
Cholesterol under oxidative stress – how lipid membranes sense oxidation as cholesterol is being...
38
Author's Accepted Manuscript Cholesterol under oxidative stress – how lipid membranes sense oxidation as cholesterol is being replaced by oxysterols Waldemar Kulig, Agnieszka Olżyńska, Piotr Jurkiewicz, Anu M. Kantola, Sanna Komulainen, Moutusi Manna, Mohsen Pourmousa, Mario Vazdar, Lukasz Cwiklik, Tomasz Rog, George Khelashvili, Daniel Harries, Ville-Veikko Telkki, Martin Hof, Ilpo Vattulainen, Pavel Jungwirth PII: S0891-5849(15)00118-5 DOI: http://dx.doi.org/10.1016/j.freeradbiomed.2015.03.006 Reference: FRB12346 To appear in: Free Radical Biology and Medicine Received date: 12 February 2015 Revised date: 6 March 2015 Accepted date: 9 March 2015 Cite this article as: Waldemar Kulig, Agnieszka Olżyńska, Piotr Jurkiewicz, Anu M. Kantola, Sanna Komulainen, Moutusi Manna, Mohsen Pourmousa, Mario Vazdar, Lukasz Cwiklik, Tomasz Rog, George Khelashvili, Daniel Harries, Ville- Veikko Telkki, Martin Hof, Ilpo Vattulainen, Pavel Jungwirth, Cholesterol under oxidative stress – how lipid membranes sense oxidation as cholesterol is being replaced by oxysterols, Free Radical Biology and Medicine, http://dx.doi. org/10.1016/j.freeradbiomed.2015.03.006 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting galley proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. www.elsevier.com/locate/freerad- biomed
Transcript of Cholesterol under oxidative stress – how lipid membranes sense oxidation as cholesterol is being...

Author's Accepted Manuscript
Cholesterol under oxidative stress – how lipidmembranes sense oxidation as cholesterol isbeing replaced by oxysterols
Waldemar Kulig, Agnieszka Olżyńska, PiotrJurkiewicz, Anu M. Kantola, Sanna Komulainen,Moutusi Manna, Mohsen Pourmousa, MarioVazdar, Lukasz Cwiklik, Tomasz Rog, GeorgeKhelashvili, Daniel Harries, Ville-Veikko Telkki,Martin Hof, Ilpo Vattulainen, Pavel Jungwirth
PII: S0891-5849(15)00118-5DOI: http://dx.doi.org/10.1016/j.freeradbiomed.2015.03.006Reference: FRB12346
To appear in: Free Radical Biology and Medicine
Received date: 12 February 2015Revised date: 6 March 2015Accepted date: 9 March 2015
Cite this article as: Waldemar Kulig, Agnieszka Olżyńska, Piotr Jurkiewicz, AnuM. Kantola, Sanna Komulainen, Moutusi Manna, Mohsen Pourmousa, MarioVazdar, Lukasz Cwiklik, Tomasz Rog, George Khelashvili, Daniel Harries, Ville-Veikko Telkki, Martin Hof, Ilpo Vattulainen, Pavel Jungwirth, Cholesterolunder oxidative stress – how lipid membranes sense oxidation as cholesterolis being replaced by oxysterols, Free Radical Biology and Medicine, http://dx.doi.org/10.1016/j.freeradbiomed.2015.03.006
This is a PDF file of an unedited manuscript that has been accepted forpublication. As a service to our customers we are providing this early version ofthe manuscript. The manuscript will undergo copyediting, typesetting, andreview of the resulting galley proof before it is published in its final citable form.Please note that during the production process errors may be discovered whichcould affect the content, and all legal disclaimers that apply to the journalpertain.
www.elsevier.com/locate/freerad-
biomed

���
Cholesterol under oxidative stress – how lipid membranes
sense oxidation as cholesterol is being replaced by oxysterols
Waldemar Kulig1*, Agnieszka Ol�y�ska2, Piotr Jurkiewicz2*, Anu M. Kantola3, Sanna
Komulainen3, Moutusi Manna1, Mohsen Pourmousa1, Mario Vazdar4,1, Lukasz Cwiklik2,5*,
Tomasz Rog1, George Khelashvili6, Daniel Harries7,�Ville-Veikko Telkki3, Martin Hof2, Ilpo
Vattulainen1,8, Pavel Jungwirth5,1
1 Department of Physics, Tampere University of Technology, P. O. Box 692, FI-33101 Tampere,
Finland
2 J. Heyrovský Institute of Physical Chemistry, Academy of Sciences of the Czech Republic, v.
v. i., Dolejskova 3, 18223 Prague 8, Czech Republic
3 Department of Physics and Chemistry, University of Oulu, P. O. Box 3000, FI-90014 Oulu,
Finland
4 Rudjer Boškovi� Institute, Division of Organic Chemistry and Biochemistry, POB 180, HR-
10002 Zagreb, Croatia
5 Institute of Organic Chemistry and Biochemistry, Academy of Sciences of the Czech Republic,
Flemingovo nám. 2, 16610 Prague 6, Czech Republic
6 Weill Cornell Medical College, New York, New York 10065, United States
7 Institute of Chemistry and the Fritz Haber Research Center, The Hebrew University of
Jerusalem, Jerusalem 91904, Israel
8 MEMPHYS-Center for Biomembrane Physics, University of Southern Denmark, Odense,
Denmark
Corresponding authors: [email protected] (WK), [email protected] (PJ), and
[email protected] (LC)

���
Abstract:
The behavior of oxysterols in phospholipid membranes and their effects on membrane properties
was investigated by means of dynamic light scattering, fluorescence spectroscopy, NMR, and
extensive atomistic simulations. Two families of oxysterols were scrutinized – tail-oxidized
sterols, which are mostly produced by enzymatic processes, and ring-oxidized sterols, formed
mostly via reactions with free radicals. The former family of sterols was found to behave
similarly to cholesterol in terms of molecular orientation, roughly parallel to the bilayer normal,
leading to increasing membrane stiffness and suppression of its membrane permeability. In
contrast, ring-oxidized sterols behave quantitatively differently from cholesterol. They acquire
tilted orientations and therefore disrupt the bilayer structure with potential implications for
signaling and other biochemical processes in the membranes.
Keywords:�phospholipid bilayers, oxysterols, molecular dynamics simulations, DPH anisotropy,
NMR measurements, Laurdan fluorescence, liposomes, tilt modulus.�
Abbreviations: LXR, liver X receptors; HDL, high-density lipoproteins; MD, molecular
dynamics; NMR, nuclear magnetic resonance; POPC, 1-palmitoyl-2-oleoyl-sn-glycero-3-
phosphocholine; POPC-d31, POPC deuterated at the sn-1 chain; chol, cholesterol; 7�-OH-chol,
7�-hydroxycholesterol; 7�-OH-chol, 7�-hydroxycholesterol; 7-keto-chol, 7-ketocholesterol;
3�5�6�-3OH-chol, 3�,5�,6�-trihydroxycholestane; 25-OH-chol, 25-hydroxycholesterol; 27-OH-
chol, 27-hydroxycholesterol; 24-OH-chol, 24-hydroxycholesterol; 24S-OH-chol, 24S-
hydroxycholesterol; HEPES, 4-(2-hydroxyethyl)piperazine-1-ethanesulfonic acid; POPOP, 2,2'-
(1,4-phenylene)bis[5-phenyl-oxazole]; DPH, 1,6-diphenyl-1,3,5-hexatriene; Laurdan, 6-lauroyl-
2-dimethylaminonaphthalene; LUVs, large unilamellar vesicles; DLS, dynamic light scattering;
UV, ultraviolet; SS, steady-state; TCSPC, time correlated single photon counting; GP,
generalized polarization; TDFS, time-dependent fluorescence shift; �r, integrated relaxation time;
��, total spectral shift; TRES, time-resolved emission spectra; OPLS, optimized parameters for
liquid simulation; NPT, isobaric-isothermic ensemble; PME, particle mesh Ewald scheme; APM,
area per molecule; |SCD|, deuterium order parameters; A�, amyloid beta peptide; 2D, two-
dimensional; FWHM(t), full-width at half-maximum;

���
INTRODUCTION
Cellular membranes consist of a bilayer composed primarily of phospholipids and sterols, in
which a large amount of proteins and other molecules is embedded. Sterols are known to affect,
e.g., the conformational order of the lipid acyl chains,[1, 2] membrane permeability,[2]
hydrophobic thickness,[1] and lateral organization of the bilayers.[1, 3-5] In contrast to the vast
abundance of phospholipid moieties, most biological membranes contain only one major sterol
type. Cholesterol is the most prevalent and essential sterol in mammalian cellular membranes
and is required for diverse cellular functions, including binding to sterol-sensing domains to
regulate protein function,[6, 7] participating in the formation of lipid rafts,[8, 9] and serving as a
precursor for bile acid and steroid hormone synthesis.[10] Because the presence of cholesterol
affects so many important functions in human cells, it is not surprising that even a small change
in chemical structure of cholesterol may significantly alter membrane functions and
dynamics.[11-13]
Oxysterols (see Figure 1) are oxidized derivatives of cholesterol with one or more additional
oxygen containing functional groups (hydroxyl, carbonyl, carboxyl or epoxy). The most
abundant oxysterols in human body (27-, 24(S)-, 7�-, 4�-hydroxycholesterol) are generated in
cells catalyzed by mitochondrial or endoplasmic reticulum cholesterol hydroxylases (part of the
cytochrome P450). Oxysterols may also arise through non-enzymatic, free-radical, and lipid
peroxide oxidative processes, usually referred to as cholesterol autoxidation processes. The most
common oxysterols produced during cholesterol autoxidation are 7�-hydroxycholesterol and 7-
ketocholesterol. Oxysterols are present in healthy human and animal tissues at low concentration
(as compared to cholesterol), but are found to be profoundly enriched in pathologic conditions
such as macrophage foam cells, atherosclerotic lesions, and cataracts.[14-17]
Oxysterols play a crucial role in many regulatory processes in human body. Oxysterols act as
intermediates in cholesterol catabolism and especially in bile acid synthesis. Therefore, they are
involved in the elimination of excess cholesterol from the body.[18, 19] They regulate lipids
metabolism as ligands of liver X receptors (LXR),[20, 21] key receptors in hepatic lipogenesis,
synthesis of nascent high-density lipoproteins (HDL), and biliary neutral sterol secretion.[22, 23]
Oxysterols also take part in pro-inflammatory signaling[24, 25] and modulation of the estrogen
receptor function.[26-28] Some oxysterols (e.g., 7-ketocholesterol) show cytotoxic and pro-

