
Different effects of calcium and lanthanum on the expression of phytochelatin synthase gene and...
10
UNCORRECTED PROOF DTD 5 Different effects of calcium and lanthanum on the expression of phytochelatin synthase gene and cadmium absorption in Lactuca sativa Zhenyan He, Jiangchuan Li, Haiyan Zhang, Mi Ma * Key Laboratory of Photosynthesis and Molecular Environmental Physiology, Institute of Botany, Chinese Academy of Sciences, Beijing, Xiangshan 100093, PR China Received 3 December 2003; received in revised form 23 June 2004; accepted 1 July 2004 Abstract Phytochelatins (PCs) have an important role in metal detoxification in plants, however the relationship of the expression of Lactuca sativa phytochelatin synthase gene, its catalyzing products—PCs and the absorption of cadmium (Cd) was still not clear. To investigate this relationship, a full-length cDNA of a phytochelatin synthase gene (LsPCS1) was cloned from L. sativa and the physiological changes were observed. When L. sativa (cabbage lettuce) plants were exposed to toxic cadmium, growth was retarded. Though L. sativa plants cadmium tolerance was enhanced when synchronously treated with lanthanum (La 3+ ) or calcium (Ca 2+ ), the effect on the expression of phytochelatin synthase gene and cadmium absorption was different. The transcript level of LsPCS1 and the PCs content in the plants grown under normal conditions were very low, and the Cd absorption was minimal. In comparison with these plants, the expression level of LsPCS1 and the absorption of Cd were enhanced both in leaf and root under Cd 2+ , Cd 2+ + Ca 2+ and Cd 2+ + La 3+ treatments. In comparison with Cd 2+ treatment, the accumulated mRNA level of LsPCS1, PCs content and the absorption of Cd were enhanced with Cd 2+ + Ca 2+ treatment. Though Cd 2+ + La 3+ treatment showed a similar expression profile with Cd 2+ + Ca 2+ treatment, the absorption of Cd of Cd 2+ + La 3+ treatment was less than that of Cd 2+ treatment. Ca 2+ alone has little effect on the expression of LsPCS1, whereas La 3+ itself enhances the mRNA and PCs accumulation. It is suggested that Ca 2+ increases the expression of PC synthase gene under Cd 2+ stress, which in turn enhances the plant tolerance and Cd accumulation. Thus, fertilization with Ca 2+ appears to be a promising strategy for increasing phytoremediation capacity. While La 3+ could decrease the accumulation of Cd, La 3+ may play an important role on reducing the accumulation of Cd of plants grown in contaminated soil. # 2004 Published by Elsevier Ireland Ltd. Keywords: Cadmium; Calcium; Lanthanum; Phytochelatin synthase; Gene expression Toxic heavy metal pollution has become an increasing worldwide environmental problem since the onset of the industry revolution. Cadmium (Cd) used as a ‘model’ non- essential metal of significant environmental relevance in many studies is one of the most serious pollutants. About 38,900 tons of cadmium is discharged annually to the environment [1]. Chronic cadmium poisoning may lead to the damage of the skeletal system and kidney and induce cancer. Recently it is reported that cadmium can act as an estrogen mimic in the whole animal, inducing conditions ranging from uterine hyperplasia to early onset of puberty, at doses as low as 5–10 mg/kg (single intraperitoneal injection) [2]. Due to the exhaust emission, waste water and garbage draining from industry, sewage irrigation, and the utilization of pesticide, chemical combination and herbicide, the heavy metals content of the plants growing in some contaminated soil has exceeded the content of food standard. The plants include two types that have tremendous effects on human health. One is directly edible by human such as the vegetables and the crops, another is indirectly utilized by animals such as the clover that is an economically most important forage legume worldwide [3]. Because of the non- degradable nature of the cadmium, they can be passed to various organisms through the food chains and lead to various diseases in humans. www.elsevier.com/locate/plantsci Plant Science xxx (2004) xxx–xxx 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 * Corresponding author. Tel.: +86 10 8259 9422; fax: +86 10 8259 9422. E-mail address: [email protected] (M. Ma). 0168-9452/$ – see front matter # 2004 Published by Elsevier Ireland Ltd. doi:10.1016/j.plantsci.2004.07.001 PSL 7000 1–10
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Different effects of calcium and lanthanum on the expression of phytochelatin synthase gene and...

DTD 5
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
2728
29
30
31
32
33
34
35
36
37
38
39
www.elsevier.com/locate/plantsci
Plant Science xxx (2004) xxx–xxx
F
Different effects of calcium and lanthanum on the expression
of phytochelatin synthase gene and cadmium
absorption in Lactuca sativa
Zhenyan He, Jiangchuan Li, Haiyan Zhang, Mi Ma*
Key Laboratory of Photosynthesis and Molecular Environmental Physiology, Institute of Botany,
Chinese Academy of Sciences, Beijing, Xiangshan 100093, PR China
Received 3 December 2003; received in revised form 23 June 2004; accepted 1 July 2004
Abstract
OTED
PR
OPhytochelatins (PCs) have an important role in metal detoxification in plants, however the relationship of the expression of Lactuca sativa
phytochelatin synthase gene, its catalyzing products—PCs and the absorption of cadmium (Cd) was still not clear. To investigate this
relationship, a full-length cDNA of a phytochelatin synthase gene (LsPCS1) was cloned from L. sativa and the physiological changes were
observed. When L. sativa (cabbage lettuce) plants were exposed to toxic cadmium, growth was retarded. Though L. sativa plants cadmium
tolerance was enhanced when synchronously treated with lanthanum (La3+) or calcium (Ca2+), the effect on the expression of phytochelatin
synthase gene and cadmium absorption was different. The transcript level of LsPCS1 and the PCs content in the plants grown under normal
conditions were very low, and the Cd absorption was minimal. In comparison with these plants, the expression level of LsPCS1 and the
absorption of Cd were enhanced both in leaf and root under Cd2+, Cd2+ + Ca2+ and Cd2+ + La3+ treatments. In comparison with Cd2+ treatment,
the accumulated mRNA level of LsPCS1, PCs content and the absorption of Cd were enhanced with Cd2+ + Ca2+ treatment. Though Cd2+ +
La3+ treatment showed a similar expression profile with Cd2+ + Ca2+ treatment, the absorption of Cd of Cd2+ + La3+ treatment was less than that
of Cd2+ treatment. Ca2+ alone has little effect on the expression of LsPCS1, whereas La3+ itself enhances the mRNA and PCs accumulation. It is
suggested that Ca2+ increases the expression of PC synthase gene under Cd2+ stress, which in turn enhances the plant tolerance and Cd
accumulation. Thus, fertilization with Ca2+ appears to be a promising strategy for increasing phytoremediation capacity. While La3+ could
decrease the accumulation of Cd, La3+ may play an important role on reducing the accumulation of Cd of plants grown in contaminated soil.
# 2004 Published by Elsevier Ireland Ltd.
Keywords: Cadmium; Calcium; Lanthanum; Phytochelatin synthase; Gene expression
EC 40
41
42
43
44
45
46
47
48
49
50
ORR
Toxic heavy metal pollution has become an increasing
worldwide environmental problem since the onset of the
industry revolution. Cadmium (Cd) used as a ‘model’ non-
essential metal of significant environmental relevance in
many studies is one of the most serious pollutants. About
38,900 tons of cadmium is discharged annually to the
environment [1]. Chronic cadmium poisoning may lead to
the damage of the skeletal system and kidney and induce
cancer. Recently it is reported that cadmium can act as an
estrogen mimic in the whole animal, inducing conditions
ranging from uterine hyperplasia to early onset of puberty, at
UN
C 51
52
53
* Corresponding author. Tel.: +86 10 8259 9422; fax: +86 10 8259 9422.
