
Development of copper based material systems for ... - UNSWorks
165
Development of copper based material systems for generating nitric oxide to control nitrifying bacterial biofilms Author: Wonoputri, Vita Publication Date: 2016 DOI: https://doi.org/10.26190/unsworks/19322 License: https://creativecommons.org/licenses/by-nc-nd/3.0/au/ Link to license to see what you are allowed to do with this resource. Downloaded from http://hdl.handle.net/1959.4/57088 in https:// unsworks.unsw.edu.au on 2022-07-17
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Development of copper based material systems for ... - UNSWorks

Development of copper based material systems forgenerating nitric oxide to control nitrifying bacterial biofilms
Author:Wonoputri, Vita
Publication Date:2016
DOI:https://doi.org/10.26190/unsworks/19322
License:https://creativecommons.org/licenses/by-nc-nd/3.0/au/Link to license to see what you are allowed to do with this resource.
Downloaded from http://hdl.handle.net/1959.4/57088 in https://unsworks.unsw.edu.au on 2022-07-17

Development of Copper Based Material
Systems for Generating Nitric Oxide to
Control Nitrifying Bacterial Biofilms
A thesis submitted to The University of New South Wales in
partial fulfilment of the degree of Doctor of Philosophy
By
Vita Wonoputri
School of Chemical Engineering
Faculty of Engineering
December 2016

i
THE UNIVERSITY OF NEW SOUTH WALES
Thesis/Dissertation Sheet Surname or Family name: WONOPUTRI
First name: VITA
Other name/s:
Abbreviation for degree as given in the University calendar:
PhD
School: Chemical Engineering
Faculty: Engineering
Title: Development of Copper Based Material Systems for Generating Nitric Oxide to Control Nitrifying Bacterial Biofilms
Abstract 350 words maximum: (PLEASE TYPE) Biofilms, which are bacteria cells aggregates that exist within a matrix of extracellular polymeric substances, are
known to cause problems in industrial water systems. In order to prevent biofilm formation and pathogens occurrence, disinfectants such as chloramine are usually added into the water. However, chloramine addition has been shown to trigger the growth of nitrifying biofilms which subsequently accelerate chloramine decay. Therefore, a new antibiofilm agent, namely nitric oxide (NO), is investigated for nitrifying biofilm control.
Herein, NO was generated from catalytic reduction of nitrite in the presence of a copper(II) complex catalyst embedded in a poly(vinyl chloride) (PVC) matrix. Ascorbic acid was added into the solution as a reducing agent to aid the formation of the active copper(I) species that will react with nitrite to generate NO. The copper-nitrite-ascorbic acid combination showed enhanced NO generation compared to that generated in the presence of nitrite-ascorbic acid alone, and subsequently, enhanced biofilm suppression was observed. The catalytically generated NO was also found to be effective in dispersing pre-formed biofilm, with simultaneous biofilm cells killing observed when a high concentration of nitrite-ascorbic acid was used.
An alternative reducing agent, namely Fe2+, was investigated for the potential to mediate reduction of copper(II) complex for generating NO. The amount of NO generated was found to be highly dependent on Fe speciation in different pH and buffer composition. Nonetheless, the NO generated in phosphate buffer pH 6 is still capable of dispersing pre-formed nitrifying biofilms, thus suggesting the robustness of NO-mediated biofilm dispersal.
In the last part of the study, an iron complex, namely FeDTTCT, was synthesised and immobilised into PVC with the copper catalyst. The iron complex was found to facilitate copper(II)/copper(I) redox cycling, subsequently enabling NO generation from nitrite. Importantly, the mixed metal system exhibited a non-toxic antibiofilm activity, whereby biofilm formation was suppressed and bacterial growth was confined to the free-floating planktonic phase. These thus imply that the mixed metal system was capable of converting nitrite endogenously produced by nitrifying bacteria to NO, hence eliminating the need to add a reducing agent and NO precursor in solution form into the system.
Declaration relating to disposition of project thesis/dissertation I hereby grant to the University of New South Wales or its agents the right to archive and to make available my thesis or dissertation in whole or in part in the University libraries in all forms of media, now or here after known, subject to the provisions of the Copyright Act 1968. I retain all property rights, such as patent rights. I also retain the right to use in future works (such as articles or books) all or part of this thesis or dissertation. I also authorise University Microfilms to use the 350 word abstract of my thesis in Dissertation Abstracts International (this is applicable to doctoral theses only). ………………………………………… Signature
……………………………………..…………….. Witness Signature
……….…………………….… Date
FOR OFFICE USE ONLY
Date of completion of requirements for Award:

ii
ORIGINALITY STATEMENT
‘I hereby declare that this submission is my own work and to the best of my knowledge it contains no materials previously published or written by another person, or substantial proportions of material which have been accepted for the award of any other degree or diploma at UNSW or any other educational institution, except where due acknowledgement is made in the thesis. Any contribution made to the research by others, with whom I have worked at UNSW or elsewhere, is explicitly acknowledged in the thesis. I also declare that the intellectual content of this thesis is the product of my own work, except to the extent that assistance from others in the project's design and conception or in style, presentation and linguistic expression is acknowledged.’
Signed ……………………………………………..............
Date ……………………………………………..............

iii
COPYRIGHT STATEMENT
‘I hereby grant the University of New South Wales or its agents the right to archive and to make available my thesis or dissertation in whole or part in the University libraries in all forms of media, now or here after known, subject to the provisions of the Copyright Act 1968. I retain all proprietary rights, such as patent rights. I also retain the right to use in future works (such as articles or books) all or part of this thesis or dissertation. I also authorise University Microfilms to use the 350 word abstract of my thesis in Dissertation Abstract International (this is applicable to doctoral theses only). I have either used no substantial portions of copyright material in my thesis or I have obtained permission to use copyright material; where permission has not been granted I have applied/will apply for a partial restriction of the digital copy of my thesis or dissertation.' Signed ……………………………………………........................... Date ……………………………………………...........................
AUTHENTICITY STATEMENT ‘I certify that the Library deposit digital copy is a direct equivalent of the final officially approved version of my thesis. No emendation of content has occurred and if there are any minor variations in formatting, they are the result of the conversion to digital format.’ Signed ……………………………………………...........................
Date ……………………………………………...........................

Abstract
iv
Abstract
Biofilms, which are bacteria cells aggregates that exist within a matrix of
extracellular polymeric substances, are known to cause problems in industrial water
systems. In order to prevent biofilm formation and pathogens occurrence, disinfectants
such as chloramine are usually added into the water. However, chloramine addition has
been shown to trigger the growth of nitrifying biofilms which subsequently accelerate
chloramine decay. Therefore, a new antibiofilm agent, namely nitric oxide (NO), is
investigated for nitrifying biofilm control.
Herein, NO was generated from catalytic reduction of nitrite in the presence of a
copper(II) complex catalyst embedded in a poly(vinyl chloride) (PVC) matrix. Ascorbic
acid was added into the solution as a reducing agent to aid the formation of the active
copper(I) species that will react with nitrite to generate NO. The copper-nitrite-ascorbic
acid combination showed enhanced NO generation compared to that generated in the
presence of nitrite-ascorbic acid alone, and subsequently, enhanced biofilm suppression
was observed. The catalytically generated NO was also found to be effective in
dispersing pre-formed biofilm, with simultaneous biofilm cells killing observed when a
high concentration of nitrite-ascorbic acid was used.
An alternative reducing agent, namely Fe2+, was investigated for the potential to
mediate reduction of copper(II) complex for generating NO. The amount of NO
generated was found to be highly dependent on Fe speciation in different pH and buffer
composition. Nonetheless, the NO generated in phosphate buffer pH 6 is still capable of

Abstract
v
dispersing pre-formed nitrifying biofilms, thus suggesting the robustness of NO-
mediated biofilm dispersal.
In the last part of the study, an iron complex, namely FeDTTCT, was
synthesised and immobilised into PVC with the copper catalyst. The iron complex was
found to facilitate copper(II)/copper(I) redox cycling, subsequently enabling NO
generation from nitrite. Importantly, the mixed metal system exhibited a non-toxic
antibiofilm activity, whereby biofilm formation was suppressed and bacterial growth
was confined to the free-floating planktonic phase. These thus imply that the mixed
metal system was capable of converting nitrite endogenously produced by nitrifying
bacteria to NO, hence eliminating the need to add a reducing agent and NO precursor in
solution form into the system.

Acknowledgements
vi
Acknowledgements
This PhD thesis would not be possible without my supervisors. My sincere
gratitude goes to Prof. Rose Amal for giving me this opportunity and for being a
wonderful mentor. Thanks to Dr. Lachlan Yee and Dr. Nicolas Barraud for all the help
in developing this lovely project and for all the constructive feedback. My gratitude also
goes to Dr. Sanly Liu for always guiding and helping me and Dr. Cindy Gunawan for all
her valuable feedback. Thank you Dr. May Lim for all the support throughout my PhD
study. This research was supported under Australian Research Council’s Linkage
Projects funding scheme (LP110100459). The financial supports from Australian Water
Quality Centre (SA Water) and Water Corporation (Western Australia) and the PVC
provision from Chemson Pacific Pty Ltd are gratefully acknowledged.
The assistance from Mark Wainwright Analytical Centre throughout this PhD
project is much appreciated. Special thanks to Dr. Bill Gong for XPS analysis, Dr.
Rabeya Akter and Dr. Dorothy Yu for ICP and ion chromatography analysis, and all the
people in BMIF and NMR lab. My sincerest gratitude to Mr. Pejhman Keshvardoust for
sharing the knowledge about nitrifying bacteria. Thank you to Dr. Victor Wong for
always helping me with the consumables ordering and John Starling for all the lab
assistance. I also would like to extend my gratitude to Mr. Jim Paschali, Mr. Phil
Thompson, and Mr. Paul Brockbank for all the technical support, especially for fixing
my incubator. Administrative support from Ms. Ik Ling Lau is much appreciated. I give
my special thanks to Dr. Mandalena Hermawan, the queen bee of Partcat gossip, for all
the support and laughter.

Acknowledgements
vii
Thank you to my Partcat friends, especially bio-lab partner Ayu, partner-in-crime
Emma, my first friend Shi Nee, gym buddy Ee Teng, and all the girls who have kept me
well fed: Hui Ling, Cui Ying, Xue Lian and Phoebe. To the Partcat boys: Tze Hao Tan,
RJ, and Hendra, thanks for enduring all the teasing. Special thanks to Peng Wang and
Amir Nashed for all the chatting session. The supports from all past and present
members of Partcat are much appreciated. My gratitude also goes to my friends in
CMBB (616).
Thank you for all the loving support from my family back in Indonesia: my dad
who always pushes me to work to the best of my ability, my mom for always praying
for me (and for sending me cute pictures of my pomeranian Science!), and my sister and
her husband for always supporting me. Finally, to my dear husband, thanks for letting
me pursue my dream.

List of Publications
viii
List of Publications
JOURNAL PUBLICATIONS:
1. Wonoputri, V.; Gunawan, C.; Liu, S.; Barraud, N.; Yee, L. H.; Lim, M.; Amal,
R. Copper complex in poly(vinyl chloride) as a nitric oxide-generating catalyst
for the control of nitrifying bacterial biofilms. ACS Applied Materials and
Interfaces (2015), 7 (40), 22148–22156.
2. Wonoputri, V.; Gunawan, C.; Liu, S.; Barraud, N.; Yee, L. H.; Lim, M.; Amal,
R. Iron complex facilitated copper redox cycling for nitric oxide generation as
non-toxic nitrifying biofilm inhibitor. ACS Applied Materials and Interfaces
(2016), 8(44), 30502-30510.
3. Wonoputri, V.; Gunawan, C.; Liu, S.; Barraud, N.; Yee, L. H.; Lim, M.; Amal,
R. Ferrous ion as reducing agent in the generation of antibiofilm nitric oxide
from copper-based catalytic system (in preparation)
CONFERENCE PRESENTATIONS:
1. Wonoputri, V.; Liu, S.; Gunawan, C.; Barraud, N.; Yee, L. H.; Lim, M.; Amal,
R. Suppression and removal of nitrifying bacteria biofilm by catalytic
generation of nitric oxide. Australasian Particle Technology Society 2nd Student
Conference, Phillip Island, Australia, 26-27 September 2015.
2. Wonoputri, V.; Liu, S.; Gunawan, C.; Barraud, N.; Yee, L. H.; Lim, M.; Amal,
R. Efficacy and mechanism of action of nitric oxide against nitrifying bacteria
biofilms at different stages of development. Asian Pacific Confederation of

List of Publications
ix
Chemical Engineering, Melbourne, Australia, 27 September-1 October 2015
(poster).
3. Wonoputri, V.; Liu, S.; Gunawan, C.; Barraud, N.; Yee, L. H.; Lim, M.; Amal,
R. Catalytic generation of nitric oxide for the control of nitrifying bacteria
biofilm. 7th ASM Conference on Biofilms, Chicago, USA, 24-29 October 2015
(poster).
4. Wonoputri, V.; Liu, S.; Gunawan, C.; Barraud, N.; Yee, L. H.; Lim, M.; Amal,
R. Nitrifying biofilm control through the utilisation of catalytically generated
nitric oxide. The University of New South Wales Engineering Postgraduate
Research Symposium, Sydney, Australia, 9-11 November 2015.
5. Wonoputri, V.; Liu, S.; Gunawan, C.; Barraud, N.; Yee, L. H.; Lim, M.; Amal,
R. Control of nitrifying bacteria biofilms through catalytically generated nitric
oxide, invited speaker for iThree Institute group presentation, University of
Technology Sydney, 9 August 2016.

x
Table of Contents
Abstract ........................................................................................................................... iv
Acknowledgements ......................................................................................................... vi
List of Publications ....................................................................................................... viii
Table of Contents ............................................................................................................ x
Chapter 1. Introduction ............................................................................................... 1
1.1. References .......................................................................................................... 4
Chapter 2. Literature review ....................................................................................... 8
2.1. Biofilms: introduction and properties ................................................................. 8
2.2. Issues with biofilms presence and difficulties in its eradication ...................... 14
2.3. Current strategies for biofilm prevention and removal .................................... 20
2.3.1. Adhesion prevention ................................................................................. 21
2.3.2. Biofilm cells killing and biofilm maturation inhibitor .............................. 26
2.3.3. Biofilm disruption ..................................................................................... 34
2.3.4. Nitric oxide as a new antibiofilm agent .................................................... 37
2.4. Current strategies in nitric oxide utilisation ..................................................... 38
2.4.1. Nitric oxide-releasing materials for application in biomedical field ........ 38
2.4.2. Application of nitric oxide donors in water industry ................................ 45
2.5. Summary........................................................................................................... 46
2.6. References ........................................................................................................ 47
Chapter 3. Nitric oxide generation from nitrite and ascorbic acid solution in
the presence of copper complex catalyst ..................................................................... 63
3.1. Introduction ...................................................................................................... 63
3.2. Experimental methods ...................................................................................... 66

Chapter 1
xi
3.2.1. Synthesis of copper dibenzo[e,k]-2,3,8,9-tetraphenyl-1,4,7,10-tetraaza-
cyclododeca-1,3,7,9-tetraene complex (CuDTTCT complex) and its
characterisation ........................................................................................................ 66
3.2.2. NO generation measurement ..................................................................... 68
3.2.3. Ascorbic acid oxidation measurement ...................................................... 69
3.2.4. Biofilm suppression assay ......................................................................... 69
3.2.5. Biofilm dispersal and metabolic activity assay on pre-formed biofilms... 71
3.3. Results and discussion ...................................................................................... 72
3.3.1. Characterisation of copper complex (CuDTTCT) powder and films ....... 72
3.3.2. Nitric oxide generation measurements ...................................................... 74
3.3.3. Nitrifying bacteria biofilm suppression by nitrite-ascorbic acid and
CuDTTCT-nitrite-ascorbic acid mixture ................................................................. 76
3.3.4. Nitrifying bacteria biofilm dispersal upon the addition of nitrite-ascorbic
acid in the presence of CuDTTCT ........................................................................... 83
3.4. Summary........................................................................................................... 88
3.5. References ........................................................................................................ 89
Chapter 4. Catalytic generation of nitric oxide: the use of ferrous ion as a
reducing agent ............................................................................................................... 94
4.1. Introduction ...................................................................................................... 94
4.2. Experimental methods ...................................................................................... 96
4.2.1. Synthesis of copper dibenzo[e,k]-2,3,8,9-tetraphenyl-1,4,7,10-tetraaza-
cyclododeca-1,3,7,9-tetraene complex (CuDTTCT complex) ................................ 96
4.2.2. NO generation measurement with Fe2+ solution as the reducing agent .... 96
4.2.3. Fe speciation analysis ................................................................................ 97
4.2.4. Biofilm dispersal assay ............................................................................. 97
4.3. Result and discussion ....................................................................................... 98
4.3.1. Effect of testing solution and pH on NO generation from CuDTTCT-
nitrite-Fe2+ mixture .................................................................................................. 98

Chapter 1
xii
4.3.2. Effect of nitrite and Fe2+ concentration on NO generation from
CuDTTCT-nitrite-Fe2+ mixture ............................................................................. 106
4.3.3. Biofilm dispersal in the presence of CuDTTCT-nitrite-Fe2+ solution .... 109
4.4. Summary......................................................................................................... 113
4.5. References ...................................................................................................... 114
Chapter 5. Iron complex facilitated copper redox cycling for nitric oxide
generation ................................................................................................................. 117
5.1. Introduction .................................................................................................... 117
5.2. Experimental methods .................................................................................... 119
5.2.1. Synthesis of dibenzo[e,k]-2,3,8,9-tetraphenyl-1,4,7,10-tetraaza-
cyclododeca-1,3,7,9-tetraene ligand and the metal complexes ............................. 119
5.2.2. Characterisation of metal complexes ...................................................... 120
5.2.3. NO generation measurement ................................................................... 121
5.2.4. Biofilm assay ........................................................................................... 122
5.3. Result and discussion ..................................................................................... 123
5.3.1. Confirmation of metal complexes formation .......................................... 123
5.3.2. Nitric oxide generation from CuDTTCT+FeDTTCT films in the presence
of nitrite ................................................................................................................. 125
5.3.3. Antibiofilm activity of CuDTTCT+FeDTTCT mixed metal film by the
involvement of catalytically generated NO ........................................................... 130
5.4. Summary......................................................................................................... 138
5.5. References ...................................................................................................... 139
Chapter 6. Conclusions and Recommendations .................................................... 143
6.1. Conclusions .................................................................................................... 143
6.2. Recommendations .......................................................................................... 145
6.3. References ...................................................................................................... 149

1
Chapter 1.
Introduction
In their natural habitat, bacteria have a tendency to form surface-attached
communities known as biofilms, where their formation is identifiable by the production
of extracellular polymeric substances that encased and protect the bacteria.1 Biofilms
formation has detrimental effects in many areas, ranging from health care to industrial
water system. For instance, biofilms can decrease heat exchanger or cooling tower
efficiency.2 Biofilm presence can foul water filtration membranes, instigating the need
for more frequent cleaning, which subsequently reduces the membrane lifetime.3
Biofilm formation on water distribution pipelines can lead to microbial induced
corrosion and offer protection for pathogenic bacteria, potentially releasing them into
the flowing water through the natural shedding cycle of biofilm or shear force.4
Moreover, environmental biofilms have been found to act as a reservoir for the spread
of antibiotics resistance genes, due to the high cell density and close cell-to-cell
proximity.5,6
In order to control microbial growth, disinfectants such as chlorine and
chloramine are usually added into the water. However, these disinfectants have been
shown to be ineffective at eradicating biofilms, due to the presence of protective EPS
barrier which increased biofilm cells resistance to antimicrobial compounds compared
to their free-floating (planktonic) counterparts.7 Chloramine disinfectant, which is

Chapter 1
2
preferred compared to chlorine because of less formation of disinfection by-products
and more stable compared to chlorine, is proven to be ineffective towards the ubiquitous
environmental biofilm of nitrifying bacteria.8,9 Moreover, nitrifying bacteria and their
soluble microbial products can accelerate the inactivation and decay of chloramine,
subsequently releasing chlorine and ammonia, where the latter act as the main nutrient
for nitrifying bacteria, further enhancing the biofilm formation.10–12 Therefore, there is a
clear need to develop a novel method to eradicate biofilms with minimal potential for
bacterial resistance development.
Several novel antibiofilm strategies have been proposed, which generally can be
divided into three different methods depending on the mechanism employed: adhesion
prevention, biofilm cells killing, and disruption of biofilm.13 However, it is believed that
a single method would not be able to eradicate biofilm thoroughly. Therefore, a
combination of different mechanisms is preferred. Nitric oxide (NO), which is a soluble
free radical gas that has been shown to be effective at controlling the formation of
antimicrobial-resistant biofilms,14 seems to be an attractive solution. NO can affect
biofilms either by inducing a toxic pathway (cell death) at high concentrations or via a
non-toxic pathway at low concentrations that cause biofilm bacteria to disperse from the
surface and revert to a free-floating phase.15–17 Importantly, it is believed that biofilm
treatment by NO will not trigger the development of resistant biofilms, due to the
several mechanisms by which NO presents toxicity to cells15,18 or the signalling
pathway involved in its non-toxic antibiofilm effect.19 However, application of NO is
still limited due to its high reactivity and short half-life. These properties of NO have
led to the development of novel strategies to control biofilms, in particular on materials
that are capable of delivering NO to the target site.

Chapter 1
3
In general, materials that are capable of delivering NO can be divided into two
types: NO-releasing materials and NO-generating materials.20–22 NO-releasing materials
depend on the use of NO donors such as N-diazeniumdiolates (NONOates) or S-
nitrosothiols (RSNO) which are incorporated into a nanomaterial delivery vehicle or
coating. Such materials have limited NO release due to the finite reservoir of NO that
can be loaded during synthesis. In contrast, NO-generating materials appear to be more
advantageous due to their ability to catalyse NO generation from endogenous sources,
such as nitrite ions which are ubiquitous in water.23 However, the ability of NO-
generating materials in eradicating biofilms has yet to be determined. Moreover, such
materials, which are usually copper(II) or selenium(II) based, are highly dependent on
the presence of appropriate reducing agents, as the reduced form is the most active
species in catalytic generation of NO.21,24 Therefore, this thesis attempts to provide
insight on the utilisation and development of copper-based NO-generating materials for
controlling the formation of biofilms. Nitrifying bacteria, commonly occurring in
chloraminated industrial water system, were chosen as a biofilm-forming model
organism. In specific, the aims and objectives of the work presented in this thesis are:
1. To assess the ability of catalytic generation of NO for the prevention and
removal of nitrifying bacteria biofilms.
2. To examine the use of Fe2+ ions as an alternative reducing agent for instigating
copper redox cycling for NO generation and its subsequent effect on biofilm
removal.
3. To develop an iron-based complex that can be embedded together with the
catalyst to facilitate copper redox cycling for generating NO, therefore
eliminating the need to add any reducing agent in the water.

Chapter 1
4
Chapter 2 provides background on biofilm properties and implications, and
reviewed the current literature on biofilm control strategies, with a detailed focus on
NO-based control strategy and materials. Chapter 3 presents an investigation on the use
of a copper complex catalyst immobilised within a poly(vinyl chloride) (PVC) matrix to
generate NO in the presence of nitrite as an NO source, and ascorbic acid as a reducing
agent. The effectivity of the system to prevent and remove nitrifying bacteria biofilm
was also investigated. Chapter 4 describes a comprehensive study on the use of Fe2+
ions as an alternative reducing agent for catalytic NO generation, including
determination of iron speciation, investigations of the effect of different parameters,
such as pH, solution composition, and nitrite-Fe2+ concentration toward NO production,
and the resulting biofilm removal efficiency. Following the result presented in Chapter
4, an iron complex that can facilitate copper redox cycling for NO generation was
synthesised and results are detailed in Chapter 5. The iron complex was subsequently
embedded together with the copper catalyst within a PVC matrix, eliminating the need
for the addition of reducing agent in solution form. PVC films containing the mixed
metal complexes were then examined for biofilm formation in the presence of nitrite
endogenously produced by the nitrifying bacteria. Chapter 6 summarises the key
findings of this work and provides recommendations for future research.
1.1. References
1. Hall-Stoodley, L., Costerton, J. W. & Stoodley, P. Bacterial biofilms: from the
natural environment to infectious diseases. Nat. Rev. Microbiol. 2, 95–108
(2004).
2. Mattila-Sandholm, T. & Wirtanen, G. Biofilm formation in the industry: A

Chapter 1
5
review. Food Rev. Int. 8, 573–603 (1992).
3. Barnes, R. J. et al. Nitric oxide treatment for the control of reverse osmosis
membrane biofouling. Appl. Environ. Microbiol. 81, 2515–2524 (2015).
4. Coetser, S. E. & Cloete, T. E. Biofouling and biocorrosion in industrial water
systems. Crit. Rev. Microbiol. 31, 213–232 (2005).
5. Balcazar, J. L., Subirats, J. & Borrego, C. M. The role of biofilms as
environmental reservoirs of antibiotic resistance. Front. Microbiol. 6, 1–9 (2015).
6. Baquero, F., Martínez, J.-L. & Cantón, R. Antibiotics and antibiotic resistance in
water environments. Curr. Opin. Biotechnol. 19, 260–265 (2008).
7. Bridier, A., Briandet, R., Thomas, V. & Dubois-Brissonnet, F. Resistance of
Bacterial Biofilms to Disinfectants: a review. Biofouling 27, 1017–1032 (2011).
8. Lee, W. H., Wahman, D. G., Bishop, P. L. & Pressman, J. G. Free chlorine and
monochloramine application to nitrifying biofilm: comparison of biofilm
penetration, activity, and viability. Environ. Sci. Technol. 45, 1412–1419 (2011).
9. Pressman, J. G., Lee, W. H., Bishop, P. L. & Wahman, D. G. Effect of free
ammonia concentration on monochloramine penetration within a nitrifying
biofilm and its effect on activity, viability, and recovery. Water Res. 46, 882–894
(2012).
10. Bal Krishna, K. C., Sathasivan, A. & Chandra Sarker, D. Evidence of soluble
microbial products accelerating chloramine decay in nitrifying bulk water
samples. Water Res. 46, 3977–3988 (2012).
11. Maestre, J. P., Wahman, D. G. & Speitel, G. E. Monochloramine cometabolism
by Nitrosomonas europaea under drinking water conditions. Water Res. 47,
4701–9 (2013).
12. Maestre, J. P., Wahman, D. G. & Speitel, G. E. Monochloramine cometabolism
by mixed-culture nitrifiers under drinking water conditions. Environ. Sci.
Technol. 50, 6240–6248 (2016).

Chapter 1
6
13. Gupta, P., Sarkar, S., Das, B., Bhattacharjee, S. & Tribedi, P. Biofilm,
pathogenesis and prevention—a journey to break the wall: a review. Arch.
Microbiol. 198, 1–15 (2015).
14. Orman, M. A. & Brynildsen, M. P. Persister formation in Escherichia coli can be
inhibited by treatment with nitric oxide. Free Radic. Biol. Med. 93, 145–154
(2016).
15. Arora, D. P., Hossain, S., Xu, Y. & Boon, E. M. Nitric Oxide Regulation of
Bacterial Biofilms. Biochemistry 54, 3717–3728 (2015).
16. Schairer, D. O., Chouake, J. S., Nosanchuk, J. D. & Friedman, A. J. The potential
of nitric oxide releasing therapies as antimicrobial agents. Virulence 3, 271–279
(2012).
17. Yang, Y., Qi, P., Yang, Z. & Huang, N. Nitric oxide based strategies for
applications of biomedical devices. Biosurface and Biotribology 1, 177–201
(2015).
18. Privett, B. J., Broadnax, A. D., Bauman, S. J., Riccio, D. A. & Schoenfisch, M.
H. Examination of bacterial resistance to exogenous nitric oxide. Nitric Oxide 26,
169–173 (2012).
19. Njoroge, J. & Sperandio, V. Jamming bacterial communication: New approaches
for the treatment of infectious diseases. EMBO Mol. Med. 1, 201–210 (2009).
20. Hwang, S. & Meyerhoff, M. E. Polyurethane with tethered copper(II)-cyclen
complex: preparation, characterization and catalytic generation of nitric oxide
from S-nitrosothiols. Biomaterials 29, 2443–2452 (2008).
21. Yang, Y. et al. Development of nitric oxide catalytic coatings by conjugating 3,3-
disulfodipropionic acid and 3,3-diselenodipropionic acid for improving
hemocompatibility. Biointerphases 10, 04A303 (2015).
22. Wu, Y. & Meyerhoff, M. E. Nitric oxide-releasing/generating polymers for the
development of implantable chemical sensors with enhanced biocompatibility.
Talanta 75, 642–650 (2008).

Chapter 1
7
23. Barraud, N., Kelso, M. J., Rice, S. A. & Kjelleberg, S. Nitric Oxide : A Key
Mediator of Biofilm Dispersal with Applications in Infectious Diseases. Curr.
Pharm. Des. 21, 31–42 (2015).
24. Oh, B. K. & Meyerhoff, M. E. Catalytic generation of nitric oxide from nitrite at
the interface of polymeric films doped with lipophilic Cu(II)-complex: a potential
route to the preparation of thromboresistant coatings. Biomaterials 25, 283–293
(2004).

8
Chapter 2.
Literature review
2.1. Biofilms: introduction and properties
Bacterial biofilms can be defined as complex microbial communities or
populations that are embedded in a self-produced matrix commonly known as
extracellular polymeric substance (EPS).1,2 In natural environments, around 99% of
bacteria in the world has been estimated to exist in biofilms.3 Bacterial growth in
biofilm form is preferred compared to the free swimming or planktonic growth mode
because of the advantages that biofilm mode of life offers. For instance, biofilm
formation allows the bacterial cells to survive in a hostile environment against a range
of stressors, such as antibiotics, host immune response, and predator.1,4 The close
proximity between cells, either single-species or multi-species microorganism, mostly
resulted in a stable synergistic interaction.5,6 For instance, ammonia oxidising bacteria,
nitrite oxidising bacteria, and heterotroph bacteria are commonly found to coexist in one
community.7 Ammonia oxidising bacteria gain energy from the conversion of ammonia
to nitrite, where the latter is subsequently used by nitrite oxidising bacteria as an energy
source.8,9 Organic matters, which are the by-product of ammonia and nitrite oxidising
bacteria (commonly known as nitrifying bacteria) metabolism, are then utilised by
heterotroph bacteria.7

Chapter 2
9
Biofilm formation is a complex process consisting of several steps (Figure 2.1)
and is affected by different factors, such as the bacteria strains and environmental
parameters.10,11 In general, the process started with diffusion and adsorption of ions and
soluble components such as sugar, proteins, lipids, fatty acids, DNA, and humic acid
onto surfaces creating conditioning film that aids in bacterial attachment.3,12 For
instance, adsorption of divalent cation on anionic substrates will enhance bacteria
attachment by reducing electrostatic repulsion between the surface and bacteria.12 Then,
attachment of planktonic bacteria onto the surface (adhesion) or to each other (cohesion)
would soon follow.11,13,14 The formation of conditioning film and the subsequent
adhesion and attachment of bacteria are regulated by surface factors such as texture,
functionality, charge, hydrophobicity, van der Waals forces, and double layer
interactions.10,12,13 Rougher surface will provide more area for adhesion and are more
difficult to clean, resulting in rapid regrowth of biofilm by multiplication.15 Bacteria
cells, which have a net negative charge, will attach rapidly to the positively charged
surface, while attachment to negatively charged surface is not stable due to the repulsive
forces of the charge.12,16 Bacteria cells prefer to attach to hydrophobic surfaces because
water displacement from the surfaces will promote close contact between cells and the
surfaces.12 However, bacteria attachment to a hydrophilic surface also has been
reported, especially if the surface tension of the bacterial cell wall is higher than the
surface tension of the surrounding liquid.12 Furthermore, it has been shown that initial
adhesion started at locations where the cells are sheltered against shear force, which
facilitates the transition from reversible to irreversible attachment.15

Chapter 2
10
Figure 2.1 Biofilm life cycle.
Cell appendages that help in bacteria motility, such as pili and flagella, also play
an important role in bacterial adhesion to surfaces and formation of biofilm, especially
in Gram-negative species.14 It has been shown that a flagellar mutant of Pseudomonas
aeruginosa is unable to colonise surfaces efficiently, while Escherichia coli bacteria
needed the aid of type I pili and curli fimbriae.11 However, it does not necessarily mean
that bacteria species that do not have cell appendages cannot form a biofilm. Instead,
protein and polysaccharides mediate the attachment of non-motile cells, such as
Staphylococcus epidermidis and Staphylococcus aureus.
When physical appendages of bacteria (such as flagella, fimbriae, and pili) are
able to overcome the repulsive force of the surface, the attachment becomes permanent
or irreversible.3,13 This irreversible attachment is also marked by the secretion of EPS
that aids the adhesion between cells and surface starts.10,12 Moreover, increased
resistance towards antimicrobial agents is observed.13
After the initial lag phase has passed, rapid growth in biofilm population
(exponential phase) begins.3 Biofilm growth could also happen by recruitment of single
cells or cell flocs from the bulk fluid, although the proportion is minor compared to the
replication.11 In the growth step, microbial cells started communicating by producing
Initial
attachment
Irreversible
attachmentGrowth Maturation Biofilm
dispersal

Chapter 2
11
autoinducer signals (commonly known as quorum sensing), such as N-acyl homoserine
lactones (AHL) and autoinducer-1 (AI-1) for gram-negative bacteria, and oligopeptides
for gram-positive bacteria.13,17 When the microbial population reached a certain number,
these autoinducers trigger gene expression that is associated with the adhesive needs of
the biofilm mode of life.3,13 For instance, motility is suppressed and production of
surface appendages is not necessary.3,11 Quorum sensing also modulates other cellular
functions such as pathogenesis, nutrient acquisition, conjugation, motility and secondary
metabolite production.12 At the same time, significant production of EPS is
expected.11,12
Biofilms are mainly comprised of EPS, which can occupy up to 75-90% of the
biofilm total volume, while cells represent only 10-25% of the volume.3,18 EPS can be
found both on the outside and the inside of biofilm,19 and usually are differentiated into
tightly bound, loosely bound, and soluble EPS.14 It is mainly composed of
polysaccharides, protein, extracellular DNA, lipids, and water.14,18,20 The composition of
EPS varies depending on the bacteria species, shear forces, temperature, nutrient
availability, and substrate type.18,19 The presence of EPS in a biofilm is very important
as it holds several functions (Table 2.1).