���
apoptotic properties.[29-31] Pathogenic effects of oxysterols have also been described in
cardiovascular diseases,[16, 32] diabetes[33] type-2, and degenerative disorders such as the
Alzheimer disease,[34-36] osteoporosis,[37] and age-related macular degeneration.[38, 39]
There are also reports describing the role of oxysterols in neurological diseases.[25, 40]
Oxysterols might play a pathological role in the Smith-Lemli-Opitz syndrome.[41]
The role of oxysterols in the above mentioned natural and pathological situations has been
confirmed, but the molecular mechanisms governing the oxysterols’ behavior in human body are
still elusive. There is limited amount of experimental[42, 43] and computational[44-46] data
describing molecular details of oxysterol interactions with other constituents of mammalian cell
membranes.
Here, we employed time-resolved fluorescence spectroscopy, dynamic light scattering
experiments, NMR measurements, and theoretical modeling based on extensive all-atom
molecular dynamics (MD) simulations to elucidate the effect of nine most common oxysterols
(see Figure 1) in the human body on model biological membranes. We found that the oxysterols
can be classified to two distinct groups depending on their effects on lipid membranes, and one
of the oxysterol groups induces membrane behavior that is distinctly different from membranes
rich in cholesterol. Given that cholesterol oxidation is one of the common processes taking place
in membranes, and cholesterol is known to interact specifically with and govern the function of
several membrane proteins and receptors,[47-49] our results suggest that cholesterol oxidation
may have severe implications for central cellular functions, such as signaling.
MATERIAL AND METHODS
Materials
1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC), ovine wool cholesterol (chol), 7�-
hydroxycholesterol (7�-OH-chol), 7�-hydroxycholesterol (7�-OH-chol), 4�-hydroxycholesterol
(4�-OH-chol), 7-ketocholesterol (7-keto-chol), and 3�,5�,6�-trihydroxycholestane (3�5�6�-
3OH-chol) were obtained from Avanti Polar Lipids, Inc. (Alabaster, AL, USA). 27-
hydroxycholesterol (27-OH-chol) and 24-hydroxycholesterol (24-OH-chol) were ordered from
Medical Isotopes, Inc. (Pelham, NH, USA). 25-hydroxycholesterol (25-OH-chol), 4-(2-

���
hydroxyethyl)piperazine-1-ethanesulfonic acid (HEPES), sodium chloride (NaCl), sodium
hydroxide (NaOH), ethylenedinitrilotetraacetic acid (EDTA), and 2,2'-(1,4-phenylene)bis[5-
phenyl-oxazole] (POPOP) were obtained from Sigma-Aldrich (St. Louis, MO, USA). 1,6-
diphenyl-1,3,5-hexatriene (DPH) and 6-lauroyl-2-dimethylaminonaphthalene (Laurdan) were
purchased from Invitrogen (Eugene, OR, USA). Buffer was prepared using Mili Q water
(Milipore, USA). Organic solvents of spectroscopic grade were supplied by Merck (Darmstadt,
Germany). All chemicals were used without further purification.
For NMR work, POPC deuterated at the sn-1 chain (POPC-d31), cholesterol, 3�,5�,6�-
trihydroxycholestane (3�5�6�-3OH-chol), and 24S-hydroxycholesterol (24S-OH-chol) were
obtained from Avanti Polar Lipids. Deuterium depleted water and organic solvents (chloroform
and methanol, ACS reagent grade) were obtained from Sigma-Aldrich.
Preparation of liposomes
Extruded large unilamellar vesicles (LUVs) were prepared as described below. The required
volumes of chloroform solutions of POPC and sterols were mixed with methanol solution of a
fluorescent probe in a glass tube. Final molar ratios of lipids:probe were 100:1 and 500:1 for
Laurdan and DPH, respectively. Solvents were evaporated under a stream of nitrogen and then
under vacuum for at least 2 hours. The dry lipid film was suspended in 10 mM HEPES buffer
(150 mM NaCl, pH 7.4, 0.2 mM EDTA). After 5 minutes of vortexing the suspension was
extruded through polycarbonate membranes (100 nm pore diameter, Avestin, Ottawa, Canada).
The final total concentration of lipids with (or without) sterols in the sample was 1 mM. The
prepared samples were transferred to a 1 cm quartz cuvette and equilibrated at 310 K (with
water-circulating thermostat; ± 0.5 K accuracy) for 15 min before each measurement (unless
otherwise specified).
For the preparation of NMR samples, POPC-d31, cholesterol, and 24S-hydroxycholesterol were
dissolved into chloroform, and 3�,5�,6�-trihydroxycholestane into a methanol:chloroform
mixture (1:6). Samples of 20 mg of POPC-d31 as well as POPC-d31 with 10 mol % of
cholesterol, 3�,5�,6�-trihydroxycholestane, or 24S-hydroxycholesterol were prepared by
pipetting and mixing appropriate solutions, removing the solvent in a rotary evaporator followed

���
by five hours (chloroform) to overnight (methanol:chloroform) evaporation in vacuum line. The
dried samples were hydrated in 250 µl of sodium phosphate buffer (pH = 7) prepared in
deuterium depleted water, shaken and vortexed for 20 minutes, and finally transferred into 225
µl KelF sealing cells for NMR measurements and let to age overnight before the experiments.
Dynamic Light Scattering (DLS)
Sizes of the liposomes were measured by means of Dynamic Light Scattering (DLS) using
Zetasizer Nano ZS (Malvern Instruments Ltd., Worcestershire, UK). Intensity-weighted size
distributions were obtained using regularized fitting implemented in Zetasizer Software 6.2. All
samples were transferred to UV grade poly(methyl methacrylate) cuvettes (Kartell, Noviglio,
Italy) and equilibrated at 298 K for 3 minutes before the measurement.
Steady-State and Time-Resolved Fluorescence instrumentation
All steady-state (SS) fluorescence measurements were performed on a Fluorolog-3
spectrofluorimeter (model FL3-11; Jobin Yvon Inc., Edison, NJ) equipped with a Xenon-arc
lamp. Fluorescence decays were recorded on a time correlated single photon counting (TCSPC)
setup (model 5000U, IBH, Glasgow, U.K.) equipped with an IBH laser diode NanoLED 11 (370
nm peak wavelength, 80 ps pulse width, 1 MHz maximum repetition rate) and a cooled
Hamamatsu R3809U-50 microchannel plate photomultiplier. Emission decays for Laurdan were
recorded at 400-540 nm wavelengths in 10 nm steps at magic angle polarization. 399 nm cutoff
filter was used to eliminate scattered light.
Laurdan fluorescence: Generalized Polarization (GP), Time-Dependent Fluorescence Shift
method (TDFS)
Steady-state and time-resolved fluorescence of Laurdan precisely located in the model lipid
bilayer[50] was used to characterize hydration and mobility of lipids at the level of lipid
carbonyls. The methodology used in this work is identical to the one published in our recent
study.[51] Briefly, Laurdan steady-state excitation spectra (EM = 440 and 490 nm) were used to
calculate excitation generalized polarization spectra, GPEX(�EX) = (I440 − I490) / (I440 + I490), where
I440 and I490 represent fluorescence intensities emitted at 440 nm and 490 nm, respectively
(excited at excitation wavelength �EX).

���
For Time-Dependent Fluorescence Shift method (TDFS), Laurdan steady-state emission spectrum (EX = 373 nm) together with fitted fluorescence decays were used to reconstruct time-resolved emission spectra (TRES).[52] From the position of TRES maximum, �(t), two main parameters were derived. Total emission shift, �� = �(0) – �(�), reflects polarity of the hydrated lipid carbonyls. The position of TRES maximum at t = 0, �(0), was estimated using the method by Fee and Maroncelli[53] to be 23800 cm-1 for all of the studied samples. Mobility of the polar moieties in the probe vicinity was evaluated in terms of the mean integrated relaxation time,
[ ]0
( ) ( ) / (0) ( )r t dtτ ν ν ν ν∞
= − ∞ − ∞� . Intrinsic uncertainties for the TDFS parameters were 50 cm-1
and 0.05 ns for �� and �r, respectively. The percentage of the relaxation process that was faster than the resolution of the instrumentation was estimated to be ~25 % for most of the measured samples (see also Discussion in the Results section) usually did not exceed 20% for any of the samples measured. Further details of TDFS method can be found in Refs. [50, 54] and in a recent review,[55] where GP and TDFS methods are compared.
DPH anisotropy
DPH anisotropy measurements[56] were used to evaluate the mobility of acyl chains in a lipid
bilayer. For details of the methodology used, please refer to our recent study.[51] Briefly, the
time-resolved anisotropy of DPH was determined using global fitting (FluoFit v.4.5 by
PicoQuant, Berlin, Germany) of the fluorescence decays measured at vertical and horizontal
orientation of the emission polarizer. Excitation/emission wavelengths were 373/466 nm,
respectively. The G-factor was measured for ethanol solution of POPOP using a tail matching
method. The anisotropy data were evaluated according to wobbling-in-cone model introduced by
Kinosita, Kawato, and Ikegami.[57, 58] DPH orientational order parameter, SDPH, was calculated
to estimate the membrane acyl chain order.[59, 60] The so-called wobbling diffusion constant,
Dw, reflecting the dynamics of the lipid bilayer interior, was calculated using the method of
Lipari and Szabo.[61]
NMR measurements
Deuterium NMR spectra were measured at 310 K on 7.05 T Bruker Avance III spectrometer
corresponding to deuterium resonance frequency of 46 MHz using solid state 7 mm variable
angle probehead by Doty Scientific, RF solenoid coil aligned perpendicular to the external field.

��
Spectra were acquired using quadrupole echo sequence with 90-degree pulse of 5.3 µs, 35 µs
interpulse delays and 1 µs dwell time. The acquired data was left shifted to the echo maximum
before Fourier transformation. For each spectrum, 100000 scans were accumulated. Sample
temperature was equilibrated for 60 minutes before starting the experiments.
Due to the rapid axially symmetric reorientation of the chain, the orientation order parameter of
the CD-bond can be written as
21
(3 1)2CD CD
S cos θ= − , (1)
where θCD is the angle between the carbon-deuterium bond and the axis of symmetry of the chain
motions. The quadrupolar splitting observable in the deuterium spectrum can then be expressed
as
2
2
3 ( )
2Q CD
e qQS P cos
hν β=� , (2)
where β is the angle between the reorientational symmetry axis and the external magnetic field
of the spectrometer, P2(cos β)=1/2(3cos2β-1) is the second order Legendre polynomial (that is 1
for β =0°), and e2qQ/h is the quadrupolar coupling constant, which is ~170 kHz for CD-bonds in
methyl and methylene groups in aliphatic chains.[62] The powder pattern arising from a
distribution of orientations in multilamellar vesicles was transformed into spectra corresponding
to a sample with unique orientation of the reorientation axis along the external field (β =0°)
using the de-Pakeing Fourier transform algorithm.[63] De-Pakeing of the spectra was performed
utilizing available code by Sani et al.[64] which implements the Fourier transform method of de-
Pakeing by McCabe and Wassall into NMRpipe software.[65] The spectral peaks were fitted
with Origin software (OriginLab) to obtain peak positions as well as intensities.
Atomistic molecular dynamics simulations
All-atom molecular dynamics (MD) simulations were performed for the POPC bilayers
containing 10 mol% cholesterol, 7-ketocholesterol, 7�-hydroxycholesterol, 7�-
hydroxycholesterol, 4�-hydroxycholesterol, 3�,5�,6�-trihydroxycholestane, 27-