E-mail address: [email protected] (M. Ma).
0168-9452/$ – see front matter # 2004 Published by Elsevier Ireland Ltd.
doi:10.1016/j.plantsci.2004.07.001
doses as low as 5–10 mg/kg (single intraperitoneal injection)
[2]. Due to the exhaust emission, waste water and garbage
draining from industry, sewage irrigation, and the utilization
of pesticide, chemical combination and herbicide, the heavy
metals content of the plants growing in some contaminated
soil has exceeded the content of food standard. The plants
include two types that have tremendous effects on human
health. One is directly edible by human such as the
vegetables and the crops, another is indirectly utilized by
animals such as the clover that is an economically most
important forage legume worldwide [3]. Because of the non-
degradable nature of the cadmium, they can be passed to
various organisms through the food chains and lead to
various diseases in humans.
PSL 7000 1–10

Z. He et al. / Plant Science xxx (2004) xxx–xxx2
DTD 5
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
It is estimated that cleanup of hazardous wastes using
conventional technologies will cost at least $200 billion in
the US alone [4]. Phytoremediation is a cost-effective and
environment-friendly technology [5–7], but there are many
problems to be solved, such as the scarcity of hyperaccu-
mulator, the complexity of the mechanisms of heavy metal
tolerance in plants, the low-efficiency of absorption for
heavy metal and so on.
Plants can produce Cys-rich peptides such as glutathione
(GSH), phytochelatins (PCs), or metallothioneins (MTs) for
detoxification or homeostasis of heavy metals [8–12]. PCs
are a kind of polypeptides with the structure (g-Glu-Cys)n-
Gly (where n = 2–11) and identified in a wide variety of plant
species and in some microorganisms [13,14]. It has been
demonstrated that PCs play an important role in metal
detoxification by transporting the metal Cd2+ to vacuole
[13], and PC synthase genes were isolated from Arabidopsis,
wheat and S. cerevisiae (AtPCS/CAD1, TaPCS and SpPCS)
in 1999 by three independent laboratories [15–17]. It is
reported that calcium (Ca) or lanthanum (La) can alleviate
cadmium toxicity in plant by anatomical and enzymological
methods [18,19]. Choi et al. [18] found that tobacco plants
actively excluded toxic Cd by forming and excreting Cd/Ca-
containing crystals through the head cells of trichomes.
Another study suggested the increased antioxidant enzymes
by La(NO3)3 enhance the antioxidant potential of the wheat
seedlings to reduce lead stress [19]. However, there are no
reports about what effects Ca and La have on the expression
of PC synthase gene, which has affinity with heavy metal
tolerance. We selected Lactuca sativa, a kind of common
vegetable, as the material to study how calcium and
lanthanum affect L. sativa cadmium tolerance and what
effects Ca and La have on the cadmium absorption in plants
and the expression of PC synthase gene, to find a way to
decrease the heavy metal accumulation in plants.
C
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
NCOR
RE
1. Materials and methods
1.1. Plant material and treatments
1.1.1. Experiment I (seedlings in plastic Petri dishes)
L. sativa seeds were sterilized by rinsing in 70% ethanol
for 30 s, then in 0.1% HgCl2 for 10 min, and subsequently in
sterile, deionized water for 30 min, all on rocking platform.
Fifty sterilized seeds were sown in each plastic Petri dish in
hydroponic conditions containing 0.05 mM CdCl2, 4 mM
CaCl2 and 0.04 mg/L La(NO3)3. After differently treated for
5 d at 25 8C under continuous dark, the seedlings were
harvested, washed, and weighed, and the length of the root
was measured.
1.1.2. Experiment II (plants on sand medium)
L. sativa seeds were grown on sand conditions with to 1/2
MS (Murashige and Skoog 1962, CaCl2 free) medium for
8 h light and 16 h dark. After 20 d at 25 8C, the sand medium
U
TED
PR
OO
F
were added 0.5 mM CdCl2, 4 mM CaCl2 and 0.04 mg/L
La(NO3)3. Leaf and root were collected from all treatments
after plants were treated 1, 2, 6, 12, 24 and 48 h. Some
materials were frozen in liquid nitrogen and stored in
�80 8C used for RT-PCR analysis and phytochelatins
determination. Others were washed in deionized water for
2 min, dried at 60 8C for Cd elemental analysis. The plants
of 48 h treatment were weighed and the length of the root
was measured only.
1.2. Elemental analysis
Leaf and root were harvested, washed in deionized water
for 2 min, dried at 60 8C and then macerated in HNO3. After
acid digestion, the concentrations of Cd in the clear extracts
were measured with graphite atomic absorption spectrometer.
1.3. Amplification of the full LsPCS1 cDNA
Total RNA from the plant of L. sativa was extracted using
Trizol Reagent (Invitrogen, Karlsruhe, Germany) followed
by chloroform extraction and isopropanol precipitation.
Polyadenylated mRNAs were obtained using Oligotex
mRNA Spin-Column (clontech). Following reverse tran-
scription from mRNA, the first-strand cDNA was used as the
template in following reaction.
PC synthase genes are very conservative in the N-
terminal of all kinds of organisms. Two degenerate primers
were designed according to the conservative domains of the
PCS cDNA from the dicotyledon: Arabidopsis, Brassica
juncea and Glycine max. The sequences were: 50-ATGGC(T/
G)ATGGCG(A/G)GTTT((A/G)TATCG-30 (up) and 50-TGCCGTCGCAG(A/G)TGCA(G/A)(T/A)ACCT-30 (down).
PCR was performed with denaturing for 30 s at 95 8C,
annealing for 45 s at 60 8C and extending for 1 min at 72 8C.
The cycle was repeated 30 times using cDNA of L. sativa as
a template. The products of PCR were ligated into pGEM-T
Easy Vector (Promega). The products were transformed into
TOP 10-E. coli cells. Plasmid DNA was isolated and
sequenced. A 50 fragment of 800 bp was firstly obtained
from L. sativa cDNA. Sequence analysis was performed
using DNAman software.
The 30 fragment was obtained using SMARTTM RACE
cDNA Amplification Kit (clontech). The gene specific
primers (GSPs) for the 30-RACE reactions is as follows:
GSP2: 50-GCTTGACTGTTGCGAGCCTTTG-30, NGSP2:
50-TGCTCGCTTCAAGTACCCTCCTC-30.
1.4. RT-PCR analysis
Leaf and root were homogenized with a mortar and pestle
in liquid nitrogen. RNA was extracted using Trizol Reagent.
