
Captive‐Housed Male Cheetahs (Acinonyx jubatus soemmeringii) Form Naturalistic Coalitions:...
10
RESEARCH ARTICLE Captive‐Housed Male Cheetahs (Acinonyx jubatus soemmeringii) Form Naturalistic Coalitions: Measuring Associations and Calculating Chance Encounters Carly L. Chadwick, 1,2 * Paul A. Rees, 2 and Barry Stevens‐Wood 1 1 Formerly School of Biology, Chemistry and Health Sciences, Manchester Metropolitan University, Chester Street, Manchester, United Kingdom 2 Ecosystems and Environment Research Centre, School of Environment and Life Sciences, University of Salford, The Crescent, Greater Manchester, United Kingdom Cheetahs are known to reproduce poorly in captivity and research suggests that the reasons for this are behavioral, rather than physiological. In the wild, male cheetahs remain in stable groups, or coalitions, throughout their lifetime. Appropriate social group housing is important in enhancing welfare and reproductive success in captivity and this study examined the effect of changes in social group composition on the behavior of four male cheetahs: two siblings and two half siblings. During the study, the cheetahs were housed both in pairs and as a group of four, before one male was relocated. The remaining cheetahs were then housed in a trio. Affiliative behaviors were frequently shown within pairs and overt aggression was seldom observed. Association indices were calculated for each cheetah pair and corrected for chance encounters based on data generated from a Monte Carlo simulation. The indices showed that two coalitions existed before the relocated male departed. Following the relocation of one of the half siblings, the remaining cheetahs appeared to form a coalition of three, as the indices of association between the unrelated male and the siblings increased and allogrooming between unrelated individuals was observed. The findings of this study indicate that natural social groupings of male cheetahs can be successfully replicated in captivity, which could potentially improve the chances of reproductive success when they are introduced to female cheetahs. Zoo Biol. 32:518–527, 2013. © 2013 Wiley Periodicals, Inc. Keywords: behavior modeling; captive breeding; felid; group housing; social behavior; vigilance behavior; zoo INTRODUCTION The rapid decline in the wild cheetah (Acinonyx jubatus) population has led to the development of coordinat- ed captive breeding programs in zoos and breeding facilities in an attempt to increase cheetah numbers [Caro, 1993; Marker‐Kraus and Kraus, 1997]. Unfortunately, many zoos have encountered problems in breeding the species, including low conception rates, irregular estrous cycles and periods of anestrus in females, and the failure of pairs recommended for breeding by the studbook keeper to reproduce [Caro, 1993; Brown et al., 1996; Asa et al., 2011; Wachter et al., 2011]. Thus, while cheetahs in the wild show no evidence of reproductive problems [Laurenson et al., 1992; Lindburg et al., 1993; Wielebnowski et al., 2002], the captive population is not self‐sustaining [Crosier et al., 2007; Bau- man et al., 2010]. During 2011 only 90 animals (44 males and 46 females), representing 5.58% of the captive cheetah population, successfully bred and of 241 facilities that maintained cheetahs, only 33 (13.7%) produced at least one cub [Marker, 2012]. Previous research into the breeding performance of captive cheetahs has indicated that there are no differences in the reproductive physiology of wild and captive cheetahs [Wildt et al., 1993; Wielebnowski and Brown, 1998], or in the physiology of breeders and non‐ breeders within the captive population [Wielebnowski and *Correspondence to: Carly L. Chadwick, Ecosystems and Environment Research Centre, School of Environment and Life Sciences, University of Salford, Salford Crescent, Greater Manchester M5 4WT, United Kingdom. E‐mail: [email protected] Received 20 March 2012; Revised 23 May 2013; Accepted 04 June 2013 DOI: 10.1002/zoo.21085 Published online 28 June 2013 in Wiley Online Library (wileyonlinelibrary.com). © 2013 Wiley Periodicals, Inc. Zoo Biology 32: 518–527 (2013)
Transcript of Captive‐Housed Male Cheetahs (Acinonyx jubatus soemmeringii) Form Naturalistic Coalitions:...

RESEARCH ARTICLE
Captive‐Housed Male Cheetahs (Acinonyx jubatussoemmeringii) Form Naturalistic Coalitions: MeasuringAssociations and Calculating Chance EncountersCarly L. Chadwick,1,2* Paul A. Rees,2 and Barry Stevens‐Wood1
1Formerly School of Biology, Chemistry and Health Sciences, Manchester Metropolitan University, Chester Street,Manchester, United Kingdom2Ecosystems and Environment Research Centre, School of Environment and Life Sciences, University of Salford,The Crescent, Greater Manchester, United Kingdom
Cheetahs are known to reproduce poorly in captivity and research suggests that the reasons for this are behavioral, rather thanphysiological. In the wild, male cheetahs remain in stable groups, or coalitions, throughout their lifetime. Appropriate socialgroup housing is important in enhancing welfare and reproductive success in captivity and this study examined the effect ofchanges in social group composition on the behavior of four male cheetahs: two siblings and two half siblings. During thestudy, the cheetahs were housed both in pairs and as a group of four, before one male was relocated. The remaining cheetahswere then housed in a trio. Affiliative behaviors were frequently shownwithin pairs and overt aggression was seldom observed.Association indices were calculated for each cheetah pair and corrected for chance encounters based on data generated from aMonte Carlo simulation. The indices showed that two coalitions existed before the relocated male departed. Following therelocation of one of the half siblings, the remaining cheetahs appeared to form a coalition of three, as the indices of associationbetween the unrelated male and the siblings increased and allogrooming between unrelated individuals was observed.The findings of this study indicate that natural social groupings of male cheetahs can be successfully replicated incaptivity, which could potentially improve the chances of reproductive success when they are introduced to female cheetahs.Zoo Biol. 32:518–527, 2013. © 2013 Wiley Periodicals, Inc.
