
Oxidative stress and phytochelatin characterisation in bread wheat exposed to cadmium excess
10
Original article Oxidative stress and phytochelatin characterisation in bread wheat exposed to cadmium excess Annamaria Ranieri a , Antonella Castagna a , Francesca Scebba a , Maria Careri b , Ingrid Zagnoni b , Giovanni Predieri b , Massimo Pagliari b , Luigi Sanità di Toppi c, * a Dipartimento di Chimica e Biotecnologie Agrarie, Università di Pisa, via del Borghetto 80, 56124 Pisa, Italy b Dipartimento di Chimica Generale ed Inorganica, Chimica Analitica, Chimica Fisica, Università di Parma, Parco Area delle Scienze 17/A, 43100 Parma, Italy c Dipartimento di Biologia Evolutiva e Funzionale, Sezione di Biologia Vegetale, Università di Parma, ParcoArea delle Scienze 11/A, 43100 Parma, Italy Received 21 September 2004; accepted 12 December 2004 Available online 07 January 2005 Abstract In this work, we first investigated if the bread wheat (Triticum aestivum L.) cv. Albimonte can be defined as “shoot cadmium excluder”—by comparing the cadmium (Cd) content in leaves and roots and by calculating the shoot-to-root Cd concentration ratio. Furthermore, we evalu- ated if the exposure to Cd excess could generate oxidative stress in leaves and roots of this cv., in terms of hydrogen peroxide (H 2 O 2 ) accumulation, NAD(P)H oxidation rate, and variations in reduced glutathione (GSH) content and peroxidase (POD, EC 1.11.1.7) activity. Finally, we surveyed possible quali- quantitative differences in thiol-peptide compound pattern between roots and leaves, in order to verify whether phytochelatins (PCs) and related thiol-peptides could contribute in limiting the Cd-induced oxidative stress. Unambiguous charac- terisation of PCs and related forms present in the root samples was obtained by electrospray ionisation mass spectrometry (ESI-MS) and ESI-tandem MS (ESI-MS/MS). Our results indicate that in leaves the stress generated by the low accumulation of Cd (due to a moderate translocation in planta) seems to be counteracted by the antioxidant response and by the PC biosynthesis. On the contrary, in roots, in spite of the elevated presence of PCs and related thiol-peptide-compounds, the excess of Cd causes a decline in the antioxidant protection of the organ, with the consequent generation of considerable amounts of H 2 O 2 , a direct agent of oxidative stress. © 2005 Elsevier SAS. All rights reserved. Keywords: Cadmium; Hydrogen peroxide; NAD(P)H oxidase; Peroxidase; Phytochelatins; Triticum aestivum; Wheat 1. Introduction Cadmium (Cd) is a non-essential heavy metal (HM), released into the environment by anthropogenic and non- anthropogenic sources [23,25,34]. Cadmium long biological half-life and its tendency to form strong complexes with S-containing peptides/proteins renders this metal a notable hazard to a wide range of organisms [39]. In fact, Cd can be transferred to animals and humans through the food chain [42], and concerns about consumption of Cd-containing foods are assuming a relevant dimension [8,21]. In many countries, increases in soil Cd concentrations over time are conspicu- ous, due to the addition of fertilisers, animal manures, sew- age sludges, amendments and the atmospheric fall-out [1,21,22]. Cd accumulation in cereals aimed for food produc- tion, i.e. Triticum aestivum L. (bread wheat), can represent indeed a worrying risk for animal and human health [8,21], and the fact that Cd is often a by-product of phosphate ferti- lisers [23,25], which are largely utilised in cereal cultivation, has further raised debates and questions on Cd risk manage- ment of these important crops. Abbreviations: Cd, cadmium; desGlyPC, desglycil-phytochelatin; DPI, diphenylene iodonium; DTT, dithiothreitol; EDTA, ethylenediaminetetraa- cetic acid; ESI-MS, electrospray ionisation mass spectrometry; ESI- MS/MS, electrospray ionisation tandem mass spectrometry; GSH, reduced glutathione; HM, heavy metal; hmPC, hydroxymethyl-phytochelatin; HPLC, high-performance liquid chromatography; PC, phytochelatin; PMSF, phenyl- methyl-sulphonyl-fluoride; POD, peroxidase; ROS, reactive oxygen spe- cies. * Corresponding author. Tel.: +39-0521-90-6411; fax: +39-0521-90-5403. E-mail address: [email protected] (L. Sanità di Toppi). Plant Physiology and Biochemistry 43 (2005) 45–54 www.elsevier.com/locate/plaphy 0981-9428/$ - see front matter © 2005 Elsevier SAS. All rights reserved. doi:10.1016/j.plaphy.2004.12.004
Transcript of Oxidative stress and phytochelatin characterisation in bread wheat exposed to cadmium excess

Original article
Oxidative stress and phytochelatin characterisation in bread wheatexposed to cadmium excess
Annamaria Ranieri a, Antonella Castagna a, Francesca Scebba a, Maria Careri b,Ingrid Zagnoni b, Giovanni Predieri b, Massimo Pagliari b, Luigi Sanità di Toppi c,*
a Dipartimento di Chimica e Biotecnologie Agrarie, Università di Pisa, via del Borghetto 80, 56124 Pisa, Italyb Dipartimento di Chimica Generale ed Inorganica, Chimica Analitica, Chimica Fisica, Università di Parma, Parco Area delle Scienze 17/A,
43100 Parma, Italyc Dipartimento di Biologia Evolutiva e Funzionale, Sezione di Biologia Vegetale, Università di Parma, Parco Area delle Scienze 11/A, 43100 Parma, Italy
Received 21 September 2004; accepted 12 December 2004
Available online 07 January 2005
Abstract
In this work, we first investigated if the bread wheat (Triticum aestivum L.) cv. Albimonte can be defined as “shoot cadmium excluder”—bycomparing the cadmium (Cd) content in leaves and roots and by calculating the shoot-to-root Cd concentration ratio. Furthermore, we evalu-ated if the exposure to Cd excess could generate oxidative stress in leaves and roots of this cv., in terms of hydrogen peroxide (H2O2)accumulation, NAD(P)H oxidation rate, and variations in reduced glutathione (GSH) content and peroxidase (POD, EC 1.11.1.7) activity.Finally, we surveyed possible quali- quantitative differences in thiol-peptide compound pattern between roots and leaves, in order to verifywhether phytochelatins (PCs) and related thiol-peptides could contribute in limiting the Cd-induced oxidative stress. Unambiguous charac-terisation of PCs and related forms present in the root samples was obtained by electrospray ionisation mass spectrometry (ESI-MS) andESI-tandem MS (ESI-MS/MS). Our results indicate that in leaves the stress generated by the low accumulation of Cd (due to a moderatetranslocation in planta) seems to be counteracted by the antioxidant response and by the PC biosynthesis. On the contrary, in roots, in spite ofthe elevated presence of PCs and related thiol-peptide-compounds, the excess of Cd causes a decline in the antioxidant protection of the organ,with the consequent generation of considerable amounts of H2O2, a direct agent of oxidative stress.© 2005 Elsevier SAS. All rights reserved.
