0.01). The difference in lytic abilities of CONT and
TABLE 2
Elimination of red cells by peritoneal cells from cholesterol-fed (CHOL) and control (CONT) guinea pigs
Experiment Diet ADCC Phagocytosis Total elimination
I12'3"CONTCHOL(6%wks)2CONTCHOL
(12wks)CONTCHOL
(13wks)29.1
9.919.121.911.533.81.55.00.911.619.9
5.118.3
±4.49.7±0.638.8
±3.522.4±1.422.4
±4.48.7±2.047.4
±14.328.8±2.160.7
±8.534.9±2.356.1
±7.228.6±3.2
12.5 x 10s PEC or 5 x IO5 PEC obtained from CONT or CHOL guinea pigs were incubated with5 x IO5red cells from a CONT guinea pig in the presence of a 103 diluton ot rabbit anti-GPrbc antiserumfor 3 hours' or 1 hour3 2Length of time on diet. 3Animals from this group were anemic. Data aremean ±SD of triplicate cultures.
by guest on March 21, 2014
jn.nutrition.orgD
ownloaded from

1676 DUWE, FITCH AND OSTWALD
60toOTOH
UOo< 30
o.
CHOL PEC
15 60 120 180
MINUTES
Fig. 2 The kinetics of phagocytosis by CHOLand CONT PEC. CONT GPrbc and CHOL (5%weeks) or CONT PEC at an E:T ratio of 1:1 plus aIO3 dilution of antiserum were incubated for theindicated time intervals. Data represent the means±so of triplicate cultures in a representative experiment.
CHOL PEC was not always readily apparent, due in part to the difficulty of obtaining accurate ADCC measurements athigh E:T ratios in this system. Phagocytosis is more rapid than ADCC (data notshown) and therefore at E:T ratios greaterthan 1:1, the majority of rbc attacked byPEC is removed early by ingestion. Incontrast to phagocytosis, significant differences in levels of ADCC betweenCONT and CHOL PEC were observedat low antiserum concentration becauseADCC was generally detectable at lowerconcentrations of antiserum than phagocytosis (data not shown).
Figure 4 demonstrates that CHOL PECrequire higher antiserum concentrationsthan CONT PEC to effect ADCC. Lysisby CHOL PEC dropped sharply as anti-serum concentration was decreased. Lysisby CONT PEC, however, did not fall offuntil antiserum dilutions greater than IO4were reached. Thus at a dilution of IO4,
ADCC by CHOL PEC was 1.9% compared to 32.6% for CONT PEC.
Erythrocytes from guinea pigs fed adiet supplemented with cholesterol con
tain double the amount of cholesterolthan rbc from control animals (1). Wetherefore compared CONT rbc andCHOL rbc for their susceptibility to Ab-dependent phagocytosis and lysis byADCC. CHOL rbc were obtained fromnon-anemic guinea pigs that had beenfed the cholesterol containing diet for atleast 4 weeks. Spurring of rbc is associated with an increase in cholesterol content (2) and is a convenient morphological test for confirming that rbc arecholesterol loaded.
As shown in table 3, there was no difference in susceptibility of CONT andCHOL rbc to phagocytosis by CONTPEC in the presence or the absence ofAb. Similarly, no difference was observed between CONT and CHOL rbc insusceptibility to lysis regardless of thepresence or absence of Ab. In addition,neither the degree of spontaneous lysisin the presence of FCS or anti GPrbc Abnor the amount of Ab bound by rbc asassessed by a hemagglutination test differed for CONT and CHOL rbc (data notshown). Time on cholesterol diet also didnot affect susceptibility of rbc to lysis.
These results indicate that in this system utilizing autologous PEC and rbc,cell cholesterol content does not affectthe susceptibility of rbc to antibody-dependent phagocytosis or lysis, while itdecreases the ability of PEC to effectthese parameters.