Keywords: behavior modeling; captive breeding; felid; group housing; social behavior; vigilance behavior; zoo
INTRODUCTION
The rapid decline in the wild cheetah (Acinonyxjubatus) population has led to the development of coordinat-ed captive breeding programs in zoos and breeding facilitiesin an attempt to increase cheetah numbers [Caro, 1993;Marker‐Kraus and Kraus, 1997]. Unfortunately, many zooshave encountered problems in breeding the species, includinglow conception rates, irregular estrous cycles and periods ofanestrus in females, and the failure of pairs recommended forbreeding by the studbook keeper to reproduce [Caro, 1993;Brown et al., 1996; Asa et al., 2011; Wachter et al., 2011].Thus, while cheetahs in the wild show no evidence ofreproductive problems [Laurenson et al., 1992; Lindburget al., 1993; Wielebnowski et al., 2002], the captivepopulation is not self‐sustaining [Crosier et al., 2007; Bau-man et al., 2010]. During 2011 only 90 animals (44 males and46 females), representing 5.58% of the captive cheetah
population, successfully bred and of 241 facilities thatmaintained cheetahs, only 33 (13.7%) produced at least onecub [Marker, 2012]. Previous research into the breedingperformance of captive cheetahs has indicated that there areno differences in the reproductive physiology of wild andcaptive cheetahs [Wildt et al., 1993; Wielebnowski andBrown, 1998], or in the physiology of breeders and non‐breeders within the captive population [Wielebnowski and
*Correspondence to: Carly L. Chadwick, Ecosystems and EnvironmentResearch Centre, School of Environment and Life Sciences, Universityof Salford, Salford Crescent, Greater Manchester M5 4WT, UnitedKingdom. E‐mail: [email protected]
Received 20 March 2012; Revised 23 May 2013; Accepted 04 June 2013
DOI: 10.1002/zoo.21085Published online 28 June 2013 in Wiley Online Library(wileyonlinelibrary.com).
© 2013 Wiley Periodicals, Inc.
Zoo Biology 32: 518–527 (2013)

Brown, 1998]. Thus, the most likely explanation for the poorreproductive performance of cheetahs in captivity is related tothe behavior and management of the species [Brownet al., 1996; Wielebnowski et al., 2002; Crosier et al., 2007].
In the wild, adult females are solitary unless accompa-nied by cubs [Caro, 1994; Wielebnowski et al., 2002; Terioet al., 2003]. During an extensive study of wild cheetahs in theSerengeti, Caro [1994] reported that cubs separated from theirmothers at approximately 18 months of age, remaining insibling groups for an average of 6.7 months with femalesseparating from their littermates between the ages of 23 and27months andmale siblings remaining together in a coalition.
Male cheetahs can be solitary, but up to 60% of wildmales remain in stable groups, or coalitions, throughout theirlifetime [Caro and Collins, 1986; Caro, 1993, 1994; Ruiz‐Miranda et al., 1998; Durant et al., 2004]. Coalition membersmaintain close proximity to one another and intra‐groupinteractions are mostly affiliative, with little overt aggressionand frequent displays of allogrooming [Caro, 1993, 1994].Coalitions almost always consist of brothers but unrelatedmales may also join up with littermates to form coalitions ofthree or four individuals [Caro, 1993, 1994]. Caro [1994]reported that coalitions of two individuals were likely toconsist only of littermates and coalitions of three individualswere likely to be composed of two littermates and one non‐relative. The formation of coalitions containing an unrelatedmale is thought to occur before 3 years of age [Caro, 1993].Coalitions are able to occupy territories for longer than singlemales [Caro and Collins, 1986; Durant et al., 2004] and morefemales are observed in territories held by coalitions[Caro, 1994; Durant et al., 2004; Gottelli et al., 2007].Furthermore, there is evidence that females mate withmultiple males in the same reproductive cycle and litters canbe of mixed paternity as a result [Gottelli et al., 2007]. Thus, itseems that the formation and maintenance of coalitions isimportant for improving the survival and reproductivesuccess of wild male cheetahs [Ruiz‐Miranda et al., 1998;Durant et al., 2004; Gottelli et al., 2007].
Animals living in zoos should, when possible, be keptin naturalistic social groups. Modern animal welfarelegislation requires that animals should be kept in such amanner that satisfies their biological requirements includingthe opportunity to exhibit normal behavior, including normalsocial behavior (e.g., Art 4 of the Council Directive 1999/22/EC of March 29, 1999 relating to the keeping of wild animalsin zoos). Appropriate social housing is an effective way ofimproving animal welfare by providing animals the context inwhich to express wild‐counterpart behavior [De Roucket al., 2005; Price and Stoinski, 2007]. This can also directlyaffect reproductive success [Mellen, 1991; Carlstead andShepherdson, 1994; Ruiz‐Miranda et al., 1998], and optimizethe use of available accommodation.
Some authors have suggested that social housing maybe important in determining reproductive success in cheetahs[Caro and Collins, 1986; Caro, 1993; Wielebnowskiet al., 2002]. Long‐term studies have been carried out into
the social behavior of wild cheetahs [Caro, 1994], howeverlittle is known about the effects of social group housingon captive cheetah behavior [Ruiz‐Miranda et al., 1998].Ruiz‐Miranda et al. [1998] studied the behavior of cheetahsin two coalitions during separation and reunion trials. Theyobserved increases in vocalization and pacing when coalitionmembers were separated from one another, and increasedaffiliative behaviors when they were reunited. The authorsconcluded that their findings indicated a degree ofpsychological attachment between coalition members[Ruiz‐Miranda et al., 1998].
Given the importance of sociality in the wild and theapparent emotional connectedness between males living incoalitions, housing male cheetahs in coalitions in captivitymay have implications for welfare and reproductive success[Caro, 1993; Ruiz‐Miranda et al., 1998]. The Association ofZoos and Aquariums (AZA) Husbandry Manual for thecheetah states that males can be housed either singly or incoalitions, and that male siblings should remain together forlife [Ziegler‐Meeks, 2009]. Consequently, there is variationin current housing practices and captive male cheetahs aremaintained singly, in pairs and in groups of three or more.However, while several institutions do house their males insocial groups, few published studies have examined thebehavior of captive coalitions in detail and even fewer havedocumented the behavioral effects of combining male pairs incaptivity.
