
Ulva lactuca Biofilters for Marine Fishpond Effluents. II. Growth Rate, Yield and C:N Ratio
8
Cohen and Neori: Ammonia uptake and nitrogen content of Viva lactuca 475 Botanica Marina Vol. 34, pp. 475-482, 1991 Ulva lactuca Biofilters for Marine Fishpond Effluents I. Ammonia Uptake Kinetics and Nitrogen Content I. Cohen 1 and A. Neori 2 Israel Oceanographic and Limnological Research, National Center for Mariculture, P.O. Box 1212 Eilat 88112, Israel (Accepted 18 July 1991) Abstract The potential of the seaweed Ulva lactuca as a biofilter for effluents of intensive marine fishponds in arid regions has been studied in Eilat, Israel. One kg (wet wt) nor 2 of Ulva in 1 m 2 tanks 60 cm deep, removed, during the day, over 90 percent of the ammonia from the fishpond effluents, at inflow fluxes up to 10 μιηο1β5 L" 1 h" 1 (0.14 mole m^d" 1 )· At fluxes of 40μπιοΐ68 L" 1 h~ 3 (0.58 mole m" 2 d" 1 ) 9 nearly 40 percent were removed. Night time ammonia removal remained efficient at low ammonia fluxes. Ammonia uptake rate and percent nitrogen (N) in dry weight (DW) of the seaweed showed hyperbolic correlations with ammonia flux through the tanks. The data fitted the Michaelis-Menten kinetics, with a maximal uptake rate of 28 μηιοΐβδ L" 1 h" 1 (0.4 mole m~ 2 d" 1 ). Pulse ammonia uptake rate was approximately four times that of continuous uptake. Maximal percent N in DW of U. lactuca, when stocked at 1 kg m" 2 , was 4.07. Increased stocking density (2, 4 and 6 kg m" 2 ) did not significantly affect ammonia uptake rate per tank. However, N-content was significantly higher, averaging 5.5 percent of DW. The data indicate that at low ammonia fluxes (below 10 μιηο1β8 L" 1 h" 1 ) U. lactuca was N-limited and that at higher fluxes the algae became gradually light limited. Our results indicate that 10m 2 of Ulva biofilter can remove from the effluents of our intensive grow-out ponds over 90 percent of the ammonia produced by 1 kg of daily feed ration or by approximately 75 kg of fish. stream eutrophication, which is particularly danger- Intensive culture of marine fishponds is being devel- ous in the highly oligotrophic and enclosed Gulf of oped at the National Center for Mariculture in Eilat, Eilat (Aqaba). Israel. The high densities of fish in the ponds cause Seaweeds have been shown to reduce efficiently the problems of water quality which in turn limit the level of seawater eutrophication in integrated mari- production (Krom et al. 1985 b). Ammonia is a major culture (Ryther et al. 1975). In a series of studies they metabolite produced by the fish and is toxic to them showed some seaweeds to have a high yield and nu- at high levels. It is also a major component in aqua- trient uptake capacity and to be very adequate for culture effluents, contributing to the eutrophication cultivation and harvesting. These advantages could of aquatic environments. Removal of excess ammonia be augmented by the economic potential of many and other inorganic nutrients can improve water qual- seaweeds in various industries. Motivated by these ity in the ponds, and at the same time reduce down- studies, Goldman (unpublished report) suggested the combination of colloid-rich seaweed culture with the fish culture effort in the arid south of Israel. Macroal- gal 'nutrient traps' have thus become a component in of "" e8rattd tert 1Mriculturc in our Corresponding author institute (Gordin 1982). Botanica Marina / Vol. 34 / 1991 / Fasc. 6 Copyright © 1991 Walter de Gruyter · Berlin · New York Brought to you by | Stockholms Universitet Authenticated | 130.237.165.22 Download Date | 9/13/12 9:10 AM
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Ulva lactuca Biofilters for Marine Fishpond Effluents. II. Growth Rate, Yield and C:N Ratio

Cohen and Neori: Ammonia uptake and nitrogen content of Viva lactuca 475
Botanica MarinaVol. 34, pp. 475-482, 1991
Ulva lactuca Biofilters for Marine Fishpond EffluentsI. Ammonia Uptake Kinetics and Nitrogen Content
I. Cohen1 and A. Neori2
Israel Oceanographic and Limnological Research, National Center for Mariculture, P.O. Box 1212Eilat 88112, Israel
(Accepted 18 July 1991)
Abstract
The potential of the seaweed Ulva lactuca as a biofilter for effluents of intensive marine fishponds in aridregions has been studied in Eilat, Israel. One kg (wet wt) nor2 of Ulva in 1 m2 tanks 60 cm deep, removed,during the day, over 90 percent of the ammonia from the fishpond effluents, at inflow fluxes up to 10 μιηο1β5L"1 h"1 (0.14 mole m^d"1)· At fluxes of 40μπιοΐ68 L"1 h~3 (0.58 mole m"2d"1)9 nearly 40 percent wereremoved. Night time ammonia removal remained efficient at low ammonia fluxes. Ammonia uptake rate andpercent nitrogen (N) in dry weight (DW) of the seaweed showed hyperbolic correlations with ammonia fluxthrough the tanks. The data fitted the Michaelis-Menten kinetics, with a maximal uptake rate of 28 μηιοΐβδL"1 h"1 (0.4 mole m~2d"1). Pulse ammonia uptake rate was approximately four times that of continuousuptake. Maximal percent N in DW of U. lactuca, when stocked at 1 kg m"2, was 4.07. Increased stockingdensity (2, 4 and 6 kg m"2) did not significantly affect ammonia uptake rate per tank. However, N-contentwas significantly higher, averaging 5.5 percent of DW. The data indicate that at low ammonia fluxes (below10 μιηο1β8 L"1 h"1) U. lactuca was N-limited and that at higher fluxes the algae became gradually light limited.Our results indicate that 10m2 of Ulva biofilter can remove from the effluents of our intensive grow-outponds over 90 percent of the ammonia produced by 1 kg of daily feed ration or by approximately 75 kg offish.
