
A Coherent, Naturalistic, and Plausible Formulation of Libertarian Free Will
Upload
univ-lyon1Category
view
2download
0
Immediate Memory for ldquowhen where and whatrdquoShort-Delay Retrieval Using Dynamic Naturalistic
Material
Sze Chai Kwok123 and Emiliano Macaluso3
1Key Laboratory of Brain Functional Genomics Ministry of Education Shanghai KeyLaboratory of Brain Functional Genomics Institute of Cognitive Neuroscience School of
Psychology and Cognitive Science East China Normal University Shanghai China2NYU-ECNU Institute of Brain and Cognitive Science NYU-Shanghai University
Shanghai China3Neuroimaging Laboratory Fondazione Santa Lucia Istituto di Ricovero e Cura a Carattere
Scientifico (IRCCS) Rome Italy
r r
Abstract We investigated the neural correlates supporting three kinds of memory judgments aftervery short delays using naturalistic material In two functional magnetic resonance imaging (fMRI)experiments subjects watched short movie clips and after a short retention (15ndash25 s) made mne-monic judgments about specific aspects of the clips In Experiment 1 subjects were presented with twoscenes and required to either choose the scene that happened earlier in the clip (ldquoscene-chronologyrdquo)or with a correct spatial arrangement (ldquoscene-layoutrdquo) or that had been shown (ldquoscene-recognitionrdquo)To segregate activity specific to seen versus unseen stimuli in Experiment 2 only one probe image waspresented (either target or foil) Across the two experiments we replicated three patterns underlyingthe three specific forms of memory judgment The precuneus was activated during temporal-orderretrieval the superior parietal cortex was activated bilaterally for spatial-related configuration judg-ments whereas the medial frontal cortex during scene recognition Conjunction analyses with a previ-ous study that used analogous retrieval tasks but a much longer delay (gt1 day) demonstrated thatthis dissociation pattern is independent of retention delay We conclude that analogous brain regionsmediate task-specific retrieval across vastly different delays consistent with the proposal of scale-invariancein episodic memory retrieval Hum Brain Mapp 362495ndash2513 2015
Key words cinematic material scale-invariance short-long-delays ldquowhat-where-whenrdquo tripartitepattern
r r
Additional Supporting Information may be found in the onlineversion of this article
Correction added after online publication 12 March 2015 Theorder of the authorsrsquo affiliations has been corrected
Contract grant sponsor European Research Council (EuropeanUnionrsquos Seventh Framework (Program FP72007-2013) Contractgrant number ERC Grant Agreement 242809 (EM) Contract grantsponsor Italian Ministry of Health (Fondazione Santa Lucia)
Correspondence to Sze Chai Kwok Neuroimaging LaboratorySanta Lucia Foundation Via Ardeatina 306 00179 Rome ItalyE-mail sze-chaikwokst-hughsoxonorg
Received for publication 28 October 2014 Revised 10 February2015 Accepted 3 March 2015
DOI 101002hbm22787Published online 12 March 2015 in Wiley Online Library(wileyonlinelibrarycom)
r Human Brain Mapping 362495ndash2513 (2015) r
VC 2015
This is an open access article under the terms of the Creative Commons Attribution-NonCommercial License which permits usedistribution and reproduction in any medium provided the original work is properly cited and is not used for commercial purposes
The Authors Human Brain Mapping Published by Wiley Periodicals Inc
VC 2015 The Authors Human Brain Mapping
Published by Wiley Periodicals Inc
INTRODUCTION
In memory research there is a long-held distinctionbetween mechanisms involved in short-term memory(STM) and long-term memory [LTM see Shallice and War-rington 1970 for classic neuropsychological data inpatients] Baddeley and Hitch posited that there are multi-ple short-term buffers as temporary memory stores eachof them separate from the stores for long-term retention[1974] More recently we have seen an expansion of evi-dence challenging this dissociation [Jonides et al 2008Nee et al 2008 Ranganath and Blumenfeld 2005] Forexample it has been put forward that different local cir-cuits mediate STM vs LTM processes within the samecortical area [Gaffan 2002 Howard et al 2005] By theseviews the distinction between STM and LTM can beexplained through differences in the intraregional neuro-nal dynamics within the same memory areas [Fuster 20062009] or through differences in the level of activity withinthese common memory areasnetworks [Jonides et al2005 Postle 2006]
We sought to contribute to this debate by characterizingtask-specific functional dissociations during the immediateretrieval of ldquowhat-where-whenrdquo information using com-plex and naturalistic material [Holland and Smulders2011] In LTM previous neuroimaging studies have inves-tigated memory retrieval based on this classification [Bur-gess et al 2001 Ekstrom and Bookheimer 2007 Ekstromet al 2011 Fujii et al 2004 Hayes et al 2004 Nyberget al 1996] However the brain regions involved inattribute-specific retrieval do not appear to correspond fol-lowing long-term retention delays [Furman et al 2012Magen et al 2009] versus short-term delays [Mohr et al2006 Munk et al 2002] We consider that the lack of suchcorrespondence could arise from the fact that paradigmsused in taxing STM and LTM retrieval had been designeddifferently to accommodate the vast difference in delays
In STM studies the use of simple stimuli enabledresearchers to accurately control for many stimulus param-eters (eg the number of objects and stimulus position)but led to experimental conditions that substantially differfrom everyday situations that require the processing ofcomplex and dynamic sensory input While memory forsimple visual displays words and spatial positions isthought to be supported by capacity-limited buffers inwhich only a few items can be held [Baddeley and Hitch1974] memory for other types of material [eg unknownfaces and possibly scenes see Shiffrin 1973] which reflectmore closely the dynamic nature of real-life situationsmay tap into the same high-capacity system as long-termepisodic memory [Shallice and Cooper 2011]
The main aim of this study was to investigate task-specificfunctional dissociations for immediate memory retrievalusing naturalistic stimuli and to compare these with theresults of a previous LTM study that used analogous materialand tasks We devised a short-delay version of the paradigmthat we previously used to investigate different types of judg-
ment during retrieval from LTM [Kwok et al 2012] In theoriginal LTM paradigm participants watched a 42-min Tele-vision (TV) film and 24 h later made discriminative choicesof scenes extracted from the film during fMRI Subjects werepresented with two scenes and required to either choose thescene that happened earlier in the film or the scene with acorrect spatial arrangement or the scene that had beenshown This paradigm revealed a tripartite dissociationwithin the retrieval network [Hayama et al 2012] compris-ing the precuneus and the angular gyrus for temporal orderjudgments dorsal frontoparietal cortices for spatial configu-ration judgments and anteromedial temporal regions forsimple recognition of objectsscenes
Here we used modified versions of the original para-digm and tested for task-specific dissociations in two fMRIexperiments that included just 15ndash25 s between the endof the encoding phase and the retrieval phase In bothexperiments we asked the participants to watch short TVcommercial clips (772ndash114 s) and after a brief delay (15ndash25 s) to make memory judgments about the ldquotemporal-chronologyrdquo the ldquospatial-layoutrdquo or ldquoscene-recognitionrdquo ofthe encoded stimuli The stimuli for encoding comprisedvideo clips of naturalistic events which are unlikely to bestored into capacity-limited memory buffers and moreclosely reflect the dynamic nature of real-life situationsBecause of the stimulus complexity memory retrievalcould be supported by a combination of various strategiespotentially with each of them utilized to a different extentduring the three tasks With respect to this some differ-ence may be expected between the current studies andprevious STM studies that have used simpler and stereo-typed stimulimdashsuch as geometrical shapes or lists ofwordsmdashwhich permit a more rigorous control over proc-esses specific to the retrieval of temporal spatial andobject details [eg Harrison et al 2010 and see Discus-sion Section) Nonetheless for consistency with our previ-ous LTM study [Kwok et al 2012] in the sections belowwe adopted the original labels of ldquoTemporalrdquo ldquoSpatialrdquoand ldquoObjectrdquo tasks
In the first experiment (Exp 1) each retrieval trialincluded two scenes (still frames) presented side-by-sideand the participants were required to either choose thescene that happened earlier in the clip (ldquoscene-chronologyrdquojudgment Temporal task) or that with a correct spatialarrangement (ldquoscene-layoutrdquo judgment Spatial task) orthat had been shown during encoding (ldquoscene-recognitionrdquojudgment Object task) Thus the retrieval cues were iden-tical to those used in the previous LTM experiment whichenabled us to assess the imaging data using exactly thesame contrastssubtractions In the second experiment(Exp 2) we again assessed task-specific dissociations dur-ing immediate retrieval but now presenting only one sin-gle probe during retrieval (ie either a ldquoseenrdquo or anldquounseenrdquo image for scene-layout and the scene-recognitiontasks see Methods) With this we could compare the threeretrieval tasks using only trials including seenold probeimages avoiding any possible confound related to the
r Kwok et al r
r 2496 r
presentation of the unseen foil images Moreover the sub-stantial changes of the task structure across experiments 1and 2 enabled us to confirm any task-specific dissociationunder different sets of task-constraints
On the assumption that memory for complex naturalis-tic events depends negligibly on capacity-limited memorybuffers we hypothesized task-specific functional dissocia-tions during retrieval after short delays were analogous tothose observed during retrieval after long delays This wastested by identifying any task-specific brain activation dur-ing retrieval after short delays (ldquotask-specificrdquo analysesExp 1 and 2) and by comparing these effects with datafrom the previous LTM experiment [ldquoconjunction analy-sesrdquo between the current Exp 1 and LTM data from Kwoket al 2012] Findings of task-specific dissociations equiva-lent to those previously reported for LTM would supportthe view that memory for naturalistic material can accom-modate a vast range of retention delays [see also Mayloret al 2001 for scale-invariance in free recall memory) Inaddition we controlled that the task-specific effects atretrieval were not merely due to the new encoding ofunencountered material (ldquoseen-onlyrdquo analyses in Exp 2)and tested whether the task-specificity was determined bythe successful retrieval outcome or rather reflected theattempt to recover relevant mnemonic details [Rugg andWilding 2000] Confirmation of our hypotheses abouttask-specific functional dissociations during retrieval aftershort delays would indicate that the processes governingmemory for naturalistic events are similarly guided bytask-specific constraints across various retention delaysand provide evidence for the notion of scale-invariance inmemory functions [Brown et al 2007 Howard andKahana 2002]
MATERIALS AND METHODS
Overview
The trial structure of Exp 1 and Exp 2 were analogousthe subject watched a short video clip and then performedone of three retrieval tasks about some details of the clipThe retrieval tasks aimed at tapping into different proc-esses associated with the judgment of scene-chronology(Temporal task) scene-layout (Spatial task) or scene-recognition (Object task) The retrieval task was cued atthe onset of the probe images This cueing procedureensured that the subjects encoded the movie clips withoutanticipating what specific stimulus detail would be probedat retrieval (see Fig 1A)
In Exp 1 the retrieval phase comprised the presentationof two images side-by-side For the Temporal task bothscenes were extracted from the clip and the subject wasasked to choose the scene that had happened at an earliertime point in the clip (target) For the Spatial and Objecttasks the judgment at retrieval was between one scenethat had been presented (target) and another one that had
not been presented (foil) We investigated task-specific dis-sociations by directly comparing each of the tasks with theaverage of the other two tasks Further we performed con-junction analyses between the current dataset of Exp 1 anda previous LTM experiment that made use of the sameretrieval procedure but with a longer movie (42-min) anda retention delay of 24 h [Kwok et al 2012] This enabledus to formally test whether there was any functional over-lap between the patterns of task-specific dissociation fol-lowing short and long delays (15ndash25 s vs 24 h)
Exp 1 was designed to match the previous LTM study[Kwok et al 2012] thus including two images (target andfoil) in each retrieval trial However with this procedurethe retrieval-display of scene-recognition Object taskincluded an unseen image and the display of the scene-layout Spatial task included an unseen image-configuration The presentation of these unseen foils mayhave contaminated any task-specific effects associated withthe Spatial and Object tasks (eg incidental encoding ofnew images or new image-configurations) This wasaddressed in Exp 2 that entailed the presentation of onesingle image at retrieval and required the subject toanswer with a yesno response In the Spatial and theObject tasks the probe was either a seenold imageextracted from the video or an unseen image The Tempo-ral task always comprised the presentation of a probeimage extracted from the clip but the task now involvedjudging whether the image belonged to the first or secondhalf of the clip We again assessed task-specific dissocia-tions during immediate retrieval but now by comparingthe three tasks using only trials including seenold probeimages to avoid any possible confound related to theunseen images
Subjects
Seventeen participants took part in Exp 1 (mean age258 21ndash33 years 8 females) and another 17 different par-ticipants took part in Exp 2 (mean age 254 20ndash35 years 7females) All participants gave written informed consenthad normal or corrected-to-normal vision without knownhistory of psychiatricneurological problems and werefree from any MRI contraindication The study wasapproved by the Fondazione Santa Lucia (Scientific Insti-tute for Research Hospitalization and Health Care) Inde-pendent Ethnics Committee in accordance with theDeclaration of Helsinki
Stimuli
Movie clips
We purchased a collection of non-Italian TV commercialclips from an Advertising Archive (httpwwwcoloribuscom) and reproduced 160 short clips using a video editingsoftware (Final Cut Pro Apple Inc) Each clip was pur-posely edited to depict a coherent storyline with multiple
r Neural Correlates of Immediate Episodic Retrieval r
r 2497 r
switches of scenes The mean length was 959 s with arange of 772ndash1140 s We preserved the background soundand used only segments without dialoguesconversationsThe whole set of 160 clips was used in Exp 1 and each clipwas assigned to one of the three retrieval tasks 96 Tempo-ral 32 Spatial and 32 Object clips For Exp 2 we selected asubset of 144 clips and each clip was used for all the threetasks counterbalanced across subjects
Clip analyses
We performed a frame-by-frame analysis to mark thechanges of scene for each clip The scene changes includedboth between-scenes cuts (no scene continuity ie edit cutfrom one scene to another) and within-scenes cuts [scenecontinuity but with a change of camera perspective Smithand Henderson 2008] This frame-by-frame analysis allowed
Figure 1
Task schema and behavioral performance for Experiments 1 and
2 (A) Trial timeline probe images and cues at retrieval Each
trial carried an identical structure consisting of presentation of a
short movie clip followed by a cued retrieval test The retrieval
test comprised either a pair of probe images (Exp 1) or one sin-
gle probe (either target or foil Exp 2) (B) Accuracy presented
in bars ( correct unit on left ordinate axes) and response
times in lines (ms right ordinate axes) for Experiment 1 (left
panel) and Experiment 2 (right panel) Error bars are standard
error of the means
r Kwok et al r
r 2498 r
us to identify boundary frames that divided segmentswithin each clip The number of segments across all clipsranged between 4 and 10 The first and last segments werealways excluded for the retrieval tests to mitigate end-of-listdistinctiveness effects [Cabeza et al 1997 Konishi et al2002] Specifically for Exp 2 we divided each clip into 20segments with equal duration The memory probe imageswere extracted from 12 of these segments 3ndash5 (1st quartile)7ndash9 (2nd quartile) 12ndash14 (3rd quartile) and 16ndash18 (4th quar-tile) (cf Temporal task)
Probe images extraction
Exp 1 After excluding the first and last segments for theTemporal trials we pseudorandomly selected two imagesfrom two different segments from each temporal clip Thisrepresented a range of temporal distances between thetwo probe images across all the Temporal trials The tem-poral distances varied between 060 and 496 s The Spatialtrials were generated by pairing a target image with itsown mirror image (by laterally flipping the target image)The Object trials were generated by pairing a target imagewith an unseen image that also contained the same objectcharacter from an edited-out part of the original clip
Exp 2 Irrespective of task we selected one probe imagefrom each clip which was used as the target image Thisimage was extracted with an equal likelihood from one ofthe 4 quartiles (see Clip analyses above) thus with half ofthem coming from the first half and the other half fromthe second half of the clip For the Spatial task the foilimages were mirror images of the targets whereas for theObject task new images were extracted from an edited-outpart of the original clip as the foil images
Trial structure
The trial structure was identical in Exp 1 and 2 eachtrial began with a green fixation cross for 05 s followedby the presentation of a clip lasting for 772ndash114 s then ablank screen for a period of 15ndash25 s This was followedby a retrieval test (Fig 1A) The two experiments differedin how memory was probed during retrieval The retrievaltest of Exp 1 contained a pair of probe images and a writ-ten cue above the images both of which were presentedon the screen for 4 s whereas that of Exp 2 contained onlyone probe image and a written cue both presented for 5 sIn both experiments the written cue specified what taskthe subject had to perform on that trial Trials were sepa-rated by a white fixation cross with jittered ITIs sampledfrom a truncated logarithmic distribution (trial durationsranged between 207 and 228 s) We presented the stimuliusing Cogent Toolbox (httpwwwvislabuclacukcogentphp) implemented in MATLAB 74 (The Math-Works Natick MA) During fMRI scanning all stimuliwere back-projected on a screen at the back of the MRbore and viewed through a mirror system (20 3 15 visual
angle) Participants were given detailed written instruc-tions and were explained of the task requirements prior tothe commencement of the experiments
Retrieval Tasks
Experiment 1
The retrieval test included three main experimental con-ditions Temporal task (96 trials) Spatial task (32 trials)and Object task (32 trials) The reason for the larger num-ber of Temporal trials was that this task comprised twodifferent trial types (Tshort and Tlong) that were classifiedmdashand modeled separately in the main fMRI analysismdashaccording to the temporal distance between the two probeimages (see also Supporting Information Table 1) This dis-tinction was motivated by previous findings showing thatthe temporal separation of the probes at encoding canaffect retrieval performance and brain activity during tem-poral order judgments with naturalistic material [Kwoket al 2012 St Jacques et al 2008]
For all three tasks each retrieval trial comprised a pairof probe images arranged with a leftright side-by-sideconfiguration one of the images was the target the otherwas the foil The left-right position of the targetfoilimages was counterbalanced within conditions Subjectsindicated the leftright stimulus by pressing either one ofthe two keys of a response box Scene-chronology judg-ment (Temporal task T) Subjects were instructed toremember the order of the events in the clip so as tochoose the image that had happened at an earlier timepoint Scene-layout judgment (Spatial task S) Subjectswere instructed to focus on the spatial layout and choosethe probe image that had the identical spatial arrangementas depicted in the clip Scene-recognition judgment (Objecttask O) Subjects were instructed to focus on the contentof the probe images and to identify the image they hadseen in the clip
It should be noted that the retrieval of complex stimulias those used here can be achieved via many differentstrategies For example to solve the Spatial task partici-pants may attend to and store the location of a specificperson in the scene the facing direction of a car and soon rather than processing the overall spatial details withinthe scene For this reason we emphasize that the ldquolabelsrdquoused here (TemporalSpatialObject) do not imply anyexclusive strategy for solving the tasks but rather aremeant to refer to the specificity of the type of judgmentimplicated during retrieval
Experiment 2
The aim of Exp 2 was to test for task-specific dissocia-tions comparing trials that included only ldquoseenrdquo images(ie images extracted from the video) The retrieval testincluded the three main conditions (Temporal task 48 tri-als Spatial task 48 trials and Object task 48 trials) but
r Neural Correlates of Immediate Episodic Retrieval r
r 2499 r
now each trial comprised one single probe image Scene-chronology (Temporal) Subjects were required to recallthe probe imagersquos temporal location with respect to themidpoint of the clip by choosing between the ldquofirst halfrdquoor the ldquosecond halfrdquo of the clip Subjects were required topress the left button if the probe image happened in thefirst half or the right button if the image happened in thesecond half Scene-layout (Spatial) Subjects wereinstructed to focus on the spatial layout and to judgewhether the probe depicted a scene with a spatial arrange-ment as identical to the clip or it was a mirror image Anidentical spatial configuration required the subject to pressthe ldquocorrectrdquo button whereas a mirror image required apress to the ldquoincorrectrdquo button The mapping of ldquocorrectincorrectrdquo (leftright) layouts was counterbalanced acrosssubjects Scene-recognition (Object) Subjects wereinstructed to focus on the content of the probe image andto indicate whether the image had been shown in the clipIf it had been shown the subject responded with theldquopresentrdquo button otherwise with the ldquoabsentrdquo buttonAgain the leftright mapping of ldquopresentabsentrdquoresponses was counterbalanced between subjects
Since only a single image was now presented atretrieval the Temporal task could not involve any factorof ldquotemporal distancerdquo (cf TshortTlong in Exp 1) Insteadin Exp 2 we found that the behavioral performance wasaffected by the ldquotemporal positionrdquo of the probe imageduring encoding (easier if extracted from the startend ofthe clip harder if extracted from around the middle)Accordingly the temporal trials were now classified andmodeled separately in the fMRI analysis as Teasy and Thard
(see also Supporting Information Table 1)
fMRI Data Acquisition and Preprocessing
The functional images were acquired on a Siemens Alle-gra (Siemens Medical Systems Erlangen Germany) 3 Teslascanner equipped for echo-planar imaging (EPI) A quad-rature volume head coil was used for radio frequencytransmission and reception Head movement was mini-mized by mild restraint and cushioning Thirty-two slicesof functional MR images were acquired using blood oxy-genation level-dependent imaging (3 3 3 mm in-plane25 mm thick 50 distance factor repetition time 5 208 secho time 5 30 ms flip angle 5 70 FOV 5 192 mm acqui-sition order 5 continuous ascending) covering the wholecortex We acquired 1648 and 1488 fMRI volumes foreach subject in Exp 1 and 2 respectively Images wereacquired across 4 fMRI runs in both experiments The totalacquisition periods lasted for 55 and 49 min respectively
Data preprocessing was performed with SPM8 (wwwfilionuclacukspm) implemented on Matlab 74 (The Math-Works Natick MA) After having discarded the first 4volumes in each run images were realigned to correct forhead movements Slice-acquisition delays were correctedusing the middle slice as a reference Images were then
normalized to the MNI EPI template resampled to 3 mmisotropic voxel size and spatially smoothed using an iso-tropic Gaussian Kernel (FWHM 5 8 mm)
fMRI Analyses
Statistical inference was based on a random effectsapproach This comprised two steps first-level analysesestimating contrasts of interest for each subject followedby second-level analyses for statistical inference at thegroup-level [Penny and Holmes 2004] For each of thetwo experiments we used several different modelsdepending on the specific questions that we sought toaddress (see below and Supporting Information Table 1)
All the first-level subject-specific models included oneregressor modeling the presentation of the video clips irre-spective of task plus several regressors that modeled theretrieval phase according to the different conditions ofinterest (see leftmost column in Supporting InformationTable 1) The regressor modeling the ldquoencoding phaserdquo(Movie-regressor) was time-locked to the onset of eachvideo and had an event duration corresponding to theduration of each specific video (ie 772ndash1140 s) All theretrieval-related regressors were time-locked to the onsetof the probe image(s) and with a duration correspondingto the probe presentation at retrieval that is 4 s in Exp 1and 5 s in Exp 2 All regressors were convolved with theSPM8 canonical hemodynamic response function Timeseries were high-pass filtered at 128 s and prewhitened bymeans of autoregressive model AR(1) The sections belowdetail how the trials were assigned to the retrieval regres-sorsconditions in the different analyses
Experiment 1
For Exp 1 we used two different sets of first-level mod-els (see Supporting Information Table 1 leftmost column)The first set of models included the encoding-regressor(Movie) plus 5 retrieval-related conditions temporal-short(Tshort) temporal-long (Tlong) spatial (S) object (O) trialsand a condition modeling all the error trials irrespective oftask (Error) The trials belonging to the Temporal taskwere modeled with two separate regressors accounting forthe factor of temporal distance [Tshort vs Tlong cf Kwoket al 2012] All temporal trials including probes with atemporal distance shorter than 19 s were modeled asTshort while those longer than 19 s modeled as Tlong
The main second-level analysis consisted of a within-subjects Analysis of variance (ANOVA) modeling the foureffects of interest Tshort Tlong S and O conditions Foreach subject and each of the four conditions a contrastimage was computed in the first-level model by averagingthe corresponding parameter estimates across the 4 fMRIruns These subject-specific contrast images were enteredin the second-level ANOVA For the identification of task-specific retrieval effects we compared each task with themean of the two other tasks averaging the two Temporal
r Kwok et al r
r 2500 r
conditions ldquoT(short1long)2gt (S 1 O)2rdquo ldquoSgt (T(short1long)2 1 O)2rdquo and ldquoOgt (T(short1long)2 1 S)2rdquo Sphericity-correction was applied to all group-level ANOVAs toaccount for any nonindependent error term for repeatedmeasures and any difference in error variance across con-ditions [Friston et al 2002] The P-values were correctedfor multiple comparisons using a cluster-level threshold ofP-FWE-corr 5 005 considering the whole brain as the vol-ume of interest The cluster size was estimated using aninitial voxel-level threshold of P-unc 5 0001 To ensurethe specificity of any task effect the main differential con-trast was inclusively masked with 2 additional effectsThese corresponded to the activation for the critical taskvs each of the two other tasks for example the Temporalcontrast ldquoT(short1long)gt (S 1 O)rdquo was inclusively maskedwith ldquoT(short1long)2gt Srdquo and ldquoT(short1long)2gtOrdquo Forthese additional and nonindependent masking contraststhe threshold was set to P-unc 5 005 Together with themain contrasts that compared the three retrieval taskswithin this ANOVA we also tested for the effect ofldquotemporal distancerdquo (TshortgtTlong)
Using the same first-level models we ran a secondwithin-subject ANOVA that now also included the averageresponse times (RT) for each condition per subject as acovariate of no interest (Supporting Information Table 1ldquoControl for task difficultyrdquo) The aim of this was to reas-sess the pattern of task-specific dissociation after havingaccounted for any differences in task difficulty (Fig 1B)This RT-controlled analysis was not independent of themain analysis and the results are reported in the Support-ing Information
Experiment 2
The aim of Exp 2 was to account for the possible influ-ence of ldquounseennewrdquo images that were presented as foilsduring the retrieval phase of the Spatial and Object tasksin Exp 1 (eg incidental encoding during retrieval) InExp 2 a single probe image was presented during theretrieval phase of the trial (Fig 1A Experiment 2) Thisprocedure imposed a new constraint on the Temporaltask which now could not include the factor of ldquotemporaldistancerdquo Instead we taxed temporal retrieval by askingparticipants to judge whether the single probe image wasextracted from the first or second half of the clip Asexpected the participants were faster and more accurate tojudge probes presented either near the beginning or theend of the clip compared with probes extracted fromaround the middle of the clip Thus we now divided thetemporal trials into Teasy and Thard accounting for thisbehavioral effect
For Exp 2 we performed one single set of first-levelmodels and a single second-level group-analysis The first-level models included the encoding-regressor (Movie) and6 retrieval-related regressors Teasy (probe extracted fromthe 1st or 4th quartile) Thard (probe extracted from the2nd or 3rd quartile) Ssame Sflipped Oold and Onew plus
one regressor modeling all the error trials (see SupportingInformation Table 1)
The group level analysis consisted of a within-subjectsANOVA that included the 6 conditions of interest TeasyThard Ssame Sflipped Oold and Onew Within this ANOVAwe first tested for the task-specific effects by comparingeach task vs the mean of the other two tasks In this initialanalysis we averaged the Teasy and Thard conditions ofthe Temporal task as well as seen and unseen images ofthe Spatial and Object tasks for example[T(easy1hard)gt (S(same1flipped) 1 O(old1new))2] The statisticalthresholds used here were the same as Exp 1 statisticalmaps were corrected for multiple comparisons using acluster-level threshold of P-FWE-corr 5 005 (whole-brain)The cluster size was estimated using an initial voxel-levelthreshold of P-unc 5 0001 Again all comparisons wereinclusively masked with the effect of the main task vseach of the two other tasks (see Exp 1 above for details)
Within the same ANOVA a second set of contrasts soughtto replicate the task-specific effects but now using onlyseenold images (Supporting Information Table 1 ldquoSeen-onlyimagesrdquo) hence excluding any possible confound associatedwith the presentation of unseen images during retrieval Weused the following contrasts [T(easy1hard)gt (Ssame 1 Oold)][Ssamegt (T(easy1hard)2 1 Oold)2] and [Ooldgt (T(easy1hard)2 1 Ssame)2] The statistical thresholds and masking proce-dure were identical to Exp 1 and Exp 2 initial analysis
In Exp 2 we also performed control tests to rule out thepossibility that our main findings of task-specific dissocia-tions merely reflected differences in task difficulty (Sup-porting Information Table 1 ldquoTask difficultyrdquo) This wasaddressed by comparing conditions that behaviorallyresulted in difficult vs easy retrieval which included thecorresponding ldquohardrdquo versus ldquoeasyrdquo trials for the Tempo-ral task as well as by trials including ldquoflippedrdquo versusldquosamerdquo images in the Spatial task (Fig 1B) The results ofthese additional analyses are reported in the SupportingInformation
Conjunction analyses Between immediate
memory and LTM
To compare the current findings about task-specificretrieval after short delays with previous data using analo-gous retrieval tasks but a much longer retention delay[24 h Kwok et al 2012] we performed ldquobetween-studiesrdquo analyses Please note that the two paradigms dif-fered not only in terms of the retention delay (which dif-fered by a factor of 43200) but also in the presenceamount of interleaving events between encoding andretrieval In the LTM version participants were exposed to24 h of new episodic experiences following encodingwhich might cause retroactive interference on the alreadyacquired information [Wohldmann et al 2008] whereassuch retroactive interference was absent in the currentstudy on immediate retrieval The video clips were alsoshorter by a factor of about 250 compared to the 42-min
r Neural Correlates of Immediate Episodic Retrieval r
r 2501 r
episode used in the LTM paradigm Because of these differ-ences our analyses focused on commonalities rather thandifferences between LTM and immediate retrieval Theseanalyses considered the current Exp 1 which included thesame retrieval tasks as the LTM experiment Separately foreach of the three tasks we computed the relevant contrasts(eg Tgt (S1O)2) in the two studies and entered these inthree separate two samples t-tests For the identification ofcommon activation across studies we performed null-conjunction analyses (eg [Tgt (S1O)2]LTM [Tgt (S1O)2]ImMem) Statistical threshold were set at P-FWE-corr 5 005 cluster-level corrected at the whole brain level(cluster size estimated and displayed at P-unc 5 0001) con-sistent with all the analyses above For completeness thecontrasts testing for any differences between studies arereported in Supporting Information Table 2
Additional control analyses Retrieval success
Finally we asked to what extent any task-specific effectsidentified in our main analyses depended on the success-ful retrieval of stored information in the clips For this wecarried out additional analyses testing for retrieval per-formance in Exp 1 (ldquoTask 3 Accuracyrdquo in T and S tasks)and for accurate performance on seenold vs unseennewstimuli in Exp 2 (ldquoTask 3 Seen-unseenrdquo in S and O tasks)Please see Supporting Information for details of these anal-yses and the corresponding results
RESULTS
Behavioral Results
Experiment 1
Subjects performed significantly above chance in allthree tasks (all Plt 0001) The percentage of correct accu-racy and response times (RT) were analyzed usingrepeated measures ANOVAs with three levels correspond-ing to the TSO tasks This showed a main effect of con-ditions both for accuracy [F(2 32) 5 8142 Plt 0001] andRT [F(232) 5 3123 Plt 0001] Post hoc t-tests revealedthat subjects performed significantly better and faster inthe Object than Spatial task [Accuracy t(16) 5 1311Plt 0001 RT t(16) 5 652 Plt 0001] and Object than Tem-poral task [Accuracy t(16) 5 979 Plt 0001 RT t(16) 5
671 Plt 0001] Subjects were also more accurate on Tem-poral trials than on Spatial trials [Accuracy t(16) 5 348Plt 0001 but no difference in RT Pgt 01] For the Tempo-ral task we also examined the effect of temporal distanceexpecting that the retrieval would be more difficult whenthe two probes were temporally close (Tshortlt 19 s) com-pared with temporally far (Tlonggt 19 s) during encodingIndeed t-tests showed that subjects responded less accu-rately and slower in short than in long distance trials[Accuracy t(16) 5 391 Plt 0005 RT t(16) 5 315 Plt 001](Fig 1B plot on the left)
Experiment 2
In Exp 2 subjects also performed better than chance inall three tasks (all Plt 0001) Repeated measures ANOVAsshowed main effects of tasks both for accuracy [F(232) 5 919 Plt 0005] and RT [F(232) 5 1766 Plt 0001]Post hoc t-tests revealed that subjects performed withhigher accuracy and shorter RT in the Object comparedwith the Spatial task [Accuracy t(16) 5 406 Plt 0001 RTt(16) 5 532 Plt 0001] and in the Object compared withthe Temporal task [Accuracy t(16) 5 277 Plt 005 RTt(16) 5 383 Plt 0005] The difference in accuracy betweenthe Temporal and Spatial tasks approached significance(Plt 009) while there was no RT difference between Tem-poral and Spatial tasks (Pgt 01) Additional t-tests directlycompared different trial types within each task For theTemporal task subjects performed more accurately andfaster in the ldquoeasyrdquo than ldquohardrdquo trials [Accuracyt(16) 5 657 Plt 0001 RT t(16) 5 527 Plt 0001)] For theSpatial task they were more accurate in recognizingldquosamerdquo than correctly rejecting ldquoflippedrdquo images [Accu-racy t(16) 5 534 Plt 0001 RT Pgt 01] For the Objecttask subjects were faster to respond to unseen Onew
images compared to seen Oold images [RT t(16) 5 294Plt 001] without any difference in accuracy (Pgt 01) (Fig1B plot on the right)
Functional Neuroimaging Task-Specific Retrieval
In both Exp 1 and 2 we assessed brain activity specificto each of the three retrieval tasks (Temporal Spatial andObject) by comparing each task versus the mean of theother two (ldquoTask specificityrdquo contrasts see SupportingInformation Table 1) In Exp 2 we also carried out addi-tional nonindependent control tests using only trials con-taining seenold images (ldquoSeen-only imagesrdquo contrasts)The latter served to ensure that any task-specific dissocia-tion did not arise from the presentation of unseennewimages during the retrieval phase of the Spatial and Objecttasks (ie flipped- and new-image foils)
Scene-chronology judgment (Temporal task)
Exp 1 The comparison of the Temporal task with theother two retrieval conditions revealed significant activa-tion in the right precuneus bilateral caudate and bilateralcuneus (see Table I and Fig 2A clusters in bluemagenta) The signal plots in Figure 2 (panels on the left)show that the precuneus and the right angular gyrus wereactivated more by the Temporal task (average of Tshort andTlong) than by the Spatial and Object tasks Note that theeffect in the right angular gyrus was detected only atuncorrected threshold (P-unclt 0001) and it is reportedhere because it was fully significant in Exp 2 (see below)and as well as in the LTM dataset [Kwok et al 2012]
Together with the main comparison for the Temporaltask in Exp 1 we assessed the effect of temporal distance
r Kwok et al r
r 2502 r
TA
BL
EI
Task
-sp
ecif
icre
trie
val
acti
vati
on
Ex
p
1E
xp
2
Ex
p
2S
een
-on
ly
Clu
ster
Vo
xel
Clu
ster
Vo
xel
Clu
ster
Vo
xel
KP
-co
rr
Zx
yz
kP
-co
rr
Zx
yz
KP
-co
rr
Zx
yz
Sce
ne-
chro
no
log
y(T
emp
ora
l)P
recu
neu
sB
258
lt00
01
71
06
254
47
2666
lt00
01
51
74
272
43
1835
lt00
01
40
06
270
41
Po
st
cin
gu
late
cort
exB
mdashmdash
mdashmdash
564
02
3027
535
62
3631
Cau
dat
eB
148
000
54
302
122
1514
mdashmdash
mdashmdash
mdashmdash
mdashmdash
Cu
neu
sL
820
052
522
215
260
17mdash
mdashmdash
mdashmdash
mdashmdash
mdashC
un
eus
R88
004
25
2118
257
20mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nto
po
lar
cort
exR
mdashmdash
mdashmdash
874
lt0
001
547
2860
970
60
001
575
2860
7S
up
erio
rfr
on
tal
cort
exR
mdashmdash
mdashmdash
685
000
15
3526
2455
889
000
15
6026
2853
An
gu
lar
gy
rus
R43
lt00
01
46
345
269
29
457
00
07
48
444
266
45
509
00
04
45
844
268
43
Sce
ne-
lay
ou
t(S
pat
ial)
Su
peri
or
pari
eta
lg
yru
sL
1142
lt00
01
69
02
21
266
59
2530
lt00
01
66
72
18
268
59
377
00
16
49
92
18
266
59
Do
rsal
occ
ipit
alco
rtex
L5
112
272
759
521
228
280
9mdash
mdashmdash
mdashS
up
ram
arg
inal
gy
rus
L6
212
602
3338
mdashmdash
mdashmdash
mdashmdash
Intr
apar
ieta
lsu
lcu
sL
577
242
239
415
562
362
4445
mdashmdash
mdashmdash
Su
peri
or
pari
eta
lg
yru
sR
1314
lt00
01
67
221
263
59
3026
lt00
01
75
516
266
59
1262
lt00
01
47
426
268
47
Do
rsal
occ
ipit
alco
rtex
R4
1230
293
237
4034
278
294
1030
274
39In
trap
arie
tal
sulc
us
R6
2536
239
4433
40
026
548
402
4045
mdashmdash
mdashmdash