This research was part of a larger project documentingthe behavior, activity and enclosure use of four young malecheetahs (Acinonyx jubatus soemmeringii), two siblings andtwo half siblings, following their arrival into a new zooexhibit [by C.C: Tetley, 2009]. During the study the cheetahswere housed initially in pairs, before being introduced to oneanother and housed as a group of four. Towards the end of thestudy, one male was relocated and the remaining cheetahswere then housed in a trio. The purpose of this study was totest the hypotheses that:
1. Related pairs of adult male cheetahs may be housed togetherin the same enclosure and will demonstrate a high level ofassociation and low level of aggression.
2. Existing social affiliations will be maintained when the twogroups are fused.
METHODS
Subjects and Housing
The subjects of the study were four‐male captive‐borncheetahs, housed at Chester Zoo (Chester, UK). One pair,Burba and Singa, were littermates and the other pair, Matrahand Shiraz, were half‐siblings from different litters with thesame sire. All four were approximately 1‐year‐old at thebeginning of the study. Burba and Singa arrived at ChesterZoo from Wadi al Safa Wildlife Centre (Dubai, United ArabEmirates) in April 2008. Matrah and Shiraz arrived from
Coalition Behavior of Male Cheetahs 519
Zoo Biology

Sharjah Breeding Centre (United Arab Emirates) inMay 2008. The cheetahs were housed in a large, naturalisticenclosure measuring approximately 3,570m2. The outdoorenclosure was divided into two large compounds at the frontof the exhibit and three smaller enclosures towards the rear.Each separate area of the enclosure contained a cubbing denand was furnished with trees and logs. The cheetahs could beviewed from a raised walkway which ran along the front ofthe exhibit and the side of compound 1, and from a coveredviewing window looking onto compound 2 (Fig. 1). Thecheetahs were in visual and auditory contact throughout thestudy, and were fed whole dead chickens or rabbits once perday at approximately 14:00 hr. Water was available adlibitum.
The study was divided into four phases, reflecting thechanges in the zoo’s management of the group. Phase 1 of thestudy began in May 2008, when Burba and Singa had accessto compound 1 only andMatrah and Shiraz only had access toenclosures A�C (Fig. 1). Only the siblings were observedduring phase 1 as the half‐siblings were not easily viewablefrom the visitor areas. Compound 2 was made available partway through the study and phase 2 began when Matrah andShiraz weremoved from enclosures A�C into compound 2 inJuly 2008, where they remained until December 3, 2008.Phase 3 began on December 3, 2008, when the gate thatseparated the compounds was opened and all four cheetahswere given access to both compounds. Phase 4 of the studybegan in May 2009, when Shiraz was moved from ChesterZoo and a female cheetah arrived from ZSL Whipsnade Zoo(Bedfordshire, UK). No observations were carried out on thisfemale as she was housed in the indoor quarters in quarantineuntil the end of the study. As far as it is possible to tell, theremaining males were not aware of her presence and it did notaffect their behavior.
Procedures
A published ethogram was amended to documentthe behavior of the cheetahs [Table 1, Caro, 1994;
Wielebnowski, 1999; Skibiel et al., 2007] and behaviorand location data were collected between June 12, 2008 andMay 22, 2009. Observations were conducted continuouslyduring zoo opening hours (approximately 10.30�16.30 hr)from the public viewing areas by one observer (C.C.). Whenthe cheetahs were housed as two separate pairs, one pair wasselected and for these animals recordings were made asfollows during each hour of the day: 0–20min, activity offocal animal A; 21–40min, activity of focal animal B; 41–60min, the locations of each animal were plotted on a 1:400scale map of the enclosure provided by Chester Zoo. Each ofthese recordings was made every minute [instantaneoussampling; Martin and Bateson, 2007] and the order of thethree 20‐min sessions was varied between visits. When thetwo pairs were kept in separate compounds, each pair wasstudied on alternate visits. When three or four animals werehoused in the same compound, this sequence of recordingswas repeated twice but with animal C replacing A and Dreplacing B in the second hour, again with the order of the 20‐min sessions being varied between visits. In addition, alloccurrences of aggression and allogrooming (as defined inTable 1) were recorded. In total, 124 hr of observation werecarried out over 23 days.
The map of the enclosure illustrated the position oflandmarks in the compounds (e.g., trees, logs, cubbing dens)and individuals were located on the map relative to theselandmarks. The position of each cheetah on the map wasindicated by his initial and a number between zero and 20corresponding to each minute of the session. These data wereused to investigate association patterns within and betweenthe pairs, and also provided an estimate of the time spent byeach cheetah in proximity to the others, defined as within 5mof each other [Caro, 1994] or occupying the same cubbingden.
Since wild coalitions maintain territories [Caro, 1994;Gottelli et al., 2007], it was anticipated that the cheetahswould bemore vigilant when an unrelated individual could beseen. In order to assess the significance of vigilance behavior,simultaneously while recording the activity of the focalanimal a record was made of whether or not the focal animalcould see either of the other pair. The recordings were madefrom an elevated area where it was possible to determinewhether or not the animals had line of sight of each other. Thiswas particularly obviously the case when the animalsthemselves were occupying elevated areas within theenclosure.
Data Analysis
Each behavior on the ethogram was assigned to one offour categories (Table 1): alert behavior, active behavior,inactive behavior, and in den (time spent in a cubbing den andnot observable). Data from focal animal sampling sessionswere summarized as the mean daily percentage of samplepoints on which each category of behavior occurred. In orderto investigate the effects of changes in housing conditions andFig. 1. Layout of the cheetah enclosure at Chester Zoo.
520 Chadwick et al.
Zoo Biology

group composition on social behavior and vigilance, dataanalysis focused particularly on aggression, allogrooming,and alert behaviors (Table 1). Friedman’s repeated measuresanalysis of variance was used to investigate changes in alertbehavior across the four phases of the study. A Wilcoxonsigned‐rank test was used to examine differences between thepercentage of sample points on which alert behavior wasobserved when the cheetahs in the other pair were out of sightand when they were visible. Daily rates of allogroomingwithin dyads were calculated by dividing the number ofoccurrences of allogrooming by the number of focal samplepoints in the day. Differences in the rates of allogroomingbetween dyads were analyzed using a Mann�Whitney U‐test. Statistical analyses were carried out using Minitab15®statistical software.