Chapter 2
12
Table 2.1 EPS functions in biofilms.
Function Relevance for biofilms
Adhesion Aids in planktonic cells colonisation and attachment
Aggregation Bridging between cells, temporary immobilisation of bacterial
populations and development of localised high cell densities
Cohesion Mechanical stability of biofilms (in conjunction with multivalent
cations) and determining biofilm architecture
Water retention Maintaining the highly hydrated environment around the
microorganisms, offer protection against desiccation
Protective barrier Displaying resistance to nonspecific and specific host defences
and tolerance to various antimicrobials agents (oxidising or
charged biocides, antibiotics, metallic cations, ultraviolet
radiation, protozoan grazers)
Sorption (sink) of
organic and
inorganic
compounds
Allows the accumulation of nutrients from the environment and
promotes ion exchange, mineral formation, and toxic metal
accumulation, thus contributing in environmental detoxification.
Storing excess carbon in unbalanced carbon to nitrogen
condition. Allows trapping of biologically active molecules,
such as cell communication signals.
Export of cell
components
Release metabolic products
Enzymatic activity Aids in the digestion of exogenous macromolecules, allows the
degradation of EPS for cells release from biofilm structure
Nutrient source Providing carbon, nitrogen, and phosphorus source
Exchange of genetic
information
Facilitates horizontal gene transfer between biofilm cells
Redox activity Acts as electron donor or acceptor Adapted from Flemming and Wingender,2010.18
As the growth increases, biofilms start to create different structures based on the
growth and environmental condition.1 The most common structure is mushroom shaped
structure known as stromatolites with a diameter between tens to hundreds of
microns.1,3 However, in areas where the water flow is strong, biofilm structure is usually
filamentous, forming a streamer structure.1 Additionally, cells inside a biofilm structure
will undergo physiological differentiation based on different factors, specifically
nutrient and oxygen gradient.21 Studies using oxygen profile measurement showed that
oxygen concentration steadily declines as the electrode travel deeper into the biofilm
structure (Figure 2.2), causing slow cells growth and low or no metabolic activity in the

Chapter 2
13
interior of a biofilm.21 On the other hand, metabolic products are more concentrated
inside the biofilm structure.21
Figure 2.2 Representative images of nutrient and metabolic products distribution inside a
biofilm. (a) Metabolic substrate concentration is higher on the interface compared to the
centre, while (b) metabolic product is more concentrated in the centre of the biofilm. (c) A
metabolic intermediate that is both consumed and produced within the biofilm will have
local maxima inside the biofilm. Adapted from Stewart and Franklin, 2008.21
When a small subpopulation does not have access to both nutrient and oxygen,
the cells become inactive or begin to die.21 The localised cell death is followed by cell
lysis which begins the active dispersal event in a biofilm.20 The initial downregulation
of flagella is halted and motile bacteria can be seen again.11 This new planktonic cells
will then migrate to a new surface and start a new cycle of biofilm formation.20
Biofilm dispersal can also be triggered by various internal and external factors.22
For instance, low nutrient availability will disperse Pseudomonas aeruginosa biofilm,
while high nutrient availability will cause Acinetobacter sp. biofilm to spread out and
disperse.20 Change in iron level has been found to disrupt Pseudomonas aeruginosa
biofilms.23 Other cell signals, including D-amino acids and the unsaturated fatty acids
cis-11-methyl-2-dodecenoic acid and cis-2-decenoic acid, were also found to modulate
biofilm dispersal and formation.20,24,25
Importantly, it was found that alteration in the level of intracellular secondary
messenger molecules known as cyclic di-GMP (c-di-GMP) is the central mechanism
involved in regulating many biofilm behaviours, including biofilm dispersal.26–28 It was
(a) Metabolic substrate (b) Metabolic product (c) Metabolic intermediate

Chapter 2
14
found that at a high level of c-di-GMP, cells mainly exist in biofilm phase, while a low
level of c-di-GMP will promote the switch to planktonic or free-floating phase. The
shift of c-di-GMP level depends on the activity diguanylate cyclases for its synthesis
and phosphodiesterases for its degradation. Several factors are known to alter c-di-GMP
levels, such as nitric oxide, glutamate, and temperature shift.26,29,30
Besides active dispersal events, some passive dispersal events can also occur.
Instead of generating motile planktonic cells (swarming dispersal), the generation of
non-motile cell aggregates that shed from the biofilm structure has been reported.1
These aggregates consist of cells with similar phenotype as biofilm cells, thus cannot be
categorised as planktonic bacteria, and they will move to a new surface following the
fluid flow of the environment.1 However, it was also reported that passive dispersal
could involve the movement of a whole biofilm structure across the surface (surface
dispersal) through shear-mediated transport.1
2.2. Issues with biofilms presence and difficulties in its eradication
Biofilm is ubiquitous in the natural and industrial environment, and it can have
both beneficial and detrimental effects. For examples, biofilms are commonly used in
the field of bioremediation, where they can utilise harmful substances for their
metabolic processes.31 They can also be used to degrade environmentally hazardous
chemicals in soil, remove metals and radionuclides contaminants, treat oil spills and
nitrogen compounds in water, and purify wastewater in bioreactors.3,14 However,
unwanted biofilm presence have caused problems in many areas, such as medical,
transportation, food processing plant, pulp and paper industry, and industrial water

Chapter 2
15
systems.32 For instance, the presence of hospital-related infection, including infection at
the site of implanted medical devices such as artificial prosthetics and catheters, is
linked to the presence of biofilms.3,13 Patients with cystic fibrosis are prone from lungs
infection caused by Pseudomonas aeruginosa biofilm.13 Treatment for biofilm-based
infections can cost more than 1 billion dollars annually.33 Biofilm with a thickness as
small as 25 µm on ship hulls can increase drag by 8%, while biofilm with a roughness
element of 50 µm can increase drag by 22%.31 Biofilm presence in the food industry has
been linked with food poisoning outbreaks,10 and longer sterilisation time requirement.32
In specific, this review will focus on the detrimental effect of biofilms in industrial
water systems.
Biofilms exist in many aspects of the water industry. The presence of biofilm on
membrane filters used in water treatment can clog the pores, causing flux reduction,
transmembrane pressure increase, higher energy consumption, and deterioration in
filtration performance.34 Biofilms cleaning can reduce membrane life significantly, thus
increasing the yearly investment cost.31,34 It was estimated that biofilm problem in
desalination plant worldwide could cost 15 billion US$ per year.31 The formation of
biofilms in a heat exchanger will significantly reduce the heat transfer efficiency, as
convective heat transfer is inhibited and only diffusive heat transfer can occur.35
Moreover, an increase in friction resistance due to biofilm growth will increase energy
consumption, subsequently increasing the cost.35 Biofilm growth on cooling tower
water system may decrease the efficiency to 10% and posed as health risks, as it can
release pathogens into the air.32 Biofilms growth on water-contacting industrial
pipelines can cause corrosion.36 It was estimated that microbial induced corrosion cost
about 16 to 17 billion US$ per year.36 In drinking water industry, biofilms presence can
reduce water quality and act as a reservoir for pathogenic bacteria.32,37 In order to

Chapter 2
16
prevent the occurrence of pathogens and biofilms, disinfectants such as chlorine or
chloramine are commonly added to the water, however this treatment is not necessarily
able to eradicate biofilms.38 Chloramine, which is preferred compared to chlorine due to
its deeper penetration into biofilms, less formation of toxic by-products, and higher
residual throughout distribution systems, is proven to be ineffective towards the
ubiquitous biofilm of nitrifying bacteria, although it was able to penetrate the biofilm
effectively.39,40 Moreover, it was found that nitrifying biofilms and their soluble
microbial products could accelerate the decay of chloramine, releasing chlorine and
ammonia, where the latter is the main nutrient needed for the growth of nitrifying
biofilms, thus promoting the formation of nitrifying biofilm.41–43
Although biofilm problems have been recognised since a long time ago, the
problems persist. It is believed that the increased tolerance or resistance that biofilm has
shown towards antimicrobials, metal cations, and disinfectants is to blame.38,44 Cells in
a biofilm have different traits that make them difficult to eradicate compared to the
planktonic counterpart.14 For instance, 100 times concentration of disinfectants or other
antimicrobials is needed to eradicate biofilm compared to their planktonic
counterpart.45,46 The treatment of cells with metals can kill planktonic bacteria easily,
however, it will trigger protective mechanism for biofilm.44,47
There are several hypotheses that are proposed for biofilm tolerance to biocidal
agents. The main hypothesis is the barrier of the matrix.1 This mechanism is especially
applicable for reactive (such as chlorine), charged (such as metal cations) and large
biocidal agents (some antibiotics).1 EPS matrix, which is mainly negatively charged, is
able to neutralise the charge of several biocides, such as metal cations, aminoglycosides,
and polypeptides, thus render them harmless.18,46 Moreover, the presence of EPS will
dilute biocidal agents to sub-lethal concentration.1 It was reported that chlorine level

Chapter 2
17
inside a biofilm is only 20-25% of the bulk chlorine concentration, with maximum
penetration of 50 to 100 µm.38,39 Moreover, chlorine and chloramine are susceptible to
reaction with organic matter which is present in the EPS, thus decreasing their
disinfection efficiency.38 EPS is also able to neutralise the enzymatic activity of several
enzymes. For instance, β-lactamase activity against Pseudomonas aeruginosa was
neutralised inside the matrix.1 It was also shown that the biofilm matrix could bind
ciproflaxin, thus reducing its concentration.33 The matrix also acts as a barrier for the
toxicity of metals. Metal ions are bound by components of the matrix, such as
extracellular polymers, cell membranes, and cell walls, thus sequestering the metals and
prevent them from interfering in important metabolic processes.44
Subpopulation with different physiological states that consists in a biofilm
structure will eventually contribute to the overall resistance of biofilm. The exposure to
a sub-lethal concentration of biocides can trigger the development of mutagenesis in a
subpopulation of cells, making them able to resist other biocides.38,46 Bacteria cells in a
biofilm also can undergo adaptive physiological changes in response to the toxic
concentration of metal species.44,47 For instance, biofilm treatment with toxic metal
triggers a protective response by upregulating exopolysaccharide production, while the
same treatment can kill planktonic cells easily.47 Some cells are able to up-regulate
genes for efflux pumps, thus cells can get rid of toxic molecules without harming the
cells.38 These factors/genes contributing to resistance can be easily transferred to other
cells due to the high cell density inside biofilm structure.48 Consequently, it was
reported that resistant genes could be detected in an environment where the parent
resistant genes are not available. For instance, vancomycin-resistant genes were
detected in drinking water systems where vancomycin-resistant enterococci are absent.48

Chapter 2
18
These will hence lead to the appearance of more biofilms that are capable of resisting
biocides.
As mentioned before, some cells in a biofilm do not have access to oxygen due
to the concentration gradient inside biofilm structure, leading to the presence of bacteria
with halted or slow metabolism.33,46 Slow growing bacteria have been shown to be more
resistant to biocides than fast growing bacteria.33 It is most likely attributed to the mode
of action of antibiotics that are only effective against bacteria with a certain degree of
activity, due to the antibiotics mechanism that only disrupts microbial processes, such as
cell wall synthesis.1,46 Moreover, some antibiotics are not effective in anaerobic
environment or at acidic pH, due to the accumulation of bacteria metabolic waste.13
Slow growing cells in anoxic environment also have increased tolerance to metal ions,
possibly due to the alteration in metal speciation, decreased metabolic ROS production,
or decrease in metal-catalysed Fenton-type reaction which requires oxygen.44
A special group of bacteria commonly known as persisters are 100 to 10000
times more commonly found inside biofilm than planktonic population.38,44 This
subpopulation of bacteria can easily enter a slow growing or starving state with reduced
metabolism during biocides treatment, rendering them highly tolerant to biocides.46,49
When biocides treatment has passed, persisters can switch back to growing state and
repopulate the biofilm (Figure 2.3),21,49 thus causing biofilm-related problem relapse.49

Chapter 2
19
Figure 2.3 Schematic of biofilm problem relapsed due to persister cells. Adapted from
Lewis, 2007.49
The interaction between bacteria, either the same species or cross-species inside
a biofilm also contributes to biofilm resistance.38,50 For instance, it has been shown that
Pseudomonas aeruginosa cells inside a biofilm can inactivate the antibiotics tobramycin
and gentamicin by secreting a periplasmic cyclic glucan that will complex with the
antibiotics.17 Another mechanism that has been suggested is the chemical interaction
between EPS from each species produce a highly viscous matrix, which will limit the
internal biocides concentration even further.38
Based on the above, it is believed that biofilm resistance to antimicrobials and
disinfectants is multifactorial, i.e. there is no one specific mechanism that solely
Antibiotic/disinfectants
treatment
Treatment stopped
Persister cells remained
Biofilm and planktonic bacteria
Biofilm repopulation

Chapter 2
20
contributes to biofilm resistance (Figure 2.4).38,46 Moreover, it is generally accepted that
the improper use of disinfectants and antibiotics would trigger the formation of bacteria
with increased resistance toward specific biocides, such as Methicillin-Resistant
Staphylococcus aureus (MRSA). The continuing problem caused by biofilm and
difficulty in eradication have driven the need for novel strategies in combating biofilm
that do not depend on the use of conventional disinfectants or antibiotics and most
importantly, do not trigger bacterial resistance.
Figure 2.4 Schematic of multifactorial biofilm resistance mechanisms.
2.3. Current strategies for biofilm prevention and removal
Antibiofilm strategies generally can be divided into three categories: materials
that can prevent biofilm adhesion onto surfaces, materials that can prevent biofilm
growth or induce cells killing, and materials that can disrupt established biofilms.13
-------
++
++
++
++
++
++
Horizontal
gene transfer
Efflux
pump
Neutralisation of
charged biocideBacteria
with
protective
response
Biocide binding by
EPS components
Persister
population
Reduction in
biocide
concentration
Slow growing
population
Anaerobic
environment
: biocide, antibiotics, or metal cations

Chapter 2
21
2.3.1. Adhesion prevention
This antibiofilm strategy is based on the modification of surface properties to
prevent the initial adhesion of bacteria. Surface wettability is the property that is most
commonly altered in designing biofilm-free surface.51 The typical method is by
incorporation of hydrophilic polymer brush such as poly(ethylene glycol) (PEG) and
poly(ethylene oxide) (PEO).12,52,53 Both of these materials could prevent the attachment
of bacteria due to the formation of water layer near the surface that prevents proteins
from attaching.54,55 However, PEG is known to undergo oxidation especially in the
presence of oxygen and transition metal ions, causing the surface to lose their
hydrophilic property.53,56,57 Another polymers that have been used before are
zwitterionic materials such as poly(sulfobetaine) (pSBMA) and poly(carboxybetaine)
(pCBMA), which have similar efficacy towards resisting protein adsorption as PEG.57
These materials have a strong electrostatically induced hydration layer that creates
superhydrophilic surfaces.53,58
Another way to increase hydrophilic property of a surface is to incorporate
inorganic nanoparticles in the surface, and since nanoparticles may also have biocide
properties, their incorporation can lead to an effective antifouling surface. For instance,
the incorporation of silver nanoparticles onto poly(vinylidene fluoride) (PVDF)
membrane improved the surface hydrophilicity. It was proposed that the silver ions
released from the nanoparticles are adsorbed on the nanoparticles in the form of
hydrated silver ions which increase surface hydrophilicity.59 The use of Mg(OH)2
nanoparticles in PVDF membrane increase the number of –OH groups on the surface,
subsequently increase the hydrophilicity.60

Chapter 2
22
The effect of surface functional groups on protein attachment also has been
investigated, and it was concluded that functional groups that can resist protein
adsorption have four similar characteristics: they contain polar functional groups or
hydrophilic groups, contain hydrogen bond acceptor groups, do not contain hydrogen
bond donor groups, and have no net charge/the overall electrical charge is neutral.61,62
Surfaces with water contact angle greater than 150° (known as
superhydrophobic) have been used as an antibiofilm surface. This material is known to
have fouling release property whereby bacteria attachment is not prevented, however,
the bond between bacteria and surface is weakened, therefore the attached cells are
more easily removed by hydrodynamic shear forces.58 Such materials are inspired by
lotus leaf, which has excellent non-wettability. The topography of lotus leaf consists of
few micrometres tall pillars and spaced approximately 20 µm between each other. These
pillars are covered with smaller scale protrusions (approximately 0.2-1 µm) and wax
(Figure 2.5).63 The properties of lotus leaf inspire the fabrication of fouling release
materials, which can be synthesised by combining micro or nanostructure with low
surface energy materials.64,65 Various methods have been used in producing surface
roughness that acts as the surface nanostructure, such as lithography, template-based
techniques, electrospinning, sol-gel synthesis, layer-by-layer deposition, etching,
chemical vapour deposition, electrochemical deposition, electroless galvanic deposition,
and anodic oxidation.65 Fluorine based chemicals are commonly used as the low surface
energy material.66 However, recent research is more focused on synthesising polymer-
nanoparticles composite, due to the toxic nature of fluorine-based chemical.
Nanoparticles are used to provide surface roughness, while polymer acts as low surface
energy material. Several polymers have been used, such as polystyrene (PS), polyvinyl

Chapter 2
23
chloride (PVC), polymethylmethacrylate (PMMA), poly(methylhydrosiloxane) (PMHS)
or polyethylene (PE).66–68
Figure 2.5 Illustration of lotus leaf topography. Adapted from Chen et al., 2012.63
A material that consists of film with lubricating liquid locked by
micro/nanoporous substrate known as SLIPS (Figure 2.6) has been synthesised and
investigated as antibiofilm surface.69–71 It was shown that SLIPS can reduce the
adhesion of Pseudomonas aeruginosa, Staphylococcus aureus, and Escherichia coli 35
times more effectively compared to PEG surface. The antibiofilm property of SLIPS
originates from the very weak adhesion of bacteria to the fluid interface, therefore
adhered bacteria can be easily removed.52,70
Figure 2.6 SLIPS synthesis. Adapted from Wong et al., 2011.69
The synthesis of SLIPS is based on 3 criteria: (1) higher chemical affinity
between lubricating fluid and solid compared to the affinity between ambient fluid and
solid, (2) stable and complete wetting of the solid by the lubricating fluid and (3)
~20 nm
200 nm – 1 µm
water
Functional or
textured solid
Locked lubricating
liquid

Chapter 2
24
lubricating fluid and ambient fluid are immiscible.70 The liquid repellency of SLIPS is
insensitive of texture geometry but highly dependent of the lubricating liquid.69
Other surface properties can also be altered to reduce bacterial adhesion.71 For
example, the topography of poly(dimethylsiloxane) (PDMS) which consists of different
riblets forming diamond-like shape pattern that was inspired by shark skin was shown to
be able to reduce biofilm formation from Staphylococcus aureus compared to the
smooth PDMS (Figure 2.7).72 Even the presence of uniform rectangular, square, or
cylindrical posts on PDMS surfaces has been shown to reduce bacterial attachment,
although the exact mechanism is not understood yet.73 Materials with high surface
energy were found to have higher attachment rates of ammonia oxidising bacteria,
specifically Nitrosomonas europaea and Nitrosospira multiformis.74
Conflicting findings on the effect of material stiffness on bacterial attachment
have been reported. For instance, attachment of Escherichia coli and Staphylococcus
aureus cells were found to correlate positively with increasing hydrogel stiffness.75
However, a negative correlation was found for Escherichia coli RP437 and
Pseudomonas aeruginosa PAO1 attachment on poly(dimethylsiloxane), i.e. increased
stiffness reduced bacterial attachment.76 The exact mechanism through which material
stiffness affects bacterial attachment is yet to be elucidated.16

Chapter 2
25
Figure 2.7 SEM images of Staphylococcus aureus attachment on smooth (left) and shark
skin inspired (known as Sharklet AFTM) PDMS surfaces on day 0 (A and B), day 2 (C and
D), day 7 (E and F), day 14 (G and H) and day 21 (I and J). Taken from Chung et al.,
2007.72
Although adhesion prevention holds the potential to be applied as an antibiofilm
strategy that will not trigger bacteria resistance, inhibition of adhesion can only be
achieved temporarily. Different species of bacteria have evolved complex mechanisms
that allow attachments in a wide range of environmental condition.14 For instance,
bacteria have been shown to be able to alter the surface interaction through the

Chapter 2
26
production of molecules that modify the physicochemical surface properties.5 Once a
pioneer bacterial species is able to overcome the repulsive force of the surface, then a
conditioning layer, which usually consists of proteins and bacteria cells, will be formed.
This conditioning layer will limit the action of the coating due to the accumulation of
cells on the surface, which will attract further bacterial attachment, and thus inactivates
the antibiofilm action of the surface.
2.3.2. Biofilm cells killing and biofilm maturation inhibitor
In this strategy, biofilm formation can be prevented by utilising novel
antimicrobial attached on surfaces that can kill biofilm cells. This material can be
differentiated into two types, surfaces that can kill cells instantly upon contact (most
commonly known as contact killing surfaces) or materials that can leach or release
antimicrobial agent that will kill the surrounding cells.73
One example of “contact-killing surface” uses cationic antimicrobials (such as
quaternary ammonium compounds or alkyl pyridium) as the active biocide, where they
can disrupt cytoplasmic membrane and release cellular content upon contact.77,78
However, the use of quaternary ammonium compounds have been proven to trigger
bacteria resistance,78,79 which is the main problem in utilisation of cell killing in
antibiofilm material. Copper and copper oxide surfaces also possess contact killing trait
and hospital trials that have been conducted showed reduction in surface
contamination.80–82
Modification in surface topography has also been reported to be able to kill
bacteria upon contact. Cicada wings are able to induce cracks in a Pseudomonas

Chapter 2
27
aeruginosa cell that leads to bacterial death solely based on the structure of cicada
wings, composed of nanopillars or nanorods, usually 200 nm tall and spaced 170 nm
apart from centre to centre. Upon contact with Pseudomonas aeruginosa cells, the
nanopillars penetrate and cause cracks on the bacterial cell wall. Eventually, the cells
rupture and sink between the nanopillars (Figure 2.8).83 However, these lysed cells will
aid in the formation of conditioning layers, which, as mentioned before, will aid in the
attachment of other cells and biofilm formation.
Figure 2.8 Cell rupture mechanism by cicada wings. Taken from Ivanova et al., 2012.83
Antibiofilm surfaces that depend on antimicrobial/biocide agent release can be
achieved either physically by soaking or impregnating the carrier material in biocides
(adsorption), or chemically by covalent attachment of the biocides with the surface.84,85
Antimicrobial peptides, which have been shown to exhibit wide antimicrobial action
towards bacteria and fungi and believed not to trigger resistance response, have been
covalently attached to stainless steel or polymer films.84,85 However, it has been
reported that attachment of antimicrobial peptides onto surfaces severely depleted its
a
b
c
d

Chapter 2
28
effectivity, due to the inability of the peptide to interact with cell membrane, and
subsequently, inability to rupture the cell membrane, which is the main mechanism
employed for cell killing by peptides.85
Overall, the long-term benefit of contact killing strategy is tricky as the surface
quickly becomes covered with dead bacteria, which will provide conditioning films for
other bacteria to attach and form biofilms, thus limiting the effect of any active
surface.14,86 For materials that depend on biocide leaching, the limited reservoir of
biocides and difficulty in controlling the leaching kinetic must be considered.
Insufficient biocide release will not be effective in eradicating the biofilms and might
trigger the formation of more resistant biofilms instead. Therefore, novel methods that
can prevent biofilm maturation but not necessarily by cell killing and do not trigger
bacterial resistance are needed.
In recent years, the application of metal-based material or nanoparticles for
antimicrobial application has increased tremendously.53,87 Silver is the most widely
known nanomaterial that exhibit cytotoxicity, however, it has been reported to trigger
bacterial resistance.88 In addition, researchers have focused on the use of metal oxide
nanoparticles because they are cheaper and considered to be non-toxic to human.53,87
Some examples of metal oxide nanoparticles with antibacterial activity are titanium
dioxide (TiO2), silicon dioxide (SiO2), magnesium oxide (MgO), copper oxide (CuO),
and zinc oxide (ZnO).53 Other non-metallic nanoparticles are also known to be
bactericidal, such as carbon nanotubes and graphene.53
Toxicity of metal and metal oxide based surfaces is highly dependent on the
release of the toxic metal ions. For instance, the toxicity of copper and copper oxide
materials depend on the release of toxic Cu2+ or Cu+ ions.81,89 On the contrary,

Chapter 2
29
cytotoxicity of metal and metal oxide nanoparticles is complex and multi-cytotoxic
origins can occur from a single source nanoparticle.87,90 For instance, attachment of
nanoparticles to the bacterial membrane from electrostatic interaction can disrupt
bacterial membrane integrity.90 Bacteria cells are also able to internalise the
nanoparticles, followed by intracellular dissolution of the nanoparticles inside the cells
resulting in further cell damage, known as Trojan horse effect.91 Nanoparticles can
locally change microenvironments near the bacteria, thus producing reactive oxygen
species (ROS) or increasing the solubility of nanoparticles or metal cations.90,92 It has
been reported that copper oxide nanoparticles toxicity against Escherichia coli
originates from complexes leached from the nanoparticles, while the nanoparticles itself
do not play any additional cytotoxic effect to the bacteria.92 However, when zinc oxide
was used, it was found that its toxicity originates not only from the leachate complex
but also the nanoparticles solid itself.91
Figure 2.9 Schematic of antimicrobial mechanisms of metal and metal oxide nanoparticles.
Adapted from Li et al., 200887 and Hajipour et al., 2012.90
ROS
Disruption of
membrane or cell
wall
Generation of reactive
oxygen species
e-
e-
M+ Release of
metal ions
Protein
oxidation
DNA damage
Interruption of
transmembrane
electron transport

Chapter 2
30
It is also known that toxicity of nanoparticles increased with decreasing size of
nanoparticles, as shown for zinc oxide, copper oxide, and silver nanoparticles.92–94 This
is probably due to the fact that smaller sized particles have higher surface charge density
as a result of the increased surface area per unit volume, which therefore imparts more
toxicity from its electrostatic interaction with the negatively charged cell membrane.93
In the case of copper oxide, copper leaching and the subsequent formation of a copper-
peptide complex is responsible for the nanoparticles toxicity, a phenomenon that was
not observed for micrometer-sized copper oxide.92 The interaction between silver
nanoparticles with the bacterial membrane is higher when the nanoparticles size is less
than 5 nm, possibly attributed to easier penetration of the nanoparticle into the cell.94
Morphology of nanoparticles was also found to affect their toxicity. For
instance, octahedral and corner-truncated octahedral cuprous oxide (Figure 2.10a, b and
Table 2.2) is more toxic against Bacillus subtilis compared to Staphylococcus aureus,
Streptococcus faecalis, Pseudomonas aeruginosa, and Enterobacter cloacae.95 On the
contrary, edge-truncated cubic (Figure 2.10j; Table 2.2) showed bacteriostatic effects to
all five bacteria tested. It was hypothesised that the difference in antibacterial activity is
attributed to differences in dominant surface facets ({111} for octahedrons and {100}
for cubes), subsequently exhibiting different adsorption and desorption behaviour
towards the bacteria.95 Similar observation was also reported for silver nanoparticles,
where truncated triangular silver nanoparticles with {111} facet as the dominant facet
have stronger antibacterial effect towards Escherichia coli compared to spherical and
rod-shape silver nanoparticles.96

Chapter 2
31
Figure 2.10 Scanning electron microscopy images of cuprous oxide crystals synthesised at
various condition by glycine-assisted mixed-solvothermal method. Shapes descriptions: (a)
octahedral, (b) corner-truncated octahedral, (c) hexa-cone, (d) hexa-rod, (e) hexa-pod, (f)
poly-cone, (g) polyhedral, (h) polyhedron, (i) cubic structure with truncated edge and/or
concave face-centres, and (j) edge-truncated cubic structure without concave face-centres.
Taken from Pang et al., 2009.95
Table 2.2 Corresponding minimum inhibitory concentrations of different shape cuprous
oxide crystals (refer to Figure 2.10). Taken from Pang et al., 2009.95
Minimum inhibitory concentration (µg/mL)
Shape Bacillus
subtilis
Staphylococcus
aureus
Streptococcus
faecalis
Pseudomonas
aeruginosa
Enterobacter
cloacae
a 12.5 >50 >50 >50 >50
b 6.25 >50 >50 >50 >50
c 6.25 >50 25 >50 25
d 12.5 6.25 25 >50 >50
e 25 >50 >50 >50 >50
f 12.5 >50 >50 >50 >50
g >50 25 >50 25 >50
h 25 25 12.5 >50 >50
i 25 12.5 25 12.5 >50
j 12.5 12.5 12.5 25 6.25
The main drawback in metal and metal oxide based materials is difficulty in
controlling the toxic metal ions release. If the metal ions release is not enough, then it
might trigger the formation of resistant biofilm instead.44 Moreover, metal ions at low
concentrations are needed to support bacterial growth. Metal-based toxicity also relies
on the surrounding condition. For instance, iron coupons exhibit different toxicity in
copper containing Na-HEPES buffer and copper containing Tris-Cl buffer due to
different complexation of copper in the buffer.89 Tris-Cl buffer also showed higher

Chapter 2
32
copper ion release compared to phosphate buffer saline solution.81 It was also reported
that copper oxide nanoparticles might exhibit high toxicity in water that is rich in
organics and low toxicity in freshwater or salty environments due to different copper
leaching behaviour.92
Utilisation of bacteriophage as antibiofilm therapy also have been suggested.97
Bacteriophage are viruses that will inject its genome into host cell’s genome, in this
case, bacteria, causing bacterial cell lysis.98 However, bacteriophages have been shown
to easily diffuse through EPS and produce enzymes that could degrade EPS, such as
depolymerases and dispersin.38,97,99 For instance, phage-induced depolymerases are able
to reduce the formation of Pantoea agglomerans, Serratia marcescens, and
Pseudomonas aeruginosa biofilms.97 Inoculation of ultrafiltration membrane system
with biofilm-forming Pseudomonas aeruginosa, Acinetobacter johnsonii, and Bacillus
subtilis and its corresponding specific phages are able to reduce the number of biofilm
by 40-60% compared the control (bacteria only).100 However, due to the bacteriophage
nature that can only infect a specific type of bacteria, a cocktail of different
bacteriophage is needed to combat multi-species biofilm, thus limiting its
application.97,98 In fact, it was shown that a three-phages cocktail is needed just to
eradicate single species biofilms of Enterobacter cloace.101 Moreover, supplemental
phage addition is sometimes needed due to the regrowth of biofilm after 24 to 48 hours,
as reported before.102 Phage-resistant biofilm was also obtained after the treatment,102
indicating that phage treatment can activate biofilm resistance.
Some acids are recently identified as effective antibiofilm agents. For instance,
acetic acid, either as liquid or dry salts, was found to eradicate Pseudomonas
aeruginosa and Staphylococcus aureus.103,104 The observed antibiofilm activity of acetic
acid is not correlated to pH drop, as a control experiment using hydrochloric acid does

Chapter 2
33
not yield similar effect. Therefore, the inhibition effect observed is caused by the acetic
acid molecules itself.103 Free nitrous acid (FNA), which is the protonated form of nitrite
(HNO2), also has been reported as an antibiofilm agent. It was found to be effective
against many water-related microbial consortiums, such as Nitrosomonas,105
Pseudomonas aeruginosa,106 and Desulfovibrio vulgaris.107 It was also found to be
effective against mixed consortiums of activated sludge,108 sewer biofilms,109 and
reverse osmosis membrane biofilms.110 FNA toxicity mechanism toward bacteria
involves multiple pathways. For instance, treatment of Pseudomonas aeruginosa with
FNA was found to inhibit denitrification activity, which subsequently led to starvation.
At the same time, cell metabolism, protein synthesis, and nucleic acid replication are
halted.111
In recent years, the use of nitric oxide as an antibacterial agent has gathered
attentions. Nitric oxide is known to be toxic against a large number of fungi, bacteria,
viruses, and parasites.112 It is also involved in wound healing regulator and has been
used by eukaryotes in combating bacterial infection.113 Nitric oxide interactions with
reactive oxygen species will produce reactive nitrogen species that can either kill
bacteria (bactericidal) or inhibit bacteria growth (bacteriostatic).112,114,115 Several
mechanisms of nitric oxide antibacterial property have been recognised, such as
interference with DNA replication and cell respiration, inhibition of enzyme function,
and lipid peroxidation.112,115 Although nitric oxide acts by cell killing, few reports have
stated that nitric oxide does not trigger bacteria resistance, which makes nitric oxide
treatment for biofilm control very attractive.113,116 Due to its multiple toxicity
mechanisms, several bacterial mutations have to occur at the same time for the bacteria
to survive,116 therefore, bacterial resistance caused by nitric oxide is difficult to occur.

