
Cytomegalovirus-Specific T Cell Immunity Is Maintained in Immunosenescent Rhesus Macaques
11
The Journal of Immunology Cytomegalovirus-Specific T Cell Immunity Is Maintained in Immunosenescent Rhesus Macaques Luka C ˇ ic ˇin-S ˇ ain,* ,†,1 Andrew W. Sylwester,* ,‡,1 Shoko I. Hagen,* ,‡ Don C. Siess, ‡ Noreen Currier,* Alfred W. Legasse, ‡ Miranda B. Fischer,* Caroline W. Koudelka, x Michael K. Axthelm,* ,‡ Janko Nikolich-Z ˇ ugich, {,‖ and Louis J. Picker* ,† Although CMV infection is largely benign in immunocompetent people, the specific T cell responses associated with control of this persistent virus are enormous and must be maintained for life. These responses may increase with advanced age and have been linked to an “immune risk profile” that is associated with poor immune responsiveness and increased mortality in aged individ- uals. Based on this association, it has been suggested that CMV-specific T cell responses might become dysfunctional with age and thereby contribute to the development of immune senescence by homeostatic disruption of other T cell populations, diminished control of CMV replication, and/or excess chronic inflammation. In this study, we use the rhesus macaque (RM) model of aging to ask whether the quantity and quality of CMV-specific T cell responses differ between healthy adult RMs and elderly RMs that manifest hallmarks of immune aging. We demonstrate that the size of the CD4 + and CD8 + CMV-specific T cell pools are similar in adult versus old RMs and show essentially identical phenotypic and functional characteristics, including a dominant effector memory phenotype, identical patterns of IFN-g, TNF-a, and IL-2 production and cytotoxic degranulation, and comparable functional avidities of optimal epitope-specific CD8 + T cells. Most importantly, the response to and protection against an in vivo CMV challenge were identical in adult and aged RMs. These data indicate that CMV-specific T cell immunity is well maintained in old RMs and argue against a primary role for progressive dysfunction of these responses in the development of immune senescence. The Journal of Immunology, 2011, 187: 1722–1732. A ging may be accompanied with a decline in immune function characterized by poor responses to vaccination and increased morbidity and mortality from infectious diseases (1–4). This functional decline is associated with complex, but characteristic, changes in both the innate and adaptive immune system, collectively referred to as “immune senescence” (5, 6). Among the most consistent and dramatic age-related changes are those that occur in T cell homeostasis and function, manifesting in blood as 1) decreased CD4/CD8 T cell ratios, 2) loss of naive cells and relative expansion of differentiated effector memory (EM) cells, 3) oligoclonality/clonal expansions, 4) poor proliferative responses, and 5) changes in cytokine secretion patterns (5, 7–12). These immunologic changes, which typically occur coordinately, have also been strongly correlated with evidence of persistent infection with the ubiquitous b-herpesvirus CMV, and together, these features constitute an immune risk profile (IRP) that in some studies has been predictive of increased mortality in aged indi- viduals (7, 10–16). These associations have led to the hypothesis that immune se- nescence may be infectious—a consequence of long-term expo- sure to, and immunologic control of, persistent infections, in particular, CMV (7, 13, 15, 17). CMV is among the most immu- nogenic of known viruses, eliciting stable frequencies of specific T cells in CMV + adults that average 10% of both the CD4 + and CD8 + memory compartments in blood (18). Frequencies of CMV- specific T cells can be even higher in aged individuals (19–21), and given that CMV-specific T cell responses are characterized by 1) a dominant EM (CD28 2 , CD27 2 , CCR7 2 ) phenotype, 2) func- tional characteristics commensurate with this phenotype (high effector cytokine production, relatively low IL-2 production, ex- pression of cytotoxic apparatus, poor in vitro proliferation), and 3) highly hierarchical clonotypic repertoires (with the top clonotypes manifesting frequencies .1%), these responses clearly underlie many of the features of the IRP (7, 13, 15, 17, 22–28). Indeed, it has been postulated that an increasingly dysfunctional, proin- flammatory, CMV-specific T cell response expands with advanced age, driving out other T cell populations and causing both the IRP and a significant component of age-associated immune deficiency (7, 13, 17, 29). However, overt CMV disease is rare in the elderly (30, 31), healthy aged individuals may also be CMV + (32, 33), and other age-related mechanisms such as thymic involution and cessation of new T cell production clearly play a major role in naive cell deficiency (17, 34, 35) and, likely, the poor re- sponsiveness of the elderly to new Ags. Thus, the large CMV- specific effector memory response may simply be better pre- *Vaccine and Gene Therapy Institute, Oregon Health & Science University, Beaver- ton, OR 97006; † Department of Vaccinology and Applied Microbiology, Helmoltz Centre for Infection Research, Braunschweig, Germany; ‡ Oregon National Primate Research Center, Oregon Health & Science University, Beaverton, OR 97006; x Bios- tatistics Shared Resource, Knight Cancer Institute at the Oregon Health & Science University, Portland, OR 97239; { Department of Immunobiology, University of Ari- zona College of Medicine, Tucson, AZ 85724; and ‖ Arizona Center on Aging, Uni- versity of Arizona College of Medicine, Tucson, AZ 85724 1 L.C ˇ .-S ˇ . and A.W.S. contributed equally to this work. Received for publication February 23, 2011. Accepted for publication June 6, 2011. This work was supported by National Institutes of Health Awards P01 AG023664 and R01 AI082529 (to L.J.P. and J.N.-Z ˇ .) and by the Oregon National Primate Research Center National Institutes of Health Core Grant P51 RR00163. Address correspondence and reprint requests to Dr. Louis J. Picker, Vaccine and Gene Therapy Institute, Oregon Health & Science University–West Campus, 505 NW 185th Avenue, Beaverton, OR 97006. E-mail address: [email protected] Abbreviations used in this article: BAL, bronchoalveolar lavage lymphocyte; CM, central memory; EM, effector memory; HCMV, human cytomegalovirus; IE, imme- diate early; IRP, immune risk profile; RhCMV, rhesus cytomegalovirus; RM, rhesus macaque; SEB, Staphylococcus enterotoxin B; TransEM, transitional effector mem- ory. Copyright Ó 2011 by The American Association of Immunologists, Inc. 0022-1767/11/$16.00 www.jimmunol.org/cgi/doi/10.4049/jimmunol.1100560
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Cytomegalovirus-Specific T Cell Immunity Is Maintained in Immunosenescent Rhesus Macaques

The Journal of Immunology
Cytomegalovirus-Specific T Cell Immunity Is Maintained inImmunosenescent Rhesus Macaques
Luka Cicin-Sain,*,†,1 Andrew W. Sylwester,*,‡,1 Shoko I. Hagen,*,‡ Don C. Siess,‡
Noreen Currier,* Alfred W. Legasse,‡ Miranda B. Fischer,* Caroline W. Koudelka,x
Michael K. Axthelm,*,‡ Janko Nikolich-Zugich,{,‖ and Louis J. Picker*,†
Although CMV infection is largely benign in immunocompetent people, the specific T cell responses associated with control of this
persistent virus are enormous and must be maintained for life. These responses may increase with advanced age and have been
linked to an “immune risk profile” that is associated with poor immune responsiveness and increased mortality in aged individ-
uals. Based on this association, it has been suggested that CMV-specific T cell responses might become dysfunctional with age and
thereby contribute to the development of immune senescence by homeostatic disruption of other T cell populations, diminished
control of CMV replication, and/or excess chronic inflammation. In this study, we use the rhesus macaque (RM) model of aging to
ask whether the quantity and quality of CMV-specific T cell responses differ between healthy adult RMs and elderly RMs that
manifest hallmarks of immune aging. We demonstrate that the size of the CD4+ and CD8+ CMV-specific T cell pools are similar in
adult versus old RMs and show essentially identical phenotypic and functional characteristics, including a dominant effector
memory phenotype, identical patterns of IFN-g, TNF-a, and IL-2 production and cytotoxic degranulation, and comparable
functional avidities of optimal epitope-specific CD8+ T cells. Most importantly, the response to and protection against an
in vivo CMV challenge were identical in adult and aged RMs. These data indicate that CMV-specific T cell immunity is well
maintained in old RMs and argue against a primary role for progressive dysfunction of these responses in the development of
immune senescence. The Journal of Immunology, 2011, 187: 1722–1732.
Aging may be accompanied with a decline in immunefunction characterized by poor responses to vaccinationand increased morbidity and mortality from infectious
diseases (1–4). This functional decline is associated with complex,but characteristic, changes in both the innate and adaptive immunesystem, collectively referred to as “immune senescence” (5, 6).Among the most consistent and dramatic age-related changes arethose that occur in T cell homeostasis and function, manifesting inblood as 1) decreased CD4/CD8 T cell ratios, 2) loss of naive cellsand relative expansion of differentiated effector memory (EM)cells, 3) oligoclonality/clonal expansions, 4) poor proliferativeresponses, and 5) changes in cytokine secretion patterns (5, 7–12).
These immunologic changes, which typically occur coordinately,have also been strongly correlated with evidence of persistentinfection with the ubiquitous b-herpesvirus CMV, and together,these features constitute an immune risk profile (IRP) that in somestudies has been predictive of increased mortality in aged indi-viduals (7, 10–16).These associations have led to the hypothesis that immune se-
nescence may be infectious—a consequence of long-term expo-sure to, and immunologic control of, persistent infections, inparticular, CMV (7, 13, 15, 17). CMV is among the most immu-nogenic of known viruses, eliciting stable frequencies of specificT cells in CMV+ adults that average 10% of both the CD4+ andCD8+ memory compartments in blood (18). Frequencies of CMV-specific T cells can be even higher in aged individuals (19–21),and given that CMV-specific T cell responses are characterized by1) a dominant EM (CD282, CD272, CCR72) phenotype, 2) func-tional characteristics commensurate with this phenotype (higheffector cytokine production, relatively low IL-2 production, ex-pression of cytotoxic apparatus, poor in vitro proliferation), and 3)highly hierarchical clonotypic repertoires (with the top clonotypesmanifesting frequencies .1%), these responses clearly underliemany of the features of the IRP (7, 13, 15, 17, 22–28). Indeed,it has been postulated that an increasingly dysfunctional, proin-flammatory, CMV-specific T cell response expands with advancedage, driving out other T cell populations and causing both the IRPand a significant component of age-associated immune deficiency(7, 13, 17, 29). However, overt CMV disease is rare in the elderly(30, 31), healthy aged individuals may also be CMV+ (32, 33), andother age-related mechanisms such as thymic involution andcessation of new T cell production clearly play a major role innaive cell deficiency (17, 34, 35) and, likely, the poor re-sponsiveness of the elderly to new Ags. Thus, the large CMV-specific effector memory response may simply be better pre-
*Vaccine and Gene Therapy Institute, Oregon Health & Science University, Beaver-ton, OR 97006; †Department of Vaccinology and Applied Microbiology, HelmoltzCentre for Infection Research, Braunschweig, Germany; ‡Oregon National PrimateResearch Center, Oregon Health & Science University, Beaverton, OR 97006; xBios-tatistics Shared Resource, Knight Cancer Institute at the Oregon Health & ScienceUniversity, Portland, OR 97239; {Department of Immunobiology, University of Ari-zona College of Medicine, Tucson, AZ 85724; and ‖Arizona Center on Aging, Uni-versity of Arizona College of Medicine, Tucson, AZ 85724
1L.C.-S. and A.W.S. contributed equally to this work.
