
Dilmun-Ancient Paradise of Bible that may have housed a Shiva Temple?

APPLIED ANIMAL BEHAVIOUR
SCIENCE
ELSEWIER Applied Animal Behaviour Science 48 (1996) 159-172
Enrichment effects on rhesus monkeys successively housed singly, in pairs, and in groups
Steven J. Schapiro *, Mollie A. Bloomsmith, Leila M. Porter, Scott A. Suarez
Department of Veterinury Sciences, The University of Texas M. D. Anderson Cancer Center, Rt. 2, Box 151 -BI, Bastrop TX 78602, USA
Accepted 2 January 1996
Abstract
Many studies have examined the short-term effects of inanimate environmental enrichment on the behavior of captive primates. Similarly, numerous studies have examined the behavioral effects of manipulations to the social environment. Few analyses have examined the long-term effects of inanimate environmental enrichment or compared the effects of inanimate and social enhancements. The behavior of control (n = 49) and enriched (n = 44) rhesus monkeys (Macaca mulutra) was observed from l-4 years, as subjects spent successive years housed singly, in pairs, and then in small groups. Social housing condition and/or age of subject significantly affected time spent inactive, grooming, playing, exploring, and behaving abnormally. Frequency of vocalization was similarly affected. Social housing resulted in more species-typical behavior patterns than did single caging. Subjects were more socially oriented when pair-housed than when living in small groups. Inanimate enrichment did not affect behavior over a 3 year period, although enhancements were well-used by singly-caged yearlings and pair-housed juveniles. Subjects used enrichment less frequently when housed in groups. The data suggest that the presence of a social partner(s) led to more beneficial changes in behavior than did the provision of inanimate enhancements for rhesus monkeys in the studied age range. Therefore, it is concluded that social enrichment should be provided when possible and appropriate. Efforts to enrich the inanimate environment will be most beneficial when focused on socially-restricted primates.
Keywords: Rhesus monkey; Housing; Primates; Environmental enrichment
’ Corresponding author: Tel. (512) 321.3991; Fax (512) 332-5208; e-mail. [email protected].
0168.1591/96/$15.00 0 1996 Elsevier Science B.V. All rights reserved PII SOl68-1591(96)01038-6

160 SJ. Schapiro et al./Applied Animal Behaviour Science 48 (1996) 159-172
1. Introduction
The potential benefits (increased levels of species-typical behaviors and decreased levels of abnormal behaviors) of inanimate environmental enrichment for captive primates housed singly (Brent et al., 1989; Line and Morgan, 1991; Bayne et al., 1991; Bayne et al., 1992) or socially (Bloomsmith et al., 1988; O’Neill et al., 1991; Novak et al., 1993; Parks and Novak, 1993; Anderson et al., 1994) have been demonstrated. Most such studies examined the effects of enrichment procedures designed to promote natural behaviors such as grooming, foraging, and playing, over intervals of weeks or months (Bayne et al., 1991; Byrne and Suomi, 1991; Pruetz and Bloomsmith, 1992). A few have sampled behavior longitudinally (Novak et al., 1992; Novak et al., 1993; Schapiro and Bloomsmith, 1994; Schapiro and Bloomsmith, 1995; Schapiro et al., 19961, but no studies have been published that continuously measured the effects of enrichment over long periods.
Manipulations of the social environment of captive primates are also known to affect behavior (Capitanio, 1986; Bloomsmith, 1989; Mason, 1991; Novak and Suomi, 1991). While early investigations of the effects of the social environment focused on the negative results of social deprivation (Harlow and Harlow, 1962; Mitchell, 1968; Rogers and Davenport, 19691, more recent studies of the social environment have focused on the beneficial outcomes of increasing species-typical stimulation via social opportunities, particularly those resulting from housing laboratory primates in pairs (Reinhardt et al., 1988; Reinhardt et al., 1995; Reinhardt, 1989; Reinhardt, 1994; Eaton et al., 1994; but see Ruppenthal et al., 1991 and Crockett et al., 1994). Behavioral effects of inanimate environmental enhancements might also be influenced by the dynamic multifaceted stimulation provided by social partners. Currently, there are no data available that compare the effects of inanimate enrichment across different social housing conditions, and only one study (Reinhardt, 1990) has compared the effects of social and inanimate enrichment.
Previous studies in this rhesus monkey colony showed that a three-phase environmen- tal enrichment program incorporating physical, feeding, and sensory enhancements to the cages of singly housed yearlings increased the amount of time the subjects spent playing and decreased the amount of time they spent self-grooming (Schapiro and Bloomsmith, 1995; Schapiro et al., 1995b). A combined program of physical and feeding enhancements increased inactivity and play and decreased time spent drinking for the same subjects when they were pair-housed as juveniles (Schapiro and Bloom- smith, 1994).
In the present study, observations of three cohorts of enriched and control monkeys as they were housed singly, in pairs, and in small groups for consecutive years are used to examine (1) the long-term effects of inanimate environmental enrichment and (2) the influence that the social environment has on the effectiveness of inanimate enrichment.