Chapter 2
34
2.3.3. Biofilm disruption
The final strategy that has been used in eradicating biofilm can be achieved by
disrupting or dispersing (removal) established biofilm. As mentioned before, EPS
consists of different components, such as proteins, polysaccharides, and extracellular
DNA. Therefore, utilisation of enzymes that can degrade specific components of EPS,
such as proteolytic (for protein hydrolysis), polysaccharases (for polysaccharide
hydrolysis), or deoxyribonuclease I (for degradation of extracellular DNA) as
antibiofilm strategy have been reported.98,117,118 The combination of bacteriophages
treatment with EPS degrading enzymes such as dispersin also have been reported.99
However, the use of enzymes as antibiofilm substances are hindered by the limited
enzyme stability and activity in different environmental condition. High temperature
will cause enzymes to denature, while low temperature can decreases enzymatic
activity.98 Enzymes are also sensitive to high salt concentration.98 Change in pH value
also has been reported to alter enzymes efficiency.117 Moreover, free nitrous acid, which
has been mentioned in the previous section, was found to degrade EPS of activated
sludge.108
Another method that has been investigated in biofilm control is to target cellular
communication (quorum sensing), which is a mechanism used by biofilms to coordinate
their communal behaviour, such as biofilm maturation and pathogenesis.13,98,119 Biofilm
maturity can be inhibited by the use of substances that can inhibit quorum sensing,
commonly known as quorum quencher. There are several methods that can be used to
inhibit quorum sensing process, such as reducing the activity of the receptor protein,
inhibition of quorum sensing signal molecule production, degradation of quorum
sensing molecule, and using quorum sensing analogues.119 Quorum sensing analogues
generally have a similar molecular structure with the real quorum sensing species,

Chapter 2
35
therefore it can bind to quorum sensing receptor and inhibit bacterial communication.98
Some of the analogues that have been used as inhibitor are halogenated furanones,
natural furanones and its derivatives, (5-oxo-2,5-dihydrofuran-3-yl)methyl alkanoate, 3-
oxo-C12-(2-aminocyclohexanone), and several polyphenolic compounds produced by
plants, such as vanillin (4-hydroxy-3-methoxybenzaldehyde).98,119 Since furanones are
known to be toxic and carcinogenic for human,120 their application for biofilm control
will be limited.
Energy is required to regulate microbial behaviour, including biofilm formation.
Energy synthesis in cell occurs through the phosphorylation of adenosine diphosphate
(ADP) to adenosine triphosphate (ATP).121 Therefore, substances that can prevent ATP
synthesis, known as chemical uncouplers, has been shown to not only disrupt the
stability of biofilms but can also inhibit biofilm formation and trigger biofilm
detachment.98 Chemical uncouplers are usually weak acids that can dissolve lipid and
carry protons across the cellular membrane. It will enter the cell while carrying
hydrogen ion and deprotonate once inside the cell due to higher pH value. This
deprotonation will deplete the amount of protons available for oxidative
phosphorylation.98 The presence of uncouplers has been shown to reduce the net
synthesis of cellular ATP by 75%, thus reducing the number of extracellular
polysaccharides and protein content in biomass,122 which as mentioned before is the
main protective mechanism employed by biofilm, thus weakening the biofilm structure
and subsequently, the biofilm itself. Chemical uncouplers are also able to cause cell
autolysis by inducing proton gradient loss across the cellular membrane, which
eventually will inhibit microbial attachment or biofilm formation.98 Moreover, the use
of chemical uncouplers can also inhibit biofilm formation through suppression of
quorum sensing substances such as autoinducer-2 (AI-2) and N-acylhomoserine

Chapter 2
36
lactones (AHL).123,124 Some examples of chemical uncouplers are 3,3’,4’,5-
tetrachlorosalicylanilide (TCS), sodium dodecyl sulphate, 2,4-dinitrophenol (DNP), and
dicyclohexylcarbodiimide.98,123,124 However, most of chemical uncouplers are classified
as toxic aromatic compounds,98 thus limiting their application.
As mentioned before (section 2.1), at the end of the life cycle, biofilm would
undergo dispersal to form planktonic bacteria that will able to colonise in a new surface.
Planktonic bacteria are easier to treat compared to biofilm, therefore biofilm removal by
activating dispersal event is an attractive alternative for antibiofilm treatment. Several
substances that have been described in the previous section, such as chemical
uncouplers and bacteriophage, are known to trigger biofilm detachment. Some
substances can cause biofilm dispersal through disruption of intracellular signalling, in
specific through alteration in c-di-GMP level (as described in section 2.1). Because of
their antibiofilm mechanism that exploits cellular signalling pathway, it is believed that
this mode of action will not trigger resistant response.116,125 Such substances are known
to be effective towards a wide spectrum of bacteria and are biodegradable.25 For
example, the fatty acid molecule known as cis-2-decanoic acid was able to induce
dispersal of wide spectrum single species biofilms.24 Some other naturally occurring
substances are also known to be able to induce biofilms dispersal, such as the
supernatant of a marine isolate Bacillus licheniformis that can disperse biofilms of
Micrococcus luteus, Bacillus subtilis, and Escherichia coli.126
Interestingly, low concentration of nitric oxide (below toxic level) was found to
induce biofilm dispersal of various monospecies biofilms, either naturally occurring
biofilms such as Nitrosomonas europaea127 or pathogens such as Pseudomonas
aeruginosa,128 Escherichia coli, and the yeast Candida albicans.129 It was also found to
be effective against multispecies biofilm naturally occurring in water systems34,129 or

Chapter 2
37
wound-relevant pathogenic biofilm.130 Additionally, treatment with nitric oxide was
found to render the biofilm susceptible to antimicrobials.128,131
2.3.4. Nitric oxide as a new antibiofilm agent
Although many strategies have been proposed, it is unlikely that one type of
action could eliminate biofilm successfully. If two different methods are used and each
individual approach is only partially effective, the combination is proposed to be more
efficient.132 For example, the antibiotics aminoglycoside tobramycin and macrolide
clarithromycin have been shown to have moderate activity against biofilms, however,
the combination of the two antibiotics can synergistically eradicate more biofilms.133
The combination of free nitrous acid and hydrogen peroxide can produce the highly
toxic reactive nitrogen intermediate such as peroxynitrite and nitrogen dioxide.134
Consequently, free nitrous acid-hydrogen peroxide combination was found to be highly
effective in the inactivation of anaerobic wastewater biofilms compared to the use of a
single agent. Additionally, it is even more desirable to achieve complete eradication of
biofilm by the combined use of treatments with different modes of action.38 For
example, biofilm treatment with chlorine and bacteriophage is able to reduce biofilm
growth and remove pre-existing biofilm.135 Nitric oxide, which can exert antibiofilm
activity through cell killing (at high concentration, typically mM) or biofilm dispersal
signal (at low concentration, typically nM),114 appears to be advantageous due to its
multi-approach as an antibiofilm agent. As mentioned before, nitric oxide has been
shown to be effective against wide range of bacterial species and it is believed that nitric
oxide will not trigger resistance.116,125 Moreover, control of nitric oxide release is made
easy due to its wide range of active concentration, as it is effective both at high or low

Chapter 2
38
concentration. Biofilm treatment with nitric oxide was also found to reduce the
formation of persisters.136 In fact, nitric oxide is being used by higher level organisms
(including humans) to combat bacterial infections, showing the natural origin of nitric
oxide as antibacterial/antibiofilm agent.137,138 For instance, mice that do not have the
ability to produce nitric oxide endogenously was found to be more susceptible to
microbial infection.139 Therefore, nitric oxide utilisation has emerged as an attractive
strategy for biofilm control.
2.4. Current strategies in nitric oxide utilisation
One of the main challenges in nitric oxide (NO) utilisation as an antibiofilm
agent is in its delivery.113 NO is a free radical with a short half-life and high reactivity,
so its action is only effective as far as 100 µm from its point of origin.112 Therefore,
most research focused on the design and synthesis of materials that can store and release
NO (commonly known as NO-releasing materials), which will be reviewed in the next
section. It should be noted that the majority of literature available on NO-releasing
materials focused on its application in biomedical fields, whereby NO is not only used
as antibiofilm agent but also being used to prevent thrombosis and infection in blood-
contacting biomedical devices.140
2.4.1. Nitric oxide-releasing materials for application in biomedical field
NO-releasing materials generally can be divided into two categories: materials
that are able to adsorb gaseous NO followed by release or materials that release NO via
NO donor pathway. Adsorption of NO gas into delivery vehicles can be achieved by

Chapter 2
39
means of zeolites and metal organic framework. Zeolite-A, which consists of alternating
SiO4 and AlO4 tetrahedra with different metal cations, has been used to chemisorb NO
and was able to release the stored NO gas by exposure to wet gas (humidity).141,142 The
amount of NO that can be stored is dependent on the species and amount of metal
cations that were used, such as cobalt, nickel, copper, manganese, zinc, and
sodium.141,142 Enhancement of NO adsorption can be achieved by utilising metal organic
framework, which has been reported to be able to adsorb 4 times more NO compared to
zeolites.143 This enhancement in NO gas adsorption was correlated to the high density of
coordinatively unsaturated metal cations, such as copper, nickel, cobalt and iron.143
Liposomes, which are vesicles consisting of a liquid core and lipid layers, also
have been used to adsorb gaseous NO.144 NO-loaded liposomes were obtained by
exposing the lipid films, which were synthesised from 1,2-dipalmitoyl-sn-glycerol-3-
ethylphosphocoline, 1-2-dioleoyl-sn-glycero-3-phosphocholine, and cholesterol, to
pressurised NO gas. This process was followed by freezing and thawing to obtain the
final material.144
Nowadays, most NO-releasing materials were formed by exploiting NO donor.
NO donors are molecules that are able to decompose and liberate NO. The most widely
used NO donor for the synthesis of NO-releasing materials is diazeniumdiolates, which
can be synthesised by reacting secondary amines with high pressure of NO gas in the
presence of sodium methoxide or acetonitrile for the deprotonation of amine backbone
and to promote its nucleophilic attack on NO (Scheme 2.1).145,146 Diazeniumdiolates are
able to generate two molecules of NO (thus also known as NONOates) spontaneously in
physiological condition (mostly triggered by pH difference) and the release is not
influenced by biological factors.146,147 As a result, NONOates are the most widely used
NO donor in the synthesis of NO-releasing materials. The NO release profile from

Chapter 2
40
NONOates can be altered by designing structural backbone of the amine-containing
parent molecule (for instance, altering the sizes or substituted cyclic or linear aliphatic
amines) or the hydrogen bonding between NONOates and a primary amine that is
attached to the backbone molecule.140,148 Moreover, NONOates have been modified so
they can liberate NO after contact with a biofilm-related enzyme (β-lactamase) or to
liberate NO and gentamicin (a type of antibiotic) at the same time.149,150 However,
toxicity concerns due to the formation of nitrosamines by-product have been raised.140
Scheme 2.1 NONOates formation and decomposition from secondary amine-bearing
compounds in the presence of sodium methoxide (top) or acetonitrile (bottom). Adapted
from Riccio and Schoenfisch, 2012.146
Another type of NO donor that is widely used is S-nitrosothiols (RSNOs).
Several types of RSNOs are known to be available endogenously in blood and plasma,
such as S-nitrosoglutathione (GSNO), S-nitrosoalbumin (AlbNO), and S-nitrosocysteine
(CysNO).151 Due to the availability of RSNOs in vivo, it is believed that RSNOs are
more biocompatible than NONOates.148,151 A widely used synthetic RSNO known as S-
nitroso-N-acetyl-D-penicillamine (SNAP) is made by the reaction of thiols with
nitrosating agents, such as sodium nitrite.146 In contrary to NONOates, NO release from
RSNOs require triggers, such as thermal or photolysis, metals ions (such as copper or
iron), and reducing agents such as ascorbate or glutathione (Scheme 2.2).146,148

Chapter 2
41
Scheme 2.2 RSNOs formation and decomposition by copper, heat or light, and
glutathione. Adapted from Riccio and Schoenfisch, 2012.146
These different NO sources were then incorporated into different delivery
vehicles to generate NO-releasing materials, either inorganic or organic based. One
example of inorganic NO-releasing nanomaterials is silica-based nanoparticles. Silica
has gathered wide attention due to their ease of preparation, size control, excellent
biocompatibility, and stability.148 NO release can be controlled by modulating the
nanoparticle size, composition, and/or surface hydrophobicity.139 Slomberg et al.
investigated the use of silica nanoparticles with different sizes and aspect ratios but with
a similar amount of overall NO released and reported that particles with decreased size
and increased aspect ratio are more effective against Pseudomonas aeruginosa and
Staphylococcus aureus biofilms due to better NO delivery exhibited by the smaller NO-
releasing particles.152 NO release from sol-gel silica nanoparticles can also be altered by
varying the amine used for NONOates synthesis.153 Amine functionalised silica
nanoparticles which were synthesised from the condensation of tetraethoxysilane
(TEOS) or tetramethoxysilane (TMOS) in the presence of different silane, namely N-(6-
aminohexyl)aminopropyltrimethoxysilane (AHAP3), (aminoethylamino-

Chapter 2
42
methyl)phenethyltrimethoxysilane (AEMP3), or N-(2-aminoethyl)-3-
aminopropyltrimethoxysilane (AEAP3) were found to have different sizes. Conversely,
when the secondary amines were converted to NONOates, the silica nanoparticles
exhibited different NO release amount and half-lives (the time needed to release half of
NO loading).153 A “pre-formation” method, where the formation of NONOates from
aminoalkoxysilanes was done before the formation of NO-releasing silica nanoparticles
also could be employed.154 This method enables silica nanoparticles to exhibit higher
NO payload and NO release (up to 6.3 and 6.7 times greater) compared to the post-
formation method.
Metal nanoparticles have gathered attention as NO-releasing nanocarriers due to
the possibility to trigger the NO release by external stimuli, such as light or magnetic
trigger.146 For example, gold nanoparticles, which exhibited plasmon resonance
property, allow the use of light as the external stimuli.155 Moreover, gold nanoparticles
are inert/biocompatible and can be easily prepared and functionalised, therefore they
have been utilised in various biomedical applications, such as nanocarriers for drug
delivery, as contrast agent, and in nanosensors.148,155 Similar with silica nanoparticles, it
was found that NO release profile from gold nanoparticles is also dependent on the
amine source, where longest half-life was obtained by using ethylene diamine, while
hexadiamine gave the highest NO loading.156 However, it was found that gold
nanoparticles solubility in water is very limited. Thus, functionalisation with tiopronin
was performed, followed by attachment of either diethylenetriamine,
tetraethylenepentamine, or pentaethylenehexamine as the secondary amine.146,156
However, these materials have a very low conversion to NONOates due to the
destabilisation of the tiopronin-containing particles from the base required for

Chapter 2
43
NONOates conversion.156 Platinum-based material that enables the use of light as a
trigger for NO release also has been reported.146
NO-release triggered by magnetic fields could be achieved by employing
superparamagnetic iron oxide nanoparticles (SPIONs).148,156 Contrary to silica and gold
nanoparticles, most reported SPIONs utilises RSNOs as the NO-donor. For instance,
SPIONs which were prepared from the co-precipitation of iron(III) chloride hexahydrate
and iron(II) chloride tetrahydrate with ammonia, were reacted with mercaptosuccinic
acid as thiol source, which was subsequently reacted with sodium nitrite yielding S-
nitrosothiols functionalised SPIONs.156
Although numerous studies have focused on the use of metal-based
nanoparticles, organic-based /polymeric NO-releasing materials make up the majority of
literature. Polymers are preferred due to their encapsulation capability and low
toxicity.147 Incorporation of NO donors onto the polymeric materials can be achieved by
physical process (encapsulation or coating) or chemical process (covalent attachment).
The simplicity of physical process makes it especially attractive for large scale
production. In the physical method, NO donors are non-covalently encapsulated or
embedded within polymeric matrices. For example, PROLI (an example of commercial
NONOates with a half-time of 1.8 s) was incorporated within poly(lactic-co-glycolic
acid) (PLGA) and poly(ethylene oxide-co-lactic acid) (PELA) microparticles.146 RSNOs
such as SNAP also can be encapsulated in PLGA microsphere.157 Biomedical grade
polymers also have been investigated for RSNOs encapsulation, such as silicone
rubber/poly(dimethylsiloxane) (PDMS), Elast-eon E2As (a mixture of PDMS,
poly(hexamethylene oxide) and methylene diphenyl isocyanate), CarboSil
(thermoplastic urethane copolymer, PDMS, and polycarbonate), Tygon (poly(vinyl

Chapter 2
44
chloride) based), and Tecophilic SP-60D-60 (aliphatic hydrophilic polyether-based
polyurethane).158–160 Liposome can be used for encapsulation of Spermine NONOate,
where the release of NO from the lipid vesicle was triggered by temperature and pH
gradient.161
Although physical encapsulation is preferred due to its simplicity, concerns
regarding NO donors premature leaching and toxic nitrosamines release from
NONOates have been raised. Therefore, covalent attachment of NONOates onto
polymeric backbones has been applied to overcome this problem. For examples,
covalent attachment of NONOates to poly(vinyl chloride) (PVC), polyurethane (PU),
silicon rubber, poly(ethylenimine) (PEI), polyester, polymethacrylate, poly(vinyl
alcohol) (PVA) and hydrogels have been reported.140 Polymer nanoparticles, such as
core cross-linked star polymers, have been successfully conjugated with Spermine
NONOates and the subsequent nanoparticle is effective in preventing biofilm
formation.162 Another study synthesised core cross-linked polymer nanoparticles
containing NONOate and gentamicin complex for biofilm eradication. These
nanoparticle complexes were reported to synergistically disperse biofilm and kill
biofilm and planktonic cells at a higher efficiency compared to the combined effect of
gentamicin and NONOates separately.150 Covalent attachment between RSNOs onto
polymer backbones has also been reported.163
Another polymeric vehicle that has been widely employed for NO-releasing
materials is dendrimers. Dendrimers are macromolecules with a spherical shape that
comprises of highly repetitive and branched structure.164 Due to the structure,
dendrimers offer numerous functionalities which, when converted to NONOates or
RSNOs, allow high loading of NO to be stored.165 It was reported that around 2 to 6
µmol NO per mg of dendrimer can be accomplished.140,156 Dual action antibacterial

Chapter 2
45
dendrimer that combines NONOates with quaternary ammonium was reported to be
effective against Pseudomonas aeruginosa and Staphylococcus aureus.166
Although dendrimers enable the synthesis of delivery vehicles with high loading
of NO, its synthesis requires a laborious process. Therefore, micelles, which are core-
shell nanostructures consisting of amphiphilic block copolymers with hydrophilic and
hydrophobic segments, have been proposed as an alternative.148,164 The hydrophobic
inner cores of micelle was proposed to be able to protect NONOates from proton
catalysed NO liberation, thus enhancing its stability.167 GSNO incorporation into the
hydrophobic core was also found to stabilise the NO donor, as the NO release half-time
was extended by 5 times.168 However, as NO donors are mostly hydrophilic, a pre-
treatment step is required in order to load the NO donors into the core.148
Scheme 2.3 NO-releasing materials synthesis strategy in the biomedical field as discussed
in this review.
2.4.2. Application of nitric oxide donors in water industry
Although most research focused on the utilisation and synthesis of NO-releasing
materials for biomedical fields, NO-based treatment does hold the potential to be
applied in the water industry. Barraud et al. reported that the application of 500 nM
NO delivery
Gas
adsorptionNO donors
Zeolites and
metal organic
framework
Liposomes Inorganic Organic
Physical
encapsulation
Covalent
attachmentSilica Metal
Polymeric
matrixLiposomes Nanoparticles Dendrimers Micelles

Chapter 2
46
sodium nitroprusside (SNP; an example of metal based NO donor) to biofilms formed in
recycle and potable water distribution pipelines led to biofilm dispersal by 47% and
60%, respectively.129 Barnes et al. have reported that the use of the NONOates PROLI
at 24 hours intervals to a laboratory scale reverse osmosis membrane system can reduce
the rate of biofouling by 92%.34 The applications of PROLI daily to a membrane
bioreactor with high transmembrane pressure for 37 days (out of 155 days of operation)
are able to reduce the fouling resistance by 56%.169 When PROLI was applied since the
commencement of 85 days operation, a reduction of 32.3% in transmembrane pressure
and delay in biofilm bacterial community development was observed.169 Although
reports on the application of NO and NO-based materials in the water industry are
limited, the existing results suggest a high potential for NO application in water
industry.
2.5. Summary
In this review, biofilm life cycle, which started by attachment of cells onto
surfaces and ended with dispersal event, was presented. Biofilm formations have mostly
negative implications throughout many different fields, including medicine and
industrial water system. Microbial biofilms are ubiquitous in nature and their increased
resistance towards conventional antimicrobial agents and disinfectants drives the need
to develop novel antibiofilm strategy. In general, antibiofilm strategy can be divided
into 3 categories depending on the mode of action, with strategy combining several
modes of action is preferred due to its higher chance of success. NO, which exerts
antibiofilm activity either by cell killing at high concentration or dispersing biofilm at
low concentration, has emerged as an attractive solution. Due to its reactivity, NO

Chapter 2
47
utilisation depends on the synthesis of materials that can store and release NO at the
point of problem. Most NO-releasing materials are focused on the encapsulation or
conjugation of NO donor. The uses of NO donor against biofilms in membrane systems
and water pipelines have shown a high potential of NO to be applied as a new biofilm
eradication method in the water industry. However, daily dosing of NO donors such as
PROLI into industrial water systems is costly and not feasible as it may release NO
prematurely. In addition, most NO-releasing materials that are mentioned in this review
only have limited NO release longevity depending on the amount of NO that can be
adsorbed or loaded during synthesis. Thus, a better alternative would be to develop a
catalytic technology that is able to release NO continuously by converting endogenous
NO source, for instance, nitrite in water. Moreover, this technology will allow the
capability to control the timing and amount of NO released. Therefore, the application
of such materials, commonly known as NO-generating materials, in controlling water-
related biofilms, in specific nitrifying bacteria, would be the focus of this thesis.
2.6. References
1. Hall-Stoodley, L., Costerton, J. W. & Stoodley, P. Bacterial biofilms: from the
natural environment to infectious diseases. Nat. Rev. Microbiol. 2, 95–108
(2004).
2. Yang, L. et al. Combating biofilms. FEMS Immunol. Med. Microbiol. 65, 146–
157 (2012).
3. Garrett, T. R., Bhakoo, M. & Zhang, Z. Bacterial adhesion and biofilms on
surfaces. Prog. Nat. Sci. 18, 1049–1056 (2008).
4. Shrout, J. D., Tolker-Nielsen, T., Givskov, M. & Parsek, M. R. The contribution
of cell-cell signaling and motility to bacterial biofilm formation. MRS Bull. 36,

Chapter 2
48
367–373 (2011).
5. Rendueles, O. & Ghigo, J. M. Multi-species biofilms: How to avoid unfriendly
neighbors. FEMS Microbiol. Rev. 36, 972–989 (2012).
6. Flemming, H.-C. in Marine and Industrial Biofouling 3–12 (2009).
7. Okabe, S., Satoh, H. & Kindaichi, T. in Methods in Enzymology 496, 163–184
(Elsevier Inc., 2011).
8. Rittmann, B. E. & Manem, J. a. Development and experimental evaluation of a
steady-state, multispecies biofilm model. Biotechnol. Bioeng. 39, 914–922
(1992).
9. Daims, H. & Wagner, M. In situ techniques and digital image analysis methods
for quantifying spatial localization patterns of nitrifiers and other
microorganisms in biofilm and flocs. Methods in enzymology 496, (Elsevier Inc.,
2011).
10. Srey, S., Jahid, I. K. & Ha, S.-D. Biofilm formation in food industries: A food
safety concern. Food Control 31, 572–585 (2013).
11. Hall-Stoodley, L. & Stoodley, P. Developmental regulation of microbial
biofilms. Curr. Opin. Biotechnol. 13, 228–233 (2002).
12. Renner, L. D. & Weibel, D. B. Physicochemical regulation of biofilm formation.
MRS Bull. 36, 347–355 (2011).
13. Gupta, P., Sarkar, S., Das, B., Bhattacharjee, S. & Tribedi, P. Biofilm,
pathogenesis and prevention—a journey to break the wall: a review. Arch.
Microbiol. 198, 1–15 (2016).
14. Hori, K. & Matsumoto, S. Bacterial adhesion: From mechanism to control.
Biochem. Eng. J. 48, 424–434 (2010).
15. Teughels, W., Assche, N., Sliepen, I. & Quirynen, M. Effect of material
characteristics and/or surface topography on biofilm development. Clin. Oral
Implants Res. 17 Suppl 2, 68–81 (2006).
16. Song, F., Koo, H. & Ren, D. Effects of Material Properties on Bacterial Adhesion
and Biofilm Formation. J. Dent. Res. 94, 1027–1034 (2015).

Chapter 2
49
17. Estrela, A. B. & Abraham, W.-R. Combining Biofilm-Controlling Compounds
and Antibiotics as a Promising New Way to Control Biofilm Infections.
Pharmaceuticals 3, 1374–1393 (2010).
18. Flemming, H.-C. & Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 8,
623–633 (2010).
19. Sheng, G.-P., Yu, H.-Q. & Li, X.-Y. Extracellular polymeric substances (EPS) of
microbial aggregates in biological wastewater treatment systems: A review.
Biotechnol. Adv. 28, 882–894 (2010).
20. McDougald, D., Rice, S. A., Barraud, N., Steinberg, P. D. & Kjelleberg, S.
Should we stay or should we go: mechanisms and ecological consequences for
biofilm dispersal. Nat. Rev. Microbiol. 10, 39–50 (2011).
21. Stewart, P. S. & Franklin, M. J. Physiological heterogeneity in biofilms. Nat.
Rev. Microbiol. 6, 199–210 (2008).
22. Karatan, E. & Watnick, P. Signals, regulatory networks, and materials that build
and break bacterial biofilms. Microbiol. Mol. Biol. Rev. 73, 310–347 (2009).
23. Musk, D. J., Banko, D. a. & Hergenrother, P. J. Iron salts perturb biofilm
formation and disrupt existing biofilms of Pseudomonas aeruginosa. Chem. Biol.
12, 789–796 (2005).
24. Davies, D. G. & Marques, C. N. H. A fatty acid messenger is responsible for
inducing dispersion in microbial biofilms. J. Bacteriol. 191, 1393–1403 (2009).
25. Shrout, J. D. & Nerenberg, R. Monitoring Bacterial Twitter: Does Quorum
Sensing Determine the Behavior of Water and Wastewater Treatment Biofilms?
Environ. Sci. Technol. 46, 1995–2005 (2012).
26. Ha, D.-G. & O’Toole, G. A. c-di-GMP and its Effects on Biofilm Formation and
Dispersion: a Pseudomonas Aeruginosa Review. Microbiol. Spectr. 3, MB–
0003–2014 (2015).
27. Barraud, N., Kjelleberg, S. & Rice, S. A. Dispersal from Microbial Biofilms.
Microbiol. Spectr. 3, MB–0015–2014 (2015).
28. Caly, D. & Bellini, D. Targeting Cyclic di-GMP Signalling: A Strategy to
Control Biofilm Formation? Curr. Pharm. Des. 21, 12–24 (2015).

Chapter 2
50
29. Barraud, N. et al. Nitric oxide signaling in Pseudomonas aeruginosa biofilms
mediates phosphodiesterase activity, decreased cyclic di-GMP levels, and
enhanced dispersal. J. Bacteriol. 191, 7333–7342 (2009).
30. Nguyen, T.-K., Duong, H. T. T., Selvanayagam, R., Boyer, C. & Barraud, N. Iron
oxide nanoparticle-mediated hyperthermia stimulates dispersal in bacterial
biofilms and enhances antibiotic efficacy. Sci. Rep. 5, 18385 (2015).
31. Flemming, H.-C. in Biofilm Highlights (eds. Flemming, H.-C., Wingender, J. &
Szewzyk, U.) 81–110 (Springer, 2011). doi:10.1007/978-3-642-19940-0
32. Mattila-Sandholm, T. & Wirtanen, G. Biofilm formation in the industry: A
review. Food Rev. Int. 8, 573–603 (1992).
33. Mah, T. & O’Toole, G. Mechanisms of biofilm resistance to antimicrobial agents.
Trends Microbiol. 9, 34–39 (2001).
34. Barnes, R. J. et al. Nitric oxide treatment for the control of reverse osmosis
membrane biofouling. Appl. Environ. Microbiol. 81, 2515–2524 (2015).
35. Flemming, H.-C. Biofouling in water systems - cases, causes and
countermeasures. Appl. Microbiol. Biotechnol. 59, 629–640 (2002).
36. Coetser, S. E. & Cloete, T. E. Biofouling and biocorrosion in industrial water
systems. Crit. Rev. Microbiol. 31, 213–232 (2005).
37. Wingender, J. & Flemming, H. C. Biofilms in drinking water and their role as
reservoir for pathogens. Int. J. Hyg. Environ. Health 214, 417–423 (2011).
38. Bridier, A., Briandet, R., Thomas, V. & Dubois-Brissonnet, F. Resistance of
Bacterial Biofilms to Disinfectants: a review. Biofouling 27, 1017–1032 (2011).
39. Lee, W. H., Wahman, D. G., Bishop, P. L. & Pressman, J. G. Free chlorine and
monochloramine application to nitrifying biofilm: comparison of biofilm
penetration, activity, and viability. Environ. Sci. Technol. 45, 1412–1419 (2011).
40. Pressman, J. G., Lee, W. H., Bishop, P. L. & Wahman, D. G. Effect of free
ammonia concentration on monochloramine penetration within a nitrifying
biofilm and its effect on activity, viability, and recovery. Water Res. 46, 882–894
(2012).
41. Bal Krishna, K. C., Sathasivan, A. & Chandra Sarker, D. Evidence of soluble

Chapter 2
51
microbial products accelerating chloramine decay in nitrifying bulk water
samples. Water Res. 46, 3977–3988 (2012).
42. Maestre, J. P., Wahman, D. G. & Speitel, G. E. Monochloramine cometabolism
by Nitrosomonas europaea under drinking water conditions. Water Res. 47,
4701–4709 (2013).
43. Maestre, J. P., Wahman, D. G. & Speitel, G. E. Monochloramine cometabolism
by mixed-culture nitrifiers under drinking water conditions. Environ. Sci.
Technol. 50, 6240–6248 (2016).
44. Harrison, J. J., Ceri, H. & Turner, R. J. Multimetal resistance and tolerance in
microbial biofilms. Nat. Rev. Microbiol. 5, 928–938 (2007).
45. Meyer, B. Approaches to prevention, removal and killing of biofilms. Int.
Biodeterior. Biodegrad. 51, 249–253 (2003).
46. Olsen, I. Biofilm-specific antibiotic tolerance and resistance. Eur. J. Clin.
Microbiol. Infect. Dis. 34, 877–886 (2015).
47. Booth, S. C. et al. Differences in metabolism between the biofilm and planktonic
response to metal stress. J. Proteome Res. 10, 3190–3199 (2011).
48. Balcazar, J. L., Subirats, J. & Borrego, C. M. The role of biofilms as
environmental reservoirs of antibiotic resistance. Front. Microbiol. 6, 1–9 (2015).
49. Lewis, K. Persister cells, dormancy and infectious disease. Nat. Rev. Microbiol.
5, 48–56 (2007).
50. Sheng, Z. & Liu, Y. Effects of silver nanoparticles on wastewater biofilms. Water
Res. 45, 6039–6050 (2011).
51. Genzer, J. & Efimenko, K. Recent developments in superhydrophobic surfaces
and their relevance to marine fouling: a review. Biofouling 22, 339–360 (2006).
52. Tan, S. Y.-E., Chew, S. C., Tan, S. Y.-Y., Givskov, M. & Yang, L. Emerging
frontiers in detection and control of bacterial biofilms. Curr. Opin. Biotechnol.
26, 1–6 (2014).
53. Banerjee, I., Pangule, R. C. & Kane, R. S. Antifouling Coatings: Recent
Developments in the Design of Surfaces That Prevent Fouling by Proteins,
Bacteria, and Marine Organisms. Adv. Mater. 23, 690–718 (2011).

Chapter 2
52
54. Mansouri, J., Harrisson, S. & Chen, V. Strategies for controlling biofouling in
membrane filtration systems: challenges and opportunities. J. Mater. Chem. 20,
4567–4586 (2010).
55. Jeon, S. . & Andrade, J. . Protein—surface interactions in the presence of
polyethylene oxide. J. Colloid Interface Sci. 142, 159–166 (1991).
56. Ostuni, E., Chapman, R. G., Holmlin, R. E., Takayama, S. & Whitesides, G. M.
A survey of structure-property relationships of surfaces that resist the adsorption
of protein. Langmuir 17, 5605–5620 (2001).
57. Zhang, Z., Chao, T., Chen, S. & Jiang, S. Superlow fouling sulfobetaine and
carboxybetaine polymers on glass slides. Langmuir 22, 10072–10077 (2006).
58. Callow, J. A. & Callow, M. E. Trends in the development of environmentally
friendly fouling-resistant marine coatings. Nat. Commun. 2, 244 (2011).
59. Li, J.-H., Shao, X.-S., Zhou, Q., Li, M.-Z. & Zhang, Q.-Q. The double effects of
silver nanoparticles on the PVDF membrane: Surface hydrophilicity and
antifouling performance. Appl. Surf. Sci. 265, 663–670 (2013).
60. Dong, C. et al. Antifouling enhancement of poly (vinylidene fluoride)
microfiltration membrane by adding Mg(OH)2 nanoparticles. J. Memb. Sci. 387,
40–47 (2012).
61. Ostuni, E. et al. Self-Assembled Monolayers That Resist the Adsorption of Cells.
Langmuir 17, 6336–6343 (2001).
62. Chapman, R. G. et al. Surveying for surfaces that resist the adsorption of
proteins. J. Am. Chem. Soc. 112, 8303–8304 (2000).
63. Chen, P. Y., McKittrick, J. & Meyers, M. A. Biological materials: Functional
adaptations and bioinspired designs. Prog. Mater. Sci. 57, 1492–1704 (2012).
64. Zhang, M., Wang, C., Wang, S. & Li, J. Fabrication of superhydrophobic cotton
textiles for water-oil separation based on drop-coating route. Carbohydr. Polym.
97, 59–64 (2013).
65. Celia, E., Darmanin, T., Taffin de Givenchy, E., Amigoni, S. & Guittard, F.
Recent advances in designing superhydrophobic surfaces. J. Colloid Interface
Sci. 402, 1–18 (2013).