Keywords: Cadmium; Hydrogen peroxide; NAD(P)H oxidase; Peroxidase; Phytochelatins; Triticum aestivum; Wheat
1. Introduction
Cadmium (Cd) is a non-essential heavy metal (HM),released into the environment by anthropogenic and non-anthropogenic sources [23,25,34]. Cadmium long biological
half-life and its tendency to form strong complexes withS-containing peptides/proteins renders this metal a notablehazard to a wide range of organisms [39]. In fact, Cd can betransferred to animals and humans through the food chain[42], and concerns about consumption of Cd-containing foodsare assuming a relevant dimension [8,21]. In many countries,increases in soil Cd concentrations over time are conspicu-ous, due to the addition of fertilisers, animal manures, sew-age sludges, amendments and the atmospheric fall-out[1,21,22]. Cd accumulation in cereals aimed for food produc-tion, i.e. Triticum aestivum L. (bread wheat), can representindeed a worrying risk for animal and human health [8,21],and the fact that Cd is often a by-product of phosphate ferti-lisers [23,25], which are largely utilised in cereal cultivation,has further raised debates and questions on Cd risk manage-ment of these important crops.
Abbreviations: Cd, cadmium; desGlyPC, desglycil-phytochelatin; DPI,diphenylene iodonium; DTT, dithiothreitol; EDTA, ethylenediaminetetraa-cetic acid; ESI-MS, electrospray ionisation mass spectrometry; ESI-MS/MS, electrospray ionisation tandem mass spectrometry; GSH, reducedglutathione; HM, heavy metal; hmPC, hydroxymethyl-phytochelatin; HPLC,high-performance liquid chromatography; PC, phytochelatin; PMSF, phenyl-methyl-sulphonyl-fluoride; POD, peroxidase; ROS, reactive oxygen spe-cies.
* Corresponding author. Tel.: +39-0521-90-6411; fax:+39-0521-90-5403.
E-mail address: [email protected] (L. Sanità di Toppi).
Plant Physiology and Biochemistry 43 (2005) 45–54
www.elsevier.com/locate/plaphy
0981-9428/$ - see front matter © 2005 Elsevier SAS. All rights reserved.doi:10.1016/j.plaphy.2004.12.004

The response to Cd stress in higher plants, and particu-larly in wheat, is a complex phenomenon [17,20,38,43]. Infact, Cd—especially at high exposure concentrations and shortexposure times—can evoke a response of parallel and/or con-secutive events, in which every mechanism could be at thesame time cause and effect of metabolic changes due to Cdstress [34]. Amongst these events, there are evidences thatCd induces antioxidant responses in all plant organs[3,13,18,30,34]. Thus, it is conceivable that, directly or indi-rectly, Cd causes the formation of reactive oxygen species(ROS), such as superoxide anion radical (O2•–), hydroxyl radi-cal (OH• ) and hydrogen peroxide (H2O2) [30,33].
Peroxidases (PODs, EC 1.11.1.7) can modulate the levelof ROS, thanks to their role in H2O2-scavenging and in H2O2-consuming physiological processes [10,27,29]. Peroxidaseshave a more elevated affinity to H2O2 than catalase [24], but,on the other hand, PODs may also produce H2O2 through theoxidation of various molecules, among which NAD(P)H [5,6].Plant cells are in fact capable of generating H2O2 followingbiotic and abiotic stresses [45]. Such an endogenous produc-tion has been mainly ascribed to a plasma membraneNAD(P)H oxidase (EC 1.23.45.3) activity [19,28,30] and/orto pH-dependent cell wall POD activity [5,6].
Both increase and decrease in the activity of many antioxi-dant enzymes has been observed in Cd-treated plants[2,13,26,33], suggesting that the antioxidant systems, besidebeing involved in detoxification process, could be sensitivetargets of Cd toxicity.
Other than influencing the antioxidant status of the plantcell, Cd induces the biosynthesis of phytochelatins PCs, theprincipal HM-complexing peptides of plants [16]. Phytoch-elatins (PCs) are peptides derived from glutathione (GSH)(c-Glu-Cys-Gly), having general structure (c-Glu-Cys)n-Gly, where n = number of repetitions of the c-Glu-Cys unit,which can vary from 2 to 11 (more commonly from 2 to 5)[16]. Interestingly, there are some noteworthy compoundssimilar but not identical to PCs, collectively named iso-PCs[34,35]: in particular, hydroxymethyl-PCs (hmPCs) are iso-PCs discovered in some Poaceae, such as wheat, oat, rice andrye [20]. Hydroxymethyl-PCs contain Ser instead of Gly [i.e.:(c-Glu-Cys)n-Ser], thus they are also called iso-PC-Ser, andare normally present simultaneously with common PCs [20].Even desGlyPCs (i.e. PCs lacking Gly) can be co-present invarying percentages in plants producing PCs and iso-PCs [4].
To separate and analyse PCs and other thiol-related com-pounds, high-performance liquid chromatography (HPLC;using post-column derivatisation with Ellman’s reagent, fol-lowed by UV–visible detection) is generally used [37].Recently, electrospray ionisation mass spectrometry (ESI-MS) and ESI-tandem mass spectrometry (ESI-MS/MS) havereached an increasing importance in PC characterisation[9,46]. These MS-based techniques lead to an increase ofspecificity with regard to the classical approaches based onother detection methods.
In this work, we first investigated if the bread wheat cv.Albimonte can be defined as “shoot Cd excluder” [38]—by
comparing the Cd content in leaves and roots and calculatingthe shoot-to-root Cd concentration ratio. Furthermore, weevaluated whether or not the exposure to Cd excess couldgenerate oxidative stress in leaves and roots of this cv., interms of H2O2 accumulation, NAD(P)H oxidation rate, andvariations in GSH content and POD activity. Finally, we sur-veyed possible quali- quantitative differences in thiol-peptide compound pattern between roots and leaves, in orderto verify whether PCs and related thiol-peptides could con-tribute in limiting the Cd-induced oxidative stress.
An elevated concentration of Cd was deliberately chosenfor three main reasons: (a) to observe clear physiological/biochemical effects in leaves and roots of Cd-exposed plants,otherwise hardly detectable; (b) to induce in these organs asufficient amount of thiol-peptide-compounds for HPLC-ESI-MS and HPLC-ESI-MS/MS characterisation; (c) to inves-tigate whether Cd is accumulated in detectable amounts notonly in the root but also in the leaf, considering the estab-lished high correlation (r = 0.95) between the concentrationof Cd in leaves and grains of wheat plants [15,21].
2. Results and discussion
2.1. Plant growth and dry weight
The adverse effect of 1 mM Cd on the overall growth ofwheat plants was already evident either 10 or 20 d after thetransferral in Cd-enriched nutrient solution (Fig. 1). In fact,the length of the leaves and roots of Cd-treated plants under-went a significant reduction (–15% and –21%, after 10 d, and–39% and –48%, after 20 d, in leaves and roots, respectively,Fig. 1). This reduction was accompanied by a decrease in theDW, particularly pronounced after 20 d of growth (–11% and–18%, after 10 d, and –71% and –142%, after 20 d, in leavesand roots, respectively, Fig. 1). Decrease in growth and inFW or DW was also reported in Cd-treated plants of wheat,barley (Hordeum vulgare L.) and rice (Oryza sativa L.)[2,36,38,43]. Particularly in wheat plants, addition of Cd tothe nutrient solution decreased shoot and root weight andlength, with a growth inhibition varying between roots andshoots and among genotypes [47].