Su
pra
mar
gin
alg
yru
sR
597
602
2435
mdashmdash
mdashmdash
mdashmdash
Su
per
ior
fro
nta
lsu
lcu
sL
213
000
16
452
243
6223
80
078
543
226
22
51mdash
mdashmdash
mdashS
up
erio
rfr
on
tal
sulc
us
R18
20
002
592
302
356
220
009
74
7028
053
mdashmdash
mdashmdash
Infe
rio
rfr
on
tal
gy
rus
L92
003
65
002
516
26mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nta
lg
yru
sR
304
lt00
01
64
054
12
11
536
00
03
60
752
12
25
238
00
78
48
050
12
25
Infe
rio
ro
ccip
ital
gy
rus
L152
00
05
48
12
57
260
210
894
lt00
01
60
32
52
264
211
591
00
02
50
32
50
266
211
Infe
rio
ro
ccip
ital
gy
rus
R186
00
02
67
754
260
213
2277
lt00
01
68
256
258
211
1161
lt00
01
48
750
258
217
Occ
ipit
alp
ole
B21
80
001
484
29
293
2mdash
mdashmdash
mdashmdash
mdashmdash
mdashS
cen
e-re
cog
nit
ion
(Ob
ject
)M
ed
ial
fro
nta
lco
rtex
B478
lt00
01
51
92
351
47
2215
lt00
01
59
62
856
39
314
00
32
46
42
10
56
39
Mid
dle
tem
po
ral
gy
rus
L490
lt00
01
59
42
60
212
225
1661
lt00
01
67
72
62
214
219
162
lt00
01
42
92
60
24
225
Mid
dle
tem
po
ral
gy
rus
R304
lt00
01
61
560
212
216
1017
lt00
01
56
960
26
223
15
lt00
02
29
662
24
223
Orb
ito
-fro
nta
lco
rtex
B95
001
45
040
512
2222
70
049
506
044
219
mdashmdash
mdashmdash
An
gu
lar
gy
rus
L22
70
001
530
251
254
29mdash
mdashmdash
mdashmdash
mdashmdash
mdashA
ng
ula
rg
yru
sR
142
000
64
9857
260
38mdash
mdashmdash
mdashmdash
mdashmdash
mdashR
SC
p
ost
ci
ng
ula
teco
rtex
B47
4lt
000
14
903
242
35mdash
mdashmdash
mdashmdash
mdashmdash
mdash
T-c
on
tras
tsco
mp
ared
each
con
dit
ion
ver
sus
the
mea
no
fth
eo
ther
two
con
dit
ion
s(e
g
T(s
ho
rt1
lon
g)
2gt
(S1
O)
2)
and
was
incl
usi
vel
ym
ask
edw
ith
the
acti
vat
ion
for
the
crit
ical
con
dit
ion
ver
sus
each
of
the
two
oth
erco
nd
itio
ns
(eg
T
(sh
ort
1lo
ng
)2gt
S2
and
T(s
ho
rt1
lon
g)
2gt
O
2)
InE
xp2
we
con
du
cted
two
sep
arat
ese
tso
fco
ntr
asts
atth
eg
rou
p-l
evel
F
irst
t-
con
tras
tsw
ere
con
du
cted
usi
ng
all
six
con
dit
ion
sto
com
par
eea
chco
nd
itio
nv
ersu
sth
em
ean
of
the
oth
ertw
oco
nd
itio
ns
(eg
S (
sam
e1
flip
ped
)gt
[T(e
asy
1h
ard
)1
O(o
ld1
new
)]
2)
see
colu
mn
sin
the
mid
dle
Se
con
d
for
Spat
ial
and
Ob
ject
con
dit
ion
sw
ew
ere
inte
rest
edin
acti
vit
yel
icit
edb
yth
ere
spo
nse
sas
soci
ated
wit
hldquos
een
rdquotr
ials
on
ly(e
g
S sa-
megt
[T(e
asy
1h
ard
)2
1O
old
]2)
se
eri
gh
tmo
stco
lum
ns
and
also
Sup
po
rtin
gIn
form
atio
nT
able
1B
rain
reg
ion
sh
igh
lig
hte
din
bo
ldco
rres
po
nd
toth
eta
sk-s
pec
ific
effe
cts
con
sist
entl
yo
bse
rved
acro
ssal
lex
per
imen
ts
anal
yse
sth
ese
reg
ion
sar
eal
sod
isp
lay
edin
Fig
ure
s2ndash
4P
-val
ues
are
corr
ecte
dfo
rm
ult
iple
com
par
iso
ns
exce
pt
for
the
rig
ht
ang
ula
rg
yru
sin
Exp
1an
dth
em
idd
lete
mp
ora
lg
yri
inE
xp2
Seen
-on
lyth
atar
ere
po
rted
ata
low
eru
nco
rrec
ted
thre
sho
ld(i
nit
alic
s)
Po
st
po
ster
ior
xy
zco
ord
inat
esin
the
stan
dar
dM
NI
spac
eo
fth
eac
tiv
atio
np
eak
sin
the
clu
ster
sk
nu
mb
ero
fv
oxe
lsin
each
clu
ster
R
SC
retr
osp
len
ial
cort
ex
L
R
B
left
ri
gh
th
emis
ph
ere
bil
ater
al
r Neural Correlates of Immediate Episodic Retrieval r
r 2503 r
by comparing Tshort vs Tlong trials In previous studiesusing LTM protocols we found that temporal distance fur-ther modulated activity in the precuneus [Kwok et al2012 2014 see also St Jacques et al 2008] Here this con-trast did not reveal any significant effect This is in con-trast to the significant behavioral cost of retrieving Tshort
vs Tlong temporal distances (see behavioral results andFig 1B plot on the left) that was analogous to our previ-ous LTM protocol Similar behavioral patterns have beenobserved in many other short-term studies [Hacker 1980Konishi et al 2002 Milner et al 1991 Muter 1979] yet toour knowledge there does not seem to exists a clear con-
sensus on the neural correlates subserving the temporaldistance effect separating two temporally very nearbyeventsitems
Exp 2 In Exp 2 we replicated the effect of the temporalretrieval task in the precuneus and found a fully signifi-cant effect in the right angular gyrus (see Fig 2 clusters inredmagenta) In addition in Exp 2 there was a significantactivation of the right superior frontal cortex and the rightinferior frontopolar cortex that were not found in Exp 1(Table I) The involvement of the precuneus and the rightangular gyrus was further confirmed using only trials
Figure 2
Clusters of brain activation and signal plots for the scene-
chronology judgment (Temporal task) Clusters of activation and
signal plots for (A) the precuneus and (B) the right angular
gyrus These regions activated selectively during the temporal
tasks in Exp 1 (in blue) Exp 2 (in red overlap between Exp 1
and 2 shown in magenta) The activation selective for the Tem-
poral task was computed with the contrast T(short1long)
2gt (S 1 O)2 inclusively masked with T(short1long)2gt S2 and
with T(short1long)2gtO2 The involvement of the precuneus and
the right angular gyrus was further confirmed using only ldquoseen
oldrdquo images (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 40 voxels Effect sizes are mean adjusted (sum to
zero) and expressed in arbitrary units (au 6 90 CI) LR left
right PreC precuneus AnG angular gyrus
r Kwok et al r
r 2504 r
including seen images (cf ldquoExp 2 Seen- onlyrdquo Table I)The signal plots on Figure 2 (panels on the right) show theactivity of the precuneus and the right angular gyrus forall the conditions of Exp 2 Both in the precuneus and theright angular gyrus the average activity in the temporaltask [T(easy1hard)2] were larger than the Ssame and theOold conditions Notably Exp 2 included the presentationof a single image and that the participants were asked tojudge the absolute temporal position of the probe ratherthan the relative temporal order Despite this importantchange of how temporal memory was probed the findingof analogous patterns of activation across both experi-ments highlights the link between immediate retrieval oftemporal information and activation of the precuneus andangular gyrus
Scene-layout judgment (Spatial task)
Exp 1 The contrast concerning the retrieval of the spatiallayout revealed activation in the superior parietal cortexbilaterally with the cluster extending to the intraparietalsulcus and the supramarginal gyrus This contrast alsoshowed activation of the superior frontal sulci the inferiorfrontal gyri plus the inferior occipital cortex (see Table Iand Fig 3 clusters displayed in bluemagenta signalsplots on the left side)
Exp 2 The same comparison in Exp 2 replicated theresults of Exp 1 showing activation in the superior parie-tal cortex bilaterally the superior frontal sulci the inferiorand dorsal occipital cortices and the right inferior frontalgyrus (Fig 3 clusters in redmagenta) When we consid-ered only the conditions including the seenold images(ldquoExp 2 Seen- onlyrdquo) we replicated the effects in the supe-rior parietal gyri the inferior occipital gyri and the rightinferior frontal gyrus (see Table I and Fig 3 signal plotson the right) The activation in the intraparietal sulci andthe superior frontal sulci did not reach statistical signifi-cance Inspection of the signal plots indicated that in theseareas there was less activation in the Ssame than Sflipped tri-als (cf bar 3 vs bar 4 signal plots of the superior fontalsulcus Fig 3A plot on the right) Nonetheless the activityin the Ssame condition was larger than those in the T andO conditions (cf bar 3 vs bars 1ndash2 and 5ndash6)
Scene-recognition judgment (Object task)
Exp 1 The Object task was associated with the activationof the medial frontal cortex (MFC) and the orbitofrontalcortex (but the latter did not fully replicate in Exp 2 seeTable I and Fig 4 clusters bluemagenta and signal plotson the left) together with bilateral activation in the middletemporal gyri the retrosplenialposterior cingulate gyrusand a portion of the angular gyrus The same contrast alsorevealed an activation of the hippocampus bilaterally Theactivation of the hippocampus was just below statisticalsignificance (P-FWE-corr 5 0052 Z 5 458 at x y z 5 2126 219 and P-FWE-corr 5 0077 Z 5 458 at x y z 5 224
29 225 considering the whole brain as the volume ofinterest) and it is reported here because the same effectwas previously found in the LTM dataset [Kwok et al2012 see also Discussion Section]
Exp 2 An almost identical pattern of activation wasfound in Exp 2 when we considered all the conditionsO(old1new)gt (T(easy1hard) 1 S(same1flipped))2 Significanteffects were found in the middle temporal gyri and alarge cluster including the medial frontal gyrus extendingventrally into the orbital-frontal cortex (Fig 4 clusters inredmagenta) However when we considered only the tri-als including seenold-images [Ooldgt (T(easy1hard)2 1 Ssame)2] the Object task was found to significantlyactivate only the MFC (Table I rightmost columns) indi-cating that this region was not affected by incidentalencoding
In summary across all three tasks the brain activationwas found to be similar across Exp 1 and Exp 2 Thescene-chronology Temporal task activated the precuneusand the right angular gyrus the scene-layout Spatial taskwas associated with activation of the posteriorsuperiorparietal cortex and the right inferior frontal gyrus whilethe scene-recognition Object task activated the MFC andthe middle temporal gurus (see Table I areas highlightedin bold) The control analysis of Exp 2 that consideredonly trials including seenold images confirmed thesetask-specific effects indicating that the pattern of dissocia-tionmdashand in particular for the Spatial and Object tasksmdashwas not merely driven by the unseennew images duringthe retrieval phase
Functional Neuroimaging Correspondence with
An Analogous LTM Dataset
We directly compared the task-specific dissociationobserved in the current dataset (Exp 1) with our previousLTM study that included the same task-specific retrievaltasks but after a much longer retention delay [ie 24 hbetween encoding and retrieval Kwok et al 2012] Com-mon across the two data sets the conjunction analysesrevealed an activation cluster in the precuneus for theTemporal task (Fig 5A and Table II) and the dorsal fron-toparietal network plus right inferior frontal gyrus andright occipital cortex for the Spatial task (Fig 5B and TableII) These formal tests show that the Temporal and Spatialtask-specific dissociation observed here for immediateretrieval matched well with the patterns observed duringretrieval from LTM By contrast for the Object task over-lapping effects in the MFC and in the hippocampus weredetected only at a more lenient uncorrected threshold ofP-uncorr 5 0001 In the hippocampus the lack of a fullysignificant ldquocommonrdquo effect across datasets was due to thefact that in the current Exp 1 the contrast comparing theObject task versus the two other tasks did not survive cor-rection for multiple comparisons considering the wholebrain as the volume of interest (P-corr 5 0052 and 0077
r Neural Correlates of Immediate Episodic Retrieval r
r 2505 r
Figure 3
Clusters of brain activation and signal plots for the Scene-layout
judgment (Spatial task) Clusters of activation and signal plots for
(A) the superior frontal cortex and the superior parietal cortex
bilaterally and (B) the right inferior occipital cortex and the right
inferior frontal cortex These regions activated for the Spatial task
in Exp 1 (blue) and in Exp 2 (red overlap between Exp 1 and Exp 2
in magenta) The activation selective for the Spatial task was com-
puted with the contrast Sgt [T(short1long)2 1 O]2 inclusively
masked with SgtT(short1long)2 and with SgtO The effects in the
superior parietal gyri the inferior occipital gyri and the right inferior
frontal gyrus were replicated when considering only sameold
images in Exp 2 (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 80 voxels Effect sizes are mean adjusted (sum to zero)
and are expressed in arbitrary units (au 6 90 CI) LR leftright
SFS superior frontal sulcus SPG superior parietal gyrus IFG infe-
rior frontal gyrus IOG inferior occipital gyrus
in the left and right hemispheres respectively cf above)Indeed the formal test assessing differences between thetwo datasets (ie ldquoTask 3 Experimentrdquo interactions seeSupporting Information Table 2) did not reveal any signifi-cant difference between LTM and immediate retrieval inthe hippocampus consistent with this region beinginvolved in the scene-recognition judgment irrespective ofthe retention delay (see also Discussion Section)
DISCUSSION
We have identified distinct sets of brain areas thatshowed task-specific activation during immediate retrievalfollowing the encoding of dynamic cinematic material Theprecuneus and the right angular gyrus were activatedwhile retrieving the temporal order of events (scene-chro-nology judgment) the superior parietal cortex wasrecruited during spatial related recall (scene-layout
Figure 4
Clusters of brain activation and signal plots for the Scene-
recognition judgment (Object task) Clusters of activation and
signal plots for (A) the MFC and (B) the MTG These regions
activated for the Object task in Exp 1 (blue) and in Exp 2 (red
overlap in magenta) The activation selective for the Object task
was computed with the contrast Ogt [T(short1long)2 1 S]2
inclusively masked with OgtT(short1long)2 and with Ogt S The
signal plots refer to the left hemisphere but analogous patterns
of activation were found in the right hemisphere (Table I) In
Exp 2 the task-specific effect of the Object task in the MFC
was replicated when considering only the sameold images
(Table I rightmost columns) Inspection of the signal plot for the
left MTG revealed less activity in the Oold than Onew trials (plots
on the right bars 5 and 6) although the Oold trials showed
numerically larger parameter estimates than the T and S condi-
tions (see p-uncorr reported in italics in Table I rightmost col-
umns) Activation clusters were estimated and displayed at P-
unc 5 0001 thresholded at a minimum of 80 voxels Effect sizes
are mean adjusted (sum to zero) and expressed in arbitrary
units (au 6 90 CI) LR leftright MTG middle temporal
gyrus MFC medial frontal cortex
r Neural Correlates of Immediate Episodic Retrieval r
r 2507 r
judgment) whereas activity in the MFC and the middletemporal gyrus (MTG) reflected processes associated withscene-recognition judgments These task-specific effectsoccurred independently of whether subjects were asked tojudge seenold or unseennew stimuli (cf ldquoSeen-onlyimagesrdquo contrasts in Exp 2) and independently ofretrieval success and general task difficulty (see Support-ing Information) With respect to the hypothesized corre-spondence between immediate retrieval and retrieval fromLTM we found that these task-specific dissociationsare independent of the informationrsquos mnemonic history(24 h vs 2 s retention ie differing by 43200 times inmagnitude)
One important qualifier for the following discussion isthat memory retrieval in all our tasks depended minimallyon memory buffers compared to other simpler itemsdis-plays that are typically used in ldquostandardrdquo short-termworking memory tasks [Shallice and Cooper 2011 Shiffrin1973] In each trial the number of items presented during
encoding was clearly well beyond the 4 6 1 items stipulatedby classical STM models [Cowan 2001] While memories ofsome materials (eg words spatial positions simple visualdisplays) are supported by limited-capacity buffers [Badde-ley and Hitch 1974] memory for other categories [egunknown faces and scenes see Shiffrin 1973] will tap intoLTM when the task requires the encoding of two or moreitems [Shallice and Cooper 2011] For this reason werestrict our discussion on the tripartite task-specific dissoci-ations with reference to highly complex naturalisticmaterial
Precuneus and Inferior Parietal Cortex Support
Temporal Order Recollection
The scene-chronology judgment task activated the pre-cuneus in both Exp 1 and 2 despite the considerablechanges in the trial structure for probing the retrieval oftemporal information (ie relative temporal order betweentwo probes in Exp 1 vs absolute temporal position of asingle probe in Exp 2) The chronology-judgment taskrequired retrieving the event sequences of many complex
Figure 5
Task-specific retrieval activation common in studies after imme-
diate delays and a longer delay Activation for the LTM study
[24 h Kwok et al 2012] is depicted in green and activation
for the current immediate retrieval study (data from current
Exp 1) depicted in red Activation clusters for both studies were
estimated and displayed at P-unc 5 0001
TABLE II Common memory retrieval activation
ImMem LTM common effects
Cluster Voxel
k P-corr Z x y z
Scene-chronology (Temporal)Precuneus B 112 0011 472 3 257 47Angular gyrus R 98 lt 0001 400 54 254 17
Scene-layout (Spatial)Superior parietal gyrus L 206 0001 595 221 269 56Supramarginal gyrus L 129 0008 346 260 236 44Intraparietal sulcus L 487 236 245 41Superior parietal gyrus R 651 lt 0001 520 24 263 59Dorsal occipital cortex R 372 30 275 29Intraparietal sulcus R 520 42 239 47Supramarginal gyrus R 482 57 233 53Superior frontal sulcus L 100 0022 482 224 3 56Superior frontal sulcus R 145 0004 481 30 0 53Inferior frontal gyrus R 95 0027 475 54 9 26Inferior occipital gyrus R 98 0024 515 54 257 210
Task-specific activation after immediate delays (current Exp 1ImMem) and a previous study that used a long delay betweenencoding and retrieval [24 h LTM Kwok et al 2012] Statisticalthreshold were set at P-FWE-corr 5 005 (cluster-level) consider-ing the whole brain as the volume of interest Note that theldquobetween-studiesrdquo overlap activation in the right angular gyruswas found at a lower uncorrected threshold (in italics) but impor-tantly within this area no differences or interactions betweenImMem and LTM studies were found (see differential effectsbetween the two studies in Supporting Information Table 2) LRB leftright hemispherebilateral
r Kwok et al r
r 2508 r
stimuli (scenes) that entail high-level conceptualhierarchi-cal relationships between them [Swallow et al 2011 Zackset al 2001] The precuneus was previously found to acti-vate in our LTM protocol that used the same temporalorder judgment [Kwok et al 2012 and see also results ofthe conjunction analyses] The current finding of precunealactivation using very different (much shorter) delays fitswell with the view that these processes are not strictlyassociated with retrieval following long retention delayssuggesting that this medial parietal region is implicatedduring temporal order retrieval irrespective of delay (Fig5A and Table II) Specifically we link the effect of chrono-logical retrieval in the precuneus with processes involvingan active reconstruction of past events [see also St Jacqueset al 2008] and possibly memory-related imagery[Fletcher et al 1995 Ishai et al 2000] These processeswould characterize temporal order judgments using com-plex naturalistic material but not order-judgment withsimpler stimulus material which previous studies haveassociated with other brain regions [eg middleprefron-tal areas Suzuki et al 2002]
Together with the precuneus temporal-related judgmentalso gave rise to activation of the right angular gyrus(AG) One possible interpretation relates the inferior parie-tal cortex to the engagement of attentional processes[Majerus et al 2007 2010] The scene-chronology judg-ment of Exp 2 provided us with a further qualifier to thiskind of temporal-related attention Although the Temporaltask in Exp 2 did not require any explicit temporal orderjudgment (only classifying a scene into either the first orsecond half of the movie) an accurate decision still neces-sitated an evaluation of the target scene with many othersegments of the movie This operation is fundamentallydifferent from the evaluation implicated in the scene-layout judgment which required decisions on comparingbetween a single scene and its own correspondentencoded snapshot The latter involves visuospatial analysisof the configuration within scenes The AG may subservedifferent types of attention among which one of them islinked with memory search operations [see Majerus et al2010 2006 also showing coactivation of these two regionsduring order retrieval of simple stimuli after short delays]
Superior Parietal Cortex Evaluates Spatial
Details Against Memory Representation
The second pattern consistently found across experi-ments (Exp 1 and Exp 2 see Fig 3) and across studieswith different retention delays (conjunction betweenimmediate retrieval and LTM studies see Fig 5B andTable II) was the activation of the superior parietal lobule(SPL) during the scene-layout judgment Previous studiesshowed sustained activation in the parietal cortex duringworking memory delays [Courtney et al 1998] and thatactivity there correlates with memory load [Todd andMaroi 2004] Here the amount of information contained
in the clips was equated in the three tasks especially inExp 2 where we counterbalanced the assignment of mov-ies across tasks The subjects also could not anticipatewhat specific type of information would be tested atretrieval and needed to keep rehearsing different aspectsof the clips Because of this we deem unlikely that theactivation of SPL that here was specific for the scene-layout tasks reflected general rehearsal processes [Magenet al 2009] andor updating operations [Bledowski et al2009 Leung et al 2007]
Instead we suggest a role for the SPL in mediating aninterface between the internal memory representation andexternal information It has been proposed that the supe-rior parietal cortex is engaged in allocating top-downattention to orient to aspects of the available visual inputand on information retrieved from memory [Ishai et al2002 Lepsien and Nobre 2007 Summerfield et al 2006]In line with this we found SPL activation when the sub-jects had to make decisions by comparing between theirmemory of the encoded clips and the scene stimuli pre-sented at retrieval This procedure might act on somevisuospatial storage which is held by the lateral occipitalcortex and mediated by the intraparietal sulcus (Fig 3B)The immediate memory task here was analogous to theSpatial task of our previous LTM study [Kwok et al2012] which also required matchingcomparing the avail-able external visual input and internal information storedin memory [see also Nobre et al 2004] This interfaceinvolves a range of top-down processes such as monitor-ing and verification [Cabeza et al 2008 Ciaramelli et al2008] which are independent of the probe imagersquos mne-monic status as shown by the equated effect sizes forldquosamerdquo and ldquoflippedrdquo images conditions (see signals plotsin Fig 3 and also ldquoRetrieval successrdquo analyses of Exp 2 inSupporting Information)
Recognition Processes in the Medial Frontal
Cortex and the Temporal Cortex
The third retrieval task involved scene-recognition judg-ments and was associated with the activation of the MFCand the MTG Moreover in Exp 1 we found a statisticaltrend in the hippocampus bilaterally an area that we pre-viously associated with scene-recognition judgment fromLTM (see next section about ldquoPossible correspondencesbetween retrieval from short- and long-term memoryrdquo)
The MFC was engaged both Exp 1 and 2 but was notactivated in the other two tasks thus a broad prefrontalcortexworking memory correspondence [Tsuchida andFellows 2009] seems insufficient to explain this task-specific effect The scene-recognition task in Exp 2 taxedboth recognition of ldquooldrdquo scenes and correct rejection ofldquonewrdquo scenes suggesting that the MFC here did not dis-tinguish between the correct rejection of new information(unseennew trials) and the recognition of old imagesOne candidate cognitive process that is common to both
r Neural Correlates of Immediate Episodic Retrieval r
r 2509 r
types of processes is feeling-of-knowing (FOK) FOKdepend on the cue specification content retrieval andappraisal of the retrieved memory fragments but are inde-pendent of the ultimate determination of content appropri-ateness [Schnyer et al 2005] Using imaging data duringsentence completion recognition Schnyer et al [2005]modeled a FOK-related directional path The path impli-cated the temporal cortex and the hippocampus andengaged the ventro-MFC at the end which is consistentwith our findings for the Object task implicating the MFCthe middle temporal cortex and at a lower statisticalthreshold the hippocampus
By contrast the activity in the MTG was larger forunseennew images than for seenold memory probes(Fig 4B) This suggests that new encoding may have con-tributed to the activation of the MTG during the retrievalphase of the Object task in Exp 1 The scene-recognitiontask required a lower precision in terms of retrievingdetails about the encoded material and could be solved bysimple recognition [Wixted and Squire 2010] Thus alter-natively larger activation for ldquonew vs oldrdquo stimuli mayrelate to some reduced activation associated with familiar-ity processes on old trials [see Henson et al 1999 whoreported reduced familiarity related activation in middletemporal gyri bilaterally and Henson and Rugg 2003 forreview] However we should point out that in Exp 2 theunseen images were extracted from edited-out parts of theoriginal clips and therefore contained the same objectcharacter depicted in the target images Therefore wedeem more likely that the relational association betweenobjects rather than their presenceabsence in the sceneshelped subjects to distinguish targets from foils We thussuggest that the activation of the MTG reflects processesutilizing the relational details among objects embedded inthe probe images for reaching the recognition decision[Diana et al 2007 Finke et al 2008 Hannula et al 2006Olson et al 2006] Future studies should consider theinclusion of some correlative behavioral measures to indexfamiliarity processes to elucidate further this issue More-over while the Temporal and Spatial tasks required mak-ing relative comparisons about aspects of the encodedvideo-clip the scene-recognition judgment merely requiredjudging whether the image presented at retrieval was pres-ent (or not) within the clip This may entail different proc-essing dynamics in the Object task compared with theother two tasks (cf also differences in retrieval times) Thelatter could be addressed using more subtle experimentalmanipulations for the scene-recognition judgment forexample by using digitally-modified foil images where asingle object in the scene has been modifiedremoved
Possible Correspondences Between Retrieval
from Short- and Long-Term Memory
The overlap between our current study and the previousLTM study using analogous tasks and materials lends
weight to the notion of scale-invariance in memory mecha-nisms [Brown et al 2007] as have been previously dem-onstrated in free recall tasks where the rate of item recallwas unvarying across recall span (day week year) andeven across from the past to the prospective future [May-lor et al 2001] Those previous behavioral findings implythat scale-invariance is a form of self-similarity in that theholistic pattern can exist at multiple levels of magnification[and even across different memory systems eg Maylor2002] Here our data affirmed the importance of the poste-rior parietal cortex in both immediate and long-termretrieval [Berryhill et al 2011 2007] and further character-ized its involvement in dealing with the spatial relationsof objects within scenes [Bricolo et al 2000 Buiatti et al2011] Our data also demonstrated such short-long-termcorrespondence exists in the precuneusangular gyrus fortemporal order retrieval (cf scene-chronology judgmenttask) In contrast the bilateral hippocampus and MFC acti-vations did not reach full statistical significance in theImMem LTM conjunction analysis At first sight thismay indicate that the scene-recognition task tapped intodistinct recognition-based mechanisms as a function ofretention delay However we deem this unlikely becausethe direct comparison between the two datasets did notshow any significant effect in these regions (see Support-ing Information Table 2) Indeed a more targeted test ask-ing the question of whether the hippocampus clusterspreviously found in LTM activated also in the currentimmediate retrieval experiments revealed significanteffects in both immediate retrieval experiments here (Exp1 x y z 5 224 29 225 P-FWE-corrlt 0001 Z 5 458 x yz 5 21 29 222 P-FWE-corr 5 0001 Z 5 451 and in Exp2 x y z 5 224 214 221 P-FWE-corr 5 0004 Z 5 396using the LTM clusters for small volume correction) Theseadditional tests point to a more general issue of sensitivitythat future studies may address using different methodo-logical approaches such as multivariate methods that canbe more sensitive than the standard univariate analysesused here Moreover the former would enable capturingtask-specific effects that are expressed as patterns of dis-tributed activation rather than local changes of activity asreported here [eg Jimura and Poldrack 2012]
Task Difficulty and Retrieval Success
We used several control analyses to show that the task-specific patterns of activation and the correspondencebetween the current data and our previous LTM study donot merely reflect some effect related to general task diffi-culty (see Supporting Information) First areas involved inoverall task difficulty should show progressively greateractivation with progressively more difficult tasks In Exp 1this would entail SgtTgtO However the patterns in theprecuneus and the angular gyrus (with TgtOgt S Fig 2A)and the MFC (Ogt SgtT Fig 4A) did not fit with simplepredictions based on overall task difficulty We also
r Kwok et al r
r 2510 r
assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r

INTRODUCTION
In memory research there is a long-held distinctionbetween mechanisms involved in short-term memory(STM) and long-term memory [LTM see Shallice and War-rington 1970 for classic neuropsychological data inpatients] Baddeley and Hitch posited that there are multi-ple short-term buffers as temporary memory stores eachof them separate from the stores for long-term retention[1974] More recently we have seen an expansion of evi-dence challenging this dissociation [Jonides et al 2008Nee et al 2008 Ranganath and Blumenfeld 2005] Forexample it has been put forward that different local cir-cuits mediate STM vs LTM processes within the samecortical area [Gaffan 2002 Howard et al 2005] By theseviews the distinction between STM and LTM can beexplained through differences in the intraregional neuro-nal dynamics within the same memory areas [Fuster 20062009] or through differences in the level of activity withinthese common memory areasnetworks [Jonides et al2005 Postle 2006]
We sought to contribute to this debate by characterizingtask-specific functional dissociations during the immediateretrieval of ldquowhat-where-whenrdquo information using com-plex and naturalistic material [Holland and Smulders2011] In LTM previous neuroimaging studies have inves-tigated memory retrieval based on this classification [Bur-gess et al 2001 Ekstrom and Bookheimer 2007 Ekstromet al 2011 Fujii et al 2004 Hayes et al 2004 Nyberget al 1996] However the brain regions involved inattribute-specific retrieval do not appear to correspond fol-lowing long-term retention delays [Furman et al 2012Magen et al 2009] versus short-term delays [Mohr et al2006 Munk et al 2002] We consider that the lack of suchcorrespondence could arise from the fact that paradigmsused in taxing STM and LTM retrieval had been designeddifferently to accommodate the vast difference in delays
In STM studies the use of simple stimuli enabledresearchers to accurately control for many stimulus param-eters (eg the number of objects and stimulus position)but led to experimental conditions that substantially differfrom everyday situations that require the processing ofcomplex and dynamic sensory input While memory forsimple visual displays words and spatial positions isthought to be supported by capacity-limited buffers inwhich only a few items can be held [Baddeley and Hitch1974] memory for other types of material [eg unknownfaces and possibly scenes see Shiffrin 1973] which reflectmore closely the dynamic nature of real-life situationsmay tap into the same high-capacity system as long-termepisodic memory [Shallice and Cooper 2011]
The main aim of this study was to investigate task-specificfunctional dissociations for immediate memory retrievalusing naturalistic stimuli and to compare these with theresults of a previous LTM study that used analogous materialand tasks We devised a short-delay version of the paradigmthat we previously used to investigate different types of judg-
ment during retrieval from LTM [Kwok et al 2012] In theoriginal LTM paradigm participants watched a 42-min Tele-vision (TV) film and 24 h later made discriminative choicesof scenes extracted from the film during fMRI Subjects werepresented with two scenes and required to either choose thescene that happened earlier in the film or the scene with acorrect spatial arrangement or the scene that had beenshown This paradigm revealed a tripartite dissociationwithin the retrieval network [Hayama et al 2012] compris-ing the precuneus and the angular gyrus for temporal orderjudgments dorsal frontoparietal cortices for spatial configu-ration judgments and anteromedial temporal regions forsimple recognition of objectsscenes
Here we used modified versions of the original para-digm and tested for task-specific dissociations in two fMRIexperiments that included just 15ndash25 s between the endof the encoding phase and the retrieval phase In bothexperiments we asked the participants to watch short TVcommercial clips (772ndash114 s) and after a brief delay (15ndash25 s) to make memory judgments about the ldquotemporal-chronologyrdquo the ldquospatial-layoutrdquo or ldquoscene-recognitionrdquo ofthe encoded stimuli The stimuli for encoding comprisedvideo clips of naturalistic events which are unlikely to bestored into capacity-limited memory buffers and moreclosely reflect the dynamic nature of real-life situationsBecause of the stimulus complexity memory retrievalcould be supported by a combination of various strategiespotentially with each of them utilized to a different extentduring the three tasks With respect to this some differ-ence may be expected between the current studies andprevious STM studies that have used simpler and stereo-typed stimulimdashsuch as geometrical shapes or lists ofwordsmdashwhich permit a more rigorous control over proc-esses specific to the retrieval of temporal spatial andobject details [eg Harrison et al 2010 and see Discus-sion Section) Nonetheless for consistency with our previ-ous LTM study [Kwok et al 2012] in the sections belowwe adopted the original labels of ldquoTemporalrdquo ldquoSpatialrdquoand ldquoObjectrdquo tasks
In the first experiment (Exp 1) each retrieval trialincluded two scenes (still frames) presented side-by-sideand the participants were required to either choose thescene that happened earlier in the clip (ldquoscene-chronologyrdquojudgment Temporal task) or that with a correct spatialarrangement (ldquoscene-layoutrdquo judgment Spatial task) orthat had been shown during encoding (ldquoscene-recognitionrdquojudgment Object task) Thus the retrieval cues were iden-tical to those used in the previous LTM experiment whichenabled us to assess the imaging data using exactly thesame contrastssubtractions In the second experiment(Exp 2) we again assessed task-specific dissociations dur-ing immediate retrieval but now presenting only one sin-gle probe during retrieval (ie either a ldquoseenrdquo or anldquounseenrdquo image for scene-layout and the scene-recognitiontasks see Methods) With this we could compare the threeretrieval tasks using only trials including seenold probeimages avoiding any possible confound related to the
r Kwok et al r
r 2496 r
presentation of the unseen foil images Moreover the sub-stantial changes of the task structure across experiments 1and 2 enabled us to confirm any task-specific dissociationunder different sets of task-constraints
On the assumption that memory for complex naturalis-tic events depends negligibly on capacity-limited memorybuffers we hypothesized task-specific functional dissocia-tions during retrieval after short delays were analogous tothose observed during retrieval after long delays This wastested by identifying any task-specific brain activation dur-ing retrieval after short delays (ldquotask-specificrdquo analysesExp 1 and 2) and by comparing these effects with datafrom the previous LTM experiment [ldquoconjunction analy-sesrdquo between the current Exp 1 and LTM data from Kwoket al 2012] Findings of task-specific dissociations equiva-lent to those previously reported for LTM would supportthe view that memory for naturalistic material can accom-modate a vast range of retention delays [see also Mayloret al 2001 for scale-invariance in free recall memory) Inaddition we controlled that the task-specific effects atretrieval were not merely due to the new encoding ofunencountered material (ldquoseen-onlyrdquo analyses in Exp 2)and tested whether the task-specificity was determined bythe successful retrieval outcome or rather reflected theattempt to recover relevant mnemonic details [Rugg andWilding 2000] Confirmation of our hypotheses abouttask-specific functional dissociations during retrieval aftershort delays would indicate that the processes governingmemory for naturalistic events are similarly guided bytask-specific constraints across various retention delaysand provide evidence for the notion of scale-invariance inmemory functions [Brown et al 2007 Howard andKahana 2002]
MATERIALS AND METHODS
Overview
The trial structure of Exp 1 and Exp 2 were analogousthe subject watched a short video clip and then performedone of three retrieval tasks about some details of the clipThe retrieval tasks aimed at tapping into different proc-esses associated with the judgment of scene-chronology(Temporal task) scene-layout (Spatial task) or scene-recognition (Object task) The retrieval task was cued atthe onset of the probe images This cueing procedureensured that the subjects encoded the movie clips withoutanticipating what specific stimulus detail would be probedat retrieval (see Fig 1A)
In Exp 1 the retrieval phase comprised the presentationof two images side-by-side For the Temporal task bothscenes were extracted from the clip and the subject wasasked to choose the scene that had happened at an earliertime point in the clip (target) For the Spatial and Objecttasks the judgment at retrieval was between one scenethat had been presented (target) and another one that had
not been presented (foil) We investigated task-specific dis-sociations by directly comparing each of the tasks with theaverage of the other two tasks Further we performed con-junction analyses between the current dataset of Exp 1 anda previous LTM experiment that made use of the sameretrieval procedure but with a longer movie (42-min) anda retention delay of 24 h [Kwok et al 2012] This enabledus to formally test whether there was any functional over-lap between the patterns of task-specific dissociation fol-lowing short and long delays (15ndash25 s vs 24 h)
Exp 1 was designed to match the previous LTM study[Kwok et al 2012] thus including two images (target andfoil) in each retrieval trial However with this procedurethe retrieval-display of scene-recognition Object taskincluded an unseen image and the display of the scene-layout Spatial task included an unseen image-configuration The presentation of these unseen foils mayhave contaminated any task-specific effects associated withthe Spatial and Object tasks (eg incidental encoding ofnew images or new image-configurations) This wasaddressed in Exp 2 that entailed the presentation of onesingle image at retrieval and required the subject toanswer with a yesno response In the Spatial and theObject tasks the probe was either a seenold imageextracted from the video or an unseen image The Tempo-ral task always comprised the presentation of a probeimage extracted from the clip but the task now involvedjudging whether the image belonged to the first or secondhalf of the clip We again assessed task-specific dissocia-tions during immediate retrieval but now by comparingthe three tasks using only trials including seenold probeimages to avoid any possible confound related to theunseen images
Subjects
Seventeen participants took part in Exp 1 (mean age258 21ndash33 years 8 females) and another 17 different par-ticipants took part in Exp 2 (mean age 254 20ndash35 years 7females) All participants gave written informed consenthad normal or corrected-to-normal vision without knownhistory of psychiatricneurological problems and werefree from any MRI contraindication The study wasapproved by the Fondazione Santa Lucia (Scientific Insti-tute for Research Hospitalization and Health Care) Inde-pendent Ethnics Committee in accordance with theDeclaration of Helsinki