Received for publication February 23, 2011. Accepted for publication June 6, 2011.
This work was supported by National Institutes of Health Awards P01 AG023664 andR01 AI082529 (to L.J.P. and J.N.-Z.) and by the Oregon National Primate ResearchCenter National Institutes of Health Core Grant P51 RR00163.
Address correspondence and reprint requests to Dr. Louis J. Picker, Vaccine and GeneTherapy Institute, Oregon Health & Science University–West Campus, 505 NW185th Avenue, Beaverton, OR 97006. E-mail address: [email protected]
Abbreviations used in this article: BAL, bronchoalveolar lavage lymphocyte; CM,central memory; EM, effector memory; HCMV, human cytomegalovirus; IE, imme-diate early; IRP, immune risk profile; RhCMV, rhesus cytomegalovirus; RM, rhesusmacaque; SEB, Staphylococcus enterotoxin B; TransEM, transitional effector mem-ory.
Copyright� 2011 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/11/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1100560

served than other T cell populations, persisting while other pop-ulations decline (36).The issue of whether CMV infection plays a causal role in im-
mune senescence and, if so, the understanding of the mechanism(s)by which this virus manifests these changes has critical implica-tions for the clinical approach to elderly individuals. Obviously,if CMV infection and/or the CMV-specific T cell response areculpable in the development of immune senescence, therapy forimmune senescence should be directed at preventing or treatingCMV infection or interfering with the mechanism by which CMVand/or CMV-specific T cell responses cause the deleteriouschanges. Alternatively, if CMV and the CMV-specific T cell re-sponse are simply bystanders to the pathogenesis of functionalimmune senescence, therapy should be directed toward the non–CMV-related causal mechanisms. When manifest in middle-agedadults, the IRP does not portend a poor prognosis (37), suggestinga progressive process that develops in late middle age to advancedage. One possibility is that the CMV-specific T cell responsedeteriorates during this time frame, undergoing slow, but progressive,dysfunction, allowing more frequent and/or higher-magnitude epi-sodes of viral replication. This antigenic stimulation might ex-pand the CMV-specific response and elicit an ever-increasing levelof dysfunction by exhausting any remaining nonsenescentT cell populations and/or stimulating production of dysfunctionalprogeny. Such a reinforcing cycle of viral replication and dys-function might then cause generalized immune dysfunction bydisplacing naive and/or central memory T cell populations (13)and/or by creating a chronic inflammatory response that nega-tively affects the homeostasis of these other crucial T cell pop-ulations (38).To address the question of whether CMV-specific T cell re-
sponses deteriorate with age and lose protective capacity, weturned to the rhesus macaque (RM) model. This model manifeststhe major features of immune senescence found in humans (35,39–42), as well as a highly homologous rhesus cytomegalovirus(RhCMV) that manifests similar immunobiology as human cyto-megalovirus (HCMV), including the same high-frequency EM-biased CD4+ and CD8+ T cell responses observed in CMV+
people (41, 43–46). Indeed, if chronic viral Ag exposure underliesdeterioration of the CMV-specific T cell responses and ultimatelythe overall T cell compartment, the RM model might be expectedto highlight this pathogenic mechanism as all colony-raisedRMs become CMV+ in the first year of life, and RhCMV, unlikeHCMV [which is shed intermittently in the chronic phase of infec-tion (47–49)], is continually shed in the urine and saliva of thevast majority, if not all, animals (44, 46, 50). In this study, wecompare in detail the CMV-specific CD4+ and CD8+ T cell respon-ses in adult and aged RMs and demonstrate that both the ex vivoand in vivo function of these responses are unchanged withadvanced age, even when other features of immune senescenceare present. These results contradict the occurrence of a generaldeterioration of CMV-specific EM T cell function with advancedage and argue against the hypothesis that such age-associateddeterioration is a requirement for development of immune senescence.
Materials and MethodsAnimals
Colony-bred male and female RMs of Indian origin were maintainedaccording to the federal, state, and local guidelines. All experiments wereapproved by the Institutional Animal Care and Use Committee at theOregon National Primate Research Center. RMs with tumors, amyloidosis,or signs of significant clinical disease at the time of initial assignment wereexcluded from the study. The overall cohort consisted of 21 young adult(average = 7.8 y, range 7–10 y; 12 males, 9 females) and 36 old (average =
22.8 y, range 19–26 y, 15 males, 21 females) RMs, which naturally ac-quired RhCMV in their first year of life. The in vivo challenge assay wasperformed on a subset of this overall cohort consisting of 11 young adult(average = 8.5 y, range 7–10 y; 6 males, 5 females) and 9 old (average =23.2 y, range 20–27 y; 4 males, 5 females) RMs. Pooled data from multipleindependent experiments is shown throughout. The number of RMs used inany particular analysis was a variable subset of the overall cohort, de-pendent on the availability of RMs at the time of analysis or (where in-dicated) on defined inclusion criteria applied to the entire RM cohort.Bronchoalveolar lavage lymphocyte (BAL) collection and sampling wasperformed as previously described (41). In the RhCMV superinfectionexperiments, RMs were challenged by s.c. injection of 107 PFU RhCMVstrain 68.1, as previously described (44).
Ags and Abs
RhCMV-specific CD4+ T cell responses were identified by the response toRhCMV (Cercopithecine herpesvirus 8) Ag preparations containing lysatesof RhCMV (strain 68.1; ATCC VR-677)-infected monolayers of telo-merized RM fibroblasts (44) combined with cell-free viral proteins con-centrated from the culture medium of these infected cells (2:1 ratio oflysate to supernatant concentrate). Proteins were concentrated from in-fected cell supernatants by precipitation with 90% saturated ammoniumsulfate, centrifugation (10,000 3 g, 30 min, 4˚C), resuspension in PBS,and finally, extensive dialysis against PBS (10,000 Da molecular massexclusion). RhCMV-specific CD8+ T cell responses were identified by theresponse to consecutive, overlapping 15-mer peptide pools (11-aa overlap)spanning the entire sequences of immediate-early (IE)-1 and IE-2 RhCMVgenes (Intavis, Reutlingen, Germany). The final concentration of eachpeptide in the pool was 2 mg/test. The mAbs L200 (CD4; AmCyan,PerCP–Cy5.5), SP34-2 (CD3; Pacific Blue, Alexa 700), SK-1 (CD8a;PerCP–Cy5.5, PE, unconjugated), CD28.2 (CD28; PE-TexasRed, PerCP–Cy5.5, unconjugated), DX2 (CD95; PE–Cy7, allophycocyanin, PE), 15053(CCR7; FITC), B56 (Ki-67; FITC, PE), FN50 (CD69; PE, PE-TexasRed),9F10 (CD49d; unconjugated), MAB11 (TNF-a; FITC, PE–Cy7), H3A3(CD107a; FITC), H4B4 (CD107b; FITC), B27 (IFN-g; allophycocyanin),and MQ1-17H12 (IL-2; PE) were obtained from BD Biosciences. mAbFN18 (CD3) was produced in-house and conjugated to Pacific blue orAlexa 700 using a conjugation kit from Life Technologies.
Cell preparation and stimulation
Citrated venous blood was centrifuged on a Ficoll-Hypaque density gra-dient (Sigma-Aldrich) to separate PBMCs. A small aliquot (100ml) of wholeblood was used at each bleed to define absolute lymphocyte counts in anAc*T 5diff hematologic analyzer (Beckman-Coulter). PBMCs and BALswere washed in HBSS and resuspended in RPMI 1640 medium supple-mented with 10% heat-inactivated FCS (HyClone), 2 mM L-glutamine(Sigma-Aldrich), 1 mM Na-pyruvate (Sigma-Aldrich), and 50 mM 2-mercaptoethanol (Sigma-Aldrich) (R10). To evaluate RhCMV-specificCD4+ T cell responses, 106 cells in 1 ml R10 were placed in 17 mm 3 100mm polystyrene tubes precoated with goat Fab fragments (2.5 mg/ml)against mouse IgG (H+L), and then coated with costimulatory CD28and CD49d Abs (2.5 mg/ml of each). Precoating and coating were done onstimulation day, in PBS, at 37˚C for 1 h, with PBS washes between pro-cedures and after coating. Six microliters of RhCMV lysate was added for6 h total, in the presence of brefeldin A (10 mg/ml) for the last 5 h.Stimulation with 15-mer peptide pools was performed in the same volumeformat, but in polypropylene tubes. Costimulatory Abs were added directlyin the tube (at 0.5 mg/ml each). To evaluate dynamic responses uponin vivo challenge, these assays were adapted to a 96-well plate format,where 5 3 105 cells in 200 ml R10 were stimulated with lysate or 15-merpeptide pools for 9 h (last 8 h in presence of brefeldin A), and costimu-lation as above. In all cases, a negative control tube that contained mediaand costimulatory Abs was set up in parallel.
Immunofluorescence staining and flow cytometric analysis
Flow cytometric analysis was performed on an LSR-II apparatus (BDBiosciences) with Pacific blue, AmCyan, FITC, PE, PE-TexasRed, PerCP–Cy5.5, PE–Cy7, allophycocyanin, Alexa 700, and allophycocyanin–Cy7 asthe available fluorescent channels. Analysis was done with FlowJo soft-ware (Tree Star, Ashland, OR). In all cases, gating on the lymphocytepopulation was followed by the separation of the CD3+ T cell subset andprogressive gating on CD4+ and CD8+ T cell subsets. Naive cells weredefined by CD28 versus CD95 expression patterns, as previously described(41). Among memory T cells, quadrant analysis of the CD28 and CCR7parameters defined the central, transitional effector, and effector andmemory subsets, as described (51). For the detection of Ag-responding
The Journal of Immunology 1723

subsets, we gated the CD41 or CD81 subsets on intracellular expression ofCD69 and any of the IFN-g, TNF-a, or IL-2 cytokines (46). Boolean gatesof any of these combinations were generated, and in select experiments,cells responding by production of any of the cytokines were phenotypedwith respect to CD28 and CCR7 expression, as described (46). For thedetection of degranulation responses to Ag stimulation, CD107a andCD107b Abs were added to the cells during the stimulation assay, so as todetect externalization of cytotoxic granule membrane proteins upongranule fusion with the cell membrane (25).