RT-PCR was manipulated using mRNA Selective PCR Kit
(Takara). PCR reactions using equal amounts of RNA
samples were performed. Cycle numbers were optimized
assure that the amplification reaction was tested in the
PSL 7000 1–10
ED
PR
OO
F
Z. He et al. / Plant Science xxx (2004) xxx–xxx 3
DTD 5
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
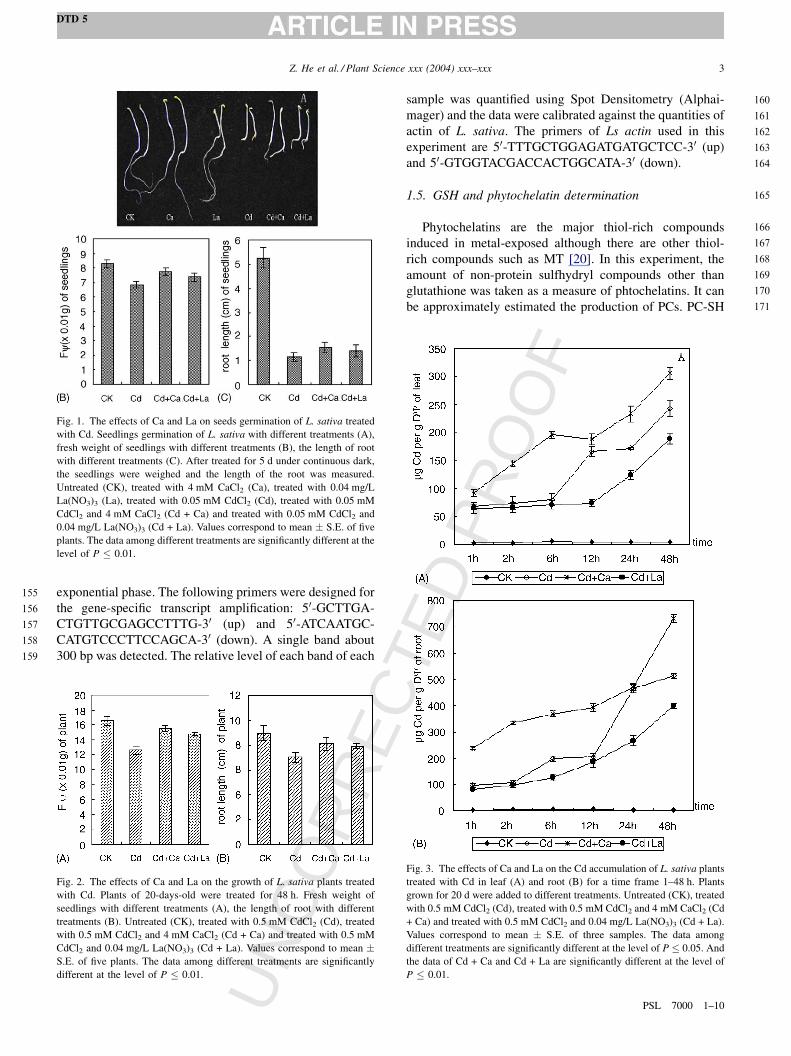
Fig. 1. The effects of Ca and La on seeds germination of L. sativa treated
with Cd. Seedlings germination of L. sativa with different treatments (A),
fresh weight of seedlings with different treatments (B), the length of root
with different treatments (C). After treated for 5 d under continuous dark,
the seedlings were weighed and the length of the root was measured.
Untreated (CK), treated with 4 mM CaCl2 (Ca), treated with 0.04 mg/L
La(NO3)3 (La), treated with 0.05 mM CdCl2 (Cd), treated with 0.05 mM
CdCl2 and 4 mM CaCl2 (Cd + Ca) and treated with 0.05 mM CdCl2 and
0.04 mg/L La(NO3)3 (Cd + La). Values correspond to mean � S.E. of five
plants. The data among different treatments are significantly different at the
level of P � 0.01.
exponential phase. The following primers were designed for
the gene-specific transcript amplification: 50-GCTTGA-
CTGTTGCGAGCCTTTG-30 (up) and 50-ATCAATGC-
CATGTCCCTTCCAGCA-30 (down). A single band about
300 bp was detected. The relative level of each band of each
UN
CO
RR
EC
Fig. 2. The effects of Ca and La on the growth of L. sativa plants treated
with Cd. Plants of 20-days-old were treated for 48 h. Fresh weight of
seedlings with different treatments (A), the length of root with different
treatments (B). Untreated (CK), treated with 0.5 mM CdCl2 (Cd), treated
with 0.5 mM CdCl2 and 4 mM CaCl2 (Cd + Ca) and treated with 0.5 mM
CdCl2 and 0.04 mg/L La(NO3)3 (Cd + La). Values correspond to mean �S.E. of five plants. The data among different treatments are significantly
different at the level of P � 0.01.
sample was quantified using Spot Densitometry (Alphai-
mager) and the data were calibrated against the quantities of
actin of L. sativa. The primers of Ls actin used in this
experiment are 50-TTTGCTGGAGATGATGCTCC-30 (up)
and 50-GTGGTACGACCACTGGCATA-30 (down).
1.5. GSH and phytochelatin determination
Phytochelatins are the major thiol-rich compounds
induced in metal-exposed although there are other thiol-
rich compounds such as MT [20]. In this experiment, the
amount of non-protein sulfhydryl compounds other than
glutathione was taken as a measure of phtochelatins. It can
be approximately estimated the production of PCs. PC-SH
T
PSL 7000 1–10
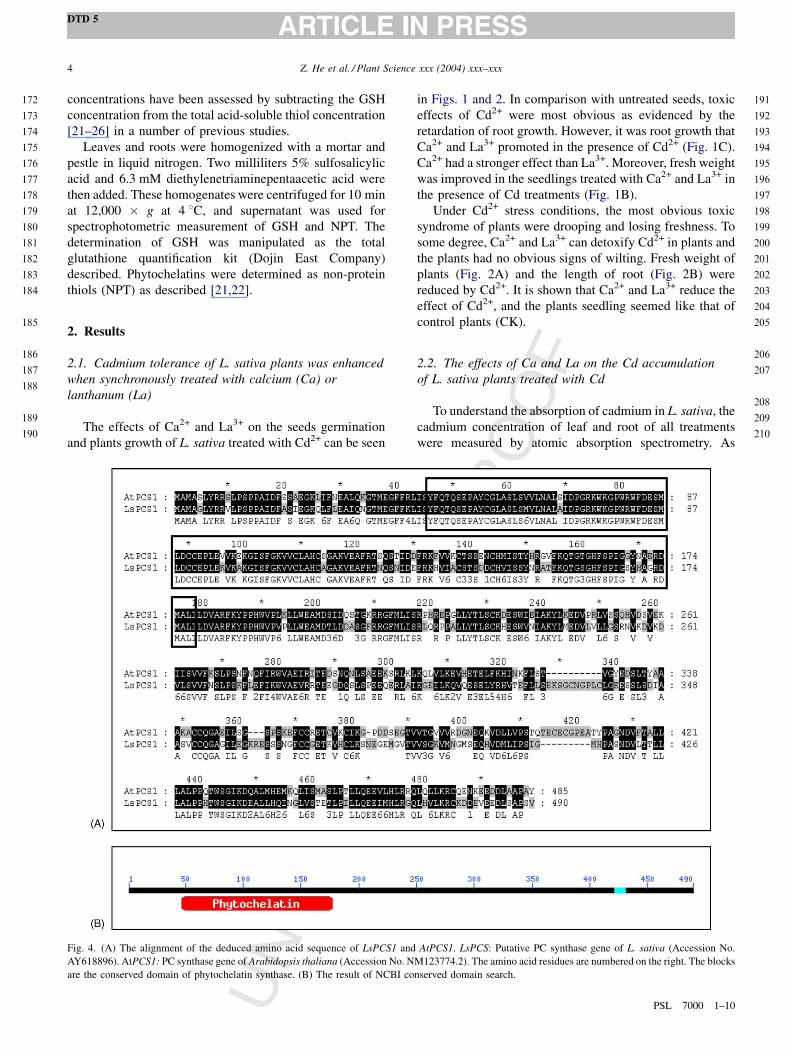
Fig. 3. The effects of Ca and La on the Cd accumulation of L. sativa plants
treated with Cd in leaf (A) and root (B) for a time frame 1–48 h. Plants
grown for 20 d were added to different treatments. Untreated (CK), treated
with 0.5 mM CdCl2 (Cd), treated with 0.5 mM CdCl2 and 4 mM CaCl2 (Cd
+ Ca) and treated with 0.5 mM CdCl2 and 0.04 mg/L La(NO3)3 (Cd + La).