2.2. Cadmium content
As it was expected, at both sampling-times, the plantsgrown in the presence of 1 mM Cd exhibited an increase inCd concentrations in both organs examined (Fig. 1). This wasparticularly evident in roots, where Cd level increased greatly,as compared to the control (+789% and +1088%, after 10 and20 d of growth, respectively), while in leaves Cd was accu-mulated at a lower, although significant, extent over the con-trol (+158% and +56%, after 10 and 20 d of growth, respec-tively). Retention/immobilisation of high amount of Cd inthe root tissue, typical of several plants, can be regarded asone important protection mechanism against the diffusion of
46 A. Ranieri et al. / Plant Physiology and Biochemistry 43 (2005) 45–54
this HM in planta [40]. For this reason, Cd concentration inroots can reach an average of 80% of the total metal taken up[43]. Likewise, in a survey on Cd distribution in various organsof several bread and durum wheat cvs., Greger and Löfstedt[15] reported that the highest level of Cd was accumulated/retained in roots. In any case, according to McLaughlin et al.[21] and Adams et al. [1], the propensity of higher plants totranslocate and accumulate Cd to edible and harvested partsdepends to a large extent on soil and climatic factors, plantgenotype and agronomic management. In particular, crop spe-cies and cvs. differ widely in their ability to absorb, accumu-late and detoxify Cd: for instance, cereals and legumes accu-mulate less Cd in leaves than do leafy crops such as lettuce(Lactuca sativa L.) and spinach (Spinacia oleracea L.) [14].
2.3. Hydrogen peroxide accumulation
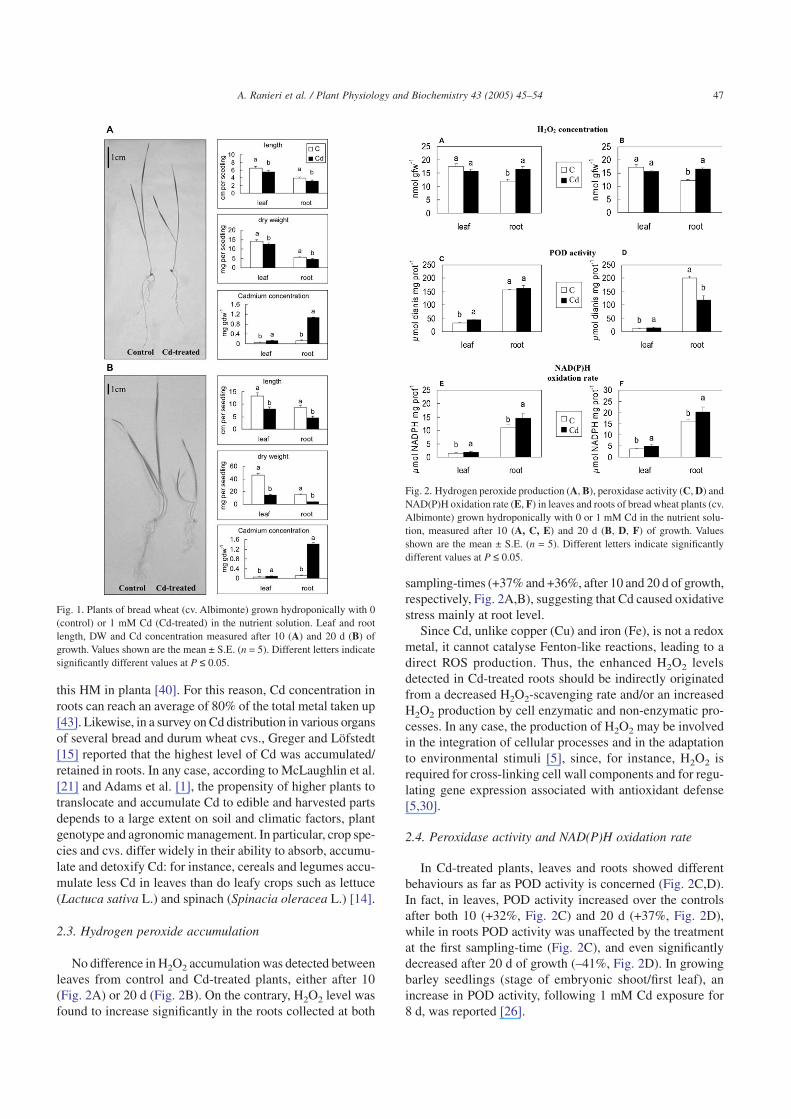
No difference in H2O2 accumulation was detected betweenleaves from control and Cd-treated plants, either after 10(Fig. 2A) or 20 d (Fig. 2B). On the contrary, H2O2 level wasfound to increase significantly in the roots collected at both
sampling-times (+37% and +36%, after 10 and 20 d of growth,respectively, Fig. 2A,B), suggesting that Cd caused oxidativestress mainly at root level.
Since Cd, unlike copper (Cu) and iron (Fe), is not a redoxmetal, it cannot catalyse Fenton-like reactions, leading to adirect ROS production. Thus, the enhanced H2O2 levelsdetected in Cd-treated roots should be indirectly originatedfrom a decreased H2O2-scavenging rate and/or an increasedH2O2 production by cell enzymatic and non-enzymatic pro-cesses. In any case, the production of H2O2 may be involvedin the integration of cellular processes and in the adaptationto environmental stimuli [5], since, for instance, H2O2 isrequired for cross-linking cell wall components and for regu-lating gene expression associated with antioxidant defense[5,30].
2.4. Peroxidase activity and NAD(P)H oxidation rate
In Cd-treated plants, leaves and roots showed differentbehaviours as far as POD activity is concerned (Fig. 2C,D).In fact, in leaves, POD activity increased over the controlsafter both 10 (+32%, Fig. 2C) and 20 d (+37%, Fig. 2D),while in roots POD activity was unaffected by the treatmentat the first sampling-time (Fig. 2C), and even significantlydecreased after 20 d of growth (–41%, Fig. 2D). In growingbarley seedlings (stage of embryonic shoot/first leaf), anincrease in POD activity, following 1 mM Cd exposure for8 d, was reported [26].
Fig. 1. Plants of bread wheat (cv. Albimonte) grown hydroponically with 0(control) or 1 mM Cd (Cd-treated) in the nutrient solution. Leaf and rootlength, DW and Cd concentration measured after 10 (A) and 20 d (B) ofgrowth. Values shown are the mean ± S.E. (n = 5). Different letters indicatesignificantly different values at P ≤ 0.05.
Fig. 2. Hydrogen peroxide production (A, B), peroxidase activity (C, D) andNAD(P)H oxidation rate (E, F) in leaves and roots of bread wheat plants (cv.Albimonte) grown hydroponically with 0 or 1 mM Cd in the nutrient solu-tion, measured after 10 (A, C, E) and 20 d (B, D, F) of growth. Valuesshown are the mean ± S.E. (n = 5). Different letters indicate significantlydifferent values at P ≤ 0.05.