Chapter 2
53
66. Guo, Y. & Wang, Q. Facile approach in fabricating superhydrophobic coatings
from silica-based nanocomposite. Appl. Surf. Sci. 257, 33–36 (2010).
67. Xu, X. et al. Fabrication of Bionic Superhydrophobic Manganese
Oxide/Polystyrene Nanocomposite Coating. J. Bionic Eng. 9, 11–17 (2012).
68. Joneydi, S., Khoddami, A. & Zadhoush, A. Novel superhydrophobic top coating
on surface modified PVC-coated fabric. Prog. Org. Coatings 76, 821–826
(2013).
69. Wong, T.-S. et al. Bioinspired self-repairing slippery surfaces with pressure-
stable omniphobicity. Nature 477, 443–447 (2011).
70. Epstein, A. K., Wong, T.-S., Belisle, R. A., Boggs, E. M. & Aizenberg, J. Liquid-
infused structured surfaces with exceptional anti-biofouling performance. Proc.
Natl. Acad. Sci. 109, 13182–13187 (2012).
71. Li, J. et al. Hydrophobic liquid-infused porous polymer surfaces for antibacterial
applications. ACS Appl. Mater. Interfaces 5, 6704–6711 (2013).
72. Chung, K. et al. Impact of engineered surface microtopography on biofilm
formation of Staphylococcus aureus. Biointerphases 2, 89–94 (2007).
73. Dobosz, K. M., Kolewe, K. W. & Schiffman, J. D. Green materials science and
engineering reduces biofouling: approaches for medical and membrane-based
technologies. Front. Microbiol. 6, 1–8 (2015).
74. Khan, M. M. T., Ista, L. K., Lopez, G. P. & Schuler, A. J. Experimental and
theoretical examination of surface energy and adhesion of nitrifying and
heterotrophic bacteria using self-assembled monolayers. Environ. Sci. Technol.
45, 1055–1060 (2011).
75. Kolewe, K. W., Peyton, S. R. & Schiffman, J. D. Fewer Bacteria Adhere to
Softer Hydrogels. ACS Appl. Mater. Interfaces 7, 19562–19569 (2015).
76. Song, F. & Ren, D. Stiffness of cross-linked poly(dimethylsiloxane) affects
bacterial adhesion and antibiotic susceptibility of attached cells. Langmuir 30,
10354–10362 (2014).
77. Murata, H., Koepsel, R. R., Matyjaszewski, K. & Russell, A. J. Permanent, non-
leaching antibacterial surfaces—2: How high density cationic surfaces kill

Chapter 2
54
bacterial cells. Biomaterials 28, 4870–4879 (2007).
78. Hegstad, K. et al. Does the wide use of quaternary ammonium compounds
enhance the selection and spread of antimicrobial resistance and thus threaten our
health? Microb. Drug Resist. 16, 91–104 (2010).
79. Hasan, J., Crawford, R. J. & Ivanova, E. P. Antibacterial surfaces: the quest for a
new generation of biomaterials. Trends Biotechnol. 31, 295–304 (2013).
80. Santo, C. E., Quaranta, D. & Grass, G. Antimicrobial metallic copper surfaces
kill Staphylococcus haemolyticus via membrane damage. Microbiologyopen 1,
46–52 (2012).
81. Hans, M. et al. Role of copper oxides in contact killing of bacteria. Langmuir 29,
16160–16166 (2013).
82. Hans, M., Mathews, S., Mücklich, F. & Solioz, M. Physicochemical properties of
copper important for its antibacterial activity and development of a unified
model. Biointerphases 11, 018902 (2016).
83. Ivanova, E. P. et al. Natural bactericidal surfaces: mechanical rupture of
Pseudomonas aeruginosa cells by cicada wings. Small 8, 2489–2494 (2012).
84. Héquet, A., Humblot, V., Berjeaud, J. M. & Pradier, C. M. Optimized grafting of
antimicrobial peptides on stainless steel surface and biofilm resistance tests.
Colloids Surfaces B Biointerfaces 84, 301–309 (2011).
85. Onaizi, S. a. & Leong, S. S. J. Tethering antimicrobial peptides: Current status
and potential challenges. Biotechnol. Adv. 29, 67–74 (2011).
86. Lebeaux, D., Ghigo, J.-M. & Beloin, C. Biofilm-Related Infections: Bridging the
Gap between Clinical Management and Fundamental Aspects of Recalcitrance
toward Antibiotics. Microbiol. Mol. Biol. Rev. 78, 510–543 (2014).
87. Li, Q. et al. Antimicrobial nanomaterials for water disinfection and microbial
control: Potential applications and implications. Water Res. 42, 4591–4602
(2008).
88. Gunawan, C., Teoh, W. Y., Marquis, C. P. & Amal, R. Induced adaptation of
Bacillus sp. to antimicrobial nanosilver. Small 9, 3554–60 (2013).
89. Mathews, S., Kumar, R. & Solioz, M. Copper reduction and contact killing of

Chapter 2
55
bacteria by iron surfaces. Appl. Environ. Microbiol. 81, 6399–6403 (2015).
90. Hajipour, M. J. et al. Antibacterial properties of nanoparticles. Trends
Biotechnol. 30, 499–511 (2012).
91. Gunawan, C., Teoh, W. Y., Marquis, C. P. & Amal, R. Zinc Oxide Nanoparticles
Induce Cell Filamentation in Escherichia coli. Part. Part. Syst. Charact. 30, 375–
380 (2013).
92. Gunawan, C., Teoh, W., Marquis, C. & Amal, R. Cytotoxic origin of copper (II)
oxide nanoparticles: comparative studies with micron-sized particles, leachate,
and metal salts. ACS Nano 5, 7214–7225 (2011).
93. Nair, S. et al. Role of size scale of ZnO nanoparticles and microparticles on
toxicity toward bacteria and osteoblast cancer cells. J. Mater. Sci. Mater. Med.
20, 235–241 (2009).
94. Choi, O. & Hu, Z. Size dependent and reactive oxygen species related nanosilver
toxicity to nitrifying bacteria. Environ. Sci. Technol. 42, 4583–4588 (2008).
95. Pang, H., Gao, F. & Lu, Q. Morphology effect on antibacterial activity of cuprous
oxide. Chem. Commun. 1076–1078 (2009).
96. Pal, S., Tak, Y. K. & Song, J. M. Does the antibacterial activity of silver
nanoparticles depend on the shape of the nanoparticle? A study of the gram-
negative bacterium Escherichia coli. J. Biol. Chem. 290, 1712–1720 (2015).
97. Donlan, R. M. Preventing biofilms of clinically relevant organisms using
bacteriophage. Trends Microbiol. 17, 66–72 (2009).
98. Xiong, Y. & Liu, Y. Biological control of microbial attachment: a promising
alternative for mitigating membrane biofouling. Appl. Microbiol. Biotechnol. 86,
825–837 (2010).
99. Lu, T. K. & Collins, J. J. Dispersing biofilms with engineered enzymatic
bacteriophage. Proc. Natl. Acad. Sci. 104, 11197–11202 (2007).
100. Goldman, G., Starosvetsky, J. & Armon, R. Inhibition of biofilm formation on
UF membrane by use of specific bacteriophages. J. Memb. Sci. 342, 145–152
(2009).
101. Tait, K., Skillman, L. C. & Sutherland, I. W. The efficacy of bacteriophage as a

Chapter 2
56
method of biofilm eradication. Biofouling 18, 305–311 (2002).
102. Fu, W. et al. Bacteriophage cocktail for the prevention of biofilm formation by
Pseudomonas aeruginosa on catheters in an in vitro model system. Antimicrob.
Agents Chemother. 54, 397–404 (2010).
103. Bjarnsholt, T. et al. Antibiofilm Properties of Acetic Acid. Adv. Wound Care 4,
363–372 (2015).
104. Halstead, F. D. et al. The antibacterial activity of acetic acid against biofilm-
producing pathogens of relevance to burns patients. PLoS One 10, 1–15 (2015).
105. Vadivelu, V. M., Keller, J. & Yuan, Z. Effect of free ammonia and free nitrous
acid concentration on the anabolic and catabolic processes of an enriched
Nitrosomonas Culture. Biotechnol. Bioeng. 95, 830–839 (2006).
106. Gao, S.-H., Fan, L., Yuan, Z. & Bond, P. L. The concentration-determined and
population-specific antimicrobial effects of free nitrous acid on Pseudomonas
aeruginosa PAO1. Appl. Microbiol. Biotechnol. 99, 2305–2312 (2015).
107. Gao, S.-H. et al. Antimicrobial effects of free nitrous acid on Desulfovibrio
vulgaris : implications for sulfide induced concrete corrosion. Appl. Environ.
Microbiol. AEM.01655–16 (2016). doi:10.1128/AEM.01655-16
108. Zhang, T., Wang, Q., Khan, J. & Yuan, Z. Free nitrous acid breaks down
extracellular polymeric substances in waste activated sludge. RSC Adv. 5, 43312–
43318 (2015).
109. Jiang, G., Gutierrez, O. & Yuan, Z. The strong biocidal effect of free nitrous acid
on anaerobic sewer biofilms. Water Res. 45, 3735–3743 (2011).
110. Filloux, E., Wang, J., Pidou, M., Gernjak, W. & Yuan, Z. Biofouling and scaling
control of reverse osmosis membrane using one-step cleaning-potential of
acidified nitrite solution as an agent. J. Memb. Sci. 495, 276–283 (2015).
111. Gao, S.-H. et al. Determining Multiple Responses of Pseudomonas aeruginosa
PAO1 to an Antimicrobial Agent, Free Nitrous Acid. Environ. Sci. Technol. 50,
2305–5312 (2016).
112. Englander, L. & Friedman, A. Nitric oxide nanoparticle technology: a novel
antimicrobial agent in the context of current treatment of skin and soft tissue

Chapter 2
57
infection. J. Clin. Aesthet. Dermatol. 3, 45–50 (2010).
113. Arora, D. P., Hossain, S., Xu, Y. & Boon, E. M. Nitric Oxide Regulation of
Bacterial Biofilms. Biochemistry 54, 3717–3728 (2015).
114. Stern, A. M. & Zhu, J. An introduction to nitric oxide sensing and response in
bacteria. Adv. Appl. Microbiol. 87, 187–220 (2014).
115. Schairer, D. O., Chouake, J. S., Nosanchuk, J. D. & Friedman, A. J. The potential
of nitric oxide releasing therapies as antimicrobial agents. Virulence 3, 271–279
(2012).
116. Privett, B. J., Broadnax, A. D., Bauman, S. J., Riccio, D. A. & Schoenfisch, M.
H. Examination of bacterial resistance to exogenous nitric oxide. Nitric Oxide 26,
169–173 (2012).
117. Loiselle, M. & Anderson, K. W. The use of cellulase in inhibiting biofilm
formation from organisms commonly found on medical implants. Biofouling 19,
77–85 (2003).
118. Taraszkiewicz, A., Fila, G., Grinholc, M. & Nakonieczna, J. Innovative
Strategies to Overcome Biofilm Resistance. Biomed Res. Int. 2013, 150653
(2013).
119. Kalia, V. C. Quorum sensing inhibitors: An overview. Biotechnol. Adv. 31, 224–
245 (2013).
120. Glinel, K., Thebault, P., Humblot, V., Pradier, C. M. & Jouenne, T. Antibacterial
surfaces developed from bio-inspired approaches. Acta Biomater. 8, 1670–1684
(2012).
121. Chen, Y. X., Ye, F. X. & Feng, X. S. The use of 3,3’,4',5-
tetrachlorosalicylanilide as a chemical uncoupler to reduce activated sludge yield.
J. Chem. Technol. Biotechnol. 79, 111–116 (2004).
122. Jiang, B. & Liu, Y. Energy uncoupling inhibits aerobic granulation. Appl.
Microbiol. Biotechnol. 85, 589–595 (2010).
123. Jiang, B. & Liu, Y. Roles of ATP-dependent N-acylhomoserine lactones (AHLs)
and extracellular polymeric substances (EPSs) in aerobic granulation.
Chemosphere 88, 1058–1064 (2012).

Chapter 2
58
124. Xu, H. & Liu, Y. Control and cleaning of membrane biofouling by energy
uncoupling and cellular communication. Environ. Sci. Technol. 45, 595–601
(2011).
125. Njoroge, J. & Sperandio, V. Jamming bacterial communication: New approaches
for the treatment of infectious diseases. EMBO Mol. Med. 1, 201–210 (2009).
126. Nijland, R., Hall, M. J. & Burgess, J. G. Dispersal of Biofilms by Secreted ,
Matrix Degrading , Bacterial DNase. 5, 1–7 (2010).
127. Schmidt, I., Steenbakkers, P. J. M., op den Camp, H. J. M., Schmidt, K. & Jetten,
M. S. M. Physiologic and Proteomic Evidence for a Role of Nitric Oxide in
Biofilm Formation by Nitrosomonas europaea and Other Ammonia Oxidizers. J.
Bacteriol. 186, 2781–2788 (2004).
128. Barraud, N. et al. Involvement of nitric oxide in biofilm dispersal of
Pseudomonas aeruginosa. J. Bacteriol. 188, 7344–7353 (2006).
129. Barraud, N. et al. Nitric oxide-mediated dispersal in single- and multi-species
biofilms of clinically and industrially relevant microorganisms. Microb.
Biotechnol. 2, 370–378 (2009).
130. Craven, M. et al. Nitric Oxide-Releasing Polyacrylonitrile Disperses Biofilms
Formed by Wound-Relevant Pathogenic Bacteria. J. Appl. Microbiol. 120, 1085–
1099 (2016).
131. Allan, R. N. et al. Low concentrations of nitric oxide modulate Streptococcus
pneumoniae biofilm metabolism and antibiotic tolerance. Antimicrob. Agents
Chemother. 60, 2456–2466 (2016).
132. Malaeb, L., Le-Clech, P., Vrouwenvelder, J. S., Ayoub, G. M. & Saikaly, P. E.
Do biological-based strategies hold promise to biofouling control in MBRs?
Water Res. 47, 5447–63 (2013).
133. Taylor, P. K., Yeung, A. T. Y. & Hancock, R. E. W. Antibiotic resistance in
Pseudomonas aeruginosa biofilms: Towards the development of novel anti-
biofilm therapies. J. Biotechnol. 191, 121–130 (2014).
134. Jiang, G. & Yuan, Z. Synergistic inactivation of anaerobic wastewater biofilm by
free nitrous acid and hydrogen peroxide. J. Hazard. Mater. 250-251, 91–98

Chapter 2
59
(2013).
135. Zhang, Y. & Hu, Z. Combined treatment of Pseudomonas aeruginosa biofilms
with bacteriophages and chlorine. Biotechnol. Bioeng. 110, 286–95 (2013).
136. Orman, M. A. & Brynildsen, M. P. Persister formation in Escherichia coli can be
inhibited by treatment with nitric oxide. Free Radic. Biol. Med. 93, 145–154
(2016).
137. Lundberg, J. O., Weitzberg, E. & Gladwin, M. T. The nitrate-nitrite-nitric oxide
pathway in physiology and therapeutics. Nat. Rev. Drug Discov. 7, 156–167
(2008).
138. Carpenter, A. W. & Schoenfisch, M. H. Nitric oxide release: Part II. Therapeutic
applications. Chem. Soc. Rev. 41, 3742 (2012).
139. Hetrick, E., Shin, J. & Stasko, N. Bactericidal efficacy of nitric oxide-releasing
silica nanoparticles. ACS Nano 2, 235–246 (2008).
140. Wo, Y., Brisbois, E. J., Bartlett, R. H. & Meyerhoff, M. E. Recent advances in
thromboresistant and antimicrobial polymers for biomedical applications: just say
yes to nitric oxide (NO). Biomater. Sci. 4, 1161–1183 (2016).
141. Wheatley, P. S. et al. NO-Releasing Zeolites and Their Antithrombotic
Properties. J. Am. Chem. Soc. 128, 502–509 (2005).
142. Neidrauer, M. et al. Antimicrobial efficacy and wound-healing property of a
topical ointment containing nitric-oxide-loaded zeolites. J. Med. Microbiol. 63,
203–209 (2014).
143. Bloch, E. D. et al. Gradual Release of Strongly Bound Nitric Oxide from
Fe2(NO)2(dobdc). J. Am. Chem. Soc. 137, 3466–3469 (2015).
144. Huang, S. L. et al. Nitric Oxide-Loaded Echogenic Liposomes for Nitric Oxide
Delivery and Inhibition of Intimal Hyperplasia. J. Am. Coll. Cardiol. 54, 652–
659 (2009).
145. Yang, Y. et al. Development of nitric oxide catalytic coatings by conjugating 3,3-
disulfodipropionic acid and 3,3-diselenodipropionic acid for improving
hemocompatibility. Biointerphases 10, 04A303 (2015).
146. Riccio, D. A. & Schoenfisch, M. H. Nitric oxide release: part I. Macromolecular

Chapter 2
60
scaffolds. Chem. Soc. Rev. 41, 3731–41 (2012).
147. Liang, H., Nacharaju, P., Friedman, A. & Friedman, J. M. Nitric oxide generating
/ releasing materials. Futur. Sci OA 1, 3065–3080. (2015).
148. Kim, J., Saravanakumar, G., Choi, H. W., Park, D. & Kim, W. J. A platform for
nitric oxide delivery. J. Mater. Chem. B 2, 341 (2014).
149. Barraud, N. et al. Cephalosporin-3′-diazeniumdiolates: Targeted NO-Donor
Prodrugs for Dispersing Bacterial Biofilms. Angew. Chemie 124, 9191–9194
(2012).
150. Nguyen, T.-K. et al. Co-delivery of nitric oxide and antibiotic using polymeric
nanoparticles. Chem. Sci. 7, 1016–1027 (2016).
151. Yang, Y., Qi, P., Yang, Z. & Huang, N. Nitric oxide based strategies for
applications of biomedical devices. Biosurface and Biotribology 1, 177–201
(2015).
152. Slomberg, D. L. et al. Role of size and shape on biofilm eradication for nitric
oxide-releasing silica nanoparticles. ACS Appl. Mater. Interfaces 5, 9322–9329
(2013).
153. Shin, J. H., Metzger, S. K. & Schoenfisch, M. H. Synthesis of nitric oxide-
releasing silica nanoparticles. J. Am. Chem. Soc. 129, 4612–9 (2007).
154. Shin, J. H. & Schoenfisch, M. H. Inorganic/organic hybrid silica nanoparticles as
a nitric oxide delivery scaffold. Chem. Mater. 20, 239–249 (2008).
155. Duong, H. T. T. et al. Functional gold nanoparticles for the storage and
controlled release of nitric oxide: applications in biofilm dispersal and
intracellular delivery. J. Mater. Chem. B 2, 5003 (2014).
156. Quinn, J. F., Whittaker, M. R. & Davis, T. P. Delivering nitric oxide with
nanoparticles. J. Control. Release 205, 190–205 (2015).
157. Lautner, G., Meyerhoff, M. E. & Schwendeman, S. P. Biodegradable poly(lactic-
co-glycolic acid) microspheres loaded with S-nitroso-N-acetyl-D-penicillamine
for controlled nitric oxide delivery. J. Control. Release 225, 133–139 (2016).
158. Brisbois, E. J., Handa, H., Major, T. C., Bartlett, R. H. & Meyerhoff, M. E.
Long-term nitric oxide release and elevated temperature stability with S-nitroso-

Chapter 2
61
N-acetylpenicillamine (SNAP)-doped Elast-eon E2As polymer. Biomaterials 34,
6957–6966 (2013).
159. Wo, Y. et al. Origin of Long-Term Storage Stability and Nitric Oxide Release
Behavior of CarboSil Polymer Doped with S-Nitroso-N-acetyl-D-penicillamine.
ACS Appl. Mater. Interfaces 7, 22218–22227 (2015).
160. Mann, M. N. et al. Plasma-modified nitric oxide-releasing polymer films exhibit
time-delayed 8-log reduction in growth of bacteria. Biointerphases 11, 031005
(2016).
161. Tai, L. A., Wang, Y. C. & Yang, C. S. Heat-activated sustaining nitric oxide
release from zwitterionic diazeniumdiolate loaded in thermo-sensitive liposomes.
Nitric Oxide 23, 60–64 (2010).
162. Duong, H. T. T. et al. Nanoparticle (star polymer) delivery of nitric oxide
effectively negates Pseudomonas aeruginosa biofilm formation.
Biomacromolecules 15, 2583–9 (2014).
163. Gierke, G. E., Nielsen, M. & Frost, M. C. S-Nitroso-N-acetyl-D-penicillamine
covalently linked to polydimethylsiloxane (SNAP–PDMS) for use as a controlled
photoinitiated nitric oxide release polymer. Sci. Technol. Adv. Mater. 12, 055007
(2011).
164. Seabra, A. B., Justo, G. Z. & Haddad, P. S. State of the art, challenges and
perspectives in the design of nitric oxide-releasing polymeric nanomaterials for
biomedical applications. Biotechnol. Adv. 33, 1370–1379 (2015).
165. Seabra, A. B., Marcato, P. D., de Paula, L. B. & Durán, N. New Strategy for
Controlled Release of Nitric Oxide. J. Nano Res. 20, 61–67 (2012).
166. Worley, B. V., Slomberg, D. L. & Schoenfisch, M. H. Nitric oxide-releasing
quaternary ammonium-modified poly(amidoamine) dendrimers as dual action
antibacterial agents. Bioconjug. Chem. 25, 918–927 (2014).
167. Jen, M. C., Serrano, M. C., van Lith, R. & Ameer, G. a. Polymer-Based Nitric
Oxide Therapies: Recent Insights for Biomedical Applications. Adv. Funct.
Mater. 22, 239–260 (2012).
168. Duong, H. T. T. et al. Intracellular nitric oxide delivery from stable NO-

Chapter 2
62
polymeric nanoparticle carriers. Chem. Commun. (Camb). 49, 4190–4192 (2013).
169. Luo, J. et al. The application of nitric oxide to control biofouling of membrane
bioreactors. Microb. Biotechnol. 8, 549–560 (2015).

63
Chapter 3.
Nitric oxide generation from nitrite
and ascorbic acid solution in the
presence of copper complex
catalyst
(This chapter is based on the work published in ACS Applied Materials and Interfaces
volume 7, issue 40, page 22148-22156, 2015.)
3.1. Introduction
Materials that are capable of delivering nitric oxide (NO) can be divided into
two types: NO-releasing materials (as reviewed in section 2.4) and NO-generating
materials.1–3 NO-generating materials appear more advantageous than NO-releasing
materials due to their ability to generate NO from endogenous sources, such as S-
nitrosothiols (RSNOs) species which are present in plasma and blood, or from nitrite
ions which are present in water.4,5 Such materials are usually copper or selenium(II)
(organoselenium) based, which can catalyse the decomposition of endogenous NO
donor to form NO.5,6 Other transition metals, such as iron(II), cobalt(II), nickel(II), and

Chapter 3
64
zinc(II), were also found to mediate NO release from S-nitroso-N-acetyl-D-
penicillamine (SNAP; a synthetic RSNOs), although at a significantly lower level
compared to copper.7 Out of all the metals reported, copper is especially attractive due
to its antibacterial activity, thus enhancing its potential as antibiofilm surface.8
The ability of copper to catalyse the reduction reaction of nitrite to NO is known
to occur in bacterial system by nitrite reductase enzyme, in specific blue copper nitrite
reductase.9 This enzyme consists of ligated copper(II) ion centre that can be reduced to
the active copper(I) ion centre, which then provides the electron for nitrite reduction.10,11
Incorporation of redox active copper species could happen by different strategies. For
instance, metallic copper nanoparticles doped inside a polymeric film can undergo slow
corrosion, thus releasing soluble oxides and other copper salts that provide a low level
of active copper(II)/copper(I) ions. These copper ions then interact with endogenous NO
sources in blood to release NO.12–14 Copper nanoparticles incorporated in
collagen/catechol thin films were shown to generate enough NO for inhibiting platelet
activation, thus showing potential application for vascular devices.15 Copper-based
metal organic framework deposited on a flexible polymer also have been used as NO-
generating materials.16 Boes et al. reported copper-containing zeolites that were used to
simultaneously store gaseous nitric oxide and catalytically generate NO from nitrite
(simultaneous NO-storing and generating material).17,18 However, utilisation of zeolites
and metal organic frameworks as NO-releasing/generating materials is not preferred.
This is mainly due to potential toxicity issue of the materials which would be a concern
for the medical application intended for their work.19 The most commonly used strategy
is by employing copper(II) complexes within a polymeric matrix that can be reduced to
copper(I) species in the presence of reducing agents, such as thiols and ascorbate, and
generate NO at the polymeric interface. Oh and Meyerhoff tested the activity of four

Chapter 3
65
different copper complexes in generating NO from sodium nitrite and ascorbate
solution.10 Out of the four complexes tested, Cu-cyclen complex and CuDTTCT
(copper(II)-dibenzo[e,k]-2,3,8,9-tetraphenyl-1,4,7,10-tetraaza-cyclododeca-1,3,7,9-
tetraene) complex exhibited the highest activities, which were found to be comparable
to the catalytic activity displayed by free copper(II) ions. Thus, covalent attachment
between Cu-cyclen within medical grade polyurethane as catheter coatings was
performed.1,20,21 Incorporation of CuDTTCT, which is more preferred over Cu-cyclen
due to its lower solubility in water, into polycarbourethane and poly(vinyl chloride)
(PVC) has been shown to generate NO in the presence of sodium nitrite-ascorbate
solution.10,22 Alternately, cathodic potentials have been used as an alternative means to
reduce copper(II)-tri(2-pyridylmethyl)amine (Cu-TPMA) inside a silicone rubber
catheters to generate NO from a nitrite reservoir.23,24
Herein, the ability of NO-generating material in suppressing biofilm formation
and removing pre-formed biofilm, either on the active material or the nearby surfaces
(the wells) where biofilm was grown, was investigated. CuDTTCT (Scheme 3.1) was
selected as the active catalytic site due to its compatibility with PVC, which is
commonly used as pipeline material for industrial water distribution. Nitrifying bacteria,
an environmentally and industrially relevant chemoautotroph microorganism frequently
found in water distribution systems, are chosen as a model organism.

Chapter 3
66
Scheme 3.1 CuDTTCT structure.
3.2. Experimental methods
3.2.1. Synthesis of copper dibenzo[e,k]-2,3,8,9-tetraphenyl-1,4,7,10-tetraaza-
cyclododeca-1,3,7,9-tetraene complex (CuDTTCT complex) and its
characterisation
Dibenzo[e,k]-2,3,8,9-tetraphenyl-1,4,7,10-tetraaza-cyclododeca-1,3,7,9-tetraene
(DTTCT) was synthesised following the method described by Oh and Meyerhoff.10 In
brief, 0.05 mol of benzil (Aldrich, 98%) and 0.05 mol of o-phenylenediamine (Aldrich,
99.5%) were dissolved in ethanol with a few drops of hydrochloric acid (Ajax
Finechem, 32%). The solution was refluxed at 80 ºC for 6 hours and cooled overnight.
On cooling overnight, light brown crystals formed. The precipitated DTTCT crystals
were filtered and washed with ethanol (resulting in a colour change to white crystalline
product) and dried in a vacuum desiccator. The compound was analysed by 1H-NMR
(Bruker Advance III 300MHz). For NMR analysis, 30 mg of dried DTTCT was
dissolved in 1 mL of deuterated DMSO.

Chapter 3
67
The CuDTTCT complex was synthesised by refluxing 0.01 mol of DTTCT and
0.05 mol of copper acetate monohydrate (Ajax APS) in ethanol at 80 ºC for 2 hours.
After cooling to room temperature overnight, washing with cold ethanol and drying in a
vacuum desiccator, a light blue precipitate was obtained, indicating the incorporation of
copper into the DTTCT structure. For NMR analysis, approximately 10 mg of
CuDTTCT was dissolved in 1 mL of deuterated DMSO.
To synthesise CuDTTCT film, CuDTTCT powder (2 mg) was dissolved in a
pre-made PVC solution in tetrahydrofuran (0.3 mL, 66 mg/mL) and sonicated in an
ultrasonic bath. The resulting solution was cast onto round glass cover slips (18 mm
diameter, ProSciTech). Before use, the glass cover slips were washed with dilute nitric
acid, acetone, and ethanol followed by drying overnight at 110 ºC. The films were dried
at 50 ºC for 12 hours and the resulting film was removed from the glass cover slips. The
amount of copper loading per film was analysed using inductive coupled plasma optical
emission spectrometry (ICP-OES, Perkin Elmer Optima 7300). Before analysis,
CuDTTCT film was weighed and then digested using concentrated nitric acid in a
commercially available microwave digestion bomb. From ICP-OES, the amount of
copper loading per film was found to be 2.88 wt%.
The immobilisation of copper complex in PVC matrix was analysed with a field-
emission scanning electron microscopy (SEM; FEI Nova NanoSEM 230) equipped with
an energy dispersive X-ray detector (EDS; Bruker Silicon Drift Energy Dispersive X-
ray detector). Prior to imaging, the samples were attached to 10 mm metal mounts using
carbon tape and sputter-coated with carbon. Data analysis was performed with Esprit
EDS software.

Chapter 3
68
X-ray photoelectron spectroscopy (XPS; ESCALAB250Xi, Thermo Scientific)
was used to determine the oxidation state of copper. Prior to XPS analysis, the
CuDTTCT films were cleaned by brief immersion in toluene to remove any
contaminants on the surface. Reduction was performed by immersing the pre-cleaned
sample in a 5 mM ascorbic acid (Sigma-Aldrich, ≥99%) solution for 5 minutes. All
samples were dried in a vacuum desiccator for two days before analysis.
3.2.2. NO generation measurement
The amount of NO that can be generated from CuDTTCT via nitrite reduction
was analysed amperometrically using Apollo TBR4100 Free Radical Analyser (World
Precision Instrument) equipped with an ISO-NOP 2 mm probe. The system was
calibrated using S-nitroso-N-acetylpenicillamine (SNAP; Sigma) and copper sulphate
solution according to the manufacturer’s protocol. Measurements were performed in
sterile nitrifying bacteria growth medium (ATCC medium 2265, which consists of three
different stock solutions – stock 1 (final composition in the medium mixture): 25 mM
(NH4)2SO4, 3 mM KH2PO4, 0.7 mM MgSO4, 0.2 mM CaCl2, 0.01 mM FeSO4, 0.02 mM
EDTA, 0.5 µM CuSO4; stock 2: 40 mM KH2PO4, 4 mM NaH2PO4, adjusted to pH 8 by
10 M NaOH; stock 3: 4 mM Na2CO3). CuDTTCT films or bare films (without any
CuDTTCT) as control samples were placed in 20 mL glass vials. Each glass vial was
then filled with 10 mL of media and stirred. After stable baseline was observed, sodium
nitrite (Ajax Finechem) and ascorbic acid were added into the solution to a final
concentration of 5 mM each.

Chapter 3
69
3.2.3. Ascorbic acid oxidation measurement
The change in ascorbic acid concentration over time was investigated via
absorbance measurement using via a colorimetric reagent (1,10-phenanthroline-iron(III)
solution) as described by Besada.25 A 10 mL aliquot of nitrifying bacteria growth
medium was added to a 20 mL glass vial containing CuDTTCT film or bare film.
Sodium nitrite and ascorbic acid at a final concentration of 5 mM were then added into
the vial. A 50 μL aliquot of samples were taken every hour and mixed with 1 mL of
colorimetric reagent and aged for 1 min. The resulting mixture was diluted with 4 mL of
water and the absorbance was recorded using UV-Vis spectrophotometer (Varian Cary
300) at 510 nm.
3.2.4. Biofilm suppression assay
Biofilm assays were performed in sterile 12-well plates (Corning). A mixed
inoculum commercially available nitrifying bacteria for aquarium water purification,
comprising Nitrosospira multiformis, Nitrospira marina and Bacillus sp. (Aquasonic
Bio-Culture), was used as a test strain. The mixed inoculum (1 mL) was added into 100
mL of nitrifying bacteria growth medium (ATCC medium 2265) and incubated for 3
days in the dark (30 ºC, 100 rpm). Three-day-old cultures were inoculated into fresh
medium and 2 mL aliquots (OD = 0.008) were added to each well of sterile 12-well
plates. Sodium nitrite and ascorbic acid in varying concentrations (ranging from 0.1 mM
to 10 mM) were added to each well. For the experiment where CuDTTCT film was
used, the CuDTTCT film was added into each well, followed by the addition of the
sodium nitrite and ascorbic acid solution. The plates were then incubated for 3 days in
the dark (30 ºC, 100 rpm).

Chapter 3
70
The amount of biomass (both planktonic and biofilm cells) was determined
using protein analysis via bicinchoninic acid method (BCA assay; Sigma). To quantify
the growth of planktonic cells, the medium was removed and centrifuged (12,000 rpm,
15 min) to recover the cells. The supernatants were removed and filtered through a 0.2
µm membrane to analyse the extent of copper leaching by ICP-OES. Two washing
stages with phosphate-buffered saline (PBS; Oxoid) were used to remove traces of
ascorbic acid from the planktonic cells. For biofilm measurements, the wells were also
washed twice with PBS to remove traces of ascorbic acid and loosely attached cells.
The BCA working reagent (2 mL) was added into each of the wells that contained
biofilm and to the centrifuge tubes that contained planktonic bacteria. All wells and
tubes were incubated for 30 minutes (37 ºC, 100 rpm) and measurements of the optical
density were performed at 562 nm (Infinite M200 Pro, Tecan). The presence of
CuDTTCT film did not interfere with the assay, as the use of CuDTTCT film in the
absence of bacteria did not result in colour change of the BCA working reagent, even
after the incubation step. Standards solution using Bovine Serum Albumin as per
manufacturer’s instruction was also used for each experiment.
Confocal laser scanning microscopy (CLSM; Olympus FluoViewTM FV1000)
with LIVE/DEAD staining was performed to quantify the bacteria surface coverage and
determine the physiological state (live vs dead) of the adherent bacterial cells. The
culture aliquots (2 mL each) with OD = 0.008 were grown in 35 mm culture dishes with
a coverglass bottom (internal glass diameter 22 mm, ProSciTech) with or without the
presence of bare film, CuDTTCT film, sodium nitrite or ascorbic acid and incubated in
the dark for three days at 30 ºC and 100 rpm. At the end of incubation, the medium was
removed and the dishes were washed twice with PBS. Adhered cells were stained with
400 µL of staining solution containing 3.34 µM of SYTO-9 and 19.97 µM of propidium

Chapter 3
71
iodide in PBS (LIVE/DEAD BacLightTM Bacterial Viability Kits L-7007, Molecular
Probes Inc.) where green stains denote viable bacteria and red or yellow stains denote
non-viable bacteria. The samples were incubated at room temperature for a minimum of
15 minutes. Surface coverage analysis from 12 images across the glass bottom was
performed on live cells (green channel) using image analysis software (Fiji/ImageJ). All
statistical analysis was performed using one-way ANOVA followed by Dunnett post
hoc analysis on Prism (GraphPad).
3.2.5. Biofilm dispersal and metabolic activity assay on pre-formed biofilms
Three-day-old cultures were inoculated into fresh medium and 2 mL aliquots
were added to each well of sterile 12-well plates. A CuDTTCT film was added into each
well and incubated for 3 days in the dark (30 ºC, 100 rpm). One hour before the
incubation period ended, sodium nitrite and ascorbic acid solution were added into each
well, and incubation continued for 1 h. The amount of biomass was then measured using
BCA assay and CLSM analysis as described before.
Adenosine triphosphate (ATP) analysis (Bactiter Glo, Promega) for biomass
metabolic activity measurements was also performed. At the end of 1 h incubation after
the addition of sodium nitrite and ascorbic acid solution, the planktonic bacteria were
collected, centrifuged, washed twice with PBS and resuspended in 100 µL of PBS. For
biofilm analysis, nitrifying biofilm samples were washed twice with PBS and
resuspended in 2 mL of PBS. To detach the cells, the plate was sonicated for 20 minutes
in a sonicating bath. A 100 µL sample of the detached cells was mixed with 100 µL of
the Bactiter Glo working reagent and incubated at room temperature for 5 min. The
luminescence was measured using a microtiter plate luminometer (Wallac Victor2).