stream eutrophication, which is particularly danger-Intensive culture of marine fishponds is being devel- ous in the highly oligotrophic and enclosed Gulf ofoped at the National Center for Mariculture in Eilat, Eilat (Aqaba).Israel. The high densities of fish in the ponds cause Seaweeds have been shown to reduce efficiently theproblems of water quality which in turn limit the level of seawater eutrophication in integrated mari-production (Krom et al. 1985 b). Ammonia is a major culture (Ryther et al. 1975). In a series of studies theymetabolite produced by the fish and is toxic to them showed some seaweeds to have a high yield and nu-at high levels. It is also a major component in aqua- trient uptake capacity and to be very adequate forculture effluents, contributing to the eutrophication cultivation and harvesting. These advantages couldof aquatic environments. Removal of excess ammonia be augmented by the economic potential of manyand other inorganic nutrients can improve water qual- seaweeds in various industries. Motivated by theseity in the ponds, and at the same time reduce down- studies, Goldman (unpublished report) suggested the
combination of colloid-rich seaweed culture with thefish culture effort in the arid south of Israel. Macroal-gal 'nutrient traps' have thus become a component inof ""e8rattd tert 1Mriculturc in our
Corresponding author institute (Gordin 1982).
Botanica Marina / Vol. 34 / 1991 / Fasc. 6Copyright © 1991 Walter de Gruyter · Berlin · New York
Brought to you by | Stockholms UniversitetAuthenticated | 130.237.165.22
Download Date | 9/13/12 9:10 AM

476 Cohen and Neori: Ammonia uptake and nitrogen content of Viva tactuca
Our current research on ways to incorporate seaweedsinto intensive mariculture, considers the seaweed yieldas a bonus in a water treatment system whose mainproducts are stable water quality and clean effluents.This attitude (shared with Langton ei al. 1977 andsome others) is different than that in most other algalbiotechnology research enterprises, in which the algaeare the main or only product. Thus, we have chosenas the first candidate the local Ulva lactuca L. (Chlo-rophyta), not very valuable but sturdy, as our primaryseaweed, rather than cope with the biological obsta-cles of culturing a more valuable but less robust plant.
Various species of the genus Ulva have been culturedin the laboratory and outdoors. They grew well inboth artificial media and wastewater, showing a highcapacity to take up nutrients (Duke et al. 1989 a,Fujita 1985, Guist and Humm 1976, Parker 1981,Rosenberg 1981). We have cultured Ulva lactuca suc-cessfully in Eilat for long periods under field condi-tions (unpublished and Vandermeulen and Gordin1990) and its strong performance suggests that thisspecies might indeed be a suitable biofilter for fish-ponds.In the present study we quantify the operational fac-tors — nutrient concentration, water exchange rateand stocking density — controlling nutrient uptakefrom effluents of intensive fishponds by Ulva lactucaunder our field conditions and compare the resultswith other studies. This is an essential step for thedesign and operation of larger scale facilities.
Material and Methods
Thalli of Ulva lactuca L. (Vandermeulen and Gordin1990), were collected from the Gulf of Eilat (Aqaba)and maintained outdoors in running seawater. Spec-imens have been deposited in the herbarium of theUniversity of La Laguna, Tenerife, Spain (Dr RicardoHaroun) and live and preserved samples have beenmaintained in our laboratory for later investigations.
All the experiments were conducted in 1 m2, 600 Ltanks (Vandermeulen 1989). The algae were grownunattached and kept suspended in the water columnby air diffusers situated at the bottom of each tank.Fishpond effluents were pumped into a large holdingtank (15 m3) and then flowed by gravity to the ex-perimental tanks. Temperatures were measured byminimum/maximum thermometers in each tank, butthe temperature record is incomplete due to the break-down of several thermometers. Light data were kindlyprovided by the Israeli Meteorological Service froma nearby facility.
In total five experiments are reported. At the begin-ning of each experiment the tanks were stocked at afixed wet weight of centrifuged thalli and each treat-ment (in triplicates, except where indicated otherwise)was provided with a fixed flow-rate of fishpond ef-fluents. The algae were harvested weekly, centrifugedin a converted washing machine and restocked at theoriginal stocking density. The results of growth rate,yield and C: N ratio will be reported in the accom-panying paper (Neori et al. 1991).Samples for dry weight determination were oven dried(60 °C, 24 hours). Samples for CHN analyses (Perkin-Elmer Model 240) were rinsed with deionized water,dried and ground to a fine powder with a mortar andpestle. The rinsing step lead to a small (< 3%) lossof dry weight (DW), probably due to the salts in theseawater. Ammonia-N concentration (practically theonly inorganic N dissolved in the fishpond water, seePorter et al. 1987 and Neori et al. 1989) was measuredat the inflow and at the outflow water of each tank2 — 3 times a week at 9:00 am. At this time the con-centration was representative of daytime values ascan be learned from the diel variation in ammoniaconcentration in the algal tanks and in the fishpondeffluents reported later. The water samples were fil-tered (GF/F, Whatmann) and analyzed for total am-monia by a Technicon AA-2 autoanalyzer (Krom etal. 1985 a). The uptake rate was determined asV = f(Si-So) where V = uptake rate (μηιο1β8 L"1
h"1), f = water flow rate (number of water exchangeshr1), Si = inflow arnmonia-N concentration and So= ammonia-N concentration at the outflow (residualN). The Vmax and Ks were calculated from the linearS/V vs S plot (Atkins and Nimmo 1980). Overnightmeasurements at 3 h intervals were made twice duringan experiment.Statistical correlations and F-tests were made by theGeneral Linear Model option of S AS. Since therewere occasionally large differences in the inflowingconcentration of ammonia between weeks, we havetreated each week as a separate experimental unit andthe data are presented accordingly. Data were aver-aged for each week in the Tables, but all the datawere used in the analyses and plotted in the Figures.