47A. Ranieri et al. / Plant Physiology and Biochemistry 43 (2005) 45–54

Our results suggest that in leaves the Cd-induced increasein POD activity may efficiently remove H2O2, thus contrib-uting to reduce the H2O2 accumulation in this organ. On thecontrary, the much more pronounced Cd accumulation foundin roots seems to adversely affect POD activity after 20 d ofCd treatment. Consequently, cells are no more fully compe-tent to remove H2O2, which possibly accumulates up to toxiclevels. Similarly, in Amaranthus lividus L. seedlings treatedwith 1 mM Cd for 7 d, a strong increase in H2O2 content,with a parallel decrease in POD activity was detected [3].According to Bhattacharjee [3], the simultaneous decrease inthe activity of POD (and catalase) can be a plausible expla-nation for the accumulation of H2O2 and the subsequent oxi-dative stress. Moreover, other studies have demonstrated thatCd-induced oxidative stress can be caused by a Cd-reducedactivity/level of enzymatic and non-enzymatic antioxidants[33]. In fact, POD as well as catalase activities have beenreported to decline in leaves of pea plants treated with 50 µMCd, which, as a consequence, showed an enhanced lipid per-oxidation and increased levels of protein carbonyl groups,indicative of oxidative stress [33]. Interestingly, a 4 mMCd-induced inhibition of a structural POD purified to homo-geneity from 15-d-old wheat plants was reported [11]; accord-ing to Converso et al. [11], this finding could explain, at leastin part, the Cd-induced oxidative stress noticed in wheatplants, particularly in the presence of acute Cd stress.
In both wheat tissues examined at the two sampling-time,the rate of NAD(P)H oxidation underwent a similar signifi-cant increase in Cd-treated plants (+30% and +32%, after 10 d,Fig. 2E, and +32% and +27%, after 20 d, Fig. 2F, in leavesand roots, respectively). However, while in leaves, owing tothe simultaneous increase in POD activity, H2O2 was not accu-mulated, in roots H2O2 produced by NAD(P)H oxidationcould not be scavenged by the unchanged (10 d of treatment)or even reduced (20 d of treatment) POD activity, thus result-ing in increased H2O2 levels.
The use of two inhibitors (DPI, diphenylene iodonium, andKCN, which inhibit NAD(P)H oxidase and POD, respec-tively) revealed that in our experiments H2O2 generation atthe expense of NAD(P)H oxidation was catalysed by PODs,as also reported by Wojtaszek [44]. In fact, while an almostcomplete inhibition of POD activity was achieved by addingKCN to the reaction medium, the addition of DPI resulted tobe ineffective (data not shown).
An increased endogenous H2O2 production at the expenseof NAD(P)H oxidation was observed following pathogenattack [5,6], as well as following exposure to oxidative stresseslike ozone or Cd [28–30].
2.5. Thiol-peptide levels
As regards thiol-peptide-compounds, in leaves of wheatplants exposed to 1 mM Cd for 10 and 20 d, hmGSH, GSHand PCs, but no hmPCs and desGly derivatives, were detected(Fig. 3). In non-exposed leaves, the only thiol-peptides presentwere GSH and traces of hmGSH (Fig. 3A,B). Cd-exposed
leaf extracts contained in particular: hmGSH, GSH, PC2, PC3
and PC4 (Fig. 3A,B). Differences between the two sampling-times were evident especially as regards GSH and total PClevels: the former was similar in controls of both sampling-times, but under Cd stress the leaves from plants exposed for20 d presented about 1.8-fold more GSH than ones fromCd-exposed plants for 10 d. Thus, an additional GSH synthe-sis probably took place after Cd addition to the nutrient solu-tion, and this synthesis increased over time (Fig. 3). The totalPC content was 4.7-fold higher in the leaves exposed for 20 dthan in the 10 d-exposed ones. Unlike in the roots, in the leavestreated for 10 d, PC3 was the thiol-peptide more abundant,followed by PC2, whereas in the leaves treated for 20 d theopposite occurred. Only traces of PC4 were detected in bothsampling-times (Fig. 3).
As far as roots are concerned, the plants exposed to 1 mMCd for 10 and 20 d, contained hmGSH, GSH, PCs, hmPCsand desGly derivatives (Fig. 4). Like in leaves, in non-exposed roots the only thiol-peptides detectable were GSHand traces of hmGSH (Fig. 4A,B), but Cd-exposed rootextracts contained hmGSH, GSH, PC2, hmPC2, desGlyPC2,PC3, hmPC3, desGlyPC3 and PC4 (Fig. 4A,B). The differ-ences in thiol content between the roots from Cd-exposedplants for 10 and 20 d were evident especially as regards GSHand total PC levels. The former was similar in the controls ofboth sampling-times, but under Cd stress the roots from plantsexposed for 20 d had a content of GSH about 3.3-fold moreelevated than the plants exposed for 10 d. PC, hmPC and des-GlyPC contents were 2.6-, 2.8- and 2.0-fold higher in the 20-dsampling-time than in the 10-d one, respectively. In bothsampling-times, PC3 and hmPC3 were the more abundantthiol-peptides, followed by PC2 and hmPC2, whereas onlytraces of PC4 were detected (Fig. 4).
The presence of hmPCs in very low concentrations wasfound by Wójcik and Tukendorf [43] in bread wheat roots
Fig. 3. Thiol-peptide content in leaves of bread wheat plants (cv. Albimonte)grown hydroponically with 0 or 1 mM Cd in the nutrient solution, measuredafter 10 (A) and 20 d (B). Values shown are the mean ± S.E. (n = 5). Diffe-rent letters indicate significantly different values at P ≤ 0.05.
48 A. Ranieri et al. / Plant Physiology and Biochemistry 43 (2005) 45–54

exposed to 50 µM Cd. However, the concentration of hmPCsrose greatly following a 250 µM Cd exposure for 7 d: thus,probably the induction of hmPCs in wheat requires higherexposure concentrations and times, as carried out in ourexperiments.
No significant differences were detected comparing theGSH and the hmGSH content, respectively, in roots and leavesof control plants. On the contrary, the increase in GSH con-tent observed both in leaves and in roots of Cd-exposed plants(except for the 10 d-treated roots) can indicate that, since PCsderive from GSH, cells subjected to acute Cd stress have torestore the GSH used to form PCs, as previously demon-strated by Rüegsegger and Brunold [31] and Rüegsegger etal. [32]. According to Sharma et al. [36], the incorporation ofGSH into PCs in barley roots and leaves could be compen-sated by an augmented sulphur uptake to support GSH bio-synthesis. An alternative hypothesis is that H2O2 accumula-tion may provoke derepression of existing mRNA encodingthe enzymes of the GSH biosynthetic pathway, with conse-quent GSH biosynthesis [45].
Finally, the total thiol-peptide contents (other than GSHand hmGSH) of Cd-exposed plants for 10 and 20 d were,respectively, 57- and 32-fold higher in roots than in leaves.The elevated Cd concentration in roots induced in this organthe biosynthesis of elevated amounts of thiol-peptides, as alsoconfirmed by Stolt et al. [38] and Wójcik and Tukendorf [43].
2.6. Thiol-peptide characterisation
HPLC-ESI-MS full scan chromatograms of all wheat rootsamples exposed to Cd showed signals corresponding to theprotonated molecule of PC3 (m/z 772), desGlyPC3 (m/z 715),hmPC3 (m/z 802), PC2 (m/z 540), desGlyPC2 (m/z 483),hmPC2 (m/z 570), as depicted in Fig. 5A. In particular, HPLC-ESI-MS profiles of the root extracts corresponding to both
sampling-times were comparable to those obtained usingUV–visible detection, whereas no signals were present in thecontrols. ESI-MS spectra of the corresponding peaks aredominated by the protonated molecules [M + H]+, which allowthe identification of each form according to its mass.