Values correspond to mean � S.E. of three samples. The data among
different treatments are significantly different at the level of P � 0.05. And
the data of Cd + Ca and Cd + La are significantly different at the level of
P � 0.01.
Z. He et al. / Plant Science xxx (2004) xxx–xxx4
DTD 5
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
concentrations have been assessed by subtracting the GSH
concentration from the total acid-soluble thiol concentration
[21–26] in a number of previous studies.
Leaves and roots were homogenized with a mortar and
pestle in liquid nitrogen. Two milliliters 5% sulfosalicylic
acid and 6.3 mM diethylenetriaminepentaacetic acid were
then added. These homogenates were centrifuged for 10 min
at 12,000 � g at 4 8C, and supernatant was used for
spectrophotometric measurement of GSH and NPT. The
determination of GSH was manipulated as the total
glutathione quantification kit (Dojin East Company)
described. Phytochelatins were determined as non-protein
thiols (NPT) as described [21,22].
2. Results
2.1. Cadmium tolerance of L. sativa plants was enhanced
when synchronously treated with calcium (Ca) or
lanthanum (La)
The effects of Ca2+ and La3+ on the seeds germination
and plants growth of L. sativa treated with Cd2+ can be seen
UN
CO
RR
EC
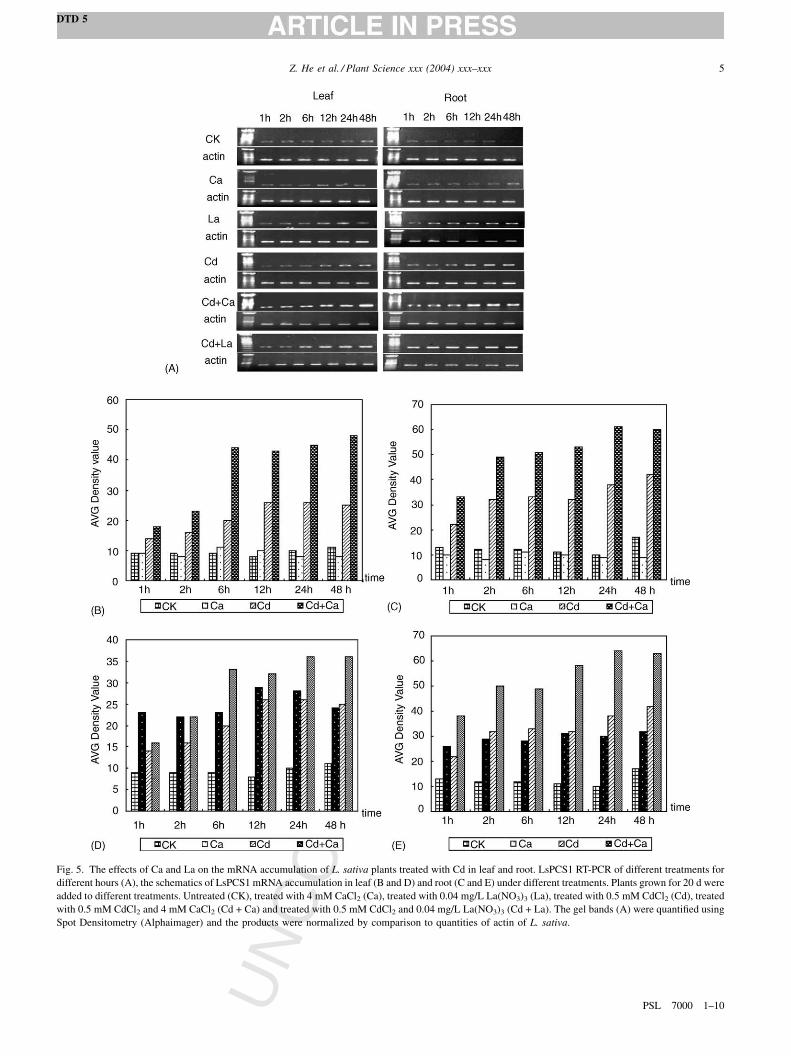
Fig. 4. (A) The alignment of the deduced amino acid sequence of LsPCS1 and
AY618896). AtPCS1: PC synthase gene of Arabidopsis thaliana (Accession No. N
are the conserved domain of phytochelatin synthase. (B) The result of NCBI co
OF
in Figs. 1 and 2. In comparison with untreated seeds, toxic
effects of Cd2+ were most obvious as evidenced by the
retardation of root growth. However, it was root growth that
Ca2+ and La3+ promoted in the presence of Cd2+ (Fig. 1C).
Ca2+ had a stronger effect than La3+. Moreover, fresh weight
was improved in the seedlings treated with Ca2+ and La3+ in
the presence of Cd treatments (Fig. 1B).
Under Cd2+ stress conditions, the most obvious toxic
syndrome of plants were drooping and losing freshness. To
some degree, Ca2+ and La3+ can detoxify Cd2+ in plants and
the plants had no obvious signs of wilting. Fresh weight of
plants (Fig. 2A) and the length of root (Fig. 2B) were
reduced by Cd2+. It is shown that Ca2+ and La3+ reduce the
effect of Cd2+, and the plants seedling seemed like that of
control plants (CK).
2.2. The effects of Ca and La on the Cd accumulation
of L. sativa plants treated with Cd
To understand the absorption of cadmium in L. sativa, the
cadmium concentration of leaf and root of all treatments
were measured by atomic absorption spectrometry. As
TED
PR
O
PSL 7000 1–10
AtPCS1. LsPCS: Putative PC synthase gene of L. sativa (Accession No.
M123774.2). The amino acid residues are numbered on the right. The blocks
nserved domain search.