2. Animals, materials and methods
2.1. Subjects and housing
This study was conducted on rhesus monkeys (Macaca mulatta) that were part of a specific pathogen-free (SPF) breeding program that was designed to rid the colony of

SJ. Schapirn et al./Applied Animal Behaviour Science 48 (1996) 159-172 161
target viruses (Herpesvirus simiae, Simian immunodeficiency virus, Simian retrovirus type D, and Simian T-lymphotropic virus; Voss et al., 1991; Schapiro et al., 1994). Subjects lived in their natal groups for 1 year, were housed in single cages for the next year, and were then housed in mixed-sex pairs for the third year. The final step in this strategy was to form small groups at the beginning of the fourth year. The process of socialization, followed by restriction, gradual resocialization, and finally full social integration was specifically designed to minimize both viral transmission and distur- bance of reproductive and parental skills (Schapiro et al., 1994; Schapiro et al., 1995a; Schapiro et al., 1995~).
Three groups of rhesus monkeys served as subjects. Group 1 consisted of 33 animals (17 males, 16 females) born in 1988, Group 2 comprised 29 animals (15 males, 14 females) born in 1989, and Group 3 consisted of 31 monkeys (15 males, 16 females) born in 1990. All subjects were observed for 1 year in each of the three housing conditions after removal from their natal groups. In each cohort, approximately half of the animals (n = 44) received an environmental enrichment program during each year of observation, while the others (n = 49) served as controls, receiving no enrichment during any of the years of study. Although control subjects could see the enrichment devices when they were brought to the experimental subjects, they could not directly view monkeys using enrichment.
All subjects were provided with monkey biscuits twice daily, were given oranges three times per week, and had water available ad libitum. When group-housed, all subjects also received fresh produce once a week and a seed/grain mixture five times a week as part of the standard colony management protocol. Subjects were housed in rack-mounted stainless steel cages (see Schapiro and Bloomsmith, 1994; Schapiro and Bloomsmith, 1995) while singly- and pair-housed, and in kennel-type runs while group-housed (Schapiro et al., 1996). Except for Group 1, subjects spent all 3 years in outdoor buildings with sensory access to other animals and exposure to natural variation in environmental conditions. Group 1 was singly-caged in indoor rooms (Schapiro and Bloomsmith, 1995; Schapiro et al., 1995b1, but spent their years of pair and group housing in the outdoor buildings.
2.2. Enrichment
Specific enrichment programs were associated with each housing condition (see Table 1) and have been described in detail previously (Schapiro and Bloomsmith, 1994; Schapiro and Bloomsmith, 1995; Schapiro et al., 1996). The three groups of subjects received three separate phases of enrichment (physical, feeding, and sensory) in different orders while singly housed (Schapiro and Bloomsmith, 1995). When housed in pairs, enriched subjects simultaneously received a combination of feeding and physical enhancements using devices and procedures that were similar to those employed during single housing (Schapiro and Bloomsmith, 1994). Physical enrichment was available continuously, and five different feeding enhancements were presented throughout each workday. During group housing, enriched monkeys were provided with a combination of physical and feeding enrichment; some was available continuously, and some intermittently (Schapiro et al., 1996).

162 SJ. Schapiro et al./AppliedAnimal Behauiour Science 48 f1996) 1.59-172
2.3. Data collection
Fifteen minute focal animal observations (Altmann, 1974) were conducted on all subjects from 1 year of age to 4 years of age. Subjects were observed approximately twice per week while singly-housed, once per week while pair-housed, and slightly less than once per week while group-housed, yielding over 2700 h of singly-caged data, 1280 h of pair-housed data, and 760 h of group-housed data. Observations were made using a Tandy 102 portable computer and The Observer (Noldus, 19911, an observa- tional software package. Data were collected between 08:30 h and 17:30 h throughout the year, were balanced for time of day, and included periods during which feeding enhancements were full, partially full, and empty. Ethograms (Schapiro and Bloom- smith, 1994; Schapiro and Bloomsmith, 1995) of mutually exclusive behaviors were very similar across housing conditions, and abnormal and social behaviors were emphasized. Interobserver reliability for cumulative durations of behaviors within observation sessions was measured monthly for a total of seven observers and averaged 88%, 85%, and 86% for single caging, pair housing, and group housing, respectively.
In addition to the behavior of the subjects, the following measures were recorded simultaneously: location in relation to the cagemate (social location); whether the behavior was directed at cagemate (social direction); and whether enrichment was being used (enrichment use). It was not possible for singly-housed subjects to engage in
Table I Enrichment procedures by housing condition
Single cage Pair cage Group run
Physical enhancements Chain perch Two chain perches Two trapezes Ball Two balls Two balls or two dog toys Dog toy Two dog toys Fleece Two fleeces Mirror Empty Puzzle
Wading pool a Hay bedding b
Feeding enhancements Small foraging mat Two small mats Two large mats Food puzzle Two food puzzles Foraging tree Foraging tree Fresh produce Produce X 2 Produce X n ’ Frozen juice Frozen juice X 2
Two liquid dispensers Sensory enhancements Primate videotapes None None Social enhancements None One cagemate Group of four to six
a Wading pools were presented twice per week when weather permitted; b hay was presented as bedding for 5 consecutive days month ’ ; ’ n,the number of monkeys in the group.