The extracted ion chromatograms of all PCs show couplesof unresolved peaks (Fig. 5A), this behaviour being particu-larly evident for PC3 and related thiol-containing peptides.This evidence could be explained by hypothesising the pres-ence of two different conformations of the same species insolution, probably due to intramolecular interactions whichslightly influence the polarity. An alternative hypothesis maybe that an adduct between the PC molecules and other spe-cies present in the matrix is formed in solution, and then it iscleaved in the ESI source.
In addition, signals at m/z 538 and 568 are observed at18.4 and 16.5 min, respectively (Fig. 5B, 1 and 2), justifyingthe presence of oxidised PC2 and hmPC2, which elute beforetheir corresponding reduced forms. This species are formedas a consequence of the intramolecular bond between the sul-phydryl groups of proximal Cys (Fig. 5B).
The thiol-containing peptides, as well as the oxidisedforms, were further characterised by tandem mass spectrom-etry, in order to induce fragmentation of the protonated mol-ecules of PCs and related thiol-peptides. Examples of ESI-MS/MS product-ion spectra obtained for the precursor ionsof PC3, desGlyPC3 and hmPC3 are shown in Fig. 5C. Ob-served product ions arise mostly from the cleavage of the pep-tide bonds to form bn ions (retaining the charge on the aminoterminus) and yn ions (retaining the charge on the carboxyterminus). Other abundant product ions derive from furtherdissociation of the bn and yn ions by loss of one or more ter-minal amino acids. In particular, the fragment at m/z 568 arisesfrom the loss of the terminal Gly residue from the y6 ion atm/z 643 (Fig. 5C, 1). A further loss of Cys and Glu residuesjustifies the presence of the fragment at m/z 336. The loss ofCys and Glu from the y5 ion at m/z 715 accounts for the pres-ence of m/z 336 in Fig. 5C (2).
The agreement between the observed and the theoreticalfragmentation patterns led to the unambiguous identificationof the PCs and other related thiol-peptide-compounds underinvestigation.
2.7. Overall comments
In leaves from bread wheat plants exposed to 1 mM Cd for10 and 20 d, the following aspects can be pointed out: the Cdcontent in leaves was about 8- and 16-fold lower than the onein roots, at 10 and 20 d sampling-times, respectively. Thus, alimited Cd translocation in planta occurred. These results con-firm the ones obtained by Hart et al. [17] and Wójcik andTukendorf [43], who noticed that bread wheat plants of vari-ous cvs. accumulated more Cd in roots than in shoots, due toa reduced root-to-shoot translocation. In fact, the transloca-tion of Cd from roots to shoots can differ significantly betweenwheat cvs. Interestingly, the proportion of Cd translocated
Fig. 4. Thiol-peptide content in roots of bread wheat plants (cv. Albimonte)grown hydroponically with 0 or 1 mM Cd in the nutrient solution, measuredafter 10 (A) and 20 d (B). Values shown are the mean ± S.E. (n = 5). Diffe-rent letters indicate significantly different values at P ≤ 0.05.
49A. Ranieri et al. / Plant Physiology and Biochemistry 43 (2005) 45–54

from roots to shoots was from 1.5- to 4.5-fold higher in breadwheat than in durum wheat [17]. Moreover, Stolt et al. [38]found that four wheat cvs. were “shoot Cd excluders”, sincethe shoot-to-root Cd concentration ratio varied from 0.09 to0.13. Also in our experiments this ratio was equal to 0.12 and0.06, respectively, in 10- and 20 d Cd-exposed plants. Thus,also the bread wheat cv.Albimonte can be regarded as a “shootCd excluder”. The low Cd accumulation in leaves is an impor-tant parameter, since in wheat there is an established highcorrelation (r = 0.95) between the concentration of Cd inleaves and in grains [21]. In this regard, also Greger and Löf-
stedt [15] found a positive correlation in some wheat cvs.between Cd levels in grains and in shoots, at least during thevegetative phase.
The relatively low level of Cd accumulation in the leaftissue can possibly explain the lack of difference in H2O2
concentration between leaves from Cd-exposed and controlplants, in spite of the elevated Cd concentration used (1 mM).In fact, the oxidative stress generated by the limited Cd pres-ence in the leaf tissue was probably counterbalanced by theincrease in POD activity and in GSH content, at both thesampling-d. Moreover, the increase in NAD(P)H oxidation
Fig. 5. Representative HPLC-ESI-MS extract-ion chromatograms of roots from bread wheat plants exposed to 1 mM Cd for 20 d: (A) ions at m/z: 1) 772 (PC3),2) 715 (desGlyPC3), 3) 802 (hmPC3), 4) 540 (PC2), 5) 483 (desGlyPC2), 6) 570 (hmPC2); (B) ions at m/z: 1) 538 and 2) 568, attributable to oxidised PC2 andhmPC2; (C) HPLC-ESI-MS/MS spectra of: 1) PC3, 2) desGlyPC3, 3) hmPC3 in the same samples.
50 A. Ranieri et al. / Plant Physiology and Biochemistry 43 (2005) 45–54

rate detected in Cd-treated leaves did not bring about an accu-mulation of H2O2, owing to the increased POD activity. Therelatively low presence of Cd in leaf tissues led also to a mod-erate thiol-peptide induction: the amount of PCs in leaveswas in fact 57- and 32-fold lower than in roots, at the twosampling-time, respectively. These results confirm the onesby Klapheck et al. [20], who found in shoots of rice and otherPoaceae a much lesser extent of thiol-peptide-compounds thanin roots. Thus, according to our experiments, the low accu-mulation of Cd in wheat leaves (due to a moderate transloca-tion in planta) seems to be counteracted by the leaf cellulardetoxification response, especially after the 10-d Cd treat-ment.
In roots, the Cd content was about 8- and 16-fold higherthan in the leaves, at 10 and 20 d sampling-times, respec-tively. Thus, an elevated immobilisation/accumulation of Cdoccurred in this organ. In experiments performed on sixteendifferent wheat genotypes, Zhang et al. [47] found that Cdconcentration in roots was 10-fold higher than in shoots.Moreover, according to Hart et al. [17], in bread wheat Cdwas retained in roots perhaps by mechanisms centred on com-partimentalisation in root cell vacuoles and/or on decreasedxylem loading of Cd ions. The accumulation of high Cd lev-els in the root may limit the accumulation of Cd in the edibleand above-ground parts of the plant [14,15], and thus it canbe particularly important in crops aimed for food production,such as wheat.
The high level of Cd immobilisation/accumulation in roots,observed in our experiments, stimulated an increase in H2O2
concentration in this organ, compared with controls, but thehigh Cd concentration did not raise the POD activity in 10-dtreated plants, and caused even a decrease in the activity ofthis enzyme after 20 d of Cd treatment. Unlike in leaves, inroots the increase in NAD(P)H oxidation rate brought aboutan accumulation of H2O2, possibly due to the constancy/decrease in POD activity. Furthermore, the high presence ofCd in root tissues led to an elevated thiol-peptide induction:the amount of PCs in roots was in fact 57- and 32-fold higherthan in leaves, at the two sampling-times, respectively. Thus,the presence in the roots of elevated concentrations of PCsand related thiols probably played a role in the retention ofhigh amounts of Cd in this tissue. However, in spite of theelevated biosynthesis of thiol-peptide-compounds, in rootsthe high Cd concentration—especially after the 20-d Cdtreatment—caused a decrease in POD activity and an accu-mulation of H2O2, with a consequent state of cellular oxida-tive stress.