Data from location recording sessions were used tomeasure the distances between individuals on the 1:400 scalemap. This information was used to calculate indices ofassociation for all possible dyads using the simple ratio index[Ginsberg and Young, 1992]:
IA ¼ x
xþ yAB þ yA þ yB
where x is the number of separate occasions when Aand B are observed together, yA is the number of separateoccasions when only A is observed, yB is the number of
separate occasions when only B is observed and yAB is thenumber of separate occasions when A and B are observed notassociated. In the present study, “separate occasions” weredefined as recordings of proximity made at 5‐min intervalsand two individuals were “together” when the distancebetween them was 5m or less [Caro, 1994], or when theywere occupying the same cubbing den.
Possible values of the index range between zero(indicating two individuals are never seen together) and one(two individuals are always seen together). This index isappropriate for studies in which the measure of association issymmetrical (i.e., dyads are either associated or notassociated), all individuals can be identified, and theidentification of individuals does not depend on whether ornot they are associated [Whitehead, 2008]. Since our datameet these assumptions, the simple ratio gives an unbiasedestimate of the proportion of time the individuals in a dyadwere seen together [Ginsberg and Young, 1992;Whitehead, 2008].
Modeling Chance Associations
It is difficult to calculate the frequency of chanceencounters. Schülke and Kappeler [2003] calculated ex-pected encounter rates in fork‐marked lemurs (Phanerfurcifer) using a random gas model, whose variables includeanimal density, rate of movement and dispersion of
TABLE 1. Cheetah behavior ethogram
Behavior Definition
ActiveAggression Growling, hissing, slapping or biting directed at another cheetahClimbing Use of raised items in the enclosure, for example, logsFeeding Eating, drinking, chewing, or licking edible substancesLocomotion Walking, runningPlaying alone Engaging in playful activities (seemingly meaningless, but non‐aggressive behavior) aloneScent marking Animal sprays urine backwards towards an objectSocial play Engaging in playful activities (seemingly meaningless, but non‐aggressive behavior)
with another cheetahStanding Flank and hindquarters off the ground, forelegs and back legs supporting the bodyUrinating/defecating Any projection of bodily fluids (except scent‐marking). Includes vomitingVocalization Auditory sound emitted by the mouth
AlertLying alert Lying with flank and hindquarters on the ground and forelegs tucked under the body.
Vigilant: head raised, eyes open and looking aroundSitting alert Sitting on back legs with forelegs vertically supporting the body. Vigilant: head raised, eyes open
and looking aroundStanding alert Flank and hindquarters off the ground, forelegs and back legs supporting the body. Vigilant: head
raised, eyes open and looking aroundInactive
Allogrooming Animal licking the fur of another cheetahGrooming Animal licking or scratching itselfLying flat out Lying prone with head on the groundLying out Lying prone with head raised. Occasional rolling over includedSitting Sitting on back legs with forelegs vertically supporting the body
In denOut of sight Animal is in cubbing den and its behavior is not observable
Adapted from Caro [1994], Wielebnowski [1999], and Skibiel et al. [2007].
Coalition Behavior of Male Cheetahs 521
Zoo Biology

individuals. The problem is more pronounced in a space‐limited captive environment, where individuals may be, forexample, mutually attracted to a food source. Despite this, noattempts appear to have been made to estimate chanceencounters for any species in captivity.
In the present study, the space available to the cheetahschanged, resulting in the possibility of differing levels ofchance association depending on the accommodationarrangements. When housed in the 715m2 compound, thesibling pair were more likely to be in proximity simply bychance than when they were given access to both compounds(2,745m2). To ascertain the effects of enclosure size on thelevels of chance associations, we devised a simple MonteCarlo model usingMicrosoft® Excel 2007 and compared ouractual observations with a randomly generated data set.
Themodel generated random pairs of points (xaya, xbyb)within a hypothetical square enclosure whose sides were thesquare root of the area of an actual enclosure. For example forcompound 1 (715m2), pairs of points were generated within ahypothetical square enclosure with sides of 26.74 units. Thepairs of points, representing animals A and B, wereconsidered to be “together” if they were �5 units apart.
If the location of animal A in two‐dimensional space isxa, ya, and the location of animal B is xb, yb, the Euclideandistance between these points is calculated using Pythagoras’Theorem:
Distance ðdÞ ¼ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiðxa � xbÞ2 þ ðya � ybÞ2
q
If this value (d) is less than the maximum distance (m)which defines association (d<m) then the animals will bedeemed to be associating together. So, if m¼ 5 units andd¼ 7 units, the animals are not associating; if d¼ 0.5 units,then they are associating. For individuals A and B locatedwithin a square enclosure where m is the maximum distancewhich defines association and 0� x�Henclosure area and0� y�Henclosure area,
if m >
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiðxa � xbÞ2 þ ðya � ybÞ2
q
then A and B are associating.For each simulation, the number of samples generated
(i.e., pairs of points plotted) equaled the actual number ofobservations made in that part of the field study to which thecalculated association index was compared. For example,when Burba and Singa were studied alone the associationindex was calculated from 103 recordings, so the model wasused to generate 103 pairs of random locations. These wereused to calculate an association index for chance encounters.We replicated the simulation 1,000 times and calculated themean index of association (and standard deviation) forcomparison with our observed index of association. This wasrepeated for each phase of the study. The model assumes thatresources are evenly distributed throughout the enclosure andthat the animals make use of the whole enclosure.
Nonetheless, it provides a basic estimate of the number oftimes animals are in proximity simply by chance.