To assess the importance of environmental differencesbetween experiments we used in experiments 1, 2 and3 a standard treatment with 1 kg of Ulva lactuca and4 daily exchanges of ambient seawater. Inorganic nu-trients, 2 mM of ammonia-N (as NH4C1) and 0.15 ΙΪΙΜof P (as NaH2PO4), were supplied weekly, while stop-ping water exchange for 18 h (Vandermeulen andGordin 1990). The rate of ammonia disappearanceduring the first 3 h of the nutrient supply was usedto calculate maximal values of 'pulse' uptake rate.
Botanica Marina / Vol. 34 / 1991 / Fasc. 6Brought to you by | Stockholms Universitet
Authenticated | 130.237.165.22Download Date | 9/13/12 9:10 AM

Cohen and Neori: Ammonia uptake and nitrogen content of Viva lactuca 477
Water exchange rate
Four experiments were conducted to determine theeffects of exchange rate of fishpond effluents in thealgal tanks on ammonia-N uptake by Ulva lactuca.Each tank was stocked with 1 kg of the seaweed. Inthe November 1987 experiment (exp. 1) water ex-change rate was 4/day. In two experiments, in Janu-ary, 1988 (exp. 2) and in February, 1988 (exp. 3) 4, 8and 16 daily water exchanges were compared (induplicates). In the last of the series, in January, 1989(exp. 4), daily water exchange rates compared were4, 8, and 12, and no CHN-samples were taken.
Stocking density
The effect of high stocking densities of Ulva lactuca(above 1 kg per tank) on the rate of ammonia-Nuptake at high ammonia-N fluxes was studied in May-June, 1988 (exp. 5). The tanks were stocked with 2, 4and 6 kg of U. lactuca and maintained at 12 dailywater exchanges of fishpond effluents.
Results
Water exchange rate
The water exchange rate experiments were all carriedout in the winter to reduce the effect of the variationin environmental factors on the results. We also foundhighly insignificant correlation (r2 = 0.0115, P = 0.7)between total daily irradiance and ammonia uptakein the combined data from the standard treatmentsof experiments 1, 2 and 3 (Table I).
Ammonia-N levels in the inflowing fishpond effluentsvaried markedly between experiments and, to a lesserextent, even between weeks in some experiments (Ta-ble II). The effect of ammonia-N concentration in theinflowing water can be seen by comparing treatmentswith similar water exchange rates between weeks andexperiments. The effect of water exchange rate, al-though not independent of ammonia-N concentra-
Table I. Total daily irradiance and pulse 18 h ammoia-N uptakein the standard treatment of three uptake experiments withUlva lactuca in 600 liter, 1 m2 tanks.
Exp.No.
1
2
3
Week
112
12
Total irradiance(Watt h m-? d-1)
4645
51603425
69106625
Pulse ammoia uptake(limoles L"1
18 h ± SE)
654 + 100
393 ± 9377 ± 21
596 + 44334 ± 8
per
tion, can be seen by comparing treatments withineach week. Outflow concentration of ammonia-N cor-related positively with both water exchange rate andinflowing ammonia-N concentration whereas percentuptake (removal efficiency) of ammonia-N by theseaweed correlated negatively with the same variables(p < 0.05). The percent uptake of ammonia-N by theseaweeds during day time was high at most levels ofwater flow rate and inflow ammonia-N concentration.Maximal removal efficiency, up to 96 percent, wasobtained at low water exchange rates and ammonia-N concentrations of experiment 2. The removal effi-ciency dropped within each experiment with increas-ing water exchange rate. It also dropped betweenexperiments with increasing ammonia concentrationin the inflowing water. Thus, the removal efficiencydropped down to 39 percent at the highest waterexchange rates and ammonia-N concentrations inexperiment 4.Nevertheless, V (the product of inflowing ammonia-N concentration, water exchange rate and the fractionof ammonia-N uptake in moles per tank, per liter orper unit area), increased both with increasing waterexchange rate in each experiment and with increasingammonia concentration in the inflowing water be-tween experiments (P < 0.05). Most likely the valueof V actually