3. Conclusion
From the overall data obtained in this work, it can beinferred that in wheat leaves the stress generated by low accu-mulation of Cd (due to a moderate translocation in planta)seems to be counteracted by the antioxidant response and bythe PC biosynthesis. On the contrary, in roots, in spite of the
elevated presence of PCs and related thiol-peptide-compounds, the high concentration of Cd caused a decline inthe antioxidant protection of the organ, with the consequentgeneration of considerable amounts of H2O2, a direct agentof oxidative stress.
4. Methods
4.1. Plant material
Seeds of wheat (T. aestivum L., cv. Albimonte) sterilisedwith 30% Na-hypochlorite were dark germinated between wetfilter papers in Petri dishes for a week, then transferred tohalf-strength Hoagland’s nutrient solution, pH 5.8, contain-ing 0 or 1 mM CdSO4 and grown under a photon flux densityof 400 µmol m–2 s–1 with a 14 h photoperiod, at a tempera-ture ranging from 18 to 25 °C and R.H. of 70%. Ten pots foreach treatment were prepared to give a total of 100 controland 100 Cd-treated plants. Growth solutions were continu-ously aerated and renewed every 3 d. After 10 or 20 d ofgrowth in a hydroponic solution in the presence of 1 mM Cd,the fully expanded leaves and the roots of 50 plants for eachtreatment were collected and immediately used for analysesor frozen in liquid nitrogen and stored at –80 °C.
4.2. Cadmium determination
The concentration of Cd in leaves and roots was deter-mined by atomic absorption using a Perkin–Elmer 373 spec-trophotometer, mainly following Sandalio et al. [33]. Imme-diately after harvesting, leaves and roots were extensivelywashed in distilled water; then, samples were oven-dried,ground to powder and stored under vacuum until acidic diges-tion, which was carried out dissolving aliquots of 100 mg ofdry matter in 5 ml flame-heated concentrated H2SO4. H2O2
drops were added until complete clarification of samples wasattained. Final volumes were adjusted to 25 ml by milliQ waterand absorption measures were carried out with a Perkin–Elmer INTENSITRON™ lamp at 228.8 nm.
4.3. Hydrogen peroxide assay
Hydrogen peroxide production was monitored fluorimetri-cally using the Amplex Red Hydrogen Peroxide Assay Kit(Molecular Probes), according to Zhou et al. [48] and to themanufacturer’s recommendations. This assay is based on thedetection of H2O2 using 10-acetyl-3,7-dihydroxy-phenoxazine (Amplex Red reagent), a highly sensitive andstable probe for H2O2. Fresh leaf and root segments (2 cmlength, about 200 mg of FW) were dark incubated in 5 ml of0.25 M Na phosphate buffer pH 7.4 for 3 h under continuousstirring. The reaction medium contained 50 µM Amplex Red,0.1 units ml–1 horseradish peroxidase and aliquots of the incu-bation buffers. After 30 min incubation at room temperaturein the dark, the red fluorescence signal was recorded using a
51A. Ranieri et al. / Plant Physiology and Biochemistry 43 (2005) 45–54

FP-750 JASCO spectrofluorometer, using excitation at 563 nmand fluorescence detection at 587 nm. Background fluores-cence, determined for a no-H2O2 control reaction, was sub-tracted from each value. The H2O2 content was determinedusing a standard curve (0–10 µM).
4.4. Enzyme extraction and activity
Leaf and root samples of control and treated plants wereground in liquid nitrogen with 10% (w/w) polyvinylpirroly-done and then homogenised in 100 mM Tricine–KOH buffer(pH 8.0), 20 mM MgCl2, 50 mM KCl, 10 mM ethylenedi-aminetetraacetic acid (EDTA), 1‰ (v/v) Triton X-100, 1 mMdithiothreitol (DTT), and 0.50 mM phenyl-methyl-sulphonyl-fluoride (PMSF).After centrifugation at 24,000 × g for 30 minat 4 °C, the supernatant was collected and dialysed overnightagainst 2.5 mM Tricine–KOH (pH 8.0). Peroxidase activitywas measured spectrophotometrically following the increasein absorbance at 460 nm due to the oxidation of o-dianisidineaccording to the method of Ranieri et al. [29]. The reactionmedium contained 20 mM Na-acetate pH 5.0, 1%o-dianisidine in methanol, 30 mM H2O2 and an adequate ali-quot of the extract. The activity was calculated using theextinction coefficient of 11.3 mM–1 cm–1 for o-dianisidineand expressed as µmol o-dianisidine oxidised min–1 mg ofprotein. The rate of NAD(P)H oxidation was measured byfollowing the decrease in absorbance at 340 nm of a reactionmedium containing 40 mM Na-acetate (pH 5.5), 250 mMsucrose, 1 mM MnCl2, 100 µM salicylhydroxamic acid,100 µM NADH and an adequate extract aliquot [41]. Theactivity was calculated using the extinction coefficient of6.22 mM–1 cm–1 for NADH. To discriminate between theactivity of NAD(P)H oxidase complex and NAD(P)H PODs,15 µM DPI or 50 µM KCN, specific inhibitors of NAD(P)Hoxidase and POD, respectively, were added to the reactionmedium. The protein content of the extracts was measuredspectrophotometrically at 595 nm according to Bradford [7],using BSA as standard.
4.5. Thiol-peptide analyses
Wheat root and leaf samples from plants exposed to 1 mMCd for 10 and 20 d were homogenised in a mortar in ice-cold5% (w/v) 5-sulphosalicylic acid, containing 6.3 mM diethyl-enetriaminepentaacetic acid, according to De Knecht et al.[12]. After centrifugation at 10,000 × g for 10 min at 4 °C,the supernatant fraction of extracts was filtered through Micro-pure 0.45 µm filter (Amicon, USA) and immediately assayedby HPLC (Beckman, USA), mainly following De Knecht etal. [12]. Thiol-containing peptides (GSH, PCs and related pep-tides) were separated by a reverse-phase C18 column (Puro-sphere, Merck, Germany), by injecting 200 µl of extract. Theseparation was obtained by using a 0–26% CH3CN gradient,with a flow-rate set at 0.7 ml min–1. Elution solutions con-tained 0.05% trifluoroacetic acid. Thiol-containing peptideswere determined using post-column derivatisation with
300 µM Ellman’s reagent [5,5′dithio(2-nitrobenzoic acid)],detected at 412 nm and measured by a calibration curve forstandard SH groups. Identification of GSH and individual PCswas based on the comparison of their retention times withstandard GSH (Merck, Germany), standard PC samples fromSilene vulgaris (Moench).