RESULTS
Social Behavior and Association
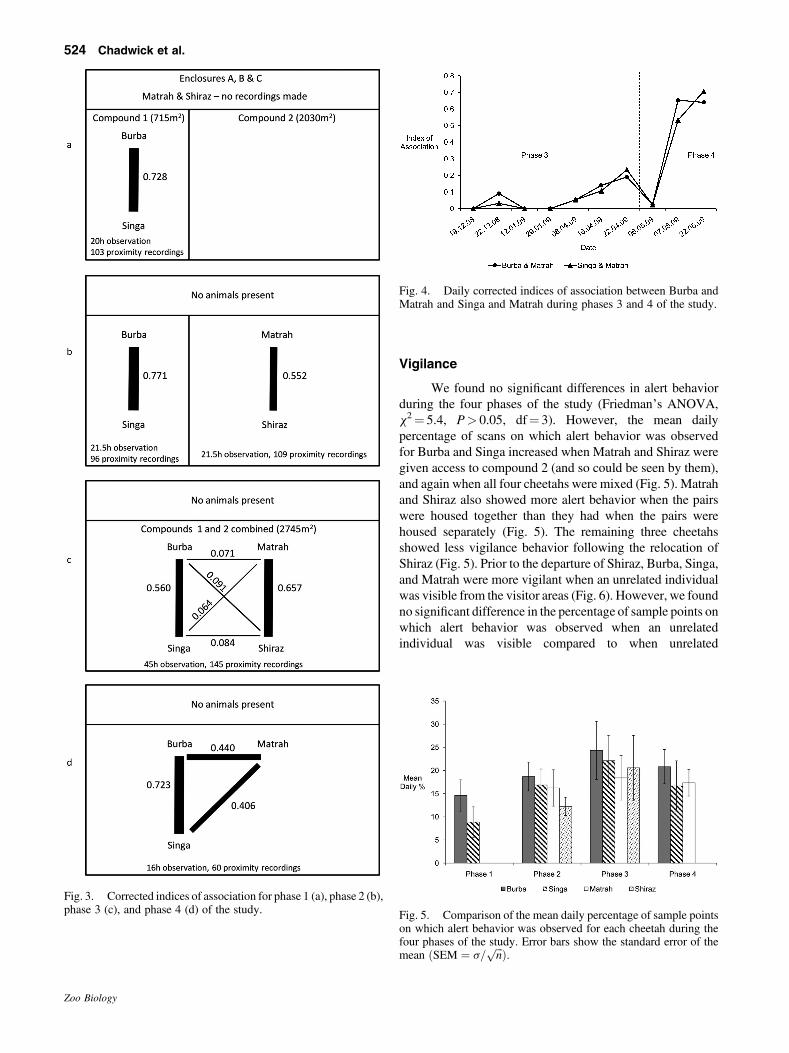
Association indices were calculated for all possibledyads for each phase of the study. These were adjusted bysubtracting the indices calculated from the model, whichrepresent the indices that would result from chanceencounters alone (Fig. 2), producing a corrected index ofassociation for each dyad (Fig. 3). Corrections ranged from adecrease of 3.6% to 28.9% of the uncorrected indices ofassociation, with larger corrections being applied to animalsin smaller enclosures (as would be expected, as chanceencounters are more likely here) and where uncorrected IAvalues were already low because the animals concerned wereonly rarely associating.
When they were housed separately, the individualsin each pair spent time in close proximity (Fig. 3a,b).When all four cheetahs were housed together and hadaccess to the same area, related individuals were moreclosely associated than unrelated individuals (Fig. 3c).Following Shiraz’s departure from Chester the indices ofassociation between the remaining three cheetahs in-creased as Matrah, who had been separated from his halfsibling, spent more time in closer proximity to the siblings(Fig. 3d). During this time, the space available to thecheetahs did not change.
Analysis of the days leading up to the departure ofShiraz revealed that the associations between Burba andMatrah and Singa and Matrah were already graduallyincreasing throughout April, before Shiraz was moved, andthat the association between Burba and Matrah increaseddramatically over the two consecutive observation days ofMay 6 andMay 7, 2009 (Fig. 4). After Shiraz left Chester, theremaining three cheetahs spent more time in closer proximityto one another even though Matrah was not related to Burbaand Singa.
During phases 1�3 of the study, there was nosignificant difference in the rates of allogrooming betweenthe brothers and the half brothers (Mann�Whitney, U¼ 3.0,P> 0.05). Allogrooming between unrelated individuals onlyoccurred during phase 4 of the study, after Shiraz had beenmoved to another collection. Following the departure ofShiraz, allogrooming between the siblings was observed 18times, Burba andMatrah were seen allogrooming seven timesand Singa and Matrah were seen allogrooming once.
During phase 3 of the study, in which all four cheetahshad access to the entire enclosure, aggressive interactionswere rare and never prolonged. Only eleven instances ofphysical aggression were recorded during 45 hr of observa-tions whilst all four cheetahs were housed together, a rate of0.06 aggressive behaviors (slaps or bites) per cheetah perhour.
522 Chadwick et al.
Zoo Biology
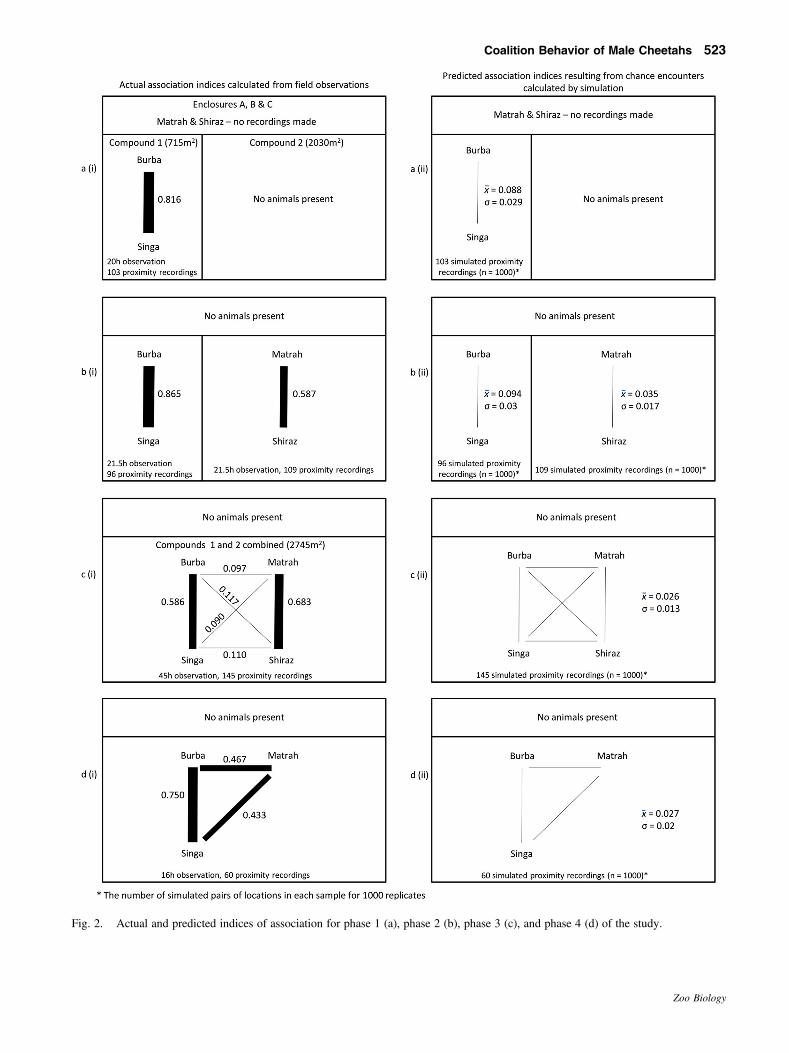
Fig. 2. Actual and predicted indices of association for phase 1 (a), phase 2 (b), phase 3 (c), and phase 4 (d) of the study.