DISCUSSION
This study has shown that supplementing the diet of guinea pigs with 1%cholesterol produces a deficiency inphagocytosis and ADCC by granulocytes,which constitute 70-80% of casein-induced PEC in guinea pigs (réf.13 andthis study). These cells play an importantrole in host defense against bacterial infection, and cholesterol-fed guinea pigsare in fact more susceptible to infectionsthat control animals (unpublished observations). Deficiency of phagocytosis bycells obtained from hypercholestero-lemic guinea pigs was demonstrated inan Ab-dependent assay system. Similarresults were obtained by Neuve et al. (7)
by guest on March 21, 2014
jn.nutrition.orgD
ownloaded from
CHOLESTEROL EFFECTS ON PHAGOCYTOSIS AND LYSIS 1677
601
OI->üO 301O<ICL
CONT PEC 60-^
30 •>
CHOL PEC \
CONT PEC
B
Rp«v,Õ\
^ \ rEiTsO.5'1 \ EtT»2'l
^ VSr-*
CHOL PEC
5xl02 IO3 5xl03 IO4 5xl04 IO3
ANTISERUM DILUTION
IO 2x10
Fig. 3 The effect of Ab concentration on phagocytosis by CHOL and CONT PEC. CONT GPrbc targetsplus CONT PEC and (A) CHOL (9 weeks) PEC at an E:T ratio of 0.5:1 or (B) CHOL (12 weeks) PEC atan E:T ratio of 0.5:1 or 2:1 were incubated with increasing dilutions of anti-GPrbc serum. Data representthe means ±so of triplicate cultures in a representative experiment.
measuring clearance of colloidal carbonfrom the blood in vivo. Hypercholester-olemic rabbits showed approximately a 2-fold decrease in their ability to clear carbon. It was suggested that this was dueto competition between cholesterol andcarbon for phagocytosis, but it is unlikelythat this mechanism could explain our results. PEC were collected after injectionof caseinate, and the large amounts of un-ingested protein present in the peritoneal exúdate fluids from both CHOL andCONT guinea pigs would have a greatercompetetive effect than cholesterol-enriched lipoproteins.
Dietary cholesterol produces in guineapigs a hypercholesterolemia accompanied by a large increase of the choles-terol/phospholipid ratios in the major lipoproteins. These abnormal lipoproteinscan transfer cholesterol to rbc in amountssufficient to cause changes in their morphology and functions (16, 17). The cholesterol content of lymphocyte membranes is also responsive to the plasma
environment (18). Cell proliferation isblocked by 7-ketocholesterol which inhibits cholesterol synthesis but this canbe reversed by addition of cholesterol-containing liposomes, demonstrating thatthese cells can obtain the cholesterol required for membrane synthesis from theirenvironment. Since granulocytes, likerbc, lack the necessary enzymes for synthesizing their own cholesterol (19, 20),they must acquire the cholesterol necessary for membrane function by exchangeequilibration via transfer from plasma lipoproteins. Thus the membrane loadingeffected by lipoproteins of CHOL guineapigs can be expected to lead to abnormallevels of cholesterol in the membranes ofthese cells.
The process of phagocytosis is dependent upon an ordered redistribution ofselective membrane receptors and lipids.During ingestion, cell surface proteinsimportant for phagocytosis, such as thereceptors for IgG (and IgG-erythrocytecomplexes) are removed by interioriza-
by guest on March 21, 2014
jn.nutrition.orgD
ownloaded from

1678 DUWE, FITCH AND OSTWALD
40
Oo
20
CONT PEC
CHOL PEC\
5xl02 IO3 5xl03 IO4 5xl04
ANTISERUM DILUTIONFig. 4 ADCC by CHOL and CONT PEC. CONT
GPrbc targets plus CHOL (6% weeks) or CONTPEC were incubated with increasing dilutions ofanti GPrbc serum at an E:T ratio of 0.5:1. Data represent the means ±SD of triplicate cultures in arepresentative experiment.
tion while transport carriers for aminoacids are excluded from phagocytic areasof the membrane (21, 22). In addition,phagocytic vesicles have a higher choles-terol-to-phospholipid ratio than wholecells (23). Thus perturbation of normalcholesterol content of (and distributionin) the plasma membrane results in abnormal receptor movement (24) whichwould lead to less efficient phagocytosis.This suggestion is supported by our observation that the decrease of phagocyticactivity by CHOL PEC was greatest whenthe number of IgG-rbc complexes available for binding to their receptors waslimited. Lowering the Ab concentrationand therefore the number of IgG molecules bound to rbc decreased phagocytosis and this was much more apparent incells from CHOL animals. This was alsotrue for ADCC.