Chapter 3
72
3.3. Results and discussion
3.3.1. Characterisation of copper complex (CuDTTCT) powder and films
The obtained spectrum from 1H-NMR spectroscopy analysis of DTTCT ligand
(Figure 3.1) is in accordance with previously reported NMR data.22 After chelation of
the copper(II) ion into the DTTCT complex, the 1H-NMR analysis was performed again
and no structural change in the DTTCT complex framework was observed (result not
shown). SEM and EDS mapping images of CuDTTCT film showed that copper can be
detected uniformly throughout the sample, albeit uneven distribution (Figure 3.2).
However, visual observation of the film does reveal uniform distribution of the copper
complex in the PVC matrix, as the film sample colour change from white (PVC only) to
blue-greenish colour (with the copper complex) without any observable darker coloured
patches. These results subsequently confirmed the presence of copper complex in the
film sample.
Figure 3.1 1H-NMR spectrum of DTTCT in d-DMSO.
8.2 8.0 7.8 7.6 7.4
Chemical shift (ppm)

Chapter 3
73
Figure 3.2 (a) SEM images of CuDTTCT in PVC film and (b) its corresponding elemental
mapping for copper. (c) EDS spectra of CuDTTCT film.
Oxidation state change of copper complex before and after the addition of
reducing agent (ascorbic acid) was analysed by XPS (Figure 3.3). Initially, Cu(II) peak
was detected at 934.8 eV, with two Cu(II) shake up peaks detected at 941.0 and 944.0
eV. The reaction between CuDTTCT with ascorbic acid caused the reduction of
copper(II)-DTTCT to copper(I)-DTTCT, as evidenced by the emergence of Cu(I) peak
at 932.8 eV (Figure 3.3 bottom).
(a) (b)
(c)

Chapter 3
74
Figure 3.3 XPS spectra of CuDTTCT polymer film before (top) and after (bottom)
reduction in 5 mM ascorbic acid solution for 5 min
3.3.2. Nitric oxide generation measurements
In order to validate the role of each chemical species in NO generation from
CuDTTCT-nitrite-ascorbic acid mixture, nitrite (5 mM) was first added into the
CuDTTCT film system, followed by an equimolar amount of ascorbic acid. The
addition of nitrite led to a small increase in NO concentration (Figure 3.4a), while
subsequent addition of ascorbic acid resulted in an abrupt rise in the NO concentration
(up to 80 nM), in good agreement with those obtained by Oh and Meyerhoff.10 It was
found that NO is catalytically formed from nitrite via Cu(II)/Cu(I) redox cycling. In
detail, ascorbic acid reduces Cu(II) to Cu(I), which subsequently reacts with nitrite to
produce NO and, in the process, regenerates Cu(II).10
948 944 940 936 932 928
Co
un
ts (
s-1)
Binding energy (eV)
Cu+
Cu2+
Cu2+
Cu2+
Cu2+
Cu2+
Cu2+

Chapter 3
75
2 Cu2+(DTTCT) + ascorbic acid ⇌ 2 Cu+(DTTCT) + dehydroascorbic acid + 2 H+ (i)
2 Cu+(DTTCT) + 2 NO2– + 2 H2O ⇌ 2 NO + 4 OH– + 2 Cu2+(DTTCT) (ii)
The subsequently formed Cu(II) can be reduced again to Cu(I) following the
proposed Cu(II)/Cu(I) redox cycling mechanism in the presence of excess ascorbic
acid.10,26 Spectrophotometric measurements showed that ascorbic acid can still be
detected in the system even after 7 hours (Figure 3.4b), therefore prolonged generation
of NO was observed.
It has been reported that in acidic conditions, nitrite solutions can form nitrous
acid (HNO2, pKa of 3.2), which can easily decompose to form NO and other nitrogen
oxides.27 As the solution pH remained between 7.3 to 7.6 upon the addition of 5 mM
nitrite-ascorbic acid, it is believed that a major proportion of NO was generated through
the redox cycling pathway, rather than through the acidification of nitrite pathway.27,28
The possible formation of HNO2 in the system which may contribute to the overall NO
release cannot be ruled out, although the system was well-mixed during the addition of
ascorbic acid. A dip in the NO level was also observed in the period between 100 s and
250 s (Figure 3.4a), which can be attributed to the oxidation of the initially generated
NO to nitrite or the oxidation of copper(I) species that lowers the NO signal
momentarily.10 In the absence of CuDTTCT, a maximum of 20 nM NO was generated,
with no prolonged release observed in an equivalent experimental setup. This confirms
the significance of CuDTTCT as an NO generation catalyst.

Chapter 3
76
Figure 3.4 (a) Amperometric measurement of NO generation versus time for control (bare
film) and CuDTTCT film (2.88 wt% of copper) in 10 mL of bacteria medium. The red
arrow indicates the time at which 5 mM of nitrite was added and the blue arrow indicates
the time at which 5 mM ascorbic acid was added. (b) Change in ascorbic acid
concentration in 10 mL of nitrifying bacteria growth medium that contains CuDTTCT
film or bare film (control).
3.3.3. Nitrifying bacteria biofilm suppression by nitrite-ascorbic acid and
CuDTTCT-nitrite-ascorbic acid mixture
The effectiveness of nitrite and ascorbic acid solution on reducing biofilm
formation was studied first in the absence of CuDTTCT. Here, the presence of biomass
was determined by measuring the protein concentration, complemented by confocal
laser scanning microscopy (CLSM) of the LIVE/DEAD-stained biomass. The latter
analysis is to account for the potential detection of biofilm EPS in the protein assay
which would lead to an overestimation of the number of bacteria on the surface.29
In the absence of CuDTTCT, an addition of 5 mM nitrite-ascorbic acid inhibited
the growth of biofilm by 40% and of planktonic biomass by 60% relative to the cultures
that did not receive any treatment (control, Figure 3.5a). Approximately 0.32 mg/L of
leached copper can be detected from this system (Table 3.2). However, it is unlikely
that the 0.32 mg/L of copper leachate detected caused the observed reduction in biofilm
growth, as even 1.5 mg/L of copper ions did not lead to significant biofilm reduction
(a) (b)
0 1 2 3 4 5 6 72
3
4
5
6
Asc. a
cid
co
nce
ntr
atio
n (
mM
)
Time (hours)
control
with CuDTTCT
0 100 200 300 400 500 600 700
-40
-20
0
20
40
60
80
100
120
NO
co
nce
ntr
atio
n (
nM
)
Time (seconds)
control
with CuDTTCT

Chapter 3
77
(Figure 3.7), which will be discussed further later. Therefore, this suggests that the
inhibition effect observed was due to the nitrite-ascorbic acid mixture. Increasing the
amount of nitrite and ascorbic acid to 10 mM led to a severe suppression of the biofilm
formation and only 15% planktonic growth as compared to the untreated controls. The
trend of the biofilm growth suppression was also observed by using LIVE/DEAD
staining in CLSM analysis (Figure 3.5b). The biofilm surface coverage was reduced by
25% after treatment with 1 mM nitrite and ascorbic acid, which is in close agreement
with the ~20% reduction in biofilm biomass measured with the protein assay after the
same treatment. Enhanced reduction in biofilm surface coverage was observed when the
nitrite and ascorbic acid concentration was increased to 5 mM, with only ~30% bacteria
detected on the surface when compared to the control. Furthermore, it was shown that
the biomass remained viable following the nitrite-ascorbic acid treatment in all samples
(Figure 3.5d and e). It appears that nitrite and ascorbic acid in this case inhibit the cell
proliferation of nitrifying bacteria. The suppressed biofilm growth in this study appears
to result from the quick burst of NO generation as observed previously in nitrite-
ascorbic acid system (Figure 3.4).26,30 Such ‘antibiofilm’ activity of the nitrite-ascorbic
acid system is in agreement with the earlier reports, in particular on Gram-negative
pathogens,28,31,32 but no study had been done previously on environmentally-relevant
bacteria, such as nitrifying bacteria.

Chapter 3
78
Figure 3.5 (a) Protein measurements of nitrifying bacteria suppression in nitrite-ascorbic
acid mixture. All values shown are normalised to the protein concentration of control
(inset). Error bars indicate standard error between replicates (n = 3); *p ≤ 0.05 against the
control. (b) Surface coverage analysis of nitrifying biofilms. All values shown are
normalised to the surface coverage of the nitrifying bacteria biofilm in medium only
(control). Error bars indicate standard error between replicates (n = 2); *p ≤ 0.05 against
the control. Confocal laser scanning microscopy images of nitrifying bacteria biofilm
grown (c) in medium only (control) and in the presence of (d) 1 mM and (e) 5 mM of
nitrite-ascorbic acid. Scale bar = 100 µm.
Table 3.1 ICP-OES measurements of copper ions in the bacteria medium after three days
incubation in the presence of bare films
Nitrite and ascorbic acid concentration (mM) Copper ions (mg/L)
0 0.08
1 0.20
5 0.32
(b)
(a) (c)
(d)
(e)
Control
1 mM
5 mM
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
**
*
Rela
tive p
rote
in c
oncentr
ation
Nitrite-ascorbic acid concentration (mM)
Biofilm
Planktonic
Control 1 5 10
*
0
125
250
Pro
tein
co
nce
ntr
atio
n
(µg
/mL
)
Control
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Rela
tive s
urf
ace c
overa
ge
Nitrite-ascorbic acid concentration (mM)
Control 1 5
*

Chapter 3
79
Recalling the prolonged and 4-fold higher release of nitric oxide (Figure 3.4),
the ‘antibiofilm’ activity of the CuDTTCT-nitrite-ascorbic acid system is investigated
with protein assay-CLSM analyses in the presence of CuDTTCT films. Incorporation of
CuDTTCT into the PVC film alone reduced the planktonic biomass to 32% of the
control culture (Figure 3.6a). This reduction is thought to be the result of copper
toxicity, specifically the leached copper ions (1.5 mg/L, Table 3.2). In order to simulate
the effect of leached copper, copper sulphate at an equivalent concentration was used
(Figure 3.7). In the presence of 1.5 mg/L of copper ions, a 65% decrease of planktonic
bacteria compared to the control was observed with no significant effect towards the
biofilm, which is in accordance with earlier observation in a CuDTTCT-only system.
The toxicity of copper to planktonic bacteria is well documented and may involve
several mechanisms such as oxidative stress induced by Fenton reaction, inactivation of
Fe-S clusters, or lipid peroxidation.33,34

Chapter 3
80
Figure 3.6 (a) Protein measurements of nitrifying bacteria suppression in CuDTTCT,
nitrite and ascorbic acid mixture. All values shown are normalised to the protein
concentration of control (inset). Error bars indicate standard error between replicates (n =
4); *p ≤ 0.05 against the control. (b) Surface coverage analysis of nitrifying biofilms. All
values shown are normalised to the surface coverage of the nitrifying biofilm grown in the
presence of bare film (control). Error bars indicate standard error between replicates (n =
2); *p ≤ 0.05 against the control. Confocal laser scanning microscopy images of nitrifying
biofilm grown in the presence of (c) bare film (control), (d) CuDTTCT film, (e) CuDTTCT
film and 1 mM of nitrite-ascorbic acid, and (f) CuDTTCT film and 5 mM of nitrite-
ascorbic acid. Scale bar = 100 µm.
Table 3.2 ICP-OES measurements of copper ions in the bacteria medium after three days
incubation in the presence of CuDTTCT films.
Nitrite and ascorbic acid
concentration (mM)
Copper ions (mg/L) %mass of copper leached out
of the CuDTTCT films
0 1.48 0.44
1 2.61 0.77
5 3.29 0.94
(a)
(b)
(c)
(d)
(e)
(f)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
*
*
**
**
Rela
tive p
rote
in c
oncentr
ation
Nitrite-ascorbic acid concentration (mM)
Biofilm
Planktonic
Control 0 0.1 0.5 1 5
*
0
200
400
Pro
tein
concentr
ation
(µg/m
L)
Control
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Rela
tive s
urf
ace c
overa
ge
Nitrite-ascorbic acid concentration (mM)
Control 0 1 5
*
Control
0 mM
1 mM
5 mM

Chapter 3
81
Figure 3.7 Protein measurements of the nitrifying bacteria after the addition of free
copper ion (in the form of copper sulphate). All values shown are normalised to the
protein concentration of control (inset). Error bars indicate standard error between
replicates (n = 3); *p ≤ 0.05 against the control.
Addition of nitrite-ascorbic acid of up to 1 mM in the presence of CuDTTCT
further reduced the planktonic and biofilm biomass to ~20% and ~43% of the control
value, respectively. Such suppression of the biofilm formation was not observed at the
same nitrite-ascorbic acid concentration in the absence of CuDTTCT. Further increase
of nitrite-ascorbic acid concentration to 5 mM in a CuDTTCT system resulted in more
than 95% reduction of biofilm biomass compared to the control and complete inhibition
of planktonic growth. The addition of 5 mM nitrite-ascorbic acid into the CuDTTCT
system was associated with detection of 3.3 mg/L of leached copper ions (Table 3.2),
which account for less than 1% of copper from CuDTTCT, suggesting the excellent
stability of CuDTTCT inside the PVC matrix. However, this observed increase in
copper leachate could alter water quality and affect the environment. Future research
directed at ligand design that can increase the stability of copper centre and prevent
leaching would be of interest.
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
*
***
Rela
tive p
rote
in c
oncentr
ation
Copper concentration (mg/L)
Biofilm
Planktonic
Control 1.5 2.6 3.3 10
*
0
100
200
300
Pro
tein
co
nce
ntr
atio
n
(µg
/mL
)
Control

Chapter 3
82
The detected 3.3 mg/L of copper ions from the system did not lead to significant
reduction in nitrifying biofilm biomass, although 77% reduction in planktonic biomass
was observed (Figure 3.7). In fact, the presence of 10 mg/L of copper ions was not able
to fully eradicate the biofilm (only 50% reduction compared to control) as opposed to
drastic biofilm reduction observed in the 5 mM CuDTTCT-nitrite-ascorbic acid system.
Therefore, the system’s high inhibition of biofilm formation cannot be solely attributed
to the leached copper ions. Taken together, it can be concluded that biofilm suppression
is predominantly caused by the CuDTTCT-nitrite-ascorbic acid mixture, most likely due
to the activity of catalytically generated NO as a cell proliferation inhibitor. In
agreement with the biofilm inhibition trend that was determined by the protein assay,
CLSM analysis revealed 25% biofilm surface coverage with the CuDTTCT system in
the presence of 1 mM nitrite-ascorbic acid relative to the control (p = 0.058, Figure
3.6b). Further increase of nitrite-ascorbic acid concentration to 5 mM significantly
inhibited biofilm formation (surface coverage reduced to 2%).
Comparing the two different systems that have been investigated, a higher
amount of nitrite and ascorbic acid was needed to achieve comparable biomass
suppression in the absence of CuDTTCT. For instance, 5 mM nitrite and ascorbic acid
in the absence of CuDTTCT were required to reduce the biofilm surface coverage by
~70% relative to the control (determined by CLSM, Figure 3.5b), while lower
concentration of nitrite and ascorbic acid (1 mM) was sufficient to acquire a comparable
biomass reduction in the presence of CuDTTCT (Figure 3.6b). Similar to the nitrite-
ascorbic acid system, the biomass remained viable following the CuDTTCT-nitrite-
ascorbic acid treatment (Figure 3.6e and f), even at the highest concentration tested (5
mM). This implies that NO generated in the CuDTTCT-nitrite-ascorbic acid system acts
as a cell proliferation inhibitor to the nitrifying bacteria. NO can acts as an antimicrobial

Chapter 3
83
agent by several mechanisms, in particular, its reactivity towards transition metals. The
NO reactivity toward iron ions has been known to inhibit bacterial respiration,35 while
the inactivation of zinc metalloproteins by NO inhibits DNA replication.36 The synthesis
and repair of DNA are also inhibited by reactions between NO and tyrosyl radicals
which limits the concentration of the DNA precursor.36
In comparison to the nitrite-ascorbic acid system, it would be reasonable to
deduce that the enhanced bacteriostatic activity of the CuDTTCT-nitrite-ascorbic acid
system is at least in part due to the 4-fold higher nitric oxide release in the quick burst
(first 200 s) along with the subsequent prolonged release of NO. It was shown in earlier
studies that higher amount of NO that was released over short durations is more
damaging to Gram-negative bacteria than a prolonged release of lower amounts of
NO.37 Similar findings were also reported on human fibroblasts, where the reduction in
fibroblast numbers exhibited by the copper-nitrite-ascorbic acid system has higher
degree of correlation to the amount of nitric oxide generated in the quick burst (first 200
s) rather than the total amount of nitric oxide over 600 s.26
3.3.4. Nitrifying bacteria biofilm dispersal upon the addition of nitrite-ascorbic acid
in the presence of CuDTTCT
The effect of NO on a pre-formed biofilm was also investigated by adding nitrite
and ascorbic acid to a CuDTTCT system following 3 days of biofilm growth. The term
“pre-formed biofilm” is used to refer to biofilms that cannot be removed through simple
washing steps alone, although biofilm maturity may not have been reached yet. The
system was found to be effective at dispersing the biofilm even with as little as 0.1 mM
nitrite-ascorbic acid, which resulted in ~40% less biofilm compared to the CuDTTCT-

Chapter 3
84
only system (Figure 3.8a). The addition of nitrite-ascorbic acid of up to 1 mM was
effective at dispersing the biofilm by 40%; however, when the concentration was
increased to 5 mM, the dispersal effect was reduced, with only 22% of biofilm dispersal
detected (p-value = 0.089 against the CuDTTCT film with no nitrite and ascorbic acid
added). Confirming the trend of biofilm dispersal determined by the protein assay,
surface coverage analysis revealed that 1 mM nitrite-ascorbic acid addition can reduce
the biofilm surface coverage by ~50% as compared to the CuDTTCT-only system with
no viability loss observed (Figure 3.8b and e). Although an increase in leached copper
concentration was observed upon the addition of nitrite-ascorbic acid into the system
(Table 3.3), the observed dispersal effect was more attributed to NO generated by the
combination of CuDTTCT-nitrite and ascorbic acid system. The ability of NO (by
addition of a NO donor or NO gas) to cause biofilm dispersal has been reported on a
wide spectrum of microorganisms, including the Gram-positive Staphylococcus
epidermidis,38 Bacillus licheniformis,38 the Gram-negative Pseudomonas aeruginosa,39
Escherichia coli,38 Serratia marcescens,38 Vibrio cholerae,38 Nitrosomonas europaea,40
as well as the Candida albicans yeast.38 In the case of Nitrosomonas europaea, which is
one of the major members of nitrifying microorganisms in aquatic environments, the
switch between biofilm and planktonic states is induced by the regulation of the motility
expression and/or chemotaxis responses by NO, which is potentially the mechanism
adopted by the test organism in this study.40

Chapter 3
85
Figure 3.8 (a) Protein measurements of the nitrifying bacteria upon addition of nitrite and
ascorbic acid on pre-formed biofilm grown in the presence of CuDTTCT. All values
shown are normalised to the protein concentration of the control (inset). Error bars
indicate standard error between replicates (n = 4); *p ≤ 0.05 against the control (bare
film); #p ≤ 0.05 against the CuDTTCT film (0 mM). (b) Surface coverage analysis of
nitrifying biofilms. All values shown are normalised to the surface coverage of nitrifying
biofilm grown in the presence of bare film (control). Error bars indicate standard error
between replicates (n = 3); * p ≤ 0.05 against the control and CuDTTCT film (0 mM).
Confocal laser scanning microscopy images of nitrifying bacteria biofilm grown in the
presence of (c) bare film (control), (d) CuDTTCT film, (e) CuDTTCT film and 1 mM of
nitrite-ascorbic acid, and (f) CuDTTCT film and 5 mM of nitrite-ascorbic acid. Nitrite and
ascorbic acid were added on pre-formed biofilm grown in the presence of CuDTTCT 1
hour before incubation ended. Scale bar = 100 µm.
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
*
**
*
*
*
Re
lative
pro
tein
co
nce
ntr
atio
n
Nitrite-ascorbic acid concentration (mM)
Biofilm
Planktonic
Control 0 1 50.1 0.5
*#*#
*#
Control0
150
300
Pro
tein
co
nce
ntr
ation
(µg
/mL
)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Rela
tive s
urf
ace c
overa
ge
Nitrite-ascorbic acid concentration (mM)Control 0 1 5
**
(a)
(b)
(c)
(d)
(e)
(f)
Control
0 mM
1 mM
5 mM

Chapter 3
86
Table 3.3 ICP-OES measurements of copper ions in the bacteria medium after the end of
second incubation step in the presence CuDTTCT films.
Nitrite and ascorbic acid concentration (mM) Copper ions (mg/L)
0 1.90
1 3.34
5 4.54
From the surface coverage analysis, the addition of 5 mM nitrite and ascorbic
acid was also found to be not as effective as for the 1 mM concentration in reducing the
biofilm biomass (Figure 3.8b,e,f). The analysis, however, revealed the presence of dead
(red) cells after addition of 5 mM nitrite-ascorbic acid. In this case, it is likely that the
cell death occurred at a faster rate relative to the biomass dispersal, thereby reducing the
efficacy of biofilm dispersal. The observed toxicity of either the increase in copper
leachate concentration or the NO generated at 5 mM nitrite-ascorbic acid is consistent
with the ATP analysis, as described below.
The level of ATP detected in biofilms grown in the presence of CuDTTCT was
60% lower than the level of ATP detected in control biofilms (Figure 3.9). Similarly,
planktonic bacteria also had 60% less ATP after exposure to CuDTTCT. Interestingly,
although protein and ATP measurements on planktonic bacteria revealed similar trends
for control (bare film) and CuDTTCT film only, a different trend was observed for the
biofilm. While no significant reduction was observed for the biofilm biomass by protein
and surface coverage analysis between the control and CuDTTCT film, 60% lower ATP
was detected in biofilms grown in the presence of CuDTTCT film compared to control
biofilm. This indicates that the presence of CuDTTCT or/and the soluble copper ion
could induce stress in the biofilm bacteria which resulted in a lower amount of ATP
being produced rather than removal of the biofilm biomass. A small and insignificant
reduction in ATP was observed after the addition of 1 mM nitrite and ascorbic acid (p-
value = 0.062 for the biofilm and p-value = 0.074 for the planktonic against the

Chapter 3
87
CuDTTCT film). However, after the addition of 5 mM nitrite-ascorbic acid, ATP levels
were significantly lowered in both biofilm and planktonic bacteria. The decrease in ATP
values in this instance could be attributed to viability loss due to cellular damage from
NO exposure as well as the increase in leached copper concentration that is triggered by
the addition of nitrite and ascorbic acid. In fact, samples containing CuDTTCT and
treated with 5 mM nitrite and ascorbic acid exhibited dead bacteria that were stained
with propidium iodide, which is an indicator of cell membrane disruption. NO is
reported to cause membrane disruption via radical lipid peroxidation.41 The
deterioration of lipid bilayer of the cell membrane could cause the increase in cell
surface roughness and membrane degradation.37 Interestingly, cell death by membrane
disruption was not observed when CuDTTCT and 5 mM of nitrite and ascorbic acid
were added before biofilm was formed (Figure 3.6f). It appears that the mode of action
of this catalytic system may be dependent on the growth stages of the bacteria. When
bacteria are exposed to nitrite, ascorbic acid and copper complex from the beginning of
incubation, i.e. during the lag phase, bacterial cells may have access to energy resources
allowing them to resist potential membrane damage and cell death induced by NO,
while being unable to trigger cell division and proliferation mechanisms. However, in
pre-formed biofilm system, bacterial cells are in a stationary phase without access to
fresh nutrients and have low metabolic activity, thus the addition of nitrite and ascorbic
acid in the presence of copper complex led to cell death by membrane disruption.
Antimicrobial action dependent on the bacterial growth phase has been widely reported
before, including for antibiotics.34,42 For instance, the polypeptide colistin, which can
solubilise the cytoplasmic membrane, was found to be more effective against stationary-
phase, dormant bacteria with low metabolic activity as compared to cells with high
metabolic activity.43 However, it is also possible that the initial nitric oxide burst

Chapter 3
88
generated before the formation of biofilm would induce membrane disruption or
reduction of metabolic activity, but the three days incubation period allowed some
bacterial cells to regain their viability.
Figure 3.9 Luminescence measurements of the nitrifying bacteria upon the addition of
nitrite and ascorbic acid on pre-formed biofilm. All values shown are normalised to the
luminescence of the control. Error bars indicate standard error between replicates (n = 2);
*p ≤ 0.05 against the control (bare film); #p ≤ 0.05 against the CuDTTCT film.
3.4. Summary
A lipophilic copper(II) complex (CuDTTCT) as an NO generating catalyst has
been successfully incorporated into PVC films and tested for the control of nitrifying
bacteria biofilm formation in the presence of nitrite and ascorbic acid. Copper(II) in the
complex was reduced to copper(I) by ascorbic acid, which subsequently reduced nitrite
to NO while being oxidized back to copper(II). Amperometric measurements revealed
an initial surge of NO generated by the copper complex system upon the addition of
ascorbic acid followed by a continuous production of NO. The potential of catalytically
generated NO to control the formation of and disperse a nitrifying bacteria biofilm was
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
*
*
Re
lative
lu
min
esce
nce
Nitrite-ascorbic acid concentration (mM)
Biofilm
Planktonic
Control 0 1 5
*
*#*#

Chapter 3
89
demonstrated. Nitrifying bacteria biofilm biomass was suppressed in the presence of
CuDTTCT-nitrite-ascorbic acid which is predominantly caused by NO. Moreover, the
system is also effective at dispersing established biofilms. No significant dead cells
were observed in the biofilm suppression studies, which indicate that prior to biofilm
formation the NO generated by copper complex-nitrite-ascorbic acid mainly inhibits cell
proliferation. Biofilm dispersal studies showed that the copper complex system with 1
mM of nitrite and ascorbic acid produced sufficient NO to effectively disperse the
accumulated biofilm. At a higher concentration of nitrite and ascorbic acid (5 mM),
biofilm dispersal effect was reduced and significant dead cells were observed. In
summary, this study highlighted the use of copper complex immobilised in PVC and
nitrite-ascorbic acid to minimise formation and disperse nitrifying bacteria biofilms
through sustained generation of NO.
3.5. References
1. Hwang, S. & Meyerhoff, M. E. Polyurethane with tethered copper(II)-cyclen
complex: preparation, characterization and catalytic generation of nitric oxide
from S-nitrosothiols. Biomaterials 29, 2443–2452 (2008).
2. Yang, Y. et al. Development of nitric oxide catalytic coatings by conjugating 3,3-
disulfodipropionic acid and 3,3-diselenodipropionic acid for improving
hemocompatibility. Biointerphases 10, 04A303 (2015).
3. Wu, Y. & Meyerhoff, M. E. Nitric oxide-releasing/generating polymers for the
development of implantable chemical sensors with enhanced biocompatibility.
Talanta 75, 642–650 (2008).
4. Barraud, N., Kelso, M. J., Rice, S. A. & Kjelleberg, S. Nitric Oxide : A Key
Mediator of Biofilm Dispersal with Applications in Infectious Diseases. Curr.

Chapter 3
90
Pharm. Des. 21, 31–42 (2015).
5. Yang, Y., Qi, P., Yang, Z. & Huang, N. Nitric oxide based strategies for
applications of biomedical devices. Biosurface and Biotribology 1, 177–201
(2015).
6. Wo, Y., Brisbois, E. J., Bartlett, R. H. & Meyerhoff, M. E. Recent advances in
thromboresistant and antimicrobial polymers for biomedical applications: just say
yes to nitric oxide (NO). Biomater. Sci. 4, 1161–1183 (2016).
7. McCarthy, C. W., Guillory II, R. J., Goldman, J. & Frost, M. C. Transition Metal
Mediated Release of Nitric Oxide (NO) from S-Nitroso-N-acetylpenicillamine
(SNAP): Potential Applications for Endogenous Release of NO on the Surface of
Stents via Corrosion Products. ACS Appl. Mater. Interfaces 8, 10128–10135
(2016).
8. Gunawan, C., Teoh, W., Marquis, C. & Amal, R. Cytotoxic origin of copper (II)
oxide nanoparticles: comparative studies with micron-sized particles, leachate,
and metal salts. ACS Nano 5, 7214–7225 (2011).
9. Cutruzzolà, F. Bacterial nitric oxide synthesis. Biochim. Biophys. Acta -
Bioenerg. 1411, 231–249 (1999).
10. Oh, B. K. & Meyerhoff, M. E. Catalytic generation of nitric oxide from nitrite at
the interface of polymeric films doped with lipophilic Cu(II)-complex: a potential
route to the preparation of thromboresistant coatings. Biomaterials 25, 283–293
(2004).
11. Opländer, C. et al. Redox-mediated mechanisms and biological responses of
copper-catalyzed reduction of the nitrite ion in vitro. Nitric Oxide 35, 152–164
(2013).
12. Major, T. C. et al. The hemocompatibility of a nitric oxide generating polymer
that catalyzes S-nitrosothiol decomposition in an extracorporeal circulation
model. Biomaterials 32, 5957–5969 (2011).
13. Wu, Y. et al. Improving blood compatibility of intravascular oxygen sensors via

Chapter 3
91
catalytic decomposition of S-nitrosothiols to generate nitric oxide in situ. Sensors
Actuators, B Chem. 121, 36–46 (2007).
14. Amoako, K. A. & Cook, K. E. Nitric oxide-generating silicone as a blood-
contacting biomaterial. ASAIO J. 57, 539–544 (2011).
15. Luo, R. et al. Copper-Incorporated Collagen/Catechol Film for in Situ Generation
of Nitric Oxide. ACS Biomater. Sci. Eng. 1, 771–779 (2015).
16. Neufeld, M. J., Harding, J. L. & Reynolds, M. M. Immobilization of Metal-
Organic Framework Copper(II) Benzene-1,3,5-tricarboxylate (CuBTC) onto
Cotton Fabric as a Nitric Oxide Release Catalyst. ACS Appl. Mater. Interfaces 7,
26742–26750 (2015).
17. Boes, A.-K., Wheatley, P. S., Xiao, B., Megson, I. L. & Morris, R. E.
Simultaneous and cooperative gas storage and gas production using bifunctional
zeolites. Chem. Commun. (Camb). 5, 6146–6148 (2008).
18. Boës, A.-K., Xiao, B., Megson, I. L. & Morris, R. E. Simultaneous Gas Storage
and Catalytic Gas Production Using Zeolites—A New Concept for Extending
Lifetime Gas Delivery. Top. Catal. 52, 35–41 (2009).
19. Seabra, A. B. & Durán, N. Nitric oxide-releasing vehicles for biomedical
applications. J. Mater. Chem. 20, 1624–1637 (2010).
20. Liu, K. & Meyerhoff, M. E. Preparation and characterization of an improved
Cu(2+)-cyclen polyurethane material that catalyzes generation of nitric oxide
from S-nitrosothiols. J. Mater. Chem. 22, 18784–18787 (2012).
21. Puiu, S. C. et al. Metal ion-mediated nitric oxide generation from polyurethanes
via covalently linked copper(II)-cyclen moieties. J. Biomed. Mater. Res. - Part B
Appl. Biomater. 91, 203–212 (2009).
22. Zhao, H., Feng, Y. & Guo, J. Polycarbonateurethane films containing complex of
copper (II) catalyzed generation of nitric oxide. J. Appl. Polym. Sci. 122, 1712–
1721 (2011).
23. Ren, H. et al. Electrochemically modulated nitric oxide (NO) releasing

Chapter 3
92
biomedical devices via copper(II)-Tri(2-pyridylmethyl)amine mediated reduction
of nitrite. ACS Appl. Mater. Interfaces 6, 3779–3783 (2014).
24. Ren, H. et al. Thromboresistant/anti-biofilm catheters via electrochemically
modulated nitric oxide release. Bioelectrochemistry 104, 10–16 (2015).
25. Besada, A. A facile and sensitive spectrophotometric determination of ascorbic
acid. Talanta 34, 731–732 (1987).
26. Opländer, C. et al. Characterization of novel nitrite-based nitric oxide generating
delivery systems for topical dermal application. Nitric Oxide 28, 24–32 (2013).
27. Lundberg, J. O., Weitzberg, E. & Gladwin, M. T. The nitrate-nitrite-nitric oxide
pathway in physiology and therapeutics. Nat. Rev. Drug Discov. 7, 156–167
(2008).
28. Carlsson, S., Wiklund, N. P., Engstrand, L., Weitzberg, E. & Lundberg, J. O.
Effects of pH, nitrite, and ascorbic acid on nonenzymatic nitric oxide generation
and bacterial growth in urine. Nitric oxide Biol. Chem. 5, 580–586 (2001).
29. Douterelo, I. et al. Methodological approaches for studying the microbial ecology
of drinking water distribution systems. Water Res. 65, 134–156 (2014).
30. Schairer, D. O., Chouake, J. S., Nosanchuk, J. D. & Friedman, A. J. The potential
of nitric oxide releasing therapies as antimicrobial agents. Virulence 3, 271–279
(2012).
31. Kishikawa, H. et al. Control of pathogen growth and biofilm formation using a
urinary catheter that releases antimicrobial nitrogen oxides. Free Radic. Biol.
Med. 65, 1257–1264 (2013).
32. Firmani, M. & Riley, L. Reactive nitrogen intermediates have a bacteriostatic
effect on Mycobacterium tuberculosis in vitro. J. Clin. Microbiol. 40, 1–6 (2002).
33. Booth, S. C. et al. Differences in metabolism between the biofilm and planktonic
response to metal stress. J. Proteome Res. 10, 3190–3199 (2011).
34. Lemire, J. A., Harrison, J. J. & Turner, R. J. Antimicrobial activity of metals:

Chapter 3
93
mechanisms, molecular targets and applications. Nat. Rev. Microbiol. 11, 371–
384 (2013).
35. Stern, A. M. & Zhu, J. An introduction to nitric oxide sensing and response in
bacteria. Adv. Appl. Microbiol. 87, 187–220 (2014).
36. Fang, F. C. Antimicrobial reactive oxygen and nitrogen species: concepts and
controversies. Nat. Rev. Microbiol. 2, 820–832 (2004).
37. Deupree, S. M. & Schoenfisch, M. H. Morphological analysis of the
antimicrobial action of nitric oxide on gram-negative pathogens using atomic
force microscopy. Acta Biomater. 5, 1405–1415 (2009).
38. Barraud, N. et al. Nitric oxide-mediated dispersal in single- and multi-species
biofilms of clinically and industrially relevant microorganisms. Microb.
Biotechnol. 2, 370–378 (2009).
39. Barraud, N. et al. Involvement of nitric oxide in biofilm dispersal of
Pseudomonas aeruginosa. J. Bacteriol. 188, 7344–7353 (2006).
40. Schmidt, I., Steenbakkers, P. J. M., op den Camp, H. J. M., Schmidt, K. & Jetten,
M. S. M. Physiologic and Proteomic Evidence for a Role of Nitric Oxide in
Biofilm Formation by Nitrosomonas europaea and Other Ammonia Oxidizers. J.
Bacteriol. 186, 2781–2788 (2004).
41. Hetrick, E., Shin, J. & Stasko, N. Bactericidal efficacy of nitric oxide-releasing
silica nanoparticles. ACS Nano 2, 235–246 (2008).
42. Pankey, G. A. & Sabath, L. D. Clinical relevance of bacteriostatic versus
bactericidal mechanisms of action in the treatment of Gram-positive bacterial
infections. Clin. Infect. Dis. 38, 864–870 (2004).
43. Pamp, S. J., Gjermansen, M., Johansen, H. K. & Tolker-Nielsen, T. Tolerance to
the antimicrobial peptide colistin in Pseudomonas aeruginosa biofilms is linked
to metabolically active cells, and depends on the pmr and mexAB-oprM genes.
Mol. Microbiol. 68, 223–240 (2008).