Functional avidity assessment
Immunodominant peptides indicated in Table I were used at graded con-centrations from 100 mg/ml to 1 pg/ml to stimulate PBMCs for 6 h (last 5 hin presence of brefeldin A). IFN-g, TNF-a, or CD107a/b responses weremeasured as detailed earlier and normalized to maximum response togenerate dose-response curves and calculate peptide concentrations cor-responding with 50% of maximum response.
RhCMV quantification using real-time PCR
RhCMVDNA in blood or BAL samples was quantified by quantitative PCRamplification of a segment of the RhCMV IE-2 gene, as previously de-scribed (44). Nucleic acid was purified from samples using a MagnapureCompact instrument (Roche Diagnostics) according to the manufacturer’sinstructions. Total nucleic acid in the samples was quantified by absor-bance at 260 nm using a Nanodrop ND-1000 spectrophotometer. Duplicate10-ml samples of purified nucleic acid were analyzed, and the resultingcopies per sample RhCMV IE-2 gene were divided by the micrograms oftotal nucleic acid in the sample.
TCRB spectratyping
TCRB transcripts were analyzed for CDR3 region length polymorphismby PCR spectratyping, as previously described (40). We monitored spec-tratypes in PBMCs over 3 y, with four time points roughly 1 y apart. AllRMs were analyzed at least twice. TCRB spectratype profiles were in-dependently evaluated for morphology by three independent researchers;profiles were defined as “Gaussian” or “clonal” when at least two as-sessments categorized a Vb as such.
Statistical analysis
Statistical analysis was performed with methods indicated in the textand using SAS software version 9.2 (SAS Institute) or Prism 5 software(GraphPad). Assessments were considered non-significant if the p valuewas .0.05.
ResultsLoss of naive cells and T cell diversity in old monkeys
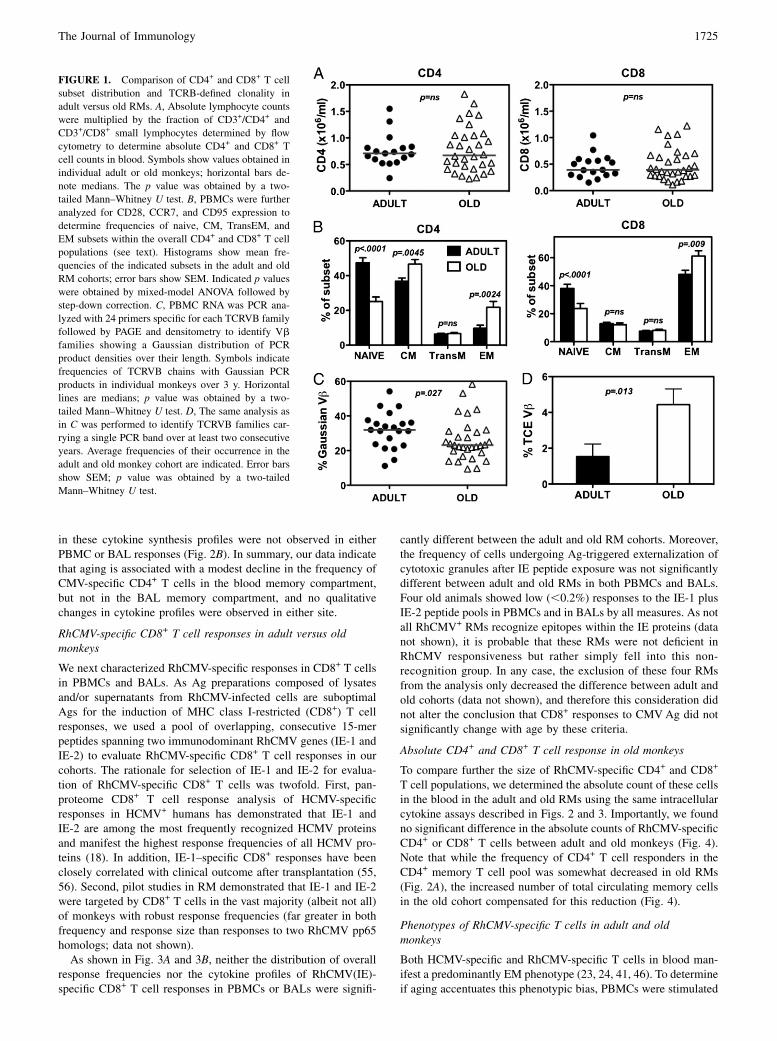
The overall goal of this project was to characterize changes in thequantity and/or quality of RhCMV-specific T cell responses as-sociated with old age. To accomplish this goal, we assembled acohort of randomly selected adult (7–10 y of age) and old (19–26 yof age) RMs (roughly corresponding with 20–30 and 60–80 y ofage, respectively, in the human life span). Our use of adult ratherthan juvenile RMs as the baseline population allowed us tocharacterize changes associated with aging and not with thematuration of the immune system. We first assessed the impact ofaging on the size and composition of the overall T cell compart-ment to provide context for changes in the RhCMV-specific T cellpopulations. As shown in Fig. 1A, the absolute counts of totalCD4+ and CD8+ T cells were comparable among the adult and oldRM cohorts, but the relative composition of these subsets wassignificantly different. The proportion of naive cells within boththe CD4+ and CD8+ T cell lineages, defined as CD28intCD95lo
(41), was significantly lower in old monkeys (Fig. 1B), consistentwith previous results (40, 41). Memory cells were subclassified bythe expression of CD28 and CCR7 into central memory (CM)subsets (CD28+CCR7+), transitional effector memory (TransEM)cells (CD28+CCR72), and EM cells (CD282CCR72) (51). Withregard to these subsets, the old RMs manifested a significant in-crease in relative size of the EM population in both lineages, as
well as a modest CD4+ T cell-specific increase in the proportionof CM cells (Fig. 1B). Notably, the proportion of TransEM cells,which include the highest fraction of proliferating cells and isa general reflection of ongoing EM cell production (51), was notdifferent between the adult and aged RMs.A key characteristic of an aging immune system is a general loss
of T cell repertoire diversity and the appearance of prominentclonal T cell expansions, reflecting both the decline of the highlydiverse naive population and the expansion or relative preservationof dominant memory clonotypes (9, 15, 52). To establish whetherour adult and aged RM cohorts differed with respect to repertoirediversity and frequency of clonal expansions, we monitored CDR3region length polymorphism of TCRB transcripts by PCR spec-tratyping (53). Cell populations exhibiting high TCRB diversityexhibit a bell-shaped (Gaussian) distribution of CDR3 regionlengths, whereas populations with narrow repertoire show dis-torted profiles and in extreme cases of repertoire reduction canshow a single PCR product band. We analyzed all 24 TCRBVchains in these adult and old RMs over 3 consecutive years andquantified their relative percentage of profiles with GaussianCDR3 size distributions and with highly skewed (single peak)distributions. As TCRBV populations with single peaks can rep-resent reversible expansions of T cells that recently encounteredan Ag (e.g., cells in the TransEM subset) or stable age-associated(potentially pathogenic) T cell clonal expansions (TCE; e.g., cellsin the EM subset) (52, 54), we determined the frequencies ofTCRBV families exhibiting the same TCE profile in at least twoconsecutive time points spaced at least a year apart. Notably, theseanalyses revealed that old RMs manifested significantly lowerfrequencies of Gaussian TCRB profiles (Fig. 1C) and a signifi-cantly higher frequency of TCRB profiles showing a stable, singlepeak (Fig. 1D). Overall, 58.8% of old RMs had at least one stableTCE, versus only 23.8% of young adult RMs, which was a sig-nificant difference (p = 0.014; two-tailed Fisher’s exact test).Taken together, these data argue for an age-associated skewing ofthe TCRB repertoire in the RMs of our old cohort.In separate studies, we have confirmed functional differences in
T cell responses in our adult versus young RM cohorts, showingthat the old cohort exhibited significantly weaker Ab and CD8+
T cell responses to an immunization with a live modified vacciniastrain Ankara (35). Thus, the old RM cohort studied in this articleexhibited significant changes in the composition and function ofthe T cell compartment, consistent with immune senescence, andreminiscent of the IRP defined in aged human subjects.
RhCMV-specific CD4+ T cell responses in adult versus oldmonkeys
RhCMV-specific CD4+ T cell responses were quantified in bothPBMCs and BALs, the latter representing an accessible mucosaleffector site (41), by in vitro stimulation with whole viral Agpreparations derived from RhCMV-infected fibroblasts and wereassessed for CD69 upregulation and IFN-g, TNF-a, and IL-2production by CD4+ memory T cells. The median frequency ofT cells within the CD4+ memory compartment in blood (PBMC)specifically responding to RhCMV Ag with a cytokine response(any of the three cytokines analyzed, in any combination) wassignificantly higher in the adult RMs than that in the old RMs (Fig.2A, left), but there was no difference in these frequencies in theBAL compartment [Fig. 2A, right; note that BAL T cells are es-sentially 100% memory in phenotype (41)]. To compare theprofile of cytokine responses in adult versus old RMs, we analyzedthe frequency of RhCMV-responding cells exhibiting any com-bination of the three cytokines and defined the contribution ofeach cytokine profile to the total response. Aging-related changes
1724 MAINTENANCE OF CMV-SPECIFIC T CELL RESPONSES IN OLD MONKEYS
in these cytokine synthesis profiles were not observed in eitherPBMC or BAL responses (Fig. 2B). In summary, our data indicatethat aging is associated with a modest decline in the frequency ofCMV-specific CD4+ T cells in the blood memory compartment,but not in the BAL memory compartment, and no qualitativechanges in cytokine profiles were observed in either site.