UN
CO
RR
EC
TED
PR
OO
F
Z. He et al. / Plant Science xxx (2004) xxx–xxx 5
DTD 5
PSL 7000 1–10
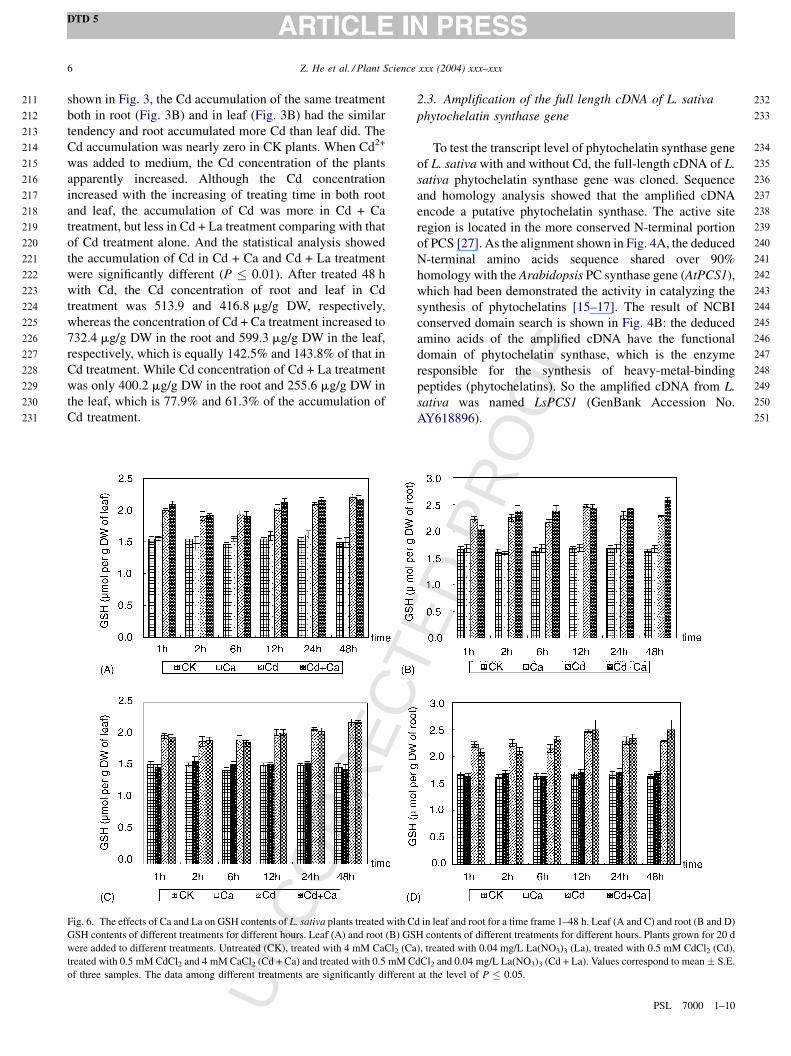
Fig. 5. The effects of Ca and La on the mRNA accumulation of L. sativa plants treated with Cd in leaf and root. LsPCS1 RT-PCR of different treatments for
different hours (A), the schematics of LsPCS1 mRNA accumulation in leaf (B and D) and root (C and E) under different treatments. Plants grown for 20 d were
added to different treatments. Untreated (CK), treated with 4 mM CaCl2 (Ca), treated with 0.04 mg/L La(NO3)3 (La), treated with 0.5 mM CdCl2 (Cd), treated
with 0.5 mM CdCl2 and 4 mM CaCl2 (Cd + Ca) and treated with 0.5 mM CdCl2 and 0.04 mg/L La(NO3)3 (Cd + La). The gel bands (A) were quantified using
Spot Densitometry (Alphaimager) and the products were normalized by comparison to quantities of actin of L. sativa.
Z. He et al. / Plant Science xxx (2004) xxx–xxx6
DTD 5
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
shown in Fig. 3, the Cd accumulation of the same treatment
both in root (Fig. 3B) and in leaf (Fig. 3B) had the similar
tendency and root accumulated more Cd than leaf did. The
Cd accumulation was nearly zero in CK plants. When Cd2+
was added to medium, the Cd concentration of the plants
apparently increased. Although the Cd concentration
increased with the increasing of treating time in both root
and leaf, the accumulation of Cd was more in Cd + Ca
treatment, but less in Cd + La treatment comparing with that
of Cd treatment alone. And the statistical analysis showed
the accumulation of Cd in Cd + Ca and Cd + La treatment
were significantly different (P � 0.01). After treated 48 h
with Cd, the Cd concentration of root and leaf in Cd
treatment was 513.9 and 416.8 mg/g DW, respectively,
whereas the concentration of Cd + Ca treatment increased to
732.4 mg/g DW in the root and 599.3 mg/g DW in the leaf,
respectively, which is equally 142.5% and 143.8% of that in
Cd treatment. While Cd concentration of Cd + La treatment
was only 400.2 mg/g DW in the root and 255.6 mg/g DW in
the leaf, which is 77.9% and 61.3% of the accumulation of
Cd treatment.
UN
CO
RR
EC
Fig. 6. The effects of Ca and La on GSH contents of L. sativa plants treated with C
GSH contents of different treatments for different hours. Leaf (A) and root (B) GS
were added to different treatments. Untreated (CK), treated with 4 mM CaCl2 (Ca
treated with 0.5 mM CdCl2 and 4 mM CaCl2 (Cd + Ca) and treated with 0.5 mM C
of three samples. The data among different treatments are significantly different
OF
2.3. Amplification of the full length cDNA of L. sativa
phytochelatin synthase gene
To test the transcript level of phytochelatin synthase gene
of L. sativa with and without Cd, the full-length cDNA of L.
sativa phytochelatin synthase gene was cloned. Sequence
and homology analysis showed that the amplified cDNA
encode a putative phytochelatin synthase. The active site
region is located in the more conserved N-terminal portion
of PCS [27]. As the alignment shown in Fig. 4A, the deduced
N-terminal amino acids sequence shared over 90%
homology with the Arabidopsis PC synthase gene (AtPCS1),
which had been demonstrated the activity in catalyzing the
synthesis of phytochelatins [15–17]. The result of NCBI
conserved domain search is shown in Fig. 4B: the deduced
amino acids of the amplified cDNA have the functional
domain of phytochelatin synthase, which is the enzyme
responsible for the synthesis of heavy-metal-binding
peptides (phytochelatins). So the amplified cDNA from L.
sativa was named LsPCS1 (GenBank Accession No.
AY618896).
TED P
RO
PSL 7000 1–10
d in leaf and root for a time frame 1–48 h. Leaf (A and C) and root (B and D)
H contents of different treatments for different hours. Plants grown for 20 d
), treated with 0.04 mg/L La(NO3)3 (La), treated with 0.5 mM CdCl2 (Cd),
dCl2 and 0.04 mg/L La(NO3)3 (Cd + La). Values correspond to mean � S.E.
at the level of P � 0.05.