94
Chapter 4.
Catalytic generation of nitric
oxide: the use of ferrous ion as a
reducing agent
4.1. Introduction
In the previous chapter, the presence of copper complex (namely CuDTTCT)
together with nitrite and ascorbic acid solution has been shown to be effective in
enhancing nitric oxide generation, which subsequently is able to suppress biofilm
formation and disperse pre-formed biofilm. However, the potential of this system to be
applied is limited due to the requirement to add a reducing agent. Therefore, it would be
highly desirable to utilise a substance that is readily available in water and capable of
reducing copper. For instance, humic substances have been shown to be able to reduce
copper(II) to copper(I) under natural water conditions.1 Despite the widespread presence
of humic substances in water, the presence of organic matters, including humic
substances, is generally unwanted in chlorinated/chloraminated water system due the
possible formation of disinfection by-products. As a result, the potential of inorganic
reducing agent, such as iron, is investigated here.

Chapter 4
95
Iron, as one of the most abundant trace elements in atmospheric waters, usually
undergoes chemical and photochemical reactions resulting in its rapid cycling between
ferrous (Fe2+) and ferric (Fe3+) ions.2,3 Iron that exists as Fe2+ ion has been shown to
reduce copper(II) species to copper(I),4 and this event has been observed in many
systems. For instance, oxidation of Fe2+ ion in seawater condition is enhanced by 0.4
log units in the presence of Cu2+ ion.5 Subsequently, a strong and rapid reduction of
Cu2+ ion to Cu+ ion was observed. Solid iron in the form of green rusts, which are
mixed iron(II)/iron(III) hydroxides, have been found to rapidly reduce aqueous Cu2+ ion
to form solid metallic copper.6 Contact killing of bacteria on solid iron surface was
observed when it is used in conjunction with Cu2+ solution, which was attributed to the
reduction of Cu2+ to form toxic Cu+ by the iron surface.7 Moreover, copper substituted
iron oxide, which is in solid form, was found to be reduced and released into solution
phase in the presence of aqueous Fe2+.8 Therefore, the interaction between copper and
iron can happen either homogeneously (both in solution) or heterogeneously (only one
species in solution form).
In this chapter, the use of Fe2+ ion in the form of ferrous chloride, as an
alternative reducing agent for the reduction of CuDTTCT and the subsequent generation
of nitric oxide (NO) was tested in different solution and pH conditions. The effect non-
equimolar addition of nitrite and Fe2+ on catalytic generation of NO was also studied.
Moreover, the effect of Fe2+ addition in dispersing pre-formed nitrifying biofilms via
NO-mediated dispersal was investigated.

Chapter 4
96
4.2. Experimental methods
4.2.1. Synthesis of copper dibenzo[e,k]-2,3,8,9-tetraphenyl-1,4,7,10-tetraaza-
cyclododeca-1,3,7,9-tetraene complex (CuDTTCT complex)
The synthesis of dibenzo[e,k]-2,3,8,9-tetraphenyl-1,4,7,10-tetraaza-cyclododeca-
1,3,7,9-tetraene (DTTCT), copper complex and its films were performed according to
the method described in the previous chapter (Section 3.2.1).
4.2.2. NO generation measurement with Fe2+ solution as the reducing agent
NO generation from the catalytic system was measured amperometrically.
CuDTTCT films or PVC films were placed at the bottom of a 20 mL glass vials
equipped with stir bars and filled with 10 mL of testing solution, either the nitrifying
bacteria growth medium (ATCC medium 2265, which consists of three different stock
solutions – stock 1 (final composition in the medium mixture): 25 mM (NH4)2SO4, 3
mM KH2PO4, 0.7 mM MgSO4, 0.2 mM CaCl2, 0.01 mM FeSO4, 0.02 mM EDTA, 0.5
µM CuSO4; stock 2: 40 mM KH2PO4, 4 mM NaH2PO4, adjusted to pH 8 by 10 M
NaOH; stock 3: 4 mM Na2CO3) or phosphate buffer (consists of 137 mM NaCl, 2.7 mM
KCl, 10 mM Na2HPO4 and 1.8 mM KH2PO4, prepared using PBS tablet from Sigma).
Growth medium at pH 6 was prepared by adjusting the pH of stock 2 from 4.5 to 5.5 by
10 M NaOH, and mixing it with stock 1 and 3. Concentrated hydrochloric acid was used
to adjust the pH of phosphate buffer from 7.4 to 7, 6.5, and 6. PVC or CuDTTCT films
were placed at the bottom of the vial and measurement probe was placed approximately
1 cm from the film. Sodium nitrite (Ajax Finechem) and iron chloride tetrahydrate
(Sigma Aldrich, ≥99%) were used as the catalytic reactant. At the end of the

Chapter 4
97
measurement, an NO scavenger namely 2-Phenyl-4,4,5,5-tetramethylimidazoline-1-oxyl
3-oxide (PTIO; Alexis Biochemicals) was added to re-establish the baseline.
4.2.3. Fe speciation analysis
The concentration of Fe2+/Fe3+ in the solution was analysed over time via
ferrozine spectroscopy method. Colour reagent ferrozine (Aldrich), reducing agent
hydroxylamine hydrochloric acid (Sigma Aldrich), and ammonium acetate buffer (Ajax)
were prepared as described by Viollier etc.9 Two mL of sample solution was added into
12 well plates (Corning) containing either bare film (PVC film without CuDTTCT) or
CuDTTCT film, followed by addition of equimolar amount of nitrite and Fe2+. Iron
speciation measurement was performed for two hours, with two 100 µL aliquots of the
sample were collected every 3 minutes or 20 minutes. The first aliquots were treated
with ferrozine and represent the concentration of Fe2+, while the second aliquots were
treated with ferrozine, reducing agent and buffer to depict the concentration of total
iron. The absorbance of the Fe2+‒ferrozine complex was measured using UV-Vis
spectrophotometer (Varian Cary 300) at 562 nm.
4.2.4. Biofilm dispersal assay
Biofilm was formed from a mixed inoculum of nitrifying bacteria that is
commercially available for aquarium water purification which comprised of
Nitrosospira multiformis, Nitrospira marina, and Bacillus sp. (Aquasonic Bio-Culture).
One mL of the mixed inoculum was added to 100 mL of the nitrifying bacteria growth
medium and incubated for 3 days in the dark (30 °C, 100 rpm). The three-day-old

Chapter 4
98
cultures were inoculated into fresh medium and 2 mL aliquots (OD = 0.008) were added
in 35 mm culture dishes with a coverglass bottom (internal glass diameter 22 mm,
ProSciTech) with the presence of PVC or CuDTTCT films. All dishes were incubated in
the dark for three days at 30 °C, 100 rpm. One hour before incubation ended, the
supernatants were removed and centrifuged at 10,000 rpm for 15 mins to separate the
planktonic bacteria. During centrifugation, phosphate buffer pH 7.4 (PBS) was added to
the biofilm to prevent the cells from drying. Planktonic bacteria were re-dispersed in
sterile phosphate buffer pH 6 solution followed by the addition of sodium nitrite and
iron(II) to trigger the production of nitric oxide and incubation was continued for one
hour. At the end of the second incubation step, planktonic bacteria were removed. The
dishes that contain the biofilm were washed twice with PBS and stained with 400 µL of
staining solution consists of 3.34 µM of SYTO-9 and 19.97 µM of propidium iodide in
PBS (LIVE/DEAD® BacLightTM Bacterial Viability Kits L-7007, Molecular Probes
Inc.). Green stains denote viable bacteria, while red and yellow stains denote non-viable
bacteria. Twelve images across the glass bottom were acquired and the surface coverage
of the biofilm was analysed using image analysis software (Fiji/ImageJ). Statistical
analysis was performed using one-way ANOVA followed by Dunnett post hoc analysis
on Prism (GraphPad).
4.3. Result and discussion
4.3.1. Effect of testing solution and pH on NO generation from CuDTTCT-nitrite-
Fe2+ mixture
In a system that contains copper(II), nitrite, and a reducing agent, generation of
nitric oxide from nitrite can occur catalytically via copper(II)/copper(I) redox cycling,

Chapter 4
99
as shown in the previous chapter and also supported by literature.10,11 Oplander et al. has
reported the use of Fe2+ as a reducing agent that is able to prompt copper redox cycling,
whereby the reaction between Fe2+ and Cu2+ can form the active Cu+ species and Fe3+,
which subsequently react with nitrite to form NO.12 Therefore, it is proposed that in the
system where CuDTTCT is used, NO generation will occur based on the following
equations:
Cu2+(DTTCT) + Fe2+ Cu+(DTTCT) + Fe3+ (i)
Cu+(DTTCT) + NO2- + H2O ⇌ NO + 2 OH- + Cu2+(DTTCT) (ii)
The amount of NO that can be generated from CuDTTCT-nitrite-Fe2+ system
using nitrifying bacteria growth medium (at initial pH of 8) was measured (Figure 4.1).
Upon the addition of Fe2+ into nitrifying bacteria growth medium that already contained
nitrite and CuDTTCT, no change in signal was observed, indicating that NO generation
could not occur in this condition. However, when the pH of the nitrifying bacteria
growth medium was decreased to 6, a significant increase in NO concentration was
observed after the addition of Fe2+. Note that no pH change was observed in the
conditions tested here. This instant increase in NO concentration is quickly followed by
a rapid decrease. Consequently, only low level of NO could be detected for the
remaining time. An NO scavenger, namely PTIO was then added (the time of addition
was denoted by black arrow; Figure 4.1). The addition of PTIO caused an instant drop
in the observed signal, confirming that the change in signal previously observed was
caused by NO. As stated earlier, NO generation via copper redox cycling is highly
dependent on the formation of the active copper(I) species. Consequently, adequate
presence of Fe2+ species as the reducing agent is vital. The extent of Fe2+ autoxidation is
increased at higher pH,5,13 thus, it is possible that this autoxidation resulted in

Chapter 4
100
diminished availability of Fe2+, subsequently, no NO generation was observed at higher
pH.
Figure 4.1 Amperometric measurement of NO generation from CuDTTCT-nitrite-Fe2+
system. Nitrifying medium pH 8 (black) and pH 6 (green) were used as testing solutions.
0.5 mM of sodium nitrite was added followed by the addition of 0.5 mM of Fe2+ solution.
PTIO was added (time of addition was denoted by a black arrow) to re-establish the
baseline.
The reduced availability of Fe2+ at higher pH and the involvement of Fe2+ in
initiating copper redox cycling is supported by the Fe2+/Fe3+ speciation study. The
amount of Fe2+ available in the absence of copper at time 0, pH 6 is around 0.33 mM
(Figure 4.2c), however, Fe2+ concentration at a similar time is significantly lower for pH
8 (0.17 mM, Figure 4.2a). It should be noted that a 10 seconds time lag generally occurs
between the addition of Fe2+ and the sampling of the first point (time 0). Therefore, the
low concentration of Fe2+ detected at pH 8 could be ascribed to the rapid oxidation of
Fe2+ to Fe3+ in the nitrifying bacteria growth medium-nitrite solution. The change in
Fe2+/Fe3+ concentration over time also supports the above postulation, as Fe2+ could not
be detected after 10 min of measurement at pH 8, while Fe2+ can still be detected up to
0 500 1000 1500-50
0
50
100
150
200
250
300N
O c
on
ce
ntr
atio
n (
nM
)
Time (s)
Growth medium pH 6
Growth medium pH 8

Chapter 4
101
18 min at pH 6. Once all Fe2+ was oxidised, only Fe3+ was detected for the remaining
time. Moreover, the presence of phosphate in the nitrifying bacteria medium could also
aid in the oxidation reaction, where the reaction between Fe2+ with phosphate can form
iron-phosphate species, such as iron-hydroxyphosphate or iron(III)-phosphate
precipitate.14–16 Indeed, a slight decrease in total iron concentration was observed at the
end of measurement (from ~0.5 mM to ~0.3 mM; Figure 4.2a and c), indicating that a
small portion of iron has precipitated out of the solution. The presence of carbonate ions
was also reported to assist iron(II) oxidation,5 and the nitrifying bacteria growth
medium used in this study contains 4 mM of carbonate ions. In addition, nitrifying
growth medium also contains EDTA (0.02 mM), which might complex with Fe2+,
therefore limiting the amount of Fe2+ even further.
Figure 4.2 Change in Fe oxidation state over time in the presence of bare film and nitrite
in nitrifying bacteria growth medium at (a) pH 8 and (c) pH 6; in the presence of
CuDTTCT film and nitrite in nitrifying bacteria medium at (b) pH 8 and (d) pH 6.
Theoretical concentration of nitrite and iron used is 0.5 mM.
(a)
(c) (d)
pH
8p
H 6
0.0
0.1
0.2
0.3
0.4
0.5
Iron c
oncentr
ation (
mM
) Fe(II)
Fe(III)
Total Fe
0 10 20 30 40
0.0
0.1
0.2
0.3
0.4
0.5
Iron c
oncentr
ation (
mM
)
Time (min)0 10 20 30 40
Time (min)
(b)
Without CuDTTCT With CuDTTCT

Chapter 4
102
When CuDTTCT is present, the oxidation of Fe2+ to Fe3+ occurs at a
significantly faster rate, as Fe2+ cannot be detected after 3 min measurement, either at
pH 6 or 8 (Figure 4.2b and d). Therefore, it appears that a reaction between the copper
complex inside the PVC matrix with the Fe2+ ions in the solution occurred. Interestingly,
the interaction between copper and Fe2+ still occur at pH 8 (Figure 4.2b), although this
interaction did not result in NO generation, which seems to be caused by the insufficient
Fe2+ available in this condition, as stated earlier (Figure 4.2a and c).
In order to delay Fe2+ oxidation rate, NO generation and Fe2+/Fe3+ speciation
testing was performed in different solution. Phosphate buffer (PB) was chosen because
of its compatibility with bacteria osmotic pressure. Although PB has been shown to
interact with Fe2+ causing its oxidation,13 the presence of phosphate, which is one of the
main species responsible for autoxidation and precipitation of Fe2+, in PB is
approximately four times lower than in nitrifying bacteria growth medium. Moreover,
carbonate ions and EDTA is not present in PB. Therefore, it was postulated that
oxidation of Fe2+ in PB would occur at a slower rate, which enables a greater amount of
NO to be generated.
The amount of NO that can be generated in PB was first tested at pH 7. In this
condition, the maximum concentration of NO that can be generated was up to 20 nM,
which was observed after ~1000 s (or ~17 min) of measurement (Figure 4.3). At
slightly lower pH of 6.5, a higher amount of NO can be generated over a longer period
of time, as up to 400 nM of NO was detected after ~2000 s (or ~33 min; Figure 4.3). At
the lowest pH tested (pH 6), a significantly higher amount of NO generated was
observed, where up to 4000 nM NO was detected after ~2500 s (or ~42 min) of

Chapter 4
103
measurement (Figure 4.3). Note that only a small pH drop (less than 0.1) was observed
upon the addition of nitrite-Fe2+ into the system at all pH tested.
Figure 4.3 Amperometric measurement of NO generation from CuDTTCT-nitrite-Fe2+
system. Phosphate buffer solution with different pH (from top to bottom: pH 7, pH 6.5
and pH 6) were used as testing solutions. 0.5 mM of sodium nitrite was added followed by
the addition of 0.5 mM of Fe2+ solution. PTIO was added (time of addition was denoted by
a black arrow) to re-establish the baseline.
Comparing the NO generation result obtained from both solutions tested at a
similar pH (pH 6), it was found that NO generation in PB is 13 times higher than in
nitrifying bacteria growth medium. Moreover, NO generation in PB occurred over a
longer period of time, as NO can still be detected even after 6000 s (100 min) of
measurement. These differences in NO generation was ascribed to the higher
0 2000 4000 6000 8000
0
1000
2000
3000
4000
0
100
200
300
400
0
10
20
30
40
Time (s)
PB pH 6
PB pH 6.5
NO
co
nce
ntr
atio
n (
nM
)
PB pH 7

Chapter 4
104
availability of Fe2+ ions in PB compared to nitrifying bacteria growth medium, as
evidenced by the speciation studies described below.
Rapid Fe2+ oxidation at higher pH was also observed when PB was used as the
testing solution (Figure 4.4). For instance, at the highest pH tested (pH 7), Fe2+ cannot
be detected after 40 min of measurement (Figure 4.4a), while at a slightly lower pH (pH
6.5) oxidation was only completed after 100 min of measurement (Figure 4.4c). As
expected, Fe2+ autoxidation was slowest at pH 6, as around 0.1 mM of iron(II) can still
be detected at the end of 120 min measurement (Figure 4.4e). Comparing the result
from PB at pH 6 with the result obtained from the use of nitrifying bacteria growth
medium at similar pH, it was observed that the oxidation of Fe2+ occurred faster in
nitrifying bacteria growth medium than in PB. Therefore, this result suggests that the
presence of complexing agents such as phosphate, carbonate, and EDTA, is a more
prominent factor in facilitating iron oxidation compared to pH.

Chapter 4
105
Figure 4.4 Change in Fe oxidation state over time in the presence of bare film and nitrite
in PB at (a) pH 7, (c) pH 6.5 and (e) pH 6; in the presence of CuDTTCT film and nitrite in
PB at (b) pH 7, (d) pH 6.5 and (f) pH 6. Theoretical concentration of nitrite and iron used
is 0.5 mM.
Similar to the observation in nitrifying bacteria medium, Fe2+ oxidation in PB
solution with the presence of CuDTTCT occurred faster compared to that in the absence
of CuDTTCT (Figure 4.4b, d, f). At pH 7, Fe2+ concentration diminished in the first 20
min of measurement (Figure 4.4b). Even at a lower pH of 6.5 and 6, Fe2+ concentrations
were depleted after 20 to 40 min of reaction (Figure 4.4d and f). Again, these results
confirmed the presence of copper-iron interaction, which was then followed by nitrite
(b)(a)
(c)
pH
7p
H 6
.5p
H 6
(d)
(e) (f)
0.0
0.1
0.2
0.3
0.4
0.5
Iro
n c
on
cen
tra
tio
n (
mM
)
Fe(II)
Fe(III)
Total Fe
0.0
0.1
0.2
0.3
0.4
0.5
Iro
n c
on
cen
tra
tio
n (
mM
)
0 20 40 60 80 100 120
0.0
0.1
0.2
0.3
0.4
0.5
Iro
n c
on
cen
tra
tio
n (
mM
)
Time (min)0 20 40 60 80 100 120
Time (min)
Without CuDTTCT With CuDTTCT

Chapter 4
106
reduction to NO in this system, as shown in NO generation measurement before (Figure
4.3). Interestingly, the reaction between copper(II) species and iron(II) species takes
place regardless of the pH value, which is in agreement with earlier report at a pH lower
than 7.5.5 Moreover, it seems that pH does not affect the reduction reaction rate of
copper, as speciation study showed that the rate of Fe2+ oxidation (around 20 to 40 min)
is similar across all pH range tested.
Although the interaction between CuDTTCT and Fe2+ was shown to happen at
all pH tested and at a similar time scale (Figure 4.4b, d, f), the amount of NO that was
generated from these systems is different (Figure 4.3), with higher NO generated at
lower pH. This therefore suggests that pH is the main factor for NO generation. It was
reported that at low pH, nitrite will form nitrous acid that could decompose and liberate
NO.17 Consequently, it is possible that the NO generated at low pH does not necessarily
originate from the interaction between the active copper(I) species with nitrite, but
could be formed as a result of the decomposition of nitrite.
4.3.2. Effect of nitrite and Fe2+ concentration on NO generation from CuDTTCT-
nitrite-Fe2+ mixture
The effect of different concentration of nitrite and Fe2+ on NO generation was
investigated in PB pH 6 (Figure 4.5). In the absence of CuDTTCT, low concentration of
NO was generated toward the end of measurement (Figure 4.5, black line), which was
possibly induced by the acidic pH of the solution or the slow decomposition of nitrite in
the presence of iron ions.12,17,18 In the presence of similar nitrite-Fe2+ concentration with
CuDTTCT, NO generation occured faster and at higher concentration (up to 200 nM of
NO was measured) compared to the system that contains nitrite-Fe2+ alone. This result

Chapter 4
107
indicates that the generation of NO in the presence of CuDTTCT predominantly occur
through nitrite reduction via copper redox cycling, instead of via acidic decomposition
of nitrite. When nitrite and Fe2+ concentrations were increased, as expected, an enhanced
generation of NO was subsequently observed. The cumulative NO generation over time
is in agreement to the NO release profile, i.e. higher NO generation with an increase in
the concentration of nitrite-Fe2+ added (second Y-axis, Figure 4.5).
The time needed to reach the maximum value differ depends on the
concentration, with higher concentration needs longer time to reach the maximum point
compared to lower concentration. For instance, the maximum value in NO generation
was reached after measurement was performed for ~1100 s for CuDTTCT-0.2 mM
nitrite-iron(II), while the peak for the CuDTTCT-0.1 mM nitrite-iron(II) system was
reached in ~850 s. After the maximum point in was achieved, the signal started to
decay. At this point, PTIO (black arrow, Figure 4.5) was added to re-establish the
baseline.

Chapter 4
108
Figure 4.5 Amperometric measurements of NO generation from CuDTTCT-nitrite-Fe2+
system (first Y-axis) and its corresponding cumulative NO generated (second Y-axis).
After stable baseline was observed, sodium nitrite was added followed by the addition of
equimolar concentration of Fe2+ solution. PTIO (black arrow) was added to re-establish
the baseline.
NO generation was also tested at a fixed nitrite concentration (0.1 mM) but at
different concentration of Fe2+ (Figure 4.6). When 0.1 mM of Fe2+ was added to nitrite-
containing PB pH 6 (red line; Figure 4.6), up to 250 nM of NO can be generated from
the mixture, with the maximum value was reached after measurement for 1000 s.
Interestingly, a further increase in Fe2+ concentration to 0.25 mM (blue line; Figure 4.6)
did not change the highest NO concentration that can be generated (up to 250 nM).
Instead, a shift in the time needed to reach this maximum NO value was observed,
whereby a longer period of 2000-3000 s was needed to reach the maximum.
Consequently, cumulative NO generation was observed to be 1.4 times higher. When
the concentration of Fe2+ added was reduced by half (0.05 mM, purple line; Figure 4.6)
or was ten times lower (0.01 mM, green line; Figure 4.6), only a slight decrease (50 nM)
in NO measurement was detected, with both systems can achieve the maximum value
0 2000 4000 6000
0
500
1000
1500
2000
NO
co
nce
ntr
atio
n (
nM
)
Time (s)
CuDTTCT-0.4 mM nit-Fe
CuDTTCT-0.2 mM nit-Fe
CuDTTCT-0.1 mM nit-Fe
0.1 mM nit-Fe
0
1E6
2E6
3E6
Cu
mu
lative
NO
ge
ne
rate
d (
nM
)

Chapter 4
109
after 1000 s of measurement. It should be noted that NO generation study was also
performed under fixed concentration of Fe2+ and different concentration of nitrite;
however, it was found that no clear trend can be observed, i.e. it behaved similarly as
when equimolar concentration of nitrite-Fe2+ was used.
Figure 4.6 Amperometric measurements of NO generation from CuDTTCT-nitrite-Fe2+
mixture (left) and its corresponding cumulative NO generated (right). Bare film (control,
black lines) or CuDTTCT film were used. After stable baseline was observed, sodium
nitrite (0.1 mM) was added followed by the addition different concentration of Fe2+
solution. After the fast decrease of NO peaks was observed, PTIO (black arrow) was
added to re-establish the baseline.
4.3.3. Biofilm dispersal in the presence of CuDTTCT-nitrite-Fe2+ solution
The effectivity of NO generated from CuDTTCT-nitrite-Fe2+ in dispersing pre-
formed nitrifying biofilm following the three days growth in nitrifying bacteria growth
medium (ATCC medium 2265) was tested. Unlike the methodology used in the
previous chapter, biofilm dispersal was studied in a PB pH 6 solution, which has been
shown to generate the maximum amount of NO. The buffer replacement from nitrifying
bacteria growth medium (approximate pH of 8) to phosphate buffer solution pH 6 do
0 2000 4000 6000
0
100
200
0
100
200
0
100
200
0
100
200
0
100
200
Time (s)
0.25 mM Fe
0.05 mM Fe
0.1 mM Fe
NO
co
nc
en
tra
tio
n (
nM
) 0.01 mM Fe
0.1 mM Fe (Control)
0 2000 4000 6000
0.0
3.0E5
6.0E5
0.0
3.0E5
6.0E5
0.0
3.0E5
6.0E5
0.0
3.0E5
6.0E5
0.0
3.0E5
6.0E5
Time (s)
Cu
mu
lati
ve N
O g
en
era
ted
(n
M)

Chapter 4
110
not cause any premature biofilm dispersal, as surface coverage analysis showed that
biofilms underwent buffer replacement are not significantly different compared to the
(untreated) control (Figure 4.7), which is in agreement with previous report.19
Figure 4.7 (a) Surface coverage analysis of nitrifying biofilm after buffer and pH change.
Both variations was grown in the absence of CuDTTCT. All values shown are normalised
to the surface coverage of control. Error bars indicate standard error between replicates
(n = 2). * p ≤ 0.05 compared to the control. Confocal laser scanning microscopy images of
nitrifying bacteria biofilm (b) untreated (control), (c) after buffer replacement with
phosphate buffer pH 6. Scale bar = 100 µm.
The addition of CuDTTCT was found not to affect biofilm growth, as the
surface coverage of biofilm that was grown in the presence of CuDTTCT is similar with
the one grown in the absence of CuDTTCT (control, Figure 4.8a-c). When the pre-
formed biofilms were treated with 0.1 mM and 0.2 mM of nitrite and Fe2+, a ~35%
reduction in biofilm surface coverage compared to the control was observed (Figure
4.8a, d-e). Enhanced biofilm surface coverage reduction was observed when 0.4 mM of
nitrite-Fe2+ was added into the wells, which resulted in a 45% reduction in biofilm
surface coverage compared to the control (Figure 4.8f). Although the addition of iron
species has been shown to disperse pre-formed Pseudomonas aeruginosa biofilm, the
effect was not observed here as the biofilms treated with Fe2+ alone at varying
(a) (b)
(c)
Control
PB pH 6
Control PB pH 60.0
0.5
1.0
1.5
Re
lative
su
rfa
ce
co
ve
rag
e
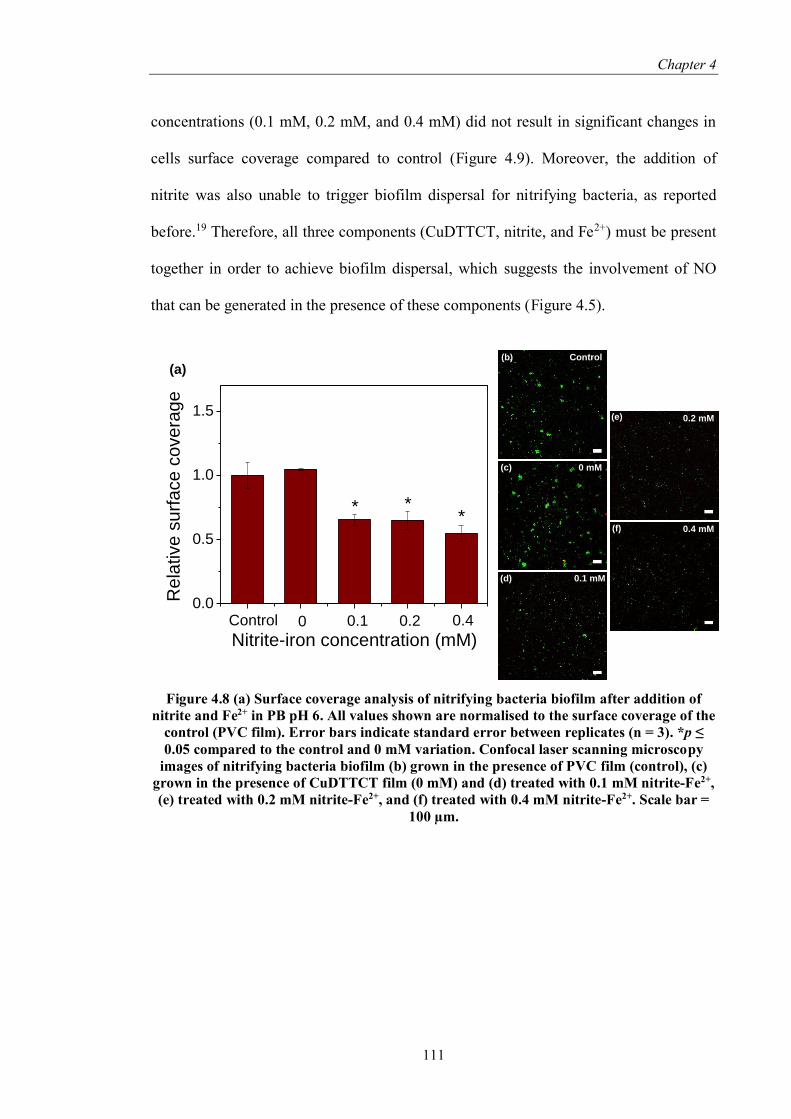
Chapter 4
111
concentrations (0.1 mM, 0.2 mM, and 0.4 mM) did not result in significant changes in
cells surface coverage compared to control (Figure 4.9). Moreover, the addition of
nitrite was also unable to trigger biofilm dispersal for nitrifying bacteria, as reported
before.19 Therefore, all three components (CuDTTCT, nitrite, and Fe2+) must be present
together in order to achieve biofilm dispersal, which suggests the involvement of NO
that can be generated in the presence of these components (Figure 4.5).
Figure 4.8 (a) Surface coverage analysis of nitrifying bacteria biofilm after addition of
nitrite and Fe2+ in PB pH 6. All values shown are normalised to the surface coverage of the
control (PVC film). Error bars indicate standard error between replicates (n = 3). *p ≤
0.05 compared to the control and 0 mM variation. Confocal laser scanning microscopy
images of nitrifying bacteria biofilm (b) grown in the presence of PVC film (control), (c)
grown in the presence of CuDTTCT film (0 mM) and (d) treated with 0.1 mM nitrite-Fe2+,
(e) treated with 0.2 mM nitrite-Fe2+, and (f) treated with 0.4 mM nitrite-Fe2+. Scale bar =
100 µm.
0.0
0.5
1.0
1.5
**
Re
lative
su
rfa
ce
co
ve
rag
e
Nitrite-iron concentration (mM)Control 0 0.1 0.2 0.4
*
(b)
(c)
(d)
(e)
(f)
Control
0 mM
0.1 mM
0.2 mM
0.4 mM
(a)

Chapter 4
112
Figure 4.9 (a) Surface coverage analysis of nitrifying biofilm after the addition of Fe2+ in
PB pH 6. All values shown are normalised to the surface coverage of the control. Error
bars indicate standard error between replicates (n = 2). *p ≤ 0.05 compared to the control.
Confocal laser scanning microscopy images of nitrifying biofilm (b) untreated (control),
(c) treated with 0.1 mM Fe2+, (d) treated with 0.2 mM Fe2+, and (e) treated with 0.4 mM
Fe2+. Scale bar = 100 µm.
The activity of NO to disperse pre-established biofilm is known for various
bacteria species, including nitrifying bacteria.19–22 NO could affect the switch between
biofilm and planktonic phase through alteration in the secondary cellular messenger
level, i.e. cyclic di-GMP level. Because NO-mediated dispersal occurs via a signalling
pathway, cell viability will still be maintained after this treatment, which is confirmed
by the observation presented in the CLSM analysis (Figure 4.8b-f). Moreover, NO is
also known to cause reduction in cell aggregation.23 Indeed, confocal images showed
that in systems without nitrite-Fe2+ treatment, both in the absence or presence of
CuDTTCT, significant cells aggregation could be seen (Figure 4.8a-b). However, when
the biofilm was treated with nitrite and Fe2+, reduction in cells aggregation was
observed (Figure 4.8d-f).
Comparing the result obtained in the previous chapter (section 3.3.4) and
presented herein, it can be concluded that NO generated catalytically via copper redox
(a)
(c) 0.1 mM
(b) Control (d) 0.2 mM
0.4 mM(e)
0.0
0.5
1.0
1.5
Re
lative
su
rfa
ce
co
ve
rag
e
Iron concentration (mM)
Control 0.1 0.2 0.4

Chapter 4
113
cycling is effective in dispersing pre-formed biofilm, regardless of the reducing agent
used. Moreover, NO-mediated dispersal was also observed under different solution
condition, i.e. nitrifying bacteria growth medium at pH 8 and PB pH 6, which implies
that biofilm dispersal by NO is not dependent on the buffer condition, and interestingly,
achievable in an environment that does not support growth such as PB.
4.4. Summary
Herein, Fe2+ species was studied as an alternative reducing agent for catalytic
generation of NO from nitrite in the presence of CuDTTCT catalyst. Amperometric
measurement showed that the combination of CuDTTCT-nitrite-Fe2+ was only able to
generate NO in nitrifying bacteria growth medium at pH 6 and not at pH 8, due to the
rapid autoxidation and precipitation of Fe2+ at high pH. Moreover, iron speciation study
showed that an interaction between CuDTTCT and Fe2+ exist at all pH conditions. The
oxidation rate of Fe2+ was faster in the presence of CuDTTCT compared to control and
at a similar kinetics for all pH conditions studied. Consequently, this interaction resulted
in the reduction of copper(II) species to copper(I), which can catalyse nitrite reduction
to form NO. Similar observation was found in PB systems, whereby higher NO was
generated at lower pH. Moreover, it was also found that the presence of complexing
agents, particularly phosphate, carbonate, and EDTA, is a more prominent factor in
influencing Fe2+ autoxidation compared to pH. Consequently, PB pH 6 was found to
generate the highest amount NO from all the different buffer medium and pH conditions
tested here. The potential of this system in dispersing pre-formed nitrifying biofilm was
performed in PB at pH 6. The NO generated in this condition is effective at dispersing
biofilm and reducing cell aggregations. The results from this and previous chapter

Chapter 4
114
therefore suggest that NO-mediated biofilm dispersal is not affected by the choice of
reducing agent or by the buffer condition.
4.5. References
1. Pham, A., Rose, A. & Waite, T. Kinetics of Cu (II) reduction by natural organic
matter. J. Phys. Chem. A 116, 6590–6599 (2012).
2. Sedlak, D. L. & Hoigné, J. The role of copper and oxalate in the redox cycling of
iron in atmospheric waters. Atmos. Environ. Part A. Gen. Top. 27, 2173–2185
(1993).
3. Willey, J. D. & Whitehead, R. F. Oxidation of Fe (II) in Rainwater. Environ. Sci.
Technol. 39, 2579–2585 (2005).
4. Matocha, C. J., Karathanasis, A. D., Rakshit, S. & Wagner, K. M. Reduction of
copper(II) by iron(II). J. Environ. Qual. 34, 1539–1546 (2005).
5. González, A. G., Pérez-Almeida, N., Magdalena Santana-Casiano, J., Millero, F.
J. & González-Dávila, M. Redox interactions of Fe and Cu in seawater. Mar.
Chem. 179, 12–22 (2016).
6. O’Loughlin, E. J., Kelly, S. D., Kemner, K. M., Csencsits, R. & Cook, R. E.
Reduction of AgI, AuIII, CuII, and Hg II by FeII/FeIII hydroxysulfate green rust.
Chemosphere 53, 437–446 (2003).
7. Mathews, S., Kumar, R. & Solioz, M. Copper reduction and contact killing of
bacteria by iron surfaces. Appl. Environ. Microbiol. 81, 6399–6403 (2015).
8. Frierdich, A. J. & Catalano, J. G. Fe(II)-mediated reduction and repartitioning of
structurally incorporated Cu, Co, and Mn in iron oxides. Environ. Sci. Technol.
46, 11070–11077 (2012).
9. Viollier, E., Inglett, P. W., Hunter, K., Roychoudhury, A. N. & Van Cappellen,
P. The ferrozine method revisited: Fe(II)/Fe(III) determination in natural waters.