��
hydroxycholesterol, 24S-hydroxycholesterol, 25-hydroxycholesterol, or 25S-cholestenoic acid,
as well as for a pure POPC bilayer. Chemical structures of all compounds are shown in Figure 1.
All the bilayers were composed of 128 molecules (lipids and sterols). The initial structure of
each bilayer was obtained by putting lipid molecules on an 8 x 8 grid, resulting in a bilayer
comprised of two monolayers with 64 lipids in each leaflet. Initial structures of the sterol-
containing bilayers were obtained by randomly exchanging 14 lipid molecules by the sterol
molecules.
Topologies of sterol molecules were built using parameters of existing molecular blocks from the
all-atom OPLS (Optimized Parameters for Liquid Simulation) force field.[66, 67] Partial charges
were also taken from the original all-atom OPLS force field. Force field parameters for lipids
were taken from Ref. [68]. Water molecules are described using the TIP3P model.[69]
The simulation temperature was kept at 310 K using the Nosé-Hoover thermostat[70, 71] with a
temperature coupling constant of 0.4 ps. The temperatures of the solute and solvent were
controlled independently. Periodic boundary conditions were used in all three directions. The
pressure was kept constant with a semi-isotropic scheme, meaning that the pressure in x and y
directions (i.e., in the membrane plane) was coupled separately from the pressure in the z
direction. The Parrinello-Rahman barostat[72, 73] was used to keep the pressure at 1 atm with a
pressure coupling constant of 1ps and a compressibility of 4.5�10-5 bar-1. Long-range electrostatic
interactions beyond a non-bonded interaction cutoff of 1.0 nm were treated by the Particle Mesh
Ewald scheme (PME)[74] with a Fourier spacing of 0.1 nm and a sixth order interpolation to the
Ewald mesh. A long-range dispersion correction to energy and pressure was added. The LINCS
algorithm[75] was used to constrain all covalent bonds, allowing a time step of 2 fs. For water,
the SETTLE method[76] was applied.
The systems were first energy minimized using the steepest descent algorithm, and then
equilibrated in the isobaric-isothermic (NPT) ensemble until stable areas per molecule were
obtained. The MD simulations of all bilayer systems were carried out over 400 ns. The first 200
ns was considered as an equilibration period, thus only the last 200 ns of each trajectory was
analyzed. All simulations were performed with the GROMACS 4.6.x software package.[77, 78]

����
Analysis of MD data
Several measurable quantities were extracted from the simulation data. The area per molecule
(APM) was calculated by dividing the total area of the simulation box in the x-y plane by the
number of all molecules (lipids and sterols) in a single leaflet. The membrane thickness was
calculated as the average head-to-head (phosphate-to-phosphate) distance between the separate
leaflets. The average values of the deuterium order parameters[79] (|SCD|) were calculated for the
acyl chains of lipid molecules using standard GROMACS scripts by averaging over all carbons
from the methylene and methyl groups of the lipid molecules tails (in POPC bilayers over C2-
C15 (sn-1 tail) or C2-C17 (sn-2 tail) atoms). The tilt angle of the sterol rings was defined as the
angle between the C3-C17 vector (see Figure 1) and the bilayer normal.
Calculations of sterol tilt modulus
To quantify the free energy cost of sterol tilting in a lipid membrane, we used a previously
establish protocol to extract the tilt modulus, , from the sterol tilt (�; � = 0 describes the sterol
tilt where its ring plane is parallel to the bilayer normal) probability distributions P(�) that are
obtained from atomistic MD simulations.[80, 81] A quadratic fit was performed to an expression
of the form ln[ ( ) / sin ]B
k T P θ θ− over an angular interval chosen to represent the best-sampled
angular region and at the same time to limit the fit to the low � regime, where the quadratic
expansion of the free energy with respect to tilt is valid. Finally, was determined from the
coefficient that corresponded to this fit (additional details of this methodology can be found in
Refs. [80, 81].
Quantification of sterol insertion into lipid membranes
To quantify insertion of different sterol molecules into lipid membranes from atomistic
simulations, the MD trajectories for each sterol were analyzed to obtain a normalized two-
dimensional (2D) probability distribution P(�, h) of finding a sterol molecule with a tilt angle �
and with its main hydroxyl (OH) group (that is present in cholesterol) at a vertical distance h
from the bilayer midplane. We define � = 0 as the state where sterol with its ring plane is parallel
to the bilayer normal, and h = 0 corresponds to the sterol OH group at the membrane midplane.

����
RESULTS
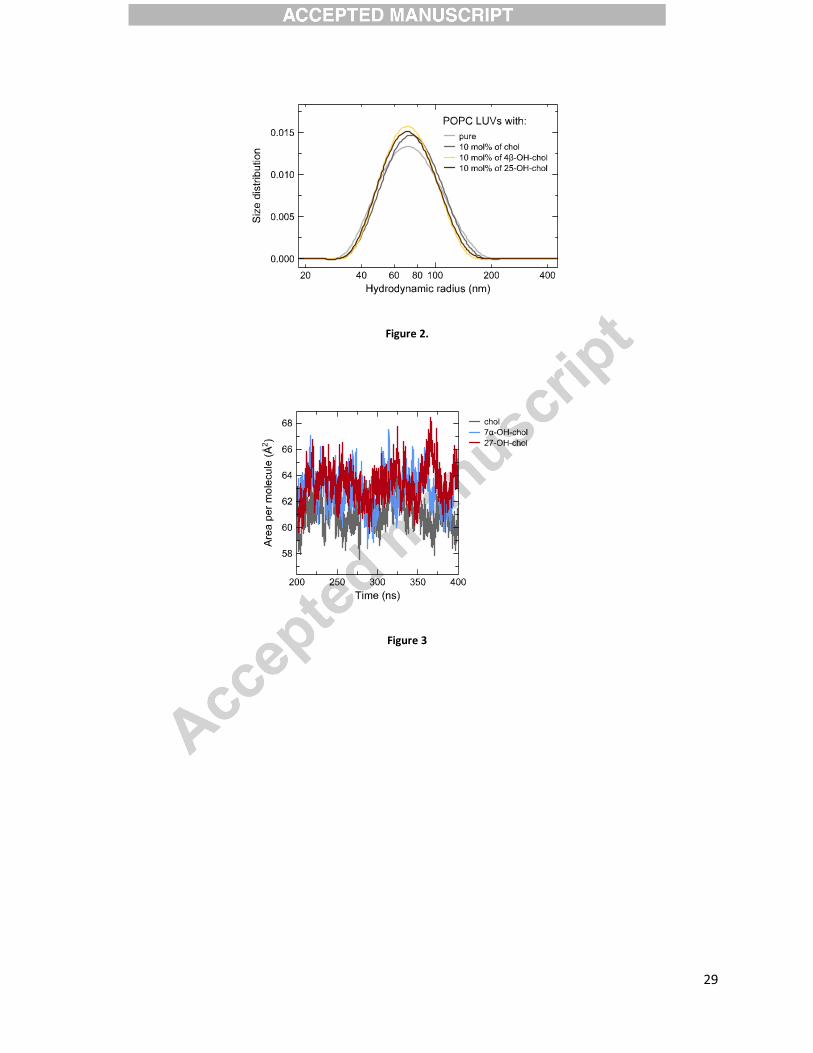
Morphology of lipid vesicles – Dynamic Light Scattering (DLS)
To study the morphology of lipid aggregates consisting of POPC and 10 mol% of one of the
sterols, DLS measurements were performed on extruded samples. The results show that addition
of the sterol does not disturb the lamellar phase of the vesicles. Size distributions are unimodal
representing a uniform population of large unilamellar vesicles (LUVs) with a hydrodynamic
radius centered at (70 ± 2) nm. It is apparent that all sterols are fully miscible with POPC at this
concentration. The distribution for pure POPC is only slightly broader than for sterol-containing
samples (Figure 2). The changes in liposome sizes upon cholesterol oxidation (i.e., between 10
mol% cholesterol and any of 10 mol% oxysterol samples) are within the uncertainty of the
method.
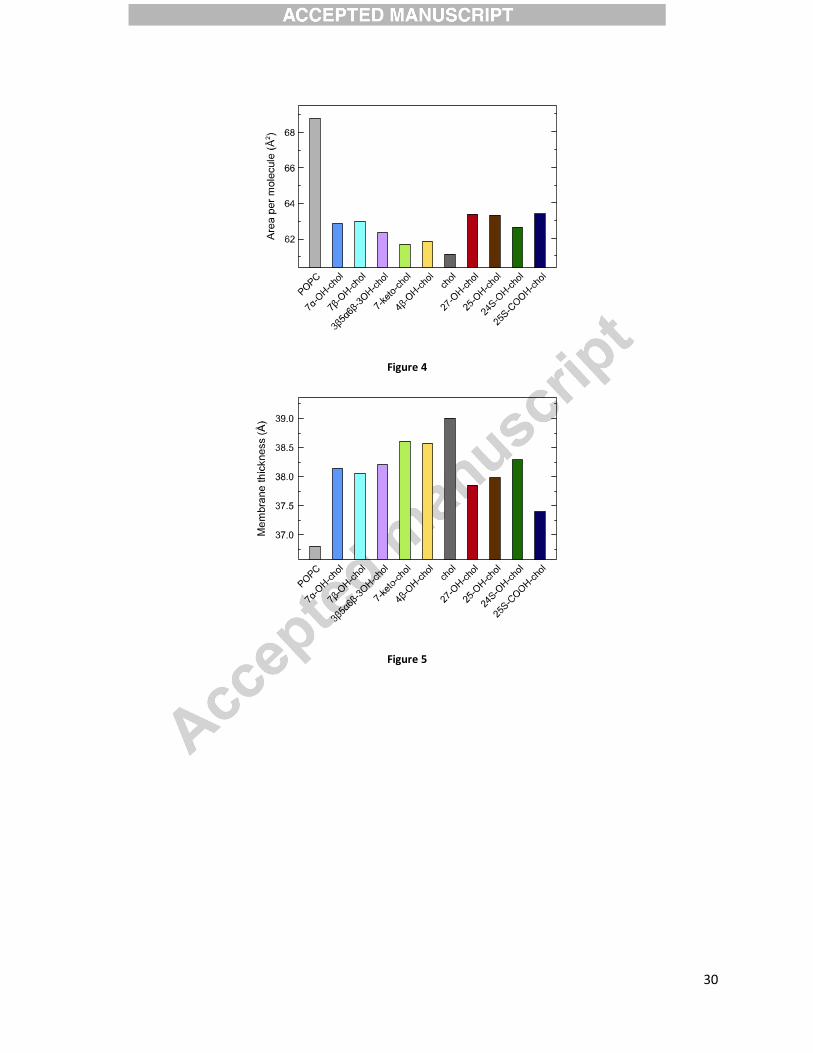
All sterols condense POPC bilayer and make it thicker
To examine in detail the structural properties of lipid/sterol membranes, we performed all-atom
MD simulations.
The time evolution (the last 200 ns out of ~400 ns simulations) of the APM of the bilayers
containing 10 mol% cholesterol, 7�-hydroxycholesterol, or 27-hydroxycholesterol is shown in
Figure 3. It shows that the APM is fairly stable indicating proper equilibration of the systems.
The time evolution of other systems studied here (not shown) is very similar, also indicating
proper equilibration. Averaged values of area per molecule for all the systems studied here are
presented in Figure 4. An average APM for the pure POPC bilayer calculated here is (68.8 ± 0.8)
Å2. This value matches well with the APM of 68.3 Å2 reported in experiments.[82] It is rather
clear that all sterols condense the bilayer as compared to the pure POPC bilayer. Cholesterol
shows the strongest condensation effect (i.e., the smallest APM value), followed by 7-
ketocholesterol and 4�-hydroxycholesterol. Oxysterols showing the weakest condensation effect
are 27-hydroxycholesterol, 25-hydroxycholesterol, and 25S-cholestenoic acid.