S.J. Schupiro et uI./AppliedAnimal Behauiour Science 48 (1996) 159-172 163
Table 2 Categories of behavior and activity for analysis
Mutually exclusive categories of behavior Analyzed by duration Analyzed by frequency of occurrence
Inactivity Sexual behavior Grooming
Self-grooming Social grooming a
Feeding Playing Abnormal behavior Exploring
Vocalizing
Non-mutually exclusive categories of activity Analyzed by duration
Socially-directed activities a Socially-located activities a Enrichment use b
a Only recorded when subjects were ‘socially’ housed; b only recorded for enriched subjects.
either socially-directed or socially-located behaviors or for control subjects to use enrichment. Interobserver reliabilities across all housing conditions averaged 96%, 94%, and 94% for social location, social direction, and enrichment use, respectively.
2.4. Analyses
Data were analyzed using multivariate analysis of variance (MANOVA) techniques within the Systat statistical package. Individual behaviors from the ethogram (Schapiro and Bloomsmith, 1994) were grouped into seven mutually exclusive categories of activity (see Table 2). The mean cumulative durations of each activity category for each subject across the entire year for each of the three housing conditions were the dependent measures used in the analyses. This resulted in three data points per subject per activity (a total of 279 data points per activity).
A 2 ( X 3) MANOVA examining the effects of treatment (enriched or control) as the between-subjects factor and housing condition (single, pair, or group) as the within-sub- jects factor on the seven mutually exclusive behavior categories was performed. In addition, a separate 2 ( X 3) analysis of variance (ANOVA) was performed using number of vocalizations as the dependent measure. Separate 2 (~2) ANOVAs were conducted for the same effects on social location and on social direction of behavior to prevent skewing of the main analysis because these categories represented combinations of the mutually exclusive activities and because subjects could not exhibit these social activi- ties while living alone. A final one-way repeated measures ANOVA for enrichment use was performed across the three housing conditions for enriched subjects only. Owing to the sequential nature of our SPF strategy, housing condition was completely confounded with subject age.
164 SJ. Schupiro et al./Applied Animal Behmiour Science 48 (1996) 159-172
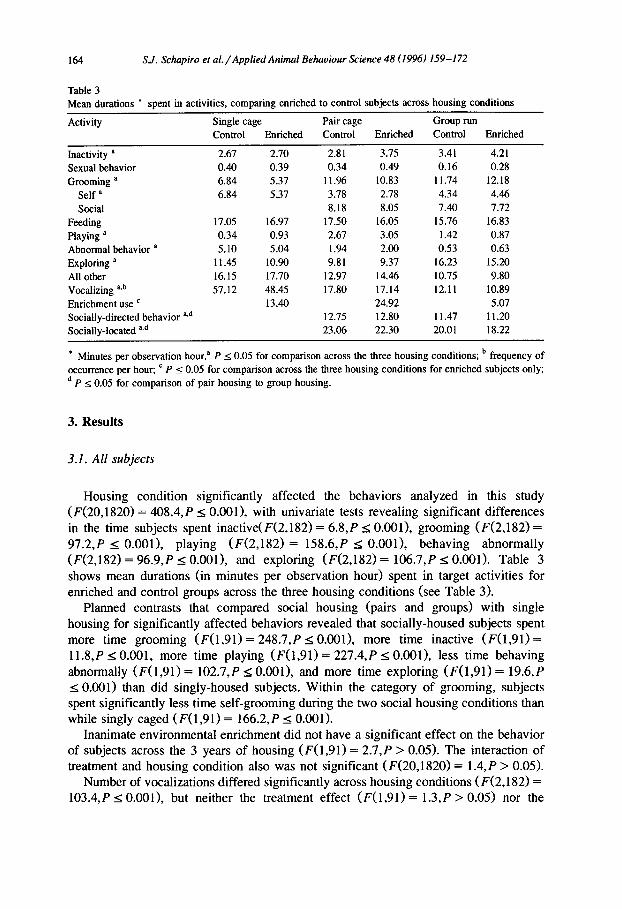
Table 3 Mean durations ?? spent in activities, comparing enriched to control subjects across housing conditions
Activity Single cage Pair cage Group run Control Enriched Control Enriched Control Enriched
Inactivity a 2.67 2.70 2.81 3.75 3.41 4.21 Sexual behavior 0.40 0.39 0.34 0.49 0.16 0.28 Grooming a 6.84 5.37 11.96 10.83 11.74 12.18
Self a 6.84 5.37 3.78 2.78 4.34 4.46 Social 8.18 8.05 7.40 7.72
Feeding 17.05 16.97 17.50 16.05 15.76 16.83 Playing a 0.34 0.93 2.67 3.05 1.42 0.87 Abnormal behavior a 5.10 5.04 1.94 2.00 0.53 0.63 Exploring ’ 11.45 10.90 9.81 9.37 16.23 15.20 All other 16.15 17.70 12.97 14.46 10.75 9.80 Vocalizing P,b 57.12 48.45 17.80 17.14 12.11 10.89 Enrichment use ’ 13.40 24.92 5.07 Socially-directed behavior ad 12.75 12.80 11.47 11.20 Socially-located qd 23.06 22.30 20.01 18.22
* Minutes per observation hour? P < 0.05 for comparison across the three housing conditions; b frequency of occurrence per hour; ’ P I 0.05 for comparison across the three housing conditions for enriched subjects only; d P s 0.05 for comparison of pair housing to group housing.