Coalition Behavior of Male Cheetahs 523
Zoo Biology
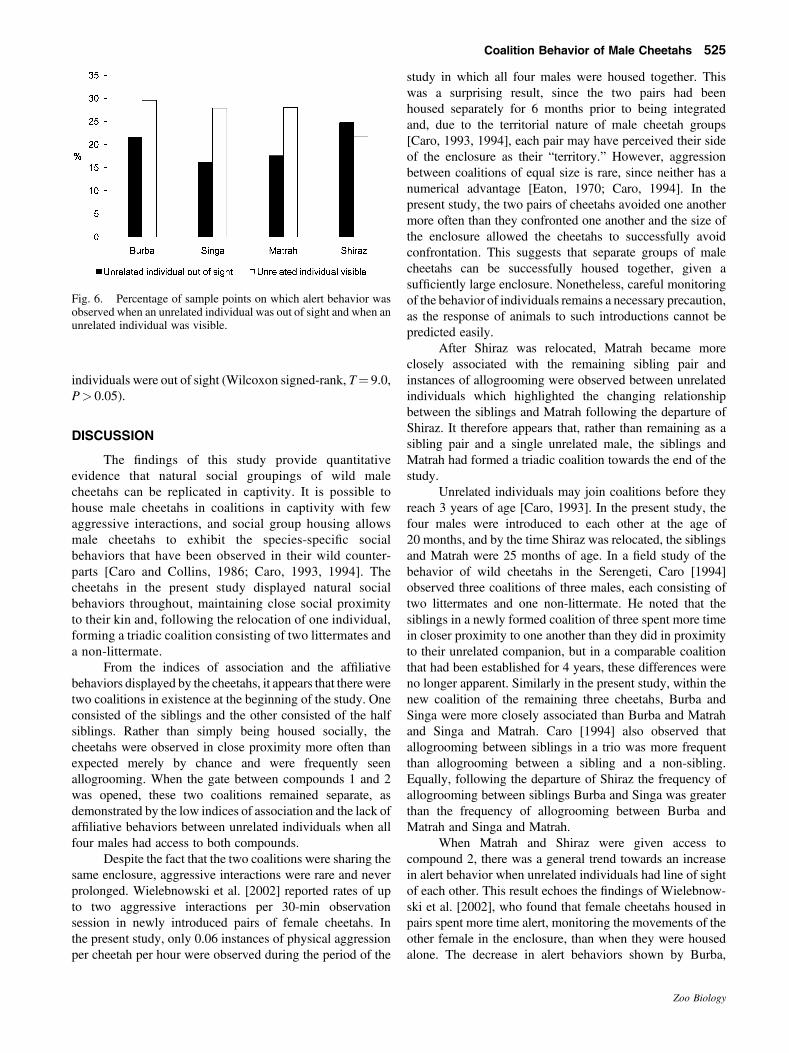
Vigilance
We found no significant differences in alert behaviorduring the four phases of the study (Friedman’s ANOVA,x2¼ 5.4, P> 0.05, df¼ 3). However, the mean dailypercentage of scans on which alert behavior was observedfor Burba and Singa increased when Matrah and Shiraz weregiven access to compound 2 (and so could be seen by them),and again when all four cheetahs were mixed (Fig. 5). Matrahand Shiraz also showed more alert behavior when the pairswere housed together than they had when the pairs werehoused separately (Fig. 5). The remaining three cheetahsshowed less vigilance behavior following the relocation ofShiraz (Fig. 5). Prior to the departure of Shiraz, Burba, Singa,and Matrah were more vigilant when an unrelated individualwas visible from the visitor areas (Fig. 6). However, we foundno significant difference in the percentage of sample points onwhich alert behavior was observed when an unrelatedindividual was visible compared to when unrelated
Fig. 3. Corrected indices of association for phase 1 (a), phase 2 (b),phase 3 (c), and phase 4 (d) of the study.
Fig. 4. Daily corrected indices of association between Burba andMatrah and Singa and Matrah during phases 3 and 4 of the study.
Fig. 5. Comparison of the mean daily percentage of sample pointson which alert behavior was observed for each cheetah during thefour phases of the study. Error bars show the standard error of themean ðSEM ¼ s=
ffiffiffin
p Þ.
524 Chadwick et al.
Zoo Biology
individuals were out of sight (Wilcoxon signed‐rank, T¼ 9.0,P> 0.05).
DISCUSSION
The findings of this study provide quantitativeevidence that natural social groupings of wild malecheetahs can be replicated in captivity. It is possible tohouse male cheetahs in coalitions in captivity with fewaggressive interactions, and social group housing allowsmale cheetahs to exhibit the species‐specific socialbehaviors that have been observed in their wild counter-parts [Caro and Collins, 1986; Caro, 1993, 1994]. Thecheetahs in the present study displayed natural socialbehaviors throughout, maintaining close social proximityto their kin and, following the relocation of one individual,forming a triadic coalition consisting of two littermates anda non‐littermate.