Z. He et al. / Plant Science xxx (2004) xxx–xxx 7
DTD 5
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
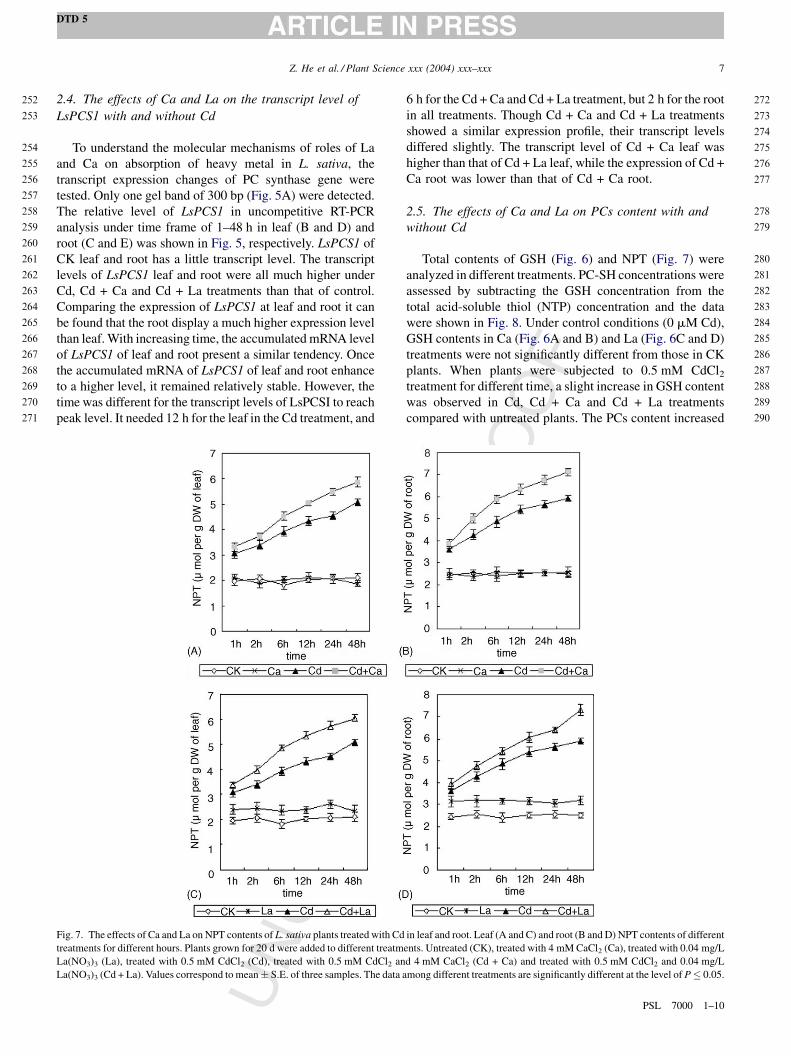
2.4. The effects of Ca and La on the transcript level of
LsPCS1 with and without Cd
To understand the molecular mechanisms of roles of La
and Ca on absorption of heavy metal in L. sativa, the
transcript expression changes of PC synthase gene were
tested. Only one gel band of 300 bp (Fig. 5A) were detected.
The relative level of LsPCS1 in uncompetitive RT-PCR
analysis under time frame of 1–48 h in leaf (B and D) and
root (C and E) was shown in Fig. 5, respectively. LsPCS1 of
CK leaf and root has a little transcript level. The transcript
levels of LsPCS1 leaf and root were all much higher under
Cd, Cd + Ca and Cd + La treatments than that of control.
Comparing the expression of LsPCS1 at leaf and root it can
be found that the root display a much higher expression level
than leaf. With increasing time, the accumulated mRNA level
of LsPCS1 of leaf and root present a similar tendency. Once
the accumulated mRNA of LsPCS1 of leaf and root enhance
to a higher level, it remained relatively stable. However, the
time was different for the transcript levels of LsPCSI to reach
peak level. It needed 12 h for the leaf in the Cd treatment, and
UN
CO
RR
EC
Fig. 7. The effects of Ca and La on NPT contents of L. sativa plants treated with Cd
treatments for different hours. Plants grown for 20 d were added to different treatm
La(NO3)3 (La), treated with 0.5 mM CdCl2 (Cd), treated with 0.5 mM CdCl2 an
La(NO3)3 (Cd + La). Values correspond to mean � S.E. of three samples. The data a
OF
6 h for the Cd + Ca and Cd + La treatment, but 2 h for the root
in all treatments. Though Cd + Ca and Cd + La treatments
showed a similar expression profile, their transcript levels
differed slightly. The transcript level of Cd + Ca leaf was
higher than that of Cd + La leaf, while the expression of Cd +
Ca root was lower than that of Cd + Ca root.
2.5. The effects of Ca and La on PCs content with and
without Cd
Total contents of GSH (Fig. 6) and NPT (Fig. 7) were
analyzed in different treatments. PC-SH concentrations were
assessed by subtracting the GSH concentration from the
total acid-soluble thiol (NTP) concentration and the data
were shown in Fig. 8. Under control conditions (0 mM Cd),
GSH contents in Ca (Fig. 6A and B) and La (Fig. 6C and D)
treatments were not significantly different from those in CK
plants. When plants were subjected to 0.5 mM CdCl2treatment for different time, a slight increase in GSH content
was observed in Cd, Cd + Ca and Cd + La treatments
compared with untreated plants. The PCs content increased
TED P
RO
PSL 7000 1–10
in leaf and root. Leaf (A and C) and root (B and D) NPT contents of different
ents. Untreated (CK), treated with 4 mM CaCl2 (Ca), treated with 0.04 mg/L
d 4 mM CaCl2 (Cd + Ca) and treated with 0.5 mM CdCl2 and 0.04 mg/L
mong different treatments are significantly different at the level of P � 0.05.
CTE
D P
RO
OF
Z. He et al. / Plant Science xxx (2004) xxx–xxx8
DTD 5
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
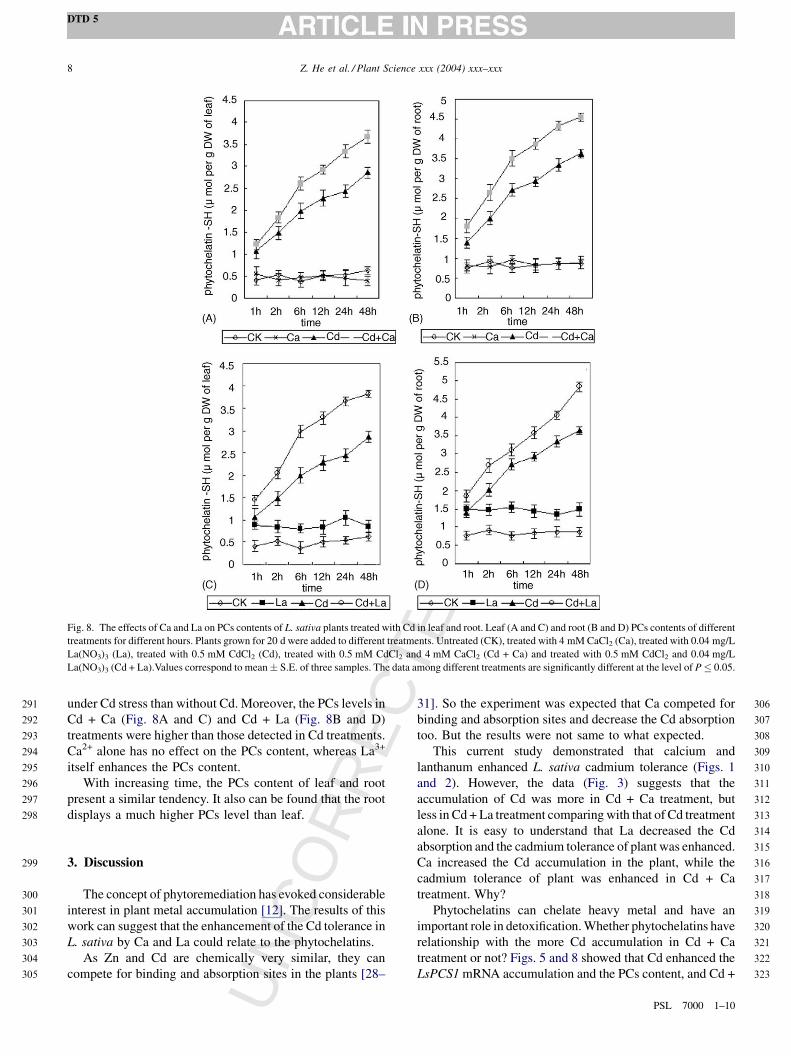
Fig. 8. The effects of Ca and La on PCs contents of L. sativa plants treated with Cd in leaf and root. Leaf (A and C) and root (B and D) PCs contents of different
treatments for different hours. Plants grown for 20 d were added to different treatments. Untreated (CK), treated with 4 mM CaCl2 (Ca), treated with 0.04 mg/L
La(NO3)3 (La), treated with 0.5 mM CdCl2 (Cd), treated with 0.5 mM CdCl2 and 4 mM CaCl2 (Cd + Ca) and treated with 0.5 mM CdCl2 and 0.04 mg/L
La(NO3)3 (Cd + La).Values correspond to mean � S.E. of three samples. The data among different treatments are significantly different at the level of P � 0.05.