depends on the in situ concentration ofnutrients in the tanks. The latter value is determinedby three parameters, V itself,-ammonia concentrationin the inflowing water and water, exchange rate. Thetwo last parameters, multiplied by each other, produceammonia-N flux as suggested by Fujita and Goldman(1985). They had found that the growth rate of Gra-cilaria tikvahiae McLachlan was predicted better byN-flux than by either flow rate or N-concentrationalone. Nutrient flux reflects the interdependency ofthe effects of flow rate and nutrient concentrationand describes better the availability of nutrients tothe algae.The dependence of V on ammonia-N flux in Ulvalactuca (Fig. la) shows a positive hyperbolic satura-tion, conforming with the Michaelis-Menten type ki-netics. The values of Vmax and Ks which form thecurve are 28 μτηο1β8 L"1 h~! and 27.5 μιηοΐ65 L""1
h"1, respectively (r2 = 0.64; n = 206). Complete am-monia removal at low ammonia fluxes is indicated bythe initial slope (Vmax/Ks) of close to unity.Plotting uptake rates as a function not of the am-monia-N flux but of the in situ ammonia-N concen-tration, or residual ammonia-N (Fig. 1 b), shows alsoa positive hyperbolic correlation which fits the Mi-
' chaelis-Menten kinetics (Vmax =13.8 μπιο1β8 L"1
h-1; Ks = 5.2 μΜ; r2 = 0.81). The points are, how-ever, much more scattered at the low concentration
Botanica Marina / Vol. 34 / 1991 / Fasc. 6 Brought to you by | Stockholms UniversitetAuthenticated | 130.237.165.22
Download Date | 9/13/12 9:10 AM
478 Cohen and Neori: Ammonia uptake and nitrogen content of Ulva lactuca
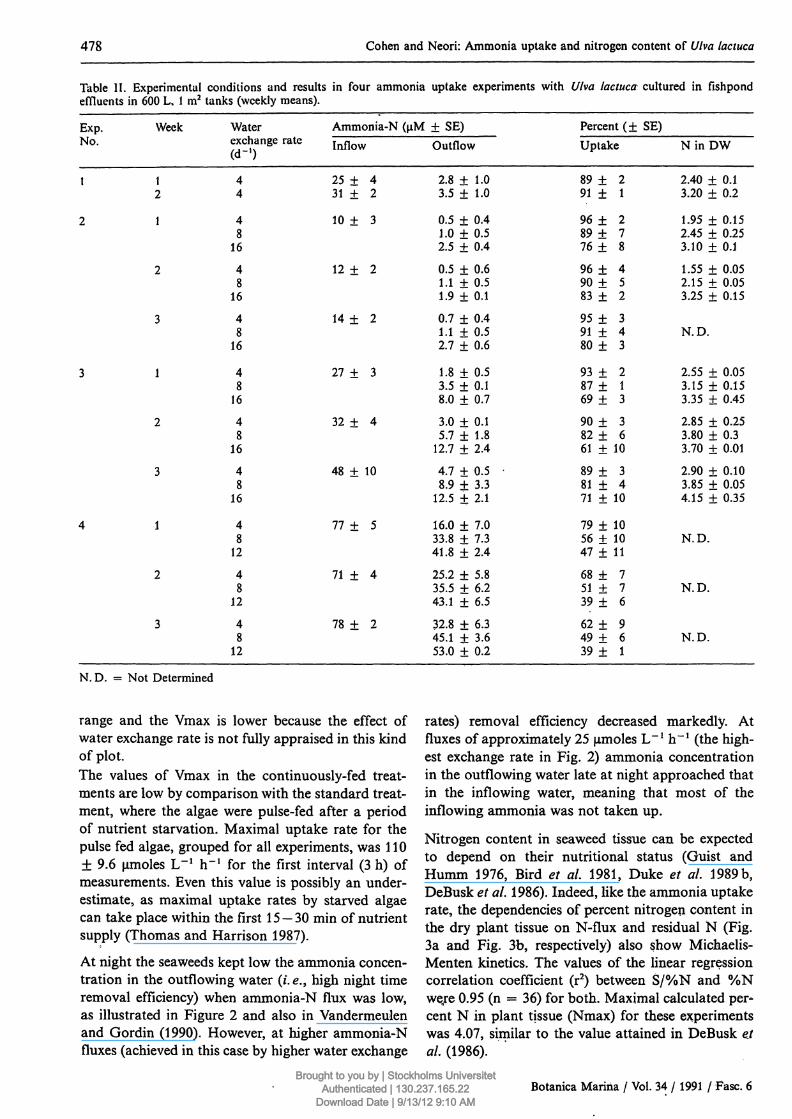
Table II. Experimental conditions and results in four ammonia uptake experimentseffluents in 600 L, 1 m2 tanks (weekly means).
Exp.No.
1
2
3
4
Week
12
1
2
3
1
2
3
1
2
3
Waterexchange r(d-1)
44
48
16
48
16
48
16
48
16
48
16
48
16
48
12
48
12
48
12
Ammonia-N (μΜ ± SE)ate Inflow Outflow
2 5 + 4 2.8 ± 1.031 ± 2 3.5 ± 1.0
10 4- 3 0.5 ± 0.41.0 ± 0.52.5 ± 0.4
1 2 + 2 0.5 ± 0.61.1 ± 0.51.9 ± 0.1
1 4 + 2 0.7 ± 0.41.1 + 0.52.7 ± 0.6
2 7 + 3 1.8 + 0.53.5 + 0.18.0 ± 0.7
3 2 + 4 3.0 ± 0.15.7 + 1.8
12.7 ± 2.4
48 + 10 4.7 + 0.58.9 ± 3.3
12.5 ± 2.1
7 7 + 5 16.0 ± 7.033.8 ± 7.341.8 ± 2.4
7 1 + 4 25.2 + 5.835.5 ± 6.243.1 ± 6.5
7 8 + 2 32.8 + 6.345.1 + 3.653.0 + 0.2
with Ulva lactuca cultured in fishpond
Percent (+ SE)Uptake
89 ± 291 + 1
9 6 + 28 9 + 77 6 + 8
9 6 + 490 ± 583 ± 2
9 5 + 391 ± 480 ± 3
9 3 + 287 + 169 ± 3
9 0 + 382 ± 661 ± 10
8 9 + 381 ± 471 + 10
79 ± 1056 + 1047 + 11
6 8 + 75 1 + 739 ± 6
6 2 + 94 9 + 639 ± 1
N i n D W
2.40 + 0.13.20 ± 0.2
1.95 + 0.152.45 + 0.253.10 ± 0.1
1.55 ± 0.052.15 ± 0.053.25 ± 0.15
N.D.
2.55 + 0.053.15 + 0.153.35 + 0.45
2.85 + 0.253.80 + 0.33.70 ± 0.01
2.90 + 0.103.85 ± 0.054.15 ± 0.35
N.D.
N.D.
N.D.
N. D. = Not Determined
range and the Vmax is lower because the effect ofwater exchange rate is not fully appraised in this kindof plot.The values of Vmax in the continuously-fed treat-ments are low by comparison with the standard treat-ment, where the algae were pulse-fed after a periodof nutrient starvation. Maximal uptake rate for thepulse fed algae, grouped for all experiments, was 110± 9.6 μπιοΐ68 L"1 h"1 for the first interval (3 h) ofmeasurements. Even this value is possibly an under-estimate, as maximal uptake rates by starved algaecan take place within the first 15 — 30 min of nutrientsupply (Thomas and Harrison 1987).
At night the seaweeds kept low the ammonia concen-tration in the outflowing water (i.e., high night timeremoval efficiency) when ammonia-N flux was low,as illustrated in Figure 2 and also in Vandermeulenand Gordin (1990). However, at higher ammonia-Nfluxes (achieved in this case by higher water exchange
rates) removal efficiency decreased markedly. Atfluxes of approximately 25 μιηοΐββ L"1 h"1 (the high-est exchange rate in Fig. 2) ammonia concentrationin the outflowing water late at night approached thatin the inflowing water, meaning that most of theinflowing ammonia was not taken up.