RhCMV-specific CD8+ T cell responses in adult versus oldmonkeys
We next characterized RhCMV-specific responses in CD8+ T cellsin PBMCs and BALs. As Ag preparations composed of lysatesand/or supernatants from RhCMV-infected cells are suboptimalAgs for the induction of MHC class I-restricted (CD8+) T cellresponses, we used a pool of overlapping, consecutive 15-merpeptides spanning two immunodominant RhCMV genes (IE-1 andIE-2) to evaluate RhCMV-specific CD8+ T cell responses in ourcohorts. The rationale for selection of IE-1 and IE-2 for evalua-tion of RhCMV-specific CD8+ T cells was twofold. First, pan-proteome CD8+ T cell response analysis of HCMV-specificresponses in HCMV+ humans has demonstrated that IE-1 andIE-2 are among the most frequently recognized HCMV proteinsand manifest the highest response frequencies of all HCMV pro-teins (18). In addition, IE-1–specific CD8+ responses have beenclosely correlated with clinical outcome after transplantation (55,56). Second, pilot studies in RM demonstrated that IE-1 and IE-2were targeted by CD8+ T cells in the vast majority (albeit not all)of monkeys with robust response frequencies (far greater in bothfrequency and response size than responses to two RhCMV pp65homologs; data not shown).As shown in Fig. 3A and 3B, neither the distribution of overall
response frequencies nor the cytokine profiles of RhCMV(IE)-specific CD8+ T cell responses in PBMCs or BALs were signifi-
cantly different between the adult and old RM cohorts. Moreover,the frequency of cells undergoing Ag-triggered externalization ofcytotoxic granules after IE peptide exposure was not significantlydifferent between adult and old RMs in both PBMCs and BALs.Four old animals showed low (,0.2%) responses to the IE-1 plusIE-2 peptide pools in PBMCs and in BALs by all measures. As notall RhCMV+ RMs recognize epitopes within the IE proteins (datanot shown), it is probable that these RMs were not deficient inRhCMV responsiveness but rather simply fell into this non-recognition group. In any case, the exclusion of these four RMsfrom the analysis only decreased the difference between adult andold cohorts (data not shown), and therefore this consideration didnot alter the conclusion that CD8+ responses to CMVAg did notsignificantly change with age by these criteria.
Absolute CD4+ and CD8+ T cell response in old monkeys
To compare further the size of RhCMV-specific CD4+ and CD8+
T cell populations, we determined the absolute count of these cellsin the blood in the adult and old RMs using the same intracellularcytokine assays described in Figs. 2 and 3. Importantly, we foundno significant difference in the absolute counts of RhCMV-specificCD4+ or CD8+ T cells between adult and old monkeys (Fig. 4).Note that while the frequency of CD4+ T cell responders in theCD4+ memory T cell pool was somewhat decreased in old RMs(Fig. 2A), the increased number of total circulating memory cellsin the old cohort compensated for this reduction (Fig. 4).
Phenotypes of RhCMV-specific T cells in adult and oldmonkeys
Both HCMV-specific and RhCMV-specific T cells in blood man-ifest a predominantly EM phenotype (23, 24, 41, 46). To determineif aging accentuates this phenotypic bias, PBMCs were stimulated
FIGURE 1. Comparison of CD4+ and CD8+ T cell
subset distribution and TCRB-defined clonality in
adult versus old RMs. A, Absolute lymphocyte counts
were multiplied by the fraction of CD3+/CD4+ and
CD3+/CD8+ small lymphocytes determined by flow
cytometry to determine absolute CD4+ and CD8+ T
cell counts in blood. Symbols show values obtained in
individual adult or old monkeys; horizontal bars de-
note medians. The p value was obtained by a two-
tailed Mann–Whitney U test. B, PBMCs were further
analyzed for CD28, CCR7, and CD95 expression to
determine frequencies of naive, CM, TransEM, and
EM subsets within the overall CD4+ and CD8+ T cell
populations (see text). Histograms show mean fre-
quencies of the indicated subsets in the adult and old
RM cohorts; error bars show SEM. Indicated p values
were obtained by mixed-model ANOVA followed by
step-down correction. C, PBMC RNA was PCR ana-
lyzed with 24 primers specific for each TCRVB family
followed by PAGE and densitometry to identify Vb
families showing a Gaussian distribution of PCR
product densities over their length. Symbols indicate
frequencies of TCRVB chains with Gaussian PCR
products in individual monkeys over 3 y. Horizontal
lines are medians; p value was obtained by a two-
tailed Mann–Whitney U test. D, The same analysis as
in C was performed to identify TCRVB families car-
rying a single PCR band over at least two consecutive
years. Average frequencies of their occurrence in the
adult and old monkey cohort are indicated. Error bars
show SEM; p value was obtained by a two-tailed
Mann–Whitney U test.
The Journal of Immunology 1725

with RhCMV Ag (whole Ag preparations and IE peptides, as inFigs. 2A and 3A), and cytokine-responding cells were categorizedby CD28 and CCR7 surface expression into CM, TransEM, orEM subsets of memory cells, as described in Fig. 1. In parallel,PBMCs were stimulated by the superantigen Staphylococcus en-terotoxin B (SEB) and similarly phenotyped to demonstrate thedistribution of a generic functional response among the CM,TransEM, or EM subsets. As shown in Fig. 5A, RhCMV-specificCD4+ T cells manifested a similar EM bias in both adult andold RMs, in contrast to the predominant CM phenotype of SEB-responsive cells. As shown in Fig. 1, the overall CD8+ memorycompartment is much more EM-biased than the CD4+ memorycompartment, and this overall bias increases with age. In keepingwith this, the SEB-responsive CD8+ T cell subset was more EM-biased in old RMs than adult RMs (Fig. 5B). Importantly, how-ever, the RhCMV(IE)-specific CD8+ T cell responses were evenmore EM-biased than the overall or SEB-responsive population,and this marked bias was similar in the adult and old RM cohorts.In conclusion, RhCMV-specific T cells are highly EM-biased inboth adult and old RMs, and aging did not increase the extent ofthis bias.
Function, phenotype, and triggering thresholds of CD8+ T cellresponses to immunodominant CMV peptide epitopes in adultand old monkeys
The 15-mer peptides composing the IE-1/IE-2 peptide mixes arenot optimal ligands for triggering CD8+ T cell responses (57), andthe responses to these mixes likely include variable combinationsof responses to different epitopes in each animal, factors that to-
gether might obscure differences in the function of RhCMV-specific CD8+ T cells in adult versus old RMs. To comparemore precisely the functional responses in adult versus old RMs,we screened our cohorts for common CD8+ T cell responses tooptimal IE-1 and IE-2 9-mer epitopes. Three epitopes with vig-orous responses common to multiple RMs in both age cohortswere identified (Table I). CD8+ T cells responding to these im-munodominant optimal epitopes displayed an even higher EMbias than the overall RhCMV(IE)-specific CD8+ T cell responses,but again, the adult and old RMs showed essentially identicalphenotype profiles (Fig. 6A). In addition, the cytokine profile ofthe CD8+ T cells responding to these immunodominant optimalpeptides were indistinguishable between the adult and old RMcohorts (Fig. 6B). As TCRs bind to peptide–MHC complexeswith varying avidity, and high-avidity responses provide better
FIGURE 3. Comparison of RhCMV(IE)-specific CD8+ T cell responses
in adult versus old RMs. A, PBMCs and BALs were assessed for CD8+ T
cell responses to mixes of consecutive, overlapping (11-aa overlap) 15-mer
peptides composing the RhCMV IE-1 and IE-2 proteins with cytokine flow
cytometry as described in Materials and Methods. The frequency of CD8+
T cells expressing CD69 and any combination of IFN-g, TNF-a, and IL-2
expression after incubation with and without RhCMV IE peptides was
determined, with the difference between these frequencies reported as the
RhCMV-specific CD8+ T cell response frequency. These values were then
normalized to the memory population size in each RM. Symbols show
individual monkeys; horizontal lines denote medians. The p value was
obtained by a two-tailed Mann–Whitney U test. B, To determine if the
pattern of IFN-g, TNF-a, and IL-2 production by RhCMV (IE) peptide-
stimulated CD8+ T cells differed in adult versus old RMs, the contribution
of each possible combination of these cytokines to the overall response
was determined by Boolean gating and statistically analyzed by a mixed-
effects regression model analysis. Histograms indicate average frequencies
of each of the indicated patterns in adult versus old responder cells; error
bars are SEM. Significance was assessed by mixed-effects regression
model analysis. C, The frequency of CD8+ T cells manifesting external-
ization of cytotoxic granules in response to RhCMV (IE) peptide versus
control stimulation in PBMCs (left) and BALs (right) was determined by
CD107a and CD107b staining. These values were then normalized to the
memory population size in each RM. Symbols show individual monkeys;
horizontal lines denote medians. The p value was obtained by two-tailed
Mann–Whitney U test.
FIGURE 2. Comparison of RhCMV-specific CD4+ T cell responses in
adult versus old RMs. A, PBMCs and BALs were assessed for CD4+ T cell
responses to RhCMV Ag preparations with cytokine flow cytometry as
described in Materials and Methods. The frequency of CD4+ T cells
expressing CD69 and any combination of IFN-g, TNF-a, and IL-2 ex-
pression after incubation with and without RhCMV Ag preparation was
determined, with the difference between these frequencies reported as the
RhCMV-specific CD4+ T cell response frequency. These values were then
normalized to the memory population size in each RM. Symbols show
individual monkeys; horizontal lines denote medians. The p value was
obtained by a two-tailed Mann–Whitney U test. B, To determine if the
pattern of IFN-g, TNF-a, and IL-2 production by RhCMVAg-stimulated
CD4+ T cells differed in adult versus old RMs, the contribution of each
possible combination of these cytokines to the overall response was de-
termined by Boolean gating and statistically analyzed by a mixed-effects
regression model analysis. Histograms indicate average frequencies of
each of the indicated patterns in adult versus old responder cells; error bars
are SEM.
1726 MAINTENANCE OF CMV-SPECIFIC T CELL RESPONSES IN OLD MONKEYS

protection against viral pathogens (58), we next investigated thefunctional avidity of CD8+ T cells to these immunodominantpeptides by determining the dependence of the cytokine and de-granulation responses on the dose of added peptide. The nor-malized dose-response curves of CD107–, IFN-g–, or TNF-a–responding CD8+ T cells upon stimulation with graded peptideconcentrations is shown in Fig. 6C. Notably, whereas dose de-pendence of the CD107 response to peptide VTTLGMALYshowed a minor shift to the right in old monkeys, other functionstriggered by this peptide and all functions triggered by a differentpeptide (SGVLPENVP) manifested statistically indistinguishabledose-response profiles. Taken together, these data argue against
a general decline in CD8+ T cell responsiveness to immunodo-minant RhCMV epitopes with aging in RMs.