RE
under Cd stress than without Cd. Moreover, the PCs levels in
Cd + Ca (Fig. 8A and C) and Cd + La (Fig. 8B and D)
treatments were higher than those detected in Cd treatments.
Ca2+ alone has no effect on the PCs content, whereas La3+
itself enhances the PCs content.
With increasing time, the PCs content of leaf and root
present a similar tendency. It also can be found that the root
displays a much higher PCs level than leaf.
R 314315
316
317
318
319
320
321
322
323
NCO3. Discussion
The concept of phytoremediation has evoked considerable
interest in plant metal accumulation [12]. The results of this
work can suggest that the enhancement of the Cd tolerance in
L. sativa by Ca and La could relate to the phytochelatins.
As Zn and Cd are chemically very similar, they can
compete for binding and absorption sites in the plants [28–
U
31]. So the experiment was expected that Ca competed for
binding and absorption sites and decrease the Cd absorption
too. But the results were not same to what expected.
This current study demonstrated that calcium and
lanthanum enhanced L. sativa cadmium tolerance (Figs. 1
and 2). However, the data (Fig. 3) suggests that the
accumulation of Cd was more in Cd + Ca treatment, but
less in Cd + La treatment comparing with that of Cd treatment
alone. It is easy to understand that La decreased the Cd
absorption and the cadmium tolerance of plant was enhanced.
Ca increased the Cd accumulation in the plant, while the
cadmium tolerance of plant was enhanced in Cd + Ca
treatment. Why?
Phytochelatins can chelate heavy metal and have an
important role in detoxification. Whether phytochelatins have
relationship with the more Cd accumulation in Cd + Ca
treatment or not? Figs. 5 and 8 showed that Cd enhanced the
LsPCS1 mRNA accumulation and the PCs content, and Cd +
PSL 7000 1–10

EC
Z. He et al. / Plant Science xxx (2004) xxx–xxx 9
DTD 5
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
425
426
427
428
429
430
431
432
RR
Ca enhanced the expression level of LsPCS1 comparing with
that of Cd treatment alone, but not Ca. It is suggested that the
enhancement of LsPCS1 expression leads to the increase of
the cadmium absorption in plants. We guess that the increased
PCs under the Ca chelated the most cadmium, then transported
to the vacuole, so Ca can decrease the toxicity of Cd.
How does Ca promote the expression of LsPCS1 in the
stress of Cd? Calcium is an important nutrient for plant
growth and Ca2+ influx and Ca2+ release from organelles
play important roles in many signaling cascades of the plant
[32,33]. We presume that Ca accelerates the transmission to
the Cd stress signal and enhances the LsPCS1 expression,
which increases the Cd absorption in plants, although further
study is needed to find the direct linkage. Pharmacology and
physics analysis will provide useful tools to study the
mechanisms. Lombi et al. [34] offered physiological
evidence for a high-affinity specific cadmium transporter
highly expressed in a Thlaspi caerulescens ecotype in the
root cell plasma membrane. It is possible that Ca induced the
high expression of LsPCS1 and cadmium transporter and led
to the enhancement of Cd absorption.
It has been known that lanthanon can enhance the
adaptability to stress conditions or increase the tolerance to
adversity such as low temperature, high temperature,
drought and salt. It was reported that lanthanon lightened
the plant damage by lead [19]. We were interesting in the
effects of La on seed germination and growth of L. sativa
under Cd stress. The results proved L. sativa plants
cadmium tolerance was enhanced when synchronously
treated with La. La itself enhances the LsPCS1 mRNA
accumulation and PCs content both in leaf and root. The
LsPCS1 expression on Cd + La treatment was higher in leaf
and root than Cd, but the Cd absorption is less than Cd
treatment. From the above results the following conclusions
could be made: The concentration of La is so low that La
cannot lead to antagonistic effects to Cd. However, the exact
mechanisms of the enhancement of LSPCS1 expression by La
itself remain unclear. It has been demonstrated by recent
research that phytochelatin have other bio-function in plants
[35]. Perhaps because La induced other mechanisms about
cadmium tolerance, which led to the decrease of Cd
absorption, La enhanced the cadmium tolerance of L. sativa
plants.
In conclusion, it was suggested that calcium and
lanthanum had different effects in Cd absorption. Further-
more, fertilization with Ca2+ appears to be a promising
strategy for increasing phytoremediation. As La can
decrease the accumulation of Cd, it may play an important
role on reducing the accumulation of Cd of the crops
growing in contaminated soil.
O433
434
435
436
437
438
439
NCAcknowledgmentsThe work was supported by the National Science
Foundation Council of PRC (Grant no. 30170086,
U
30370127), the Special Project of Transgenic Research
(JY03A2001) and Hi-tech Research and Development
Program of China (2001AA645010-5).
TED
PR
OO
F
References
[1] G.M. Gadd, C. White, Microbial treatment of metal pollution—a
working biotechnology? Trends Biotechnol. 11 (1993) 353–359.
[2] M.D. Johnson, N. Kenney, A. Stoica, L. Hilakivi-Clarke, B. Singh, G.R.
Chepko, Clarke, P.F. Sholler, A.A. Lirio, C. Foss, R. Reiter, B. Trock, S.
Paik, M.B. Martin, Cadmium mimics the in vivo effects of estrogen in
the uterus and mammary gland, Nat. Med. 9 (2003) 1081–1084.
[3] Y.H. Lin, E. Ludlow, R. Kalla, C. Pallaghy, M. Emmerling, G.
Spangenberg, Organ-specific, developmentally-regulated and abiotic
stress-induced activities of four Arabidopsis thaliana promoters in
transgenic white clover (Trifolium repens L.), Plant Sci. 165 (2003)
1437–1444.
[4] US Environmental Protection Agency (EPA), Environmental Invest-
ments: The Cost of a Clean Environment, vol. 5, US Government
Printing Office, Washington, DC, 1990 (ed EPA-230-11-90-083).
[5] C. Gisbert, R. Ros, A.D. Haro, D.J. Walker, M.P. Bernal, R. Serrano, J.
Navarro-Avin, A plant genetically modified that accumulates Pb is
especially promising for phytoremediation, Biochem. Biophys. Res.
Commun. 303 (2003) 440–445.
[6] R.L. Chaney, M. Malik, Y.M. Li, S.L. Brown, E.P. Brewer, J.S. Angle,
A.J.M. Baker, Phytoremediation of soil metals, Curr. Opin. Biotech-
nol. 8 (1997) 279–284.
[7] D.E. Salt, R.D. Smith, I. Raskin, Phytoremediation, Annu. Rev. Plant
Physiol. Plant Mol. Biol. 49 (1998) 643–668.