Stimuli
Movie clips
We purchased a collection of non-Italian TV commercialclips from an Advertising Archive (httpwwwcoloribuscom) and reproduced 160 short clips using a video editingsoftware (Final Cut Pro Apple Inc) Each clip was pur-posely edited to depict a coherent storyline with multiple
r Neural Correlates of Immediate Episodic Retrieval r
r 2497 r
switches of scenes The mean length was 959 s with arange of 772ndash1140 s We preserved the background soundand used only segments without dialoguesconversationsThe whole set of 160 clips was used in Exp 1 and each clipwas assigned to one of the three retrieval tasks 96 Tempo-ral 32 Spatial and 32 Object clips For Exp 2 we selected asubset of 144 clips and each clip was used for all the threetasks counterbalanced across subjects
Clip analyses
We performed a frame-by-frame analysis to mark thechanges of scene for each clip The scene changes includedboth between-scenes cuts (no scene continuity ie edit cutfrom one scene to another) and within-scenes cuts [scenecontinuity but with a change of camera perspective Smithand Henderson 2008] This frame-by-frame analysis allowed
Figure 1
Task schema and behavioral performance for Experiments 1 and
2 (A) Trial timeline probe images and cues at retrieval Each
trial carried an identical structure consisting of presentation of a
short movie clip followed by a cued retrieval test The retrieval
test comprised either a pair of probe images (Exp 1) or one sin-
gle probe (either target or foil Exp 2) (B) Accuracy presented
in bars ( correct unit on left ordinate axes) and response
times in lines (ms right ordinate axes) for Experiment 1 (left
panel) and Experiment 2 (right panel) Error bars are standard
error of the means
r Kwok et al r
r 2498 r
us to identify boundary frames that divided segmentswithin each clip The number of segments across all clipsranged between 4 and 10 The first and last segments werealways excluded for the retrieval tests to mitigate end-of-listdistinctiveness effects [Cabeza et al 1997 Konishi et al2002] Specifically for Exp 2 we divided each clip into 20segments with equal duration The memory probe imageswere extracted from 12 of these segments 3ndash5 (1st quartile)7ndash9 (2nd quartile) 12ndash14 (3rd quartile) and 16ndash18 (4th quar-tile) (cf Temporal task)
Probe images extraction
Exp 1 After excluding the first and last segments for theTemporal trials we pseudorandomly selected two imagesfrom two different segments from each temporal clip Thisrepresented a range of temporal distances between thetwo probe images across all the Temporal trials The tem-poral distances varied between 060 and 496 s The Spatialtrials were generated by pairing a target image with itsown mirror image (by laterally flipping the target image)The Object trials were generated by pairing a target imagewith an unseen image that also contained the same objectcharacter from an edited-out part of the original clip
Exp 2 Irrespective of task we selected one probe imagefrom each clip which was used as the target image Thisimage was extracted with an equal likelihood from one ofthe 4 quartiles (see Clip analyses above) thus with half ofthem coming from the first half and the other half fromthe second half of the clip For the Spatial task the foilimages were mirror images of the targets whereas for theObject task new images were extracted from an edited-outpart of the original clip as the foil images
Trial structure
The trial structure was identical in Exp 1 and 2 eachtrial began with a green fixation cross for 05 s followedby the presentation of a clip lasting for 772ndash114 s then ablank screen for a period of 15ndash25 s This was followedby a retrieval test (Fig 1A) The two experiments differedin how memory was probed during retrieval The retrievaltest of Exp 1 contained a pair of probe images and a writ-ten cue above the images both of which were presentedon the screen for 4 s whereas that of Exp 2 contained onlyone probe image and a written cue both presented for 5 sIn both experiments the written cue specified what taskthe subject had to perform on that trial Trials were sepa-rated by a white fixation cross with jittered ITIs sampledfrom a truncated logarithmic distribution (trial durationsranged between 207 and 228 s) We presented the stimuliusing Cogent Toolbox (httpwwwvislabuclacukcogentphp) implemented in MATLAB 74 (The Math-Works Natick MA) During fMRI scanning all stimuliwere back-projected on a screen at the back of the MRbore and viewed through a mirror system (20 3 15 visual
angle) Participants were given detailed written instruc-tions and were explained of the task requirements prior tothe commencement of the experiments
Retrieval Tasks
Experiment 1
The retrieval test included three main experimental con-ditions Temporal task (96 trials) Spatial task (32 trials)and Object task (32 trials) The reason for the larger num-ber of Temporal trials was that this task comprised twodifferent trial types (Tshort and Tlong) that were classifiedmdashand modeled separately in the main fMRI analysismdashaccording to the temporal distance between the two probeimages (see also Supporting Information Table 1) This dis-tinction was motivated by previous findings showing thatthe temporal separation of the probes at encoding canaffect retrieval performance and brain activity during tem-poral order judgments with naturalistic material [Kwoket al 2012 St Jacques et al 2008]
For all three tasks each retrieval trial comprised a pairof probe images arranged with a leftright side-by-sideconfiguration one of the images was the target the otherwas the foil The left-right position of the targetfoilimages was counterbalanced within conditions Subjectsindicated the leftright stimulus by pressing either one ofthe two keys of a response box Scene-chronology judg-ment (Temporal task T) Subjects were instructed toremember the order of the events in the clip so as tochoose the image that had happened at an earlier timepoint Scene-layout judgment (Spatial task S) Subjectswere instructed to focus on the spatial layout and choosethe probe image that had the identical spatial arrangementas depicted in the clip Scene-recognition judgment (Objecttask O) Subjects were instructed to focus on the contentof the probe images and to identify the image they hadseen in the clip
It should be noted that the retrieval of complex stimulias those used here can be achieved via many differentstrategies For example to solve the Spatial task partici-pants may attend to and store the location of a specificperson in the scene the facing direction of a car and soon rather than processing the overall spatial details withinthe scene For this reason we emphasize that the ldquolabelsrdquoused here (TemporalSpatialObject) do not imply anyexclusive strategy for solving the tasks but rather aremeant to refer to the specificity of the type of judgmentimplicated during retrieval
Experiment 2
The aim of Exp 2 was to test for task-specific dissocia-tions comparing trials that included only ldquoseenrdquo images(ie images extracted from the video) The retrieval testincluded the three main conditions (Temporal task 48 tri-als Spatial task 48 trials and Object task 48 trials) but
r Neural Correlates of Immediate Episodic Retrieval r
r 2499 r
now each trial comprised one single probe image Scene-chronology (Temporal) Subjects were required to recallthe probe imagersquos temporal location with respect to themidpoint of the clip by choosing between the ldquofirst halfrdquoor the ldquosecond halfrdquo of the clip Subjects were required topress the left button if the probe image happened in thefirst half or the right button if the image happened in thesecond half Scene-layout (Spatial) Subjects wereinstructed to focus on the spatial layout and to judgewhether the probe depicted a scene with a spatial arrange-ment as identical to the clip or it was a mirror image Anidentical spatial configuration required the subject to pressthe ldquocorrectrdquo button whereas a mirror image required apress to the ldquoincorrectrdquo button The mapping of ldquocorrectincorrectrdquo (leftright) layouts was counterbalanced acrosssubjects Scene-recognition (Object) Subjects wereinstructed to focus on the content of the probe image andto indicate whether the image had been shown in the clipIf it had been shown the subject responded with theldquopresentrdquo button otherwise with the ldquoabsentrdquo buttonAgain the leftright mapping of ldquopresentabsentrdquoresponses was counterbalanced between subjects
Since only a single image was now presented atretrieval the Temporal task could not involve any factorof ldquotemporal distancerdquo (cf TshortTlong in Exp 1) Insteadin Exp 2 we found that the behavioral performance wasaffected by the ldquotemporal positionrdquo of the probe imageduring encoding (easier if extracted from the startend ofthe clip harder if extracted from around the middle)Accordingly the temporal trials were now classified andmodeled separately in the fMRI analysis as Teasy and Thard
(see also Supporting Information Table 1)
fMRI Data Acquisition and Preprocessing
The functional images were acquired on a Siemens Alle-gra (Siemens Medical Systems Erlangen Germany) 3 Teslascanner equipped for echo-planar imaging (EPI) A quad-rature volume head coil was used for radio frequencytransmission and reception Head movement was mini-mized by mild restraint and cushioning Thirty-two slicesof functional MR images were acquired using blood oxy-genation level-dependent imaging (3 3 3 mm in-plane25 mm thick 50 distance factor repetition time 5 208 secho time 5 30 ms flip angle 5 70 FOV 5 192 mm acqui-sition order 5 continuous ascending) covering the wholecortex We acquired 1648 and 1488 fMRI volumes foreach subject in Exp 1 and 2 respectively Images wereacquired across 4 fMRI runs in both experiments The totalacquisition periods lasted for 55 and 49 min respectively
Data preprocessing was performed with SPM8 (wwwfilionuclacukspm) implemented on Matlab 74 (The Math-Works Natick MA) After having discarded the first 4volumes in each run images were realigned to correct forhead movements Slice-acquisition delays were correctedusing the middle slice as a reference Images were then
normalized to the MNI EPI template resampled to 3 mmisotropic voxel size and spatially smoothed using an iso-tropic Gaussian Kernel (FWHM 5 8 mm)
fMRI Analyses
Statistical inference was based on a random effectsapproach This comprised two steps first-level analysesestimating contrasts of interest for each subject followedby second-level analyses for statistical inference at thegroup-level [Penny and Holmes 2004] For each of thetwo experiments we used several different modelsdepending on the specific questions that we sought toaddress (see below and Supporting Information Table 1)
All the first-level subject-specific models included oneregressor modeling the presentation of the video clips irre-spective of task plus several regressors that modeled theretrieval phase according to the different conditions ofinterest (see leftmost column in Supporting InformationTable 1) The regressor modeling the ldquoencoding phaserdquo(Movie-regressor) was time-locked to the onset of eachvideo and had an event duration corresponding to theduration of each specific video (ie 772ndash1140 s) All theretrieval-related regressors were time-locked to the onsetof the probe image(s) and with a duration correspondingto the probe presentation at retrieval that is 4 s in Exp 1and 5 s in Exp 2 All regressors were convolved with theSPM8 canonical hemodynamic response function Timeseries were high-pass filtered at 128 s and prewhitened bymeans of autoregressive model AR(1) The sections belowdetail how the trials were assigned to the retrieval regres-sorsconditions in the different analyses
Experiment 1
For Exp 1 we used two different sets of first-level mod-els (see Supporting Information Table 1 leftmost column)The first set of models included the encoding-regressor(Movie) plus 5 retrieval-related conditions temporal-short(Tshort) temporal-long (Tlong) spatial (S) object (O) trialsand a condition modeling all the error trials irrespective oftask (Error) The trials belonging to the Temporal taskwere modeled with two separate regressors accounting forthe factor of temporal distance [Tshort vs Tlong cf Kwoket al 2012] All temporal trials including probes with atemporal distance shorter than 19 s were modeled asTshort while those longer than 19 s modeled as Tlong
The main second-level analysis consisted of a within-subjects Analysis of variance (ANOVA) modeling the foureffects of interest Tshort Tlong S and O conditions Foreach subject and each of the four conditions a contrastimage was computed in the first-level model by averagingthe corresponding parameter estimates across the 4 fMRIruns These subject-specific contrast images were enteredin the second-level ANOVA For the identification of task-specific retrieval effects we compared each task with themean of the two other tasks averaging the two Temporal
r Kwok et al r
r 2500 r
conditions ldquoT(short1long)2gt (S 1 O)2rdquo ldquoSgt (T(short1long)2 1 O)2rdquo and ldquoOgt (T(short1long)2 1 S)2rdquo Sphericity-correction was applied to all group-level ANOVAs toaccount for any nonindependent error term for repeatedmeasures and any difference in error variance across con-ditions [Friston et al 2002] The P-values were correctedfor multiple comparisons using a cluster-level threshold ofP-FWE-corr 5 005 considering the whole brain as the vol-ume of interest The cluster size was estimated using aninitial voxel-level threshold of P-unc 5 0001 To ensurethe specificity of any task effect the main differential con-trast was inclusively masked with 2 additional effectsThese corresponded to the activation for the critical taskvs each of the two other tasks for example the Temporalcontrast ldquoT(short1long)gt (S 1 O)rdquo was inclusively maskedwith ldquoT(short1long)2gt Srdquo and ldquoT(short1long)2gtOrdquo Forthese additional and nonindependent masking contraststhe threshold was set to P-unc 5 005 Together with themain contrasts that compared the three retrieval taskswithin this ANOVA we also tested for the effect ofldquotemporal distancerdquo (TshortgtTlong)
Using the same first-level models we ran a secondwithin-subject ANOVA that now also included the averageresponse times (RT) for each condition per subject as acovariate of no interest (Supporting Information Table 1ldquoControl for task difficultyrdquo) The aim of this was to reas-sess the pattern of task-specific dissociation after havingaccounted for any differences in task difficulty (Fig 1B)This RT-controlled analysis was not independent of themain analysis and the results are reported in the Support-ing Information
Experiment 2
The aim of Exp 2 was to account for the possible influ-ence of ldquounseennewrdquo images that were presented as foilsduring the retrieval phase of the Spatial and Object tasksin Exp 1 (eg incidental encoding during retrieval) InExp 2 a single probe image was presented during theretrieval phase of the trial (Fig 1A Experiment 2) Thisprocedure imposed a new constraint on the Temporaltask which now could not include the factor of ldquotemporaldistancerdquo Instead we taxed temporal retrieval by askingparticipants to judge whether the single probe image wasextracted from the first or second half of the clip Asexpected the participants were faster and more accurate tojudge probes presented either near the beginning or theend of the clip compared with probes extracted fromaround the middle of the clip Thus we now divided thetemporal trials into Teasy and Thard accounting for thisbehavioral effect
For Exp 2 we performed one single set of first-levelmodels and a single second-level group-analysis The first-level models included the encoding-regressor (Movie) and6 retrieval-related regressors Teasy (probe extracted fromthe 1st or 4th quartile) Thard (probe extracted from the2nd or 3rd quartile) Ssame Sflipped Oold and Onew plus
one regressor modeling all the error trials (see SupportingInformation Table 1)
The group level analysis consisted of a within-subjectsANOVA that included the 6 conditions of interest TeasyThard Ssame Sflipped Oold and Onew Within this ANOVAwe first tested for the task-specific effects by comparingeach task vs the mean of the other two tasks In this initialanalysis we averaged the Teasy and Thard conditions ofthe Temporal task as well as seen and unseen images ofthe Spatial and Object tasks for example[T(easy1hard)gt (S(same1flipped) 1 O(old1new))2] The statisticalthresholds used here were the same as Exp 1 statisticalmaps were corrected for multiple comparisons using acluster-level threshold of P-FWE-corr 5 005 (whole-brain)The cluster size was estimated using an initial voxel-levelthreshold of P-unc 5 0001 Again all comparisons wereinclusively masked with the effect of the main task vseach of the two other tasks (see Exp 1 above for details)
Within the same ANOVA a second set of contrasts soughtto replicate the task-specific effects but now using onlyseenold images (Supporting Information Table 1 ldquoSeen-onlyimagesrdquo) hence excluding any possible confound associatedwith the presentation of unseen images during retrieval Weused the following contrasts [T(easy1hard)gt (Ssame 1 Oold)][Ssamegt (T(easy1hard)2 1 Oold)2] and [Ooldgt (T(easy1hard)2 1 Ssame)2] The statistical thresholds and masking proce-dure were identical to Exp 1 and Exp 2 initial analysis
In Exp 2 we also performed control tests to rule out thepossibility that our main findings of task-specific dissocia-tions merely reflected differences in task difficulty (Sup-porting Information Table 1 ldquoTask difficultyrdquo) This wasaddressed by comparing conditions that behaviorallyresulted in difficult vs easy retrieval which included thecorresponding ldquohardrdquo versus ldquoeasyrdquo trials for the Tempo-ral task as well as by trials including ldquoflippedrdquo versusldquosamerdquo images in the Spatial task (Fig 1B) The results ofthese additional analyses are reported in the SupportingInformation
Conjunction analyses Between immediate
memory and LTM
To compare the current findings about task-specificretrieval after short delays with previous data using analo-gous retrieval tasks but a much longer retention delay[24 h Kwok et al 2012] we performed ldquobetween-studiesrdquo analyses Please note that the two paradigms dif-fered not only in terms of the retention delay (which dif-fered by a factor of 43200) but also in the presenceamount of interleaving events between encoding andretrieval In the LTM version participants were exposed to24 h of new episodic experiences following encodingwhich might cause retroactive interference on the alreadyacquired information [Wohldmann et al 2008] whereassuch retroactive interference was absent in the currentstudy on immediate retrieval The video clips were alsoshorter by a factor of about 250 compared to the 42-min
r Neural Correlates of Immediate Episodic Retrieval r
r 2501 r
episode used in the LTM paradigm Because of these differ-ences our analyses focused on commonalities rather thandifferences between LTM and immediate retrieval Theseanalyses considered the current Exp 1 which included thesame retrieval tasks as the LTM experiment Separately foreach of the three tasks we computed the relevant contrasts(eg Tgt (S1O)2) in the two studies and entered these inthree separate two samples t-tests For the identification ofcommon activation across studies we performed null-conjunction analyses (eg [Tgt (S1O)2]LTM [Tgt (S1O)2]ImMem) Statistical threshold were set at P-FWE-corr 5 005 cluster-level corrected at the whole brain level(cluster size estimated and displayed at P-unc 5 0001) con-sistent with all the analyses above For completeness thecontrasts testing for any differences between studies arereported in Supporting Information Table 2
Additional control analyses Retrieval success
Finally we asked to what extent any task-specific effectsidentified in our main analyses depended on the success-ful retrieval of stored information in the clips For this wecarried out additional analyses testing for retrieval per-formance in Exp 1 (ldquoTask 3 Accuracyrdquo in T and S tasks)and for accurate performance on seenold vs unseennewstimuli in Exp 2 (ldquoTask 3 Seen-unseenrdquo in S and O tasks)Please see Supporting Information for details of these anal-yses and the corresponding results
RESULTS
Behavioral Results
Experiment 1
Subjects performed significantly above chance in allthree tasks (all Plt 0001) The percentage of correct accu-racy and response times (RT) were analyzed usingrepeated measures ANOVAs with three levels correspond-ing to the TSO tasks This showed a main effect of con-ditions both for accuracy [F(2 32) 5 8142 Plt 0001] andRT [F(232) 5 3123 Plt 0001] Post hoc t-tests revealedthat subjects performed significantly better and faster inthe Object than Spatial task [Accuracy t(16) 5 1311Plt 0001 RT t(16) 5 652 Plt 0001] and Object than Tem-poral task [Accuracy t(16) 5 979 Plt 0001 RT t(16) 5
671 Plt 0001] Subjects were also more accurate on Tem-poral trials than on Spatial trials [Accuracy t(16) 5 348Plt 0001 but no difference in RT Pgt 01] For the Tempo-ral task we also examined the effect of temporal distanceexpecting that the retrieval would be more difficult whenthe two probes were temporally close (Tshortlt 19 s) com-pared with temporally far (Tlonggt 19 s) during encodingIndeed t-tests showed that subjects responded less accu-rately and slower in short than in long distance trials[Accuracy t(16) 5 391 Plt 0005 RT t(16) 5 315 Plt 001](Fig 1B plot on the left)
Experiment 2
In Exp 2 subjects also performed better than chance inall three tasks (all Plt 0001) Repeated measures ANOVAsshowed main effects of tasks both for accuracy [F(232) 5 919 Plt 0005] and RT [F(232) 5 1766 Plt 0001]Post hoc t-tests revealed that subjects performed withhigher accuracy and shorter RT in the Object comparedwith the Spatial task [Accuracy t(16) 5 406 Plt 0001 RTt(16) 5 532 Plt 0001] and in the Object compared withthe Temporal task [Accuracy t(16) 5 277 Plt 005 RTt(16) 5 383 Plt 0005] The difference in accuracy betweenthe Temporal and Spatial tasks approached significance(Plt 009) while there was no RT difference between Tem-poral and Spatial tasks (Pgt 01) Additional t-tests directlycompared different trial types within each task For theTemporal task subjects performed more accurately andfaster in the ldquoeasyrdquo than ldquohardrdquo trials [Accuracyt(16) 5 657 Plt 0001 RT t(16) 5 527 Plt 0001)] For theSpatial task they were more accurate in recognizingldquosamerdquo than correctly rejecting ldquoflippedrdquo images [Accu-racy t(16) 5 534 Plt 0001 RT Pgt 01] For the Objecttask subjects were faster to respond to unseen Onew
images compared to seen Oold images [RT t(16) 5 294Plt 001] without any difference in accuracy (Pgt 01) (Fig1B plot on the right)
Functional Neuroimaging Task-Specific Retrieval
In both Exp 1 and 2 we assessed brain activity specificto each of the three retrieval tasks (Temporal Spatial andObject) by comparing each task versus the mean of theother two (ldquoTask specificityrdquo contrasts see SupportingInformation Table 1) In Exp 2 we also carried out addi-tional nonindependent control tests using only trials con-taining seenold images (ldquoSeen-only imagesrdquo contrasts)The latter served to ensure that any task-specific dissocia-tion did not arise from the presentation of unseennewimages during the retrieval phase of the Spatial and Objecttasks (ie flipped- and new-image foils)
Scene-chronology judgment (Temporal task)
Exp 1 The comparison of the Temporal task with theother two retrieval conditions revealed significant activa-tion in the right precuneus bilateral caudate and bilateralcuneus (see Table I and Fig 2A clusters in bluemagenta) The signal plots in Figure 2 (panels on the left)show that the precuneus and the right angular gyrus wereactivated more by the Temporal task (average of Tshort andTlong) than by the Spatial and Object tasks Note that theeffect in the right angular gyrus was detected only atuncorrected threshold (P-unclt 0001) and it is reportedhere because it was fully significant in Exp 2 (see below)and as well as in the LTM dataset [Kwok et al 2012]
Together with the main comparison for the Temporaltask in Exp 1 we assessed the effect of temporal distance
r Kwok et al r
r 2502 r
TA
BL
EI
Task
-sp
ecif
icre
trie
val
acti
vati
on
Ex
p
1E
xp
2
Ex
p
2S
een
-on
ly
Clu
ster
Vo
xel
Clu
ster
Vo
xel
Clu
ster
Vo
xel
KP
-co
rr
Zx
yz
kP
-co
rr
Zx
yz
KP
-co
rr
Zx
yz
Sce
ne-
chro
no
log
y(T
emp
ora
l)P
recu
neu
sB
258
lt00
01
71
06
254
47
2666
lt00
01
51
74
272
43
1835
lt00
01
40
06
270
41
Po
st
cin
gu
late
cort
exB
mdashmdash
mdashmdash
564
02
3027
535
62
3631
Cau
dat
eB
148
000
54
302
122
1514
mdashmdash
mdashmdash
mdashmdash
mdashmdash
Cu
neu
sL
820
052
522
215
260
17mdash
mdashmdash
mdashmdash
mdashmdash
mdashC
un
eus
R88
004
25
2118
257
20mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nto
po
lar
cort
exR
mdashmdash
mdashmdash
874
lt0
001
547
2860
970
60
001
575
2860
7S
up
erio
rfr
on
tal
cort
exR
mdashmdash
mdashmdash
685
000
15
3526
2455
889
000
15
6026
2853
An
gu
lar
gy
rus
R43
lt00
01
46
345
269
29
457
00
07
48
444
266
45
509
00
04
45
844
268
43
Sce
ne-
lay
ou
t(S
pat
ial)
Su
peri
or
pari
eta
lg
yru
sL
1142
lt00
01
69
02
21
266
59
2530
lt00
01
66
72
18
268
59
377
00
16
49
92
18
266
59
Do
rsal
occ
ipit
alco
rtex
L5
112
272
759
521
228
280
9mdash
mdashmdash
mdashS
up
ram
arg
inal
gy
rus
L6
212
602
3338
mdashmdash
mdashmdash
mdashmdash
Intr
apar
ieta
lsu
lcu
sL
577
242
239
415
562
362
4445
mdashmdash
mdashmdash
Su
peri
or
pari
eta
lg
yru
sR
1314
lt00
01
67
221
263
59
3026
lt00
01
75
516
266
59
1262
lt00
01
47
426
268
47
Do
rsal
occ
ipit
alco
rtex
R4
1230
293
237
4034
278
294
1030
274
39In
trap
arie
tal
sulc
us
R6
2536
239
4433
40
026
548
402
4045
mdashmdash
mdashmdash
Su
pra
mar
gin
alg
yru
sR
597
602
2435
mdashmdash
mdashmdash
mdashmdash
Su
per
ior
fro
nta
lsu
lcu
sL
213
000
16
452
243
6223
80
078
543
226
22
51mdash
mdashmdash
mdashS
up
erio
rfr
on
tal
sulc
us
R18
20
002
592
302
356
220
009
74
7028
053
mdashmdash
mdashmdash
Infe
rio
rfr
on
tal
gy
rus
L92
003
65
002
516
26mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nta
lg
yru
sR
304
lt00
01
64
054
12
11
536
00
03
60
752
12
25
238
00
78
48
050
12
25
Infe
rio
ro
ccip
ital
gy
rus
L152
00
05
48
12
57
260
210
894
lt00
01
60
32
52
264
211
591
00
02
50
32
50
266
211
Infe
rio
ro
ccip
ital
gy
rus
R186
00
02
67
754
260
213
2277
lt00
01
68
256
258
211
1161
lt00
01
48
750
258
217
Occ
ipit
alp
ole
B21
80
001
484
29
293
2mdash
mdashmdash
mdashmdash
mdashmdash
mdashS
cen
e-re
cog
nit
ion
(Ob
ject
)M
ed
ial
fro
nta
lco
rtex
B478
lt00
01
51
92
351
47
2215
lt00
01
59
62
856
39
314
00
32
46
42
10
56
39
Mid
dle
tem
po
ral
gy
rus
L490
lt00
01
59
42
60
212
225
1661
lt00
01
67
72
62
214
219
162
lt00
01
42
92
60
24
225
Mid
dle
tem
po
ral
gy
rus
R304
lt00
01
61
560
212
216
1017
lt00
01
56
960
26
223
15
lt00
02
29
662
24
223
Orb
ito
-fro
nta
lco
rtex
B95
001
45
040
512
2222
70
049
506
044
219
mdashmdash
mdashmdash
An
gu
lar
gy
rus
L22
70
001
530
251
254
29mdash
mdashmdash
mdashmdash
mdashmdash
mdashA
ng
ula
rg
yru
sR
142
000
64
9857
260
38mdash
mdashmdash
mdashmdash
mdashmdash
mdashR
SC
p
ost
ci
ng
ula
teco
rtex
B47
4lt
000
14
903
242
35mdash
mdashmdash
mdashmdash
mdashmdash
mdash
T-c
on
tras
tsco
mp
ared
each
con
dit
ion
ver
sus
the
mea
no
fth
eo
ther
two
con
dit
ion
s(e
g
T(s
ho
rt1
lon
g)
2gt
(S1
O)
2)
and
was
incl
usi
vel
ym
ask
edw
ith
the
acti
vat
ion
for
the
crit
ical
con
dit
ion
ver
sus
each
of
the
two
oth
erco
nd
itio
ns
(eg
T
(sh
ort
1lo
ng
)2gt
S2
and
T(s
ho
rt1
lon
g)
2gt
O
2)
InE
xp2
we
con
du
cted
two
sep
arat
ese
tso
fco
ntr
asts
atth
eg
rou
p-l
evel
F
irst
t-
con
tras
tsw
ere
con
du
cted
usi
ng
all
six
con
dit
ion
sto
com
par
eea
chco
nd
itio
nv
ersu
sth
em
ean
of
the
oth
ertw
oco
nd
itio
ns
(eg
S (
sam
e1
flip
ped
)gt
[T(e
asy
1h
ard
)1
O(o
ld1
new
)]
2)
see
colu
mn
sin
the
mid
dle
Se
con
d
for
Spat
ial
and
Ob
ject
con
dit
ion
sw
ew
ere
inte
rest
edin
acti
vit
yel
icit
edb
yth
ere
spo
nse
sas
soci
ated
wit
hldquos
een
rdquotr
ials
on
ly(e
g
S sa-
megt
[T(e
asy
1h
ard
)2
1O
old
]2)
se
eri
gh
tmo
stco
lum
ns
and
also
Sup
po
rtin
gIn
form
atio
nT
able
1B
rain
reg
ion
sh
igh
lig
hte
din
bo
ldco
rres
po
nd
toth
eta
sk-s
pec
ific
effe
cts
con
sist
entl
yo
bse
rved
acro
ssal
lex
per
imen
ts
anal
yse
sth
ese
reg
ion
sar
eal
sod
isp
lay
edin
Fig
ure
s2ndash
4P
-val
ues
are
corr
ecte
dfo
rm
ult
iple
com
par
iso
ns
exce
pt
for
the
rig
ht
ang
ula
rg
yru
sin
Exp
1an
dth
em
idd
lete
mp
ora
lg
yri
inE
xp2
Seen
-on
lyth
atar
ere
po
rted
ata
low
eru
nco
rrec
ted
thre
sho
ld(i
nit
alic
s)
Po
st
po
ster
ior
xy
zco
ord
inat
esin
the
stan
dar
dM
NI
spac
eo
fth
eac
tiv
atio
np
eak
sin
the
clu
ster
sk
nu
mb
ero
fv
oxe
lsin
each
clu
ster
R
SC
retr
osp
len
ial
cort
ex
L
R
B
left
ri
gh
th
emis
ph
ere
bil
ater
al
r Neural Correlates of Immediate Episodic Retrieval r
r 2503 r
by comparing Tshort vs Tlong trials In previous studiesusing LTM protocols we found that temporal distance fur-ther modulated activity in the precuneus [Kwok et al2012 2014 see also St Jacques et al 2008] Here this con-trast did not reveal any significant effect This is in con-trast to the significant behavioral cost of retrieving Tshort
vs Tlong temporal distances (see behavioral results andFig 1B plot on the left) that was analogous to our previ-ous LTM protocol Similar behavioral patterns have beenobserved in many other short-term studies [Hacker 1980Konishi et al 2002 Milner et al 1991 Muter 1979] yet toour knowledge there does not seem to exists a clear con-
sensus on the neural correlates subserving the temporaldistance effect separating two temporally very nearbyeventsitems
Exp 2 In Exp 2 we replicated the effect of the temporalretrieval task in the precuneus and found a fully signifi-cant effect in the right angular gyrus (see Fig 2 clusters inredmagenta) In addition in Exp 2 there was a significantactivation of the right superior frontal cortex and the rightinferior frontopolar cortex that were not found in Exp 1(Table I) The involvement of the precuneus and the rightangular gyrus was further confirmed using only trials
Figure 2
Clusters of brain activation and signal plots for the scene-
chronology judgment (Temporal task) Clusters of activation and
signal plots for (A) the precuneus and (B) the right angular
gyrus These regions activated selectively during the temporal
tasks in Exp 1 (in blue) Exp 2 (in red overlap between Exp 1
and 2 shown in magenta) The activation selective for the Tem-
poral task was computed with the contrast T(short1long)
2gt (S 1 O)2 inclusively masked with T(short1long)2gt S2 and
with T(short1long)2gtO2 The involvement of the precuneus and
the right angular gyrus was further confirmed using only ldquoseen
oldrdquo images (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 40 voxels Effect sizes are mean adjusted (sum to
zero) and expressed in arbitrary units (au 6 90 CI) LR left
right PreC precuneus AnG angular gyrus
r Kwok et al r
r 2504 r
including seen images (cf ldquoExp 2 Seen- onlyrdquo Table I)The signal plots on Figure 2 (panels on the right) show theactivity of the precuneus and the right angular gyrus forall the conditions of Exp 2 Both in the precuneus and theright angular gyrus the average activity in the temporaltask [T(easy1hard)2] were larger than the Ssame and theOold conditions Notably Exp 2 included the presentationof a single image and that the participants were asked tojudge the absolute temporal position of the probe ratherthan the relative temporal order Despite this importantchange of how temporal memory was probed the findingof analogous patterns of activation across both experi-ments highlights the link between immediate retrieval oftemporal information and activation of the precuneus andangular gyrus
Scene-layout judgment (Spatial task)
Exp 1 The contrast concerning the retrieval of the spatiallayout revealed activation in the superior parietal cortexbilaterally with the cluster extending to the intraparietalsulcus and the supramarginal gyrus This contrast alsoshowed activation of the superior frontal sulci the inferiorfrontal gyri plus the inferior occipital cortex (see Table Iand Fig 3 clusters displayed in bluemagenta signalsplots on the left side)
Exp 2 The same comparison in Exp 2 replicated theresults of Exp 1 showing activation in the superior parie-tal cortex bilaterally the superior frontal sulci the inferiorand dorsal occipital cortices and the right inferior frontalgyrus (Fig 3 clusters in redmagenta) When we consid-ered only the conditions including the seenold images(ldquoExp 2 Seen- onlyrdquo) we replicated the effects in the supe-rior parietal gyri the inferior occipital gyri and the rightinferior frontal gyrus (see Table I and Fig 3 signal plotson the right) The activation in the intraparietal sulci andthe superior frontal sulci did not reach statistical signifi-cance Inspection of the signal plots indicated that in theseareas there was less activation in the Ssame than Sflipped tri-als (cf bar 3 vs bar 4 signal plots of the superior fontalsulcus Fig 3A plot on the right) Nonetheless the activityin the Ssame condition was larger than those in the T andO conditions (cf bar 3 vs bars 1ndash2 and 5ndash6)
Scene-recognition judgment (Object task)
Exp 1 The Object task was associated with the activationof the medial frontal cortex (MFC) and the orbitofrontalcortex (but the latter did not fully replicate in Exp 2 seeTable I and Fig 4 clusters bluemagenta and signal plotson the left) together with bilateral activation in the middletemporal gyri the retrosplenialposterior cingulate gyrusand a portion of the angular gyrus The same contrast alsorevealed an activation of the hippocampus bilaterally Theactivation of the hippocampus was just below statisticalsignificance (P-FWE-corr 5 0052 Z 5 458 at x y z 5 2126 219 and P-FWE-corr 5 0077 Z 5 458 at x y z 5 224
29 225 considering the whole brain as the volume ofinterest) and it is reported here because the same effectwas previously found in the LTM dataset [Kwok et al2012 see also Discussion Section]
Exp 2 An almost identical pattern of activation wasfound in Exp 2 when we considered all the conditionsO(old1new)gt (T(easy1hard) 1 S(same1flipped))2 Significanteffects were found in the middle temporal gyri and alarge cluster including the medial frontal gyrus extendingventrally into the orbital-frontal cortex (Fig 4 clusters inredmagenta) However when we considered only the tri-als including seenold-images [Ooldgt (T(easy1hard)2 1 Ssame)2] the Object task was found to significantlyactivate only the MFC (Table I rightmost columns) indi-cating that this region was not affected by incidentalencoding
In summary across all three tasks the brain activationwas found to be similar across Exp 1 and Exp 2 Thescene-chronology Temporal task activated the precuneusand the right angular gyrus the scene-layout Spatial taskwas associated with activation of the posteriorsuperiorparietal cortex and the right inferior frontal gyrus whilethe scene-recognition Object task activated the MFC andthe middle temporal gurus (see Table I areas highlightedin bold) The control analysis of Exp 2 that consideredonly trials including seenold images confirmed thesetask-specific effects indicating that the pattern of dissocia-tionmdashand in particular for the Spatial and Object tasksmdashwas not merely driven by the unseennew images duringthe retrieval phase
Functional Neuroimaging Correspondence with
An Analogous LTM Dataset
We directly compared the task-specific dissociationobserved in the current dataset (Exp 1) with our previousLTM study that included the same task-specific retrievaltasks but after a much longer retention delay [ie 24 hbetween encoding and retrieval Kwok et al 2012] Com-mon across the two data sets the conjunction analysesrevealed an activation cluster in the precuneus for theTemporal task (Fig 5A and Table II) and the dorsal fron-toparietal network plus right inferior frontal gyrus andright occipital cortex for the Spatial task (Fig 5B and TableII) These formal tests show that the Temporal and Spatialtask-specific dissociation observed here for immediateretrieval matched well with the patterns observed duringretrieval from LTM By contrast for the Object task over-lapping effects in the MFC and in the hippocampus weredetected only at a more lenient uncorrected threshold ofP-uncorr 5 0001 In the hippocampus the lack of a fullysignificant ldquocommonrdquo effect across datasets was due to thefact that in the current Exp 1 the contrast comparing theObject task versus the two other tasks did not survive cor-rection for multiple comparisons considering the wholebrain as the volume of interest (P-corr 5 0052 and 0077
r Neural Correlates of Immediate Episodic Retrieval r
r 2505 r
Figure 3
Clusters of brain activation and signal plots for the Scene-layout
judgment (Spatial task) Clusters of activation and signal plots for
(A) the superior frontal cortex and the superior parietal cortex
bilaterally and (B) the right inferior occipital cortex and the right
inferior frontal cortex These regions activated for the Spatial task
in Exp 1 (blue) and in Exp 2 (red overlap between Exp 1 and Exp 2
in magenta) The activation selective for the Spatial task was com-
puted with the contrast Sgt [T(short1long)2 1 O]2 inclusively
masked with SgtT(short1long)2 and with SgtO The effects in the
superior parietal gyri the inferior occipital gyri and the right inferior
frontal gyrus were replicated when considering only sameold
images in Exp 2 (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 80 voxels Effect sizes are mean adjusted (sum to zero)
and are expressed in arbitrary units (au 6 90 CI) LR leftright
SFS superior frontal sulcus SPG superior parietal gyrus IFG infe-
rior frontal gyrus IOG inferior occipital gyrus
in the left and right hemispheres respectively cf above)Indeed the formal test assessing differences between thetwo datasets (ie ldquoTask 3 Experimentrdquo interactions seeSupporting Information Table 2) did not reveal any signifi-cant difference between LTM and immediate retrieval inthe hippocampus consistent with this region beinginvolved in the scene-recognition judgment irrespective ofthe retention delay (see also Discussion Section)
DISCUSSION
We have identified distinct sets of brain areas thatshowed task-specific activation during immediate retrievalfollowing the encoding of dynamic cinematic material Theprecuneus and the right angular gyrus were activatedwhile retrieving the temporal order of events (scene-chro-nology judgment) the superior parietal cortex wasrecruited during spatial related recall (scene-layout
Figure 4
Clusters of brain activation and signal plots for the Scene-
recognition judgment (Object task) Clusters of activation and
signal plots for (A) the MFC and (B) the MTG These regions
activated for the Object task in Exp 1 (blue) and in Exp 2 (red
overlap in magenta) The activation selective for the Object task
was computed with the contrast Ogt [T(short1long)2 1 S]2
inclusively masked with OgtT(short1long)2 and with Ogt S The
signal plots refer to the left hemisphere but analogous patterns
of activation were found in the right hemisphere (Table I) In
Exp 2 the task-specific effect of the Object task in the MFC
was replicated when considering only the sameold images
(Table I rightmost columns) Inspection of the signal plot for the
left MTG revealed less activity in the Oold than Onew trials (plots
on the right bars 5 and 6) although the Oold trials showed
numerically larger parameter estimates than the T and S condi-
tions (see p-uncorr reported in italics in Table I rightmost col-
umns) Activation clusters were estimated and displayed at P-
unc 5 0001 thresholded at a minimum of 80 voxels Effect sizes
are mean adjusted (sum to zero) and expressed in arbitrary
units (au 6 90 CI) LR leftright MTG middle temporal
gyrus MFC medial frontal cortex
r Neural Correlates of Immediate Episodic Retrieval r
r 2507 r
judgment) whereas activity in the MFC and the middletemporal gyrus (MTG) reflected processes associated withscene-recognition judgments These task-specific effectsoccurred independently of whether subjects were asked tojudge seenold or unseennew stimuli (cf ldquoSeen-onlyimagesrdquo contrasts in Exp 2) and independently ofretrieval success and general task difficulty (see Support-ing Information) With respect to the hypothesized corre-spondence between immediate retrieval and retrieval fromLTM we found that these task-specific dissociationsare independent of the informationrsquos mnemonic history(24 h vs 2 s retention ie differing by 43200 times inmagnitude)