It is possible that the decrease inphagocytosis may not be due to intrinsicdeficiencies in the CHOL cells but to inhibitory factors other than cholesterolpresent in the serum. A number of lipo-
protein factors affects various lymphoidcell functions. A subpopulation of humanlow density lipoprotein (LDL) has beencharacterized, which inhibits both lectinand allogeneic cell-induced lymphocyteproliferation (25). Another specific LDLpresent in some hepatitis B sera blockssheep erythrocyte rosette formation by Tlymphocytes (26). A third LDL present inmouse tumor ascites fluid reduces ADCCto tumor cell targets (27). Furthermore,lipids have been reported to affect phagocytosis. Cholesterol oleate and methylpalmitate depress phagocytic function invivo (5, 28). Thus serum constituentspresent in CHOL animals but not inCONT guinea pigs may be responsiblefor the decrease in phagocytosis andADCC. This would necessarily have tobe a relatively permanent inhibition sincecells from CHOL guinea pigs show a decrease in phagocytosis even after repeated washing and overnight storage at4°(data not shown).
The deficient phagocytosis shown byhypercholesterolemic animals in vivo (7)and in vitro (this study) would result indecreased clearance of bacteria from theblood and lead to a higher incidence ofdisease. This could explain the greatlyincreased susceptibility of our cholesterol-fed guinea pigs to infection, dictating that special precautions be takenwhen feeding animals this type of diet.More importantly, an abnormally highcholesterol level of lymphocytes couldimpair host defenses to infection in man,particularly in populations such as the
TABLE 3
ADCC and phagocytosis of red cells fromcholesterol-fed (CHOL) and control
(CONT) guinea pigs'
Expt. Diet ADCC % Phagocytosis
No AbCONTCHOL1.1
± 2.0(10)1.3 ± 3.5(10)11.9
± 5.9(12)13.5 ± 6.6(13)
Plus Ab CONT 22.3 ± 7.5(9) 48.5 ±16.1 (9)CHOL 20.0 ±12.5 (9) 43.5 ± 3.8 (9)
15 x 10»CONT PEC were incubated for 3 hours at 37°with 5 x 10s CONT or CHOL rbc plus or minus a 10sdilution of rabbit anti-GPrbc antiserum. Data expressed asmeans ±SD (number of animals per group).
by guest on March 21, 2014
jn.nutrition.orgD
ownloaded from

CHOLESTEROL EFFECTS ON PHAGOCYTOSIS AND LYSIS 1679
aged and ill where defenses are alreadylow.
We found no difference in susceptibility to Ab-dependent phagocytosis ofCONT and CHOL rbc. Guinea pigs feda cholesterol-supplemented diet havegrossly enlarged livers and spleens anddie of an anemia that develops about8-12 weeks after initiation of cholesterolfeeding, whereas the rbc of these CHOLanimals show a spurred morphologywithin days (2). It has been suggestedthat the anemia associated with certainhepatic disorders is caused by increasedclearance of rbc by the reticuloendothe-lial system due to the spur shape of therbc caused by their increased cholesterolcontent (29). Our results do not supportthis conclusion at least for the guinea pigsystem. CHOL rbc have the spur morphology due to increased cholesterolcontent but are not phagocytosed to agreater extent than normal rbc in the Ab-dependent assay. However, a small percentage of sensitive cells would not havebeen detected by this method.
Increasing the cholesterol content ofsheep erythrocyte and liposomal membranes has been reported to decreasecomplement-mediated hemolysis and release of intraliposomal contents, presumably due to inhibition of transmembranechannel formation (10). Since complement-mediated and cell-mediated membrane damage share a number of essential features and are probably due to thesame mechanisms (30), we might haveexpected a decreased ADCC of CHOLrbc. We found no difference betweenCONT and CHOL rbc in this Ab-dependent and cell-mediated assay, eventhough increasing the cholesterol contentof membranes has been reported to increase Ab binding (31).