3. Results
3.1. All subjects
Housing condition significantly affected the behaviors analyzed in this study (F(20,1820) = 408.4, P s O.OOl), with univariate tests revealing significant differences in the time subjects spent inactive(F(2,182) = 6.8, P I; O.OOl>, grooming (F(2,182) = 97.2,P s O.OOl), playing (F(2,182) = 158.6,P < O.OOl>, behaving abnormally (F(2,182) = 96.9,P s O.OOl), and exploring (F(2,182) = 106.7,P 4 0.001). Table 3 shows mean durations (in minutes per observation hour) spent in target activities for enriched and control groups across the three housing conditions (see Table 3).
Planned contrasts that compared social housing (pairs and groups) with single housing for significantly affected behaviors revealed that socially-housed subjects spent more time grooming (F(1,91) = 248.7,P 5 O.OOl>, more time inactive (F(1,91) = 11.8,P ZG 0.001, more time playing (F(l,91) = 227.4,P I O.OOl), less time behaving abnormally (F(1,91) = 102.7,P 5 O.OOl), and more time exploring (F(l,91) = 19.6,P < 0.001) than did singly-housed subjects. Within the category of grooming, subjects spent significantly less time self-grooming during the two social housing conditions than while singly caged (F(1,91) = 166.2,P I 0.001).
Inanimate environmental enrichment did not have a significant effect on the behavior of subjects across the 3 years of housing (F(1,91> = 2.7, P > 0.05). The interaction of treatment and housing condition also was not significant (F(20,1820) = 1.4,P > 0.05).
Number of vocalizations differed significantly across housing conditions (F(2,182) = 103.4,P I O.OOl>, but neither the treatment effect (F(1,91) = 1.3,P > 0.05) nor the

SJ. Schapiro et al./Applied Animal Behauiour Science 48 (1996) 159-172 16.5
treatment by housing condition interaction (F(2,182) = 1 .O,P > 0.05) were significant. By planned comparison, number of vocalizations was significantly higher during single housing than during pair and group housing (F(1,91) = 109.6,P I 0.001).
Subjects spent significantly more time directing behavior toward and/or located near a cagemate when pair-housed than when group-housed (F(1,91) = 7.4, P < 0.01 and F(1,91) = 16.3,P s 0.001, respectively). The treatment effects (F(1,91) = 0.03,P > 0.05; F(1,91) = 2.1, P > 0.05) and the treatment by housing interactions (F(1,91) = 0.1, P > 0.05; F(1,91) = 0.3, P > 0.05) were not significant.
3.2. Enriched subjects only
For enriched subjects only, the amount of time spent using enrichment differed significantly across housing conditions (F(2,86) = 329.5,P I O.OOl), with the highest levels of use occurring during pair housing (almost 25 min h-l) and the lowest levels during group housing (5 min hh ’ >.
4. Discussion
The data clearly demonstrate that housing condition, age of subject, or both affected the behavior of the young rhesus monkeys in this study. Housing condition was confounded by age throughout, and potential influences of age on each behavior will be discussed in turn. In general, increasing age, within the range that we studied (l-4 years old), was unlikely to solely account for the observed changes in behavior.
Subjects spent more time playing, less time self-grooming and behaving abnormally, and vocalized less frequently while living in pairs and groups than they did while housed alone. Increased play and decreased abnormal behavior, self-grooming, and vocalizations are changes that more closely approximate species-typical patterns (Lin- dburg, 1971; Teas et al., 1980). Behavioral changes similar to these have been identified as representing an improvement in psychological well-being (Line, 1987; Novak and Suomi, 1988; Rosenblum and Andrews, 1995). Expression of more species-typical behaviors was an expected outcome of living with a social partner or partners (Bramb- lett, 1989; Mason, 1991; Novak and Suomi, 1991; de Waal, 1991). The lone observed contradiction to this beneficial trend of social housing was an increase in the amount of time that socially-housed, older subjects spent inactive. Increased inactivity is typical of older primates (Line and Morgan, 1991; Novak et al., 1992) but would not be expected in the age range that we were studying. An increase in inactivity exclusively as a function of social housing would also be unexpected. Given the previously mentioned constellation of benefits, the overall positive interpretation of the observed behavioral changes should not be compromised by this single presumably negative outcome.
The observed decrease in self-grooming while subjects were living in pairs and in small groups is of particular interest because it supports the suggestion that abnormally high levels of self-grooming by singly-caged rhesus may be amenable to reduction via enrichment attempts (Bayne et al., 1994; Eaton et al., 1994; Schapiro and Bloomsmith, 1995; Schapiro et al., 1995b). There is little reason to expect that time spent self-groom-

166 SJ. Schupiro et d/Applied Animul Behaviour Science 48 (1996) 159-172
ing should change significantly as young rhesus age. However, others have demon- strated that when primates lacked naturalistic stimulation, more self-directed grooming was exhibited (Harlow and Harlow, 1962; Capitanio, 1986) and that when naturalistic enhancements were provided, self-grooming diminished (Bayne et al., 1994; Eaton et al., 1994; Schapiro et al., 1995b). Inanimate and social enrichment each provide some naturalistic stimulation and thus lead to decreased levels of self-grooming.