From the indices of association and the affiliativebehaviors displayed by the cheetahs, it appears that there weretwo coalitions in existence at the beginning of the study. Oneconsisted of the siblings and the other consisted of the halfsiblings. Rather than simply being housed socially, thecheetahs were observed in close proximity more often thanexpected merely by chance and were frequently seenallogrooming. When the gate between compounds 1 and 2was opened, these two coalitions remained separate, asdemonstrated by the low indices of association and the lack ofaffiliative behaviors between unrelated individuals when allfour males had access to both compounds.
Despite the fact that the two coalitions were sharing thesame enclosure, aggressive interactions were rare and neverprolonged. Wielebnowski et al. [2002] reported rates of upto two aggressive interactions per 30‐min observationsession in newly introduced pairs of female cheetahs. Inthe present study, only 0.06 instances of physical aggressionper cheetah per hour were observed during the period of the
study in which all four males were housed together. Thiswas a surprising result, since the two pairs had beenhoused separately for 6 months prior to being integratedand, due to the territorial nature of male cheetah groups[Caro, 1993, 1994], each pair may have perceived their sideof the enclosure as their “territory.” However, aggressionbetween coalitions of equal size is rare, since neither has anumerical advantage [Eaton, 1970; Caro, 1994]. In thepresent study, the two pairs of cheetahs avoided one anothermore often than they confronted one another and the size ofthe enclosure allowed the cheetahs to successfully avoidconfrontation. This suggests that separate groups of malecheetahs can be successfully housed together, given asufficiently large enclosure. Nonetheless, careful monitoringof the behavior of individuals remains a necessary precaution,as the response of animals to such introductions cannot bepredicted easily.
After Shiraz was relocated, Matrah became moreclosely associated with the remaining sibling pair andinstances of allogrooming were observed between unrelatedindividuals which highlighted the changing relationshipbetween the siblings and Matrah following the departure ofShiraz. It therefore appears that, rather than remaining as asibling pair and a single unrelated male, the siblings andMatrah had formed a triadic coalition towards the end of thestudy.
Unrelated individuals may join coalitions before theyreach 3 years of age [Caro, 1993]. In the present study, thefour males were introduced to each other at the age of20 months, and by the time Shiraz was relocated, the siblingsand Matrah were 25 months of age. In a field study of thebehavior of wild cheetahs in the Serengeti, Caro [1994]observed three coalitions of three males, each consisting oftwo littermates and one non‐littermate. He noted that thesiblings in a newly formed coalition of three spent more timein closer proximity to one another than they did in proximityto their unrelated companion, but in a comparable coalitionthat had been established for 4 years, these differences wereno longer apparent. Similarly in the present study, within thenew coalition of the remaining three cheetahs, Burba andSinga were more closely associated than Burba and Matrahand Singa and Matrah. Caro [1994] also observed thatallogrooming between siblings in a trio was more frequentthan allogrooming between a sibling and a non‐sibling.Equally, following the departure of Shiraz the frequency ofallogrooming between siblings Burba and Singa was greaterthan the frequency of allogrooming between Burba andMatrah and Singa and Matrah.
When Matrah and Shiraz were given access tocompound 2, there was a general trend towards an increasein alert behavior when unrelated individuals had line of sightof each other. This result echoes the findings of Wielebnow-ski et al. [2002], who found that female cheetahs housed inpairs spent more time alert, monitoring the movements of theother female in the enclosure, than when they were housedalone. The decrease in alert behaviors shown by Burba,
Fig. 6. Percentage of sample points on which alert behavior wasobserved when an unrelated individual was out of sight and when anunrelated individual was visible.
Coalition Behavior of Male Cheetahs 525
Zoo Biology

Singa, and Matrah after Shiraz left provides further evidencefor the formation of a triadic coalition, rather than the threecheetahs simply remaining as a single male and a sibling pairhoused in the same enclosure. If that was the case, the pair andthe single male would have continued to monitor each other’smovements and no decrease in vigilance behavior wouldhave been expected.
With the arrival of the female cheetah at the end of thestudy, there will inevitably be management implications in theevent that she is introduced to the males for breeding. Femalereproductive activity may be stimulated in the presence of agroup of males [Brown et al., 1996], as most matings observedin the wild have occurred between females and coalitionmembers [Caro, 1993] and ovulation in female cheetahs is ofteninduced [Wildt et al., 1993; Brown et al., 1996; Wielebnowskiet al., 2002]. Indeed, Bircher and Noble [1997] reported the useof male�male interactions to stimulate breeding at St. LouisZoo. Two males were placed in adjacent enclosures andaggressively chased one another along the fence. Two females,in adjacent enclosures to the males, came into estrus as a resultof using this technique and one subsequently produced a litter.Wielebnowski et al. [2002] also observed two separateinstances of ovulation in two females in response to thecourtship displays of males housed in nearby enclosures.Additionally, Ziegler‐Meeks [2009] suggested that coalitions ofmales are better able to investigate a female’s enclosure forsigns of estrus, and they appear more “behaviorally confident”than single males [Ziegler‐Meeks, 2009, p. 26]. Thus, thepresence of coalitions in captivity may be important forencouraging natural courtship behavior in both males andfemales [Brown et al., 1996]. Our results show that coalitions ofmale cheetahs can be successfully introduced andmaintained incaptivity. Further research is required in order to fullyunderstand the potential benefits of this practice for bothmale and female reproductive success.
CONCLUSIONS
1. Male cheetahs can be successfully introduced to formcoalitions in captivity.
2. Groups of male cheetahs can be housed in the same enclosurewith few aggressive interactions.
3. Housing male cheetahs in social groups allows them toexpress species‐specific social behavior.
4. Social group housing may have implications for cheetahwelfare and the success of captive breeding programs.
5. Social studies of captive animals using an association indexshould take into account chance encounters, especially whenenclosure size is small.