Response of adult and old RMs to CMV challenge
Although ex vivo functional assays of RhCMV-specific T cellsrevealed few differences in the quantity or quality of theseresponses in adult versus old RMs, these data do not rule out adefect in the ability of aged RhCMV-specific T cells to respond toin vivo challenge. To address directly the in vivo capabilities ofRhCMV-specific T cell responses, we challenged a subset of theadult (n = 11) and old (n = 9) RM cohorts with infectious RhCMVand compared the ability of adult versus old RMs to respond toand control RhCMV superinfection (a superphysiologic challengedesigned to reveal otherwise subclinical deficits in anti-RhCMVimmunity). As a measure of such superinfection, levels ofRhCMV DNA were monitored by quantitative PCR of bothPBMCs and BALs. RhCMV DNAwas not detectable in either cellpopulation in any RM prior to challenge, and even after challenge,PBMCs remained negative in both adult and old RMs, consistentwith previous studies of RhCMV superinfection of adult RMs(44). However, RhCMV DNA appeared in the BALs of both adultand old RMs after challenge, starting at day 14 and peaking atpostinfection day 70, prior to a slow decline (Fig. 7). Importantly,the kinetics and levels of RhCMV replication in BALs were re-markably similar in adult and old RMs, strongly arguing thataging does not impair the ability of RhCMV-specific T cellresponses to control RhCMV. This in vivo challenge also allowedassessment of the T cell proliferative response to RhCMV-Agstimulation in vivo, as determined by the change in the fre-quency of RhCMV-specific T cells in blood and BALs, as well asthe induction of the proliferation-associated marker Ki-67 onRhCMV-specific T cells postchallenge (41). In this regard, al-though the average frequency of RhCMV-specific CD4+ T cells inblood was slightly, but significantly, higher in adult RMs com-pared with that in old RMs, both the fold increase in these
FIGURE 5. Comparison of the phenotypically defined differentiation state of RhCMV-specific T cell responses in adult versus old RMs. PBMCs were
stimulated with SEB (200 ng/ml), whole RhCMVAg preparations, RhCMV(IE) peptide mixes, or no Ag, as described in Figs. 2 and 3. Cells were analyzed
for the expression of CD3, CD4, CD8, CD69, IFN-g, TNF-a, and IL-2, as described in Figs. 2 and 3, and in addition, the surface markers CD28 and CCR7,
with the CD28 versus CCR7 phenotype (CM, TransEM, and EM designations as in Fig. 1) determined for all responding cells (CD69+ plus any combination
of the three cytokines). To provide sufficient events for accurate phenotypic analysis, the data set was restricted to RMs with specific response frequencies of
.0.4%. A, Analysis of CD4+ T cells. B, Analysis of CD8+ T cells. Horizontal lines show medians. Significance assessed by Kruskal–Wallis test, with Dunn’s
postanalysis. Of note, parametric statistical analysis by ANOVA with Bonferroni postanalysis revealed an age-related difference between the CD8+ T cell
populations responding to SEB, at a significance of p , 0.05, but not CD8+ T cell populations to IE, or any of the responding CD4+ T cell populations.
FIGURE 4. Comparison of absolute counts of RhCMV-specific CD4+
and CD8+ T cells in adult versus old RMs. PBMCs were stimulated to
elicit cytokine responses in CD4+ or CD8+ T cells as shown in Figs. 2 and
3, respectively. Blood lymphocytes were counted in parallel, and this value
was multiplied by the fraction of CD4+ or CD8+ RhCMV-specific lym-
phocytes to define the absolute count of RhCMV-specific CD4+ and
CD8+ T cells. Symbols show individual monkeys; horizontal lines denote
medians. The p value was obtained by a two-tailed Mann–Whitney U test.
The Journal of Immunology 1727

frequencies and the kinetics and level of Ki-67 induction on thesecells were indistinguishable in adult versus old RMs (Fig. 8A–C).In keeping with their marked EM bias (59), frequencies ofRhCMV(IE)-specific CD8+ T cells did not increase after chal-lenge, and Ki-67 induction was modest; however, these profileswere also essentially identical in the adult versus old RM cohorts(Fig. 8D–F). Thus, the response to and control of RhCMV chal-lenge was equivalent in the adult and old RM cohorts.
DiscussionPrimate CMVs have evolved a unique immunobiologic relationshipwith their hosts. In the absence of disease and in most cases overtinfection, these viruses elicit and maintain remarkably high fre-quency T cell responses, considerably higher than those of othercommon viral infections and similar to those of overtly pathogenicchronic infections such as HIV (18, 60–64). Moreover, theseresponses manifest a distinctive highly EM-biased phenotype (22–25, 28, 59). Although the basis of this unique immunogenicityremains to be experimentally confirmed, HCMV is thought to, andRhCMV is known to, persist as a very low level continuous orfrequently intermittent infection, rather than as a completely orpredominantly quiescent, latent infection (44, 65, 66). This char-acteristic would provide chronic, low-level Ag exposure, whichmight drive the characteristic EM differentiation of CMV-specificT cells and maintain the very high CMV-specific T cell responsefrequencies over the lifetime of the host. Note that this chronic Ag
exposure is not so high as to induce the T cell “functional ex-haustion” that characterizes pathogenic chronic viral infection (67,68), as CMV-specific T cells are highly functional (23, 25, 46, 60,69). Indeed, the rapidity of CMV reactivation after the onset ofpotent immunosuppressive therapy, the potential seriousness ofCMV disease in this setting, and the fact that the CMV is almostinvariably controlled without overt disease in immunocompetentindividuals, even into old age, attests to the in vivo function ofthese T cell responses (30, 31, 47, 65). Moreover, the fact that thehighly EM-biased SIV-specific T cell responses elicited byRhCMV vectors encoding SIVAgs have shown promising efficacyin protection against pathogenic SIV challenge in RM suggeststhat “CMV-like” T cell responses can effectively function againstheterologous pathogens (46). However, as indicated above, theassociation of CMV infection and the IRP has led to the hypoth-esis that lifelong control of CMV might have adverse con-sequences in old age, with the massive immune responses againstCMV possibly causing or materially contributing to the immunesenescence and the consequent increased morbidity and mortalityof infectious diseases in the elderly (7, 11–13, 15, 17, 21). Morerecently, it has also been suggested that CMV control itself mightbe compromised in the elderly, leading to occult infection andassociated chronic inflammation that might compromise functionof diverse organs and physiologic processes (14, 70).Perhaps the most likely mechanism that could link CMV in-
fection or anti-CMV immunity to aging pathogenesis would be thedevelopment in old age of an intrinsic senescence of the CMV-specific T cell response itself. Given that CMV is often acquiredin early childhood and that CMV-specific T cells are, as suggestedabove, almost certainly exposed to chronic antigenic stimulation, itis reasonable to speculate that after decades of “use,” includingmany years without a functional thymus to provide new naive cellsto replenish and diversify these responses (71), CMV-specificT cells might develop critically short telomeres, accumulatemutations, and thus simply wear out (15, 19, 20, 29, 72). Hypo-thetically, this might lead to diminished control of CMV repli-
FIGURE 6. Comparison of the phenotype, cytokine profile, and avidity of CD8+ T cells responding to immunodominant peptides in adult versus old
RMs. A, CD8+ cells responding to the immunodominant 9-mer peptides shown in Table I were phenotyped by CD28 and CCR7 surface expression as in Fig.
5. Symbols show individual adult versus old RMs; horizontal lines are medians. Significance was assessed by Kruskal–Wallis test, with Dunn’s post-
analysis. B, Cytokine profiles of CD8+ T cells responding to immunodominant 9-mer peptides shown in Table I were determined as in Fig. 3. Histograms
indicate average values for adult versus old RMs; error bars are SEM. Significance was assessed by mixed-effects regression model analysis. C, PBMCs
were stimulated with serial dilutions of the VTTLGMALY (top panels) or SGVLPENVP (bottom panels) peptides. The percentage of CD8+ T cells
responding to these peptides with CD107 surface expression or intracellular production of IFN-g or TNF-a was determined as in Fig. 3 and was normalized
to the maximum response for each RM. Dose-response curves were defined by regression analysis in each RM, and adult and old RMs were compared by
mixed-model analysis.
Table I. CD8+ T cell response to optimal IE peptides (% responding inthe blood memory population)
PeptideAdult
Mean 6 SD (n)Old
Mean 6 SD (n)Kruskal–WallisTest p Value
VTTLGMALY 0.92 6 0.98 (4) 5.19 6 4.53 (8) .0.05SGVLPENVP 3.38 6 2.41 (4) 7.63 6 10.7 (5) .0.05SEDLQMTVI 6.57 6 9.23 (2) 6.43 6 8.59 (2) n/a
n/a, not applicable, given n = 2 for adult and aged groups.