Nitrogen content in seaweed tissue can be expectedto depend on their nutritional status (Guist andHumm 1976, Bird et al. 1981, Duke et al. 1989 b,DeBusk et al. 1986). Indeed, like the ammonia uptakerate, the dependencies of percent nitrogen content inthe dry plant tissue on N-flux and residual N (Fig.3a and Fig. 3b, respectively) also show Michaelis-Menten kinetics. The values of the linear regressioncorrelation coefficient (r2) between S/%N and %Nwejre 0.95 (n = 36) for both. Maximal calculated per-cent N in plant tissue (Nmax) for these experimentswas 4.07, similar to the value attained in DeBusk etal. (1986).
Botanica Marina / Vol. 34 /1991 / Fasc. 6Brought to you by | Stockholms Universitet
Authenticated | 130.237.165.22Download Date | 9/13/12 9:10 AM
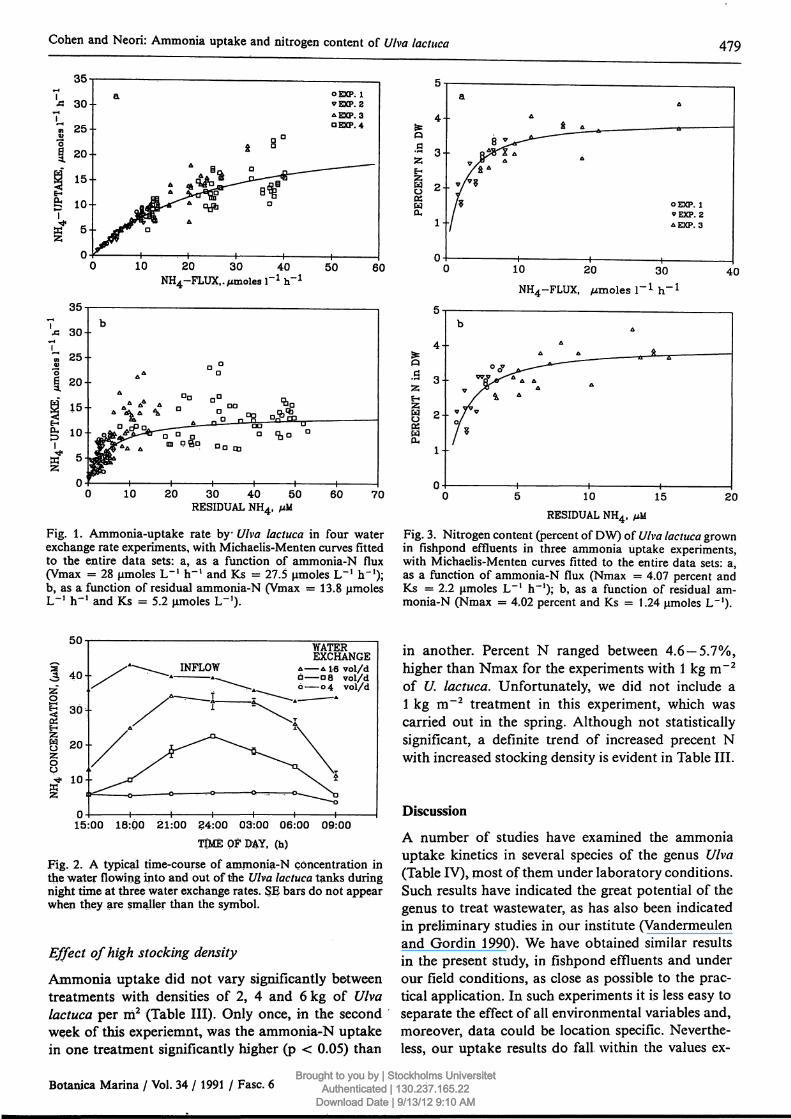
Cohen and Neori: Ammonia uptake and nitrogen content of Ulva lactuca 479
10 20 30 40NH4-FLUX..Mmoles Γ1 h"1
50 60
10 20 30 40 50RESIDUAL NH4, μΜ
60 70
Fig. 1. Ammonia-uptake rate by Ulva lactuca in four waterexchange rate experiments, with Michaelis-Menten curves fittedto the entire data sets: a, as a function of ammonia-N flux(Vmax = 28 μηιο1ε5 L"1 h"1 and Ks = 27.5 μιηο1β8 L"1 h"1);b, as a function of residual ammonia-N (Vmax = 13.8L"1 h-1 and Ks = 5.2 μπιοΐεβ L'1).
10 20 30
NH4-FLUX, pinoles r1 h"140
RESIDUAL NH4, yuM
Fig. 3. Nitrogen content (percent of DW) of Ulva lactuca grownin fishpond effluents in three ammonia uptake experiments,with Michaelis-Menten curves fitted to the entire data sets: a,as a function of ammonia-N flux (Nmax = 4.07 percent andKs = 2.2 μιηοΐ65 L"1 h""1); b, as a function of residual am-monia-N (Nmax = 4.02 percent and Ks = 1.24 μπκ>1β$ L"1)·
WATEREXCHANGE
Δ Δ 16 vol/dα α 8 vol/dό o4 vol/d
15:00 18:00 21:00 24:00 03:00 06:00 09:00TIME OF DAY, (h)
Fig. 2. A typical time-course of ammonia-N concentration inthe water flowing into and put of the Ulva lactuca tanks duringnight time at three water exchange rates. SE bars do not appearwhen they are smaller than the symbol.