ACKNOWLEDGMENTS
We would like to thank Sonya Hill, Katalin Csatadi,and Mary Lherbier in the Chester Zoo Research Departmentand carnivore keepers Alan Woodward, Andy Wolfenden
and Laura Kelly for providing valuable information about thecheetah exhibit throughout the study. We are grateful to SeanO’Hara and two anonymous reviewers for their insightfulcomments on an earlier version of the manuscript.
REFERENCESAsa CS, Traylor‐Holzer K, Lacy RC. 2011. Can conservation‐breedingprograms be improved by incorporating mate choice? Int Zoo Yearbook45:203–212.
BaumanK, Blumer E, Crosier A, et al. 2010. Global cheetah ex situ planning:Linking Managed Populations Working Group. Conser Breed SpecialistGroup News 21:1–4.
Bircher JS, Noble GA. 1997. Management of cheetah (Acinonyx jubatus) atSaint Louis Zoological Park. Int Zoo Yearbook 35:51–58.
Brown JL, Wildt DE, Wielebnowski N, et al. 1996. Reproductive activity incaptive female cheetahs (Acinonyx jubatus) assessed by fecal steroids. JReprod Fertility 106:337–346.
Carlstead K, Shepherdson D. 1994. Effects of environmental enrichment onreproduction. Zoo Biol 13:447–458.
Caro TM. 1993. Behavioral solutions to breeding cheetahs in captivity:insights from the wild. Zoo Biol 12:19–30.
Caro TM. 1994. Cheetahs of the Serengeti plains: group living in an asocialspecies. Chicago: University of Chicago Press.
Caro TM, Collins DA. 1986. Male cheetahs of the Serengeti. Natl Geogr Res2:75–86.
Crosier AE,Marker L, Howard J, et al. 2007. Ejaculate traits in the Namibiancheetah (Acinonyx jubatus): influence of age, season and captivity. ReprodFertility Develop 19:370–382.
De RouckM, Kitchener AC, LawG, Nelissen M. 2005. A comparative studyof the influence of social housing conditions on the behavior of captivetigers (Panthera tigris). Anim Welf 14:229–238.
Durant SM, Kelly M, Caro T. 2004. Factors affecting life and deathin Serengeti cheetahs: environment, age and sociality. Behav Ecol 15:11–22.
Eaton R. 1970. Group interactions, spacing and territoriality in cheetahs. JCompar Ethol 21:481–491.
Ginsberg JR, Young TP. 1992.Measuring association between individuals orgroups in behavioral studies. Anim Behav 44:377–379.
Gottelli D, Wang J, Bashir S, Durant SM. 2007. Genetic analysis revealspromiscuity among female cheetahs. Proc R Soc B Biol Sci 274:1993–2001.
Laurenson MK, Caro T, Borner M. 1992. Patterns of female reproduction inwild cheetahs: implications for conservation. Natl Geogr Res Explor 8:64–75.
Lindburg DG, Durrant BS, Millard SE, Oosterhuis JE. 1993. Fertilityassessment of cheetahmales with poor quality semen. Zoo Biol 12:97–103.
Marker L. 2012. International cheetah (Acinonyx jubatus) studbook 2011.Otjiwarongo, Namibia: Cheetah Conservation Fund.
Marker‐Kraus L, Kraus D. 1997. Conservation strategies for the long‐termsurvival of the cheetah Acinonyx jubatus by the Cheetah ConservationFund, Windhoek. Int Zoo Yearbook 35:59–66.
Martin P, Bateson P. 2007. Measuring behavior: an introductory guide. 3rdedition. Cambridge: Cambridge University Press.
Mellen JD. 1991. Factors influencing reproductive success in small captiveexotic felids (Felis spp.): a multiple regression analysis. Zoo Biol 10:95–110.
Price EE, Stoinski TS. 2007. Group size: determinants in the wild andimplications for the captive housing of wild mammals in zoos. Appl AnimBehav Sci 103:255–264.
Ruiz‐Miranda CR,Wells SA, Golden R, Seidensticker J. 1998. Vocalizationsand other behavioral responses of male cheetahs (Acinonyx jubatus) duringexperimental separation and reunion trials. Zoo Biol 17:1–16.
Schülke O, Kappeler PM. 2003. So near and yet so far: territorial pairs butlow cohesion between pair partners in a nocturnal lemur, Phaner furcifer.Anim Behav 65:331–343.
Skibiel AL, Trevino HS, Naugher K. 2007. Comparison of several types ofenrichment for captive felids. Zoo Biol 26:371–381.
Terio KA, Marker L, Overstrom EW, Brown JL. 2003. Analysis of ovarianand adrenal activity in Namibian cheetahs. South Afr J Wildl Res 33:71–78.
526 Chadwick et al.
Zoo Biology

Tetley CL. 2009. Social group behavior of captive male cheetahs (Acinonyxjubatus soemmeringii) during the development of a new exhibit at ChesterZoo [MSc thesis]. Manchester: Manchester Metropolitan University.
Wachter B, Thalwitzer S, Hofer H, et al. 2011. Reproductive history andabsence of predators are important determinants of reproductive fitness: thecheetah controversy revisited. Conser Lett 4:47–54.
Whitehead H. 2008. Analyzing animal societies: quantitative methods forvertebrate social analysis. Chicago, Illinois: University of Chicago Press.
Wielebnowski NC. 1999. Behavioral differences as predictors of breedingstatus in captive cheetahs. Zoo Biol 18:335–349.
Wielebnowski NC, Brown JL. 1998. Behavioral correlates of physiologicalestrus in cheetahs. Zoo Biol 17:193–209.
Wielebnowski NC, Ziegler K,Wildt DE, Lukas J, Brown JL. 2002. Impact ofsocial management on reproductive, adrenal and behavioral activity in thecheetah (Acinonyx jubatus). Animal Conser 5:291–301.
Wildt DE, Brown JL, Bush M, et al. 1993. Reproductive status of cheetahs(Acinonyx jubatus) in North American Zoos: the benefits of physiologicalsurveys for strategic planning. Zoo Biol 12:45–80.
Ziegler‐Meeks K. 2009. Husbandry manual for the cheetah. Silver Spring,MD: AZA.
Coalition Behavior of Male Cheetahs 527
Zoo Biology




