1728 MAINTENANCE OF CMV-SPECIFIC T CELL RESPONSES IN OLD MONKEYS

cation, increasing the frequency and/or magnitude of reactivationepisodes, which in turn might result in direct pathogenicity byfrequent or continuous occult infection(s) and chronic inflam-mation in critical tissues and/or indirect pathogenicity by drivingexpansion of an abnormal CMV-specific T cell response or bystimulating chronic immune activation, thereby displacing or dys-regulating the homeostasis of other crucial T cell populations.Conceivably, these conditions might set up a reinforcing spiral ofdeterioration in which failing CMV-specific immunity leads to
CMV replication, which further compromises both the CMV-specific response and, as collateral damage, the overall immunesystem, leading to further CMV replication (as well as other in-fectious complications), and further immune compromise.This concept has been sustained by reports that either the CMV-
specific T cell response itself and/or the CD282 EM populationit dominates comprise the major proportion of PBLs in elderlyindividuals (particularly those with the highest short-term mor-tality), that these populations manifest replicative senescence andother functional “abnormalities,” that CMV detection in secretionsis more common in the elderly, and that elderly individuals withthe highest quartile of CMV-specific Ab levels have increasedmortality (7, 13, 15–17, 19, 20, 29, 70, 72). There are, however,several crucial caveats in interpreting these observations. First, thedomination of CMV-specific EM cells in the elderly has only beenreported as a relative dominance in peripheral blood. The overallnumber of CMV-specific T cells in the body has not been enu-merated in adult versus elderly individuals, and thus the domi-nance of CMV-specific T cells might result from loss of non-EMpopulations around a stable EM population that includes the vastmajority of the CMV-specific T cells. CMV2 seronegative indi-viduals would lack the benchmark of the large CMV-specificpopulation by which the relative loss of other populations inblood can be visualized. In tissue, CMV-specific T cells arepresent at high frequency in extralymphoid effector sites, but thesecells constitute only a minor component of T cells in the lymphnodes supporting naive and CM T cell homeostasis [(73) andL. Picker, unpublished observations], arguing against a directdisplacement of naive and CM T cells by CMV-specific T cells.The replicative senescence and other “abnormal” functions at-tributed to CMV-specific T cells (low IL-2, etc.) are in fact in-trinsic characteristics of EM cell differentiation and typify CMV-specific T cell populations from individuals of all ages, includingchildren, and even heterologous responses elicited by CMV vec-tors in the RM model (22, 46, 59). Moreover, the skewed clono-typic hierarchies of CMV-specific T cells are also characteristic ofmature CMV-specific T cell responses regardless of age (22, 26,44, 74). Finally, the association of CMV secretion or elevatedlevels of CMV Abs with sick or “soon to be sick” elderly indi-viduals is not surprising given CMV replication can increase withstress (65, 75). It is therefore possible that CMV replication andincreased levels of CMV-specific immunity in such end-of-lifeindividuals is initiated by their other age-associated disease,rather than vice versa.A crucial issue in the investigation of a possible causal role
of CMV in immune senescence is the question of whether CMV-specific T cell populations do or do not become dysfunctionalwith age and therefore are less able to control the virus. In thisstudy, we addressed this question in the RM model, which includesboth a human-like immune senescence syndrome and a CMV virus,RhCMV, which is biologically highly homologous to HCMV andwhich allows for infectious challenge with RhCMV. We comparedthe frequency of RhCMV-specific CD4+ and CD8+ T cells in bloodand lung airspace (a representative mucosal interface effectorsite), the absolute number and differentiation phenotype of thesecells in blood, and both ex vivo and in vivo function of these cellsin cohorts of adult and old RMs, the latter cohort showing labo-ratory and functional evidence of immune senescence. Overall,these studies failed to show biologically significant differences inadult versus old CMV-specific T cell responses, most importantlyincluding any difference in the response to and control of in-fectious RhCMV challenge. In a previously reported study, wefound that the clonotypic hierarchies of RhCMV-specific CD4+
T cell responses were similar in adult and old RMs (44). Taken
FIGURE 8. Comparison of the in vivo proliferation response of
RhCMV-specific T cells in adult versus old RMs after infectious RhCMV
challenge. The adult and old RMs challenged as shown in Fig. 7 were
followed over time for frequencies of RhCMV-specific T cells in the cir-
culating memory compartment by cytokine flow cytometry. Responding
cells were defined by IFN-g and/or TNF-a expression in CD69+ cells. A–
C, CD4+ T cells responding to whole RhCMVAg preparations. D–F, CD8+
T cells responding to RhCMV(IE) peptide mixes. Response frequencies
are shown both as average 6 SEM frequencies (A, D) and as average 6SEM fold change from the prechallenge baseline (average of day 27 and
day 0 values) (B, E). Responding cells were simultaneously analyzed for
expression of the proliferation marker Ki-67 (41) with results presented as
average 6 SEM for percent Ki-67+ of the RhCMV-specific T cells (C, F).
Statistical analysis was by repeated-measures ANOVA.
FIGURE 7. Comparison of RhCMV kinetics in adult and old monkeys
after infectious RhCMV challenge. Adult (n = 11) or old (n = 9) RMs were
challenged by s.c. injection of 107 PFU RhCMV. RhCMV genomes in
PBMCs or BALs were quantified by quantitative PCR and standardized to
genomic host DNA. Average 6 SEM of RhCMV copy numbers (y-axis) at
indicated postchallenge days (x-axis) are indicated. LD, limit of detection.
The Journal of Immunology 1729

together, these results are not consistent with a general de-terioration of RhCMV-specific T cells with old age and thereforeargue against a major contribution of such deterioration to im-mune senescence in this model. There are, however, several cav-eats to these analyses. First, we did observe a minor but sta-tistically significant shift to the left in one functional response toone immunodominant peptide. Second, our analysis of RhCMV-specific CD8+ T cells was limited to those specific for IE-1 andIE-2. Although this specificity is undoubtedly an important com-ponent of the overall RhCMV-specific response (55, 56), manyother CMV proteins are targeted by CD8+ T cells (18), and theIE-1– and IE-2–specific CD8+ T cell response may not be rep-resentative of CD8+ T cell responses to all RhCMV proteins.However, given the equivalence of adult and old RMs in thecontrol of, and functional response to, a superphysiologic in vivoRhCMV challenge, we do not believe these caveats change ouroverall interpretation that cellular immunity to RhCMV is gen-erally well preserved in old age. In this regard, it is notable thattranscriptional analysis of highly purified CD8+/CD282 cells inyoung and elderly people revealed remarkable similarity, whereasanalysis of CD8+/CD28+ populations revealed changes in the olddonors (76), results that also suggest relative preservation of EMT cell integrity with age compared with naive and CM T cells.These data do not, by any means, rule out the participation of
CMV infection in the pathogenesis of immune senescence inhumans, but they do help refine the possible mechanisms by whichsuch participation might occur. For example, because healthy el-derly individuals may harbor CMV infection and the associationbetween the IRP and aging mortality is not uniform across allhuman populations (32, 33), it is possible that other genetic orenvironmental predispositions might be required for failure ofCMV-specific immunity or other age-associated contribution ofCMV to aging pathogenesis. RMs and human populations thatlack such putative predispositions would lack CMV-mediatedacceleration of immune senescence, but still would be suscepti-ble to other age-related immune deterioration such as lack of thy-mopoiesis and loss of naive populations (17). In addition, CMVmight participate in the pathogenesis of immune senescence bymechanisms that do not require general deterioration of the CMV-specific T cell response. In this regard, chronic, nonpathogenicviral infections have been shown to reset the innate immunesystem in animal models, in some instances providing augmentedinnate immune-mediated protection against heterologous patho-gens (77). This innate immune resetting may have a negative ef-fect on immune homeostasis when maintained over long periodsof time and/or when associated with other pathology such asthymic involution (78). Finally, CMV may only participate inaging pathogenesis when associated with multisystem failure,providing an additional insult to a collapsing overall physiology(75). In this scenario, dysfunction of the CMV-specific T cell re-sponse might only occur in old individuals with significant on-going disease and would not be observed in otherwise healthyelderly subjects. As RMs manifesting significant disease at time ofassignment were excluded from the old RM cohort studied here,our study would not have identified such secondary failure ofRhCMV-specific immunity.The b-herpesviruses in general, and the CMV family of viruses
specifically, emerged prior to mammalian radiation, and therefore,these viruses have coevolved with their mammalian hosts, leadingto an evolutionarily “negotiated” balance between the host im-mune system and the virus. With this balance, the virus enjoyspervasiveness in a large host population that could not occur withunchecked pathogenicity. For the host, the prevention of patho-genicity within reproductively relevant members of the population
eliminates the evolutionary cost of carrying the infection. Theunique high frequency of EM-biased T cell responses elicited byprimate CMVs are clearly an integral part of this balance in thesespecies and should be considered normal and adaptive in children,adults, and most elderly. This, of course, does not preclude theinvolvement of CMV or these protective T cell responses in dis-ease in the elderly, a common theme in aging (7, 40), but doessuggest that the mechanisms responsible for any such involvementare likely to be complex. Our data demonstrate a remarkablepreservation of CMV-specific T cell responses in a primate model,arguing against a simple model in which progressive deteriorationor dysregulation of these responses releases CMV from immunecontrol in old individuals and/or otherwise contributes to immunedysfunction. Further work in both the human system and non-human primate models will be required to confirm or rule outother potential mechanisms linking CMV infection or CMV-specific immunity to age-related disease and mortality and/or todefine cofactors that might facilitate CMV-mediated pathogenesisin a subset of susceptible elderly individuals.
AcknowledgmentsWe thank Nate Whizin for cytometry assistance with data collection and
Tomi Mori for assistance with statistical analysis.
DisclosuresThe authors have no financial conflicts of interest.
References1. Ginaldi, L., M. F. Loreto, M. P. Corsi, M. Modesti, and M. De Martinis. 2001.
Immunosenescence and infectious diseases. Microbes Infect. 3: 851–857.2. High, K. P. 2004. Infection as a cause of age-related morbidity and mortality.
Ageing Res. Rev. 3: 1–14.3. Hakim, F. T., and R. E. Gress. 2007. Immunosenescence: deficits in adaptive
immunity in the elderly. Tissue Antigens 70: 179–189.4. Aspinall, R., G. Del Giudice, R. B. Effros, B. Grubeck-Loebenstein, and
S. Sambhara. 2007. Challenges for vaccination in the elderly. Immun. Ageing 4:9.
5. Weiskopf, D., B. Weinberger, and B. Grubeck-Loebenstein. 2009. The aging ofthe immune system. Transpl. Int. 22: 1041–1050.
6. Linton, P. J., and K. Dorshkind. 2004. Age-related changes in lymphocyte de-velopment and function. Nat. Immunol. 5: 133–139.
7. Derhovanessian, E., A. Larbi, and G. Pawelec. 2009. Biomarkers of humanimmunosenescence: impact of cytomegalovirus infection. Curr. Opin. Immunol.21: 440–445.
8. Kovaiou, R. D., and B. Grubeck-Loebenstein. 2006. Age-associated changeswithin CD4+ T cells. Immunol. Lett. 107: 8–14.
9. Posnett, D. N., R. Sinha, S. Kabak, and C. Russo. 1994. Clonal populations ofT cells in normal elderly humans: the T cell equivalent to “benign monoclonalgammapathy.” J. Exp. Med. 179: 609–618.
10. Wikby, A., P. Maxson, J. Olsson, B. Johansson, and F. G. Ferguson. 1998.Changes in CD8 and CD4 lymphocyte subsets, T cell proliferation responses andnon-survival in the very old: the Swedish longitudinal OCTO-immune study.Mech. Ageing Dev. 102: 187–198.
11. Olsson, J., A. Wikby, B. Johansson, S. Lofgren, B. O. Nilsson, andF. G. Ferguson. 2000. Age-related change in peripheral blood T-lymphocytesubpopulations and cytomegalovirus infection in the very old: the Swedishlongitudinal OCTO immune study. Mech. Ageing Dev. 121: 187–201.