����
Figure 5 shows the average thickness calculated from the MD trajectories as an average inter-
leaflet head-to-head (i.e., phosphorus-phosphorus) distance of the bilayers. Figure 5 reveals that
all sterols thicken the POPC bilayer but the strength of this effect depends on the type of the
sterol. Indeed, as compared to cholesterol, oxysterols with the additional oxygen-containing
functional group localized in the tail (27-hydroxycholesterol, 25-hydroxycholesterol, and 25S-
cholestenoic acid, see also Figure 1) have the strongest effect on the membrane thickness, while
7-ketocholesterol and 4�-hydroxycholesterol have almost the same effect as cholesterol.
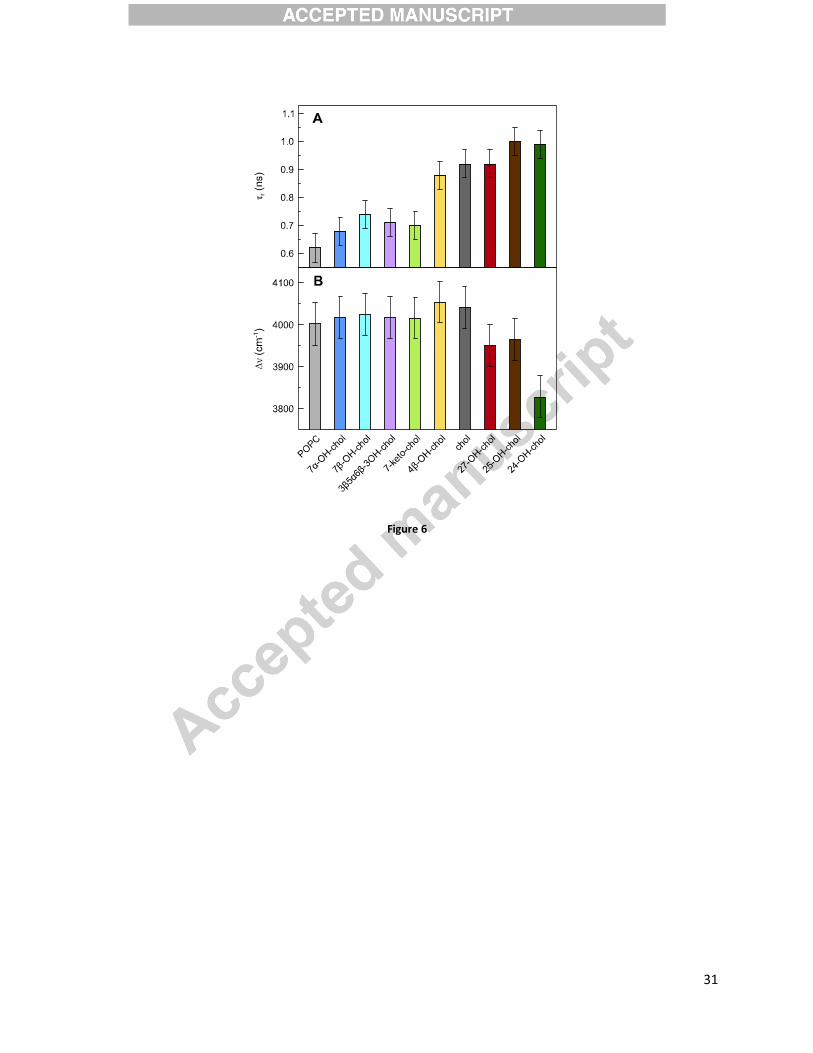
Sterols slow down lipids’ mobility
TDFS was used to obtain two physical parameters describing i) local lipid mobility,
corresponding to the integrated relaxation time �r, and ii) local polarity, corresponding to the total
spectral shift ��. Laurdan probe, which has a well-defined location in the bilayer at the carbonyl
level of lipid molecules, was used to probe the studied systems.[83] As shown in Figure 6A,
addition of 10 mol% sterol prolongs the relaxation probed by Laurdan. The difference in r value
between POPC and 24-OH-chol samples is as large as ~40 %. Furthermore, oxidation of
cholesterol causes different effects depending on the position of the additional oxygen atom.
Ring-oxidized sterols (light color bars in Figure 6A) significantly increase the lipid mobility at
the glycerol level, except for 4�-OH-chol, for which r is comparable to that of cholesterol. Tail-
oxidized sterols (dark color bars in Figure 6A) do not change (27-OH-chol) or even slightly
increase (24-OH-chol and 25-OH-chol) the obtained �r values. It is important to mention here
that for tail-oxidized sterols the relaxation process, characterized by the time �r, is not completed
within the experimental time window (see below the discussion of �� values). This lowers the
accuracy of TDFS parameters obtained for the tail-oxidized sterols.
It was shown that �r probed using Laurdan is usually inversely proportional to APM.[50] This is
because the lateral expansion either homogenously increases lipid mobility or creates voids in
which the probe can reorient more freely. This relation between APM and �r holds for the action
of sterols in comparison to pure POPC – the presence of a sterol compresses bilayer laterally,
which also hinders local mobility at POPC carbonyls. Oxidation of cholesterol hampers its
condensing function, which is observed as an increase of APM for all oxysterol samples in

����
comparison with cholesterol. This corresponds to the increased carbonyl mobility, for the ring-
oxidized sterols. For tail-oxysterols an increased mobility is not detected, which can be caused
by the problems discussed above.
The total spectral shift of Laurdan is proportional to the local polarity at POPC carbonyls.[50]
Figure 6B shows that substitution of 10 mol% of POPC molecules with cholesterol or ring-
oxidized sterols does not change it. Significantly lower �� values were obtained only for the tail-
oxidized sterols. In this respect it is important to mention the results of the inspection of the
Time-Resolved Emission Spectra. The time profile of their Full-Width at Half-Maximum,
FWHM(t), is usually very characteristic. Namely, there is a well-pronounced single maximum of
FWHM(t) at the time t corresponding to the mean relaxation time. This is what was observed for
most of the samples discussed herein (Figure S1). In the case of tail-oxidized sterols, however, a
secondary increase of FWHM(t) was observed at about 10-15 ns after electronic excitation. This
indicates the presence of additional slow relaxation process that is not well captured in our
experiment due to limited fluorescence lifetime of Laurdan (~4 ns). Therefore, the obtained ��
values are falsely lowered and we cannot properly estimate their uncertainties.
Sterols’ orientation
To elucidate the sterol orientation in the lipid bilayer, we calculated the tilt angle of the sterol
rings. Figure 7 shows the tilt angle distributions for all the bilayers taken into consideration in
this study. Sterols, where the oxidation is in the ring, are more tilted in the POPC bilayer than
cholesterol. Their tilt angle distributions are wider, with the maxima shifted towards larger
angles (Figure 7A). Tail-oxidized sterols have their tilt angle distributions rather narrow, with the
maxima below 20° (Figure 7B). Interestingly, the tail-oxidized sterols can adopt to a nearly
perpendicular orientation with respect to bilayer normal in the lipid bilayer (see the non-
vanishing tails of the distributions for larger angles in Figure 7B), while ring-oxidized ones do
not show this feature. This suggests that the position of the oxidation leads to distinct oxysterol
behavior, resulting in different properties of the corresponding lipid bilayers.
Figure 8 shows the partial mass density profiles of the hydroxyl group containing an oxygen
atom (see Figure 1; in this manuscript referred to as the cholesterol hydroxyl group). Mass

����
density profiles of different systems were shifted such that the distributions of phosphate groups
overlap, which allows one to compare the depth of the penetration into the bilayer by the sterols.
The profiles indicate that all oxysterols tend to be localized such that the cholesterol hydroxyl
group is pointing out into the aqueous interface (see the maximum of the distribution around 1.5
nm from the bilayer center). This arrangement is very similar to that in cholesterol-containing
bilayers. Nevertheless, in the case of tail-oxidized sterols additional maxima in partial mass
density profiles are observed. These maxima correspond to the interior of the bilayer (0.8 and 0
nm from the center of the bilayer) and indicate that the cholesterol hydroxyl group may also be
located inside the lipid bilayer, as described in detail below.
To better quantify the positioning of different sterol molecules inside a POPC membrane, we
constructed from MD trajectories the 2D probability distributions P(�,h) for the sterol
orientations and positions. Figure 9A compares the distributions for cholesterol and 27-
hydroxycholesterol. The positioning of the two sterols is strikingly different. Indeed, whereas
cholesterol inserts into POPC membrane in a more upright orientation (seen as the location of the
dark shades on Fig. 9A), 27-hydroxycholesterol is accommodated inside the POPC bilayer in
multiple ways: i) in an upright position (see Fig. 9B), similar to cholesterol (region on Fig. 9A
corresponding to small � and large h values); this orientation is the most frequent and common to
all oxysterols; ii) in a lying-down positioning (see Fig. 9C) close to lipid head-group region
(region of intermediate h values, between 12 and 16Å, and high � values on Fig. 9A); iii) in
interleaflet orientation (see Fig. 9D) in which the sterol molecule is located in-between two
leaflets of the bilayer such that both hydroxyl groups point out into opposite water interfaces
thinning the bilayer at the same time (on Fig. 9A in the region of small h and � values); iv) in
flipped orientation (see Fig. 9E), whereby its main hydroxyl group is located near the membrane
midplane (strong density on Fig. 9A in the region of small h and � values). Interestingly, there is
non-zero density for small h and high � values which might be interpreted as a heavily tilted
flipped orientation similar to the one showed in Fig 9E. Last three orientations (lying-down,
interleaflet and flipped) have been observed only in tail-oxidized sterol systems.
These unusual orientations of tail-oxidized sterols are responsible for additional maxima in the
cholesterol hydroxyl group mass density profiles (Figure 8). For example, the interleaflet