One important qualifier for the following discussion isthat memory retrieval in all our tasks depended minimallyon memory buffers compared to other simpler itemsdis-plays that are typically used in ldquostandardrdquo short-termworking memory tasks [Shallice and Cooper 2011 Shiffrin1973] In each trial the number of items presented during
encoding was clearly well beyond the 4 6 1 items stipulatedby classical STM models [Cowan 2001] While memories ofsome materials (eg words spatial positions simple visualdisplays) are supported by limited-capacity buffers [Badde-ley and Hitch 1974] memory for other categories [egunknown faces and scenes see Shiffrin 1973] will tap intoLTM when the task requires the encoding of two or moreitems [Shallice and Cooper 2011] For this reason werestrict our discussion on the tripartite task-specific dissoci-ations with reference to highly complex naturalisticmaterial
Precuneus and Inferior Parietal Cortex Support
Temporal Order Recollection
The scene-chronology judgment task activated the pre-cuneus in both Exp 1 and 2 despite the considerablechanges in the trial structure for probing the retrieval oftemporal information (ie relative temporal order betweentwo probes in Exp 1 vs absolute temporal position of asingle probe in Exp 2) The chronology-judgment taskrequired retrieving the event sequences of many complex
Figure 5
Task-specific retrieval activation common in studies after imme-
diate delays and a longer delay Activation for the LTM study
[24 h Kwok et al 2012] is depicted in green and activation
for the current immediate retrieval study (data from current
Exp 1) depicted in red Activation clusters for both studies were
estimated and displayed at P-unc 5 0001
TABLE II Common memory retrieval activation
ImMem LTM common effects
Cluster Voxel
k P-corr Z x y z
Scene-chronology (Temporal)Precuneus B 112 0011 472 3 257 47Angular gyrus R 98 lt 0001 400 54 254 17
Scene-layout (Spatial)Superior parietal gyrus L 206 0001 595 221 269 56Supramarginal gyrus L 129 0008 346 260 236 44Intraparietal sulcus L 487 236 245 41Superior parietal gyrus R 651 lt 0001 520 24 263 59Dorsal occipital cortex R 372 30 275 29Intraparietal sulcus R 520 42 239 47Supramarginal gyrus R 482 57 233 53Superior frontal sulcus L 100 0022 482 224 3 56Superior frontal sulcus R 145 0004 481 30 0 53Inferior frontal gyrus R 95 0027 475 54 9 26Inferior occipital gyrus R 98 0024 515 54 257 210
Task-specific activation after immediate delays (current Exp 1ImMem) and a previous study that used a long delay betweenencoding and retrieval [24 h LTM Kwok et al 2012] Statisticalthreshold were set at P-FWE-corr 5 005 (cluster-level) consider-ing the whole brain as the volume of interest Note that theldquobetween-studiesrdquo overlap activation in the right angular gyruswas found at a lower uncorrected threshold (in italics) but impor-tantly within this area no differences or interactions betweenImMem and LTM studies were found (see differential effectsbetween the two studies in Supporting Information Table 2) LRB leftright hemispherebilateral
r Kwok et al r
r 2508 r
stimuli (scenes) that entail high-level conceptualhierarchi-cal relationships between them [Swallow et al 2011 Zackset al 2001] The precuneus was previously found to acti-vate in our LTM protocol that used the same temporalorder judgment [Kwok et al 2012 and see also results ofthe conjunction analyses] The current finding of precunealactivation using very different (much shorter) delays fitswell with the view that these processes are not strictlyassociated with retrieval following long retention delayssuggesting that this medial parietal region is implicatedduring temporal order retrieval irrespective of delay (Fig5A and Table II) Specifically we link the effect of chrono-logical retrieval in the precuneus with processes involvingan active reconstruction of past events [see also St Jacqueset al 2008] and possibly memory-related imagery[Fletcher et al 1995 Ishai et al 2000] These processeswould characterize temporal order judgments using com-plex naturalistic material but not order-judgment withsimpler stimulus material which previous studies haveassociated with other brain regions [eg middleprefron-tal areas Suzuki et al 2002]
Together with the precuneus temporal-related judgmentalso gave rise to activation of the right angular gyrus(AG) One possible interpretation relates the inferior parie-tal cortex to the engagement of attentional processes[Majerus et al 2007 2010] The scene-chronology judg-ment of Exp 2 provided us with a further qualifier to thiskind of temporal-related attention Although the Temporaltask in Exp 2 did not require any explicit temporal orderjudgment (only classifying a scene into either the first orsecond half of the movie) an accurate decision still neces-sitated an evaluation of the target scene with many othersegments of the movie This operation is fundamentallydifferent from the evaluation implicated in the scene-layout judgment which required decisions on comparingbetween a single scene and its own correspondentencoded snapshot The latter involves visuospatial analysisof the configuration within scenes The AG may subservedifferent types of attention among which one of them islinked with memory search operations [see Majerus et al2010 2006 also showing coactivation of these two regionsduring order retrieval of simple stimuli after short delays]
Superior Parietal Cortex Evaluates Spatial
Details Against Memory Representation
The second pattern consistently found across experi-ments (Exp 1 and Exp 2 see Fig 3) and across studieswith different retention delays (conjunction betweenimmediate retrieval and LTM studies see Fig 5B andTable II) was the activation of the superior parietal lobule(SPL) during the scene-layout judgment Previous studiesshowed sustained activation in the parietal cortex duringworking memory delays [Courtney et al 1998] and thatactivity there correlates with memory load [Todd andMaroi 2004] Here the amount of information contained
in the clips was equated in the three tasks especially inExp 2 where we counterbalanced the assignment of mov-ies across tasks The subjects also could not anticipatewhat specific type of information would be tested atretrieval and needed to keep rehearsing different aspectsof the clips Because of this we deem unlikely that theactivation of SPL that here was specific for the scene-layout tasks reflected general rehearsal processes [Magenet al 2009] andor updating operations [Bledowski et al2009 Leung et al 2007]
Instead we suggest a role for the SPL in mediating aninterface between the internal memory representation andexternal information It has been proposed that the supe-rior parietal cortex is engaged in allocating top-downattention to orient to aspects of the available visual inputand on information retrieved from memory [Ishai et al2002 Lepsien and Nobre 2007 Summerfield et al 2006]In line with this we found SPL activation when the sub-jects had to make decisions by comparing between theirmemory of the encoded clips and the scene stimuli pre-sented at retrieval This procedure might act on somevisuospatial storage which is held by the lateral occipitalcortex and mediated by the intraparietal sulcus (Fig 3B)The immediate memory task here was analogous to theSpatial task of our previous LTM study [Kwok et al2012] which also required matchingcomparing the avail-able external visual input and internal information storedin memory [see also Nobre et al 2004] This interfaceinvolves a range of top-down processes such as monitor-ing and verification [Cabeza et al 2008 Ciaramelli et al2008] which are independent of the probe imagersquos mne-monic status as shown by the equated effect sizes forldquosamerdquo and ldquoflippedrdquo images conditions (see signals plotsin Fig 3 and also ldquoRetrieval successrdquo analyses of Exp 2 inSupporting Information)
Recognition Processes in the Medial Frontal
Cortex and the Temporal Cortex
The third retrieval task involved scene-recognition judg-ments and was associated with the activation of the MFCand the MTG Moreover in Exp 1 we found a statisticaltrend in the hippocampus bilaterally an area that we pre-viously associated with scene-recognition judgment fromLTM (see next section about ldquoPossible correspondencesbetween retrieval from short- and long-term memoryrdquo)
The MFC was engaged both Exp 1 and 2 but was notactivated in the other two tasks thus a broad prefrontalcortexworking memory correspondence [Tsuchida andFellows 2009] seems insufficient to explain this task-specific effect The scene-recognition task in Exp 2 taxedboth recognition of ldquooldrdquo scenes and correct rejection ofldquonewrdquo scenes suggesting that the MFC here did not dis-tinguish between the correct rejection of new information(unseennew trials) and the recognition of old imagesOne candidate cognitive process that is common to both
r Neural Correlates of Immediate Episodic Retrieval r
r 2509 r
types of processes is feeling-of-knowing (FOK) FOKdepend on the cue specification content retrieval andappraisal of the retrieved memory fragments but are inde-pendent of the ultimate determination of content appropri-ateness [Schnyer et al 2005] Using imaging data duringsentence completion recognition Schnyer et al [2005]modeled a FOK-related directional path The path impli-cated the temporal cortex and the hippocampus andengaged the ventro-MFC at the end which is consistentwith our findings for the Object task implicating the MFCthe middle temporal cortex and at a lower statisticalthreshold the hippocampus
By contrast the activity in the MTG was larger forunseennew images than for seenold memory probes(Fig 4B) This suggests that new encoding may have con-tributed to the activation of the MTG during the retrievalphase of the Object task in Exp 1 The scene-recognitiontask required a lower precision in terms of retrievingdetails about the encoded material and could be solved bysimple recognition [Wixted and Squire 2010] Thus alter-natively larger activation for ldquonew vs oldrdquo stimuli mayrelate to some reduced activation associated with familiar-ity processes on old trials [see Henson et al 1999 whoreported reduced familiarity related activation in middletemporal gyri bilaterally and Henson and Rugg 2003 forreview] However we should point out that in Exp 2 theunseen images were extracted from edited-out parts of theoriginal clips and therefore contained the same objectcharacter depicted in the target images Therefore wedeem more likely that the relational association betweenobjects rather than their presenceabsence in the sceneshelped subjects to distinguish targets from foils We thussuggest that the activation of the MTG reflects processesutilizing the relational details among objects embedded inthe probe images for reaching the recognition decision[Diana et al 2007 Finke et al 2008 Hannula et al 2006Olson et al 2006] Future studies should consider theinclusion of some correlative behavioral measures to indexfamiliarity processes to elucidate further this issue More-over while the Temporal and Spatial tasks required mak-ing relative comparisons about aspects of the encodedvideo-clip the scene-recognition judgment merely requiredjudging whether the image presented at retrieval was pres-ent (or not) within the clip This may entail different proc-essing dynamics in the Object task compared with theother two tasks (cf also differences in retrieval times) Thelatter could be addressed using more subtle experimentalmanipulations for the scene-recognition judgment forexample by using digitally-modified foil images where asingle object in the scene has been modifiedremoved
Possible Correspondences Between Retrieval
from Short- and Long-Term Memory
The overlap between our current study and the previousLTM study using analogous tasks and materials lends
weight to the notion of scale-invariance in memory mecha-nisms [Brown et al 2007] as have been previously dem-onstrated in free recall tasks where the rate of item recallwas unvarying across recall span (day week year) andeven across from the past to the prospective future [May-lor et al 2001] Those previous behavioral findings implythat scale-invariance is a form of self-similarity in that theholistic pattern can exist at multiple levels of magnification[and even across different memory systems eg Maylor2002] Here our data affirmed the importance of the poste-rior parietal cortex in both immediate and long-termretrieval [Berryhill et al 2011 2007] and further character-ized its involvement in dealing with the spatial relationsof objects within scenes [Bricolo et al 2000 Buiatti et al2011] Our data also demonstrated such short-long-termcorrespondence exists in the precuneusangular gyrus fortemporal order retrieval (cf scene-chronology judgmenttask) In contrast the bilateral hippocampus and MFC acti-vations did not reach full statistical significance in theImMem LTM conjunction analysis At first sight thismay indicate that the scene-recognition task tapped intodistinct recognition-based mechanisms as a function ofretention delay However we deem this unlikely becausethe direct comparison between the two datasets did notshow any significant effect in these regions (see Support-ing Information Table 2) Indeed a more targeted test ask-ing the question of whether the hippocampus clusterspreviously found in LTM activated also in the currentimmediate retrieval experiments revealed significanteffects in both immediate retrieval experiments here (Exp1 x y z 5 224 29 225 P-FWE-corrlt 0001 Z 5 458 x yz 5 21 29 222 P-FWE-corr 5 0001 Z 5 451 and in Exp2 x y z 5 224 214 221 P-FWE-corr 5 0004 Z 5 396using the LTM clusters for small volume correction) Theseadditional tests point to a more general issue of sensitivitythat future studies may address using different methodo-logical approaches such as multivariate methods that canbe more sensitive than the standard univariate analysesused here Moreover the former would enable capturingtask-specific effects that are expressed as patterns of dis-tributed activation rather than local changes of activity asreported here [eg Jimura and Poldrack 2012]
Task Difficulty and Retrieval Success
We used several control analyses to show that the task-specific patterns of activation and the correspondencebetween the current data and our previous LTM study donot merely reflect some effect related to general task diffi-culty (see Supporting Information) First areas involved inoverall task difficulty should show progressively greateractivation with progressively more difficult tasks In Exp 1this would entail SgtTgtO However the patterns in theprecuneus and the angular gyrus (with TgtOgt S Fig 2A)and the MFC (Ogt SgtT Fig 4A) did not fit with simplepredictions based on overall task difficulty We also
r Kwok et al r
r 2510 r
assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r

presentation of the unseen foil images Moreover the sub-stantial changes of the task structure across experiments 1and 2 enabled us to confirm any task-specific dissociationunder different sets of task-constraints
On the assumption that memory for complex naturalis-tic events depends negligibly on capacity-limited memorybuffers we hypothesized task-specific functional dissocia-tions during retrieval after short delays were analogous tothose observed during retrieval after long delays This wastested by identifying any task-specific brain activation dur-ing retrieval after short delays (ldquotask-specificrdquo analysesExp 1 and 2) and by comparing these effects with datafrom the previous LTM experiment [ldquoconjunction analy-sesrdquo between the current Exp 1 and LTM data from Kwoket al 2012] Findings of task-specific dissociations equiva-lent to those previously reported for LTM would supportthe view that memory for naturalistic material can accom-modate a vast range of retention delays [see also Mayloret al 2001 for scale-invariance in free recall memory) Inaddition we controlled that the task-specific effects atretrieval were not merely due to the new encoding ofunencountered material (ldquoseen-onlyrdquo analyses in Exp 2)and tested whether the task-specificity was determined bythe successful retrieval outcome or rather reflected theattempt to recover relevant mnemonic details [Rugg andWilding 2000] Confirmation of our hypotheses abouttask-specific functional dissociations during retrieval aftershort delays would indicate that the processes governingmemory for naturalistic events are similarly guided bytask-specific constraints across various retention delaysand provide evidence for the notion of scale-invariance inmemory functions [Brown et al 2007 Howard andKahana 2002]
MATERIALS AND METHODS
Overview
The trial structure of Exp 1 and Exp 2 were analogousthe subject watched a short video clip and then performedone of three retrieval tasks about some details of the clipThe retrieval tasks aimed at tapping into different proc-esses associated with the judgment of scene-chronology(Temporal task) scene-layout (Spatial task) or scene-recognition (Object task) The retrieval task was cued atthe onset of the probe images This cueing procedureensured that the subjects encoded the movie clips withoutanticipating what specific stimulus detail would be probedat retrieval (see Fig 1A)
In Exp 1 the retrieval phase comprised the presentationof two images side-by-side For the Temporal task bothscenes were extracted from the clip and the subject wasasked to choose the scene that had happened at an earliertime point in the clip (target) For the Spatial and Objecttasks the judgment at retrieval was between one scenethat had been presented (target) and another one that had
not been presented (foil) We investigated task-specific dis-sociations by directly comparing each of the tasks with theaverage of the other two tasks Further we performed con-junction analyses between the current dataset of Exp 1 anda previous LTM experiment that made use of the sameretrieval procedure but with a longer movie (42-min) anda retention delay of 24 h [Kwok et al 2012] This enabledus to formally test whether there was any functional over-lap between the patterns of task-specific dissociation fol-lowing short and long delays (15ndash25 s vs 24 h)
Exp 1 was designed to match the previous LTM study[Kwok et al 2012] thus including two images (target andfoil) in each retrieval trial However with this procedurethe retrieval-display of scene-recognition Object taskincluded an unseen image and the display of the scene-layout Spatial task included an unseen image-configuration The presentation of these unseen foils mayhave contaminated any task-specific effects associated withthe Spatial and Object tasks (eg incidental encoding ofnew images or new image-configurations) This wasaddressed in Exp 2 that entailed the presentation of onesingle image at retrieval and required the subject toanswer with a yesno response In the Spatial and theObject tasks the probe was either a seenold imageextracted from the video or an unseen image The Tempo-ral task always comprised the presentation of a probeimage extracted from the clip but the task now involvedjudging whether the image belonged to the first or secondhalf of the clip We again assessed task-specific dissocia-tions during immediate retrieval but now by comparingthe three tasks using only trials including seenold probeimages to avoid any possible confound related to theunseen images
Subjects
Seventeen participants took part in Exp 1 (mean age258 21ndash33 years 8 females) and another 17 different par-ticipants took part in Exp 2 (mean age 254 20ndash35 years 7females) All participants gave written informed consenthad normal or corrected-to-normal vision without knownhistory of psychiatricneurological problems and werefree from any MRI contraindication The study wasapproved by the Fondazione Santa Lucia (Scientific Insti-tute for Research Hospitalization and Health Care) Inde-pendent Ethnics Committee in accordance with theDeclaration of Helsinki
Stimuli
Movie clips
We purchased a collection of non-Italian TV commercialclips from an Advertising Archive (httpwwwcoloribuscom) and reproduced 160 short clips using a video editingsoftware (Final Cut Pro Apple Inc) Each clip was pur-posely edited to depict a coherent storyline with multiple
r Neural Correlates of Immediate Episodic Retrieval r
r 2497 r
switches of scenes The mean length was 959 s with arange of 772ndash1140 s We preserved the background soundand used only segments without dialoguesconversationsThe whole set of 160 clips was used in Exp 1 and each clipwas assigned to one of the three retrieval tasks 96 Tempo-ral 32 Spatial and 32 Object clips For Exp 2 we selected asubset of 144 clips and each clip was used for all the threetasks counterbalanced across subjects
Clip analyses
We performed a frame-by-frame analysis to mark thechanges of scene for each clip The scene changes includedboth between-scenes cuts (no scene continuity ie edit cutfrom one scene to another) and within-scenes cuts [scenecontinuity but with a change of camera perspective Smithand Henderson 2008] This frame-by-frame analysis allowed
Figure 1
Task schema and behavioral performance for Experiments 1 and
2 (A) Trial timeline probe images and cues at retrieval Each
trial carried an identical structure consisting of presentation of a
short movie clip followed by a cued retrieval test The retrieval
test comprised either a pair of probe images (Exp 1) or one sin-
gle probe (either target or foil Exp 2) (B) Accuracy presented
in bars ( correct unit on left ordinate axes) and response
times in lines (ms right ordinate axes) for Experiment 1 (left
panel) and Experiment 2 (right panel) Error bars are standard
error of the means
r Kwok et al r
r 2498 r
us to identify boundary frames that divided segmentswithin each clip The number of segments across all clipsranged between 4 and 10 The first and last segments werealways excluded for the retrieval tests to mitigate end-of-listdistinctiveness effects [Cabeza et al 1997 Konishi et al2002] Specifically for Exp 2 we divided each clip into 20segments with equal duration The memory probe imageswere extracted from 12 of these segments 3ndash5 (1st quartile)7ndash9 (2nd quartile) 12ndash14 (3rd quartile) and 16ndash18 (4th quar-tile) (cf Temporal task)
Probe images extraction
Exp 1 After excluding the first and last segments for theTemporal trials we pseudorandomly selected two imagesfrom two different segments from each temporal clip Thisrepresented a range of temporal distances between thetwo probe images across all the Temporal trials The tem-poral distances varied between 060 and 496 s The Spatialtrials were generated by pairing a target image with itsown mirror image (by laterally flipping the target image)The Object trials were generated by pairing a target imagewith an unseen image that also contained the same objectcharacter from an edited-out part of the original clip
Exp 2 Irrespective of task we selected one probe imagefrom each clip which was used as the target image Thisimage was extracted with an equal likelihood from one ofthe 4 quartiles (see Clip analyses above) thus with half ofthem coming from the first half and the other half fromthe second half of the clip For the Spatial task the foilimages were mirror images of the targets whereas for theObject task new images were extracted from an edited-outpart of the original clip as the foil images
Trial structure
The trial structure was identical in Exp 1 and 2 eachtrial began with a green fixation cross for 05 s followedby the presentation of a clip lasting for 772ndash114 s then ablank screen for a period of 15ndash25 s This was followedby a retrieval test (Fig 1A) The two experiments differedin how memory was probed during retrieval The retrievaltest of Exp 1 contained a pair of probe images and a writ-ten cue above the images both of which were presentedon the screen for 4 s whereas that of Exp 2 contained onlyone probe image and a written cue both presented for 5 sIn both experiments the written cue specified what taskthe subject had to perform on that trial Trials were sepa-rated by a white fixation cross with jittered ITIs sampledfrom a truncated logarithmic distribution (trial durationsranged between 207 and 228 s) We presented the stimuliusing Cogent Toolbox (httpwwwvislabuclacukcogentphp) implemented in MATLAB 74 (The Math-Works Natick MA) During fMRI scanning all stimuliwere back-projected on a screen at the back of the MRbore and viewed through a mirror system (20 3 15 visual
angle) Participants were given detailed written instruc-tions and were explained of the task requirements prior tothe commencement of the experiments
Retrieval Tasks
Experiment 1
The retrieval test included three main experimental con-ditions Temporal task (96 trials) Spatial task (32 trials)and Object task (32 trials) The reason for the larger num-ber of Temporal trials was that this task comprised twodifferent trial types (Tshort and Tlong) that were classifiedmdashand modeled separately in the main fMRI analysismdashaccording to the temporal distance between the two probeimages (see also Supporting Information Table 1) This dis-tinction was motivated by previous findings showing thatthe temporal separation of the probes at encoding canaffect retrieval performance and brain activity during tem-poral order judgments with naturalistic material [Kwoket al 2012 St Jacques et al 2008]
For all three tasks each retrieval trial comprised a pairof probe images arranged with a leftright side-by-sideconfiguration one of the images was the target the otherwas the foil The left-right position of the targetfoilimages was counterbalanced within conditions Subjectsindicated the leftright stimulus by pressing either one ofthe two keys of a response box Scene-chronology judg-ment (Temporal task T) Subjects were instructed toremember the order of the events in the clip so as tochoose the image that had happened at an earlier timepoint Scene-layout judgment (Spatial task S) Subjectswere instructed to focus on the spatial layout and choosethe probe image that had the identical spatial arrangementas depicted in the clip Scene-recognition judgment (Objecttask O) Subjects were instructed to focus on the contentof the probe images and to identify the image they hadseen in the clip
It should be noted that the retrieval of complex stimulias those used here can be achieved via many differentstrategies For example to solve the Spatial task partici-pants may attend to and store the location of a specificperson in the scene the facing direction of a car and soon rather than processing the overall spatial details withinthe scene For this reason we emphasize that the ldquolabelsrdquoused here (TemporalSpatialObject) do not imply anyexclusive strategy for solving the tasks but rather aremeant to refer to the specificity of the type of judgmentimplicated during retrieval
Experiment 2
The aim of Exp 2 was to test for task-specific dissocia-tions comparing trials that included only ldquoseenrdquo images(ie images extracted from the video) The retrieval testincluded the three main conditions (Temporal task 48 tri-als Spatial task 48 trials and Object task 48 trials) but
r Neural Correlates of Immediate Episodic Retrieval r
r 2499 r
now each trial comprised one single probe image Scene-chronology (Temporal) Subjects were required to recallthe probe imagersquos temporal location with respect to themidpoint of the clip by choosing between the ldquofirst halfrdquoor the ldquosecond halfrdquo of the clip Subjects were required topress the left button if the probe image happened in thefirst half or the right button if the image happened in thesecond half Scene-layout (Spatial) Subjects wereinstructed to focus on the spatial layout and to judgewhether the probe depicted a scene with a spatial arrange-ment as identical to the clip or it was a mirror image Anidentical spatial configuration required the subject to pressthe ldquocorrectrdquo button whereas a mirror image required apress to the ldquoincorrectrdquo button The mapping of ldquocorrectincorrectrdquo (leftright) layouts was counterbalanced acrosssubjects Scene-recognition (Object) Subjects wereinstructed to focus on the content of the probe image andto indicate whether the image had been shown in the clipIf it had been shown the subject responded with theldquopresentrdquo button otherwise with the ldquoabsentrdquo buttonAgain the leftright mapping of ldquopresentabsentrdquoresponses was counterbalanced between subjects
Since only a single image was now presented atretrieval the Temporal task could not involve any factorof ldquotemporal distancerdquo (cf TshortTlong in Exp 1) Insteadin Exp 2 we found that the behavioral performance wasaffected by the ldquotemporal positionrdquo of the probe imageduring encoding (easier if extracted from the startend ofthe clip harder if extracted from around the middle)Accordingly the temporal trials were now classified andmodeled separately in the fMRI analysis as Teasy and Thard
(see also Supporting Information Table 1)
fMRI Data Acquisition and Preprocessing
The functional images were acquired on a Siemens Alle-gra (Siemens Medical Systems Erlangen Germany) 3 Teslascanner equipped for echo-planar imaging (EPI) A quad-rature volume head coil was used for radio frequencytransmission and reception Head movement was mini-mized by mild restraint and cushioning Thirty-two slicesof functional MR images were acquired using blood oxy-genation level-dependent imaging (3 3 3 mm in-plane25 mm thick 50 distance factor repetition time 5 208 secho time 5 30 ms flip angle 5 70 FOV 5 192 mm acqui-sition order 5 continuous ascending) covering the wholecortex We acquired 1648 and 1488 fMRI volumes foreach subject in Exp 1 and 2 respectively Images wereacquired across 4 fMRI runs in both experiments The totalacquisition periods lasted for 55 and 49 min respectively
Data preprocessing was performed with SPM8 (wwwfilionuclacukspm) implemented on Matlab 74 (The Math-Works Natick MA) After having discarded the first 4volumes in each run images were realigned to correct forhead movements Slice-acquisition delays were correctedusing the middle slice as a reference Images were then
normalized to the MNI EPI template resampled to 3 mmisotropic voxel size and spatially smoothed using an iso-tropic Gaussian Kernel (FWHM 5 8 mm)
fMRI Analyses
Statistical inference was based on a random effectsapproach This comprised two steps first-level analysesestimating contrasts of interest for each subject followedby second-level analyses for statistical inference at thegroup-level [Penny and Holmes 2004] For each of thetwo experiments we used several different modelsdepending on the specific questions that we sought toaddress (see below and Supporting Information Table 1)
All the first-level subject-specific models included oneregressor modeling the presentation of the video clips irre-spective of task plus several regressors that modeled theretrieval phase according to the different conditions ofinterest (see leftmost column in Supporting InformationTable 1) The regressor modeling the ldquoencoding phaserdquo(Movie-regressor) was time-locked to the onset of eachvideo and had an event duration corresponding to theduration of each specific video (ie 772ndash1140 s) All theretrieval-related regressors were time-locked to the onsetof the probe image(s) and with a duration correspondingto the probe presentation at retrieval that is 4 s in Exp 1and 5 s in Exp 2 All regressors were convolved with theSPM8 canonical hemodynamic response function Timeseries were high-pass filtered at 128 s and prewhitened bymeans of autoregressive model AR(1) The sections belowdetail how the trials were assigned to the retrieval regres-sorsconditions in the different analyses
Experiment 1
For Exp 1 we used two different sets of first-level mod-els (see Supporting Information Table 1 leftmost column)The first set of models included the encoding-regressor(Movie) plus 5 retrieval-related conditions temporal-short(Tshort) temporal-long (Tlong) spatial (S) object (O) trialsand a condition modeling all the error trials irrespective oftask (Error) The trials belonging to the Temporal taskwere modeled with two separate regressors accounting forthe factor of temporal distance [Tshort vs Tlong cf Kwoket al 2012] All temporal trials including probes with atemporal distance shorter than 19 s were modeled asTshort while those longer than 19 s modeled as Tlong
The main second-level analysis consisted of a within-subjects Analysis of variance (ANOVA) modeling the foureffects of interest Tshort Tlong S and O conditions Foreach subject and each of the four conditions a contrastimage was computed in the first-level model by averagingthe corresponding parameter estimates across the 4 fMRIruns These subject-specific contrast images were enteredin the second-level ANOVA For the identification of task-specific retrieval effects we compared each task with themean of the two other tasks averaging the two Temporal
r Kwok et al r
r 2500 r
conditions ldquoT(short1long)2gt (S 1 O)2rdquo ldquoSgt (T(short1long)2 1 O)2rdquo and ldquoOgt (T(short1long)2 1 S)2rdquo Sphericity-correction was applied to all group-level ANOVAs toaccount for any nonindependent error term for repeatedmeasures and any difference in error variance across con-ditions [Friston et al 2002] The P-values were correctedfor multiple comparisons using a cluster-level threshold ofP-FWE-corr 5 005 considering the whole brain as the vol-ume of interest The cluster size was estimated using aninitial voxel-level threshold of P-unc 5 0001 To ensurethe specificity of any task effect the main differential con-trast was inclusively masked with 2 additional effectsThese corresponded to the activation for the critical taskvs each of the two other tasks for example the Temporalcontrast ldquoT(short1long)gt (S 1 O)rdquo was inclusively maskedwith ldquoT(short1long)2gt Srdquo and ldquoT(short1long)2gtOrdquo Forthese additional and nonindependent masking contraststhe threshold was set to P-unc 5 005 Together with themain contrasts that compared the three retrieval taskswithin this ANOVA we also tested for the effect ofldquotemporal distancerdquo (TshortgtTlong)
Using the same first-level models we ran a secondwithin-subject ANOVA that now also included the averageresponse times (RT) for each condition per subject as acovariate of no interest (Supporting Information Table 1ldquoControl for task difficultyrdquo) The aim of this was to reas-sess the pattern of task-specific dissociation after havingaccounted for any differences in task difficulty (Fig 1B)This RT-controlled analysis was not independent of themain analysis and the results are reported in the Support-ing Information
Experiment 2
The aim of Exp 2 was to account for the possible influ-ence of ldquounseennewrdquo images that were presented as foilsduring the retrieval phase of the Spatial and Object tasksin Exp 1 (eg incidental encoding during retrieval) InExp 2 a single probe image was presented during theretrieval phase of the trial (Fig 1A Experiment 2) Thisprocedure imposed a new constraint on the Temporaltask which now could not include the factor of ldquotemporaldistancerdquo Instead we taxed temporal retrieval by askingparticipants to judge whether the single probe image wasextracted from the first or second half of the clip Asexpected the participants were faster and more accurate tojudge probes presented either near the beginning or theend of the clip compared with probes extracted fromaround the middle of the clip Thus we now divided thetemporal trials into Teasy and Thard accounting for thisbehavioral effect
For Exp 2 we performed one single set of first-levelmodels and a single second-level group-analysis The first-level models included the encoding-regressor (Movie) and6 retrieval-related regressors Teasy (probe extracted fromthe 1st or 4th quartile) Thard (probe extracted from the2nd or 3rd quartile) Ssame Sflipped Oold and Onew plus
one regressor modeling all the error trials (see SupportingInformation Table 1)
The group level analysis consisted of a within-subjectsANOVA that included the 6 conditions of interest TeasyThard Ssame Sflipped Oold and Onew Within this ANOVAwe first tested for the task-specific effects by comparingeach task vs the mean of the other two tasks In this initialanalysis we averaged the Teasy and Thard conditions ofthe Temporal task as well as seen and unseen images ofthe Spatial and Object tasks for example[T(easy1hard)gt (S(same1flipped) 1 O(old1new))2] The statisticalthresholds used here were the same as Exp 1 statisticalmaps were corrected for multiple comparisons using acluster-level threshold of P-FWE-corr 5 005 (whole-brain)The cluster size was estimated using an initial voxel-levelthreshold of P-unc 5 0001 Again all comparisons wereinclusively masked with the effect of the main task vseach of the two other tasks (see Exp 1 above for details)
Within the same ANOVA a second set of contrasts soughtto replicate the task-specific effects but now using onlyseenold images (Supporting Information Table 1 ldquoSeen-onlyimagesrdquo) hence excluding any possible confound associatedwith the presentation of unseen images during retrieval Weused the following contrasts [T(easy1hard)gt (Ssame 1 Oold)][Ssamegt (T(easy1hard)2 1 Oold)2] and [Ooldgt (T(easy1hard)2 1 Ssame)2] The statistical thresholds and masking proce-dure were identical to Exp 1 and Exp 2 initial analysis
In Exp 2 we also performed control tests to rule out thepossibility that our main findings of task-specific dissocia-tions merely reflected differences in task difficulty (Sup-porting Information Table 1 ldquoTask difficultyrdquo) This wasaddressed by comparing conditions that behaviorallyresulted in difficult vs easy retrieval which included thecorresponding ldquohardrdquo versus ldquoeasyrdquo trials for the Tempo-ral task as well as by trials including ldquoflippedrdquo versusldquosamerdquo images in the Spatial task (Fig 1B) The results ofthese additional analyses are reported in the SupportingInformation
Conjunction analyses Between immediate
memory and LTM
To compare the current findings about task-specificretrieval after short delays with previous data using analo-gous retrieval tasks but a much longer retention delay[24 h Kwok et al 2012] we performed ldquobetween-studiesrdquo analyses Please note that the two paradigms dif-fered not only in terms of the retention delay (which dif-fered by a factor of 43200) but also in the presenceamount of interleaving events between encoding andretrieval In the LTM version participants were exposed to24 h of new episodic experiences following encodingwhich might cause retroactive interference on the alreadyacquired information [Wohldmann et al 2008] whereassuch retroactive interference was absent in the currentstudy on immediate retrieval The video clips were alsoshorter by a factor of about 250 compared to the 42-min
r Neural Correlates of Immediate Episodic Retrieval r
r 2501 r
episode used in the LTM paradigm Because of these differ-ences our analyses focused on commonalities rather thandifferences between LTM and immediate retrieval Theseanalyses considered the current Exp 1 which included thesame retrieval tasks as the LTM experiment Separately foreach of the three tasks we computed the relevant contrasts(eg Tgt (S1O)2) in the two studies and entered these inthree separate two samples t-tests For the identification ofcommon activation across studies we performed null-conjunction analyses (eg [Tgt (S1O)2]LTM [Tgt (S1O)2]ImMem) Statistical threshold were set at P-FWE-corr 5 005 cluster-level corrected at the whole brain level(cluster size estimated and displayed at P-unc 5 0001) con-sistent with all the analyses above For completeness thecontrasts testing for any differences between studies arereported in Supporting Information Table 2
Additional control analyses Retrieval success
Finally we asked to what extent any task-specific effectsidentified in our main analyses depended on the success-ful retrieval of stored information in the clips For this wecarried out additional analyses testing for retrieval per-formance in Exp 1 (ldquoTask 3 Accuracyrdquo in T and S tasks)and for accurate performance on seenold vs unseennewstimuli in Exp 2 (ldquoTask 3 Seen-unseenrdquo in S and O tasks)Please see Supporting Information for details of these anal-yses and the corresponding results
RESULTS
Behavioral Results
Experiment 1