4.6. Thiol-peptide characterisation by HPLC–MSand HPLC–MS/MS
HPLC elution was performed on a Luna C18(2) narrow-bore column (250 × 2.1 mm, 5 µm) (Phenomenex, Torrance,CA, USA) at room temperature and using aqueous trifluoro-acetic acid 0.05% (v/v, pH 2.8; solvent A) and acetonitrile/water (50/50, v/v) acidified with 0.05% (v/v) trifluoroaceticacid (solvent B) as the mobile phase, mainly following Chas-saigne et al. [9]. The linear gradient was set as follows: from0 to 12 min 98% A, at 15 min 89% A, at 31 min 74% A, at38 min 0% A and from 40 to 50 min reequilibrating at 98%A. The flow-rate was 200 µl min–1. The mobile phase wasdelivered by a Waters 2690 series Alliance quaternary pump(Waters, Milford, MA, USA) equipped with a 120-vial capac-ity sample management system. The injection volume was30 µl. A Quattro LC triple quadrupole instrument (Micro-mass, Manchester, UK) equipped with an electrospray inter-face and a Masslynx v. 3.4 software (Micromass) was usedfor data acquisition and processing. The nebulising gas (nitro-gen, 99.999% purity) and the desolvation gas (nitrogen,99.998% purity) were delivered at a flow-rate of 105 and536 l h–1, respectively. The operating parameters of the ESIinterface were as follows: electrospray voltage 3.0 kV, conevoltage 30 V, r.f. lens 0.3 V, source temperature 110 °C, des-olvation temperature 150 °C. The quadrupoles were tuned tounit mass resolution. Qualitative LC–MS analysis were per-formed by operating the mass spectrometer in positive ion(PI) mode acquiring mass spectra over the scan range m/z300–2000 using a step size of 0.1 Da and a scan duration of2.2 s. Operating in MS/MS mode for confirmation purpose,product-ion scan mass spectra of the protonated molecules ofPC2, desGlyPC2, hmPC2, PC3, desGlyPC3, and hmPC3 wererecorded in the 30–820 Da mass range with a scan durationof 1 s and an interscan delay of 0.05 s. Collision energy wasset at 25 eV (argon pressure 2.56 e–3 mbar).
4.7. Chemicals
All the reagents used were of analytical grade and obtainedfrom Aldrich (Steinheim, Germany), BDH (Poole, UK),Merck (Darmstadt, Germany), Serva (Heidelberg, Ger-many), Sigma (St. Louis, MO) and Carlo Erba (Milan, Italy).Water was purified with a Milli-Q Element A-10 water puri-fication system (Millipore, Bedford, MA, USA).
4.8. Statistical analyses
All treatments and analytical procedures were carried outon at least three replicates. Values shown in the Figs. 1–4 are
52 A. Ranieri et al. / Plant Physiology and Biochemistry 43 (2005) 45–54

the means of five determinations ± standard error (S.E.). Com-parison between controls and Cd-treated samples was evalu-ated by one-way ANOVA. Different letters denote significantdifferences at P ≤ 0.05.
Acknowledgements
The technical assistance of Zulema Vattuone, Universityof Parma, is gratefully acknowledged.
References
[1] M.L. Adams, F.J. Zhao, S.P. McGrath, F.A. Nicholson, B.J. Cham-bers, Predicting cadmium concentrations in wheat and barley grainusing soil properties, J. Environ. Qual. 33 (2004) 532–541.
[2] M.B. Ali, H.S. Chun, B.K. Kim, C.B. Lee, Cadmium-induced changesin antioxidant enzyme activities in rice (Oryza sativa L. cv. Dongjin),J. Plant Biol. 45 (2002) 134–140.
[3] S. Bhattacharjee, Membrane lipid peroxidation, free radical scaven-gers and ethylene evolution in Amaranthus as affected by lead andcadmium, Biol. Plant. 40 (1997/98) 131–135.
[4] W.R. Bernhard, J.H.R. Kägi, Purification and characterization ofatypical cadmium-binding polypeptides from Zea mays, Experientia52 (1987) 309–315.
[5] C.S. Bestwick, I.R. Brown, J.W. Mansfield, Localized changes inperoxidase activity accompany hydrogen peroxide generation duringthe development of a non-host hypersensitive reaction in lettuce, PlantPhysiol. 118 (1998) 1067–1078.
[6] G.P. Bolwell, Role of active oxygen species and nitric oxide in plantdefense responses, Curr. Opin. Plant Biol. 2 (1999) 287–294.
[7] M.M. Bradford, A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein–dyebinding, Anal. Biochem. 72 (1976) 248–254.
[8] R.L. Chaney, J.A. Ryan, Y.-M. Li, S.L. Brown, Soil cadmium as athreat to human health, in: M.J. McLaughlin, B.R. Singh (Eds.),Cadmium in Soils and Plants, Kluwer Academic Publishers, Dor-drecht, 1999, pp. 219–256.
[9] H. Chassaigne, V. Vacchina, T.M. Kutchan, M.H. Zenk, Identificationof phytochelatin-related peptides in maize seedlings exposed to cad-mium and obtained enzymatically in vitro, Phytochemistry 56 (2001)657–668.
[10] J.H. Christensen, G. Bauw, K.G. Welinder, M. Van Montagu, W. Boer-jan, Purification and characterization of peroxidases correlated withlignification in poplar xylem, Plant Physiol. 118 (1998) 125–135.
[11] D.A. Converso, M.E. Fernandez, M.L. Tomaro, Cadmium inhibitionof a structural wheat peroxidase, J. Enzyme Inhibit. 15 (2000) 171–183.
[12] J.A. De Knecht, M. Van Dillen, P.L.M. Koevoets, H. Schat,J.A.C. Verkleji, W.H.O. Ernst, Phytochelatins in cadmium-sensitiveand cadmium-tolerant Silene vulgaris: chain length distribution andsulphide incorporation, Plant Physiol. 104 (1994) 255–261.
[13] S.M. Gallego, M.P. Benavides, M.L. Tomaro, Effect of heavy metalion excess on sunflower leaves: evidences for involvement of oxida-tive stress, Plant Sci. 121 (1996) 151–159.
[14] C.A. Grant, L.D. Bailey, M.J. McLaughlin, B.R. Singh, Managementfactors which influence cadmium concentrations in crops, in:M.J. McLaughlin, B.R. Singh (Eds.), Cadmium in Soils and Plants,Kluwer Academic Publishers, Dordrecht, 1999, pp. 151–198.
[15] M. Greger, M. Löfstedt, Comparison of uptake and distribution ofcadmium in different cultivars of bread and durum wheat, Crop Sci. 44(2004) 501–507.
[16] E. Grill, E.-L. Winnacker, M.H. Zenk, Phytochelatins: the principalheavy-metal complexing peptides of higher plants, Science 230(1985) 674–676.
[17] J.J. Hart, R.M. Welch, W.A. Norvell, L.A. Sullivan, L.V. Kochian,Characterization of cadmium binding, uptake, and translocation inintact seedlings of bread and durum wheat cultivars, Plant Physiol.116 (1998) 1413–1420.
[18] M.A. Iannelli, F. Pietrini, L. Fiore, L. Petrilli, A. Massacci, Antioxi-dant response to cadmium stress in Phragmites australis plants, PlantPhysiol. Biochem. 40 (2002) 977–982.
[19] T. Keller, H.G. Damude, D. Werner, P. Doerner, R.A. Dixon, C. Lamb,A plant homolog of the neutrophil NADPH oxidase gp91phox subunitgene encodes a plasma membrane protein with Ca2+ binding motifs,Plant Cell 10 (1998) 255–266.