����
orientation gives rise to the maximum centered at 0.8 nm from the bilayer center, while the
reversed orientation is responsible for the maximum localized around the center of the bilayer.
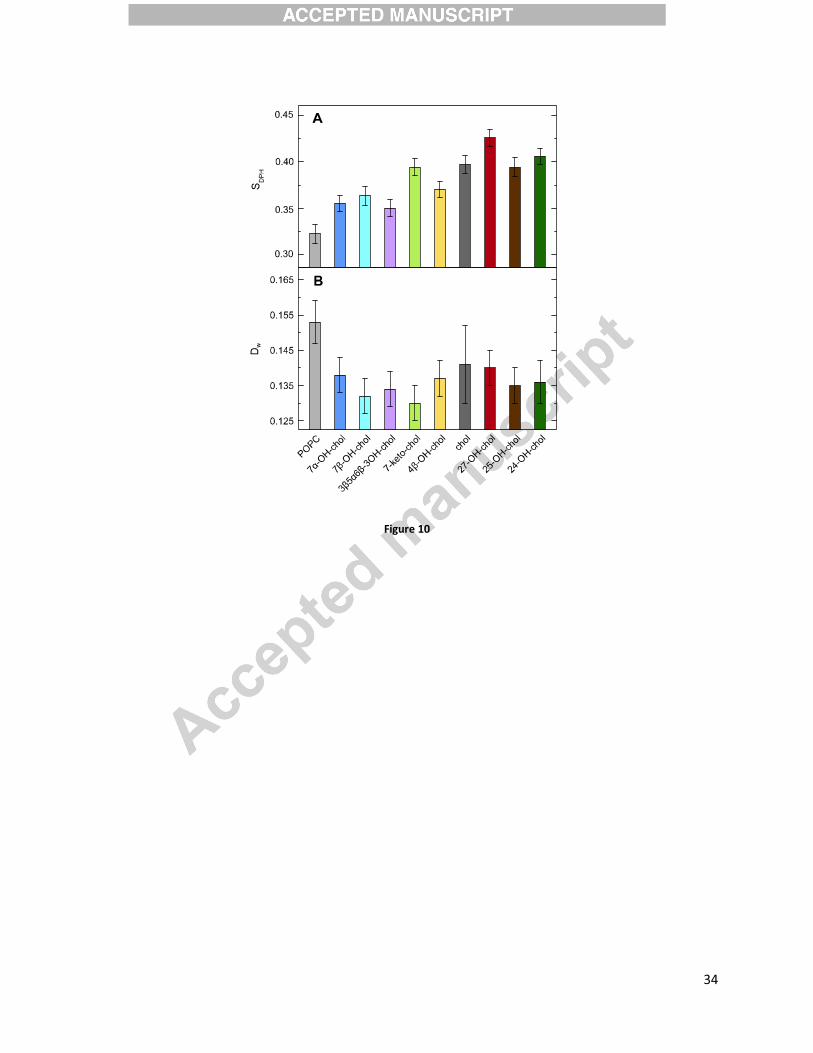
Ordering effect
Time-resolved fluorescence anisotropy of DPH was used to measure the influence of the sterols
(10 mol%) on the order and mobility of the acyl chain region of a POPC bilayer. The DPH order
parameter SDPH (Figure 10A), which gives a simple estimate of the POPC acyl chain order, is
increased in all the sterol-containing systems. On average, the ring-oxidized sterols (light color
bars in Figure 10A) order the hydrophobic core of the POPC bilayer less than cholesterol does.
From this group only 7-keto-chol has similar SDPH values compared to those of cholesterol.
Oxidation of the cholesterol tail (dark color bars in Figure 10A) does not change the order of the
acyl chain region, or even slightly increases it for 27-OH-chol. The DPH wobbling diffusion
coefficients (Figure 10B) are slightly lowered for all the sterols, suggesting that their presence
not only orders POPC chains, but also slows down rotational mobility of the DPH probe. This
rather typical dependence between SDPH and Dw parameters becomes, however, more
complicated when oxysterols and cholesterol are compared to each other. Surprisingly, ring-
oxidized sterols order the hydrophobic core of POPC bilayer less than cholesterol does, but slow
down rotational diffusion of DPH slightly more compared to cholesterol. This discrepancy
between the order and dynamics of DPH rotation might originate either from the different
geometrical restrains the ring-oxidized sterols impose (i.e. their lying-down positioning depicted
in Fig. 9C) or by specific interactions between the oxysterols and DPH probe.
We also measured deuterium NMR spectra of the POPC bilayer containing 10 mol% cholesterol,
24S-hydroxycholesterol, 3�,5�,6�-trihydroxycholestane, as well as of the pure POPC bilayer.
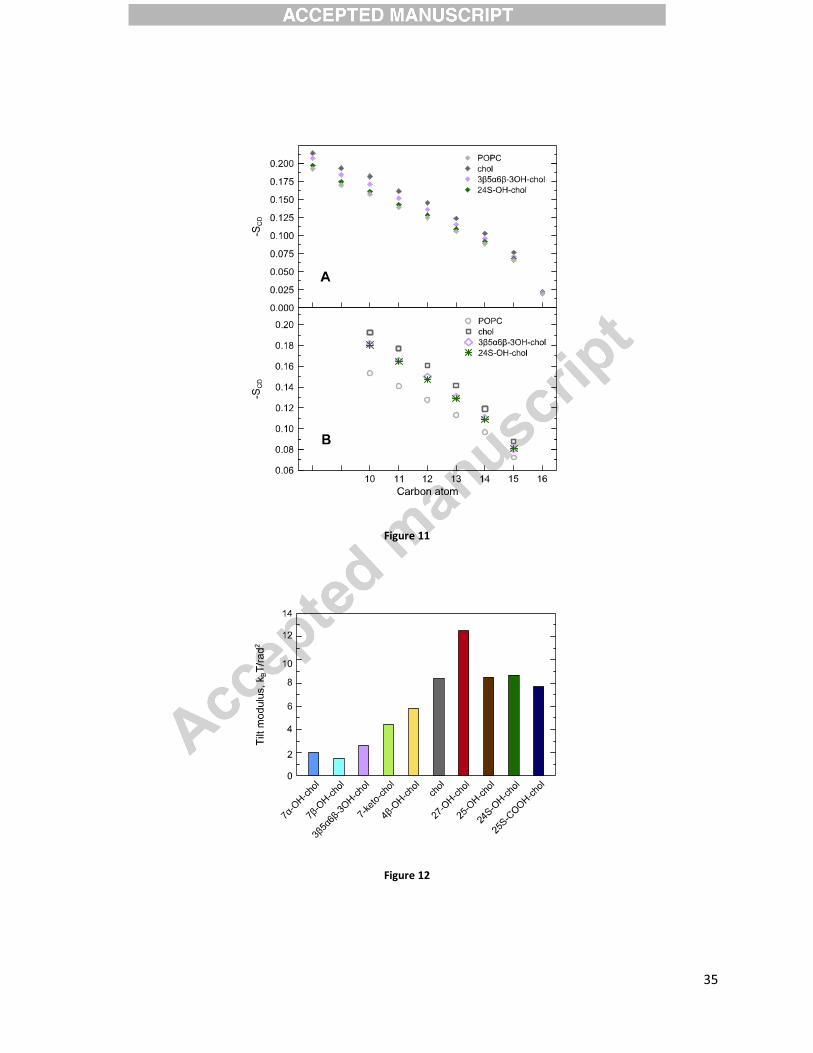
Figure 11A presents experimental values of the deuterium order parameters measured at 310 K.
In the analysis it is assumed that the end of the sn-1 chain (carbons 10-16) exhibits smaller
orientation than the parts of the chain closer to the head group and that the order decreases
monotonically toward the terminal methyl group. This behavior has been previously confirmed
by 1H-13C solid state NMR experiments for pure POPC and for POPC with low concentrations of
cholesterol.[84] The two points in Figure 11A not identified with any particular carbon
correspond to average orientations obtained from the two outermost broader peaks in the

����
spectrum. These signals arise from deuterons attached to carbons 2-9, which cannot be
unambiguously assigned from the deuterium NMR experiments. Experimental values of
deuterium order parameters measured for carbons 10-16 in the sn-1 chain of POPC (see Figure
1) agree well with the theoretical values obtained from MD simulations (presented in Figure
11B) as well as with experimental DPH measurements. Both theory and experiment show that
cholesterol orders the POPC bilayer the most (as compared to the pure POPC bilayer) followed
by 3�,5�,6�-trihydroxycholestane and 24S-hydroxycholesterol.
Order of the lipid chains, surface area per lipid, membrane thickness, and cholesterol tilt are
interconnected parameters. In most cases the increase of area per lipid is associated with
decrease of membrane thickness and decrease of order parameter. Smaller cholesterol tilt was
shown to induce higher ordering and condensing effect[81, 85] and could be seen as a predictive
parameter for strength of the membrane modification due to presence of a particular sterol
type.[86] It is worth noticing that in the case of sterols with modifications in the chain region, the
above-described dependencies between parameters are not followed. Area per lipid in these
sterols is generally larger than in the case of sterols with a modification in the ring, which
corresponds to lower thickness. However, order of the lipid chains is similar or larger to both
cholesterol and ring-oxidized sterols, corresponding to a smaller sterol tilt. This might be
explained by the tendency of oxysterol with a modified tail to link opposite interfaces (see Fig. 9
D), inducing local thinning of the bilayer.
Energetics of sterol tilt
To quantify the energetics of tilting different sterols in POPC membranes, we used the tilt angle
distributions showed in Figure 7 to calculate the tilt modulus, , using a previously established
protocol (see the Material and Methods section and references therein). Specifically, in the low
tilt angle limit, quantifies the free energy cost of tilting a sterol molecule by a unit angle with
respect to the membrane normal. Figure 12 shows for all the studied sterols, revealing that the
tilt modulus for sterols that are oxidized at their tails is relatively large, and comparable to that of
cholesterol. In contrast, for the sterols with oxidized rings is significantly lower (compare, e.g.,
for cholesterol of ~4 kcal/mol vs. ~1 kcal/mol for 7�-hydroxycholesterol). This result suggests
that the energy cost of tilting sterols oxidized at their tails is significantly higher than in sterols

����
where oxidization takes place in their rings. This trend is directly related to the differences in
sterol orientation in the membranes. The ring oxidized sterols significantly broaden the P(�)
distribution (Figure 7), suggesting that their fluctuations are higher, hence implying that tilting
these sterols comes at a lower free energy cost. We note that a similar effect is observed with
other sterol ring modifications, such as the effect of bond saturation on sterol tilt (see e.g.
differences between tilts of cholesterol and 7-dehydrocholesterol in Ref. [87]). The tail oxidized
sterols show a very different P(�) distribution. Specifically, most of these sterols show a bimodal
tilt distribution. Since it represents the low order quadratic term in the tilt energy, the tilt
modulus reports exclusively on the low-� regime, for which the tail oxidized sterols do not show
a large difference from cholesterol tilting. The second lobe at higher angles for these sterols is
thus not captured by the tilt modulus.
DISCUSSION
Oxysterols are oxidative derivatives of cholesterol playing a crucial role in many regulatory
processes in the human body. They are produced naturally from cholesterol during an enzymatic
side-chain hydroxylation (catalyzed by cytochrome P450) or non-enzymatic oxidation (upon
direct reactive oxygen species attack on cholesterol molecule). Biological oxysterols fall into two
main categories; ring- and tail-oxidized. Generally, ring-oxidized sterols are formed non-
enzymatically, whereas tail-oxidized sterols usually have an enzymatic provenience. However,
there are exceptions to this rule; for example 4�-hydroxycholesterol is produced in an enzymatic
way, while 25-hydroxycholesterol and 7�-hydroxycholesterol can be produced through both
enzymatic and non-enzymatic pathways.[88] In general, there are significant biological and
chemical differences between ring- and tail-oxidized oxysterols. For example, tail-oxidized
oxysterols act actively as liver X receptor and Insig ligands as well as in HMG-CoA reductase
(i.e., a rate-controlling enzyme in the metabolic pathway that produces cholesterol and other
isoprenoids) degradation, while ring-oxidized oxysterols do not participate in these processes or
their role is very limited.[88, 89]
Oxysterols can specifically influence physiological processes either directly, by modifying
interactions between oxysterol-sensing proteins and other proteins (e.g. ABC transporters[90]) or