Social housing led to significant decreases in the amount of time that subjects spent in abnormal activities. Abnormal behaviors, particularly those that develop while very young rhesus monkeys are socially restricted, are extremely persistent components of those animals’ lifetime behavioral repertoires, diminishing little as a function of age (Harlow and Harlow, 1962; Mitchell, 1968; Capitanio, 1986). Appropriate socialization, on the other hand, does diminish abnormal behavior, because conspecifics provide a wide variety of species-typical stimuli that serve a therapeutic function (Harlow and Suomi, 1971; Mason, 1991; Eaton et al., 1994). The significant decrease in abnormal behavior observed across housing conditions highlights an extremely important way in which social enrichment may be superior to inanimate enrichment. Abnormal behavior was not affected by any of our inanimate enrichment procedures (Schapiro and Bloomsmith, 1994; Schapiro and Bloomsmith, 1995) or by many similar enhancements studied by others (Byrne and Suomi, 1991; Line and Morgan, 1991; Bayne et al., 1994; but see Bloomsmith et al., 1988 and Bayne et al., 1991). Whereas inanimate enrichment increased some species-typical activities, social enrichment both increased species-typi- cal activity and diminished abnormal activity.
Cagemates were important foci of activity; 20% of observed behavior was socially- directed when subjects were socially housed. Approximately two-thirds of this behavior was socially-directed grooming. Subjects were more socially oriented when they were younger and pair-housed than they were when older and living in small groups. We attribute this to a combination of factors from the social and physical environments. Our subjects, having spent the previous year alone, were likely to be extremely motivated to affiliate. As in prior studies of rhesus monkeys (Reinhardt et al., 1988; Reinhardt, 1989; Reinhardt, 19941, when pairs were formed, affiliative interactions commenced almost immediately. In contrast, the more complex affiliative and agonistic interactions neces- sary for successful integration into a social group with four to seven same-aged strangers, without the influence of natural matrilines, took longer to develop. Although overt agonistic interactions were observed to account for only about 0.5 min h-l for group-housed subjects, this was over four times the amount of agonism observed among paired monkeys. The higher potential for agonism in larger social groups may have led to increased avoidance of social interactions in groups than in pairs.
Infants in the SPF program were housed in their natal groups until 1 year of age specifically to prevent the social inadequacy that is a common outcome of early restriction (Mitchell, 1968; Capitanio, 1986). Most study animals were socially compe- tent, as the high mean levels of social grooming attest, but subjectively, some appeared to be socially ‘awkward’, particularly when responding to the submissive gestures of other monkeys. Groups in which 3 year old males served as breeders (Schapiro et al., 1995~) were the most affected. Males this young, even without histories of social restriction, are unlikely to possess the social inclinations and maturity necessary to

S.J. Schapiro et al./AppliedAnimal Behaviour Science 48 (1996) 159-172 167
interact in adult species-typical fashion (Pusey and Packer, 1987; Bernstein et al., 1993). Analyses of pregnancy rates (Schapiro et al., 199%) showed that this problem seemed to diminish as males and females aged, suggesting an improvement in social compatibility.
Available space may also have affected social interactions in this study (an average of 21 min h-’ was spent within 8 cm of a cagemate). While subjects were group-housed, they had access to 30 times more volume than they did while pair-housed, providing more space for solitary activity. Hoff and colleagues (Hoff et al., 1994) have demon- strated that social interactions were diminished by large increases in available space, but this is not a universal finding (Nieuwenhuijsen and de Waal, 1982; Chamove and Rohrhuber, 1989; Novak et al., 1992).
Overall, enriched and control subjects did not differ behaviorally, despite the fact that the enriched subjects received 3 years of intensive enrichment. Enriched subjects did differ behaviorally from controls while they were singly and pair housed (Schapiro and Bloomsmith, 1994; Schapiro and Bloomsmith, 1995), but did not during group housing. The types of inanimate enhancements employed, although used by subjects in all housing conditions, may not have long-term value for monkeys that are similar to those studied (in terms of age, social experience, etc.).
Subjects used enrichment for much less time when housed in groups than during the other conditions. Several factors may have led to this outcome. First, monkeys may have habituated to the enrichment devices and procedures, because many of the enhancements were similar throughout the 3 years of the study. However, the very high level of enrichment use during pair housing, when enhancements were presented every workday for 1 year, weakens this possibility somewhat. Second, group runs contained at least 30 times the volume of cages and enrichment in runs occupied a smaller percentage of available volume than in cages. It was therefore easier for subjects to be away from enrichment in group runs than in cages. Finally, in the group run, unlike in the single cage, all subjects did not have free access to the enhancements. Pairs and groups were always provided with two of each enhancement in an attempt to limit competition; this strategy was more successful in pairs than in groups. While monkeys were rarely observed to engage in agonistic encounters over access to enrichment, they may have learned to avoid enhancements to avoid agonism, even though aggressive competition is reported to be relatively rare among rhesus monkeys of this age (de Waal, 1993). Increased subject age over single, pair, and group housing conditions is unlikely to explain the observed decrease in enrichment use (O’Neill and Price, 1991; Novak et al.. 1993).
Despite our inability to measure behavioral change as a function of long-term use of inanimate enrichment in this study, we still feel that feeding, physical, and occupational enhancements can be valuable for captive primates. We have previously identified positive behavioral effects and high levels of use of inanimate enrichment for these subjects while singly and pair housed (Schapiro and Bloomsmith, 1994, Schapiro and Bloomsmith, 1995). Numerous other studies have documented similar positive effects and high levels of enrichment use for primates in a variety of social settings (Blooms- mith et al., 1988; Line and Morgan, 1991; O’Neill et al., 1991; Bayne et al., 1992; Parks and Novak, 1993; Anderson et al., 1994).