Subjects performed significantly above chance in allthree tasks (all Plt 0001) The percentage of correct accu-racy and response times (RT) were analyzed usingrepeated measures ANOVAs with three levels correspond-ing to the TSO tasks This showed a main effect of con-ditions both for accuracy [F(2 32) 5 8142 Plt 0001] andRT [F(232) 5 3123 Plt 0001] Post hoc t-tests revealedthat subjects performed significantly better and faster inthe Object than Spatial task [Accuracy t(16) 5 1311Plt 0001 RT t(16) 5 652 Plt 0001] and Object than Tem-poral task [Accuracy t(16) 5 979 Plt 0001 RT t(16) 5
671 Plt 0001] Subjects were also more accurate on Tem-poral trials than on Spatial trials [Accuracy t(16) 5 348Plt 0001 but no difference in RT Pgt 01] For the Tempo-ral task we also examined the effect of temporal distanceexpecting that the retrieval would be more difficult whenthe two probes were temporally close (Tshortlt 19 s) com-pared with temporally far (Tlonggt 19 s) during encodingIndeed t-tests showed that subjects responded less accu-rately and slower in short than in long distance trials[Accuracy t(16) 5 391 Plt 0005 RT t(16) 5 315 Plt 001](Fig 1B plot on the left)
Experiment 2
In Exp 2 subjects also performed better than chance inall three tasks (all Plt 0001) Repeated measures ANOVAsshowed main effects of tasks both for accuracy [F(232) 5 919 Plt 0005] and RT [F(232) 5 1766 Plt 0001]Post hoc t-tests revealed that subjects performed withhigher accuracy and shorter RT in the Object comparedwith the Spatial task [Accuracy t(16) 5 406 Plt 0001 RTt(16) 5 532 Plt 0001] and in the Object compared withthe Temporal task [Accuracy t(16) 5 277 Plt 005 RTt(16) 5 383 Plt 0005] The difference in accuracy betweenthe Temporal and Spatial tasks approached significance(Plt 009) while there was no RT difference between Tem-poral and Spatial tasks (Pgt 01) Additional t-tests directlycompared different trial types within each task For theTemporal task subjects performed more accurately andfaster in the ldquoeasyrdquo than ldquohardrdquo trials [Accuracyt(16) 5 657 Plt 0001 RT t(16) 5 527 Plt 0001)] For theSpatial task they were more accurate in recognizingldquosamerdquo than correctly rejecting ldquoflippedrdquo images [Accu-racy t(16) 5 534 Plt 0001 RT Pgt 01] For the Objecttask subjects were faster to respond to unseen Onew
images compared to seen Oold images [RT t(16) 5 294Plt 001] without any difference in accuracy (Pgt 01) (Fig1B plot on the right)
Functional Neuroimaging Task-Specific Retrieval
In both Exp 1 and 2 we assessed brain activity specificto each of the three retrieval tasks (Temporal Spatial andObject) by comparing each task versus the mean of theother two (ldquoTask specificityrdquo contrasts see SupportingInformation Table 1) In Exp 2 we also carried out addi-tional nonindependent control tests using only trials con-taining seenold images (ldquoSeen-only imagesrdquo contrasts)The latter served to ensure that any task-specific dissocia-tion did not arise from the presentation of unseennewimages during the retrieval phase of the Spatial and Objecttasks (ie flipped- and new-image foils)
Scene-chronology judgment (Temporal task)
Exp 1 The comparison of the Temporal task with theother two retrieval conditions revealed significant activa-tion in the right precuneus bilateral caudate and bilateralcuneus (see Table I and Fig 2A clusters in bluemagenta) The signal plots in Figure 2 (panels on the left)show that the precuneus and the right angular gyrus wereactivated more by the Temporal task (average of Tshort andTlong) than by the Spatial and Object tasks Note that theeffect in the right angular gyrus was detected only atuncorrected threshold (P-unclt 0001) and it is reportedhere because it was fully significant in Exp 2 (see below)and as well as in the LTM dataset [Kwok et al 2012]
Together with the main comparison for the Temporaltask in Exp 1 we assessed the effect of temporal distance
r Kwok et al r
r 2502 r
TA
BL
EI
Task
-sp
ecif
icre
trie
val
acti
vati
on
Ex
p
1E
xp
2
Ex
p
2S
een
-on
ly
Clu
ster
Vo
xel
Clu
ster
Vo
xel
Clu
ster
Vo
xel
KP
-co
rr
Zx
yz
kP
-co
rr
Zx
yz
KP
-co
rr
Zx
yz
Sce
ne-
chro
no
log
y(T
emp
ora
l)P
recu
neu
sB
258
lt00
01
71
06
254
47
2666
lt00
01
51
74
272
43
1835
lt00
01
40
06
270
41
Po
st
cin
gu
late
cort
exB
mdashmdash
mdashmdash
564
02
3027
535
62
3631
Cau
dat
eB
148
000
54
302
122
1514
mdashmdash
mdashmdash
mdashmdash
mdashmdash
Cu
neu
sL
820
052
522
215
260
17mdash
mdashmdash
mdashmdash
mdashmdash
mdashC
un
eus
R88
004
25
2118
257
20mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nto
po
lar
cort
exR
mdashmdash
mdashmdash
874
lt0
001
547
2860
970
60
001
575
2860
7S
up
erio
rfr
on
tal
cort
exR
mdashmdash
mdashmdash
685
000
15
3526
2455
889
000
15
6026
2853
An
gu
lar
gy
rus
R43
lt00
01
46
345
269
29
457
00
07
48
444
266
45
509
00
04
45
844
268
43
Sce
ne-
lay
ou
t(S
pat
ial)
Su
peri
or
pari
eta
lg
yru
sL
1142
lt00
01
69
02
21
266
59
2530
lt00
01
66
72
18
268
59
377
00
16
49
92
18
266
59
Do
rsal
occ
ipit
alco
rtex
L5
112
272
759
521
228
280
9mdash
mdashmdash
mdashS
up
ram
arg
inal
gy
rus
L6
212
602
3338
mdashmdash
mdashmdash
mdashmdash
Intr
apar
ieta
lsu
lcu
sL
577
242
239
415
562
362
4445
mdashmdash
mdashmdash
Su
peri
or
pari
eta
lg
yru
sR
1314
lt00
01
67
221
263
59
3026
lt00
01
75
516
266
59
1262
lt00
01
47
426
268
47
Do
rsal
occ
ipit
alco
rtex
R4
1230
293
237
4034
278
294
1030
274
39In
trap
arie
tal
sulc
us
R6
2536
239
4433
40
026
548
402
4045
mdashmdash
mdashmdash
Su
pra
mar
gin
alg
yru
sR
597
602
2435
mdashmdash
mdashmdash
mdashmdash
Su
per
ior
fro
nta
lsu
lcu
sL
213
000
16
452
243
6223
80
078
543
226
22
51mdash
mdashmdash
mdashS
up
erio
rfr
on
tal
sulc
us
R18
20
002
592
302
356
220
009
74
7028
053
mdashmdash
mdashmdash
Infe
rio
rfr
on
tal
gy
rus
L92
003
65
002
516
26mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nta
lg
yru
sR
304
lt00
01
64
054
12
11
536
00
03
60
752
12
25
238
00
78
48
050
12
25
Infe
rio
ro
ccip
ital
gy
rus
L152
00
05
48
12
57
260
210
894
lt00
01
60
32
52
264
211
591
00
02
50
32
50
266
211
Infe
rio
ro
ccip
ital
gy
rus
R186
00
02
67
754
260
213
2277
lt00
01
68
256
258
211
1161
lt00
01
48
750
258
217
Occ
ipit
alp
ole
B21
80
001
484
29
293
2mdash
mdashmdash
mdashmdash
mdashmdash
mdashS
cen
e-re
cog
nit
ion
(Ob
ject
)M
ed
ial
fro
nta
lco
rtex
B478
lt00
01
51
92
351
47
2215
lt00
01
59
62
856
39
314
00
32
46
42
10
56
39
Mid
dle
tem
po
ral
gy
rus
L490
lt00
01
59
42
60
212
225
1661
lt00
01
67
72
62
214
219
162
lt00
01
42
92
60
24
225
Mid
dle
tem
po
ral
gy
rus
R304
lt00
01
61
560
212
216
1017
lt00
01
56
960
26
223
15
lt00
02
29
662
24
223
Orb
ito
-fro
nta
lco
rtex
B95
001
45
040
512
2222
70
049
506
044
219
mdashmdash
mdashmdash
An
gu
lar
gy
rus
L22
70
001
530
251
254
29mdash
mdashmdash
mdashmdash
mdashmdash
mdashA
ng
ula
rg
yru
sR
142
000
64
9857
260
38mdash
mdashmdash
mdashmdash
mdashmdash
mdashR
SC
p
ost
ci
ng
ula
teco
rtex
B47
4lt
000
14
903
242
35mdash
mdashmdash
mdashmdash
mdashmdash
mdash
T-c
on
tras
tsco
mp
ared
each
con
dit
ion
ver
sus
the
mea
no
fth
eo
ther
two
con
dit
ion
s(e
g
T(s
ho
rt1
lon
g)
2gt
(S1
O)
2)
and
was
incl
usi
vel
ym
ask
edw
ith
the
acti
vat
ion
for
the
crit
ical
con
dit
ion
ver
sus
each
of
the
two
oth
erco
nd
itio
ns
(eg
T
(sh
ort
1lo
ng
)2gt
S2
and
T(s
ho
rt1
lon
g)
2gt
O
2)
InE
xp2
we
con
du
cted
two
sep
arat
ese
tso
fco
ntr
asts
atth
eg
rou
p-l
evel
F
irst
t-
con
tras
tsw
ere
con
du
cted
usi
ng
all
six
con
dit
ion
sto
com
par
eea
chco
nd
itio
nv
ersu
sth
em
ean
of
the
oth
ertw
oco
nd
itio
ns
(eg
S (
sam
e1
flip
ped
)gt
[T(e
asy
1h
ard
)1
O(o
ld1
new
)]
2)
see
colu
mn
sin
the
mid
dle
Se
con
d
for
Spat
ial
and
Ob
ject
con
dit
ion
sw
ew
ere
inte
rest
edin
acti
vit
yel
icit
edb
yth
ere
spo
nse
sas
soci
ated
wit
hldquos
een
rdquotr
ials
on
ly(e
g
S sa-
megt
[T(e
asy
1h
ard
)2
1O
old
]2)
se
eri
gh
tmo
stco
lum
ns
and
also
Sup
po
rtin
gIn
form
atio
nT
able
1B
rain
reg
ion
sh
igh
lig
hte
din
bo
ldco
rres
po
nd
toth
eta
sk-s
pec
ific
effe
cts
con
sist
entl
yo
bse
rved
acro
ssal
lex
per
imen
ts
anal
yse
sth
ese
reg
ion
sar
eal
sod
isp
lay
edin
Fig
ure
s2ndash
4P
-val
ues
are
corr
ecte
dfo
rm
ult
iple
com
par
iso
ns
exce
pt
for
the
rig
ht
ang
ula
rg
yru
sin
Exp
1an
dth
em
idd
lete
mp
ora
lg
yri
inE
xp2
Seen
-on
lyth
atar
ere
po
rted
ata
low
eru
nco
rrec
ted
thre
sho
ld(i
nit
alic
s)
Po
st
po
ster
ior
xy
zco
ord
inat
esin
the
stan
dar
dM
NI
spac
eo
fth
eac
tiv
atio
np
eak
sin
the
clu
ster
sk
nu
mb
ero
fv
oxe
lsin
each
clu
ster
R
SC
retr
osp
len
ial
cort
ex
L
R
B
left
ri
gh
th
emis
ph
ere
bil
ater
al
r Neural Correlates of Immediate Episodic Retrieval r
r 2503 r
by comparing Tshort vs Tlong trials In previous studiesusing LTM protocols we found that temporal distance fur-ther modulated activity in the precuneus [Kwok et al2012 2014 see also St Jacques et al 2008] Here this con-trast did not reveal any significant effect This is in con-trast to the significant behavioral cost of retrieving Tshort
vs Tlong temporal distances (see behavioral results andFig 1B plot on the left) that was analogous to our previ-ous LTM protocol Similar behavioral patterns have beenobserved in many other short-term studies [Hacker 1980Konishi et al 2002 Milner et al 1991 Muter 1979] yet toour knowledge there does not seem to exists a clear con-
sensus on the neural correlates subserving the temporaldistance effect separating two temporally very nearbyeventsitems
Exp 2 In Exp 2 we replicated the effect of the temporalretrieval task in the precuneus and found a fully signifi-cant effect in the right angular gyrus (see Fig 2 clusters inredmagenta) In addition in Exp 2 there was a significantactivation of the right superior frontal cortex and the rightinferior frontopolar cortex that were not found in Exp 1(Table I) The involvement of the precuneus and the rightangular gyrus was further confirmed using only trials
Figure 2
Clusters of brain activation and signal plots for the scene-
chronology judgment (Temporal task) Clusters of activation and
signal plots for (A) the precuneus and (B) the right angular
gyrus These regions activated selectively during the temporal
tasks in Exp 1 (in blue) Exp 2 (in red overlap between Exp 1
and 2 shown in magenta) The activation selective for the Tem-
poral task was computed with the contrast T(short1long)
2gt (S 1 O)2 inclusively masked with T(short1long)2gt S2 and
with T(short1long)2gtO2 The involvement of the precuneus and
the right angular gyrus was further confirmed using only ldquoseen
oldrdquo images (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 40 voxels Effect sizes are mean adjusted (sum to
zero) and expressed in arbitrary units (au 6 90 CI) LR left
right PreC precuneus AnG angular gyrus
r Kwok et al r
r 2504 r
including seen images (cf ldquoExp 2 Seen- onlyrdquo Table I)The signal plots on Figure 2 (panels on the right) show theactivity of the precuneus and the right angular gyrus forall the conditions of Exp 2 Both in the precuneus and theright angular gyrus the average activity in the temporaltask [T(easy1hard)2] were larger than the Ssame and theOold conditions Notably Exp 2 included the presentationof a single image and that the participants were asked tojudge the absolute temporal position of the probe ratherthan the relative temporal order Despite this importantchange of how temporal memory was probed the findingof analogous patterns of activation across both experi-ments highlights the link between immediate retrieval oftemporal information and activation of the precuneus andangular gyrus
Scene-layout judgment (Spatial task)
Exp 1 The contrast concerning the retrieval of the spatiallayout revealed activation in the superior parietal cortexbilaterally with the cluster extending to the intraparietalsulcus and the supramarginal gyrus This contrast alsoshowed activation of the superior frontal sulci the inferiorfrontal gyri plus the inferior occipital cortex (see Table Iand Fig 3 clusters displayed in bluemagenta signalsplots on the left side)
Exp 2 The same comparison in Exp 2 replicated theresults of Exp 1 showing activation in the superior parie-tal cortex bilaterally the superior frontal sulci the inferiorand dorsal occipital cortices and the right inferior frontalgyrus (Fig 3 clusters in redmagenta) When we consid-ered only the conditions including the seenold images(ldquoExp 2 Seen- onlyrdquo) we replicated the effects in the supe-rior parietal gyri the inferior occipital gyri and the rightinferior frontal gyrus (see Table I and Fig 3 signal plotson the right) The activation in the intraparietal sulci andthe superior frontal sulci did not reach statistical signifi-cance Inspection of the signal plots indicated that in theseareas there was less activation in the Ssame than Sflipped tri-als (cf bar 3 vs bar 4 signal plots of the superior fontalsulcus Fig 3A plot on the right) Nonetheless the activityin the Ssame condition was larger than those in the T andO conditions (cf bar 3 vs bars 1ndash2 and 5ndash6)
Scene-recognition judgment (Object task)
Exp 1 The Object task was associated with the activationof the medial frontal cortex (MFC) and the orbitofrontalcortex (but the latter did not fully replicate in Exp 2 seeTable I and Fig 4 clusters bluemagenta and signal plotson the left) together with bilateral activation in the middletemporal gyri the retrosplenialposterior cingulate gyrusand a portion of the angular gyrus The same contrast alsorevealed an activation of the hippocampus bilaterally Theactivation of the hippocampus was just below statisticalsignificance (P-FWE-corr 5 0052 Z 5 458 at x y z 5 2126 219 and P-FWE-corr 5 0077 Z 5 458 at x y z 5 224
29 225 considering the whole brain as the volume ofinterest) and it is reported here because the same effectwas previously found in the LTM dataset [Kwok et al2012 see also Discussion Section]
Exp 2 An almost identical pattern of activation wasfound in Exp 2 when we considered all the conditionsO(old1new)gt (T(easy1hard) 1 S(same1flipped))2 Significanteffects were found in the middle temporal gyri and alarge cluster including the medial frontal gyrus extendingventrally into the orbital-frontal cortex (Fig 4 clusters inredmagenta) However when we considered only the tri-als including seenold-images [Ooldgt (T(easy1hard)2 1 Ssame)2] the Object task was found to significantlyactivate only the MFC (Table I rightmost columns) indi-cating that this region was not affected by incidentalencoding
In summary across all three tasks the brain activationwas found to be similar across Exp 1 and Exp 2 Thescene-chronology Temporal task activated the precuneusand the right angular gyrus the scene-layout Spatial taskwas associated with activation of the posteriorsuperiorparietal cortex and the right inferior frontal gyrus whilethe scene-recognition Object task activated the MFC andthe middle temporal gurus (see Table I areas highlightedin bold) The control analysis of Exp 2 that consideredonly trials including seenold images confirmed thesetask-specific effects indicating that the pattern of dissocia-tionmdashand in particular for the Spatial and Object tasksmdashwas not merely driven by the unseennew images duringthe retrieval phase
Functional Neuroimaging Correspondence with
An Analogous LTM Dataset
We directly compared the task-specific dissociationobserved in the current dataset (Exp 1) with our previousLTM study that included the same task-specific retrievaltasks but after a much longer retention delay [ie 24 hbetween encoding and retrieval Kwok et al 2012] Com-mon across the two data sets the conjunction analysesrevealed an activation cluster in the precuneus for theTemporal task (Fig 5A and Table II) and the dorsal fron-toparietal network plus right inferior frontal gyrus andright occipital cortex for the Spatial task (Fig 5B and TableII) These formal tests show that the Temporal and Spatialtask-specific dissociation observed here for immediateretrieval matched well with the patterns observed duringretrieval from LTM By contrast for the Object task over-lapping effects in the MFC and in the hippocampus weredetected only at a more lenient uncorrected threshold ofP-uncorr 5 0001 In the hippocampus the lack of a fullysignificant ldquocommonrdquo effect across datasets was due to thefact that in the current Exp 1 the contrast comparing theObject task versus the two other tasks did not survive cor-rection for multiple comparisons considering the wholebrain as the volume of interest (P-corr 5 0052 and 0077
r Neural Correlates of Immediate Episodic Retrieval r
r 2505 r
Figure 3
Clusters of brain activation and signal plots for the Scene-layout
judgment (Spatial task) Clusters of activation and signal plots for
(A) the superior frontal cortex and the superior parietal cortex
bilaterally and (B) the right inferior occipital cortex and the right
inferior frontal cortex These regions activated for the Spatial task
in Exp 1 (blue) and in Exp 2 (red overlap between Exp 1 and Exp 2
in magenta) The activation selective for the Spatial task was com-
puted with the contrast Sgt [T(short1long)2 1 O]2 inclusively
masked with SgtT(short1long)2 and with SgtO The effects in the
superior parietal gyri the inferior occipital gyri and the right inferior
frontal gyrus were replicated when considering only sameold
images in Exp 2 (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 80 voxels Effect sizes are mean adjusted (sum to zero)
and are expressed in arbitrary units (au 6 90 CI) LR leftright
SFS superior frontal sulcus SPG superior parietal gyrus IFG infe-
rior frontal gyrus IOG inferior occipital gyrus
in the left and right hemispheres respectively cf above)Indeed the formal test assessing differences between thetwo datasets (ie ldquoTask 3 Experimentrdquo interactions seeSupporting Information Table 2) did not reveal any signifi-cant difference between LTM and immediate retrieval inthe hippocampus consistent with this region beinginvolved in the scene-recognition judgment irrespective ofthe retention delay (see also Discussion Section)
DISCUSSION
We have identified distinct sets of brain areas thatshowed task-specific activation during immediate retrievalfollowing the encoding of dynamic cinematic material Theprecuneus and the right angular gyrus were activatedwhile retrieving the temporal order of events (scene-chro-nology judgment) the superior parietal cortex wasrecruited during spatial related recall (scene-layout
Figure 4
Clusters of brain activation and signal plots for the Scene-
recognition judgment (Object task) Clusters of activation and
signal plots for (A) the MFC and (B) the MTG These regions
activated for the Object task in Exp 1 (blue) and in Exp 2 (red
overlap in magenta) The activation selective for the Object task
was computed with the contrast Ogt [T(short1long)2 1 S]2
inclusively masked with OgtT(short1long)2 and with Ogt S The
signal plots refer to the left hemisphere but analogous patterns
of activation were found in the right hemisphere (Table I) In
Exp 2 the task-specific effect of the Object task in the MFC
was replicated when considering only the sameold images
(Table I rightmost columns) Inspection of the signal plot for the
left MTG revealed less activity in the Oold than Onew trials (plots
on the right bars 5 and 6) although the Oold trials showed
numerically larger parameter estimates than the T and S condi-
tions (see p-uncorr reported in italics in Table I rightmost col-
umns) Activation clusters were estimated and displayed at P-
unc 5 0001 thresholded at a minimum of 80 voxels Effect sizes
are mean adjusted (sum to zero) and expressed in arbitrary
units (au 6 90 CI) LR leftright MTG middle temporal
gyrus MFC medial frontal cortex
r Neural Correlates of Immediate Episodic Retrieval r
r 2507 r
judgment) whereas activity in the MFC and the middletemporal gyrus (MTG) reflected processes associated withscene-recognition judgments These task-specific effectsoccurred independently of whether subjects were asked tojudge seenold or unseennew stimuli (cf ldquoSeen-onlyimagesrdquo contrasts in Exp 2) and independently ofretrieval success and general task difficulty (see Support-ing Information) With respect to the hypothesized corre-spondence between immediate retrieval and retrieval fromLTM we found that these task-specific dissociationsare independent of the informationrsquos mnemonic history(24 h vs 2 s retention ie differing by 43200 times inmagnitude)
One important qualifier for the following discussion isthat memory retrieval in all our tasks depended minimallyon memory buffers compared to other simpler itemsdis-plays that are typically used in ldquostandardrdquo short-termworking memory tasks [Shallice and Cooper 2011 Shiffrin1973] In each trial the number of items presented during
encoding was clearly well beyond the 4 6 1 items stipulatedby classical STM models [Cowan 2001] While memories ofsome materials (eg words spatial positions simple visualdisplays) are supported by limited-capacity buffers [Badde-ley and Hitch 1974] memory for other categories [egunknown faces and scenes see Shiffrin 1973] will tap intoLTM when the task requires the encoding of two or moreitems [Shallice and Cooper 2011] For this reason werestrict our discussion on the tripartite task-specific dissoci-ations with reference to highly complex naturalisticmaterial
Precuneus and Inferior Parietal Cortex Support
Temporal Order Recollection
The scene-chronology judgment task activated the pre-cuneus in both Exp 1 and 2 despite the considerablechanges in the trial structure for probing the retrieval oftemporal information (ie relative temporal order betweentwo probes in Exp 1 vs absolute temporal position of asingle probe in Exp 2) The chronology-judgment taskrequired retrieving the event sequences of many complex
Figure 5
Task-specific retrieval activation common in studies after imme-
diate delays and a longer delay Activation for the LTM study
[24 h Kwok et al 2012] is depicted in green and activation
for the current immediate retrieval study (data from current
Exp 1) depicted in red Activation clusters for both studies were
estimated and displayed at P-unc 5 0001
TABLE II Common memory retrieval activation
ImMem LTM common effects
Cluster Voxel
k P-corr Z x y z
Scene-chronology (Temporal)Precuneus B 112 0011 472 3 257 47Angular gyrus R 98 lt 0001 400 54 254 17
Scene-layout (Spatial)Superior parietal gyrus L 206 0001 595 221 269 56Supramarginal gyrus L 129 0008 346 260 236 44Intraparietal sulcus L 487 236 245 41Superior parietal gyrus R 651 lt 0001 520 24 263 59Dorsal occipital cortex R 372 30 275 29Intraparietal sulcus R 520 42 239 47Supramarginal gyrus R 482 57 233 53Superior frontal sulcus L 100 0022 482 224 3 56Superior frontal sulcus R 145 0004 481 30 0 53Inferior frontal gyrus R 95 0027 475 54 9 26Inferior occipital gyrus R 98 0024 515 54 257 210
Task-specific activation after immediate delays (current Exp 1ImMem) and a previous study that used a long delay betweenencoding and retrieval [24 h LTM Kwok et al 2012] Statisticalthreshold were set at P-FWE-corr 5 005 (cluster-level) consider-ing the whole brain as the volume of interest Note that theldquobetween-studiesrdquo overlap activation in the right angular gyruswas found at a lower uncorrected threshold (in italics) but impor-tantly within this area no differences or interactions betweenImMem and LTM studies were found (see differential effectsbetween the two studies in Supporting Information Table 2) LRB leftright hemispherebilateral
r Kwok et al r
r 2508 r
stimuli (scenes) that entail high-level conceptualhierarchi-cal relationships between them [Swallow et al 2011 Zackset al 2001] The precuneus was previously found to acti-vate in our LTM protocol that used the same temporalorder judgment [Kwok et al 2012 and see also results ofthe conjunction analyses] The current finding of precunealactivation using very different (much shorter) delays fitswell with the view that these processes are not strictlyassociated with retrieval following long retention delayssuggesting that this medial parietal region is implicatedduring temporal order retrieval irrespective of delay (Fig5A and Table II) Specifically we link the effect of chrono-logical retrieval in the precuneus with processes involvingan active reconstruction of past events [see also St Jacqueset al 2008] and possibly memory-related imagery[Fletcher et al 1995 Ishai et al 2000] These processeswould characterize temporal order judgments using com-plex naturalistic material but not order-judgment withsimpler stimulus material which previous studies haveassociated with other brain regions [eg middleprefron-tal areas Suzuki et al 2002]
Together with the precuneus temporal-related judgmentalso gave rise to activation of the right angular gyrus(AG) One possible interpretation relates the inferior parie-tal cortex to the engagement of attentional processes[Majerus et al 2007 2010] The scene-chronology judg-ment of Exp 2 provided us with a further qualifier to thiskind of temporal-related attention Although the Temporaltask in Exp 2 did not require any explicit temporal orderjudgment (only classifying a scene into either the first orsecond half of the movie) an accurate decision still neces-sitated an evaluation of the target scene with many othersegments of the movie This operation is fundamentallydifferent from the evaluation implicated in the scene-layout judgment which required decisions on comparingbetween a single scene and its own correspondentencoded snapshot The latter involves visuospatial analysisof the configuration within scenes The AG may subservedifferent types of attention among which one of them islinked with memory search operations [see Majerus et al2010 2006 also showing coactivation of these two regionsduring order retrieval of simple stimuli after short delays]
Superior Parietal Cortex Evaluates Spatial
Details Against Memory Representation
The second pattern consistently found across experi-ments (Exp 1 and Exp 2 see Fig 3) and across studieswith different retention delays (conjunction betweenimmediate retrieval and LTM studies see Fig 5B andTable II) was the activation of the superior parietal lobule(SPL) during the scene-layout judgment Previous studiesshowed sustained activation in the parietal cortex duringworking memory delays [Courtney et al 1998] and thatactivity there correlates with memory load [Todd andMaroi 2004] Here the amount of information contained
in the clips was equated in the three tasks especially inExp 2 where we counterbalanced the assignment of mov-ies across tasks The subjects also could not anticipatewhat specific type of information would be tested atretrieval and needed to keep rehearsing different aspectsof the clips Because of this we deem unlikely that theactivation of SPL that here was specific for the scene-layout tasks reflected general rehearsal processes [Magenet al 2009] andor updating operations [Bledowski et al2009 Leung et al 2007]
Instead we suggest a role for the SPL in mediating aninterface between the internal memory representation andexternal information It has been proposed that the supe-rior parietal cortex is engaged in allocating top-downattention to orient to aspects of the available visual inputand on information retrieved from memory [Ishai et al2002 Lepsien and Nobre 2007 Summerfield et al 2006]In line with this we found SPL activation when the sub-jects had to make decisions by comparing between theirmemory of the encoded clips and the scene stimuli pre-sented at retrieval This procedure might act on somevisuospatial storage which is held by the lateral occipitalcortex and mediated by the intraparietal sulcus (Fig 3B)The immediate memory task here was analogous to theSpatial task of our previous LTM study [Kwok et al2012] which also required matchingcomparing the avail-able external visual input and internal information storedin memory [see also Nobre et al 2004] This interfaceinvolves a range of top-down processes such as monitor-ing and verification [Cabeza et al 2008 Ciaramelli et al2008] which are independent of the probe imagersquos mne-monic status as shown by the equated effect sizes forldquosamerdquo and ldquoflippedrdquo images conditions (see signals plotsin Fig 3 and also ldquoRetrieval successrdquo analyses of Exp 2 inSupporting Information)
Recognition Processes in the Medial Frontal
Cortex and the Temporal Cortex
The third retrieval task involved scene-recognition judg-ments and was associated with the activation of the MFCand the MTG Moreover in Exp 1 we found a statisticaltrend in the hippocampus bilaterally an area that we pre-viously associated with scene-recognition judgment fromLTM (see next section about ldquoPossible correspondencesbetween retrieval from short- and long-term memoryrdquo)
The MFC was engaged both Exp 1 and 2 but was notactivated in the other two tasks thus a broad prefrontalcortexworking memory correspondence [Tsuchida andFellows 2009] seems insufficient to explain this task-specific effect The scene-recognition task in Exp 2 taxedboth recognition of ldquooldrdquo scenes and correct rejection ofldquonewrdquo scenes suggesting that the MFC here did not dis-tinguish between the correct rejection of new information(unseennew trials) and the recognition of old imagesOne candidate cognitive process that is common to both
r Neural Correlates of Immediate Episodic Retrieval r
r 2509 r
types of processes is feeling-of-knowing (FOK) FOKdepend on the cue specification content retrieval andappraisal of the retrieved memory fragments but are inde-pendent of the ultimate determination of content appropri-ateness [Schnyer et al 2005] Using imaging data duringsentence completion recognition Schnyer et al [2005]modeled a FOK-related directional path The path impli-cated the temporal cortex and the hippocampus andengaged the ventro-MFC at the end which is consistentwith our findings for the Object task implicating the MFCthe middle temporal cortex and at a lower statisticalthreshold the hippocampus
By contrast the activity in the MTG was larger forunseennew images than for seenold memory probes(Fig 4B) This suggests that new encoding may have con-tributed to the activation of the MTG during the retrievalphase of the Object task in Exp 1 The scene-recognitiontask required a lower precision in terms of retrievingdetails about the encoded material and could be solved bysimple recognition [Wixted and Squire 2010] Thus alter-natively larger activation for ldquonew vs oldrdquo stimuli mayrelate to some reduced activation associated with familiar-ity processes on old trials [see Henson et al 1999 whoreported reduced familiarity related activation in middletemporal gyri bilaterally and Henson and Rugg 2003 forreview] However we should point out that in Exp 2 theunseen images were extracted from edited-out parts of theoriginal clips and therefore contained the same objectcharacter depicted in the target images Therefore wedeem more likely that the relational association betweenobjects rather than their presenceabsence in the sceneshelped subjects to distinguish targets from foils We thussuggest that the activation of the MTG reflects processesutilizing the relational details among objects embedded inthe probe images for reaching the recognition decision[Diana et al 2007 Finke et al 2008 Hannula et al 2006Olson et al 2006] Future studies should consider theinclusion of some correlative behavioral measures to indexfamiliarity processes to elucidate further this issue More-over while the Temporal and Spatial tasks required mak-ing relative comparisons about aspects of the encodedvideo-clip the scene-recognition judgment merely requiredjudging whether the image presented at retrieval was pres-ent (or not) within the clip This may entail different proc-essing dynamics in the Object task compared with theother two tasks (cf also differences in retrieval times) Thelatter could be addressed using more subtle experimentalmanipulations for the scene-recognition judgment forexample by using digitally-modified foil images where asingle object in the scene has been modifiedremoved
Possible Correspondences Between Retrieval
from Short- and Long-Term Memory
The overlap between our current study and the previousLTM study using analogous tasks and materials lends
weight to the notion of scale-invariance in memory mecha-nisms [Brown et al 2007] as have been previously dem-onstrated in free recall tasks where the rate of item recallwas unvarying across recall span (day week year) andeven across from the past to the prospective future [May-lor et al 2001] Those previous behavioral findings implythat scale-invariance is a form of self-similarity in that theholistic pattern can exist at multiple levels of magnification[and even across different memory systems eg Maylor2002] Here our data affirmed the importance of the poste-rior parietal cortex in both immediate and long-termretrieval [Berryhill et al 2011 2007] and further character-ized its involvement in dealing with the spatial relationsof objects within scenes [Bricolo et al 2000 Buiatti et al2011] Our data also demonstrated such short-long-termcorrespondence exists in the precuneusangular gyrus fortemporal order retrieval (cf scene-chronology judgmenttask) In contrast the bilateral hippocampus and MFC acti-vations did not reach full statistical significance in theImMem LTM conjunction analysis At first sight thismay indicate that the scene-recognition task tapped intodistinct recognition-based mechanisms as a function ofretention delay However we deem this unlikely becausethe direct comparison between the two datasets did notshow any significant effect in these regions (see Support-ing Information Table 2) Indeed a more targeted test ask-ing the question of whether the hippocampus clusterspreviously found in LTM activated also in the currentimmediate retrieval experiments revealed significanteffects in both immediate retrieval experiments here (Exp1 x y z 5 224 29 225 P-FWE-corrlt 0001 Z 5 458 x yz 5 21 29 222 P-FWE-corr 5 0001 Z 5 451 and in Exp2 x y z 5 224 214 221 P-FWE-corr 5 0004 Z 5 396using the LTM clusters for small volume correction) Theseadditional tests point to a more general issue of sensitivitythat future studies may address using different methodo-logical approaches such as multivariate methods that canbe more sensitive than the standard univariate analysesused here Moreover the former would enable capturingtask-specific effects that are expressed as patterns of dis-tributed activation rather than local changes of activity asreported here [eg Jimura and Poldrack 2012]
Task Difficulty and Retrieval Success
We used several control analyses to show that the task-specific patterns of activation and the correspondencebetween the current data and our previous LTM study donot merely reflect some effect related to general task diffi-culty (see Supporting Information) First areas involved inoverall task difficulty should show progressively greateractivation with progressively more difficult tasks In Exp 1this would entail SgtTgtO However the patterns in theprecuneus and the angular gyrus (with TgtOgt S Fig 2A)and the MFC (Ogt SgtT Fig 4A) did not fit with simplepredictions based on overall task difficulty We also
r Kwok et al r
r 2510 r
assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r

switches of scenes The mean length was 959 s with arange of 772ndash1140 s We preserved the background soundand used only segments without dialoguesconversationsThe whole set of 160 clips was used in Exp 1 and each clipwas assigned to one of the three retrieval tasks 96 Tempo-ral 32 Spatial and 32 Object clips For Exp 2 we selected asubset of 144 clips and each clip was used for all the threetasks counterbalanced across subjects
Clip analyses
We performed a frame-by-frame analysis to mark thechanges of scene for each clip The scene changes includedboth between-scenes cuts (no scene continuity ie edit cutfrom one scene to another) and within-scenes cuts [scenecontinuity but with a change of camera perspective Smithand Henderson 2008] This frame-by-frame analysis allowed
Figure 1
Task schema and behavioral performance for Experiments 1 and
2 (A) Trial timeline probe images and cues at retrieval Each
trial carried an identical structure consisting of presentation of a
short movie clip followed by a cued retrieval test The retrieval
test comprised either a pair of probe images (Exp 1) or one sin-
gle probe (either target or foil Exp 2) (B) Accuracy presented
in bars ( correct unit on left ordinate axes) and response
times in lines (ms right ordinate axes) for Experiment 1 (left
panel) and Experiment 2 (right panel) Error bars are standard
error of the means
r Kwok et al r
r 2498 r
us to identify boundary frames that divided segmentswithin each clip The number of segments across all clipsranged between 4 and 10 The first and last segments werealways excluded for the retrieval tests to mitigate end-of-listdistinctiveness effects [Cabeza et al 1997 Konishi et al2002] Specifically for Exp 2 we divided each clip into 20segments with equal duration The memory probe imageswere extracted from 12 of these segments 3ndash5 (1st quartile)7ndash9 (2nd quartile) 12ndash14 (3rd quartile) and 16ndash18 (4th quar-tile) (cf Temporal task)
Probe images extraction
Exp 1 After excluding the first and last segments for theTemporal trials we pseudorandomly selected two imagesfrom two different segments from each temporal clip Thisrepresented a range of temporal distances between thetwo probe images across all the Temporal trials The tem-poral distances varied between 060 and 496 s The Spatialtrials were generated by pairing a target image with itsown mirror image (by laterally flipping the target image)The Object trials were generated by pairing a target imagewith an unseen image that also contained the same objectcharacter from an edited-out part of the original clip
Exp 2 Irrespective of task we selected one probe imagefrom each clip which was used as the target image Thisimage was extracted with an equal likelihood from one ofthe 4 quartiles (see Clip analyses above) thus with half ofthem coming from the first half and the other half fromthe second half of the clip For the Spatial task the foilimages were mirror images of the targets whereas for theObject task new images were extracted from an edited-outpart of the original clip as the foil images
Trial structure
The trial structure was identical in Exp 1 and 2 eachtrial began with a green fixation cross for 05 s followedby the presentation of a clip lasting for 772ndash114 s then ablank screen for a period of 15ndash25 s This was followedby a retrieval test (Fig 1A) The two experiments differedin how memory was probed during retrieval The retrievaltest of Exp 1 contained a pair of probe images and a writ-ten cue above the images both of which were presentedon the screen for 4 s whereas that of Exp 2 contained onlyone probe image and a written cue both presented for 5 sIn both experiments the written cue specified what taskthe subject had to perform on that trial Trials were sepa-rated by a white fixation cross with jittered ITIs sampledfrom a truncated logarithmic distribution (trial durationsranged between 207 and 228 s) We presented the stimuliusing Cogent Toolbox (httpwwwvislabuclacukcogentphp) implemented in MATLAB 74 (The Math-Works Natick MA) During fMRI scanning all stimuliwere back-projected on a screen at the back of the MRbore and viewed through a mirror system (20 3 15 visual
angle) Participants were given detailed written instruc-tions and were explained of the task requirements prior tothe commencement of the experiments
Retrieval Tasks
Experiment 1
The retrieval test included three main experimental con-ditions Temporal task (96 trials) Spatial task (32 trials)and Object task (32 trials) The reason for the larger num-ber of Temporal trials was that this task comprised twodifferent trial types (Tshort and Tlong) that were classifiedmdashand modeled separately in the main fMRI analysismdashaccording to the temporal distance between the two probeimages (see also Supporting Information Table 1) This dis-tinction was motivated by previous findings showing thatthe temporal separation of the probes at encoding canaffect retrieval performance and brain activity during tem-poral order judgments with naturalistic material [Kwoket al 2012 St Jacques et al 2008]
For all three tasks each retrieval trial comprised a pairof probe images arranged with a leftright side-by-sideconfiguration one of the images was the target the otherwas the foil The left-right position of the targetfoilimages was counterbalanced within conditions Subjectsindicated the leftright stimulus by pressing either one ofthe two keys of a response box Scene-chronology judg-ment (Temporal task T) Subjects were instructed toremember the order of the events in the clip so as tochoose the image that had happened at an earlier timepoint Scene-layout judgment (Spatial task S) Subjectswere instructed to focus on the spatial layout and choosethe probe image that had the identical spatial arrangementas depicted in the clip Scene-recognition judgment (Objecttask O) Subjects were instructed to focus on the contentof the probe images and to identify the image they hadseen in the clip