���
lipids, or by regulating gene expression. Both of these mechanisms, however, require oxysterol-
protein interactions that were not addressed in this study. For instance oxysterols were shown to
increase penetration of the amyloid beta (A�) peptide into a lipid bilayer.[91] Oxysterols can also
regulate cell function by modification of physical properties of lipid membranes, as investigated
in the present work. Oxidation of membrane cholesterol or introduction of oxysterols from other
sources is likely to change the structure and the dynamics of a lipid bilayer. This can result in a
number of biologically important effects. First, the mechanical properties of the lipid bilayer are
to large extent governed by the sterols and their interactions with lipids. As we have shown,
oxysterol presence can tune lipid order in a manner that depends on the oxysterol orientation.
The ring-oxidized sterols that can adopt tilted conformations disturb the membrane structure
more significantly in comparison with cholesterol and tail-oxidized sterols. As a result, they
decrease sterol tilt modulus, increase mobility of lipid carbonyls, and decrease order probed by
DPH.
Cholesterol is also known to play an important role in lateral membrane organization. It is well
justified to hypothesize that the above mentioned physical changes caused by the ring-oxidized
sterols will also influence the size and existence of lipid rafts. This, in turn, may disturb the
function of the signaling proteins associated with rafts. Finally, cholesterol strongly suppresses
permeability of membranes to water, oxygen, ions, and other small solutes. The observed fast
reorientation of the tail-oxidized sterols is a potential breach of this permeability barrier. Similar
to truncated oxidized lipids, tail-oxidized sterols might also reduce lipid membrane asymmetry
by lowering the energetic barrier for the flip-flop of phospholipids. These later hypotheses need
to be tested.
CONCLUSIONS
Using several experimental techniques, including dynamic light scattering, fluorescence
spectroscopy, and NMR, together with atomistic molecular dynamics simulations and a
quantitative description of molecular tilt, we have characterized the behavior of a family of
oxysterols in phospholipid membranes and compared the resulting data to that of cholesterol. We
found that the two distinct groups of oxysterols, i.e., ring-oxidized sterols (mostly produced by

���
free radicals) and tail-oxidized sterols (mostly produced enzymatically) behave differently in a
lipid membrane, influencing it in different ways. Unlike tail-oxidized sterols or cholesterol, ring-
oxidized sterols can efficiently acquire tilted orientations in the bilayer leading to a stronger
disruption of the membrane structure. These changes in physical properties of the bilayers may
have implications for membrane biochemistry. Namely, tail-oxidized sterols behave similarly to
cholesterol in terms of membrane stiffening, indicating that their effect on biochemical processes
in membranes will be similar to that of cholesterol. In contrast, ring-oxidized sterols exhibit
behavior distinct from cholesterol, implying potential disruptions in membrane functionalities
including signaling processes. The presence of oxysterols in biological membranes may also
affect protein-lipid and protein-sterol interactions, but this cannot be clarified by the results of
this work, which does not contain protein explicitly.
ACKNOWLEDGEMENTS
We thank the Academy of Finland for financial support (the Finland Distinguished Professor
(FiDiPro, grant number 263410) program (WK, PJ), and Center of Excellence (WK, MM, MP,
TR, IV) funding (grant number 272130). IV thanks the European Research Council (Advanced
Grant CROWDED-PRO-LIPIDS). AO, PJ, MH, LC, PJ thank the Czech Science Foundation
(grant P208/12/G016). MV thanks the Croatian Academy of Sciences and Arts for financial
support. CSC — IT Centre for Science (Espoo, Finland) is acknowledged for excellent
computational resources (project number tty3995). AMK wishes to acknowledge the Academy
of Finland for funding. PJ acknowledges the Academy of Sciences of the Czech Republic for the
Praemium Academiae award.

����
REFERENCES
�� � ������������������������������������� ���!���"����#��� $�%&��'���!�(&���&�����)�*�+���(���,��&&���$�(���%�*���&-��&���.�$�%+��&(�+*-��&-��(�$+*���*�������*�%�**���/�&��*�&����(�����"����*�&(���*����"�&(���*���$�*���&(���*0� �&%�**1��"*�����(����&��((����"�&(�$+�������������������� ������������������� �������1����������� � ������2������*&�����������3�*(������4-&�����������)*��%������5��,��%�����������������(&����6��7��8��%�*���&(���*�(�����*�&(���*0�#(���(���*��/�*�(������$�$�..����(��*��..��(&����*�-�$�9�*�+��&�������� �������������������1�������������� � :���:��2���)�((%����;���<�-��(��*��7������&&*����<���=�*���>���'���2��$����?��?..��(��.�(���&(���(�����.���(���*�&(���*&���$�&-���"�*�-�$&����(���.��%�(�����.���$���$�&-���"�*�-�$@&(���*�$�%���&�A��.(&B����������������������������� ������������1��������������� � ;�"��������&��,��3��>17����*�������=�((�*�������4���!��((���������6�$����"��..��(&��.����*�&(���*���$��(&����*�"��&�������������������� ������������������� �������1������������ � ;C"������=�((�*�������4��'��*�&(���*��&-���"�*�-�$&����$�"*+��*�-�$&0�D��(�$��3��,��3��9��(�(�������*�������.(1*�,��%�%9����&E������ ���������� �� ��������� ������1������������� � ?-��$��;�����'��*�&(���*���$�(�����(����(�����.�-��(���&�3�(��%�%9�����$�%���&�������� ������������ ����������1������������ � !�3�9��������?���2�9���&&����������&(���*1&��&��"�$�%���0�%�*(�-*��.�%�*��&�������F�����*�E������ ���������� ������1������������ � 4����$������������?�(����#��'��*�&(���*����&�"��*�(���&$��(������������ ����������������������������1������������ � ?$�$�����������&(�(���.�*�-�$���.(&0�8��%�%�$�*�%�%9����&�(����**&�����������!��"���������� �� ������������������#������������1������������� � '����"�����G��2��;�"�*�(�����.�9�*�����$�&+�(��&�&0�-�(�3�+&�����*��������-(��&����$�%������&%&�������������$��������������1�������������� � #�%��&��!���4,������?��'�**�9��*�"+�1���3���**&����$*�����*�&(���*��#���������������1��������������� � =�������#������&��������!��/�&�*�������;�"������!��((����������=�((�*�������4���4,������?��#�"��.��������.�&(���*�&(���(���*�&-���.���(+�1�<�&%�&(���*������(���-*�������*�&(���*����*�-�$���.(&����������������������������� ���������1�������������� � ;�"��������&��,��3��>17����*�������=�((�*�������4���!��((���������D��(���--��&��.����*�&(���*��&�%�$��&%��(���0�4%-��(������.�%�(�+*�&�9&(�(���(&�������*�&(���*����"�&(���(�������-��&-��(�$+*���*���1&(���*���(����(����������� ��������������������1��������������� � ��/�((�����)��6H+&(���*&0���/�*�9��*�"�����*�&�.���(�����&(����(��+��#����� �����1������������� � ��*���7���)��&���8���2�����$�>>���7��6H+&(���*&����(���-�(��"���&�&��.�%�I�����������$�&��&�&������%��������������1�������������� � )��3��� ��������&&�-��D��6H+&(���*&���$��(����&�*���&�&������� ����� � ������1��������� � 6*,,������=������2��(������6H+&(���*&���$��H+&(���*�9��$��"�-��(���&0���*�����*�-�$�%�(�9�*�&%���$��(����&�*���&�&������� ����&�������������1������������� � #�����-.����7�����6H+&(���*&0���$�*�(��&��.����*�&(���*�%�(�9�*�&%���$��(����-����&&�&����� ������������!��" �������1������������� � )I��,��%��4��8�/��$���$�&�3�(���H+&(���*&����������������1�������������� � #��/�$$����7���)�**��(���8���=��$�%��*���7��#�1��1�6H+&(���*&����(��������&(����.�*�/�����**�%�(�9�*�&%��'������������������������&���������� ���������� ��#���������

����
��� � )I��,��%��4�� ���&�$�1�������H�$�>�$��H+&(���*&���"�*�(��&��*&�����/�/�E�������������������� ��������#���1#������������ � ���(���>���������"�*&$��.��<�����2�/���:�����-(���&�"��*��"�-�(�3�+&�������$��/�&��*���$�&��&���&��������������������������1������������ � 5�*������������(���>�����2�/���:�����-(��&��&���(�"��(��&��.�%�(�9�*�����$���.*�%%�(��+�&�"��*��"����������������������(�!� ��������������1�������������� � ?���$"���'���D�99��<������#-��,�((��'�������1�+$��H+���*�&(���*���91�+$��H+���*�&(���*���$��1,�(����*�&(���*��-��"�*�(����(��*��,��1��H-��&&������$�-��$��(*+��.���**1*�,������-(����������������&�"��**��"������%���%����-��"�&��'�������������� ������������1�������������� � 2�%����1?3��"��#��������(��'������(��"�������=�I�H�� ���)��(������ ���)�&&�$���7���'����&��2���7�%9��(���������*��<���2�>��$��7��'�%-���&����.�(����+(�(�H����-��1�H�$��(���$�-��1��.*�%%�(��+�������(���&(��&��.�$�..����(��H+&(���*&���������������������%������������1�������������� � <�#�**��'��<���J%�(���������#���*�����D������"�*&$��.��<��������<����**��<�������1�+$��H+���*�&(���*��&������$�"����&�&�*��(�/���&(��"�������-(���%�$�*�(����&���������������������������1������������ � J%�(���������<�%�(�������7��%*�+�� ��!���G��������4��#���'�%%��&��'��2�����/�((�����)���!�������!��#���#���*�����D������"�*&$��.��<�������1�+$��H+���*�&(���*��&������$�"����&�#?;��(��(�����9�(&�(������$��/�&��*����..��(&��.��&(��"����)�����&�������������1������������� � J%�(���������#���*�����D����1�+$��H+���*�&(���*0�(���.��&(��$��(�.��$���$�"����&�#?;�������� ���������������������&������ ��������1������������� � 2����'��#������,��D�����������?��#���)��"�����<�..����(��*�%�$�*�(�����.��1,�(����*�&(���*�(�H���(+��"���&(��'�����**&�9+���*%�$�*�����(�"���&(&���$�'��K�������*�9*��,��&��)��������������� ���������1����������� � 2�(�����#���<��"������7��%�I��� ��2���'�3�������!�%��<��D������,����� ���!�--��%�����)��<���!����+�����'���1!�(����*�&(���*���(�/�(�&���&-�&�&1�@��1����$1��������%���%����/�&��*�����$�(��*��*���**&����/�(����&����!� �������� �����������1������������� � 2���D���2�&,���� ���#�*(���������6&%����?���7��&�������2���L���G�����:�1���'�**�$��(����$���$�9+��1�H+&(���*&�/���*+&�&�%�*���$�%�(�����$���*�-�(�3�+&��&�-��1$�-��$��(��'������������������������&�������������1��������������� � *,�>�%���<���?"�*��$��7���=�+���������*(>����#���!�9�3��#��6H+&(���*��&������,����.� (����"�����<+&*�-�$�%������ $�*�&����������������������������������������*�&������ �������1������������ � ���$�*���7���)��(�*�����<������(���*���'�������$$�������4�*������2���7�**���8��2�-�,���&���$��H+&(���*&0���/�*��$�-�&�1$���/�$�*�-�$����%���&�*��,��"��$�-�&��$+&.���(������$���&�*�����&�&(������'������������������������&������������1������������� � 2������=��6H+&(���*&��&�%��,��&��.������*�"���*�$�&��&��1�����/��3��#�������!������������������������*�����������(�!� ������������1������������ � !�*&��������2�(I�������<���/���)��"%�����!���������;��������*���.���#1�+$��H+���*�&(���*���� *>���%��M&�$�&��&�������+�����������������,�������*�����������1������������� � 7�&&�*�(��8���#���(1��*������8����(��2��?..��(&��.��H+&(���*&����(���9*��$19�����9������0�4%-*���(���&�.��� *>���%��M&�$�&��&������������������������� ������� ������������������ �������1������������� � 2�����������G�����2���:���#������D��"��!���5���"�����2��'��*�&(���1��9�(�����*-�����9�(�1(���*�����9�(&��&(��9*�&(���$�..����(��(������$�-��%�(�&��-�-(�&�&��.���(�9����%����3�&(��%�*���**&������������������������������ �������1������������ � ��*/�((���2������(��"���������..����'���=�I�H�� ������>���'���)����� ���'���>�(17��������'���2�>��$��7�� ��*�"��&�9�(3�����(����&�*���&�&���$��"�1��*�(�$�%���*�-�(�+0��H-��(�$���*�&��.��H+&(���*&����������'������ �-� ������������������1����������

����
�� � ;�$��"��>��4��;���2����+�>��4�����'��*�&(���*��H�$�(�������(�����(���0��%-*���(���&��.��!'��.��%�(�����������������.*�%%�(������$��"�1��*�(�$�%���*���$�"�����(������������������������ �����������1�������������� � =�I�H�� ���2�>��$��7��'+(�(�H����..��(&��.��H+&(���*&��&&����(�$�3�(����%���$�&��&�&0�4�$��(�����.���**�$��(��A�-�-(�&�&���$@�������&�&B���H�$�(�/����$���.*�%%�(��+���(�/�(��&����$�-��&-��*�-�$�&�&��&���������� ��� ����&�������������1������������� � !���$���5���:���2���#��*(����;������(������� ��)��*�"���*���(�/�(��&��.��1$��+$�����*�&(���*1$���/�$��H+&(���*&0��%-*���(���&�.���#%�(�12�%*�16-�(>�&+�$��%���������������������� �����������1�������������� � 7�*���#��?���D�&(�/����?������<�$*�+������!��&������!������*����#���#���������<��?��������:��2���5�����:������)���,%�������2���)��3���;��?���'�/�+��<��8���#���..�������?���#��*�&��"��������6�+��<��#��#�$��'�����6H+"���(�$�'��*�&(���*�;�"�*�(�&�'�**�*���'��*�&(���*���%��&(�&�&�(����"��<����(�#(���*1��%9�����4�(����(���&����������������������������� �����������1�������������� � ���(>����?���'���*�&��7���7��$����#��4�(����(�����.�(3���H+&(���*&���1,�(����*�&(���*���$���1�+$��H+���*�&(���*��3�(��-��&-��(�$+*���*������$�&-���"�%+�*������%�$�*�%�%9����&������� ���������� �� ��������� ������1������������� � 6*&����)������#��*�&��"�����������)�,������� �����(��9�(���&��.���%9�����#(���(����9+�'��*�&(���*���$�'��*�&(���*�<���/�(�/�&� ���<�(��%���$�9+�#(���*�6����(�(�������������������������������������#������������1������������� � 6*&����)������#��*�&��"��������)�,������� �����(��9�(�����.���%9�����#(���(����9+�6H+&(���*&�������� ������������������ 1�� ����������� � 6*&����)������#��*�&��"�����������6�+��<��#���)�,������� ����1�+$��H+���*�&(���*�4�����&�&�(��� /��*�9�*�(+��.�'��*�&(���*�������&-��*�-�$���%9����&�������� ������������������1������������� � ���*���G��<�����3�����#���'��((�-�$�+�+�� �� ���&-���.����������*������*�&(���*�9��$��"�&�(�&�-��&��(����71-��(�������-*�$�����-(��&E������������������� ������������������� ��������1������������ � 2��"3��$��<���)�����"(����)���;�"������=�((�*�������4���7�>+9�,������'�&,����J���2��"3��$��'�� ���#�%��&��!��'��*�&(���*�%�$�*�(�&�"*+��*�-�$����.��%�(������$�����-(�����(�/�(+��)���������������������������1������������� � �������������,��-�$�+�+��'��'��*�&(���*�$��/����*(���(�����.�(������.��%�(������$�$+��%��&��.�-��&-��*�%9������%�$�*�%�%9����&����� ���������� ��������������� �� ������1�������������� � ���,��3��>������'3�,*�,��2������"3��(���������.�����2�-�$��+$��(������$�%�9�*�(+0� ����(��-*�+�9�(3����.*����&������&�*/��(���*�H�(�����H-���%��(&���$�%�*���*���$+��%��&�&�%�*�(���&�����������������1������������� � !�*�"��D������,��3��>������6*N+O&,��� ����+�,,+�����������/������������������������;�"��������.������=�((�*�������4������"3��(������?H-���%��(�*�$�(��%���(������$���%-�(�(����*���(��-��(�(�����.�9��-�+&���*�-��-��(��&��.�*�-�$�9�*�+��&��������$�9+����*�&(��+*���%�&������(���������������������� ��������.���/�������������� ���������1�������������� � ����"�����2���7��$��,������ �����-�>+���� ����������**������#�9-���&����$����&���%��(&��.���*���#�*/�(����<+��%��&�1�'��%����1����;�/�&�(�$���������������� ���������� �����������1�������������� � 8����;��#����������**������?&(�%�(��"�(�����%�15����#-��(��%������%�1;�&�*/�$�?%�&&�������&���%��(&��.�#�*/�(����<+��%��&�������������� �� �������1������������ � ���,��3��>������#+,���������6*>+�&,��� �����%-*��,�/���������.�����#�*/��(���*�H�(�������-��&-��*�-�$�9�*�+��&0�������-*�&���$������(��--*���(���&�������������'����� ���������1���������

����
��� � %���������P���*��;������,��3��>������'��(������ �������(���������.�������%�1;�&�*/�$�8*����&���������2�-�$�)�*�+��&0�#�*��(�$� --*���(���&���$� $/��(�"�&��/���#(��$+�#(�(��������� ���������������������1��������������� � #����(>,+������)������*>��G��82J4<4�G�� ; �?�?;#�68�24�4<�;?746�#�<?�?;�4�?<�)G�82J6;?#'?�'?��62 ;45 �46��������������������� �������������1����������� � !�3�(���#���!���&�(���!���4,�"�%��� ��<+��%���#(���(�����.�2�-�$�)�*�+��&�#(�$��$�9+�����&����$�8*����&������������F��&���������� ���������1������������� � !���&�(���!���4,�"�%��� ���!�3�(���#��6��(���D�99*��"1��1'���� ��*+&�&��.�8*����&������ ��&�(��-+�<���+�������� ������������������1����������� � �Q���"��8��#(���(���*�6�$����.�2�-�$&���$����(���&������%9����&�1�?/�*��(�����.�8*����&������ ��&�(��-+�<�(������������� �������)������������������#������ �������0�����#�� �������������������1������������� � ��+���������<�(��%���(�����.�*�-�$�6�$�������%�(��&���$�;�(�(����*�'����*�(������%�&�.��%�8*����&������<�-�*���>�(����?H-���%��(&��'�#����� �������1������������ � 2�-�����7���#>�9��� ��?88?'��68�24); �46� 2��6�46��6��82J6;?#'?�'?�<?�62 ;45 �46�� �<��J'2? ;�� 7�?�4'1;?#6� �'?�;?2 : �46��4��� ';6�62?'J2?#� �<��?�); �?#�������� ����������������1������������ � )����((��2��������**����)�����<��(�����L��$��-�*��'��-*��"�'��&(��(&����������#�*�$�<��(���(�$�����..����+$�����9��&0�'�<���'�<����'�<������������������������������� �� ����1������������� � ��'�9������ ���D�&&�**��#��;��;�-�$�$����/�*�(�����.���;�-�3$���&-��(���9+�3��"�(�$�.�&(�8�������(���&.��%�(�����#�����#���)�������&��������� ������������1����������� � #�������1 ���D�9����<��!���<�*�"*����8���#�-���/����8���7��%�������<�� �-���(���*��%-*�%��(�(�����.�$�1��,�1��"�/���3��"�(�$�8�������(���&.��%�(����������������1�������������� � <�*�"*����8���7�>�&��,��#���=��&(����7��D���5����7����.��.��������)�H�� ����;�4�?�1� ��J2�4<4�?�#46� 2�#�?'�; 2��;6'?##4�7�#G#�?��) #?<�6��J�4:��4�?#��������������������������)��������1����������� � ���"��&����D��2�������$���/�&���������6�2#���(��(��*�8���(���&�.������(���&�1�?���"+�����%�>�(���&�.���'�+&(�*&��.�'+�*��1��-(�$�&���$�'��%9������������������������������������#��������������1������������ � '���$��&�,���������#���$��&���������"��&����D��2��?..�����(��H-*���(�����.����.��%�(����*�&-�����&��"�(���&(����&(���&������%�(��$0� --*���(����(��9�(�1-�-(�$���*�"�%��&������������������������������ ����������1�������������� � �����I�3&,��� �����&��,��3��>17����*�������'��%�������6���=�((�*�������4���;�"�����;�.���$�6�2#� **1 (�%�8�����8��*$�.���#�(���(�$����&-��(�$+*���*����)�*�+��&��(�8�**��+$��(����������������������� ���������� ��1�����������1������������� � ���"��&����D��2���'���$��&�,�����������$��������<���4%-�+��;��D���!*��������2��'6�� ;4#6��68�#4��2?��6�?��4 2�8J�'�46�#�86;�#4�J2 �4�7�24LJ4<�D �?;������������������������� �� ������1����������� � ��&���#�� ���*���*��1<+��%��&���(��$�.���#�%�*�(���&����(���'�������*�?�&�%9*���&������������ �� ������1����������� � ���/����D��7��' �6�4' 2�<G� �4'#�1�?LJ424);4J���� #?1#� '?�<4#�;4)J�46�#����� �������!��"��������1������������ � �������**�������;��%���� ����*+%��-��������&�(���&����#��"*�1'�+&(�*&�1�����3���*���*��1<+��%��&���(��$������������������������ �� ������1������������ � ��&���#���!*��������2��'��&(��(����&&������*���*��1<+��%��&�.�����*���*��1#+&(�%&��&������������ �� �������1����������

����
��� � ?&&%�����J�����������2���)��,�3�(>�����2���<��$��������2����������$��&����2��7�� �#%��(�����(��*����&��?3�*$���(��$������������������������� �� �������1����������� � ��&&��)���)�,,��������)����$&����������'���8����I������7��?�����24�'#0� �*���������&(����(�&�*/���.���%�*���*���&�%�*�(���&������������������������������ ����������1������������� � ��+�%�(���#���!�**%������� ��#?��2?�1� �� ��*+(���*�=��&�����.�(���#� !?���$�; ��2?� *"���(�%�.���;�"�$�D�(�����$�*&������������������������������ �������1����������� � 2��$��*��?�����&&��)���/���$���#-��*��<��7;6� '#����0���-��,�"��.���%�*���*���&�%�*�(������$�(��I��(��+����*+&�&�������������&���������&�������������1������������� � D���%��&��8���#����$*�������<�/�*�-%��(��.���%�*(�-���(�%�$�*�.���&�*.������-��(���&0� ���3�-���%�(��>�(����&���%��(����-��$������"�1*�/�*��9����(�����(����(��������"��&������������������������������ ��������1������������ � 8�*�,��?�����(��������!��((�����������+/�������������=�((�*�������4��2�&&��&��.�&*����"�%�%9����&0�4�(��-*�+��.�-��,��"��.������������$�*�(���*�$�..�&�������-��&-��*�-�$@���*�&(���*�9�*�+��&�������� ��������������������1������������� � !��*�&�/�*���7���!�**%�(>����)�����.(9��"����������9&(��7���������&��<��'�*��*�(��"�(���)��$��"���$�*�&�.�����*(���%-����(�2�-�$���%9����&����<�..����(�����%�$+��%������&�&������������������������������������������������1������������� � !��*�&�/�*���7���������&��<����3�&(���*�(�*(���"�*�(�&�-��-��(��&���$���"���>�(�����.�*�-�$�%�%9����&���$�%�%9�������&��(���&������� ���������� �� ��������� ��������1������������� � !����,����������&(��%1��"*���#�����"*������8��#(���(�����.�.�**+��+$��(�$�.*��$�-��&��*�-�$�9�*�+��&�3�(��%�����&�(���(�$������&�������������&����������������������1������������� � ���,��3��>������6*>+�&,��� ���2��"�����������.��������$"���-��+$��(������$�%�9�*�(+��.�<6� �@<6�'�9�*�+��&0� �.*����&������&�*/��(���*�H�(����&(�$+�����������������1������������ � 8����������������'���(�17�%�&��8���6**�*���6�����#������������������=�>��D��2��'�����-"���$��<��'��*�&(���*���$��6�'�&�"%��(�*���$���-���%�(��&����*�-�$�%�%9����&0�&�*�$�&(�(���1�1'1�����;���$��<�&�%�*�(����&(�$��&����� ���������� ��������������� �� ������1����������� � !��*�&�/�*���7���������������5�����7���������&��<���#��((�����2����*���*������"��&��.�9��$��"���"�$�(+����*�-�$&�3�(���&�*�(�$���$����I�"�(�$�$��9*��9��$&0������..��(��.����*�&(���*������� ���������� �� ��������� ������1������������ � �((����%����������%�*Q�����#����+/R������������!��((����������=�((�*�������4��4�&�"�(���(��(�����(�(�/��#-���.���4�(����(���&�9�(3����'��*�&(���*��#-���"�%+�*������$���*%�(�+*16*��+*����&-��(�$+*���*����������� ��������������������1�������������� � !��*�&�/�*���7���;�--�*(������'�����#�1D�����9&(��7���������&��<��4%-��(��.�&(���*�(�*(����%�%9�����9��$��"���"�$�(+�������*�&(���*���$��<�'1���(�����"�<��'�%�%9����&��#���&���������1�������������� � 7�**��#���'��3��;���)��3��� �����#(���*���"�*�(��&��.����*�&(���*���%��&(�&�&���$�9�+��$0������H+&(���*��+-�(��&�&���/�&�(�$���$���/�&�$�������� ������������ �����������1����������� � 6*,,������=�������+�+�����;��4�(����(���&��.��H+&(���*&�3�(��%�%9����&���$�-��(���&��&���������� ��� ����&�������������1������������ � ;��>�����2������8�����$�&��2��;���2�/+��<���)+$*�3&,���#�����4�(����*�(���&��-�9�(3���� ��19��$��"���&&�((��(���&-��(��&���$��H+&(���*&�������������������������������1���������� � ������������������(�����������(�������G�$���������%�$�������=�&(��"���$�����$��'�����,�"�����������..��(��.��H+&(���*&����(�����(����(�����.� *>���%��M&��%+*��$�9�(��3�(��%�$�*�%�%9����&�������������������� ������������������� ���������1�����������
�
�

����
FIGURE CAPTIONS
Figure 1. Chemical structures of 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC),
cholesterol, 7-ketocholesterol, 7�-hydroxycholesterol, 7�-hydroxycholesterol, 4�-
hydroxycholesterol, 3�,5�,6�-trihydroxycholestane, 27-hydroxycholesterol, 24S-
hydroxycholesterol, 25-hydroxycholesterol and 25S-cholestenoic acid. Abbreviations used in this
study are presented in brackets.
Figure 2. Size distributions of extruded liposomes with different bilayer composition: pure
POPC (light grey); POPC with 10 mol% cholesterol (dark grey); POPC with 10 mol% 4�-
hydroxycholesterol (yellow); POPC with 10 mol% 25-hydroxycholesterol (brown) obtained from
DLS measurements. Distributions are normalized (areas equal one).
Figure 3. The time evolution of the area per molecule of the POPC bilayers containing 10 mol%
cholesterol, 7�-hydroxycholesterol, or 27-hydroxycholesterol. The time evolution of other
systems studied here (not shown) is very similar.
Figure 4. Average areas per molecule of the POPC bilayers containing 10 mol% cholesterol
(chol), 7�-hydroxycholesterol (7�-OH-chol), 7�-hydroxycholesterol (7�-OH-chol), 7-
ketocholesterol (7-keto-chol), 4�-hydroxycholesterol (4�-OH-chol), 3�,5�,6�-
trihydroxycholestane (3�,5�,6�-3OH-chol), 27-hydroxychoelsterol (27-OH-chol), 25-
hydroxycholesterol (25-OH-chol), 24S-hydroxycholesterol (24S-OH-chol), or 25S-cholestenoic
acid (25S-COOH-chol). The error of the APM estimation is below 0.6 Å2.
Figure 5. Average membrane thickness of the POPC bilayers containing 10 mol% cholesterol
(chol), 7�-hydroxycholesterol (7�-OH-chol), 7�-hydroxycholesterol (7�-OH-chol), 7-
ketocholesterol (7-keto-chol), 4�-hydroxycholesterol (4�-OH-chol), 3�,5�,6�-
trihydroxycholestane (3�5�6�-3OH-chol), 27-hydroxychoelsterol (27-OH-chol), 25-
hydroxycholesterol (25-OH-chol), 24S-hydroxycholesterol (24S-OH-chol), or 25S-cholestenoic
acid (25S-COOH-chol). The error of the membrane thickness estimation is lower than 0.5 Å.
Figure 6. Time-Dependent Fluorescence Shift parameters: (A) integrated relaxation time �r and
(B) total spectral shift ��; measured for pure POPC and for a POPC bilayer containing 10 mol%

����
sterol LUVs. As a probe, Laurdan located at carbonyl level was used. Measurements were done
at 310K.
Figure 7. The tilt angle distribution of sterol rings with respect to the bilayer normal.
Figure 8. Partial mass density profiles of a hydroxyl group containing the oxygen (O1 in the
simulation model) atom (see Figure 1) along the bilayer normal. Profiles were shifted so that the
distributions of the phosphate groups in phosphatidylcholine headgroups overlap. Membrane
depth equal to zero corresponds to the center of the bilayer.
Figure 9. (A) The normalized two-dimensional (2D) probability distribution, P(�, h), from MD
trajectories, of finding a sterol molecule (chol or 27-OH-chol in left or right panels, respectively)
with a tilt angle � and with cholesterol hydroxyl group at the vertical distance h from the bilayer
midplane. � = 0 describes the sterol tilt where its ring plane is parallel to the bilayer normal, and
h = 0 corresponds to the sterol position in which its OH group is located at the membrane
midplane. (B-E) Snapshots from the MD simulation of the POPC bilayer containing 10 mol%
27-hydroxycholesterol, showing possible orientations of oxysterols in the lipid bilayer: B)
Standard orientation of ring-oxidized sterols and cholesterol; C) Tilted orientation of tail-
oxidized sterols; D) Interleaflet orientation of tail-oxidized sterols; E) Reversed orientation of
tail-oxidized sterols. Lipid tails as well as other oxysterol molecules have been removed from the
figure for clarity. Nitrogen and phosphorus atoms from the lipid headgroups are shown as blue
and tan balls, respectively, to show the thickness of the lipid bilayers.
Figure 10. Time-resolved fluorescence anisotropy of diphenylhexatriene (DPH) embedded in
large unilamellar vesicles (LUVs) composed of pure POPC, and of POPC with 10 mol % sterol.
Measurements were done at 310K. SDPH is the DPH order parameter, while DW id the DPH
wobbling diffusion constant.
Figure 11. (A) Experimental values of the deuterium order parameters measured for the POPC
bilayers containing 10 mol% cholesterol, 24S-hydroxycholeaterol, or 3�,5�,6�-
trihydroxycholestane, as well as for the pure POPC bilayer. The two points in Figure 11A not
identified with any particular carbon correspond to average orientations of deuterons attached to
carbons 2-9, which cannot be unambiguously assigned. (B) Theoretical values of the deuterium

����
order parameters calculated for the POPC bilayers containing 10 mol% cholesterol, 24S-
hydroxycholeaterol, or 3�,5�,6�-trihydroxycholestane, as well as for the pure POPC bilayer.
Figure 12. Tilt modulus, , for different sterols obtained from the tilt probability distribution
profiles in Figure 7. The uncertainties in the reported values for all the systems, calculated from
the standard deviations obtained from the fits performed for different angular intervals, are <7%,
with the exception of 7�-OH-chol and 7�-OH-chol for which the error bars were 20%.

���
FIGURES
�
������ ��
���
�
������ ��
�
������
�
����
�
������ �
�
������
�
����
�
������ �
�

����
�
������ �
�
�
������ �
�

����
�
������ �
�
����
�
������ ��
����
�
�
������ ��
�
�
������ ��
�
�

����
HIGHLIGHTS
Oxidation by-products of cholesterol (oxysterols) change properties of cell membranes
Tail-oxidized oxysterols behave similarly to cholesterol
Ring-oxidized sterols disrupt cell membrane structure more than tail-oxidized oxysterols
�
Graphical Abstract (for review)




