[20] S. Klapheck, W. Fliegner, I. Zimmer, Hydroxymethyl-phytochelatins[(c-glutamylcysteine)n-serine] are metal-induced peptides of thePoaceae, Plant Physiol. 104 (1994) 1325–1332.
[21] M.J. McLaughlin, D.R. Parker, J.M. Clarke, Metals andmicronutrients—food safety issues, Field Crop Res. 60 (1999) 143–163.
[22] M.J. McLaughlin, B.R. Singh, Cadmium in soils and plants, in:M.J. McLaughlin, B.R. Singh (Eds.), Cadmium in Soils and Plants,Kluwer Academic Publishers, Dordrecht, 1999, pp. 1–9.
[23] G.N. Mhatre, C.E. Pankhurst, Bioindicators to detect contaminationof soils with special reference to heavy metals, in: C.E. Pankhurst,B.M. Doube, V.V.S.R. Gupta (Eds.), Biological Indicators of SoilHealth and Sustainable Productivity, CAB International, New York,1997, pp. 349–369.
[24] G. Noctor, C.H. Foyer, Ascorbate and glutathione: keeping activeoxygen under control, Annu. Rev. Plant Physiol. Plant Mol. Biol. 49(1998) 249–279.
[25] J.O. Nriagu, J.M. Pacyna, Quantitative assessment of worldwide con-tamination of air, water and soils with trace metals, Nature 333 (1988)134–139.
[26] J. Patra, B.B. Panda, A comparison of biochemical responses tooxidative and metal stress in plants of barley, Hordeum vulgare L,Environ. Pollut. 101 (1998) 99–105.
[27] C. Penel, S. Carpin, M. Crevecoeur, P. Simon, H. Greppin, Binding ofperoxidases to Ca2+-pectate: possible significance for peroxidasefunction in cell wall, Plant Perox, Newslett. 14 (2000) 33–40.
[28] A. Ranieri, A. Castagna, J. Pacini, B. Baldan, A. Mensuali-Sodi,G.F. Soldatini, Early production and scavenging of hydrogen peroxidein the apoplast of sunflower plants exposed to ozone, J. Exp. Bot. 54(2003) 2529–2540.
[29] A. Ranieri, F. Petacco, A. Castagna, G.F. Soldatini, Redox state andperoxidase system in sunflower plants exposed to ozone, Plant Sci.159 (2000) 159–168.
[30] M.C. Romero-Puertas, M. Rodriguez-Serrano, F.J. Corpas,M. Gomez, L.A. Del Rio, Cadmium-induced subcellular accumula-tion of O2
– and H2O2 in pea leaves, Plant Cell Environ. 27 (2004)1122–1134.
[31] A. Rüegsegger, C. Brunold, Effects of cadmium on c-glumilcysteinyl-glycine synthesis in maize seedlings, Plant Physiol. 99 (1992) 428–433.
[32] A. Rüegsegger, D. Schmutz, C. Brunold, Regulation of glutathionesynthesis by cadmium in Pisum sativum L, Plant Physiol. 93 (1990)1579–1584.
[33] L.M. Sandalio, H.C. Dalurzo, M. Gomez, M.C. Romero-Puertas,L.A. Del Rio, Cadmium-induced changes in the growth and oxidativemetabolism of pea plants, J. Exp. Bot. 52 (2001) 2115–2126.
[34] L. Sanità di Toppi, R. Gabbrielli, Response to cadmium in higherplants, Environ. Exp. Bot. 41 (1999) 105–130.
[35] L. Sanità di Toppi, P. Gremigni, B. Pawlik-Skowronska,M.N.V. Prasad, C.S. Cobbett, Response to heavy metals in plants: amolecular approach, in: L. Sanità di Toppi, B. Pawlik-Skowronska(Eds.), Abiotic Stresses in Plants, Kluwer Academic Publishers, Dor-drecht, 2003, pp. 133–156.
53A. Ranieri et al. / Plant Physiology and Biochemistry 43 (2005) 45–54

[36] S.S. Sharma, S. Kaul, A. Metwally, K.C. Goyal, I. Finkemeier,K.-J. Dietz, Cadmium toxicity to barley (Hordeum vulgare) asaffected by varying Fe nutritional status, Plant Sci. 166 (2004) 1287–1295.
[37] F.E.C. Sneller, L.M. Van Heerwaarden, P.L.M. Koevoets, R. Vooijs,H. Schat, J.A.C. Verkleij, Derivatization of phytochelatins from Silenevulgaris induced upon exposure to arsenate and cadmium. Compari-son of derivatization with Ellman’s reagent and monobromobimane, J.Agric. Food Chem. 48 (2000) 4014–4019.
[38] J.P. Stolt, F.E.C. Sneller, T. Bryngelsson, T. Lundborg, H. Schat,Phytochelatin and cadmium accumulation in wheat, Environ. Exp.Bot. 49 (2003) 21–28.
[39] S.J. Traina, The environmental chemistry of cadmium, in:M.J. McLaughlin, B.R. Singh (Eds.), Cadmium in Soils and Plants,Kluwer Academic Publishers, Dordrecht, 1999, pp. 11–37.
[40] J.A.C. Verkleij, H. Schat, Mechanisms of metal tolerance in higherplants, in: A.J. Shaw (Ed.), Heavy Metal Tolerance in Plants: Evolu-tionary Aspects, CRC Press, Boca Raton, 1990, pp. 179–193.
[41] A. Vianello, M. Zancani, G. Nagy, F. Macrì, Guaiacol peroxidaseassociated to soybean root plasma membranes oxidises ascorbate, J.Plant Physiol. 150 (1997) 573–577.
[42] G.J. Wagner, Accumulation of cadmium in crop plants and its conse-quences to human health, Adv. Agron. 51 (1993) 173–212.
[43] M. Wójcik, A. Tukendorf, Cd-tolerance of maize, rye and wheatseedlings, Acta Physiol. Plant. 21 (1999) 99–107.
[44] P. Wojtaszek, The oxidative burst: a plant’s early response againstinfection, Biochem. J. 322 (1997) 681–692.
[45] C. Xiang, D.J. Oliver, Glutathione metabolic genes co-ordinatelyrespond to heavy metals and jasmonic acid in Arabidopsis, Plant Cell10 (1998) 1539–1550.
[46] T.Y. Yen, J.A. Villa, J.G. DeWitt, Analysis of phytochelatin-cadmiumcomplexes from plant tissue culture using nano-electrospray ioniza-tion tandem mass spectrometry and capillary liquid chromatography/electrospray ionization tandem mass spectrometry, J. Mass Spectrom.34 (1999) 930–941.
[47] G. Zhang, M. Fukami, H. Sekimoto, Genotypic differences in effectsof cadmium on growth and nutrient compositions in wheat, J. PlantNutr. 23 (2000) 1337–1350.
[48] M. Zhou, Z. Diwu, N. Panchuk-Voloshina, R.P. Haughland, A stablenonfluorescent derivative of resofurin for the fluorometric determina-tion of trace hydrogen peroxide: applications in detecting the activityof phagocyte NADPH oxidase and other oxidases, Anal. Biochem.253 (1997) 162–168.
54 A. Ranieri et al. / Plant Physiology and Biochemistry 43 (2005) 45–54




