It should be noted that the retrieval of complex stimulias those used here can be achieved via many differentstrategies For example to solve the Spatial task partici-pants may attend to and store the location of a specificperson in the scene the facing direction of a car and soon rather than processing the overall spatial details withinthe scene For this reason we emphasize that the ldquolabelsrdquoused here (TemporalSpatialObject) do not imply anyexclusive strategy for solving the tasks but rather aremeant to refer to the specificity of the type of judgmentimplicated during retrieval
Experiment 2
The aim of Exp 2 was to test for task-specific dissocia-tions comparing trials that included only ldquoseenrdquo images(ie images extracted from the video) The retrieval testincluded the three main conditions (Temporal task 48 tri-als Spatial task 48 trials and Object task 48 trials) but
r Neural Correlates of Immediate Episodic Retrieval r
r 2499 r
now each trial comprised one single probe image Scene-chronology (Temporal) Subjects were required to recallthe probe imagersquos temporal location with respect to themidpoint of the clip by choosing between the ldquofirst halfrdquoor the ldquosecond halfrdquo of the clip Subjects were required topress the left button if the probe image happened in thefirst half or the right button if the image happened in thesecond half Scene-layout (Spatial) Subjects wereinstructed to focus on the spatial layout and to judgewhether the probe depicted a scene with a spatial arrange-ment as identical to the clip or it was a mirror image Anidentical spatial configuration required the subject to pressthe ldquocorrectrdquo button whereas a mirror image required apress to the ldquoincorrectrdquo button The mapping of ldquocorrectincorrectrdquo (leftright) layouts was counterbalanced acrosssubjects Scene-recognition (Object) Subjects wereinstructed to focus on the content of the probe image andto indicate whether the image had been shown in the clipIf it had been shown the subject responded with theldquopresentrdquo button otherwise with the ldquoabsentrdquo buttonAgain the leftright mapping of ldquopresentabsentrdquoresponses was counterbalanced between subjects
Since only a single image was now presented atretrieval the Temporal task could not involve any factorof ldquotemporal distancerdquo (cf TshortTlong in Exp 1) Insteadin Exp 2 we found that the behavioral performance wasaffected by the ldquotemporal positionrdquo of the probe imageduring encoding (easier if extracted from the startend ofthe clip harder if extracted from around the middle)Accordingly the temporal trials were now classified andmodeled separately in the fMRI analysis as Teasy and Thard
(see also Supporting Information Table 1)
fMRI Data Acquisition and Preprocessing
The functional images were acquired on a Siemens Alle-gra (Siemens Medical Systems Erlangen Germany) 3 Teslascanner equipped for echo-planar imaging (EPI) A quad-rature volume head coil was used for radio frequencytransmission and reception Head movement was mini-mized by mild restraint and cushioning Thirty-two slicesof functional MR images were acquired using blood oxy-genation level-dependent imaging (3 3 3 mm in-plane25 mm thick 50 distance factor repetition time 5 208 secho time 5 30 ms flip angle 5 70 FOV 5 192 mm acqui-sition order 5 continuous ascending) covering the wholecortex We acquired 1648 and 1488 fMRI volumes foreach subject in Exp 1 and 2 respectively Images wereacquired across 4 fMRI runs in both experiments The totalacquisition periods lasted for 55 and 49 min respectively
Data preprocessing was performed with SPM8 (wwwfilionuclacukspm) implemented on Matlab 74 (The Math-Works Natick MA) After having discarded the first 4volumes in each run images were realigned to correct forhead movements Slice-acquisition delays were correctedusing the middle slice as a reference Images were then
normalized to the MNI EPI template resampled to 3 mmisotropic voxel size and spatially smoothed using an iso-tropic Gaussian Kernel (FWHM 5 8 mm)
fMRI Analyses
Statistical inference was based on a random effectsapproach This comprised two steps first-level analysesestimating contrasts of interest for each subject followedby second-level analyses for statistical inference at thegroup-level [Penny and Holmes 2004] For each of thetwo experiments we used several different modelsdepending on the specific questions that we sought toaddress (see below and Supporting Information Table 1)
All the first-level subject-specific models included oneregressor modeling the presentation of the video clips irre-spective of task plus several regressors that modeled theretrieval phase according to the different conditions ofinterest (see leftmost column in Supporting InformationTable 1) The regressor modeling the ldquoencoding phaserdquo(Movie-regressor) was time-locked to the onset of eachvideo and had an event duration corresponding to theduration of each specific video (ie 772ndash1140 s) All theretrieval-related regressors were time-locked to the onsetof the probe image(s) and with a duration correspondingto the probe presentation at retrieval that is 4 s in Exp 1and 5 s in Exp 2 All regressors were convolved with theSPM8 canonical hemodynamic response function Timeseries were high-pass filtered at 128 s and prewhitened bymeans of autoregressive model AR(1) The sections belowdetail how the trials were assigned to the retrieval regres-sorsconditions in the different analyses
Experiment 1
For Exp 1 we used two different sets of first-level mod-els (see Supporting Information Table 1 leftmost column)The first set of models included the encoding-regressor(Movie) plus 5 retrieval-related conditions temporal-short(Tshort) temporal-long (Tlong) spatial (S) object (O) trialsand a condition modeling all the error trials irrespective oftask (Error) The trials belonging to the Temporal taskwere modeled with two separate regressors accounting forthe factor of temporal distance [Tshort vs Tlong cf Kwoket al 2012] All temporal trials including probes with atemporal distance shorter than 19 s were modeled asTshort while those longer than 19 s modeled as Tlong
The main second-level analysis consisted of a within-subjects Analysis of variance (ANOVA) modeling the foureffects of interest Tshort Tlong S and O conditions Foreach subject and each of the four conditions a contrastimage was computed in the first-level model by averagingthe corresponding parameter estimates across the 4 fMRIruns These subject-specific contrast images were enteredin the second-level ANOVA For the identification of task-specific retrieval effects we compared each task with themean of the two other tasks averaging the two Temporal
r Kwok et al r
r 2500 r
conditions ldquoT(short1long)2gt (S 1 O)2rdquo ldquoSgt (T(short1long)2 1 O)2rdquo and ldquoOgt (T(short1long)2 1 S)2rdquo Sphericity-correction was applied to all group-level ANOVAs toaccount for any nonindependent error term for repeatedmeasures and any difference in error variance across con-ditions [Friston et al 2002] The P-values were correctedfor multiple comparisons using a cluster-level threshold ofP-FWE-corr 5 005 considering the whole brain as the vol-ume of interest The cluster size was estimated using aninitial voxel-level threshold of P-unc 5 0001 To ensurethe specificity of any task effect the main differential con-trast was inclusively masked with 2 additional effectsThese corresponded to the activation for the critical taskvs each of the two other tasks for example the Temporalcontrast ldquoT(short1long)gt (S 1 O)rdquo was inclusively maskedwith ldquoT(short1long)2gt Srdquo and ldquoT(short1long)2gtOrdquo Forthese additional and nonindependent masking contraststhe threshold was set to P-unc 5 005 Together with themain contrasts that compared the three retrieval taskswithin this ANOVA we also tested for the effect ofldquotemporal distancerdquo (TshortgtTlong)
Using the same first-level models we ran a secondwithin-subject ANOVA that now also included the averageresponse times (RT) for each condition per subject as acovariate of no interest (Supporting Information Table 1ldquoControl for task difficultyrdquo) The aim of this was to reas-sess the pattern of task-specific dissociation after havingaccounted for any differences in task difficulty (Fig 1B)This RT-controlled analysis was not independent of themain analysis and the results are reported in the Support-ing Information
Experiment 2
The aim of Exp 2 was to account for the possible influ-ence of ldquounseennewrdquo images that were presented as foilsduring the retrieval phase of the Spatial and Object tasksin Exp 1 (eg incidental encoding during retrieval) InExp 2 a single probe image was presented during theretrieval phase of the trial (Fig 1A Experiment 2) Thisprocedure imposed a new constraint on the Temporaltask which now could not include the factor of ldquotemporaldistancerdquo Instead we taxed temporal retrieval by askingparticipants to judge whether the single probe image wasextracted from the first or second half of the clip Asexpected the participants were faster and more accurate tojudge probes presented either near the beginning or theend of the clip compared with probes extracted fromaround the middle of the clip Thus we now divided thetemporal trials into Teasy and Thard accounting for thisbehavioral effect
For Exp 2 we performed one single set of first-levelmodels and a single second-level group-analysis The first-level models included the encoding-regressor (Movie) and6 retrieval-related regressors Teasy (probe extracted fromthe 1st or 4th quartile) Thard (probe extracted from the2nd or 3rd quartile) Ssame Sflipped Oold and Onew plus
one regressor modeling all the error trials (see SupportingInformation Table 1)
The group level analysis consisted of a within-subjectsANOVA that included the 6 conditions of interest TeasyThard Ssame Sflipped Oold and Onew Within this ANOVAwe first tested for the task-specific effects by comparingeach task vs the mean of the other two tasks In this initialanalysis we averaged the Teasy and Thard conditions ofthe Temporal task as well as seen and unseen images ofthe Spatial and Object tasks for example[T(easy1hard)gt (S(same1flipped) 1 O(old1new))2] The statisticalthresholds used here were the same as Exp 1 statisticalmaps were corrected for multiple comparisons using acluster-level threshold of P-FWE-corr 5 005 (whole-brain)The cluster size was estimated using an initial voxel-levelthreshold of P-unc 5 0001 Again all comparisons wereinclusively masked with the effect of the main task vseach of the two other tasks (see Exp 1 above for details)
Within the same ANOVA a second set of contrasts soughtto replicate the task-specific effects but now using onlyseenold images (Supporting Information Table 1 ldquoSeen-onlyimagesrdquo) hence excluding any possible confound associatedwith the presentation of unseen images during retrieval Weused the following contrasts [T(easy1hard)gt (Ssame 1 Oold)][Ssamegt (T(easy1hard)2 1 Oold)2] and [Ooldgt (T(easy1hard)2 1 Ssame)2] The statistical thresholds and masking proce-dure were identical to Exp 1 and Exp 2 initial analysis
In Exp 2 we also performed control tests to rule out thepossibility that our main findings of task-specific dissocia-tions merely reflected differences in task difficulty (Sup-porting Information Table 1 ldquoTask difficultyrdquo) This wasaddressed by comparing conditions that behaviorallyresulted in difficult vs easy retrieval which included thecorresponding ldquohardrdquo versus ldquoeasyrdquo trials for the Tempo-ral task as well as by trials including ldquoflippedrdquo versusldquosamerdquo images in the Spatial task (Fig 1B) The results ofthese additional analyses are reported in the SupportingInformation
Conjunction analyses Between immediate
memory and LTM
To compare the current findings about task-specificretrieval after short delays with previous data using analo-gous retrieval tasks but a much longer retention delay[24 h Kwok et al 2012] we performed ldquobetween-studiesrdquo analyses Please note that the two paradigms dif-fered not only in terms of the retention delay (which dif-fered by a factor of 43200) but also in the presenceamount of interleaving events between encoding andretrieval In the LTM version participants were exposed to24 h of new episodic experiences following encodingwhich might cause retroactive interference on the alreadyacquired information [Wohldmann et al 2008] whereassuch retroactive interference was absent in the currentstudy on immediate retrieval The video clips were alsoshorter by a factor of about 250 compared to the 42-min
r Neural Correlates of Immediate Episodic Retrieval r
r 2501 r
episode used in the LTM paradigm Because of these differ-ences our analyses focused on commonalities rather thandifferences between LTM and immediate retrieval Theseanalyses considered the current Exp 1 which included thesame retrieval tasks as the LTM experiment Separately foreach of the three tasks we computed the relevant contrasts(eg Tgt (S1O)2) in the two studies and entered these inthree separate two samples t-tests For the identification ofcommon activation across studies we performed null-conjunction analyses (eg [Tgt (S1O)2]LTM [Tgt (S1O)2]ImMem) Statistical threshold were set at P-FWE-corr 5 005 cluster-level corrected at the whole brain level(cluster size estimated and displayed at P-unc 5 0001) con-sistent with all the analyses above For completeness thecontrasts testing for any differences between studies arereported in Supporting Information Table 2
Additional control analyses Retrieval success
Finally we asked to what extent any task-specific effectsidentified in our main analyses depended on the success-ful retrieval of stored information in the clips For this wecarried out additional analyses testing for retrieval per-formance in Exp 1 (ldquoTask 3 Accuracyrdquo in T and S tasks)and for accurate performance on seenold vs unseennewstimuli in Exp 2 (ldquoTask 3 Seen-unseenrdquo in S and O tasks)Please see Supporting Information for details of these anal-yses and the corresponding results
RESULTS
Behavioral Results
Experiment 1
Subjects performed significantly above chance in allthree tasks (all Plt 0001) The percentage of correct accu-racy and response times (RT) were analyzed usingrepeated measures ANOVAs with three levels correspond-ing to the TSO tasks This showed a main effect of con-ditions both for accuracy [F(2 32) 5 8142 Plt 0001] andRT [F(232) 5 3123 Plt 0001] Post hoc t-tests revealedthat subjects performed significantly better and faster inthe Object than Spatial task [Accuracy t(16) 5 1311Plt 0001 RT t(16) 5 652 Plt 0001] and Object than Tem-poral task [Accuracy t(16) 5 979 Plt 0001 RT t(16) 5
671 Plt 0001] Subjects were also more accurate on Tem-poral trials than on Spatial trials [Accuracy t(16) 5 348Plt 0001 but no difference in RT Pgt 01] For the Tempo-ral task we also examined the effect of temporal distanceexpecting that the retrieval would be more difficult whenthe two probes were temporally close (Tshortlt 19 s) com-pared with temporally far (Tlonggt 19 s) during encodingIndeed t-tests showed that subjects responded less accu-rately and slower in short than in long distance trials[Accuracy t(16) 5 391 Plt 0005 RT t(16) 5 315 Plt 001](Fig 1B plot on the left)
Experiment 2
In Exp 2 subjects also performed better than chance inall three tasks (all Plt 0001) Repeated measures ANOVAsshowed main effects of tasks both for accuracy [F(232) 5 919 Plt 0005] and RT [F(232) 5 1766 Plt 0001]Post hoc t-tests revealed that subjects performed withhigher accuracy and shorter RT in the Object comparedwith the Spatial task [Accuracy t(16) 5 406 Plt 0001 RTt(16) 5 532 Plt 0001] and in the Object compared withthe Temporal task [Accuracy t(16) 5 277 Plt 005 RTt(16) 5 383 Plt 0005] The difference in accuracy betweenthe Temporal and Spatial tasks approached significance(Plt 009) while there was no RT difference between Tem-poral and Spatial tasks (Pgt 01) Additional t-tests directlycompared different trial types within each task For theTemporal task subjects performed more accurately andfaster in the ldquoeasyrdquo than ldquohardrdquo trials [Accuracyt(16) 5 657 Plt 0001 RT t(16) 5 527 Plt 0001)] For theSpatial task they were more accurate in recognizingldquosamerdquo than correctly rejecting ldquoflippedrdquo images [Accu-racy t(16) 5 534 Plt 0001 RT Pgt 01] For the Objecttask subjects were faster to respond to unseen Onew
images compared to seen Oold images [RT t(16) 5 294Plt 001] without any difference in accuracy (Pgt 01) (Fig1B plot on the right)
Functional Neuroimaging Task-Specific Retrieval
In both Exp 1 and 2 we assessed brain activity specificto each of the three retrieval tasks (Temporal Spatial andObject) by comparing each task versus the mean of theother two (ldquoTask specificityrdquo contrasts see SupportingInformation Table 1) In Exp 2 we also carried out addi-tional nonindependent control tests using only trials con-taining seenold images (ldquoSeen-only imagesrdquo contrasts)The latter served to ensure that any task-specific dissocia-tion did not arise from the presentation of unseennewimages during the retrieval phase of the Spatial and Objecttasks (ie flipped- and new-image foils)
Scene-chronology judgment (Temporal task)
Exp 1 The comparison of the Temporal task with theother two retrieval conditions revealed significant activa-tion in the right precuneus bilateral caudate and bilateralcuneus (see Table I and Fig 2A clusters in bluemagenta) The signal plots in Figure 2 (panels on the left)show that the precuneus and the right angular gyrus wereactivated more by the Temporal task (average of Tshort andTlong) than by the Spatial and Object tasks Note that theeffect in the right angular gyrus was detected only atuncorrected threshold (P-unclt 0001) and it is reportedhere because it was fully significant in Exp 2 (see below)and as well as in the LTM dataset [Kwok et al 2012]
Together with the main comparison for the Temporaltask in Exp 1 we assessed the effect of temporal distance
r Kwok et al r
r 2502 r
TA
BL
EI
Task
-sp
ecif
icre
trie
val
acti
vati
on
Ex
p
1E
xp
2
Ex
p
2S
een
-on
ly
Clu
ster
Vo
xel
Clu
ster
Vo
xel
Clu
ster
Vo
xel
KP
-co
rr
Zx
yz
kP
-co
rr
Zx
yz
KP
-co
rr
Zx
yz
Sce
ne-
chro
no
log
y(T
emp
ora
l)P
recu
neu
sB
258
lt00
01
71
06
254
47
2666
lt00
01
51
74
272
43
1835
lt00
01
40
06
270
41
Po
st
cin
gu
late
cort
exB
mdashmdash
mdashmdash
564
02
3027
535
62
3631
Cau
dat
eB
148
000
54
302
122
1514
mdashmdash
mdashmdash
mdashmdash
mdashmdash
Cu
neu
sL
820
052
522
215
260
17mdash
mdashmdash
mdashmdash
mdashmdash
mdashC
un
eus
R88
004
25
2118
257
20mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nto
po
lar
cort
exR
mdashmdash
mdashmdash
874
lt0
001
547
2860
970
60
001
575
2860
7S
up
erio
rfr
on
tal
cort
exR
mdashmdash
mdashmdash
685
000
15
3526
2455
889
000
15
6026
2853
An
gu
lar
gy
rus
R43
lt00
01
46
345
269
29
457
00
07
48
444
266
45
509
00
04
45
844
268
43
Sce
ne-
lay
ou
t(S
pat
ial)
Su
peri
or
pari
eta
lg
yru
sL
1142
lt00
01
69
02
21
266
59
2530
lt00
01
66
72
18
268
59
377
00
16
49
92
18
266
59
Do
rsal
occ
ipit
alco
rtex
L5
112
272
759
521
228
280
9mdash
mdashmdash
mdashS
up
ram
arg
inal
gy
rus
L6
212
602
3338
mdashmdash
mdashmdash
mdashmdash
Intr
apar
ieta
lsu
lcu
sL
577
242
239
415
562
362
4445
mdashmdash
mdashmdash
Su
peri
or
pari
eta
lg
yru
sR
1314
lt00
01
67
221
263
59
3026
lt00
01
75
516
266
59
1262
lt00
01
47
426
268
47
Do
rsal
occ
ipit
alco
rtex
R4
1230
293
237
4034
278
294
1030
274
39In
trap
arie
tal
sulc
us
R6
2536
239
4433
40
026
548
402
4045
mdashmdash
mdashmdash
Su
pra
mar
gin
alg
yru
sR
597
602
2435
mdashmdash
mdashmdash
mdashmdash
Su
per
ior
fro
nta
lsu
lcu
sL
213
000
16
452
243
6223
80
078
543
226
22
51mdash
mdashmdash
mdashS
up
erio
rfr
on
tal
sulc
us
R18
20
002
592
302
356
220
009
74
7028
053
mdashmdash
mdashmdash
Infe
rio
rfr
on
tal
gy
rus
L92
003
65
002
516
26mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nta
lg
yru
sR
304
lt00
01
64
054
12
11
536
00
03
60
752
12
25
238
00
78
48
050
12
25
Infe
rio
ro
ccip
ital
gy
rus
L152
00
05
48
12
57
260
210
894
lt00
01
60
32
52
264
211
591
00
02
50
32
50
266
211
Infe
rio
ro
ccip
ital
gy
rus
R186
00
02
67
754
260
213
2277
lt00
01
68
256
258
211
1161
lt00
01
48
750
258
217
Occ
ipit
alp
ole
B21
80
001
484
29
293
2mdash
mdashmdash
mdashmdash
mdashmdash
mdashS
cen
e-re
cog
nit
ion
(Ob
ject
)M
ed
ial
fro
nta
lco
rtex
B478
lt00
01
51
92
351
47
2215
lt00
01
59
62
856
39
314
00
32
46
42
10
56
39
Mid
dle
tem
po
ral
gy
rus
L490
lt00
01
59
42
60
212
225
1661
lt00
01
67
72
62
214
219
162
lt00
01
42
92
60
24
225
Mid
dle
tem
po
ral
gy
rus
R304
lt00
01
61
560
212
216
1017
lt00
01
56
960
26
223
15
lt00
02
29
662
24
223
Orb
ito
-fro
nta
lco
rtex
B95
001
45
040
512
2222
70
049
506
044
219
mdashmdash
mdashmdash
An
gu
lar
gy
rus
L22
70
001
530
251
254
29mdash
mdashmdash
mdashmdash
mdashmdash
mdashA
ng
ula
rg
yru
sR
142
000
64
9857
260
38mdash
mdashmdash
mdashmdash
mdashmdash
mdashR
SC
p
ost
ci
ng
ula
teco
rtex
B47
4lt
000
14
903
242
35mdash
mdashmdash
mdashmdash
mdashmdash
mdash
T-c
on
tras
tsco
mp
ared
each
con
dit
ion
ver
sus
the
mea
no
fth
eo
ther
two
con
dit
ion
s(e
g
T(s
ho
rt1
lon
g)
2gt
(S1
O)
2)
and
was
incl
usi
vel
ym
ask
edw
ith
the
acti
vat
ion
for
the
crit
ical
con
dit
ion
ver
sus
each
of
the
two
oth
erco
nd
itio
ns
(eg
T
(sh
ort
1lo
ng
)2gt
S2
and
T(s
ho
rt1
lon
g)
2gt
O
2)
InE
xp2
we
con
du
cted
two
sep
arat
ese
tso
fco
ntr
asts
atth
eg
rou
p-l
evel
F
irst
t-
con
tras
tsw
ere
con
du
cted
usi
ng
all
six
con
dit
ion
sto
com
par
eea
chco
nd
itio
nv
ersu
sth
em
ean
of
the
oth
ertw
oco
nd
itio
ns
(eg
S (
sam
e1
flip
ped
)gt
[T(e
asy
1h
ard
)1
O(o
ld1
new
)]
2)
see
colu
mn
sin
the
mid
dle
Se
con
d
for
Spat
ial
and
Ob
ject
con
dit
ion
sw
ew
ere
inte
rest
edin
acti
vit
yel
icit
edb
yth
ere
spo
nse
sas
soci
ated
wit
hldquos
een
rdquotr
ials
on
ly(e
g
S sa-
megt
[T(e
asy
1h
ard
)2
1O
old
]2)
se
eri
gh
tmo
stco
lum
ns
and
also
Sup
po
rtin
gIn
form
atio
nT
able
1B
rain
reg
ion
sh
igh
lig
hte
din
bo
ldco
rres
po
nd
toth
eta
sk-s
pec
ific
effe
cts
con
sist
entl
yo
bse
rved
acro
ssal
lex
per
imen
ts
anal
yse
sth
ese
reg
ion
sar
eal
sod
isp
lay
edin
Fig
ure
s2ndash
4P
-val
ues
are
corr
ecte
dfo
rm
ult
iple
com
par
iso
ns
exce
pt
for
the
rig
ht
ang
ula
rg
yru
sin
Exp
1an
dth
em
idd
lete
mp
ora
lg
yri
inE
xp2
Seen
-on
lyth
atar
ere
po
rted
ata
low
eru
nco
rrec
ted
thre
sho
ld(i
nit
alic
s)
Po
st
po
ster
ior
xy
zco
ord
inat
esin
the
stan
dar
dM
NI
spac
eo
fth
eac
tiv
atio
np
eak
sin
the
clu
ster
sk
nu
mb
ero
fv
oxe
lsin
each
clu
ster
R
SC
retr
osp
len
ial
cort
ex
L
R
B
left
ri
gh
th
emis
ph
ere
bil
ater
al
r Neural Correlates of Immediate Episodic Retrieval r
r 2503 r
by comparing Tshort vs Tlong trials In previous studiesusing LTM protocols we found that temporal distance fur-ther modulated activity in the precuneus [Kwok et al2012 2014 see also St Jacques et al 2008] Here this con-trast did not reveal any significant effect This is in con-trast to the significant behavioral cost of retrieving Tshort
vs Tlong temporal distances (see behavioral results andFig 1B plot on the left) that was analogous to our previ-ous LTM protocol Similar behavioral patterns have beenobserved in many other short-term studies [Hacker 1980Konishi et al 2002 Milner et al 1991 Muter 1979] yet toour knowledge there does not seem to exists a clear con-
sensus on the neural correlates subserving the temporaldistance effect separating two temporally very nearbyeventsitems
Exp 2 In Exp 2 we replicated the effect of the temporalretrieval task in the precuneus and found a fully signifi-cant effect in the right angular gyrus (see Fig 2 clusters inredmagenta) In addition in Exp 2 there was a significantactivation of the right superior frontal cortex and the rightinferior frontopolar cortex that were not found in Exp 1(Table I) The involvement of the precuneus and the rightangular gyrus was further confirmed using only trials
Figure 2
Clusters of brain activation and signal plots for the scene-
chronology judgment (Temporal task) Clusters of activation and
signal plots for (A) the precuneus and (B) the right angular
gyrus These regions activated selectively during the temporal
tasks in Exp 1 (in blue) Exp 2 (in red overlap between Exp 1
and 2 shown in magenta) The activation selective for the Tem-
poral task was computed with the contrast T(short1long)
2gt (S 1 O)2 inclusively masked with T(short1long)2gt S2 and
with T(short1long)2gtO2 The involvement of the precuneus and
the right angular gyrus was further confirmed using only ldquoseen
oldrdquo images (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 40 voxels Effect sizes are mean adjusted (sum to
zero) and expressed in arbitrary units (au 6 90 CI) LR left
right PreC precuneus AnG angular gyrus
r Kwok et al r
r 2504 r
including seen images (cf ldquoExp 2 Seen- onlyrdquo Table I)The signal plots on Figure 2 (panels on the right) show theactivity of the precuneus and the right angular gyrus forall the conditions of Exp 2 Both in the precuneus and theright angular gyrus the average activity in the temporaltask [T(easy1hard)2] were larger than the Ssame and theOold conditions Notably Exp 2 included the presentationof a single image and that the participants were asked tojudge the absolute temporal position of the probe ratherthan the relative temporal order Despite this importantchange of how temporal memory was probed the findingof analogous patterns of activation across both experi-ments highlights the link between immediate retrieval oftemporal information and activation of the precuneus andangular gyrus
Scene-layout judgment (Spatial task)
Exp 1 The contrast concerning the retrieval of the spatiallayout revealed activation in the superior parietal cortexbilaterally with the cluster extending to the intraparietalsulcus and the supramarginal gyrus This contrast alsoshowed activation of the superior frontal sulci the inferiorfrontal gyri plus the inferior occipital cortex (see Table Iand Fig 3 clusters displayed in bluemagenta signalsplots on the left side)
Exp 2 The same comparison in Exp 2 replicated theresults of Exp 1 showing activation in the superior parie-tal cortex bilaterally the superior frontal sulci the inferiorand dorsal occipital cortices and the right inferior frontalgyrus (Fig 3 clusters in redmagenta) When we consid-ered only the conditions including the seenold images(ldquoExp 2 Seen- onlyrdquo) we replicated the effects in the supe-rior parietal gyri the inferior occipital gyri and the rightinferior frontal gyrus (see Table I and Fig 3 signal plotson the right) The activation in the intraparietal sulci andthe superior frontal sulci did not reach statistical signifi-cance Inspection of the signal plots indicated that in theseareas there was less activation in the Ssame than Sflipped tri-als (cf bar 3 vs bar 4 signal plots of the superior fontalsulcus Fig 3A plot on the right) Nonetheless the activityin the Ssame condition was larger than those in the T andO conditions (cf bar 3 vs bars 1ndash2 and 5ndash6)
Scene-recognition judgment (Object task)
Exp 1 The Object task was associated with the activationof the medial frontal cortex (MFC) and the orbitofrontalcortex (but the latter did not fully replicate in Exp 2 seeTable I and Fig 4 clusters bluemagenta and signal plotson the left) together with bilateral activation in the middletemporal gyri the retrosplenialposterior cingulate gyrusand a portion of the angular gyrus The same contrast alsorevealed an activation of the hippocampus bilaterally Theactivation of the hippocampus was just below statisticalsignificance (P-FWE-corr 5 0052 Z 5 458 at x y z 5 2126 219 and P-FWE-corr 5 0077 Z 5 458 at x y z 5 224
29 225 considering the whole brain as the volume ofinterest) and it is reported here because the same effectwas previously found in the LTM dataset [Kwok et al2012 see also Discussion Section]
Exp 2 An almost identical pattern of activation wasfound in Exp 2 when we considered all the conditionsO(old1new)gt (T(easy1hard) 1 S(same1flipped))2 Significanteffects were found in the middle temporal gyri and alarge cluster including the medial frontal gyrus extendingventrally into the orbital-frontal cortex (Fig 4 clusters inredmagenta) However when we considered only the tri-als including seenold-images [Ooldgt (T(easy1hard)2 1 Ssame)2] the Object task was found to significantlyactivate only the MFC (Table I rightmost columns) indi-cating that this region was not affected by incidentalencoding
In summary across all three tasks the brain activationwas found to be similar across Exp 1 and Exp 2 Thescene-chronology Temporal task activated the precuneusand the right angular gyrus the scene-layout Spatial taskwas associated with activation of the posteriorsuperiorparietal cortex and the right inferior frontal gyrus whilethe scene-recognition Object task activated the MFC andthe middle temporal gurus (see Table I areas highlightedin bold) The control analysis of Exp 2 that consideredonly trials including seenold images confirmed thesetask-specific effects indicating that the pattern of dissocia-tionmdashand in particular for the Spatial and Object tasksmdashwas not merely driven by the unseennew images duringthe retrieval phase
Functional Neuroimaging Correspondence with
An Analogous LTM Dataset
We directly compared the task-specific dissociationobserved in the current dataset (Exp 1) with our previousLTM study that included the same task-specific retrievaltasks but after a much longer retention delay [ie 24 hbetween encoding and retrieval Kwok et al 2012] Com-mon across the two data sets the conjunction analysesrevealed an activation cluster in the precuneus for theTemporal task (Fig 5A and Table II) and the dorsal fron-toparietal network plus right inferior frontal gyrus andright occipital cortex for the Spatial task (Fig 5B and TableII) These formal tests show that the Temporal and Spatialtask-specific dissociation observed here for immediateretrieval matched well with the patterns observed duringretrieval from LTM By contrast for the Object task over-lapping effects in the MFC and in the hippocampus weredetected only at a more lenient uncorrected threshold ofP-uncorr 5 0001 In the hippocampus the lack of a fullysignificant ldquocommonrdquo effect across datasets was due to thefact that in the current Exp 1 the contrast comparing theObject task versus the two other tasks did not survive cor-rection for multiple comparisons considering the wholebrain as the volume of interest (P-corr 5 0052 and 0077
r Neural Correlates of Immediate Episodic Retrieval r
r 2505 r
Figure 3
Clusters of brain activation and signal plots for the Scene-layout
judgment (Spatial task) Clusters of activation and signal plots for
(A) the superior frontal cortex and the superior parietal cortex
bilaterally and (B) the right inferior occipital cortex and the right
inferior frontal cortex These regions activated for the Spatial task
in Exp 1 (blue) and in Exp 2 (red overlap between Exp 1 and Exp 2
in magenta) The activation selective for the Spatial task was com-
puted with the contrast Sgt [T(short1long)2 1 O]2 inclusively
masked with SgtT(short1long)2 and with SgtO The effects in the
superior parietal gyri the inferior occipital gyri and the right inferior
frontal gyrus were replicated when considering only sameold
images in Exp 2 (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 80 voxels Effect sizes are mean adjusted (sum to zero)
and are expressed in arbitrary units (au 6 90 CI) LR leftright
SFS superior frontal sulcus SPG superior parietal gyrus IFG infe-
rior frontal gyrus IOG inferior occipital gyrus
in the left and right hemispheres respectively cf above)Indeed the formal test assessing differences between thetwo datasets (ie ldquoTask 3 Experimentrdquo interactions seeSupporting Information Table 2) did not reveal any signifi-cant difference between LTM and immediate retrieval inthe hippocampus consistent with this region beinginvolved in the scene-recognition judgment irrespective ofthe retention delay (see also Discussion Section)
DISCUSSION
We have identified distinct sets of brain areas thatshowed task-specific activation during immediate retrievalfollowing the encoding of dynamic cinematic material Theprecuneus and the right angular gyrus were activatedwhile retrieving the temporal order of events (scene-chro-nology judgment) the superior parietal cortex wasrecruited during spatial related recall (scene-layout
Figure 4
Clusters of brain activation and signal plots for the Scene-
recognition judgment (Object task) Clusters of activation and
signal plots for (A) the MFC and (B) the MTG These regions
activated for the Object task in Exp 1 (blue) and in Exp 2 (red
overlap in magenta) The activation selective for the Object task
was computed with the contrast Ogt [T(short1long)2 1 S]2
inclusively masked with OgtT(short1long)2 and with Ogt S The
signal plots refer to the left hemisphere but analogous patterns
of activation were found in the right hemisphere (Table I) In
Exp 2 the task-specific effect of the Object task in the MFC
was replicated when considering only the sameold images
(Table I rightmost columns) Inspection of the signal plot for the
left MTG revealed less activity in the Oold than Onew trials (plots
on the right bars 5 and 6) although the Oold trials showed
numerically larger parameter estimates than the T and S condi-
tions (see p-uncorr reported in italics in Table I rightmost col-
umns) Activation clusters were estimated and displayed at P-
unc 5 0001 thresholded at a minimum of 80 voxels Effect sizes
are mean adjusted (sum to zero) and expressed in arbitrary
units (au 6 90 CI) LR leftright MTG middle temporal
gyrus MFC medial frontal cortex
r Neural Correlates of Immediate Episodic Retrieval r
r 2507 r
judgment) whereas activity in the MFC and the middletemporal gyrus (MTG) reflected processes associated withscene-recognition judgments These task-specific effectsoccurred independently of whether subjects were asked tojudge seenold or unseennew stimuli (cf ldquoSeen-onlyimagesrdquo contrasts in Exp 2) and independently ofretrieval success and general task difficulty (see Support-ing Information) With respect to the hypothesized corre-spondence between immediate retrieval and retrieval fromLTM we found that these task-specific dissociationsare independent of the informationrsquos mnemonic history(24 h vs 2 s retention ie differing by 43200 times inmagnitude)
One important qualifier for the following discussion isthat memory retrieval in all our tasks depended minimallyon memory buffers compared to other simpler itemsdis-plays that are typically used in ldquostandardrdquo short-termworking memory tasks [Shallice and Cooper 2011 Shiffrin1973] In each trial the number of items presented during
encoding was clearly well beyond the 4 6 1 items stipulatedby classical STM models [Cowan 2001] While memories ofsome materials (eg words spatial positions simple visualdisplays) are supported by limited-capacity buffers [Badde-ley and Hitch 1974] memory for other categories [egunknown faces and scenes see Shiffrin 1973] will tap intoLTM when the task requires the encoding of two or moreitems [Shallice and Cooper 2011] For this reason werestrict our discussion on the tripartite task-specific dissoci-ations with reference to highly complex naturalisticmaterial
Precuneus and Inferior Parietal Cortex Support
Temporal Order Recollection
The scene-chronology judgment task activated the pre-cuneus in both Exp 1 and 2 despite the considerablechanges in the trial structure for probing the retrieval oftemporal information (ie relative temporal order betweentwo probes in Exp 1 vs absolute temporal position of asingle probe in Exp 2) The chronology-judgment taskrequired retrieving the event sequences of many complex
Figure 5
Task-specific retrieval activation common in studies after imme-
diate delays and a longer delay Activation for the LTM study
[24 h Kwok et al 2012] is depicted in green and activation
for the current immediate retrieval study (data from current
Exp 1) depicted in red Activation clusters for both studies were
estimated and displayed at P-unc 5 0001
TABLE II Common memory retrieval activation
ImMem LTM common effects
Cluster Voxel
k P-corr Z x y z
Scene-chronology (Temporal)Precuneus B 112 0011 472 3 257 47Angular gyrus R 98 lt 0001 400 54 254 17
Scene-layout (Spatial)Superior parietal gyrus L 206 0001 595 221 269 56Supramarginal gyrus L 129 0008 346 260 236 44Intraparietal sulcus L 487 236 245 41Superior parietal gyrus R 651 lt 0001 520 24 263 59Dorsal occipital cortex R 372 30 275 29Intraparietal sulcus R 520 42 239 47Supramarginal gyrus R 482 57 233 53Superior frontal sulcus L 100 0022 482 224 3 56Superior frontal sulcus R 145 0004 481 30 0 53Inferior frontal gyrus R 95 0027 475 54 9 26Inferior occipital gyrus R 98 0024 515 54 257 210
Task-specific activation after immediate delays (current Exp 1ImMem) and a previous study that used a long delay betweenencoding and retrieval [24 h LTM Kwok et al 2012] Statisticalthreshold were set at P-FWE-corr 5 005 (cluster-level) consider-ing the whole brain as the volume of interest Note that theldquobetween-studiesrdquo overlap activation in the right angular gyruswas found at a lower uncorrected threshold (in italics) but impor-tantly within this area no differences or interactions betweenImMem and LTM studies were found (see differential effectsbetween the two studies in Supporting Information Table 2) LRB leftright hemispherebilateral
r Kwok et al r
r 2508 r
stimuli (scenes) that entail high-level conceptualhierarchi-cal relationships between them [Swallow et al 2011 Zackset al 2001] The precuneus was previously found to acti-vate in our LTM protocol that used the same temporalorder judgment [Kwok et al 2012 and see also results ofthe conjunction analyses] The current finding of precunealactivation using very different (much shorter) delays fitswell with the view that these processes are not strictlyassociated with retrieval following long retention delayssuggesting that this medial parietal region is implicatedduring temporal order retrieval irrespective of delay (Fig5A and Table II) Specifically we link the effect of chrono-logical retrieval in the precuneus with processes involvingan active reconstruction of past events [see also St Jacqueset al 2008] and possibly memory-related imagery[Fletcher et al 1995 Ishai et al 2000] These processeswould characterize temporal order judgments using com-plex naturalistic material but not order-judgment withsimpler stimulus material which previous studies haveassociated with other brain regions [eg middleprefron-tal areas Suzuki et al 2002]
Together with the precuneus temporal-related judgmentalso gave rise to activation of the right angular gyrus(AG) One possible interpretation relates the inferior parie-tal cortex to the engagement of attentional processes[Majerus et al 2007 2010] The scene-chronology judg-ment of Exp 2 provided us with a further qualifier to thiskind of temporal-related attention Although the Temporaltask in Exp 2 did not require any explicit temporal orderjudgment (only classifying a scene into either the first orsecond half of the movie) an accurate decision still neces-sitated an evaluation of the target scene with many othersegments of the movie This operation is fundamentallydifferent from the evaluation implicated in the scene-layout judgment which required decisions on comparingbetween a single scene and its own correspondentencoded snapshot The latter involves visuospatial analysisof the configuration within scenes The AG may subservedifferent types of attention among which one of them islinked with memory search operations [see Majerus et al2010 2006 also showing coactivation of these two regionsduring order retrieval of simple stimuli after short delays]
Superior Parietal Cortex Evaluates Spatial
Details Against Memory Representation
The second pattern consistently found across experi-ments (Exp 1 and Exp 2 see Fig 3) and across studieswith different retention delays (conjunction betweenimmediate retrieval and LTM studies see Fig 5B andTable II) was the activation of the superior parietal lobule(SPL) during the scene-layout judgment Previous studiesshowed sustained activation in the parietal cortex duringworking memory delays [Courtney et al 1998] and thatactivity there correlates with memory load [Todd andMaroi 2004] Here the amount of information contained