Our data indicate that social enrichment is more effective and more powerful than

168 SJ. Schapiro et al./Applied Animal Behaviour Science 48 (1996) 159-172
inanimate enrichment, eclipsing any effects of physical, feeding, or sensory enhance- ments. The data suggest that in many situations, social enrichment should be preferen- tially implemented. Others have advocated the benefits of social enrichment (see Bramblett, 1989; Reinhardt et al., 1988; Reinhardt et al., 1995; Reinhardt, 1989: Reinhardt, 1994; Eaton et al., 19941, but few have conducted studies which simultane- ously compared inanimate and social enhancements (only Reinhardt (1990)). Social enrichment will not, however, be appropriate for all primates in all captive housing conditions (Coe, 1991; Ruppenthal et al., 1991; Crockett et al., 1994). Because inanimate enrichment does appear to be used by and behaviorally benefit many singly-housed primates, animals that can not be provided with social enrichment opportunities should receive inanimate enrichment, and the programs that they receive should include a combination of a variety of physical, feeding, and occupational enhancements. Monkeys that are already benefitting from multiple social partners may be appropriate candidates to receive inanimate enrichment programs that include fewer and simpler enhancements. Additionally, socially-restricted subjects that will soon be resocialized (animals which the data from this study suggest may enjoy short-term benefits from inanimate enrichment but are unlikely to demonstrate behavioral benefits in the long run) may also warrant less intense inanimate enrichment programs.
5. Conclusions
Social housing condition and/or subject age affected the amount of time that monkeys spent inactive, grooming, playing, exploring, and behaving abnormally, and the number of times they vocalized. Increased species-typical behaviors and decreased abnormal behaviors when subjects were pair- and group-housed compared to when they were singly-housed suggest that the social enrichment provided by a groupmate was beneficial. The inanimate enrichment procedures that we employed resulted in behav- ioral benefits while subjects were singly-caged, but had no long-term effects. Inanimate enrichment was used more by subjects when they were single- and pair-housed than when they were group-housed. To maximize the behavioral benefits of enrichment for captive primates, social enrichment should be provided when possible and appropriate, and inanimate enrichment efforts should be focused on primates with restricted social opportunities.
Acknowledgements
The authors wish to thank A. Kessel, M. McDonald, D. Oubari, and D. Bushong for dedicated collection of data, K. Vela and C. J. Maliniemi for data management efforts, and J. McMahan, D. Glasgow, and M. Smith for enrichment efforts. Animals are maintained in facilities approved by the American Association for Accreditation of Laboratory Animal Care and in accordance with current United States Department of Agriculture, Department of Health and Human Services, and National Institutes of

SJ. Schqiro et d/Applied Animal Behaviour Science 48 (1996) 159-172 169
Health regulations and standards. Financial support for this project came from the National Institutes of Health/NCRR grants U42-RR05080 and ROl -RRO5092.
References
Altmann, J. (1974) Observational study of behavior: Sampling methods, Behaviour, (Vol. 491, 227-267. Anderson, J.R., Rortais, A. and Guillemein, S. (1994) Diving and underwater swimming as enrichment
activities for captive rhesus macaques (Macaw mulurfa), Anim. Welf., (Vol. 3), 275-283. Bayne, K., Mainzer, H., Dexter, S., Campbell, Cl., Yamada, F. and Suomi, S. (1991) The reduction of
abnormal behaviors in individually housed rhesus monkeys (Macaw muluttd with a foraging/grooming board, Am. J. Primatol., (Vol. 231, 23-35.
Bayne, K., Dexter, S., Mainzer. H., McCully, C., Campbell, Cl. and Yamada, F. (1992) The use of artificial turf as a foraging substrate for individually housed rhesus monkeys (Mucaca mulattu), Anim. Welf., (Vol. I), 39-53.
Bayne, K.A.L., Strange, G.M. and Dexter, S.L. (1994) Influence of food enrichment on cage side preference, Lab. Anim. Sci., (Vol. 441, 624-629.
Bernstein, I.S., Judge, P.G. and Ruehlmann, T.E. (1993) Sex differences in adolescent rhesus monkey (Macaca muluttd behavior, Am. J. Primatol., (Vol. 31). 197-210.
Bloomsmith, M.A. (1989) Interactions between adult male and immature captive chimpanzees: Implications for housing chimpanzees, Am. J. Primatol., Suppl., (Vol. l), 93-99.
Bloomsmith, M.A., Alford, P.L. and Maple, T.L. (19881 Successful feeding enrichment for captive chim- panzees, Am. J. Primatol., (Vol. 161, 155-164.
Bramblett, C., 1989. Mental well-being in anthropoids. In: E.F. Segal (Editor), Housing, Care and Psychologi- cal Wellbeing of Captive and Laboratory Primates. Noyes Publications, Park Ridge, NJ, pp. I- 11.
Brent, L., Lee, D.R. and Eichberg, J.W. (1989) Evaluation of two environmental enrichment devices for singly caged chimpanzees (Pan troglodytes), Am. J. Primatol., Suppl., (Vol. I), 65-70.