Chapter 4
115
Appl. Geochemistry 15, 785–790 (2000).
10. Opländer, C. et al. Characterization of novel nitrite-based nitric oxide generating
delivery systems for topical dermal application. Nitric Oxide 28, 24–32 (2013).
11. Oh, B. K. & Meyerhoff, M. E. Catalytic generation of nitric oxide from nitrite at
the interface of polymeric films doped with lipophilic Cu(II)-complex: a potential
route to the preparation of thromboresistant coatings. Biomaterials 25, 283–293
(2004).
12. Opländer, C. et al. Redox-mediated mechanisms and biological responses of
copper-catalyzed reduction of the nitrite ion in vitro. Nitric Oxide 35, 152–164
(2013).
13. Welch, K. D., Davis, T. Z. & Aust, S. D. Iron autoxidation and free radical
generation: effects of buffers, ligands, and chelators. Arch. Biochem. Biophys.
397, 360–369 (2002).
14. Van Der Grift, B., Rozemeijer, J. C., Griffioen, J. & Van Der Velde, Y. Iron
oxidation kinetics and phosphate immobilization along the flow-path from
groundwater into surface water. Hydrol. Earth Syst. Sci. 18, 4687–4702 (2014).
15. Grift, B. van der et al. Fe hydroxyphosphate precipitation and Fe ( II ) oxidation
kinetics upon aeration of Fe ( II ) and phosphate-containing synthetic and natural
solutions. Geochim. Cosmochim. Acta 186, 71–90 (2016).
16. Baken, S., Moens, C., van der Grift, B. & Smolders, E. Phosphate binding by
natural iron-rich colloids in streams. Water Res. 98, 326–333 (2016).
17. Lundberg, J. O., Weitzberg, E. & Gladwin, M. T. The nitrate-nitrite-nitric oxide
pathway in physiology and therapeutics. Nat. Rev. Drug Discov. 7, 156–167
(2008).
18. Van Cleemput, O. & Baert, L. Nitrite stability influenced by iron compounds.
Soil Biol. Biochem. 15, 137–140 (1983).
19. Schmidt, I., Steenbakkers, P. J. M., op den Camp, H. J. M., Schmidt, K. & Jetten,
M. S. M. Physiologic and Proteomic Evidence for a Role of Nitric Oxide in

Chapter 4
116
Biofilm Formation by Nitrosomonas europaea and Other Ammonia Oxidizers. J.
Bacteriol. 186, 2781–2788 (2004).
20. McDougald, D., Rice, S. A., Barraud, N., Steinberg, P. D. & Kjelleberg, S.
Should we stay or should we go: mechanisms and ecological consequences for
biofilm dispersal. Nat. Rev. Microbiol. 10, 39–50 (2011).
21. Craven, M. et al. Nitric Oxide-Releasing Polyacrylonitrile Disperses Biofilms
Formed by Wound-Relevant Pathogenic Bacteria. J. Appl. Microbiol. 120, 1085–
1099 (2016).
22. Barnes, R. J. et al. Nitric oxide treatment for the control of reverse osmosis
membrane biofouling. Appl. Environ. Microbiol. 81, 2515–2524 (2015).
23. Schlag, S., Nerz, C., Birkenstock, T. A., Altenberend, F. & Götz, F. Inhibition of
staphylococcal biofilm formation by nitrite. J. Bacteriol. 189, 7911–7919 (2007).

117
Chapter 5.
Iron complex facilitated copper
redox cycling for nitric oxide
generation
(This chapter is based on the work published in ACS Applied Materials and Interfaces
volume 8, issue 44, page 30502-30510, 2016.)
5.1. Introduction
The studies presented in the previous chapters (Chapter 3 and 4) have shown that
aqueous reducing agents are required for nitric oxide (NO) generation. However, these
reducing agents are not always readily available in water. The need for continuous
addition of an external reducing agent is not desirable as it is costly, difficult to control
the dose and the added reducing agent might affect water composition and quality.
Moreover, the presence of oxidants chlorine and chloramine, which are commonly
added to water systems as disinfectants, might neutralise the reducing agent.
Consequently, a reducing agent that can be immobilised together with the copper
catalyst within a poly(vinyl chloride) (PVC) matrix will be highly beneficial. In
particular, the utilisation of iron complex is of interest.

Chapter 5
118
Iron species, which is capable of reducing copper catalyst as shown in Chapter 4,
is known to assist various redox processes. For instance, an iron-humic acid complex
was found to accelerate the microbial reduction of iron(III) oxide.1 Several parameters
should be considered in ligand selection suitable for complex formation. For instance,
iron ligands can be either oxygen, nitrogen, or sulphur atoms-based. Oxygen-based
ligands, such as citrate, prefer to complex with iron(III), consequently, these ligands
will promote the oxidation of iron(II) to iron(III) and a decrease in iron reduction
potential can be observed.2 On the other hand, nitrogen- and sulphur-based ligands can
stabilise iron(II), therefore they can inhibit iron(II) oxidation and cause an increase in
iron reduction potential.2,3 Therefore, nitrogen- or sulphur-based ligands are preferred
in this study, due to the required presence of iron(II) species in facilitating
copper(II)/copper(I) redox cycling for NO generation. Moreover, the ligand should also
able to accommodate the change in metal ion size from reduction/oxidation process that
will occur.4 DTTCT (dibenzo[e,k]-2,3,8,9-tetraphenyl-1,4,7,10-tetraaza-cyclododeca-
1,3,7,9-tetraene), which is used as the ligand for the copper complex catalyst employed
in this study, is a nitrogen-based ligand that has been proven capable of accommodating
metal ion size change, as evidenced by x-ray photoelectron spectroscopy (XPS) study
presented in Chapter 3. Interestingly, it was reported that ligands which prefer
copper(II) species will possess an affinity for other bivalent metals, such as iron(II).3
Therefore, DTTCT seems to be a suitable ligand for iron complexation in this study.
Herein, iron and copper complexes (FeDTTCT and CuDTTCT, respectively)
were immobilised within a PVC matrix. The ability of the immobilised FeDTTCT in
facilitating the reduction of copper(II) in CuDTTCT to the active copper(I) species and
its subsequent generation of NO via nitrite reduction was assessed. The effectiveness of

Chapter 5
119
the mixed metal system in inhibiting nitrifying bacteria biofilm formation without any
additional reducing agent was also examined.
Scheme 5.1 DTTCT structure
5.2. Experimental methods
5.2.1. Synthesis of dibenzo[e,k]-2,3,8,9-tetraphenyl-1,4,7,10-tetraaza-cyclododeca-
1,3,7,9-tetraene ligand and the metal complexes
The synthesis of dibenzo[e,k]-2,3,8,9-tetraphenyl-1,4,7,10-tetraaza-cyclododeca-
1,3,7,9-tetraene (DTTCT) and copper complex were performed according to the method
described in the previous chapter (Section 3.2.1 and Section 4.2.1). Synthesis of iron
complex was carried out by mixing iron(III) chloride hexahydrate (0.05 mol; Ajax APS)
with DTTCT (0.01 mol) in ethanol and refluxed at 85 °C for 6 hours. After cooling
overnight, the resultant precipitate was separated by centrifugation, washed with cold
ethanol and dried in vacuum desiccator yielding a dark brown powder.
The films were synthesised by mixing the metal complex with pre-dissolved
poly(vinyl chloride) (PVC) in tetrahydrofuran (THF, Chem-Supply) solution (0.3 mL,
66 mg/L) in an ultrasonic bath. Round glass cover slips (18 mm diameter, ProSciTech)

Chapter 5
120
were washed with dilute nitric acid, acetone, and ethanol followed by overnight drying
at 110 °C before being used as the cast template. Bare film (PVC without any metal
complexes, typical weight ~20 mg) was used as the control. To prepare the films that
contain a mixture of the CuDTTCT and FeDTTCT metal complexes, CuDTTCT (0.25
mg, 0.5 mg, 0.75 mg, and 1 mg; corresponding to 1.25 wt%, 2.5 wt%, 3.5 wt%, and 5
wt%, respectively) and FeDTTCT (equal loading) were mixed with the pre-dissolved
PVC in THF solution in an ultrasonic bath, and then casted on the round glass cover
slips. All films were dried at 50 °C for 12 hours and peeled off from the glass cover
slips.
5.2.2. Characterisation of metal complexes
Analysis of the structure was performed in deuterated dimethyl sulfoxide solvent
by 1H-NMR spectroscopy using a Bruker Avance III HD 400 MHz. Thermal analysis of
the ligands and metal complexes were carried out by Differential Scanning Calorimetry
(DSC; Perkin Elmer) to confirm the metal complex formation. Around 3 to 6 mg of the
sample were placed in a closed aluminium pan, with empty closed aluminium pan as the
reference. The temperature was increased at a rate of 10 °C/mins under nitrogen
environment (50 mL/min). Results shown are a representative thermogram obtained
from duplicates.
A field emission scanning electron microscopy (SEM; FEI Nova NanoSEM 230)
equipped with an energy dispersive X-ray detector (EDS; Bruker Silicon Drift Energy
Dispersive X-ray detector) was used to show the immobilisation of copper and iron
complexes in PVC matrix. Prior to imaging, the samples were attached to 10 mm metal

Chapter 5
121
mounts by carbon tape and sputter coated with chromium. Data analysis was performed
with Esprit EDS software.
XPS measurements were performed using an ESCALAB 220iXL spectrometer
with monochromated Al Kα (energy 1486.6 eV). Film samples were cleaned by brief
immersion in toluene prior to analysis. Reaction was performed by immersing the pre-
cleaned sample in a 10 mM phosphate buffer solution pH 6 (Sigma, pH adjusted by
concentrated hydrochloric acid) containing 1 mM sodium nitrite (Ajax Finechem) for 10
min. All film samples were dried in a vacuum desiccator for a minimum of 2 days
before analysis. A random spot on the film with diameter around 0.5 mm was chosen
for the analysis. All spectra were calibrated to the C1s peak signal at 284.8 eV.
5.2.3. NO generation measurement
NO generation from the system was measured amperometrically by using an
Apollo TBR4100 Free Radical Analyser (World Precision Instrument) equipped with
ISO-NOP 2 mm probe. Calibration was performed using S-nitroso-N-
acetylpenicillamine (SNAP; Sigma) and copper sulphate solution according to
manufacturer’s protocol. The test film was placed at the bottom of a 20 mL glass vial
equipped with stir bars and filled with 10 mL of 10 mM phosphate buffer solution pH 6.
After a stable baseline was observed, 1 mM sodium nitrite was added to the glass vial.
At the end of the measurement, an NO scavenger namely 2-phenyl-4,4,5,5-
tetramethylimidazoline-1-oxyl 3-oxide (PTIO; Alexis Biochemicals) was added to re-
establish the baseline. All experiments were performed in the presence of ambient
oxygen. Result shown are a representative graph obtained from a minimum of three
replicates.

Chapter 5
122
5.2.4. Biofilm assay
The effect of the mixed metal complex films on nitrifying biofilms was assessed
as previously described (Section 3.2.4). Biofilm was formed from a mixed inoculum of
nitrifying bacteria that is available commercially for the purification of aquarium water
and is comprised of ammonia-oxidising bacteria Nitrosospira multiformis, nitrite-
oxidising bacteria Nitrospira marina, and heterotrophic bacteria Bacillus sp. (Aquasonic
Bio-Culture). One millilitre of the mixed inoculum was added into 100 mL of the
nitrifying growth medium (ATCC medium 2265 consisting of three different stock
solutions – stock 1 (final composition in the medium mixture): 25 mM (NH4)2SO4, 3
mM KH2PO4, 0.7 mM MgSO4, 0.2 mM CaCl2, 0.01 mM FeSO4, 0.02 mM EDTA, 0.5
µM CuSO4; stock 2: 40 mM KH2PO4, 4 mM NaH2PO4, adjusted to pH 8 by 10 M
NaOH; stock 3: 4 mM Na2CO3) and incubated for 3 days in the dark (30 °C, 100 rpm).
The three-day-old cultures were inoculated into fresh medium and 2 mL aliquots (OD =
0.008) were added into 12-well plates with the presence of a bare film (control) or a
mixed metal film. The plates were then incubated in the dark for 3 days at 30 °C, 100
rpm. At the end of incubation, the bacteria supernatants were removed and centrifuged
at 12,000 rpm for 15 min and washed in phosphate buffer saline (PBS; Sigma) once to
separate planktonic bacteria. The supernatants were filtered through a 0.2 µm membrane
and used to measure the extent of copper and iron leaching from the films by ICP-OES
(PerkinElmer OPTIMA 7300). The biofilms were washed twice to remove the loosely
attached cells. The biomass was measured by quantifying total protein content by using
the bicinchoninic acid method (BCA assay; Sigma). BCA working reagent (2 mL) was
added into each of the biofilm and planktonic cells, followed by incubation for 30 mins
at 37 °C, 100 rpm. A preliminary experiment has shown that the presence of a mixed

Chapter 5
123
metal film did not interfere with the assay, even after the incubation step. Standard
solutions as per manufacturer’s instructions were used for each experiment.
Surface coverage analysis and cell viability assessment were performed using
confocal laser scanning microscopy (Olympus FluoViewTM FV1000) of LIVE/DEAD-
stained biofilm cells. For microscopic observation, an aliquot of the nitrifying culture
was added into a 35 mm culture dishes with coverglass bottom (internal glass diameter
22 mm, ProSciTech) with the presence of a bare film (control) or a mixed metal film,
followed by incubation in the dark for three days at 30 °C, 100 rpm. At the end of
incubation, the supernatants were removed and biofilms attached on the glass
substratum were washed twice with PBS. Adhered cells were stained with SYTO-9 and
propidium iodide according to manufacturer’s instruction (LIVE/DEAD BacLightTM
bacterial viability kits L-7007, Molecular Probes Inc.) and incubated at room
temperature for a minimum of 15 min. Twelve pictures across the glass bottom were
obtained and surface coverage analysis was performed on live cells (green channel)
using image analysis software (Fiji/ImageJ). All statistical analyses were performed on
GraphPad Prism 6 (GraphPad Software) using one-way ANOVA method followed by
Dunnett’s posthoc analysis.
5.3. Result and discussion
5.3.1. Confirmation of metal complexes formation
Copper complex (CuDTTCT) and iron complex (FeDTTCT) were synthesised
and characterised. 1H-NMR analysis of the metal complexes shows similar spectra as
the one observed for DTTCT, thus confirming the presence of DTTCT in the metal

Chapter 5
124
complex (Figure 3.1). Thermal analysis by DSC shows ligand DTTCT only exhibited
one strong endothermic peak at 124 °C. Further complexation with copper generated
four endothermic peaks at 121 °C, 156 °C, 204 °C and 269 °C (Figure 5.1). The
sharpest peak of 269 °C could be ascribed to the melting point-decomposition of
CuDTTCT sample. This shift in melting point from 124 °C for DTTCT to 269 °C
confirmed the successful formation of the complex. For FeDTTCT sample, one
exothermic peak at 153 °C was observed, followed by 3 endothermic peaks at 165 °C,
193 °C, and 248 °C. The exothermic peak at 153 °C is possibly due to the
decomposition of the sample, as observed previously by Breviglieri et al.5 The absence
of sharp endothermic peak at 124 °C again confirmed the formation of metal complex.5–
7 Finally, SEM-EDS mapping was performed on the PVC film that contains both
CuDTTCT and FeDTTCT (mixed metal film). Copper was uniformly detected
throughout the samples, while iron mapping showed aggregated distribution (Figure
5.2). Visual observation of the films also confirms this as dark brown aggregates (the
iron complex) were observed on the surface of the samples. These results confirmed the
presence of copper and iron in the film, and subsequently, in the complexes.
Figure 5.1 DSC thermograms of DTTCT, CuDTTCT, and FeDTTCT with a heating rate
of 10 °C/min.
0 100 200 300-12
-8
-4
0
4
8
Heat flo
w (
mW
)
Temperature (°C)
DTTCT
0 100 200 300
Temperature (°C)
CuDTTCT
0 100 200 300
Temperature (°C)
FeDTTCT

Chapter 5
125
Figure 5.2 (a) SEM images of mixed metal film (CuDTTCT+FeDTTCT) and its
corresponding elemental mapping of (b) copper and (c) iron. (d) EDS spectra of the mixed
metal film. Other peak labels are removed for clarity.
5.3.2. Nitric oxide generation from CuDTTCT+FeDTTCT films in the presence of
nitrite
Evaluation of NO release in systems containing mixed CuDTTCT+FeDTTCT
metal complexes films (along with their associated set of controls) was performed
amperometrically, where NO peak can be observed upon the addition of 1 mM nitrite
into the systems (Figure 5.3). Low and stable levels of NO was generated in the various
control systems. In the case of the bare PVC film (without metal complex) and film with
1 mg FeDTTCT loading, up to 25 nM NO were generated, which is thought to result
from the formation of nitrous acid from nitrite and its subsequent decomposition in
acidic pH.8 A higher level of 50 nM NO was generated in the presence of film with 1
mg CuDTTCT loading. The interactions of copper(II) with various NO donors, such as
(a) (b) (c)
(d)

Chapter 5
126
nitrite and S-nitrosothiols, have been reported to generate low levels of NO.9–11
Interestingly, when the combination of CuDTTCT+FeDTTCT mixed metal film at
(0.5+0.5) mg loading was used, a surge of up to 125 nM NO was generated in the first
~750 s of measurement, followed by a slow decrease in NO concentration. The addition
of the NO scavenger PTIO at the end of the measurement caused a sudden decrease in
the signal, confirming the generation of NO. The significant generation of NO from
nitrite in the presence of the mixed metal complexes is most likely facilitated by
copper(II)/copper(I) redox cycling, with iron complex acting as reducing agent, as
suggested by the following equations:
Cu2+(DTTCT) + Fe2+(DTTCT) ⇌ Cu+(DTTCT) + Fe3+(DTTCT) (i)
Cu+(DTTCT) + NO2– + H2O ⇌ NO + 2 OH‒ + Cu2+(DTTCT) (ii)
As shown in previous chapters, NO generation from nitrite can occur via
copper(II)/copper(I) redox cycling in the presence of a reducing agent (such as ascorbic
acid and Fe2+ ions). Copper(I) species, which was formed from reduction of copper(II)
species by the reducing agent, is the most active species for NO production from
nitrite.12 Copper(II) reduction by iron species and importantly, the generation of NO
from nitrite via reaction with the formed copper(I) species, have been observed by
several studies,9,13–17 including the one shown in Chapter 4. However, up to this stage,
copper(II) reduction by iron(II) has been reported in solution form, either
homogeneously (both metals as dissolved species) or heterogeneously (with one metal
in solution form).16 Eventually, when the formed copper(I) is oxidised to copper(II) and
iron-facilitated reduction of copper(II) does not happen anymore, NO generation will
stop.

Chapter 5
127
Doubling the mixed metal complexes loading to (1+1) mg caused an up to 2-
fold increase in NO generation in the presence of nitrite, which was detected within
similar time frame as that of the lower loading of metal complexes (in the first 750 s).
This was again followed by a similar slow decrease in NO concentration prior to PTIO
addition. This enhanced NO generation was due to the presence of more catalytic active
sites with the increasing amount of metal complexes.
Figure 5.3 Amperometric measurements of NO generation from mixed metal films
containing a mixture of metal complex (CuDTTCT+FeDTTCT; 1+1 and 0.5+0.5) with the
addition of 1 mM nitrite. Three different controls were used (bare film, 1 mg CuDTTCT
film, 1 mg FeDTTCT film) in the presence of 1 mM nitrite solution. PTIO (100 µM; black
arrow) was added at the end of measurement to re-establish the baseline.
The iron complex-facilitated copper(II)/copper(I) redox cycling is supported by
the observed changes in the binding energy of iron and copper species, as revealed by
XPS analysis of the mixed metal complexes before and after addition of nitrite (Figure
5.4). The XPS spectra of FeDTTCT in the mixed metal film can be deconvoluted into
two Fe 2p3/2 peaks at 711.3 eV and 713.9 eV, with a satellite peak at 719.4 eV (Figure
5.4b, before reaction), which is similar to that of the as-synthesised FeDTTCT (Figure
0 1000 2000 3000 4000
-50
0
50
100
150
200
250
NO
co
nce
ntr
atio
n (
nM
)
Time (s)
(1+1) mg
(0.5+0.5) mg
Cu 1 mg
Fe 1 mg
bare

Chapter 5
128
5.4a). Note that there is currently no published data on the XPS spectra of FeDTTCT,
and the position of iron(II) and iron(III) peaks in Fe 2p region could differ depending on
the ligand species.18 The CuDTTCT in the mixed metal film mainly exists as copper(II)
species, with detection of Cu 2p3/2 peak at 934.7 eV along with two satellite peaks at
940.1 eV and 943.7 eV (Figure 5.4c, before reaction). Following NO generation in the
presence of nitrite, a significant Fe 2p3/2 peak shift from 711.3 eV to 712.1 eV was
observed, which was followed by the shift in the corresponding satellite peak to 718.6
eV and the disappearance of the second Fe 2p3/2 peak (Figure 5.4b, after reaction). This
shift of Fe 2p3/2 peak to higher binding energy indicates oxidation of the iron
complex.18,19 Post reaction XPS analysis also revealed the emergence of copper(I) peak
at 933.9 eV (Figure 5.4c, after reaction), which as stated previously, is the active species
in NO generation reaction. Therefore, this observation implies the role of the iron
complex as a reducing agent, facilitating the catalytic activity of copper through
copper(II)/copper(I) redox cycling for generation NO from nitrite. Further studies on
catalytic NO generation in solutions containing nitrite and ascorbic acid (as reducing
agent) detected a significantly higher NO concentration in the presence of CuDTTCT
than that of FeDTTCT (Figure 5.5). This further validates the role of FeDTTCT in
facilitating the reduction of copper(II) to copper(I) in the mixed metal systems, rather
than catalysing the reduction of nitrite to NO.

Chapter 5
129
Figure 5.4 XPS spectra of Fe 2p region of (a) as synthesised FeDTTCT, (b)
CuDTTCT+FeDTTCT (mixed) film before and after reaction. (c) XPS spectra of Cu 2p
region of CuDTTCT+FeDTTCT (mixed) film before and after reaction. Reaction was
performed in the presence of 1 mM of nitrite for 10 mins.
740 730 720 710 700
Counts
(s
-1)
Binding energy (eV)
Before reaction
After reaction
970 960 950 940 930 920
Cu2+
After reaction
Binding energy (eV)
Cu+
Cu2+
Before reaction
(b) (c)
(a)
740 730 720 710 700
Co
un
ts (
s-1)
Binding energy (eV)
FeDTTCT as synthesised

Chapter 5
130
Figure 5.5 Amperometric measurements of NO generation from CuDTTCT film (1 mg
loading) or FeDTTCT film (1 mg loading) in the presence of 1 mM nitrite and 1 mM
ascorbic acid. Bare film (PVC film without the addition of metal complex) was used as the
control. Black arrow denotes the addition of ascorbic acid.
5.3.3. Antibiofilm activity of CuDTTCT+FeDTTCT mixed metal film by the
involvement of catalytically generated NO
The potential of CuDTTCT+FeDTTCT mixed metal complexes as NO-
generating material for antibiofilm application was investigated against the
ubiquitously-occurring nitrifying bacterial biofilms. Nitrifying bacteria of the
Nitrosospira genera are commonly known as ammonia oxidising bacteria or AOB,
capable of oxidising ammonia to nitrite. Nitrite production from ammonia occurs via a
two-step process involving the conversion of ammonia to hydroxylamine catalysed by
the enzyme ammonia monooxygenase (Amo), which then followed by the conversion of
hydroxylamine to nitrite by the enzyme hydroxylamine oxidoreductase (Hao). A mixed
inoculum containing nitrifying bacteria was used to grow nitrifying biofilms for three
days in a specific ammonia oxidising bacteria culture medium in the presence of bare
0 1000 2000 3000 4000 5000 6000
0
1000
2000
3000
4000
5000
NO
concentr
ation (
nM
)
Time (s)
CuDTTCT
Bare
FeDTTCT

Chapter 5
131
(PVC-only) film, whereby 16 mM nitrite was detected in the system (by ion
chromatography) after 3 days. Low level of 0.1 mM nitrate was also detected in the
system, potentially due to the presence of nitrite oxidising bacteria (NOB) Nitrospira
spp. in the inoculum, which are capable of oxidising nitrite to nitrate. The biofilm
growth was also characterised by detection of ~300 µg/mL protein on the well and bare
film surfaces, while ~100 µg/mL of protein was detected in the suspended planktonic
phase. In this self-produced nitrite environment, the mixed metal systems exhibited a
unique anti-biofilm effect, as described in the following.
The extent of biofilm and planktonic growth in the presence of increasing mixed
metal loading of (0.25 mg Cu + 0.25 mg Fe), (0.5 mg Cu + 0.5 mg Fe) and (0.75 mg Cu
+ 0.75 mg Fe) along with their corresponding copper and iron leached ions are
presented in Figure 5.6 and Table 5.1, respectively. The presence of (0.25+0.25) mg
mixed metal complexes did not affect bacterial growth, as comparable biofilm and
planktonic biomass as those detected in the control was observed (Figure 5.6a).
Increasing the metal loading to (0.5+0.5) mg caused a 20% reduction in biofilm biomass
relative to the control growth and correspondingly, a 20% increase in planktonic
biomass. Ultimately, at (0.75+0.75) mg metal loading, a 36% decrease in biofilm
biomass was observed, which was followed by the detection of a 70% increase in
planktonic biomass. It should be noted that a comparable total leaching of ~0.5 mg/L of
Cu and Fe was detected at all mixed metal loading (Table 5.1). Surface coverage
analysis by confocal laser scanning microscopy (CLSM) (Figure 5.6b) further
confirmed the abovementioned trend from the protein assay, with a 56% overall
reduction in biofilm surface coverage observed for the (0.75+0.75) mg mixed metal
system relative to the control. Note that the CLSM analysis was carried out to account
for potential interference of biofilm EPS in the protein assay.

Chapter 5
132
Figure 5.6 (a) Protein measurements of nitrifying bacteria biomass grown in the presence
of various loading of mixed metal film. All values shown are normalised to the bacteria
biomass grown in the presence of bare film (control, inset). Error bars indicate standard
error between replicates (n = 4); *p ≤ 0.05 against the control. (b) Surface coverage
analysis of nitrifying biofilm. All values shown are normalised to the surface coverage of
control. Error bars indicate standard error between replicates (n = 3); *p ≤ 0.05 against
the control. Confocal laser scanning images of nitrifying biofilm grown in the presence of
(c) bare film, (d) mixed metal film with (0.25+0.25) mg loading, (e) (0.5+0.5) mg loading,
and (f) (0.75+0.75) mg loading.
(a)
(e)
(f)
(b)
(c)
(d)
0.0
0.5
1.0
1.5
2.0*
*
Rela
tive p
rote
in c
oncentr
ation
CuDTTCT+FeDTTCT loading (mg)
Biofilm
Planktonic
*
Control (0.25+0.25) (0.5+0.5) (0.75+0.75)
0
100
200
300
Pro
tein
co
nc
en
tra
tio
n
(µg
/mL
)
Control
0.0
0.5
1.0
1.5
2.0
Rela
tive s
urf
ace c
overa
ge
CuDTTCT+FeDTTCT loading (mg)
Control (0.25+0.25) (0.5+0.5) (0.75+0.75)
*
Control
(0.25+0.25)
(0.5+0.5)
(0.75+0.75)

Chapter 5
133
Table 5.1 Corresponding ICP-OES measurements of leached copper and iron ions from
the mixed metal films after 3 days incubation in 2 mL of nitrifying bacteria culture.
Samples Leached copper
(mg/L)
Leached iron
(mg/L)
%mass of copper
leached out
%mass of iron
leached out
0.25+0.25 0.11 0.34 0.27 0.82
0.5+0.5 0.21 0.28 0.25 0.40
0.75+0.75 0.40 0.12 0.29 0.09
The effect of metals toxicity was also investigated by testing individual copper
or iron films alone (along with the corresponding leached soluble species). Overall, the
presence of up to 2 mg FeDTTCT (with 0.6 mg/L leached Fe, Table 5.2) alone, or up to
2 mg CuDTTCT (with 1.2 mg/L leached Cu, Table 5.3) alone did not alter the extent of
biofilm growth and the presence of planktonic biomass relative to the control (Figure
5.7). Therefore, this ‘dispersal-type’ antibiofilm activity does not originate from metal
toxicity, but most likely result from the presence of NO, which seems to be catalytically
generated by the mixed metal complexes from nitrite endogenously produced by the
AOB biofilm.