12. Wikby, A., B. Johansson, J. Olsson, S. Lofgren, B. O. Nilsson, and F. Ferguson.2002. Expansions of peripheral blood CD8 T-lymphocyte subpopulations and anassociation with cytomegalovirus seropositivity in the elderly: the SwedishNONA immune study. Exp. Gerontol. 37: 445–453.
13. Pawelec, G., A. Akbar, C. Caruso, R. Effros, B. Grubeck-Loebenstein, andA. Wikby. 2004. Is immunosenescence infectious? Trends Immunol. 25: 406–410.
14. Vescovini, R., C. Biasini, A. R. Telera, M. Basaglia, A. Stella, F. Magalini,L. Bucci, D. Monti, T. Lazzarotto, P. Dal Monte, et al. 2010. Intense anti-extracellular adaptive immune response to human cytomegalovirus in very oldsubjects with impaired health and cognitive and functional status. J. Immunol.184: 3242–3249.
15. Hadrup, S. R., J. Strindhall, T. Køllgaard, T. Seremet, B. Johansson, G. Pawelec,P. thor Straten, and A. Wikby. 2006. Longitudinal studies of clonally expandedCD8 T cells reveal a repertoire shrinkage predicting mortality and an increasednumber of dysfunctional cytomegalovirus-specific T cells in the very elderly. J.Immunol. 176: 2645–2653.
16. Wang, G. C., W. H. Kao, P. Murakami, Q. L. Xue, R. B. Chiou, B. Detrick,J. F. McDyer, R. D. Semba, V. Casolaro, J. D. Walston, and L. P. Fried. 2010.
1730 MAINTENANCE OF CMV-SPECIFIC T CELL RESPONSES IN OLD MONKEYS

Cytomegalovirus infection and the risk of mortality and frailty in older women:a prospective observational cohort study. Am. J. Epidemiol. 171: 1144–1152.
17. Brunner, S., D. Herndler-Brandstetter, B. Weinberger, and B. Grubeck-Loe-benstein. 2011. Persistent viral infections and immune aging. Ageing Res. Rev.10: 362–369.
18. Sylwester, A. W., B. L. Mitchell, J. B. Edgar, C. Taormina, C. Pelte, F. Ruchti,P. R. Sleath, K. H. Grabstein, N. A. Hosken, F. Kern, et al. 2005. Broadly tar-geted human cytomegalovirus-specific CD4+ and CD8+ T cells dominate thememory compartments of exposed subjects. J. Exp. Med. 202: 673–685.
19. Ouyang, Q., W. M. Wagner, A. Wikby, S. Walter, G. Aubert, A. I. Dodi,P. Travers, and G. Pawelec. 2003. Large numbers of dysfunctional CD8+T lymphocytes bearing receptors for a single dominant CMV epitope in the veryold. J. Clin. Immunol. 23: 247–257.
20. Ouyang, Q., W. M. Wagner, W. Zheng, A. Wikby, E. J. Remarque, andG. Pawelec. 2004. Dysfunctional CMV-specific CD8(+) T cells accumulate inthe elderly. Exp. Gerontol. 39: 607–613.
21. Vescovini, R., C. Biasini, F. F. Fagnoni, A. R. Telera, L. Zanlari, M. Pedrazzoni,L. Bucci, D. Monti, M. C. Medici, C. Chezzi, et al. 2007. Massive load offunctional effector CD4+ and CD8+ T cells against cytomegalovirus in very oldsubjects. J. Immunol. 179: 4283–4291.
22. Komatsu, H., A. Inui, T. Sogo, T. Fujisawa, H. Nagasaka, S. Nonoyama,S. Sierro, J. Northfield, M. Lucas, A. Vargas, and P. Klenerman. 2006. Largescale analysis of pediatric antiviral CD8+ T cell populations reveals sustained,functional and mature responses. Immun. Ageing 3: 11.
23. Kern, F., E. Khatamzas, I. Surel, C. Frommel, P. Reinke, S. L. Waldrop,L. J. Picker, and H. D. Volk. 1999. Distribution of human CMV-specific memoryT cells among the CD8pos. subsets defined by CD57, CD27, and CD45 isoforms.Eur. J. Immunol. 29: 2908–2915.
24. Appay, V., P. R. Dunbar, M. Callan, P. Klenerman, G. M. Gillespie, L. Papagno,G. S. Ogg, A. King, F. Lechner, C. A. Spina, et al. 2002. Memory CD8+ T cellsvary in differentiation phenotype in different persistent virus infections. Nat.Med. 8: 379–385.
25. Casazza, J. P., M. R. Betts, D. A. Price, M. L. Precopio, L. E. Ruff,J. M. Brenchley, B. J. Hill, M. Roederer, D. C. Douek, and R. A. Koup. 2006.Acquisition of direct antiviral effector functions by CMV-specific CD4+T lymphocytes with cellular maturation. J. Exp. Med. 203: 2865–2877.
26. Bitmansour, A. D., S. L. Waldrop, C. J. Pitcher, E. Khatamzas, F. Kern,V. C. Maino, and L. J. Picker. 2001. Clonotypic structure of the human CD4+memory T cell response to cytomegalovirus. J. Immunol. 167: 1151–1163.
27. Almanzar, G., S. Schwaiger, B. Jenewein, M. Keller, D. Herndler-Brandstetter,R. Wurzner, D. Schonitzer, and B. Grubeck-Loebenstein. 2005. Long-term cy-tomegalovirus infection leads to significant changes in the composition of theCD8+ T-cell repertoire, which may be the basis for an imbalance in the cytokineproduction profile in elderly persons. J. Virol. 79: 3675–3683.
28. Sacre, K., G. Carcelain, N. Cassoux, A. M. Fillet, D. Costagliola, D. Vittecoq,D. Salmon, Z. Amoura, C. Katlama, and B. Autran. 2005. Repertoire, diversity,and differentiation of specific CD8 T cells are associated with immune pro-tection against human cytomegalovirus disease. J. Exp. Med. 201: 1999–2010.
29. Fletcher, J. M., M. Vukmanovic-Stejic, P. J. Dunne, K. E. Birch, J. E. Cook,S. E. Jackson, M. Salmon, M. H. Rustin, and A. N. Akbar. 2005. Cytomeg-alovirus-specific CD4+ T cells in healthy carriers are continuously driven toreplicative exhaustion. J. Immunol. 175: 8218–8225.
30. Rafailidis, P. I., E. G. Mourtzoukou, I. C. Varbobitis, and M. E. Falagas. 2008.Severe cytomegalovirus infection in apparently immunocompetent patients:a systematic review. Virol. J. 5: 47.
31. Morunglav, M., I. Theate, G. Bertin, and P. Hantson. 2010. CMV enteritiscausing massive intestinal hemorrhage in an elderly patient. Case Report Med.2010: pii: 385795.
32. Colonna-Romano, G., A. N. Akbar, A. Aquino, M. Bulati, G. Candore, D. Lio,P. Ammatuna, J. M. Fletcher, C. Caruso, and G. Pawelec. 2007. Impact of CMVand EBV seropositivity on CD8 T lymphocytes in an old population from West-Sicily. Exp. Gerontol. 42: 995–1002.
33. Derhovanessian, E., A. B. Maier, R. Beck, G. Jahn, K. Hahnel, P. E. Slagboom,A. J. de Craen, R. G. Westendorp, and G. Pawelec. 2010. Hallmark features ofimmunosenescence are absent in familial longevity. J. Immunol. 185: 4618–4624.
34. Douek, D. C., R. D. McFarland, P. H. Keiser, E. A. Gage, J. M. Massey,B. F. Haynes, M. A. Polis, A. T. Haase, M. B. Feinberg, J. L. Sullivan, et al.1998. Changes in thymic function with age and during the treatment of HIVinfection. Nature 396: 690–695.
35. Cicin-Sain, L., S. Smyk-Pearson, N. Currier, L. Byrd, C. Koudelka, T. Robinson,G. Swarbrick, S. Tackitt, A. Legasse, M. Fischer, et al. 2010. Loss of naiveT cells and repertoire constriction predict poor response to vaccination in oldprimates. J. Immunol. 184: 6739–6745.
36. Nikolich-Zugich, J., and B. D. Rudd. 2010. Immune memory and aging: aninfinite or finite resource? Curr. Opin. Immunol. 22: 535–540.
37. Wikby, A., I. A. Mansson, B. Johansson, J. Strindhall, and S. E. Nilsson. 2008.The immune risk profile is associated with age and gender: findings from threeSwedish population studies of individuals 20-100 years of age. Biogerontology9: 299–308.
38. Bruunsgaard, H., M. Pedersen, and B. K. Pedersen. 2001. Aging and proin-flammatory cytokines. Curr. Opin. Hematol. 8: 131–136.
39. Jankovic, V., I. Messaoudi, and J. Nikolich-Zugich. 2003. Phenotypic andfunctional T-cell aging in rhesus macaques (Macaca mulatta): differential be-havior of CD4 and CD8 subsets. Blood 102: 3244–3251.
40. Cicin-Sain, L., I. Messaoudi, B. Park, N. Currier, S. Planer, M. Fischer,S. Tackitt, D. Nikolich-Zugich, A. Legasse, M. K. Axthelm, et al. 2007. Dra-
matic increase in naive T cell turnover is linked to loss of naive T cells from oldprimates. Proc. Natl. Acad. Sci. USA 104: 19960–19965.
41. Pitcher, C. J., S. I. Hagen, J. M. Walker, R. Lum, B. L. Mitchell, V. C. Maino,M. K. Axthelm, and L. J. Picker. 2002. Development and homeostasis of T cellmemory in rhesus macaque. J. Immunol. 168: 29–43.
42. Haberthur, K., F. Engelman, A. Barron, and I. Messaoudi. 2010. Immune se-nescence in aged nonhuman primates. Exp. Gerontol. 45: 655–661.
43. Powers, C., and K. Fruh. 2008. Rhesus CMV: an emerging animal model forhuman CMV. Med. Microbiol. Immunol. (Berl.) 197: 109–115.
44. Price, D. A., A. D. Bitmansour, J. B. Edgar, J. M. Walker, M. K. Axthelm,D. C. Douek, and L. J. Picker. 2008. Induction and evolution of cytomegalovirus-specific CD4+ T cell clonotypes in rhesus macaques. J. Immunol. 180: 269–280.
45. Kaur, A., M. D. Daniel, D. Hempel, D. Lee-Parritz, M. S. Hirsch, andR. P. Johnson. 1996. Cytotoxic T-lymphocyte responses to cytomegalovirus innormal and simian immunodeficiency virus-infected rhesus macaques. J. Virol.70: 7725–7733.