Byrne, G.D. and Suomi, S. (1991) Effects of woodchips and buried food on behavior patterns and psychological well-being of captive rhesus monkeys, Am. J. Primatol., (Vol. 23), 141- 151.
Capitanio, J., 1986. Behavioral pathology. In: G. Mitchell and J. Erwin (Editors), Comparative Primate Biology Vol. 2A: Behavior, Conservation and Ecology. Liss, New York, pp. 41 l-454.
Chamove, A.S. and Rohrhuber, B. (1989) Moving Callitrichid monkeys from cages to outside areas, Zoo Biol., (Vol. 81, 151-163.
Coe, C.L., 1991. Is social housing of primates always the optimal choice? In: M.A. Novak and A.J. Petto (Editors), Through the Looking Glass: Issues of Psychological Well-being in Captive Nonhuman Primates. American Psychological Association, Washington, DC, pp. 78-92.
Crockett, CM., Bowers, C.L., Bowden, D.M. and Sackett, G.P. (1994) Sex differences in compatibility of pair-housed adult longtailed macaques, Am. J. Primatol., (Vol. 32). 73-94.
de Waal, F.B.M., 1991. The social nature of primates. In: M.A. Novak and A.J. Petto @ditorsI, Through the Looking Glass: Issues of Psychological Well-being in Captive Nonhuman Primates. American Psychologi- cal Association, Washington, DC, pp. 69-77.
de WaaI, F.B.M., 1993. &development of dominance relations and affiliative bonds in rhesus monkeys. In: M.E. Pemira and L.A. Fairbanks (Editors), Juvenile Primates: Life History, Development, and Behavior. Oxford University Press, New York, pp. 259-270.
Eaton, G.G., Kelley, S.T., Axthelm, M.K., Iliff-Sizemore, S.A. and Shiigi, S.M. (1994) Psychological well-being in paired adult female rhesus (Macaca mdatta), Am. J. Primatol., (Vol. 33). 89-99.
Harlow, H.F. and Harlow, M.K. (1962) Social deprivation in monkeys, Sci. Am., (Vol. 2071, 137-146. Harlow, H.F. and Suomi, S.J. (1971) Social recovery by isolation-reared monkeys, Proc. Nat. Acad. Sci., (Vol.
681, 1534-1538. Hoff, M.P., Forthman, D.L. and Maple, T.L. (1994) Dyadic interactions of infant lowland gorillas in an
outdoor exhibit compared to an indoor holding area, Zoo Biol., (Vol. 13), 245-256.

170 SJ. Schapiro et al./Applied Animal Behaviour Science 48 (1996) 159-172
Lindburg, D.G., 1971. The rhesus monkey in North India. In: L.A. Rosenblum (Editor), Primate Behavior: Developments in Field and Laboratory Research Vol. 2. Academic Press, New York, pp. l-106.
Line, S.W. (1987) Environmental enrichment for laboratory primates, J. Am. Vet. Med. Assoc., (Vol. 190). 854-859.
Line, S.W. and Morgan, K.N. (1991) The effects of two novel objects on the behavior of singly caged adult rhesus macaques, Lab. Anim. Sci., (Vol. 41). 365-369.
Mason, W.A. (1991) Effects of social interaction on well-being: Development aspects, Lab. Anim. Sci., (Vol. 41). 323-328.
Mitchell, G. (I 968) Persistent behavior pathology in rhesus monkeys following early social isolation, Folia Primatol., (Vol. 81, 132- 147.
Nieuwenhuijsen, K. and de Waal, F.B.M. (1982) Effects of spatial crowding on social behavior in a chimpanzee colony, Zoo Biol., (Vol. l), 5-28.
Noldus, L.P.J.J. (1991) The observer: A software system for collection and analysis of observational data, Beh. Res. Meth. Instr. Comp., (Vol. 23), 415--429.
Novak, M.A., Musante, A., Munroe, H., O’Neill, P.L., Price, C. and Suomi, S.J. (1993) Old, socially housed rhesus monkeys manipulate objects, Zoo Biol., (Vol. 12), 285-298.
Novak, M.A., O’Neill, P. and Suomi, S.J. (1992) Adjustments and adaptations to indoor and outdoor environments: Continuity and change in young adult rhesus monkeys, Am. J. Primatol., (Vol. 28). 125- 138.
Novak, M.A. and Suomi, S.J. (1988) Psychological well-being of primates in captivity, Am. Psychol., (Vol. 43), 765-773.
Novak, M.A. and Suomi, S.J. (1991) Social interaction in nonhuman primates: An underlying theme for primate research, Lab. Anim. Sci., (Vol. 41), 308-314.
O’Neill, P.L., Novak, M.A. and Suomi, S.J. (1991) Normalizing laboratory-reared rhesus macaque (Macaca mulatra) behavior with exposure to complex outdoor enclosures, Zoo Biol., (Vol. IO), 237-245.
O’Neill, P.L. and Price, C.S. (1991) Customizing an enrichment program: Rhesus monkeys, Lab. Anim., (Vol. 201, 29-41.
Parks, K.A. and Novak, M.A. (1993) Observations of increased activity and tool use in captive rhesus monkeys exposed to troughs of water, Am. J. Primatol., (Vol. 29). 13-25.