We also found no difference in Abbinding using a hemagglutination assay,but this test is relatively insensitive. Thelack of any difference between controland cholesterol enriched rbc in susceptibility to phagocytosis and Ab dependent lysis may be due to the fact thatCHOL rbc matured in vivo in a cholesterol-enriched environment as opposedto being cholesterol-enriched in vitro as
a mature cell. This suggests that cholesterol may affect membrane function differently when it is present during cellmaturation than when it is inserted intoan existing membrane in vitro. The observation that enrichment of membranecholesterol in a mycoplasma by growth ofthe cells in a cholesterol-enriched medium does not affect complement mediated lysis (12) supports this concept.
ACKNOWLEDGMENTS
The authors wish to thank Gayle Hoffman for technical assistance.
LITERATURE CITED
1. Ostwald, R. & Shannon, A. (1964)Composition of tissue lipids and anaemia ofguinea pigs in response to dietary cholesterol.Biochem. J. 91, 146-154.
2. Aswad, C. & Ostwald, R. (1976) Spiculatederythrocytes of cholesterol-fed guinea pigs:changes of morphology, composition, and osmotic fragility. Proc. Soc. Exp. Biol. Med. 153,505-510.
3. Davis, B. D., Dulbecco, R., Eisen, H. N., Ginsberg, H. S., Wood, W. B. & McCarty, M.,eds. (1973) Microbiology, pp. 643-648, Harper and Row, Hagerstown, MD.
4. Berken, A. & Benecerraf, B. (1968)Depression of reticuloendothelial systemphagocytic function by ingested lipids. Proc.Soc. Exp. Biol. Med. 128, 793-795.
5. Di Luzio, N. R. & Wooles, W. R. (1964)Depression of phagocytic activity and immuneresponse by methyl palmitate. Am. J. Physiol.206, 939-943.
6. Dianzani, M. U., Tornelli, M. V., Canuto, R. A.,Garcea, R. & Feo, F. (1976) The influenceof enrichment with cholesterol on the phagocytic activity of rat macrophages. J. Path. 118,193-199.
7. Neveu, T., Biozzi, G., Benacerraf, B., Stiffel, C.& Halpem, B. N. (1956) Role of reticulo-en-dothelial system in blood clearance of cholesterol. Am. J. Physiol. 187, 269-274.
8. Griffin, F. M. & Silverstein, S. C.(1974) Segmental response of the macrophageplasma membrane to a phagocytic stimulus. J.Exp. Med. 139, 323-336.
9. Ostwald, R., Yamanaka, W., Irwin, I. D.,Hansma, H., Light, M. & Tom, K.(1971) Effects of dietary modifications on cholesterol-induced anemia in guinea pigs. J. Nutr.101, 699-712.
10. Shin, M. L., Paznekas, W. A. & Mayer, M.M. (1978) On the mechanism of membranedamage by complement: the effect of lengthand unsaturation of the acyl chains in liposomal
by guest on March 21, 2014
jn.nutrition.orgD
ownloaded from

1680 DUWE, FITCH AND OSTWALD
bilayers and the effect of cholesterol concentration in sheep erythrocyte and liposomalmembranes. J. Immunol. 120, 1996-2002.
11. Ohanian, S. H., Schlager, S. L, Yamazaki, M.& Ishida, B. (1979) Effect of specific phos-pholipids on the antibody-complement-mediated killing of nucleated cells. J. Immunol.123, 1014-1019.
12. Dahl, J. S., Dahl, C. E. & Levine, R. P.(1979) Role of lipid fatty acyl composition andmembrane fluidity in the resistance of Acho-leplasma laidlawii to complement-mediatedkilling. J. Immunol. 123, 104-108.