in the clips was equated in the three tasks especially inExp 2 where we counterbalanced the assignment of mov-ies across tasks The subjects also could not anticipatewhat specific type of information would be tested atretrieval and needed to keep rehearsing different aspectsof the clips Because of this we deem unlikely that theactivation of SPL that here was specific for the scene-layout tasks reflected general rehearsal processes [Magenet al 2009] andor updating operations [Bledowski et al2009 Leung et al 2007]
Instead we suggest a role for the SPL in mediating aninterface between the internal memory representation andexternal information It has been proposed that the supe-rior parietal cortex is engaged in allocating top-downattention to orient to aspects of the available visual inputand on information retrieved from memory [Ishai et al2002 Lepsien and Nobre 2007 Summerfield et al 2006]In line with this we found SPL activation when the sub-jects had to make decisions by comparing between theirmemory of the encoded clips and the scene stimuli pre-sented at retrieval This procedure might act on somevisuospatial storage which is held by the lateral occipitalcortex and mediated by the intraparietal sulcus (Fig 3B)The immediate memory task here was analogous to theSpatial task of our previous LTM study [Kwok et al2012] which also required matchingcomparing the avail-able external visual input and internal information storedin memory [see also Nobre et al 2004] This interfaceinvolves a range of top-down processes such as monitor-ing and verification [Cabeza et al 2008 Ciaramelli et al2008] which are independent of the probe imagersquos mne-monic status as shown by the equated effect sizes forldquosamerdquo and ldquoflippedrdquo images conditions (see signals plotsin Fig 3 and also ldquoRetrieval successrdquo analyses of Exp 2 inSupporting Information)
Recognition Processes in the Medial Frontal
Cortex and the Temporal Cortex
The third retrieval task involved scene-recognition judg-ments and was associated with the activation of the MFCand the MTG Moreover in Exp 1 we found a statisticaltrend in the hippocampus bilaterally an area that we pre-viously associated with scene-recognition judgment fromLTM (see next section about ldquoPossible correspondencesbetween retrieval from short- and long-term memoryrdquo)
The MFC was engaged both Exp 1 and 2 but was notactivated in the other two tasks thus a broad prefrontalcortexworking memory correspondence [Tsuchida andFellows 2009] seems insufficient to explain this task-specific effect The scene-recognition task in Exp 2 taxedboth recognition of ldquooldrdquo scenes and correct rejection ofldquonewrdquo scenes suggesting that the MFC here did not dis-tinguish between the correct rejection of new information(unseennew trials) and the recognition of old imagesOne candidate cognitive process that is common to both
r Neural Correlates of Immediate Episodic Retrieval r
r 2509 r
types of processes is feeling-of-knowing (FOK) FOKdepend on the cue specification content retrieval andappraisal of the retrieved memory fragments but are inde-pendent of the ultimate determination of content appropri-ateness [Schnyer et al 2005] Using imaging data duringsentence completion recognition Schnyer et al [2005]modeled a FOK-related directional path The path impli-cated the temporal cortex and the hippocampus andengaged the ventro-MFC at the end which is consistentwith our findings for the Object task implicating the MFCthe middle temporal cortex and at a lower statisticalthreshold the hippocampus
By contrast the activity in the MTG was larger forunseennew images than for seenold memory probes(Fig 4B) This suggests that new encoding may have con-tributed to the activation of the MTG during the retrievalphase of the Object task in Exp 1 The scene-recognitiontask required a lower precision in terms of retrievingdetails about the encoded material and could be solved bysimple recognition [Wixted and Squire 2010] Thus alter-natively larger activation for ldquonew vs oldrdquo stimuli mayrelate to some reduced activation associated with familiar-ity processes on old trials [see Henson et al 1999 whoreported reduced familiarity related activation in middletemporal gyri bilaterally and Henson and Rugg 2003 forreview] However we should point out that in Exp 2 theunseen images were extracted from edited-out parts of theoriginal clips and therefore contained the same objectcharacter depicted in the target images Therefore wedeem more likely that the relational association betweenobjects rather than their presenceabsence in the sceneshelped subjects to distinguish targets from foils We thussuggest that the activation of the MTG reflects processesutilizing the relational details among objects embedded inthe probe images for reaching the recognition decision[Diana et al 2007 Finke et al 2008 Hannula et al 2006Olson et al 2006] Future studies should consider theinclusion of some correlative behavioral measures to indexfamiliarity processes to elucidate further this issue More-over while the Temporal and Spatial tasks required mak-ing relative comparisons about aspects of the encodedvideo-clip the scene-recognition judgment merely requiredjudging whether the image presented at retrieval was pres-ent (or not) within the clip This may entail different proc-essing dynamics in the Object task compared with theother two tasks (cf also differences in retrieval times) Thelatter could be addressed using more subtle experimentalmanipulations for the scene-recognition judgment forexample by using digitally-modified foil images where asingle object in the scene has been modifiedremoved
Possible Correspondences Between Retrieval
from Short- and Long-Term Memory
The overlap between our current study and the previousLTM study using analogous tasks and materials lends
weight to the notion of scale-invariance in memory mecha-nisms [Brown et al 2007] as have been previously dem-onstrated in free recall tasks where the rate of item recallwas unvarying across recall span (day week year) andeven across from the past to the prospective future [May-lor et al 2001] Those previous behavioral findings implythat scale-invariance is a form of self-similarity in that theholistic pattern can exist at multiple levels of magnification[and even across different memory systems eg Maylor2002] Here our data affirmed the importance of the poste-rior parietal cortex in both immediate and long-termretrieval [Berryhill et al 2011 2007] and further character-ized its involvement in dealing with the spatial relationsof objects within scenes [Bricolo et al 2000 Buiatti et al2011] Our data also demonstrated such short-long-termcorrespondence exists in the precuneusangular gyrus fortemporal order retrieval (cf scene-chronology judgmenttask) In contrast the bilateral hippocampus and MFC acti-vations did not reach full statistical significance in theImMem LTM conjunction analysis At first sight thismay indicate that the scene-recognition task tapped intodistinct recognition-based mechanisms as a function ofretention delay However we deem this unlikely becausethe direct comparison between the two datasets did notshow any significant effect in these regions (see Support-ing Information Table 2) Indeed a more targeted test ask-ing the question of whether the hippocampus clusterspreviously found in LTM activated also in the currentimmediate retrieval experiments revealed significanteffects in both immediate retrieval experiments here (Exp1 x y z 5 224 29 225 P-FWE-corrlt 0001 Z 5 458 x yz 5 21 29 222 P-FWE-corr 5 0001 Z 5 451 and in Exp2 x y z 5 224 214 221 P-FWE-corr 5 0004 Z 5 396using the LTM clusters for small volume correction) Theseadditional tests point to a more general issue of sensitivitythat future studies may address using different methodo-logical approaches such as multivariate methods that canbe more sensitive than the standard univariate analysesused here Moreover the former would enable capturingtask-specific effects that are expressed as patterns of dis-tributed activation rather than local changes of activity asreported here [eg Jimura and Poldrack 2012]
Task Difficulty and Retrieval Success
We used several control analyses to show that the task-specific patterns of activation and the correspondencebetween the current data and our previous LTM study donot merely reflect some effect related to general task diffi-culty (see Supporting Information) First areas involved inoverall task difficulty should show progressively greateractivation with progressively more difficult tasks In Exp 1this would entail SgtTgtO However the patterns in theprecuneus and the angular gyrus (with TgtOgt S Fig 2A)and the MFC (Ogt SgtT Fig 4A) did not fit with simplepredictions based on overall task difficulty We also
r Kwok et al r
r 2510 r
assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r

us to identify boundary frames that divided segmentswithin each clip The number of segments across all clipsranged between 4 and 10 The first and last segments werealways excluded for the retrieval tests to mitigate end-of-listdistinctiveness effects [Cabeza et al 1997 Konishi et al2002] Specifically for Exp 2 we divided each clip into 20segments with equal duration The memory probe imageswere extracted from 12 of these segments 3ndash5 (1st quartile)7ndash9 (2nd quartile) 12ndash14 (3rd quartile) and 16ndash18 (4th quar-tile) (cf Temporal task)
Probe images extraction
Exp 1 After excluding the first and last segments for theTemporal trials we pseudorandomly selected two imagesfrom two different segments from each temporal clip Thisrepresented a range of temporal distances between thetwo probe images across all the Temporal trials The tem-poral distances varied between 060 and 496 s The Spatialtrials were generated by pairing a target image with itsown mirror image (by laterally flipping the target image)The Object trials were generated by pairing a target imagewith an unseen image that also contained the same objectcharacter from an edited-out part of the original clip
Exp 2 Irrespective of task we selected one probe imagefrom each clip which was used as the target image Thisimage was extracted with an equal likelihood from one ofthe 4 quartiles (see Clip analyses above) thus with half ofthem coming from the first half and the other half fromthe second half of the clip For the Spatial task the foilimages were mirror images of the targets whereas for theObject task new images were extracted from an edited-outpart of the original clip as the foil images
Trial structure
The trial structure was identical in Exp 1 and 2 eachtrial began with a green fixation cross for 05 s followedby the presentation of a clip lasting for 772ndash114 s then ablank screen for a period of 15ndash25 s This was followedby a retrieval test (Fig 1A) The two experiments differedin how memory was probed during retrieval The retrievaltest of Exp 1 contained a pair of probe images and a writ-ten cue above the images both of which were presentedon the screen for 4 s whereas that of Exp 2 contained onlyone probe image and a written cue both presented for 5 sIn both experiments the written cue specified what taskthe subject had to perform on that trial Trials were sepa-rated by a white fixation cross with jittered ITIs sampledfrom a truncated logarithmic distribution (trial durationsranged between 207 and 228 s) We presented the stimuliusing Cogent Toolbox (httpwwwvislabuclacukcogentphp) implemented in MATLAB 74 (The Math-Works Natick MA) During fMRI scanning all stimuliwere back-projected on a screen at the back of the MRbore and viewed through a mirror system (20 3 15 visual
angle) Participants were given detailed written instruc-tions and were explained of the task requirements prior tothe commencement of the experiments
Retrieval Tasks
Experiment 1
The retrieval test included three main experimental con-ditions Temporal task (96 trials) Spatial task (32 trials)and Object task (32 trials) The reason for the larger num-ber of Temporal trials was that this task comprised twodifferent trial types (Tshort and Tlong) that were classifiedmdashand modeled separately in the main fMRI analysismdashaccording to the temporal distance between the two probeimages (see also Supporting Information Table 1) This dis-tinction was motivated by previous findings showing thatthe temporal separation of the probes at encoding canaffect retrieval performance and brain activity during tem-poral order judgments with naturalistic material [Kwoket al 2012 St Jacques et al 2008]
For all three tasks each retrieval trial comprised a pairof probe images arranged with a leftright side-by-sideconfiguration one of the images was the target the otherwas the foil The left-right position of the targetfoilimages was counterbalanced within conditions Subjectsindicated the leftright stimulus by pressing either one ofthe two keys of a response box Scene-chronology judg-ment (Temporal task T) Subjects were instructed toremember the order of the events in the clip so as tochoose the image that had happened at an earlier timepoint Scene-layout judgment (Spatial task S) Subjectswere instructed to focus on the spatial layout and choosethe probe image that had the identical spatial arrangementas depicted in the clip Scene-recognition judgment (Objecttask O) Subjects were instructed to focus on the contentof the probe images and to identify the image they hadseen in the clip
It should be noted that the retrieval of complex stimulias those used here can be achieved via many differentstrategies For example to solve the Spatial task partici-pants may attend to and store the location of a specificperson in the scene the facing direction of a car and soon rather than processing the overall spatial details withinthe scene For this reason we emphasize that the ldquolabelsrdquoused here (TemporalSpatialObject) do not imply anyexclusive strategy for solving the tasks but rather aremeant to refer to the specificity of the type of judgmentimplicated during retrieval
Experiment 2
The aim of Exp 2 was to test for task-specific dissocia-tions comparing trials that included only ldquoseenrdquo images(ie images extracted from the video) The retrieval testincluded the three main conditions (Temporal task 48 tri-als Spatial task 48 trials and Object task 48 trials) but
r Neural Correlates of Immediate Episodic Retrieval r
r 2499 r
now each trial comprised one single probe image Scene-chronology (Temporal) Subjects were required to recallthe probe imagersquos temporal location with respect to themidpoint of the clip by choosing between the ldquofirst halfrdquoor the ldquosecond halfrdquo of the clip Subjects were required topress the left button if the probe image happened in thefirst half or the right button if the image happened in thesecond half Scene-layout (Spatial) Subjects wereinstructed to focus on the spatial layout and to judgewhether the probe depicted a scene with a spatial arrange-ment as identical to the clip or it was a mirror image Anidentical spatial configuration required the subject to pressthe ldquocorrectrdquo button whereas a mirror image required apress to the ldquoincorrectrdquo button The mapping of ldquocorrectincorrectrdquo (leftright) layouts was counterbalanced acrosssubjects Scene-recognition (Object) Subjects wereinstructed to focus on the content of the probe image andto indicate whether the image had been shown in the clipIf it had been shown the subject responded with theldquopresentrdquo button otherwise with the ldquoabsentrdquo buttonAgain the leftright mapping of ldquopresentabsentrdquoresponses was counterbalanced between subjects
Since only a single image was now presented atretrieval the Temporal task could not involve any factorof ldquotemporal distancerdquo (cf TshortTlong in Exp 1) Insteadin Exp 2 we found that the behavioral performance wasaffected by the ldquotemporal positionrdquo of the probe imageduring encoding (easier if extracted from the startend ofthe clip harder if extracted from around the middle)Accordingly the temporal trials were now classified andmodeled separately in the fMRI analysis as Teasy and Thard
(see also Supporting Information Table 1)
fMRI Data Acquisition and Preprocessing
The functional images were acquired on a Siemens Alle-gra (Siemens Medical Systems Erlangen Germany) 3 Teslascanner equipped for echo-planar imaging (EPI) A quad-rature volume head coil was used for radio frequencytransmission and reception Head movement was mini-mized by mild restraint and cushioning Thirty-two slicesof functional MR images were acquired using blood oxy-genation level-dependent imaging (3 3 3 mm in-plane25 mm thick 50 distance factor repetition time 5 208 secho time 5 30 ms flip angle 5 70 FOV 5 192 mm acqui-sition order 5 continuous ascending) covering the wholecortex We acquired 1648 and 1488 fMRI volumes foreach subject in Exp 1 and 2 respectively Images wereacquired across 4 fMRI runs in both experiments The totalacquisition periods lasted for 55 and 49 min respectively
Data preprocessing was performed with SPM8 (wwwfilionuclacukspm) implemented on Matlab 74 (The Math-Works Natick MA) After having discarded the first 4volumes in each run images were realigned to correct forhead movements Slice-acquisition delays were correctedusing the middle slice as a reference Images were then
normalized to the MNI EPI template resampled to 3 mmisotropic voxel size and spatially smoothed using an iso-tropic Gaussian Kernel (FWHM 5 8 mm)
fMRI Analyses
Statistical inference was based on a random effectsapproach This comprised two steps first-level analysesestimating contrasts of interest for each subject followedby second-level analyses for statistical inference at thegroup-level [Penny and Holmes 2004] For each of thetwo experiments we used several different modelsdepending on the specific questions that we sought toaddress (see below and Supporting Information Table 1)
All the first-level subject-specific models included oneregressor modeling the presentation of the video clips irre-spective of task plus several regressors that modeled theretrieval phase according to the different conditions ofinterest (see leftmost column in Supporting InformationTable 1) The regressor modeling the ldquoencoding phaserdquo(Movie-regressor) was time-locked to the onset of eachvideo and had an event duration corresponding to theduration of each specific video (ie 772ndash1140 s) All theretrieval-related regressors were time-locked to the onsetof the probe image(s) and with a duration correspondingto the probe presentation at retrieval that is 4 s in Exp 1and 5 s in Exp 2 All regressors were convolved with theSPM8 canonical hemodynamic response function Timeseries were high-pass filtered at 128 s and prewhitened bymeans of autoregressive model AR(1) The sections belowdetail how the trials were assigned to the retrieval regres-sorsconditions in the different analyses
Experiment 1
For Exp 1 we used two different sets of first-level mod-els (see Supporting Information Table 1 leftmost column)The first set of models included the encoding-regressor(Movie) plus 5 retrieval-related conditions temporal-short(Tshort) temporal-long (Tlong) spatial (S) object (O) trialsand a condition modeling all the error trials irrespective oftask (Error) The trials belonging to the Temporal taskwere modeled with two separate regressors accounting forthe factor of temporal distance [Tshort vs Tlong cf Kwoket al 2012] All temporal trials including probes with atemporal distance shorter than 19 s were modeled asTshort while those longer than 19 s modeled as Tlong
The main second-level analysis consisted of a within-subjects Analysis of variance (ANOVA) modeling the foureffects of interest Tshort Tlong S and O conditions Foreach subject and each of the four conditions a contrastimage was computed in the first-level model by averagingthe corresponding parameter estimates across the 4 fMRIruns These subject-specific contrast images were enteredin the second-level ANOVA For the identification of task-specific retrieval effects we compared each task with themean of the two other tasks averaging the two Temporal
r Kwok et al r
r 2500 r
conditions ldquoT(short1long)2gt (S 1 O)2rdquo ldquoSgt (T(short1long)2 1 O)2rdquo and ldquoOgt (T(short1long)2 1 S)2rdquo Sphericity-correction was applied to all group-level ANOVAs toaccount for any nonindependent error term for repeatedmeasures and any difference in error variance across con-ditions [Friston et al 2002] The P-values were correctedfor multiple comparisons using a cluster-level threshold ofP-FWE-corr 5 005 considering the whole brain as the vol-ume of interest The cluster size was estimated using aninitial voxel-level threshold of P-unc 5 0001 To ensurethe specificity of any task effect the main differential con-trast was inclusively masked with 2 additional effectsThese corresponded to the activation for the critical taskvs each of the two other tasks for example the Temporalcontrast ldquoT(short1long)gt (S 1 O)rdquo was inclusively maskedwith ldquoT(short1long)2gt Srdquo and ldquoT(short1long)2gtOrdquo Forthese additional and nonindependent masking contraststhe threshold was set to P-unc 5 005 Together with themain contrasts that compared the three retrieval taskswithin this ANOVA we also tested for the effect ofldquotemporal distancerdquo (TshortgtTlong)
Using the same first-level models we ran a secondwithin-subject ANOVA that now also included the averageresponse times (RT) for each condition per subject as acovariate of no interest (Supporting Information Table 1ldquoControl for task difficultyrdquo) The aim of this was to reas-sess the pattern of task-specific dissociation after havingaccounted for any differences in task difficulty (Fig 1B)This RT-controlled analysis was not independent of themain analysis and the results are reported in the Support-ing Information
Experiment 2
The aim of Exp 2 was to account for the possible influ-ence of ldquounseennewrdquo images that were presented as foilsduring the retrieval phase of the Spatial and Object tasksin Exp 1 (eg incidental encoding during retrieval) InExp 2 a single probe image was presented during theretrieval phase of the trial (Fig 1A Experiment 2) Thisprocedure imposed a new constraint on the Temporaltask which now could not include the factor of ldquotemporaldistancerdquo Instead we taxed temporal retrieval by askingparticipants to judge whether the single probe image wasextracted from the first or second half of the clip Asexpected the participants were faster and more accurate tojudge probes presented either near the beginning or theend of the clip compared with probes extracted fromaround the middle of the clip Thus we now divided thetemporal trials into Teasy and Thard accounting for thisbehavioral effect
For Exp 2 we performed one single set of first-levelmodels and a single second-level group-analysis The first-level models included the encoding-regressor (Movie) and6 retrieval-related regressors Teasy (probe extracted fromthe 1st or 4th quartile) Thard (probe extracted from the2nd or 3rd quartile) Ssame Sflipped Oold and Onew plus
one regressor modeling all the error trials (see SupportingInformation Table 1)
The group level analysis consisted of a within-subjectsANOVA that included the 6 conditions of interest TeasyThard Ssame Sflipped Oold and Onew Within this ANOVAwe first tested for the task-specific effects by comparingeach task vs the mean of the other two tasks In this initialanalysis we averaged the Teasy and Thard conditions ofthe Temporal task as well as seen and unseen images ofthe Spatial and Object tasks for example[T(easy1hard)gt (S(same1flipped) 1 O(old1new))2] The statisticalthresholds used here were the same as Exp 1 statisticalmaps were corrected for multiple comparisons using acluster-level threshold of P-FWE-corr 5 005 (whole-brain)The cluster size was estimated using an initial voxel-levelthreshold of P-unc 5 0001 Again all comparisons wereinclusively masked with the effect of the main task vseach of the two other tasks (see Exp 1 above for details)
Within the same ANOVA a second set of contrasts soughtto replicate the task-specific effects but now using onlyseenold images (Supporting Information Table 1 ldquoSeen-onlyimagesrdquo) hence excluding any possible confound associatedwith the presentation of unseen images during retrieval Weused the following contrasts [T(easy1hard)gt (Ssame 1 Oold)][Ssamegt (T(easy1hard)2 1 Oold)2] and [Ooldgt (T(easy1hard)2 1 Ssame)2] The statistical thresholds and masking proce-dure were identical to Exp 1 and Exp 2 initial analysis
In Exp 2 we also performed control tests to rule out thepossibility that our main findings of task-specific dissocia-tions merely reflected differences in task difficulty (Sup-porting Information Table 1 ldquoTask difficultyrdquo) This wasaddressed by comparing conditions that behaviorallyresulted in difficult vs easy retrieval which included thecorresponding ldquohardrdquo versus ldquoeasyrdquo trials for the Tempo-ral task as well as by trials including ldquoflippedrdquo versusldquosamerdquo images in the Spatial task (Fig 1B) The results ofthese additional analyses are reported in the SupportingInformation
Conjunction analyses Between immediate
memory and LTM
To compare the current findings about task-specificretrieval after short delays with previous data using analo-gous retrieval tasks but a much longer retention delay[24 h Kwok et al 2012] we performed ldquobetween-studiesrdquo analyses Please note that the two paradigms dif-fered not only in terms of the retention delay (which dif-fered by a factor of 43200) but also in the presenceamount of interleaving events between encoding andretrieval In the LTM version participants were exposed to24 h of new episodic experiences following encodingwhich might cause retroactive interference on the alreadyacquired information [Wohldmann et al 2008] whereassuch retroactive interference was absent in the currentstudy on immediate retrieval The video clips were alsoshorter by a factor of about 250 compared to the 42-min
r Neural Correlates of Immediate Episodic Retrieval r
r 2501 r
episode used in the LTM paradigm Because of these differ-ences our analyses focused on commonalities rather thandifferences between LTM and immediate retrieval Theseanalyses considered the current Exp 1 which included thesame retrieval tasks as the LTM experiment Separately foreach of the three tasks we computed the relevant contrasts(eg Tgt (S1O)2) in the two studies and entered these inthree separate two samples t-tests For the identification ofcommon activation across studies we performed null-conjunction analyses (eg [Tgt (S1O)2]LTM [Tgt (S1O)2]ImMem) Statistical threshold were set at P-FWE-corr 5 005 cluster-level corrected at the whole brain level(cluster size estimated and displayed at P-unc 5 0001) con-sistent with all the analyses above For completeness thecontrasts testing for any differences between studies arereported in Supporting Information Table 2
Additional control analyses Retrieval success
Finally we asked to what extent any task-specific effectsidentified in our main analyses depended on the success-ful retrieval of stored information in the clips For this wecarried out additional analyses testing for retrieval per-formance in Exp 1 (ldquoTask 3 Accuracyrdquo in T and S tasks)and for accurate performance on seenold vs unseennewstimuli in Exp 2 (ldquoTask 3 Seen-unseenrdquo in S and O tasks)Please see Supporting Information for details of these anal-yses and the corresponding results
RESULTS
Behavioral Results
Experiment 1
Subjects performed significantly above chance in allthree tasks (all Plt 0001) The percentage of correct accu-racy and response times (RT) were analyzed usingrepeated measures ANOVAs with three levels correspond-ing to the TSO tasks This showed a main effect of con-ditions both for accuracy [F(2 32) 5 8142 Plt 0001] andRT [F(232) 5 3123 Plt 0001] Post hoc t-tests revealedthat subjects performed significantly better and faster inthe Object than Spatial task [Accuracy t(16) 5 1311Plt 0001 RT t(16) 5 652 Plt 0001] and Object than Tem-poral task [Accuracy t(16) 5 979 Plt 0001 RT t(16) 5
671 Plt 0001] Subjects were also more accurate on Tem-poral trials than on Spatial trials [Accuracy t(16) 5 348Plt 0001 but no difference in RT Pgt 01] For the Tempo-ral task we also examined the effect of temporal distanceexpecting that the retrieval would be more difficult whenthe two probes were temporally close (Tshortlt 19 s) com-pared with temporally far (Tlonggt 19 s) during encodingIndeed t-tests showed that subjects responded less accu-rately and slower in short than in long distance trials[Accuracy t(16) 5 391 Plt 0005 RT t(16) 5 315 Plt 001](Fig 1B plot on the left)
Experiment 2
In Exp 2 subjects also performed better than chance inall three tasks (all Plt 0001) Repeated measures ANOVAsshowed main effects of tasks both for accuracy [F(232) 5 919 Plt 0005] and RT [F(232) 5 1766 Plt 0001]Post hoc t-tests revealed that subjects performed withhigher accuracy and shorter RT in the Object comparedwith the Spatial task [Accuracy t(16) 5 406 Plt 0001 RTt(16) 5 532 Plt 0001] and in the Object compared withthe Temporal task [Accuracy t(16) 5 277 Plt 005 RTt(16) 5 383 Plt 0005] The difference in accuracy betweenthe Temporal and Spatial tasks approached significance(Plt 009) while there was no RT difference between Tem-poral and Spatial tasks (Pgt 01) Additional t-tests directlycompared different trial types within each task For theTemporal task subjects performed more accurately andfaster in the ldquoeasyrdquo than ldquohardrdquo trials [Accuracyt(16) 5 657 Plt 0001 RT t(16) 5 527 Plt 0001)] For theSpatial task they were more accurate in recognizingldquosamerdquo than correctly rejecting ldquoflippedrdquo images [Accu-racy t(16) 5 534 Plt 0001 RT Pgt 01] For the Objecttask subjects were faster to respond to unseen Onew
images compared to seen Oold images [RT t(16) 5 294Plt 001] without any difference in accuracy (Pgt 01) (Fig1B plot on the right)
Functional Neuroimaging Task-Specific Retrieval
In both Exp 1 and 2 we assessed brain activity specificto each of the three retrieval tasks (Temporal Spatial andObject) by comparing each task versus the mean of theother two (ldquoTask specificityrdquo contrasts see SupportingInformation Table 1) In Exp 2 we also carried out addi-tional nonindependent control tests using only trials con-taining seenold images (ldquoSeen-only imagesrdquo contrasts)The latter served to ensure that any task-specific dissocia-tion did not arise from the presentation of unseennewimages during the retrieval phase of the Spatial and Objecttasks (ie flipped- and new-image foils)
Scene-chronology judgment (Temporal task)
Exp 1 The comparison of the Temporal task with theother two retrieval conditions revealed significant activa-tion in the right precuneus bilateral caudate and bilateralcuneus (see Table I and Fig 2A clusters in bluemagenta) The signal plots in Figure 2 (panels on the left)show that the precuneus and the right angular gyrus wereactivated more by the Temporal task (average of Tshort andTlong) than by the Spatial and Object tasks Note that theeffect in the right angular gyrus was detected only atuncorrected threshold (P-unclt 0001) and it is reportedhere because it was fully significant in Exp 2 (see below)and as well as in the LTM dataset [Kwok et al 2012]
Together with the main comparison for the Temporaltask in Exp 1 we assessed the effect of temporal distance
r Kwok et al r
r 2502 r
TA
BL
EI
Task
-sp
ecif
icre
trie
val
acti
vati
on
Ex
p
1E
xp
2
Ex
p
2S
een
-on
ly
Clu
ster
Vo
xel
Clu
ster
Vo
xel
Clu
ster
Vo
xel
KP
-co
rr
Zx
yz
kP
-co
rr
Zx
yz
KP
-co
rr
Zx
yz
Sce
ne-
chro
no
log
y(T
emp
ora
l)P
recu
neu
sB
258
lt00
01
71
06
254
47
2666
lt00
01
51
74
272
43
1835
lt00
01
40
06
270
41
Po
st
cin
gu
late
cort
exB
mdashmdash
mdashmdash
564
02
3027
535
62
3631
Cau
dat
eB
148
000
54
302
122
1514
mdashmdash
mdashmdash
mdashmdash
mdashmdash
Cu
neu
sL
820
052
522
215
260
17mdash
mdashmdash
mdashmdash
mdashmdash
mdashC
un
eus
R88
004
25
2118
257
20mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nto
po
lar
cort
exR
mdashmdash
mdashmdash
874
lt0
001
547
2860
970
60
001
575
2860
7S
up
erio
rfr
on
tal
cort
exR
mdashmdash
mdashmdash
685
000
15
3526
2455
889
000
15
6026
2853
An
gu
lar
gy
rus
R43
lt00
01
46
345
269
29
457
00
07
48
444
266
45
509
00
04
45
844
268
43
Sce
ne-
lay
ou
t(S
pat
ial)
Su
peri
or
pari
eta
lg
yru
sL
1142
lt00
01
69
02
21
266
59
2530
lt00
01
66
72
18
268
59
377
00
16
49
92
18
266
59
Do
rsal
occ
ipit
alco
rtex
L5
112
272
759
521
228
280
9mdash
mdashmdash
mdashS
up
ram
arg
inal
gy
rus
L6
212
602
3338
mdashmdash
mdashmdash
mdashmdash
Intr
apar
ieta
lsu
lcu
sL
577
242
239
415
562
362
4445
mdashmdash
mdashmdash
Su
peri
or
pari
eta
lg
yru
sR
1314
lt00
01
67
221
263
59
3026
lt00
01
75
516
266
59
1262
lt00
01
47
426
268
47
Do
rsal
occ
ipit
alco
rtex
R4
1230
293
237
4034
278
294
1030
274
39In
trap
arie
tal
sulc
us
R6
2536
239
4433
40
026
548
402
4045
mdashmdash
mdashmdash
Su
pra
mar
gin
alg
yru
sR
597
602
2435
mdashmdash
mdashmdash
mdashmdash
Su
per
ior
fro
nta
lsu
lcu
sL
213
000
16
452
243
6223
80
078
543
226
22
51mdash
mdashmdash
mdashS
up
erio
rfr
on
tal
sulc
us
R18
20
002
592
302
356
220
009
74
7028
053
mdashmdash
mdashmdash
Infe
rio
rfr
on
tal
gy
rus
L92
003
65
002
516
26mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nta
lg
yru
sR
304
lt00
01
64
054
12
11
536
00
03
60
752
12
25
238
00
78
48
050
12
25
Infe
rio
ro
ccip
ital
gy
rus
L152
00
05
48
12
57
260
210
894
lt00
01
60
32
52
264
211
591
00
02
50
32
50
266
211
Infe
rio
ro
ccip
ital
gy
rus
R186
00
02
67
754
260
213
2277
lt00
01
68
256
258
211
1161
lt00
01
48
750
258
217
Occ
ipit
alp
ole
B21
80
001
484
29
293
2mdash
mdashmdash
mdashmdash
mdashmdash
mdashS
cen
e-re
cog
nit
ion
(Ob
ject
)M
ed
ial
fro
nta
lco
rtex
B478
lt00
01
51
92
351
47
2215
lt00
01
59
62
856
39
314
00
32
46
42
10
56
39
Mid
dle
tem
po
ral
gy
rus
L490
lt00
01
59
42
60
212
225
1661
lt00
01
67
72
62
214
219
162
lt00
01
42
92
60
24
225
Mid
dle
tem
po
ral
gy
rus
R304
lt00
01
61
560
212
216
1017
lt00
01
56
960
26
223
15
lt00
02
29
662
24
223
Orb
ito
-fro
nta
lco
rtex
B95
001
45
040
512
2222
70
049
506
044
219
mdashmdash
mdashmdash
An
gu
lar
gy
rus
L22
70
001
530
251
254
29mdash
mdashmdash
mdashmdash
mdashmdash
mdashA
ng
ula
rg
yru
sR
142
000
64
9857
260
38mdash
mdashmdash
mdashmdash
mdashmdash
mdashR
SC
p
ost
ci
ng
ula
teco
rtex
B47
4lt
000
14
903
242
35mdash
mdashmdash
mdashmdash
mdashmdash
mdash
T-c
on
tras
tsco
mp
ared
each
con
dit
ion
ver
sus
the
mea
no
fth
eo
ther
two
con
dit
ion
s(e
g
T(s
ho
rt1
lon
g)
2gt
(S1
O)
2)
and
was
incl
usi
vel
ym
ask
edw
ith
the
acti
vat
ion
for
the
crit
ical
con
dit
ion
ver
sus
each
of
the
two
oth
erco
nd
itio
ns
(eg
T
(sh
ort
1lo
ng
)2gt
S2
and
T(s
ho
rt1
lon
g)
2gt
O
2)
InE
xp2
we
con
du
cted
two
sep
arat
ese
tso
fco
ntr
asts
atth
eg
rou
p-l
evel
F
irst
t-
con
tras
tsw
ere
con
du
cted
usi
ng
all
six
con
dit
ion
sto
com
par
eea
chco
nd
itio
nv
ersu
sth
em
ean
of
the
oth
ertw
oco
nd
itio
ns
(eg
S (
sam
e1
flip
ped
)gt
[T(e
asy
1h
ard
)1
O(o
ld1
new
)]
2)
see
colu
mn
sin
the
mid
dle
Se
con
d
for
Spat
ial
and
Ob
ject
con
dit
ion
sw
ew
ere
inte
rest
edin
acti
vit
yel
icit
edb
yth
ere
spo
nse
sas
soci
ated
wit
hldquos
een
rdquotr
ials
on
ly(e
g
S sa-
megt
[T(e
asy
1h
ard
)2
1O
old
]2)
se
eri
gh
tmo
stco
lum
ns
and
also
Sup
po
rtin
gIn
form
atio
nT
able
1B
rain
reg
ion
sh
igh
lig
hte
din
bo
ldco
rres
po
nd
toth
eta
sk-s
pec
ific
effe
cts
con
sist
entl
yo
bse
rved
acro
ssal
lex
per
imen
ts
anal
yse
sth
ese
reg
ion
sar
eal
sod
isp
lay
edin
Fig
ure
s2ndash
4P
-val
ues
are
corr
ecte
dfo
rm
ult
iple
com
par
iso
ns
exce
pt
for
the
rig
ht
ang
ula
rg
yru
sin
Exp
1an
dth
em
idd
lete
mp
ora
lg
yri
inE
xp2
Seen
-on
lyth
atar
ere
po
rted
ata
low
eru
nco
rrec
ted
thre
sho
ld(i
nit
alic
s)
Po
st
po
ster
ior
xy
zco
ord
inat
esin
the
stan
dar
dM
NI
spac
eo
fth
eac
tiv
atio
np
eak
sin
the
clu
ster
sk
nu
mb
ero
fv
oxe
lsin
each
clu
ster
R
SC
retr
osp
len
ial
cort
ex
L
R
B
left
ri
gh
th
emis
ph
ere
bil
ater
al
r Neural Correlates of Immediate Episodic Retrieval r
r 2503 r
by comparing Tshort vs Tlong trials In previous studiesusing LTM protocols we found that temporal distance fur-ther modulated activity in the precuneus [Kwok et al2012 2014 see also St Jacques et al 2008] Here this con-trast did not reveal any significant effect This is in con-trast to the significant behavioral cost of retrieving Tshort
vs Tlong temporal distances (see behavioral results andFig 1B plot on the left) that was analogous to our previ-ous LTM protocol Similar behavioral patterns have beenobserved in many other short-term studies [Hacker 1980Konishi et al 2002 Milner et al 1991 Muter 1979] yet toour knowledge there does not seem to exists a clear con-
sensus on the neural correlates subserving the temporaldistance effect separating two temporally very nearbyeventsitems
Exp 2 In Exp 2 we replicated the effect of the temporalretrieval task in the precuneus and found a fully signifi-cant effect in the right angular gyrus (see Fig 2 clusters inredmagenta) In addition in Exp 2 there was a significantactivation of the right superior frontal cortex and the rightinferior frontopolar cortex that were not found in Exp 1(Table I) The involvement of the precuneus and the rightangular gyrus was further confirmed using only trials
Figure 2
Clusters of brain activation and signal plots for the scene-
chronology judgment (Temporal task) Clusters of activation and
signal plots for (A) the precuneus and (B) the right angular
gyrus These regions activated selectively during the temporal
tasks in Exp 1 (in blue) Exp 2 (in red overlap between Exp 1
and 2 shown in magenta) The activation selective for the Tem-
poral task was computed with the contrast T(short1long)
2gt (S 1 O)2 inclusively masked with T(short1long)2gt S2 and
with T(short1long)2gtO2 The involvement of the precuneus and
the right angular gyrus was further confirmed using only ldquoseen
oldrdquo images (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 40 voxels Effect sizes are mean adjusted (sum to
zero) and expressed in arbitrary units (au 6 90 CI) LR left
right PreC precuneus AnG angular gyrus
r Kwok et al r
r 2504 r
including seen images (cf ldquoExp 2 Seen- onlyrdquo Table I)The signal plots on Figure 2 (panels on the right) show theactivity of the precuneus and the right angular gyrus forall the conditions of Exp 2 Both in the precuneus and theright angular gyrus the average activity in the temporaltask [T(easy1hard)2] were larger than the Ssame and theOold conditions Notably Exp 2 included the presentationof a single image and that the participants were asked tojudge the absolute temporal position of the probe ratherthan the relative temporal order Despite this importantchange of how temporal memory was probed the findingof analogous patterns of activation across both experi-ments highlights the link between immediate retrieval oftemporal information and activation of the precuneus andangular gyrus
Scene-layout judgment (Spatial task)
Exp 1 The contrast concerning the retrieval of the spatiallayout revealed activation in the superior parietal cortexbilaterally with the cluster extending to the intraparietalsulcus and the supramarginal gyrus This contrast alsoshowed activation of the superior frontal sulci the inferiorfrontal gyri plus the inferior occipital cortex (see Table Iand Fig 3 clusters displayed in bluemagenta signalsplots on the left side)
Exp 2 The same comparison in Exp 2 replicated theresults of Exp 1 showing activation in the superior parie-tal cortex bilaterally the superior frontal sulci the inferiorand dorsal occipital cortices and the right inferior frontalgyrus (Fig 3 clusters in redmagenta) When we consid-ered only the conditions including the seenold images(ldquoExp 2 Seen- onlyrdquo) we replicated the effects in the supe-rior parietal gyri the inferior occipital gyri and the rightinferior frontal gyrus (see Table I and Fig 3 signal plotson the right) The activation in the intraparietal sulci andthe superior frontal sulci did not reach statistical signifi-cance Inspection of the signal plots indicated that in theseareas there was less activation in the Ssame than Sflipped tri-als (cf bar 3 vs bar 4 signal plots of the superior fontalsulcus Fig 3A plot on the right) Nonetheless the activityin the Ssame condition was larger than those in the T andO conditions (cf bar 3 vs bars 1ndash2 and 5ndash6)
Scene-recognition judgment (Object task)