[8] W.E. Rauser, Structure and function of metal chelators produced by
plants: the case for organic acids, amino acids, phytin and metal-
lothioneins, Cell Biochem. Biophys. 32 (1999) 19–48;
C.S. Cobbett, A family of phytochelatin synthase genes from plant,
fungal and animal species, Trends Plant Sci. 4 (1999) 335–337.
[9] C. Cobbett, Phytochelatins and their roles in heavy metal detoxifica-
tion, Plant Physiol. 123 (2000) 825–832.
[10] C. Cobbett, Phytochelatin biosynthesis and function in heavy-metal
detoxification, Curr. Opin. Plant Biol. 3 (2000) 211–216.
[11] S. Lee, J.S. Moon, T.S. Ko, D. Petros, P. Goldsbrough, S.S. Korban,
Overexpression of Arabidopsis phytochelatin synthase paradoxically
leads to hypersensitivity to cadmium stress 1, Plant Physiol. 131
(2003) 656–663.
[12] M. Ma, P.S. Lau, Y.T. Jia, W.K. Tsang, S.K.S. Lam, N.F.Y. Tam, Y.S.
Wong, The isolation and characterization of Type 1 metallothionein
(MT) cDNA from a heavy-metal-tolerant plant, Festuca rubra cv.
Merlin, Plant Sci. 164 (2003) 51–60.
[13] C. Cobbett, P. Goldsbrough, Phytochelatins and metallothioneins:
roles in heavy metal detoxification and homeostasis, Annu. Rev. Plant
Biol. 53 (2002) 159–182.
[14] W.E. Rauser, Phytochelatins and related peptides: structure, biosynth-
esis, and function, Plant Physiol. 109 (1995) 1141–1149.
[15] S.B. Ha, A.P. Smith, R. Howden, W.M. Dietrich, S. Bugg, M.J.
O’Connell, P.B. Goldsbrough, C.S. Cobbett, Phytochelatin synthase
genes from Arabidopsis and the yeast, Schizosaccharomyces pombe,
Plant Cell 11 (1999) 1153–1164.
[16] S. Clemens, E.J. Kim, D. Neumann, J.I. Schroeder, Tolerance to toxic
metals by a gene family of phytochelatin synthases from plants and
yeast, EMBO J. 18 (1999) 3325–3333.
[17] O.K. Vatamaniuk, S. Mari, Y.P. Lu, P.A. Rea, AtPCS1, a phytochelatin
synthase from Arabidopsis: isolation and in vitro reconstitution, Proc.
Natl. Acad. Sci. U.S.A. 96 (1999) 7110–7115.
[18] Y.E. Choi, E. Harada, M. Wada, H. Tsuboi, Y. Morita, T. Kusano, H.
Sano, Detoxification of cadmium in tobacco plants: formation and
active excretion of crystals containing cadmium and calcium through
trichomes, Planta 213 (2001) 45–50.
PSL 7000 1–10

Z. He et al. / Plant Science xxx (2004) xxx–xxx10
DTD 5
440
441
442
443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
472
473
474
475
476
477
478
479
480
481
482
483
484
485
486
487
488
489
490
491
492
493
[19] X. Pang, D.H. Wang, A. Peng, Effect of La on the activities of
antioxidant enzymes in wheat seedlings under lead stress, Environ.
Chem. 21 (2002) 318–323 (in Chinese).
[20] J.C. Steffens, The heavy metal-binding peptides of plants, Annu. Rev.
Plant Physiol. Plant Mol. Biol. 41 (1990) 553–575.
[21] C.H.R. De Vos, M.J. Vonk, R. Vooijs, H. Schat, Glutathione depletion
due to copper-induced phytochelatin synthesis causes oxidative stress
in Silene cucubalus, Plant Physiol. 98 (1992) 853–858.
[22] M.J. May, T. Vernoux, R. Sanchez-Fernandez, M. Van Montagu, D.
Inze, Evidence for posttranscriptional activation of g-glutamylcys-
teine synthetase during plant stress response, Proc. Natl. Acad. Sci. 95
(1998) 12049–12054.
[23] H.V. Scheller, B. Huang, E. Hatch, P.B. Goldsbrough, Phytochelatin
synthesis and glutathione levels in response to heavy metals in tomato
cells, Plant Physiol. 85 (1987) 1031–1035.
[24] R. Vogeli-Lange, G.J. Wagner, Subcellular localization of cadmium
and cadmium-binding peptides in tobacco leaf, Plant Physiol. l92
(1990) 1086–1093.
[25] J.A. De Knecht, P.L.M. Koevoets, J.A.C. Verkleij, W.H.O. Ernst,
Evidence against a role for phytochelatins in naturally selected
increased cadmium tolerance in Silene vulgaris (Moench) Garcke,
New Phytol. l22 (1992) 681–688.
[26] H. Schat, M.M.A. Kalff, Are phytochelatins involved in differential
metal tolerance or do they merely reflect metal-imposed strain? Plant
Physiol. 99 (1992) 1475–1480.
[27] R. Ruotolo, A. Peracchi, A. Bolchi, G. Infusini, A. Amoresano, S.
Ottonello, Domain organization of phytochelatin synthase: functional
UN
CO
RR
EC
OF
properties of truncated enzyme species identified by limited proteo-
lysis, J. Biol. Chem. 279 (2004) 14686–14693.
[28] L. Ozturk, S. Karanlik, F. Ozkutlu, I. Cakmak, L.V. Kochian, Shoot
biomass and zinc/cadmium uptake for hyperaccumulator and non-
accumulator Thlaspi species in response to growth on a zinc-deficient
calcareous soil, Plant Sci. 164 (2003) 1095–1101.
[29] C.A. Grant, W.T. Buckley, L.D. Bailey, F. Selles, Cadmium accumu-
lation in crops, Can. J. Plant Sci. 78 (1998) 1–17.
[30] I. Cakmak, R.M. Welch, B. Erenoglu, V. Romheld, W.A. Norvell, L.V.
Kochian, Influence of varied zinc supply on retranslocation of cad-
mium (109Cd) and rubidium (86Rb) applied on mature leaf of durum
wheat seedlings, Plant Soil 219 (2000) 279–284.
[31] J.J. Hart, R.M. Welch, W.A. Norvell, L.V. Kochian, Transport inter-
actions between cadmium and zinc in root of bread and durum wheat
seedlings, Physiol. Plant. 116 (2002) 73–78.
[32] D.S. Bush, Calcium regulation in plant cells and its role in signaling,
Annu. Rev. Plant Physiol. Plant Mol. Biol. 46 (1995) 95–122.
[33] A.A.R. Webb, M.R. McAinsh, J.E. Taylor, A.M. Hetherington, Cal-
cium ions as intracellular second messengers in higher plants, Adv.
Bot. Res. 22 (1996) 45–96.
[34] E. Lombi, F.J. Zhao, S.P. McGrath, S.D. Young, G.A. Sacchi, Phy-
siological evidence for a high-affinity cadmium transporter highly
expressed in a Thlaspi caerulescens ecotype, New Phytol. 149 (2001)
50–53.
[35] A. Beck, K. Lendzian, M. Oven, Christmann, E. Grill, Phytochelatin
synthase catalyzes key step in turnover of glutathione conjugates,
Phytochemistry 6 (2003) 423–431.
TED
PR
O
PSL 7000 1–10
494




