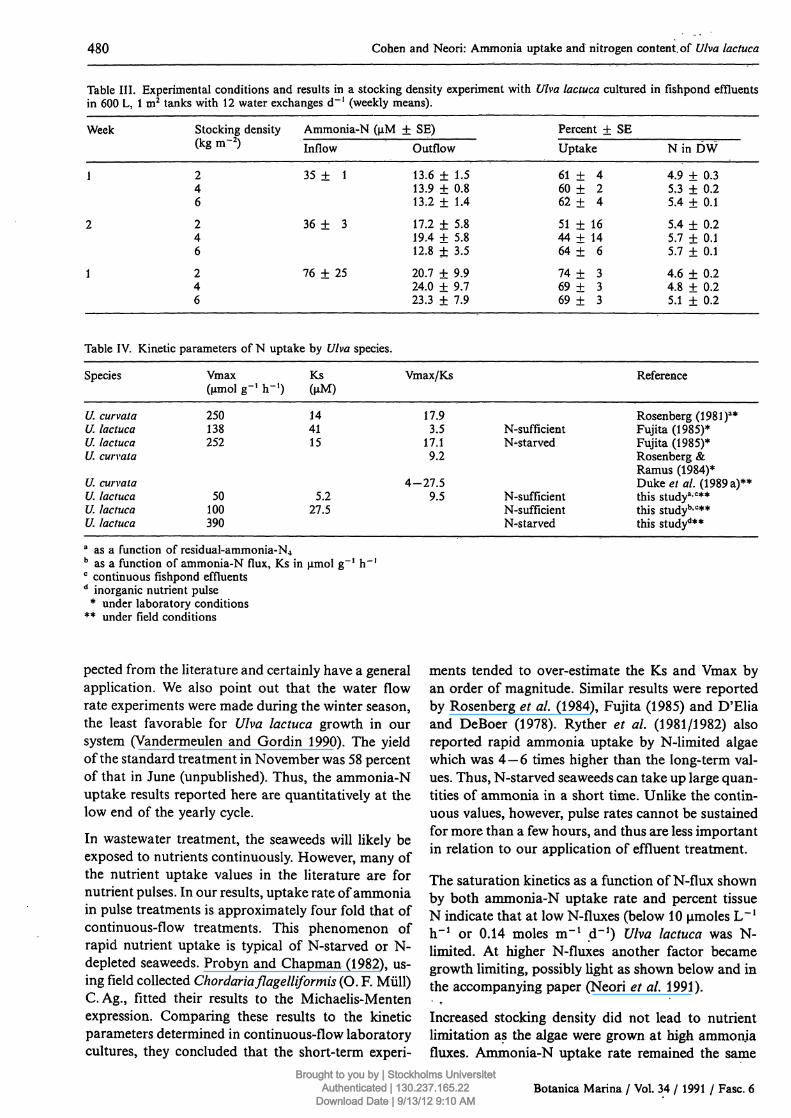
Effect of high stocking density
Ammonia uptake did not vary significantly betweentreatments with densities of 2, 4 and 6 kg of Ulvalactuca per m2 (Table III). Only once, in the secondweek of this experiemnt, was the ammonia-N uptakein one treatment significantly higher (p < 0.05) than
in another. Percent Ν ranged between 4.6 — 5.7%,higher than Nmax for the experiments with 1 kg m~2
of U. lactuca. Unfortunately, we did not include a1 kg m~2 treatment in this experiment, which wascarried out in the spring. Although not statisticallysignificant, a definite trend of increased precent Νwith increased stocking density is evident in Table III.
Discussion
A number of studies have examined the ammoniauptake kinetics in several species of the genus Ulva(Table IV), most of them under laboratory conditions.Such results have indicated the great potential of thegenus to treat wastewater, as has also been indicatedin preliminary studies in our institute (Vandermeulenand Gordin 1990). We have obtained similar resultsin the present study, in fishpond effluents and underour field conditions, as close as possible to the prac-tical application. In such experiments it is less easy toseparate the effect of all environmental variables and,moreover, data could be location specific. Neverthe-less, our uptake results do fall within the values ex-
Botanica Marina / Vol. 34 / 1991 / Fasc. 6Brought to you by | Stockholms Universitet
Authenticated | 130.237.165.22Download Date | 9/13/12 9:10 AM
480 Cohen and Neori: Ammonia uptake and nitrogen content, of Uha lactuca
Table III. Experimental conditions and results in a stocking density experiment with Ulva lactuca cultured in fishpond effluentsin 600 L, l m2 tanks with 12 water exchanges d~! (weekly means).
Week
1
2
1
Stocking density(kg m-2)
246
246
246
Ammonia-N (μΜInflow
35 ± 1
36 ± 3
76 ± 25
±SE)Outflow
13.6 ± 1.513.9 ± 0.813.2 ± 1.4
17.2 + 5.819.4 + 5.812.8 ± 3.5
20.7 + 9.924.0 + 9.723.3 + 7.9
Percent ± SEUptake
6 1 + 46 0 + 262 ± 4
51 ± 1644 + 1464 ± 6
7 4 + 36 9 + 36 9 + 3
N i n D W
4.9 + 0.35.3 + 0.25.4 + 0.1
5.4 + 0.25.7 + 0.15.7 ± 0.1
4.6 ± 0,24.8 + 0.25.1 + 0.2
Table IV. Kinetic parameters of Ν uptake by Ulva species.
Species
{/. curvataU. lactucaU. lactucaU. curvata
U. curvataU. lactucaU. lactucaU. lactuca
Vmax(μπιοί g""1 h"1)
250138252
50100390
Ks(μΜ)
144115
5.227.5
Vmax/Ks
17.93.5
17.19.2
4-27.59.5
N-sufficientN^starved
N-sufficientN-sufflcientN-starved
Reference
Rosenberg (1 98 l)a*Fujita (1985)*Fujita (1985)*Rosenberg &Ramus (1984)*Duke et al. (1989 a)**this study3·0**this studyb-c**this studyd**
a as a function of residual-ammonia-N4b as a function of ammonia-N flux, Ks in μπιοί g"1 h~ !c continuous fishpond effluentsd inorganic nutrient pulse
* under laboratory conditions** under field conditions
pected from the literature and certainly have a generalapplication. We also point out that the water flowrate experiments were made during the winter season,the least favorable for Ulva lactuca growth in oursystem (Vandermeulen and Gordin 1990). The yieldof the standard treatment in November was 58 percentof that in June (unpublished). Thus, the ammonia-Nuptake results reported here are quantitatively at thelow end of the yearly cycle.