Pruetz, J.D. and Bloomsmith, M.A. (1992) Comparing two manipulable objects as enrichment for captive chimpanzees, Anim. Welf., (Vol. 1). 127-137.
Pusey, A.E. and Packer, C., 1987. Dispersal and philopatry. In: B.B. Smuts, D.L. Cheney, R.M. Seyfarth, R.W. Wrangham and T.T. Struhsaker (Editors), Primate Societies. University of Chicago Press, Chicago, pp. 250-266.
Reinhardt, V. (1989) Behavioral response of unrelated adult male rhesus monkeys familiarized and paired for the purpose of environmental enrichment, Am. J. Primatol., (Vol. 17), 243-248.
Reinhardt, V. (1990) Time budget of caged rhesus monkeys exposed to a companion, a PVC perch, and a piece of wood for an extended time, Am. J. Primatol., (Vol. 201, 51-56.
Reinhardt, V. (1994) Pair-housing rather than single-housing for laboratory rhesus macaques, J. Med. Primatol., (Vol. 23). 426-43 1.
Reinhardt, V., Houser, D., Eisele, S., Cowley, D. and Vertein, R. (1988) Behavioral responses of unrelated rhesus monkey females paired for the purpose of environmental enrichment, Am. J. Primatol., (Vol. 14). 135-140.
Reinhardt, V., Liss, C. and Stevens, C. (1995) Social housing of previously single-caged macaques: What are the options and the risks?, Auim. Welf., (Vol. 41, 307-328.
Rogers, C.M. and Davenport, R.K. (1969) Effects of restricted rearing on sexual behavior of chimpanzees, Dcv. Psych., (Vol. 1). 200-204.
Rosenblum, L.A. and Andrews, M.W., 1995. Environmental enrichment and psychological well-being of nonhuman primates. In: B.T. Bennett, C.R. Abee and R. Henrickson (Editors), Nonhuman Primates in Biomedical Research: Biology and Management. Academic Press, New York, pp. 101-l 12.
Ruppenthal, G.C., Walker, C.G. and Sackett, G.P. (1991) Rearing infant monkeys (Macaca nemestrina) in pairs produces deficient social development compared with rearing in single cages, Am. J. Primatol., (Vol. 25). 103-I 13.

SJ. Schapiro et d./AppliedAnimul Behaviour Science 48 (1996) 159-172 171
Schapiro, S.J. and Bloomsmith, M.A. (1994) Behavioral effects of enrichment on pair-housed juvenile rhesus monkeys, Am. J. Primatol., (Vol. 32), 159-170.
Schapiro, S.J. and Bloomsmith, M.A. (1995) Behavioral effects of enrichment on singly-housed, yearling rhesus monkeys: An analysis including three enrichment conditions and a control group, Am. J. Primatol., (Vol. 35). 89-101.
Schapiro, S.J., Bloomsmith, M.A., Suarcz, S.A. and Porter, L.M. (1995) Maternal behavior of primiparous rhesus monkeys: Effects of limited social restriction and inanimate environmental enrichment, Appl. Anim. Behav. Sci., (Vol. 45), 141-152.
Schapiro, S.J., Lee-Pat&z, D.E., Taylor, L.L., Watson, L., Bloomsmith, M.A. and Petto, A. (1994) Behavioral management of specific pathogen-free (SPF) rhesus macaques: Group formation, reproduction, and parental competence, Lab. Anim. Sci., (Vol. 441, 229-234.
Schapiro, S.J., Porter, L.M., Suarez, S.A. and Bloomsmith, M.A. (1995) The behaviour of singly-caged, yearling rhesus monkeys is affected by the environment outside of the cage, Appl. Anim. Behav. Sci., (Vol. 45). 153-166.
Schapiro, S.J., Porter, L.M., Suarez, S.A. and Bloomsmith, M.A. (1995) Age and social experience affect the breeding performance of subadult male rhesus monkeys (Mucucu mulard, Cont. Top. Lab. Anim. Sci., (Vol. 34). 93-95.
Schapiro, S.J., Bloomsmith, M.A., Suarez, S.A. and Porter, L.M., 1996. A comparison of the effects of simple versus complex environmental enrichment on the behaviour of group-housed, subadult rhesus macaques. Anim. Welf., in press.
Teas, J.. Richie, T., Taylor, H. and Southwick, C., 1980. Population patterns and behavioral ecology of rhesus monkeys (Macucu muluffu) in Nepal. In: D. Lindburg (Editor), The Macaques: Studies in Ecology, Behavior, and Evolution. Van Nostrand Reinhold, New York, pp. 247-262.
Voss, W.R., Buchl, S.J., Keeling, M.E., Hill&d, J., Lcrche, N., Schapiro, S.J., Bloomsmith, M.A., 1991. Derivation strategy for establishing a Macaca mulatta (rhesus) colony specifically pathogen-free of Herpes B virus and simian retrovirus. AALAS Bull., 30(4), 18-19, 1991.
Copyright © 2022 FDOKUMEN



![18F]Fluoroazabenzoxazoles as potential amyloid plaque PET tracers: synthesis and in vivo evaluation in rhesus monkey](https://static.fdokumen.com/doc/165x107/631f7e5b3b43b66d3c0fcb6e/18ffluoroazabenzoxazoles-as-potential-amyloid-plaque-pet-tracers-synthesis-and.jpg)
