46. Hansen, S. G., C. Vieville, N. Whizin, L. Coyne-Johnson, D. C. Siess,D. D. Drummond, A. W. Legasse, M. K. Axthelm, K. Oswald, C. M. Trubey,et al. 2009. Effector memory T cell responses are associated with protection ofrhesus monkeys from mucosal simian immunodeficiency virus challenge. Nat.Med. 15: 293–299.
47. Pass, R. F. 2001. Cytomegalovirus. In Fields Virology, 4th Ed. . D. M. Knipe, andP. M. Howley, eds., eds. Lippincott/The Williams and Wilkins Co., Philadelphia,PA, p. 2675–2705.
48. Clarke, L. M., A. Duerr, J. Feldman, M. F. Sierra, B. J. Daidone, andS. H. Landesman. 1996. Factors associated with cytomegalovirus infectionamong human immunodeficiency virus type 1-seronegative and -seropositivewomen from an urban minority community. J. Infect. Dis. 173: 77–82.
49. Arora, N., Z. Novak, K. B. Fowler, S. B. Boppana, and S. A. Ross. 2010. Cy-tomegalovirus viruria and DNAemia in healthy seropositive women. J. Infect.Dis. 202: 1800–1803.
50. Hansen, S. G., C. J. Powers, R. Richards, A. B. Ventura, J. C. Ford, D. Siess,M. K. Axthelm, J. A. Nelson, M. A. Jarvis, L. J. Picker, and K. Fruh. 2010.Evasion of CD8+ T cells is critical for superinfection by cytomegalovirus.Science 328: 102–106.
51. Picker, L. J., E. F. Reed-Inderbitzin, S. I. Hagen, J. B. Edgar, S. G. Hansen,A. Legasse, S. Planer, M. Piatak, Jr., J. D. Lifson, V. C. Maino, et al. 2006. IL-15induces CD4 effector memory T cell production and tissue emigration in non-human primates. J. Clin. Invest. 116: 1514–1524.
52. Clambey, E. T., J. W. Kappler, and P. Marrack. 2007. CD8 T cell clonalexpansions & aging: a heterogeneous phenomenon with a common outcome.Exp. Gerontol. 42: 407–411.
53. Mosley, R. L., M. M. Koker, and R. A. Miller. 1998. Idiosyncratic alterations ofTCR size distributions affecting both CD4 and CD8 T cell subsets in aging mice.Cell. Immunol. 189: 10–18.
54. Messaoudi, I., J. Lemaoult, J. A. Guevara-Patino, B. M. Metzner, andJ. Nikolich-Zugich. 2004. Age-related CD8 T cell clonal expansions constrictCD8 T cell repertoire and have the potential to impair immune defense. J. Exp.Med. 200: 1347–1358.
55. Nickel, P., G. Bold, F. Presber, D. Biti, N. Babel, S. Kreutzer, J. Pratschke,C. Schonemann, F. Kern, H. D. Volk, and P. Reinke. 2009. High levels of CMV-IE-1-specific memory T cells are associated with less alloimmunity and im-proved renal allograft function. Transpl. Immunol. 20: 238–242.
56. Bunde, T., A. Kirchner, B. Hoffmeister, D. Habedank, R. Hetzer, G. Cherepnev,S. Proesch, P. Reinke, H. D. Volk, H. Lehmkuhl, and F. Kern. 2005. Protectionfrom cytomegalovirus after transplantation is correlated with immediate early 1-specific CD8 T cells. J. Exp. Med. 201: 1031–1036.
57. Maecker, H. T., H. S. Dunn, M. A. Suni, E. Khatamzas, C. J. Pitcher, T. Bunde,N. Persaud, W. Trigona, T. M. Fu, E. Sinclair, et al. 2001. Use of overlappingpeptide mixtures as antigens for cytokine flow cytometry. J. Immunol. Methods255: 27–40.
58. Messaoudi, I., J. A. Guevara Patino, R. Dyall, J. LeMaoult, and J. Nikolich-Zugich. 2002. Direct link between mhc polymorphism, T cell avidity, and di-versity in immune defense. Science 298: 1797–1800.
59. Lanzavecchia, A., and F. Sallusto. 2005. Understanding the generation andfunction of memory T cell subsets. Curr. Opin. Immunol. 17: 326–332.
60. Waldrop, S. L., C. J. Pitcher, D. M. Peterson, V. C. Maino, and L. J. Picker. 1997.Determination of antigen-specific memory/effector CD4+ T cell frequencies byflow cytometry: evidence for a novel, antigen-specific homeostatic mechanism inHIV-associated immunodeficiency. J. Clin. Invest. 99: 1739–1750.
61. Asanuma, H., M. Sharp, H. T. Maecker, V. C. Maino, and A. M. Arvin. 2000.Frequencies of memory T cells specific for varicella-zoster virus, herpes simplexvirus, and cytomegalovirus by intracellular detection of cytokine expression. J.Infect. Dis. 181: 859–866.
62. Sester, M., U. Sester, S. Alarcon Salvador, G. Heine, S. Lipfert, M. Girndt,B. Gartner, and H. Kohler. 2002. Age-related decrease in adenovirus-specificT cell responses. J. Infect. Dis. 185: 1379–1387.
63. Hammarlund, E., M. W. Lewis, S. G. Hansen, L. I. Strelow, J. A. Nelson,G. J. Sexton, J. M. Hanifin, and M. K. Slifka. 2003. Duration of antiviral im-munity after smallpox vaccination. Nat. Med. 9: 1131–1137.
64. Betts, M. R., D. R. Ambrozak, D. C. Douek, S. Bonhoeffer, J. M. Brenchley,J. P. Casazza, R. A. Koup, and L. J. Picker. 2001. Analysis of total human im-munodeficiency virus (HIV)-specific CD4(+) and CD8(+) T-cell responses: re-lationship to viral load in untreated HIV infection. J. Virol. 75: 11983–11991.
65. Reinke, P., S. Prosch, F. Kern, and H. D. Volk. 1999. Mechanisms of humancytomegalovirus (HCMV) (re)activation and its impact on organ transplantpatients. Transpl. Infect. Dis. 1: 157–164.
The Journal of Immunology 1731

66. Sinclair, J., and P. Sissons. 2006. Latency and reactivation of human cytomeg-alovirus. J. Gen. Virol. 87: 1763–1779.
67. Barber, D. L., E. J. Wherry, D. Masopust, B. Zhu, J. P. Allison, A. H. Sharpe,G. J. Freeman, and R. Ahmed. 2006. Restoring function in exhausted CD8T cells during chronic viral infection. Nature 439: 682–687.
68. Day, C. L., D. E. Kaufmann, P. Kiepiela, J. A. Brown, E. S. Moodley, S. Reddy,E. W. Mackey, J. D. Miller, A. J. Leslie, C. DePierres, et al. 2006. PD-1 ex-pression on HIV-specific T cells is associated with T-cell exhaustion and diseaseprogression. Nature 443: 350–354.
69. Makedonas, G., N. Hutnick, D. Haney, A. C. Amick, J. Gardner, G. Cosma,A. R. Hersperger, D. Dolfi, E. J. Wherry, G. Ferrari, and M. R. Betts. 2010.Perforin and IL-2 upregulation define qualitative differences among highlyfunctional virus-specific human CD8 T cells. PLoS Pathog. 6: e1000798.
70. Stowe, R. P., E. V. Kozlova, D. L. Yetman, D. M. Walling, J. S. Goodwin, andR. Glaser. 2007. Chronic herpesvirus reactivation occurs in aging. Exp. Gerontol.42: 563–570.
71. Vezys, V., D. Masopust, C. C. Kemball, D. L. Barber, L. A. O’Mara,C. P. Larsen, T. C. Pearson, R. Ahmed, and A. E. Lukacher. 2006. Continuousrecruitment of naive T cells contributes to heterogeneity of antiviral CD8 T cellsduring persistent infection. J. Exp. Med. 203: 2263–2269.
72. Monteiro, J., F. Batliwalla, H. Ostrer, and P. K. Gregersen. 1996. Shortenedtelomeres in clonally expanded CD28-CD8+ T cells imply a replicative historythat is distinct from their CD28+CD8+ counterparts. J. Immunol. 156: 3587–3590.
73. Ellefsen, K., A. Harari, P. Champagne, P. A. Bart, R. P. Sekaly, and G. Pantaleo.
2002. Distribution and functional analysis of memory antiviral CD8 T cell responses
in HIV-1 and cytomegalovirus infections. Eur. J. Immunol. 32: 3756–3764.74. Price, D. A., J. M. Brenchley, L. E. Ruff, M. R. Betts, B. J. Hill, M. Roederer,
R. A. Koup, S. A. Migueles, E. Gostick, L. Wooldridge, et al. 2005. Avidity for
antigen shapes clonal dominance in CD8+ T cell populations specific for per-
sistent DNA viruses. J. Exp. Med. 202: 1349–1361.75. Limaye, A. P., K. A. Kirby, G. D. Rubenfeld, W. M. Leisenring, E. M. Bulger,
M. J. Neff, N. S. Gibran, M. L. Huang, T. K. Santo Hayes, L. Corey, and
M. Boeckh. 2008. Cytomegalovirus reactivation in critically ill immunocom-
petent patients. JAMA 300: 413–422.76. Lazuardi, L., D. Herndler-Brandstetter, S. Brunner, G. T. Laschober, G. Lepper-
dinger, and B. Grubeck-Loebenstein. 2009. Microarray analysis reveals simi-
larity between CD8+CD28- T cells from young and elderly persons, but not of
CD8+CD28+ T cells. Biogerontology 10: 191–202.77. Barton, E. S., D. W. White, J. S. Cathelyn, K. A. Brett-McClellan, M. Engle,
M. S. Diamond, V. L. Miller, and H. W. Virgin IV. 2007. Herpesvirus latency
confers symbiotic protection from bacterial infection. Nature 447: 326–329.78. Sauce, D., M. Larsen, S. Fastenackels, A. Duperrier, M. Keller, B. Grubeck-
Loebenstein, C. Ferrand, P. Debre, D. Sidi, and V. Appay. 2009. Evidence of
premature immune aging in patients thymectomized during early childhood. J.
Clin. Invest. 119: 3070–3078.
1732 MAINTENANCE OF CMV-SPECIFIC T CELL RESPONSES IN OLD MONKEYS




