In wastewater treatment, the seaweeds will likely beexposed to nutrients continuously. However, many ofthe nutrient uptake values in the literature are fornutrient pulses. In our results, uptake rate of ammoniain pulse treatments is approximately four fold that ofcontinuous-flow treatments. This phenomenon ofrapid nutrient uptake is typical of N-starved or N-depleted seaweeds. Probyn and Chapman (1982), us-ing field collected Chordariaflagelliformis (O. F. M ll)C.Ag., fitted their results to the Michaelis-Mentenexpression. Comparing these results to the kineticparameters determined in continuous-flow laboratorycultures, they concluded that the short-term experi-
ments tended to over-estimate the Ks and Vmax byan order of magnitude. Similar results were reportedby Rosenberg et al. (1984), Fujita (1985) and D'Eliaand DeBoer (1978). Ryther et al. (1981/1982) alsoreported rapid ammonia uptake by N-limited algaewhich was 4—6 times higher than the long-term val-ues. Thus, N-starved seaweeds can take up large quan-tities of ammonia in a short time. Unlike the contin-uous values, however, pulse rates cannot be sustainedfor more than a few hours, and thus are less importantin relation to our application of effluent treatment.
The saturation kinetics as a function of N-flux shownby both ammonia-N uptake rate and percent tissueN indicate that at low N-fluxes (below 10 μιηοΐεβ L"1
h"1 or 0.14 moles m""1 d""1) Ulva lactuca was N-limited. At higher N-fluxes another factor becamegrowth limiting, possibly light as shown below and inthe accompanying paper (Neori et al. 1991).
Increased stocking density did not lead to nutrientlimitation as the algae were grown at high ammoniafluxes. Ammonia-N uptake rate remained the same
Botanica Marina / Vol. 34 / 1991 / Fasc. 6Brought to you by | Stockholms Universitet
Authenticated | 130.237.165.22Download Date | 9/13/12 9:10 AM

Cohen and Neori: Ammonia uptake and nitrogen content of Ulva lactuca 481
for all treatments in the stocking density experiment.However, percent N in algal tissue here was higherthan for the 1 kg m~2 experiments and a positivecorrelation was suggested between percent N andstocking density. Neori et al (1991) found stockingdensity did not influence percent carbon in the sameexperiment. Thus the carbon limitation of the algae,suggested for this experimental system by Vander-meulen and Gordin (1990), is not supported by ourresults. The seasonal difference between spring andwinter is also unlikely to account for the observedhigh percent tissue N in the stocking density experi-ment compared with the water exchange rate experi-ments. Actually, if the effects were only seasonal thevalues of percent tissue N should have decreased asa result of the better temperature and light conditionsin the spring and not increased, as observed here.Duke et al. (1986 and 1989 a) showed in studies withUlva curvata (K tz.) de Toni and Codiitm decorticatum(Woodw.) Howe, that improved light and temperatureconditions decreased percent tissue N in the seaweeds.These findings can be reconciled with the increasedpercent N in our spring experiment, based on thestudy of Vymazal (1987) on optimal stocking density.The super-optimal stocking densities (2, 4 and 6 kgm~2) in our experiment No.· 5 must have led to re-duced mean light availability in the tanks, in spite ofit taking place in the spring, relative to the optimally-stocked tanks of the winter experiments 1—4. Van-dermeulen and Gordin (1990), using the very sametanks, indeed reported a reduction in photon fluxdensity at the bottom of 89% and 100% for tankswith 1 kg and 4 kg of Ulva, respectively.
In conclussion, the robust seaweed Ulva lactuca couldcontinuously and efficiently strip nearly all the am-monia from fishpond effluents at low nutrient fluxes(below 10 iimoles L"1 h~1 or 0.14 moles m~2 d"1)·
The rate of ammonia uptake increased at higher am-monia fluxes to 28 μπιο1β8 L"1 h""1 (or 0.4 moles m~2
d"1), at the cost of reduced removal efficiency, espe-cially at night. Nitrogen content of the seaweeds in-creased with increased ammonia flux and with in-creased stocking Density. Ammonia uptake rate pertank was insensitive to algal stocking density in therange of 1 to 6 kg m ~2.
Ulva biofilters can operate both as recirculating unitsof intensive fishponds and as units for polishing ef-fluents before their release. The data in this paperallow the design of pilot-size Ulva biofilters for inten-sive seawater fishponds. Our calculations show that10 m2 of the seaweed biofilter can take up completelyand in one stage the ammonia produced in the fish-pond from 1 kg of daily feed. This quantity feedsapproximately 75 kg of Spams aurata L., the fishgrown in our grow-out ponds. A multi-stage biofiltersystem of similar size (10m2), designed to use thehigher uptake capacity of the seaweed at high am-monia fluxes, can treat the effluents of even twicethat weight of fish.
Acknowledgements
We are grateful to A. Davidson for chemical analyses,to H. Voet for help with statistical analyses and tothe staff of the NCM for their assistance. H. Gordinprovided encouragement and facilitated this research.M. Friedlander and C. Dawes critically reviewed ear-lier versions of this manuscript. This research wassupported by the Israeli Ministry for Energy andInfrastructure Mariculture Project, The Arava Re-search and Development Fund, BARD research grantNo. 1-1634-89 and grant No. 3035189 from the IsraeliMinistry for Science and Technology.
ReferencesAtkins, G. L. and I. A. Nimmo. 1980. Current trends in the
estimation of Michaelis-Menten parameters. Anal. Biochem.104: 1-9.
Bird, K. T., M. D. Hanisak and J. H. Ryther. 1981. Chemicalquality and production of agars extracted from Gracilariatikvahiae grown in different nitrogen enrichment conditions.Bot. Mar. 7*441-444.
DeBusk, Τ. Α., M. Blakeslee and J. H. Ryther. 1986. Studieson the outdoor cultivation of Ulva lactuca L. Bot. Mar. 24:381-386.
D'Elia, D. R and J. A. DeBoer. 1978. Nutritional studies oftwo red algae. II. kinetics of ammonium and nitrate uptake./. Phycol. 14: 266-272.