Chapter 5
134
Figure 5.7 Protein measurements of nitrifying bacteria biomass grown in the presence of
various loading of (a) FeDTTCT films and (b) CuDTTCT films. All values shown are
normalised to the bacteria biomass grown in the presence of bare film (control, inset).
Error bars indicate standard error between replicates (n = 4); *p ≤ 0.05 against the
control.
Table 5.2 Corresponding ICP-OES measurements of leached iron ions from the FeDTTCT
films after 3 days incubation in 2 mL of nitrifying bacteria culture.
FeDTTCT loading
(mg) Leached iron (mg/L)
%mass of iron
leached out
0.1 0.58 3.37
0.25 0.59 1.37
0.5 0.59 0.69
1 0.52 0.30
2 0.55 0.16
0.0
0.5
1.0
1.5
2.0
2.5
*
Rela
tive p
rote
in c
oncentr
ation
CuDTTCT loading (mg)
Biofilm
Planktonic
Control 0.1 0.25 0.5 1 2
0
100
200
Pro
tein
co
nc
en
tra
tio
n
(µg
/mL
)
Control
0.0
0.5
1.0
1.5
2.0
2.5
Rela
tive p
rote
in c
oncentr
ation
FeDTTCT loading (mg)
Biofilm
Planktonic
Control 0.1 0.25 0.5 1 2
0
100
200
Pro
tein
co
nc
en
tra
tio
n
(µg
/mL
)
Control
(a)
(b)
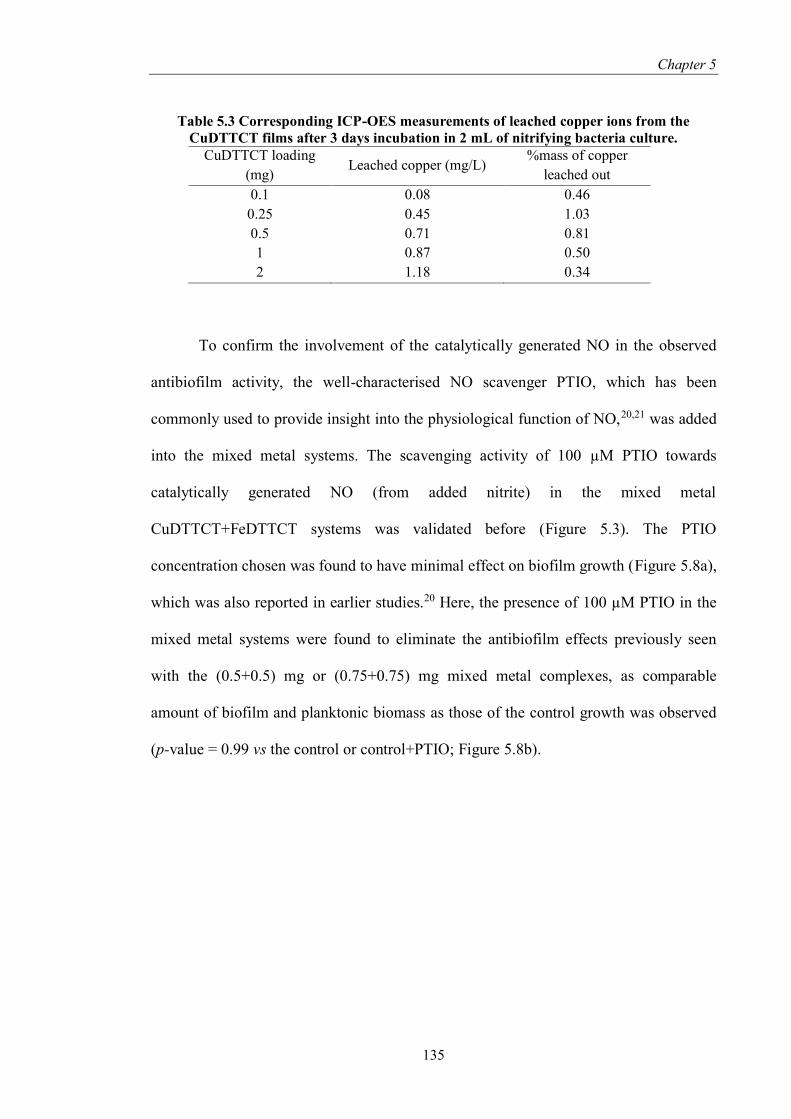
Chapter 5
135
Table 5.3 Corresponding ICP-OES measurements of leached copper ions from the
CuDTTCT films after 3 days incubation in 2 mL of nitrifying bacteria culture.
CuDTTCT loading
(mg) Leached copper (mg/L)
%mass of copper
leached out
0.1 0.08 0.46
0.25 0.45 1.03
0.5 0.71 0.81
1 0.87 0.50
2 1.18 0.34
To confirm the involvement of the catalytically generated NO in the observed
antibiofilm activity, the well-characterised NO scavenger PTIO, which has been
commonly used to provide insight into the physiological function of NO,20,21 was added
into the mixed metal systems. The scavenging activity of 100 µM PTIO towards
catalytically generated NO (from added nitrite) in the mixed metal
CuDTTCT+FeDTTCT systems was validated before (Figure 5.3). The PTIO
concentration chosen was found to have minimal effect on biofilm growth (Figure 5.8a),
which was also reported in earlier studies.20 Here, the presence of 100 µM PTIO in the
mixed metal systems were found to eliminate the antibiofilm effects previously seen
with the (0.5+0.5) mg or (0.75+0.75) mg mixed metal complexes, as comparable
amount of biofilm and planktonic biomass as those of the control growth was observed
(p-value = 0.99 vs the control or control+PTIO; Figure 5.8b).
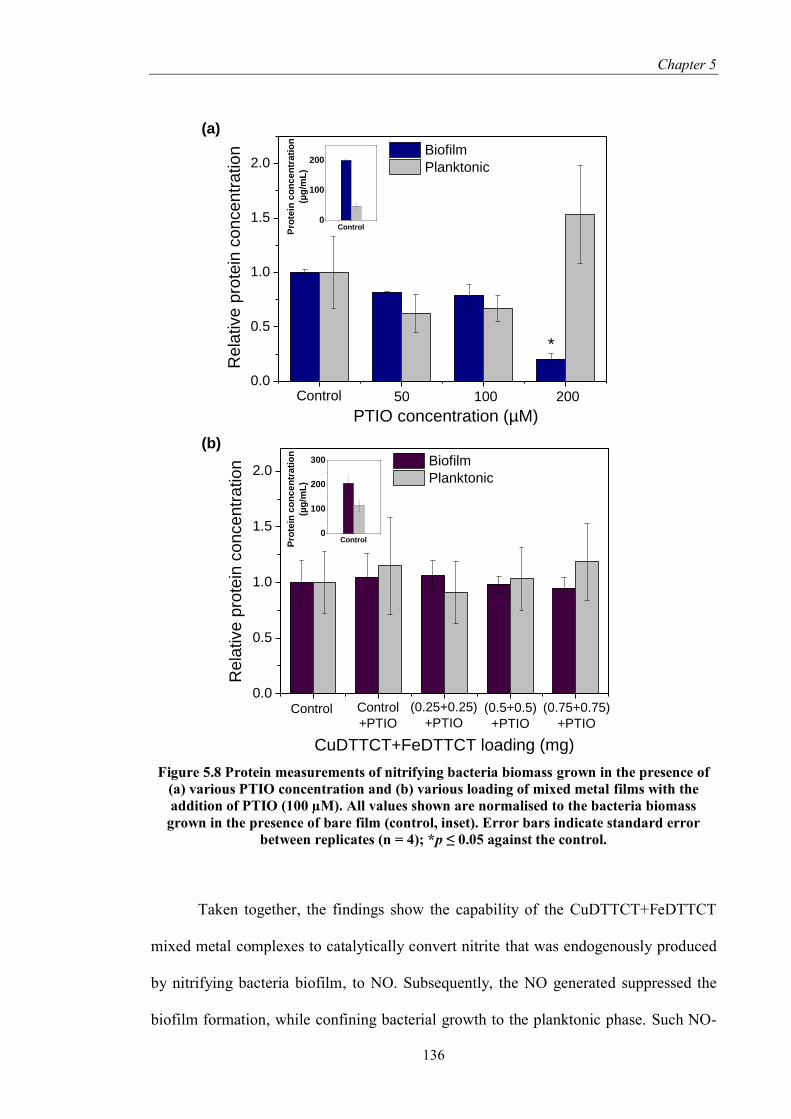
Chapter 5
136
Figure 5.8 Protein measurements of nitrifying bacteria biomass grown in the presence of
(a) various PTIO concentration and (b) various loading of mixed metal films with the
addition of PTIO (100 µM). All values shown are normalised to the bacteria biomass
grown in the presence of bare film (control, inset). Error bars indicate standard error
between replicates (n = 4); *p ≤ 0.05 against the control.
Taken together, the findings show the capability of the CuDTTCT+FeDTTCT
mixed metal complexes to catalytically convert nitrite that was endogenously produced
by nitrifying bacteria biofilm, to NO. Subsequently, the NO generated suppressed the
biofilm formation, while confining bacterial growth to the planktonic phase. Such NO-
0.0
0.5
1.0
1.5
2.0
Re
lative
pro
tein
co
nce
ntr
atio
n
CuDTTCT+FeDTTCT loading (mg)
Biofilm
Planktonic
Control Control
+PTIO
(0.25+0.25)
+PTIO
(0.5+0.5)
+PTIO
(0.75+0.75)
+PTIO
0
100
200
300
Pro
tein
co
nc
en
tra
tio
n
(µg
/mL
)
Control
0.0
0.5
1.0
1.5
2.0
Rela
tive p
rote
in c
oncentr
ation
PTIO concentration (µM)
Biofilm
Planktonic
*
Control 50 100 200
0
100
200
Pro
tein
co
nc
en
tra
tio
n
(µg
/mL
)
Control
(a)
(b)

Chapter 5
137
mediated biofilm dispersal has been reported in a wide spectrum of biofilm-forming
microorganisms.22–25 A common mechanism underlying the transition from biofilm to
planktonic phase is via the NO-regulated alteration of intracellular levels of the
secondary messenger cyclic di-GMP (c-di-GMP), already observed in various bacteria,
including Pseudomonas aeruginosa and Shewanella oneidensis.26–28 In general, low
cellular c-di-GMP levels tend to induce biofilm dispersal, activate cell’s motility and
promote planktonic growth, while an increase in cellular c-di-GMP level promotes cell
attachment to surfaces and biofilm formation.29 In Nitrosomonas europaea, which is
one of the major members of AOB, the link between c-di-GMP signaling and NO-
mediated dispersal has yet been fully established. Instead, NO-mediated dispersal in the
bacteria has been shown to involve up-regulations of 11 proteins, such as motility
(flagellar) and flagellar assembly proteins.30 Due to the involvement of signalling
pathways in NO-mediated dispersal, cellular membrane damage is not expected to
occur, as confirmed by confocal images shown earlier (Figure 5.6c-f, no red (dead) cells
were observed). Such ‘non-toxic’ mode of biofilm dispersal, often associates with a
unique characteristic of NO release, as described in the following.
The antibiofilm effects were observed over relatively long period of 3 days and
given the short half-life of NO in biological systems, this suggests the continual slow
release of low concentrations of NO by the mixed CuDTTCT+FeDTTCT metal
systems. The slow release is a likely consequence of interactions between the redox
cycling of the complexes’ metallic centres and the activity of the slow growing AOB as
nitrite producer. Further, the iron centre could act as an electron mediator that receive
electrons from bacteria,1 such as Bacillus spp (also present in the bacterial inoculum),
which have been shown to utilise (“breathe”) insoluble substrates such as iron(III).31
Such cellular respiration can occur via secretion and subsequent transfer of extracellular

Chapter 5
138
redox metabolites from the bacteria known as electron shuttles, to the iron substrate, or
alternatively can involve direct contact between cell appendages (e.g. pili proteins or
EPS components) with the iron substrate.32 These effects could restart the redox cycling
of iron and copper and subsequently, the generation of NO from nitrite produced by the
AOB. As observed here, the indicated slow release of NO at low concentrations tend to
induce a dispersal effect converting bacteria to the free-floating planktonic phase, which
will otherwise induce lethal effects on biofilms if rapidly generated at high levels.33–35
Due to its non-growth inhibiting or non-cell killing antibiofilm effect, the development
of bacterial resistance to NO is unlikely.36,37 This importantly implies minimal
emergence of highly tolerant biofilm with the use of CuDTTCT+FeDTTCT metal
complexes as NO-based antibiofilm technology.
5.4. Summary
An iron complex (FeDTTCT) was successfully synthesised and incorporated
into PVC matrix together with an active copper species (CuDTTCT) for catalytic
generation of NO from nitrite. Amperometric measurement showed that the mixed
metal complexes (CuDTTCT+FeDTTCT) in nitrite solution was able to generate NO
without additional reducing agent. NO was generated from nitrite through
copper(II)/copper(I) redox cycling, whereby nitrite reduction to NO is via reaction with
copper(I), the latter was formed from reduction of copper(II) facilitated by the iron
complex, as shown by XPS analysis. It was found here that the mixed metal complexes
were capable of converting nitrite that was endogenously produced by nitrifying
bacteria biofilms, into NO. The generated NO subsequently exhibited a ‘dispersal-type’
antibiofilm activity or in other words, suppressing the biofilm formation without

Chapter 5
139
inhibiting planktonic bacterial growth. The involvement of NO in the antibiofilm
activity was validated by the use of NO scavenger, whereby its presence in the mixed
metal system reverted the antibiofilm effects.
5.5. References
1. Zhang, C. et al. Insoluble Fe-humic acid complex as a solid-phase electron
mediator for microbial reductive dechlorination. Environ. Sci. Technol. 48, 6318–
6325 (2014).
2. Welch, K. D., Davis, T. Z., van Eden, M. E. & Aust, S. D. Deleterious Iron-
Mediated Oxidation of Biomolecules. Free Radic. Biol. Med. 32, 577–583
(2002).
3. Liu, Z. D. & Hider, R. C. Design of iron chelators with therapeutic application.
Coord. Chem. Rev. 232, 151–171 (2002).
4. Hwang, S. Nitric Oxide Generating Polymers (NOGPs). (The University of
Michigan, 2007).
5. Breviglieri, S. T., Cavalheiro, E. T. G. & Chierice, G. O. Correlation between
ionic radius and thermal decomposition of Fe (II), Co (II), Ni (II), Cu (II) and Zn
(II) diethanoldithiocarbamates. Thermochim. Acta 356, 79–84 (2000).
6. Yang, Y., Ding, H., Hao, S., Zhang, Y. & Kan, Q. Iron(III), cobalt(II) and
copper(II) complexes bearing 8-quinolinol encapsulated in zeolite-Y for the
aerobic oxidation of styrene. Appl. Organomet. Chem. 25, 262–269 (2011).
7. Saghatforoush, L. A., Aminkhani, A. & Chalabian, F. Iron(III) Schiff base
complexes with asymmetric tetradentate ligands: synthesis, spectroscopy, and
antimicrobial properties. Transit. Met. Chem. 34, 899–904 (2009).
8. Lundberg, J. O., Weitzberg, E. & Gladwin, M. T. The nitrate-nitrite-nitric oxide
pathway in physiology and therapeutics. Nat. Rev. Drug Discov. 7, 156–167

Chapter 5
140
(2008).
9. Opländer, C. et al. Redox-mediated mechanisms and biological responses of
copper-catalyzed reduction of the nitrite ion in vitro. Nitric Oxide 35, 152–164
(2013).
10. McCarthy, C. W., Guillory II, R. J., Goldman, J. & Frost, M. C. Transition Metal
Mediated Release of Nitric Oxide (NO) from S-Nitroso-N-acetylpenicillamine
(SNAP): Potential Applications for Endogenous Release of NO on the Surface of
Stents via Corrosion Products. ACS Appl. Mater. Interfaces 8, 10128–10135
(2016).
11. Bartha, B., Kolbert, Z. & Erdei, L. Nitric oxide production induced by heavy
metals in Brassica juncea L. Czern and Pisum sativum L. Acta Biol. Szeged. 49,
9–12 (2005).
12. Oh, B. K. & Meyerhoff, M. E. Catalytic generation of nitric oxide from nitrite at
the interface of polymeric films doped with lipophilic Cu(II)-complex: a potential
route to the preparation of thromboresistant coatings. Biomaterials 25, 283–293
(2004).
13. Matocha, C. J., Karathanasis, A. D., Rakshit, S. & Wagner, K. M. Reduction of
copper(II) by iron(II). J. Environ. Qual. 34, 1539–1546 (2005).
14. Tao, L. & Li, F. Electrochemical evidence of Fe(II)/Cu(II) interaction on titanium
oxide for 2-nitrophenol reductive transformation. Appl. Clay Sci. 64, 84–89
(2012).
15. González, A. G., Pérez-Almeida, N., Magdalena Santana-Casiano, J., Millero, F.
J. & González-Dávila, M. Redox interactions of Fe and Cu in seawater. Mar.
Chem. 179, 12–22 (2016).
16. Frierdich, A. J. & Catalano, J. G. Fe(II)-mediated reduction and repartitioning of
structurally incorporated Cu, Co, and Mn in iron oxides. Environ. Sci. Technol.
46, 11070–11077 (2012).
17. O’Loughlin, E. J., Kelly, S. D., Kemner, K. M., Csencsits, R. & Cook, R. E.

Chapter 5
141
Reduction of AgI, AuIII, CuII, and Hg II by FeII/FeIII hydroxysulfate green rust.
Chemosphere 53, 437–446 (2003).
18. Grosvenor, A. P., Kobe, B. A., Biesinger, M. C. & McIntyre, N. S. Investigation
of multiplet splitting of Fe 2p XPS spectra and bonding in iron compounds. Surf.
Interface Anal. 36, 1564–1574 (2004).
19. Yamashita, T. & Hayes, P. Analysis of XPS spectra of Fe2+ and Fe3+ ions in
oxide materials. Appl. Surf. Sci. 254, 2441–2449 (2008).
20. Sauder, L. A., Ross, A. A. & Neufeld, J. D. Nitric oxide scavengers differentially
inhibit ammonia oxidation in ammonia-oxidizing archaea and bacteria. FEMS
Microbiol. Lett. 363, 1–7 (2016).
21. Goldstein, S., Russo, A. & Samuni, A. Reactions of PTIO and carboxy-PTIO
with *NO, *NO2, and O2-*. J. Biol. Chem. 278, 50949–50955 (2003).
22. Barraud, N. et al. Nitric oxide-mediated dispersal in single- and multi-species
biofilms of clinically and industrially relevant microorganisms. Microb.
Biotechnol. 2, 370–378 (2009).
23. Craven, M. et al. Nitric Oxide-Releasing Polyacrylonitrile Disperses Biofilms
Formed by Wound-Relevant Pathogenic Bacteria. J. Appl. Microbiol. 120, 1085–
1099 (2016).
24. Arora, D. P., Hossain, S., Xu, Y. & Boon, E. M. Nitric Oxide Regulation of
Bacterial Biofilms. Biochemistry 54, 3717–3728 (2015).
25. Barnes, R. J. et al. Nitric oxide treatment for the control of reverse osmosis
membrane biofouling. Appl. Environ. Microbiol. 81, 2515–2524 (2015).
26. Barraud, N. et al. Nitric oxide signaling in Pseudomonas aeruginosa biofilms
mediates phosphodiesterase activity, decreased cyclic di-GMP levels, and
enhanced dispersal. J. Bacteriol. 191, 7333–7342 (2009).
27. Ha, D.-G. & O’Toole, G. A. c-di-GMP and its Effects on Biofilm Formation and
Dispersion: a Pseudomonas aeruginosa Review. Microbiol. Spectr. 3, MB–
0003–2014 (2015).

Chapter 5
142
28. Plate, L. & Marletta, M. A. Nitric Oxide Modulates Bacterial Biofilm Formation
through a Multicomponent Cyclic-di-GMP Signaling Network. Mol. Cell 46,
449–460 (2012).
29. Liu, S. et al. Understanding, Monitoring and Controlling Biofilm Growth in
Drinking Water Distribution Systems. Environ. Sci. Technol. DOI:
10.1021/acs.est.6b00835 (2016).
30. Schmidt, I., Steenbakkers, P. J. M., op den Camp, H. J. M., Schmidt, K. & Jetten,
M. S. M. Physiologic and Proteomic Evidence for a Role of Nitric Oxide in
Biofilm Formation by Nitrosomonas europaea and Other Ammonia Oxidizers. J.
Bacteriol. 186, 2781–2788 (2004).
31. Nealson, K. H. & Saffarini, D. Iron and manganese in anaerobic respiration:
environmental significance, physiology, and regulation. Annu. Rev. Microbiol.
48, 311–343 (1994).
32. Nealson, K. H. & Finkel, S. E. Electron flow and biofilms. MRS Bull. 36, 380–
384 (2011).
33. Barraud, N., Kelso, M. J., Rice, S. A. & Kjelleberg, S. Nitric Oxide : A Key
Mediator of Biofilm Dispersal with Applications in Infectious Diseases. Curr.
Pharm. Des. 21, 31–42 (2015).
34. Barraud, N. et al. Involvement of nitric oxide in biofilm dispersal of
Pseudomonas aeruginosa. J. Bacteriol. 188, 7344–7353 (2006).
35. Deupree, S. M. & Schoenfisch, M. H. Morphological analysis of the
antimicrobial action of nitric oxide on gram-negative pathogens using atomic
force microscopy. Acta Biomater. 5, 1405–1415 (2009).
36. Privett, B. J., Broadnax, A. D., Bauman, S. J., Riccio, D. A. & Schoenfisch, M.
H. Examination of bacterial resistance to exogenous nitric oxide. Nitric Oxide 26,
169–173 (2012).
37. Njoroge, J. & Sperandio, V. Jamming bacterial communication: New approaches
for the treatment of infectious diseases. EMBO Mol. Med. 1, 201–210 (2009).

143
Chapter 6.
Conclusions and Recommendations
6.1. Conclusions
The research presented herein aims to assess and develop a catalytic system that
can generate nitric oxide (NO) for nitrifying bacteria biofilm control in industrial water
system. A copper complex, namely CuDTTCT, was synthesised and embedded within a
PVC matrix and tested as an NO-generating catalyst in the presence of ascorbic acid as
a reducing agent and nitrite as an NO source. The copper(II) complex was reduced to
copper(I) by ascorbic acid, which subsequently catalysed the reduction of nitrite to NO
while being oxidised back to copper(II). Enhanced and prolonged NO production was
observed when CuDTTCT was used compared to the addition of nitrite-ascorbic acid
alone. The system was found to be effective in suppressing the formation of nitrifying
bacteria biofilm, where it was revealed that the observed biofilm suppression is
predominantly caused by produced NO. Significant cell death was not observed,
indicating that prior to biofilm formation, the NO generated acts as a cell proliferation
inhibitor. Induction of NO production by the copper complex system also triggered the
dispersal of pre-formed biofilms. Additionally, simultaneous biofilm dispersal and
biofilm cells killing were observed when a high concentration of nitrite-ascorbic acid (5
mM) was used along with the CuDTTCT catalyst. Therefore, mechanism of action of
NO is complex and can occur via multiple pathways depending on the biofilm growth

Chapter 6
144
stage. This study highlights the potential of catalytically generated NO for nitrifying
bacteria biofilm control.
In the subsequent study, the ubiquitous Fe2+ ions were employed as an
alternative reducing agent for initiating the copper(II)/copper(I) redox cycling. The
combination of CuDTTCT-nitrite-Fe2+ was found to be capable of NO generation.
However, the amount of NO generated is highly dependent on the Fe2+ speciation in
different pH condition and importantly, buffer composition. The presence of
complexing agents such as phosphate, carbonate, and EDTA was found to be a more
prominent factor in influencing Fe2+ autoxidation compared to pH. Nonetheless, the
system is still effective in generating sufficient NO for biofilm dispersal and reduction
of cell aggregations. Comparing the system used in this study and that used in Chapter
3, these result thus imply that biofilm dispersal mediated by NO is not affected by the
selection of reducing agent. Importantly, buffer condition was also found to have
minimal impact on NO-mediated dispersal, thus it may have a higher possibility of
success when applied in real water condition. The findings of this study hence
emphasised the robustness of NO-mediated dispersal method for biofilm control.
In order to eliminate the need for the addition of a reducing agent in solution
form, an iron complex, namely FeDTTCT, was synthesised and incorporated into the
PVC matrix with the CuDTTCT catalyst. FeDTTCT was shown to facilitate
copper(II)/copper(I) redox cycling, thus expediting NO generation from nitrite without
the need for additional reducing agent. Moreover, the mixed metal complexes system
was capable of suppressing biofilm formation while confining bacteria growth to the
free-floating planktonic phase over three days incubation, which suggests the
conversion of nitrite endogenously produced by the nitrifying bacteria to NO.
Importantly, this ‘dispersal-type’ antibiofilm activity does not involve cell inhibition or

Chapter 6
145
cell killing pathway, and therefore could minimise the possible emergence of bacterial
resistance. The involvement of NO in exerting antibiofilm activity was confirmed by the
addition of an NO scavenger, whereby its presence with the mixed metal system
reverted the antibiofilm effect. This work highlights the development of self-sustained
metal-based material that could be potentially applied in industrial water systems.
6.2. Recommendations
As mentioned earlier, the presence of reducing agents is paramount in the
copper(II)/copper(I) redox cycling for NO generation. The studies presented here show
the use of ascorbic acid, Fe2+ ions, and iron complex as reducing agents. In addition to
the above, there are numerous attractive reducing agent candidates that can be tested.
For instance, Ren et al. reported the use of cathodic potentials as an alternative means to
reduce copper(II)-tri(2-pyridylmethyl)amine (Cu-TPMA) within silicone rubber
catheters to generate NO from a nitrite reservoir.1,2 Moreover, it was found that
copper(I) species can be generated from the reduction of copper(II) species by hydrogen
peroxide, which is produced from photochemical reactions of organic compounds in
natural waters.3,4 Other possible reducing agents include hydroxylamine5 and glucose.6
The study on effectiveness and suitability of different reducing agents is crucial to assist
in understanding the selection criteria for suitable reducing agents for
copper(II)/copper(I) redox cycling.
The study presented here also proved that NO can trigger biofilm dispersal on
the model organism nitrifying bacteria employed here. Although it was mentioned that
regulation of motility expression has been linked as the underlying mechanism involved

Chapter 6
146
for the switch between biofilm to planktonic phase in Nitrosomonas europaea,7 which is
one of the major member of ammonia oxidisers, the underlying mechanism for the test
bacteria used in this study, specifically Nitrosospira multiformis, Nitrospira marina, and
Bacillus sp. has yet to be clarified. Proteomic approach, which has been used by
Schmidt et al. to investigate protein regulation in different phase of bacteria cells,7 can
be used as the first step towards in-depth understanding of NO-mediated dispersal in the
test organisms used in study. Moreover, measurement of the intracellular level of cyclic-
di-GMP is suggested, as it is believed that this secondary messenger molecule is the
central mechanism involved in regulating biofilm dispersal in many bacterial species.8–
10
In this study, nitrifying bacteria are chosen because they are commonly found in
chloraminated water.11 While this study is aimed for the application in various industrial
water system, biofilm-forming bacteria can go beyond nitrifying bacteria group. For
instance, Legionella is commonly found in air conditioners, water systems, cooling
towers, water fountains, and hospital equipments.12 In membrane systems,
Sphingomonas,13 Acinetobacter,14 and Staphylococcus14 have been identified as the
pioneer biofilm-forming bacteria. Moreover, anaerobic bacteria such as Desulfovibrio
would be the dominant species in water systems where oxygen is not readily available.
Therefore, it is suggested that biofilm control by catalytically generated NO can be
tested against wider water-related bacterial consortiums. Additionally, antibiofilm
activity testing against natural occurring microbial consortiums formed from real water
sample would be of interest.
The study presented in Chapter 5 shows the use of a mixed metal-based material
for biofilm control for a period of three days. However, studies have reported that
increased biofilm resistance is observed in mature/thicker biofilms than early

Chapter 6
147
stage/thinner biofilms.15 Therefore, investigation over longer incubation periods (up to
one week) is recommended. In addition, continuous systems that will allow thicker
biofilm formation, such as flow cells, can be used to study the impact of biofilm
thickness (maturity) against the mixed metal system effectivity.
Although the mixed metal system has been proven to be effective in suppressing
nitrifying biofilm formation in a non-toxic manner, the underlying mechanism, i.e. the
NO release profile and NO effective concentration is yet to be established. The widely
employed chemiluminescence for prolonged measurement of NO can be used to
understand NO release profile from the mixed metal system during the incubation
period.16 The effective concentration of NO that affected the cells can be measured by
employing the intracellular NO-sensitive dye, 4,5-diaminofluorescein diacetate (DAF-2
DA).17,18 Additionally, in-depth understanding on the postulated mechanism for
prolonged NO mechanism presented in Chapter 5, whereby the oxidised iron complex
was reduced via extracellular microbial respiration, can be investigated. The above
knowledge will allow better understanding towards the applicability of the mixed metal
catalyst system for long-term application.
Barraud et al. reported that biofilms which receive treatment with NO donors
exhibit higher sensitivity towards chlorine.19 Thus, investigation on the combined
biofilm removal efficiency of catalytically generated NO with chlorine or chloramine is
also suggested. It should be noted that chlorine/chloramine, which are oxidants, can
neutralise any reducing agents available in water. Therefore, the study on the mixed
metal system developed in Chapter 5 can be extended with this investigation.
For future applications, it is important to study industrial scale production of
NO-generating materials which does not involve the use of organic solvent as shown in

Chapter 6
148
this thesis. Consequently, the production of PVC-copper complex material by the
commonly used extrusion method is recommended. Because a simple physical mixing
process between the PVC and the metal complexes will be sufficient for the
incorporation of the metal complexes, the cost associated with the synthesis of PVC
containing metal complex is expected to be minimal. Nevertheless, economic
assessment regarding this process is needed.
Future research also should be directed towards the design of novel ligands that
can increase the stability of the metal centre to prevent metal ion leaching (as mentioned
in Chapter 3) and prevent aggregation of the metal complexes (especially iron complex)
in PVC matrix. Moreover, novel ligand design that can ‘mask’ copper species de-
stabilising influence towards the polymeric chain without affecting the
copper(II)/copper(I) redox activity. This research could also be expanded with studies
on other transition metals that can catalytically reduce nitrite to NO and have higher
compatibility with PVC. For instance, cobalt, nickel, or zinc have been found to mediate
NO generation from S-nitroso-N-acetylpenicillamine (SNAP; a synthetic NO donor).20
In the long term, the materials developed in this study can be easily adapted to
different applications with minor adjustments. For instance, in clinical settings, the use
of glutathione (a reducing agent commonly available in plasma and blood) to reduce the
copper complex used in this study can be investigated. It is also recommended to test
NO-generating materials effectivity against some targeted pathogens such as
Pseudomonas aeruginosa, Staphylococcus aureus, and Escherichia coli.

Chapter 6
149
6.3. References
1. Ren, H. et al. Electrochemically modulated nitric oxide (NO) releasing
biomedical devices via copper(II)-Tri(2-pyridylmethyl)amine mediated reduction
of nitrite. ACS Appl. Mater. Interfaces 6, 3779–3783 (2014).
2. Ren, H. et al. Thromboresistant/anti-biofilm catheters via electrochemically
modulated nitric oxide release. Bioelectrochemistry 104, 10–16 (2015).
3. Millero, F. J., Sharma, V. K. & Karn, B. The rate of reduction of copper(II) with
hydrogen peroxide in seawater. Mar. Chem. 36, 71–83 (1991).
4. Millero, F. J., Johnson, R. L., Vega, C. A., Sharma, V. K. & Sotolongo, S. Effect
of Ionic Interactions on the Rates of Reduction of Cu(II) with H2O2 in Aqueous
Solutions. J. Solution Chem. 21, 1271 – 1287 (1993).
5. Lee, H. et al. Activation of Oxygen and Hydrogen Peroxide by Copper(II)
Coupled with Hydroxylamine for Oxidation of Organic Contaminants. Environ.
Sci. Technol. acs.est.6b02067 (2016). doi:10.1021/acs.est.6b02067
6. Jin, M. et al. Shape-controlled synthesis of copper nanocrystals in an aqueous
solution with glucose as a reducing agent and hexadecylamine as a capping
agent. Angew. Chemie - Int. Ed. 50, 10560–10564 (2011).
7. Schmidt, I., Steenbakkers, P. J. M., op den Camp, H. J. M., Schmidt, K. & Jetten,
M. S. M. Physiologic and Proteomic Evidence for a Role of Nitric Oxide in
Biofilm Formation by Nitrosomonas europaea and Other Ammonia Oxidizers. J.
Bacteriol. 186, 2781–2788 (2004).
8. Ha, D.-G. & O’Toole, G. A. c-di-GMP and its Effects on Biofilm Formation and
Dispersion: a Pseudomonas aeruginosa Review. Microbiol. Spectr. 3, MB–
0003–2014 (2015).
9. Barraud, N., Kjelleberg, S. & Rice, S. A. Dispersal from Microbial Biofilms.
Microbiol. Spectr. 3, MB–0015–2014 (2015).
10. Caly, D. & Bellini, D. Targeting Cyclic di-GMP Signalling: A Strategy to

Chapter 6
150
Control Biofilm Formation? Curr. Pharm. Des. 21, 12–24 (2015).
11. Martiny, A. C., Albrechtsen, H.-J., Arvin, E. & Molin, S. Identification of
Bacteria in Biofilm and Bulk Water Samples from a Nonchlorinated Model
Drinking Water Distribution System: Detection of a Large Nitrite-Oxidizing
Population associated with Nitrospira spp. Appl. Environ. Microbiol. 71, 8611–
8617 (2005).
12. Coniglio, M. A., Melada, S. & Yassin, M. H. Monochloramine for controlling
Legionella in biofilms : how much we know ? J. Nat. Sci. 1, e44 (2015).
13. Bereschenko, L. A., Stams, A. J. M., Euverink, G. J. W. & Van Loosdrecht, M.
C. M. Biofilm formation on reverse osmosis membranes is initiated and
dominated by Sphingomonas spp. Appl. Environ. Microbiol. 76, 2623–2632
(2010).
14. Zhang, K., Choi, H., Dionysiou, D. D., Sorial, G. a. & Oerther, D. B. Identifying
pioneer bacterial species responsible for biofouling membrane bioreactors.
Environ. Microbiol. 8, 433–440 (2006).
15. Liu, S. et al. Understanding, Monitoring and Controlling Biofilm Growth in
Drinking Water Distribution Systems. Environ. Sci. Technol. DOI:
10.1021/acs.est.6b00835 (2016).
16. Hunter, R. a, Storm, W. L., Coneski, P. N. & Schoenfisch, M. H. Inaccuracies of
nitric oxide measurement methods in biological media. Anal. Chem. 85, 1957–63
(2013).
17. Sun, B., Slomberg, D. L., Chudasama, S. L., Lu, Y. & Schoenfisch, M. H. Nitric
oxide-releasing dendrimers as antibacterial agents. Biomacromolecules 13, 3343–
3354 (2012).
18. Slomberg, D. L. et al. Role of size and shape on biofilm eradication for nitric
oxide-releasing silica nanoparticles. ACS Appl. Mater. Interfaces 5, 9322–9329
(2013).
19. Barraud, N. et al. Nitric oxide-mediated dispersal in single- and multi-species

Chapter 6
151
biofilms of clinically and industrially relevant microorganisms. Microb.
Biotechnol. 2, 370–378 (2009).
20. McCarthy, C. W., Guillory II, R. J., Goldman, J. & Frost, M. C. Transition Metal
Mediated Release of Nitric Oxide (NO) from S-Nitroso-N-acetylpenicillamine
(SNAP): Potential Applications for Endogenous Release of NO on the Surface of
Stents via Corrosion Products. ACS Appl. Mater. Interfaces 8, 10128–10135
(2016).




