13. Stahelin, H., Suter, E. & Karnovsky, M.(1956) Studies on the interaction betweenphagocytes and tubercle bacilli. 1. Observations on the metabolism of guinea pig leukocytes and the influence of phagocytosis. J. Exp.Med. 104, 121-136.
14. Walker, W. S. & Demus, A. (1975) Antibody-dependent cytolysis of chicken erythrocytes byan in vitro-established line of mouse peritonealmacrophages. J. Immunol. 114, 765-769.
15. Ostwald, R., Yamanaka, W., Light, M. & Kroes,J. (1977) The time course of metabolicchanges induced by dietary cholesterol inguinea pigs. Atherosclerosis 26, 41-53.
16. Sardet, C., Hansma, H. & Ostwald, R.(1972) Effects of plasma lipoproteins fromcontrol and cholesterol-fed guinea pigs on redcell morphology and cholesterol content: an invitro study. J. Lipid Res. 13, 705-715.
17. Kroes, J. & Ostwald, R. (1971) Erythrocytemembranes—effect of increased cholesterolcontent on permeability. Biochem. Biophys.Acta 249, 647-650.
18. Ip, S. H. C., Abrahm, J., & Cooper, R.A. (1980) Enhancement of blastogenesis incholesterol-enriched lyphocytes. J. Immunol.124, 87-93.
19. Fogelman, A. M., Seager, J., Edwards, P. A.,Hokom, M. & Popjak, G. (1977) Cholesterolbiosynthesis in human lymphocytes, mono-cytes and granulocytes. Biochem. Biophys. Res.Commun. 76, 167-173.
20. Van Deenen, L. L. M. & deGier, J.(1964) Chemical composition and metabolism
of lipids in red cells of various animal species.In: The Red Blood Cell (Bishop, C. & Surge-nor, D. M., eds.), pp. 243-307, Academic Press,New York.
21. Tsan, M. F. & Berlin, R. D. (1971) Effect ofphagocytosis on membrane transport of non-electrolytes. J. Exp. Med. 134, 1016-1035.
22. Schmidt, M. E. & Douglas, S. D.(1972) Disappearance and recovery of humanmonocyte IgG receptor activity after phagocytosis. J. Immunol. ¿09,914-917.
23. Mason, R. J., Stessei, T. P. & Vaughan,M. (1972) Lipids of alveolar macrophages,polymorphonuclear leucocytes, and theirphagocytic vesicles. J. Clin. Invest. 51,2399-2407.
24. Edelman, G. M., Yahara, I. & Wang, J.L. (1973) Receptor mobility and receptor-cytoplasmic interactions in lymphocytes. Proc.Nati. Acad. Sci. U.S.A. 70, 1442-1446
25. Curtiss, L. K. & Edgington, T. S.(1976) Regulatory serum lipoproteins: regulation of lymphocyte stimulation by a speciesof low density lipoprotein. J. Immunol. 116,1452-1458.
26. Chisari, F. V. & Edgington, T. S.(1975) Lymphocyte E-rosette inhibitory factor: a regulatory serum lipoprotein. J. Exp.Med. 142, 1092-1107.
27. Yamazaki, M., Shinoda, H., Hattori, R. & Mi-zuno, D. (1977) Inhibition of macrophage-mediated cytolysis by lipoproteins from cell-free tumorous ascites. Gann 68, 513-516.
28. Stuart, A. E. (1962) Effect of cholesterololeate on the phagocytic function of the reti-culo-endothelial system. Nature 196, 78-79.
29. Shohet, S. B. (1972) Hemolysis and changesin erythrocyte membrane lipids. N. Engl. J.Med. 286, 638-644.
30. Mayer, M. M. (1977) Mechanism of cytolysisby lymphocytes: a comparison with complement. J. Immunol. 119, 1195-1203.
31. Humphries, G. M. K. & McConnell, H.M. (1975) Antigen mobility in membranesand complement-mediated immune attack. Proc.Nati. Acad. Sci. U.S.A. 72, 2483-2487.
by guest on March 21, 2014
jn.nutrition.orgD
ownloaded from




