Duke, C. S., B. E. Lap inte and J. Ramus. 1986. Effects oflight on growth, RuBPase activity and chemical compositionof Ulva species (Chlorophyta)". /. Phycol. 22: 362-370.
Botanica Marina / Vol. 34 / 1991 / Fasc. 6
Duke, C. S., W. Litaker and J. Ramus. %1989a. Effects of thetemperature, nitrogen supply and tissue nitrogen on am-monium uptake rates of the Chlorophyte seaweeds Ulvacurvata and Codium decorticatum. J. Phycol. 25: 113 — 120.
Duke, C. S., W. Litaker and J. Ramus. 1989b. Effect of tem-perature on nitrogen limited growth rate and chemical com-position of Ulva curvata (UlVales: Chlorophyta). Mar. Biol.100: 143-150.
Fujita, R. M. 1985. The role of nitrogen status in regulatingtransient ammonium uptake and nitrogen storage by ma-croalgae. /. Exp. Mar. Biol. Ecol. 92: 283-301.
Fujita, R. M. and J. C. Goldman. 1985. Nutrient flux andgrowth of the red alga Gracilaria tikvahiae McLachlan (Rho-dophyta). Bot. Mar. 28: 265-268.
Gordin, H. 1982. Aquaculture: potential developemnt. In: (RG. Brewer, ed) Oceanography: Present and Future. Springer-Verlag, New York. pp. 347-361.
Brought to you by | Stockholms UniversitetAuthenticated | 130.237.165.22
Download Date | 9/13/12 9:10 AM

482 Cohen and Neori: Ammonia uptake and nitrogen content of Ulva lactuca
Guist, G. G. Jr. and H. J. Humm. 1976. Effects of Sewageeffluents on growth of Ulva lactuca. BioL Sei. 4: 267-271.
Krom, M. D., S. Grayer and A. Davidson. 1985 a. An auto-mated method of ammonia determination for use in mari-cullure. Aquaculture 44: 153 — 160.
Kjom, M. D., C. B. Porter and H. Gordin. 1985 b. Causes offish mortalities in scmi-intensivcly operated seawater pondsin Eilat, Israel. Aquaculture 49: 159-177.
Langton, R. W, K. C. Haines and R. E. Lyon. 1977. Ammonia-nitrogen produced by the bivalve mollusc Tapes japonicaand its recovery by the red seaweed Hypnca musciformis ina tropical mariculture system. Helgoländer wiss. Meeresun-fers. 30:211-229.
Neori. A., I. Cohen and H. Gordin. 1991. Ulva laciuca biofiltersfor marine fishpond effluents. II. growth rate, yield andC : N ratio. Bot. Mar. 34: 483-489.
Neori, A., M. D. Krom, I. Cohen and H. Gordin. 1989. Waterquality conditions and paniculate chlorophyll a of newintensive seawater fishponds in Eijat, Israel: daily and dielvariations. Aquaculture 80: 63 — 78.
Parker, H. S. 1981. Influence of relative water motion on thegrowth, ammonium uptake and carbon composition of Ulvalaciuca (Chlorophyta). Mar. Biol. 63: 309-318.
Porter, C. B., M. D. Krom, M. G. Robbins, L. Brickeil and A.Davidson. 1987. Ammonia excretion and total N budgetfor gilthead seabrcam (Spams aurata) and its effect on waterquality conditions. Aquaculture 66: 287 — 297.
Probyn, T. A. and A. R. O. Chapman. 1982. Nitrogen uptakecharacteristics of Chordaria flagelliformis (Phaeophyta) inbatch mode and continuous mode experiments. Mar. Biol.71: 129-133.
Rosenberg, G. 1981. Ecological growth strategies in the redseaweed Gracilariafoliifera (Rhodophyceae). Ph.D. thesis,Yale University, pp. 151.
Rosenberg, G., T. A. Probyn and K. H. Mann. 1984. Nutrientuptake and growth kinetics in brown seaweeds: response tocontinuous and single additions of ammonium. J. Exp. Mar.BioLEcol.80: 125-146.
Ryther, J. H., J. C. Goldman, C. E. Gifford, J. E. Huguenin,A. S. Wing, J. P. Clarner, L. D. Williams and B. E. Lapointc.1975. Physical models of integrated waste recycling marinepolyculture systems. Aquaculture 5: 163 — 177.
Ryther, J. H., N. Corwin, T. A. DeBusk and L. D. Williams.1981/1982. Nitrogen uptake and storage by the red algaGracilaria tikvahiae (McLachlan, 1979). Aquaculture 26:107-115.
Thomas, T. E. and P. J. Harrison. 1987. Rapid ammoniumuptake and nitrogen interactions in five intertidal seaweedsgrown under field conditions. Exp. Mar. Biol. Ecol. 107:1-8.
Vandermeulen, H. 1989. A low-maintenance tank for massculture of seaweeds. Aqua Eng. 8: 67—71.
Vandermeulen, H. and H. Gordin. 1990. Ammonium uptakeusing Ulva (Chlorophyta) in intensive fishpond systems:mass culture and treatment of effluent. /. Appl. Phycol. 2:363-374.
Vymazal, J. 1987. Ammonium uptake and biomass interactionin Chladophora glomerata (Chlorophyta). Br. Phycol. J. 22:163-167.
Botanica Marina / Vol. 34 / 1991 / Fasc. 6Brought to you by | Stockholms Universitet
Authenticated | 130.237.165.22Download Date | 9/13/12 9:10 AM







![Tetra-kis(μ(2)-cyanido-κ(2)C:N)dicyanido-tetra-kis-[tris-(2-amino-eth-yl)amine-κ(3)N,N',N'',N''']tetra-copper(II)iron(II) bis[pentacyanidonitrosoferrate(II)] hexahydrate](https://static.fdokumen.com/doc/165x107/634199118718ae62200b4f38/tetra-kism2-cyanido-k2cndicyanido-tetra-kis-tris-2-amino-eth-ylamine-k3nnnntetra-copperiiironii.jpg)