Exp 1 The Object task was associated with the activationof the medial frontal cortex (MFC) and the orbitofrontalcortex (but the latter did not fully replicate in Exp 2 seeTable I and Fig 4 clusters bluemagenta and signal plotson the left) together with bilateral activation in the middletemporal gyri the retrosplenialposterior cingulate gyrusand a portion of the angular gyrus The same contrast alsorevealed an activation of the hippocampus bilaterally Theactivation of the hippocampus was just below statisticalsignificance (P-FWE-corr 5 0052 Z 5 458 at x y z 5 2126 219 and P-FWE-corr 5 0077 Z 5 458 at x y z 5 224
29 225 considering the whole brain as the volume ofinterest) and it is reported here because the same effectwas previously found in the LTM dataset [Kwok et al2012 see also Discussion Section]
Exp 2 An almost identical pattern of activation wasfound in Exp 2 when we considered all the conditionsO(old1new)gt (T(easy1hard) 1 S(same1flipped))2 Significanteffects were found in the middle temporal gyri and alarge cluster including the medial frontal gyrus extendingventrally into the orbital-frontal cortex (Fig 4 clusters inredmagenta) However when we considered only the tri-als including seenold-images [Ooldgt (T(easy1hard)2 1 Ssame)2] the Object task was found to significantlyactivate only the MFC (Table I rightmost columns) indi-cating that this region was not affected by incidentalencoding
In summary across all three tasks the brain activationwas found to be similar across Exp 1 and Exp 2 Thescene-chronology Temporal task activated the precuneusand the right angular gyrus the scene-layout Spatial taskwas associated with activation of the posteriorsuperiorparietal cortex and the right inferior frontal gyrus whilethe scene-recognition Object task activated the MFC andthe middle temporal gurus (see Table I areas highlightedin bold) The control analysis of Exp 2 that consideredonly trials including seenold images confirmed thesetask-specific effects indicating that the pattern of dissocia-tionmdashand in particular for the Spatial and Object tasksmdashwas not merely driven by the unseennew images duringthe retrieval phase
Functional Neuroimaging Correspondence with
An Analogous LTM Dataset
We directly compared the task-specific dissociationobserved in the current dataset (Exp 1) with our previousLTM study that included the same task-specific retrievaltasks but after a much longer retention delay [ie 24 hbetween encoding and retrieval Kwok et al 2012] Com-mon across the two data sets the conjunction analysesrevealed an activation cluster in the precuneus for theTemporal task (Fig 5A and Table II) and the dorsal fron-toparietal network plus right inferior frontal gyrus andright occipital cortex for the Spatial task (Fig 5B and TableII) These formal tests show that the Temporal and Spatialtask-specific dissociation observed here for immediateretrieval matched well with the patterns observed duringretrieval from LTM By contrast for the Object task over-lapping effects in the MFC and in the hippocampus weredetected only at a more lenient uncorrected threshold ofP-uncorr 5 0001 In the hippocampus the lack of a fullysignificant ldquocommonrdquo effect across datasets was due to thefact that in the current Exp 1 the contrast comparing theObject task versus the two other tasks did not survive cor-rection for multiple comparisons considering the wholebrain as the volume of interest (P-corr 5 0052 and 0077
r Neural Correlates of Immediate Episodic Retrieval r
r 2505 r
Figure 3
Clusters of brain activation and signal plots for the Scene-layout
judgment (Spatial task) Clusters of activation and signal plots for
(A) the superior frontal cortex and the superior parietal cortex
bilaterally and (B) the right inferior occipital cortex and the right
inferior frontal cortex These regions activated for the Spatial task
in Exp 1 (blue) and in Exp 2 (red overlap between Exp 1 and Exp 2
in magenta) The activation selective for the Spatial task was com-
puted with the contrast Sgt [T(short1long)2 1 O]2 inclusively
masked with SgtT(short1long)2 and with SgtO The effects in the
superior parietal gyri the inferior occipital gyri and the right inferior
frontal gyrus were replicated when considering only sameold
images in Exp 2 (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 80 voxels Effect sizes are mean adjusted (sum to zero)
and are expressed in arbitrary units (au 6 90 CI) LR leftright
SFS superior frontal sulcus SPG superior parietal gyrus IFG infe-
rior frontal gyrus IOG inferior occipital gyrus
in the left and right hemispheres respectively cf above)Indeed the formal test assessing differences between thetwo datasets (ie ldquoTask 3 Experimentrdquo interactions seeSupporting Information Table 2) did not reveal any signifi-cant difference between LTM and immediate retrieval inthe hippocampus consistent with this region beinginvolved in the scene-recognition judgment irrespective ofthe retention delay (see also Discussion Section)
DISCUSSION
We have identified distinct sets of brain areas thatshowed task-specific activation during immediate retrievalfollowing the encoding of dynamic cinematic material Theprecuneus and the right angular gyrus were activatedwhile retrieving the temporal order of events (scene-chro-nology judgment) the superior parietal cortex wasrecruited during spatial related recall (scene-layout
Figure 4
Clusters of brain activation and signal plots for the Scene-
recognition judgment (Object task) Clusters of activation and
signal plots for (A) the MFC and (B) the MTG These regions
activated for the Object task in Exp 1 (blue) and in Exp 2 (red
overlap in magenta) The activation selective for the Object task
was computed with the contrast Ogt [T(short1long)2 1 S]2
inclusively masked with OgtT(short1long)2 and with Ogt S The
signal plots refer to the left hemisphere but analogous patterns
of activation were found in the right hemisphere (Table I) In
Exp 2 the task-specific effect of the Object task in the MFC
was replicated when considering only the sameold images
(Table I rightmost columns) Inspection of the signal plot for the
left MTG revealed less activity in the Oold than Onew trials (plots
on the right bars 5 and 6) although the Oold trials showed
numerically larger parameter estimates than the T and S condi-
tions (see p-uncorr reported in italics in Table I rightmost col-
umns) Activation clusters were estimated and displayed at P-
unc 5 0001 thresholded at a minimum of 80 voxels Effect sizes
are mean adjusted (sum to zero) and expressed in arbitrary
units (au 6 90 CI) LR leftright MTG middle temporal
gyrus MFC medial frontal cortex
r Neural Correlates of Immediate Episodic Retrieval r
r 2507 r
judgment) whereas activity in the MFC and the middletemporal gyrus (MTG) reflected processes associated withscene-recognition judgments These task-specific effectsoccurred independently of whether subjects were asked tojudge seenold or unseennew stimuli (cf ldquoSeen-onlyimagesrdquo contrasts in Exp 2) and independently ofretrieval success and general task difficulty (see Support-ing Information) With respect to the hypothesized corre-spondence between immediate retrieval and retrieval fromLTM we found that these task-specific dissociationsare independent of the informationrsquos mnemonic history(24 h vs 2 s retention ie differing by 43200 times inmagnitude)
One important qualifier for the following discussion isthat memory retrieval in all our tasks depended minimallyon memory buffers compared to other simpler itemsdis-plays that are typically used in ldquostandardrdquo short-termworking memory tasks [Shallice and Cooper 2011 Shiffrin1973] In each trial the number of items presented during
encoding was clearly well beyond the 4 6 1 items stipulatedby classical STM models [Cowan 2001] While memories ofsome materials (eg words spatial positions simple visualdisplays) are supported by limited-capacity buffers [Badde-ley and Hitch 1974] memory for other categories [egunknown faces and scenes see Shiffrin 1973] will tap intoLTM when the task requires the encoding of two or moreitems [Shallice and Cooper 2011] For this reason werestrict our discussion on the tripartite task-specific dissoci-ations with reference to highly complex naturalisticmaterial
Precuneus and Inferior Parietal Cortex Support
Temporal Order Recollection
The scene-chronology judgment task activated the pre-cuneus in both Exp 1 and 2 despite the considerablechanges in the trial structure for probing the retrieval oftemporal information (ie relative temporal order betweentwo probes in Exp 1 vs absolute temporal position of asingle probe in Exp 2) The chronology-judgment taskrequired retrieving the event sequences of many complex
Figure 5
Task-specific retrieval activation common in studies after imme-
diate delays and a longer delay Activation for the LTM study
[24 h Kwok et al 2012] is depicted in green and activation
for the current immediate retrieval study (data from current
Exp 1) depicted in red Activation clusters for both studies were
estimated and displayed at P-unc 5 0001
TABLE II Common memory retrieval activation
ImMem LTM common effects
Cluster Voxel
k P-corr Z x y z
Scene-chronology (Temporal)Precuneus B 112 0011 472 3 257 47Angular gyrus R 98 lt 0001 400 54 254 17
Scene-layout (Spatial)Superior parietal gyrus L 206 0001 595 221 269 56Supramarginal gyrus L 129 0008 346 260 236 44Intraparietal sulcus L 487 236 245 41Superior parietal gyrus R 651 lt 0001 520 24 263 59Dorsal occipital cortex R 372 30 275 29Intraparietal sulcus R 520 42 239 47Supramarginal gyrus R 482 57 233 53Superior frontal sulcus L 100 0022 482 224 3 56Superior frontal sulcus R 145 0004 481 30 0 53Inferior frontal gyrus R 95 0027 475 54 9 26Inferior occipital gyrus R 98 0024 515 54 257 210
Task-specific activation after immediate delays (current Exp 1ImMem) and a previous study that used a long delay betweenencoding and retrieval [24 h LTM Kwok et al 2012] Statisticalthreshold were set at P-FWE-corr 5 005 (cluster-level) consider-ing the whole brain as the volume of interest Note that theldquobetween-studiesrdquo overlap activation in the right angular gyruswas found at a lower uncorrected threshold (in italics) but impor-tantly within this area no differences or interactions betweenImMem and LTM studies were found (see differential effectsbetween the two studies in Supporting Information Table 2) LRB leftright hemispherebilateral
r Kwok et al r
r 2508 r
stimuli (scenes) that entail high-level conceptualhierarchi-cal relationships between them [Swallow et al 2011 Zackset al 2001] The precuneus was previously found to acti-vate in our LTM protocol that used the same temporalorder judgment [Kwok et al 2012 and see also results ofthe conjunction analyses] The current finding of precunealactivation using very different (much shorter) delays fitswell with the view that these processes are not strictlyassociated with retrieval following long retention delayssuggesting that this medial parietal region is implicatedduring temporal order retrieval irrespective of delay (Fig5A and Table II) Specifically we link the effect of chrono-logical retrieval in the precuneus with processes involvingan active reconstruction of past events [see also St Jacqueset al 2008] and possibly memory-related imagery[Fletcher et al 1995 Ishai et al 2000] These processeswould characterize temporal order judgments using com-plex naturalistic material but not order-judgment withsimpler stimulus material which previous studies haveassociated with other brain regions [eg middleprefron-tal areas Suzuki et al 2002]
Together with the precuneus temporal-related judgmentalso gave rise to activation of the right angular gyrus(AG) One possible interpretation relates the inferior parie-tal cortex to the engagement of attentional processes[Majerus et al 2007 2010] The scene-chronology judg-ment of Exp 2 provided us with a further qualifier to thiskind of temporal-related attention Although the Temporaltask in Exp 2 did not require any explicit temporal orderjudgment (only classifying a scene into either the first orsecond half of the movie) an accurate decision still neces-sitated an evaluation of the target scene with many othersegments of the movie This operation is fundamentallydifferent from the evaluation implicated in the scene-layout judgment which required decisions on comparingbetween a single scene and its own correspondentencoded snapshot The latter involves visuospatial analysisof the configuration within scenes The AG may subservedifferent types of attention among which one of them islinked with memory search operations [see Majerus et al2010 2006 also showing coactivation of these two regionsduring order retrieval of simple stimuli after short delays]
Superior Parietal Cortex Evaluates Spatial
Details Against Memory Representation
The second pattern consistently found across experi-ments (Exp 1 and Exp 2 see Fig 3) and across studieswith different retention delays (conjunction betweenimmediate retrieval and LTM studies see Fig 5B andTable II) was the activation of the superior parietal lobule(SPL) during the scene-layout judgment Previous studiesshowed sustained activation in the parietal cortex duringworking memory delays [Courtney et al 1998] and thatactivity there correlates with memory load [Todd andMaroi 2004] Here the amount of information contained
in the clips was equated in the three tasks especially inExp 2 where we counterbalanced the assignment of mov-ies across tasks The subjects also could not anticipatewhat specific type of information would be tested atretrieval and needed to keep rehearsing different aspectsof the clips Because of this we deem unlikely that theactivation of SPL that here was specific for the scene-layout tasks reflected general rehearsal processes [Magenet al 2009] andor updating operations [Bledowski et al2009 Leung et al 2007]
Instead we suggest a role for the SPL in mediating aninterface between the internal memory representation andexternal information It has been proposed that the supe-rior parietal cortex is engaged in allocating top-downattention to orient to aspects of the available visual inputand on information retrieved from memory [Ishai et al2002 Lepsien and Nobre 2007 Summerfield et al 2006]In line with this we found SPL activation when the sub-jects had to make decisions by comparing between theirmemory of the encoded clips and the scene stimuli pre-sented at retrieval This procedure might act on somevisuospatial storage which is held by the lateral occipitalcortex and mediated by the intraparietal sulcus (Fig 3B)The immediate memory task here was analogous to theSpatial task of our previous LTM study [Kwok et al2012] which also required matchingcomparing the avail-able external visual input and internal information storedin memory [see also Nobre et al 2004] This interfaceinvolves a range of top-down processes such as monitor-ing and verification [Cabeza et al 2008 Ciaramelli et al2008] which are independent of the probe imagersquos mne-monic status as shown by the equated effect sizes forldquosamerdquo and ldquoflippedrdquo images conditions (see signals plotsin Fig 3 and also ldquoRetrieval successrdquo analyses of Exp 2 inSupporting Information)
Recognition Processes in the Medial Frontal
Cortex and the Temporal Cortex
The third retrieval task involved scene-recognition judg-ments and was associated with the activation of the MFCand the MTG Moreover in Exp 1 we found a statisticaltrend in the hippocampus bilaterally an area that we pre-viously associated with scene-recognition judgment fromLTM (see next section about ldquoPossible correspondencesbetween retrieval from short- and long-term memoryrdquo)
The MFC was engaged both Exp 1 and 2 but was notactivated in the other two tasks thus a broad prefrontalcortexworking memory correspondence [Tsuchida andFellows 2009] seems insufficient to explain this task-specific effect The scene-recognition task in Exp 2 taxedboth recognition of ldquooldrdquo scenes and correct rejection ofldquonewrdquo scenes suggesting that the MFC here did not dis-tinguish between the correct rejection of new information(unseennew trials) and the recognition of old imagesOne candidate cognitive process that is common to both
r Neural Correlates of Immediate Episodic Retrieval r
r 2509 r
types of processes is feeling-of-knowing (FOK) FOKdepend on the cue specification content retrieval andappraisal of the retrieved memory fragments but are inde-pendent of the ultimate determination of content appropri-ateness [Schnyer et al 2005] Using imaging data duringsentence completion recognition Schnyer et al [2005]modeled a FOK-related directional path The path impli-cated the temporal cortex and the hippocampus andengaged the ventro-MFC at the end which is consistentwith our findings for the Object task implicating the MFCthe middle temporal cortex and at a lower statisticalthreshold the hippocampus
By contrast the activity in the MTG was larger forunseennew images than for seenold memory probes(Fig 4B) This suggests that new encoding may have con-tributed to the activation of the MTG during the retrievalphase of the Object task in Exp 1 The scene-recognitiontask required a lower precision in terms of retrievingdetails about the encoded material and could be solved bysimple recognition [Wixted and Squire 2010] Thus alter-natively larger activation for ldquonew vs oldrdquo stimuli mayrelate to some reduced activation associated with familiar-ity processes on old trials [see Henson et al 1999 whoreported reduced familiarity related activation in middletemporal gyri bilaterally and Henson and Rugg 2003 forreview] However we should point out that in Exp 2 theunseen images were extracted from edited-out parts of theoriginal clips and therefore contained the same objectcharacter depicted in the target images Therefore wedeem more likely that the relational association betweenobjects rather than their presenceabsence in the sceneshelped subjects to distinguish targets from foils We thussuggest that the activation of the MTG reflects processesutilizing the relational details among objects embedded inthe probe images for reaching the recognition decision[Diana et al 2007 Finke et al 2008 Hannula et al 2006Olson et al 2006] Future studies should consider theinclusion of some correlative behavioral measures to indexfamiliarity processes to elucidate further this issue More-over while the Temporal and Spatial tasks required mak-ing relative comparisons about aspects of the encodedvideo-clip the scene-recognition judgment merely requiredjudging whether the image presented at retrieval was pres-ent (or not) within the clip This may entail different proc-essing dynamics in the Object task compared with theother two tasks (cf also differences in retrieval times) Thelatter could be addressed using more subtle experimentalmanipulations for the scene-recognition judgment forexample by using digitally-modified foil images where asingle object in the scene has been modifiedremoved
Possible Correspondences Between Retrieval
from Short- and Long-Term Memory
The overlap between our current study and the previousLTM study using analogous tasks and materials lends
weight to the notion of scale-invariance in memory mecha-nisms [Brown et al 2007] as have been previously dem-onstrated in free recall tasks where the rate of item recallwas unvarying across recall span (day week year) andeven across from the past to the prospective future [May-lor et al 2001] Those previous behavioral findings implythat scale-invariance is a form of self-similarity in that theholistic pattern can exist at multiple levels of magnification[and even across different memory systems eg Maylor2002] Here our data affirmed the importance of the poste-rior parietal cortex in both immediate and long-termretrieval [Berryhill et al 2011 2007] and further character-ized its involvement in dealing with the spatial relationsof objects within scenes [Bricolo et al 2000 Buiatti et al2011] Our data also demonstrated such short-long-termcorrespondence exists in the precuneusangular gyrus fortemporal order retrieval (cf scene-chronology judgmenttask) In contrast the bilateral hippocampus and MFC acti-vations did not reach full statistical significance in theImMem LTM conjunction analysis At first sight thismay indicate that the scene-recognition task tapped intodistinct recognition-based mechanisms as a function ofretention delay However we deem this unlikely becausethe direct comparison between the two datasets did notshow any significant effect in these regions (see Support-ing Information Table 2) Indeed a more targeted test ask-ing the question of whether the hippocampus clusterspreviously found in LTM activated also in the currentimmediate retrieval experiments revealed significanteffects in both immediate retrieval experiments here (Exp1 x y z 5 224 29 225 P-FWE-corrlt 0001 Z 5 458 x yz 5 21 29 222 P-FWE-corr 5 0001 Z 5 451 and in Exp2 x y z 5 224 214 221 P-FWE-corr 5 0004 Z 5 396using the LTM clusters for small volume correction) Theseadditional tests point to a more general issue of sensitivitythat future studies may address using different methodo-logical approaches such as multivariate methods that canbe more sensitive than the standard univariate analysesused here Moreover the former would enable capturingtask-specific effects that are expressed as patterns of dis-tributed activation rather than local changes of activity asreported here [eg Jimura and Poldrack 2012]
Task Difficulty and Retrieval Success
We used several control analyses to show that the task-specific patterns of activation and the correspondencebetween the current data and our previous LTM study donot merely reflect some effect related to general task diffi-culty (see Supporting Information) First areas involved inoverall task difficulty should show progressively greateractivation with progressively more difficult tasks In Exp 1this would entail SgtTgtO However the patterns in theprecuneus and the angular gyrus (with TgtOgt S Fig 2A)and the MFC (Ogt SgtT Fig 4A) did not fit with simplepredictions based on overall task difficulty We also
r Kwok et al r
r 2510 r
assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r

now each trial comprised one single probe image Scene-chronology (Temporal) Subjects were required to recallthe probe imagersquos temporal location with respect to themidpoint of the clip by choosing between the ldquofirst halfrdquoor the ldquosecond halfrdquo of the clip Subjects were required topress the left button if the probe image happened in thefirst half or the right button if the image happened in thesecond half Scene-layout (Spatial) Subjects wereinstructed to focus on the spatial layout and to judgewhether the probe depicted a scene with a spatial arrange-ment as identical to the clip or it was a mirror image Anidentical spatial configuration required the subject to pressthe ldquocorrectrdquo button whereas a mirror image required apress to the ldquoincorrectrdquo button The mapping of ldquocorrectincorrectrdquo (leftright) layouts was counterbalanced acrosssubjects Scene-recognition (Object) Subjects wereinstructed to focus on the content of the probe image andto indicate whether the image had been shown in the clipIf it had been shown the subject responded with theldquopresentrdquo button otherwise with the ldquoabsentrdquo buttonAgain the leftright mapping of ldquopresentabsentrdquoresponses was counterbalanced between subjects
Since only a single image was now presented atretrieval the Temporal task could not involve any factorof ldquotemporal distancerdquo (cf TshortTlong in Exp 1) Insteadin Exp 2 we found that the behavioral performance wasaffected by the ldquotemporal positionrdquo of the probe imageduring encoding (easier if extracted from the startend ofthe clip harder if extracted from around the middle)Accordingly the temporal trials were now classified andmodeled separately in the fMRI analysis as Teasy and Thard
(see also Supporting Information Table 1)
fMRI Data Acquisition and Preprocessing
The functional images were acquired on a Siemens Alle-gra (Siemens Medical Systems Erlangen Germany) 3 Teslascanner equipped for echo-planar imaging (EPI) A quad-rature volume head coil was used for radio frequencytransmission and reception Head movement was mini-mized by mild restraint and cushioning Thirty-two slicesof functional MR images were acquired using blood oxy-genation level-dependent imaging (3 3 3 mm in-plane25 mm thick 50 distance factor repetition time 5 208 secho time 5 30 ms flip angle 5 70 FOV 5 192 mm acqui-sition order 5 continuous ascending) covering the wholecortex We acquired 1648 and 1488 fMRI volumes foreach subject in Exp 1 and 2 respectively Images wereacquired across 4 fMRI runs in both experiments The totalacquisition periods lasted for 55 and 49 min respectively
Data preprocessing was performed with SPM8 (wwwfilionuclacukspm) implemented on Matlab 74 (The Math-Works Natick MA) After having discarded the first 4volumes in each run images were realigned to correct forhead movements Slice-acquisition delays were correctedusing the middle slice as a reference Images were then
normalized to the MNI EPI template resampled to 3 mmisotropic voxel size and spatially smoothed using an iso-tropic Gaussian Kernel (FWHM 5 8 mm)
fMRI Analyses
Statistical inference was based on a random effectsapproach This comprised two steps first-level analysesestimating contrasts of interest for each subject followedby second-level analyses for statistical inference at thegroup-level [Penny and Holmes 2004] For each of thetwo experiments we used several different modelsdepending on the specific questions that we sought toaddress (see below and Supporting Information Table 1)
All the first-level subject-specific models included oneregressor modeling the presentation of the video clips irre-spective of task plus several regressors that modeled theretrieval phase according to the different conditions ofinterest (see leftmost column in Supporting InformationTable 1) The regressor modeling the ldquoencoding phaserdquo(Movie-regressor) was time-locked to the onset of eachvideo and had an event duration corresponding to theduration of each specific video (ie 772ndash1140 s) All theretrieval-related regressors were time-locked to the onsetof the probe image(s) and with a duration correspondingto the probe presentation at retrieval that is 4 s in Exp 1and 5 s in Exp 2 All regressors were convolved with theSPM8 canonical hemodynamic response function Timeseries were high-pass filtered at 128 s and prewhitened bymeans of autoregressive model AR(1) The sections belowdetail how the trials were assigned to the retrieval regres-sorsconditions in the different analyses
Experiment 1
For Exp 1 we used two different sets of first-level mod-els (see Supporting Information Table 1 leftmost column)The first set of models included the encoding-regressor(Movie) plus 5 retrieval-related conditions temporal-short(Tshort) temporal-long (Tlong) spatial (S) object (O) trialsand a condition modeling all the error trials irrespective oftask (Error) The trials belonging to the Temporal taskwere modeled with two separate regressors accounting forthe factor of temporal distance [Tshort vs Tlong cf Kwoket al 2012] All temporal trials including probes with atemporal distance shorter than 19 s were modeled asTshort while those longer than 19 s modeled as Tlong
The main second-level analysis consisted of a within-subjects Analysis of variance (ANOVA) modeling the foureffects of interest Tshort Tlong S and O conditions Foreach subject and each of the four conditions a contrastimage was computed in the first-level model by averagingthe corresponding parameter estimates across the 4 fMRIruns These subject-specific contrast images were enteredin the second-level ANOVA For the identification of task-specific retrieval effects we compared each task with themean of the two other tasks averaging the two Temporal
r Kwok et al r
r 2500 r
conditions ldquoT(short1long)2gt (S 1 O)2rdquo ldquoSgt (T(short1long)2 1 O)2rdquo and ldquoOgt (T(short1long)2 1 S)2rdquo Sphericity-correction was applied to all group-level ANOVAs toaccount for any nonindependent error term for repeatedmeasures and any difference in error variance across con-ditions [Friston et al 2002] The P-values were correctedfor multiple comparisons using a cluster-level threshold ofP-FWE-corr 5 005 considering the whole brain as the vol-ume of interest The cluster size was estimated using aninitial voxel-level threshold of P-unc 5 0001 To ensurethe specificity of any task effect the main differential con-trast was inclusively masked with 2 additional effectsThese corresponded to the activation for the critical taskvs each of the two other tasks for example the Temporalcontrast ldquoT(short1long)gt (S 1 O)rdquo was inclusively maskedwith ldquoT(short1long)2gt Srdquo and ldquoT(short1long)2gtOrdquo Forthese additional and nonindependent masking contraststhe threshold was set to P-unc 5 005 Together with themain contrasts that compared the three retrieval taskswithin this ANOVA we also tested for the effect ofldquotemporal distancerdquo (TshortgtTlong)
Using the same first-level models we ran a secondwithin-subject ANOVA that now also included the averageresponse times (RT) for each condition per subject as acovariate of no interest (Supporting Information Table 1ldquoControl for task difficultyrdquo) The aim of this was to reas-sess the pattern of task-specific dissociation after havingaccounted for any differences in task difficulty (Fig 1B)This RT-controlled analysis was not independent of themain analysis and the results are reported in the Support-ing Information
Experiment 2
The aim of Exp 2 was to account for the possible influ-ence of ldquounseennewrdquo images that were presented as foilsduring the retrieval phase of the Spatial and Object tasksin Exp 1 (eg incidental encoding during retrieval) InExp 2 a single probe image was presented during theretrieval phase of the trial (Fig 1A Experiment 2) Thisprocedure imposed a new constraint on the Temporaltask which now could not include the factor of ldquotemporaldistancerdquo Instead we taxed temporal retrieval by askingparticipants to judge whether the single probe image wasextracted from the first or second half of the clip Asexpected the participants were faster and more accurate tojudge probes presented either near the beginning or theend of the clip compared with probes extracted fromaround the middle of the clip Thus we now divided thetemporal trials into Teasy and Thard accounting for thisbehavioral effect
For Exp 2 we performed one single set of first-levelmodels and a single second-level group-analysis The first-level models included the encoding-regressor (Movie) and6 retrieval-related regressors Teasy (probe extracted fromthe 1st or 4th quartile) Thard (probe extracted from the2nd or 3rd quartile) Ssame Sflipped Oold and Onew plus
one regressor modeling all the error trials (see SupportingInformation Table 1)
The group level analysis consisted of a within-subjectsANOVA that included the 6 conditions of interest TeasyThard Ssame Sflipped Oold and Onew Within this ANOVAwe first tested for the task-specific effects by comparingeach task vs the mean of the other two tasks In this initialanalysis we averaged the Teasy and Thard conditions ofthe Temporal task as well as seen and unseen images ofthe Spatial and Object tasks for example[T(easy1hard)gt (S(same1flipped) 1 O(old1new))2] The statisticalthresholds used here were the same as Exp 1 statisticalmaps were corrected for multiple comparisons using acluster-level threshold of P-FWE-corr 5 005 (whole-brain)The cluster size was estimated using an initial voxel-levelthreshold of P-unc 5 0001 Again all comparisons wereinclusively masked with the effect of the main task vseach of the two other tasks (see Exp 1 above for details)
Within the same ANOVA a second set of contrasts soughtto replicate the task-specific effects but now using onlyseenold images (Supporting Information Table 1 ldquoSeen-onlyimagesrdquo) hence excluding any possible confound associatedwith the presentation of unseen images during retrieval Weused the following contrasts [T(easy1hard)gt (Ssame 1 Oold)][Ssamegt (T(easy1hard)2 1 Oold)2] and [Ooldgt (T(easy1hard)2 1 Ssame)2] The statistical thresholds and masking proce-dure were identical to Exp 1 and Exp 2 initial analysis
In Exp 2 we also performed control tests to rule out thepossibility that our main findings of task-specific dissocia-tions merely reflected differences in task difficulty (Sup-porting Information Table 1 ldquoTask difficultyrdquo) This wasaddressed by comparing conditions that behaviorallyresulted in difficult vs easy retrieval which included thecorresponding ldquohardrdquo versus ldquoeasyrdquo trials for the Tempo-ral task as well as by trials including ldquoflippedrdquo versusldquosamerdquo images in the Spatial task (Fig 1B) The results ofthese additional analyses are reported in the SupportingInformation
Conjunction analyses Between immediate
memory and LTM
To compare the current findings about task-specificretrieval after short delays with previous data using analo-gous retrieval tasks but a much longer retention delay[24 h Kwok et al 2012] we performed ldquobetween-studiesrdquo analyses Please note that the two paradigms dif-fered not only in terms of the retention delay (which dif-fered by a factor of 43200) but also in the presenceamount of interleaving events between encoding andretrieval In the LTM version participants were exposed to24 h of new episodic experiences following encodingwhich might cause retroactive interference on the alreadyacquired information [Wohldmann et al 2008] whereassuch retroactive interference was absent in the currentstudy on immediate retrieval The video clips were alsoshorter by a factor of about 250 compared to the 42-min
r Neural Correlates of Immediate Episodic Retrieval r
r 2501 r
episode used in the LTM paradigm Because of these differ-ences our analyses focused on commonalities rather thandifferences between LTM and immediate retrieval Theseanalyses considered the current Exp 1 which included thesame retrieval tasks as the LTM experiment Separately foreach of the three tasks we computed the relevant contrasts(eg Tgt (S1O)2) in the two studies and entered these inthree separate two samples t-tests For the identification ofcommon activation across studies we performed null-conjunction analyses (eg [Tgt (S1O)2]LTM [Tgt (S1O)2]ImMem) Statistical threshold were set at P-FWE-corr 5 005 cluster-level corrected at the whole brain level(cluster size estimated and displayed at P-unc 5 0001) con-sistent with all the analyses above For completeness thecontrasts testing for any differences between studies arereported in Supporting Information Table 2
Additional control analyses Retrieval success
Finally we asked to what extent any task-specific effectsidentified in our main analyses depended on the success-ful retrieval of stored information in the clips For this wecarried out additional analyses testing for retrieval per-formance in Exp 1 (ldquoTask 3 Accuracyrdquo in T and S tasks)and for accurate performance on seenold vs unseennewstimuli in Exp 2 (ldquoTask 3 Seen-unseenrdquo in S and O tasks)Please see Supporting Information for details of these anal-yses and the corresponding results
RESULTS
Behavioral Results
Experiment 1
Subjects performed significantly above chance in allthree tasks (all Plt 0001) The percentage of correct accu-racy and response times (RT) were analyzed usingrepeated measures ANOVAs with three levels correspond-ing to the TSO tasks This showed a main effect of con-ditions both for accuracy [F(2 32) 5 8142 Plt 0001] andRT [F(232) 5 3123 Plt 0001] Post hoc t-tests revealedthat subjects performed significantly better and faster inthe Object than Spatial task [Accuracy t(16) 5 1311Plt 0001 RT t(16) 5 652 Plt 0001] and Object than Tem-poral task [Accuracy t(16) 5 979 Plt 0001 RT t(16) 5
671 Plt 0001] Subjects were also more accurate on Tem-poral trials than on Spatial trials [Accuracy t(16) 5 348Plt 0001 but no difference in RT Pgt 01] For the Tempo-ral task we also examined the effect of temporal distanceexpecting that the retrieval would be more difficult whenthe two probes were temporally close (Tshortlt 19 s) com-pared with temporally far (Tlonggt 19 s) during encodingIndeed t-tests showed that subjects responded less accu-rately and slower in short than in long distance trials[Accuracy t(16) 5 391 Plt 0005 RT t(16) 5 315 Plt 001](Fig 1B plot on the left)
Experiment 2
In Exp 2 subjects also performed better than chance inall three tasks (all Plt 0001) Repeated measures ANOVAsshowed main effects of tasks both for accuracy [F(232) 5 919 Plt 0005] and RT [F(232) 5 1766 Plt 0001]Post hoc t-tests revealed that subjects performed withhigher accuracy and shorter RT in the Object comparedwith the Spatial task [Accuracy t(16) 5 406 Plt 0001 RTt(16) 5 532 Plt 0001] and in the Object compared withthe Temporal task [Accuracy t(16) 5 277 Plt 005 RTt(16) 5 383 Plt 0005] The difference in accuracy betweenthe Temporal and Spatial tasks approached significance(Plt 009) while there was no RT difference between Tem-poral and Spatial tasks (Pgt 01) Additional t-tests directlycompared different trial types within each task For theTemporal task subjects performed more accurately andfaster in the ldquoeasyrdquo than ldquohardrdquo trials [Accuracyt(16) 5 657 Plt 0001 RT t(16) 5 527 Plt 0001)] For theSpatial task they were more accurate in recognizingldquosamerdquo than correctly rejecting ldquoflippedrdquo images [Accu-racy t(16) 5 534 Plt 0001 RT Pgt 01] For the Objecttask subjects were faster to respond to unseen Onew
images compared to seen Oold images [RT t(16) 5 294Plt 001] without any difference in accuracy (Pgt 01) (Fig1B plot on the right)
Functional Neuroimaging Task-Specific Retrieval
In both Exp 1 and 2 we assessed brain activity specificto each of the three retrieval tasks (Temporal Spatial andObject) by comparing each task versus the mean of theother two (ldquoTask specificityrdquo contrasts see SupportingInformation Table 1) In Exp 2 we also carried out addi-tional nonindependent control tests using only trials con-taining seenold images (ldquoSeen-only imagesrdquo contrasts)The latter served to ensure that any task-specific dissocia-tion did not arise from the presentation of unseennewimages during the retrieval phase of the Spatial and Objecttasks (ie flipped- and new-image foils)
Scene-chronology judgment (Temporal task)
Exp 1 The comparison of the Temporal task with theother two retrieval conditions revealed significant activa-tion in the right precuneus bilateral caudate and bilateralcuneus (see Table I and Fig 2A clusters in bluemagenta) The signal plots in Figure 2 (panels on the left)show that the precuneus and the right angular gyrus wereactivated more by the Temporal task (average of Tshort andTlong) than by the Spatial and Object tasks Note that theeffect in the right angular gyrus was detected only atuncorrected threshold (P-unclt 0001) and it is reportedhere because it was fully significant in Exp 2 (see below)and as well as in the LTM dataset [Kwok et al 2012]
Together with the main comparison for the Temporaltask in Exp 1 we assessed the effect of temporal distance
r Kwok et al r
r 2502 r
TA
BL
EI
Task
-sp
ecif
icre
trie
val
acti
vati
on
Ex
p
1E
xp
2
Ex
p
2S
een
-on
ly
Clu
ster
Vo
xel
Clu
ster
Vo
xel
Clu
ster
Vo
xel
KP
-co
rr
Zx
yz
kP
-co
rr
Zx
yz
KP
-co
rr
Zx
yz
Sce
ne-
chro
no
log
y(T
emp
ora
l)P
recu
neu
sB
258
lt00
01
71
06
254
47
2666
lt00
01
51
74
272
43
1835
lt00
01
40
06
270
41
Po
st
cin
gu
late
cort
exB
mdashmdash
mdashmdash
564
02
3027
535
62
3631
Cau
dat
eB
148
000
54
302
122
1514
mdashmdash
mdashmdash
mdashmdash
mdashmdash
Cu
neu
sL
820
052
522
215
260
17mdash
mdashmdash
mdashmdash
mdashmdash
mdashC
un
eus
R88
004
25
2118
257
20mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nto
po
lar
cort
exR
mdashmdash
mdashmdash
874
lt0
001
547
2860
970
60
001
575
2860
7S
up
erio
rfr
on
tal
cort
exR
mdashmdash
mdashmdash
685
000
15
3526
2455
889
000
15
6026
2853
An
gu
lar
gy
rus
R43
lt00
01
46
345
269
29
457
00
07
48
444
266
45
509
00
04
45
844
268
43
Sce
ne-
lay
ou
t(S
pat
ial)
Su
peri
or
pari
eta
lg
yru
sL
1142
lt00
01
69
02
21
266
59
2530
lt00
01
66
72
18
268
59
377
00
16
49
92
18
266
59
Do
rsal
occ
ipit
alco
rtex
L5
112
272
759
521
228
280
9mdash
mdashmdash
mdashS
up
ram
arg
inal
gy
rus
L6
212
602
3338
mdashmdash
mdashmdash
mdashmdash
Intr
apar
ieta
lsu
lcu
sL
577
242
239
415
562
362
4445
mdashmdash
mdashmdash
Su
peri
or
pari
eta
lg
yru
sR
1314
lt00
01
67
221
263
59
3026
lt00
01
75
516
266
59
1262
lt00
01
47
426
268
47
Do
rsal
occ
ipit
alco
rtex
R4
1230
293
237
4034
278
294
1030
274
39In
trap
arie
tal
sulc
us
R6
2536
239
4433
40
026
548
402
4045
mdashmdash
mdashmdash
Su
pra
mar
gin
alg
yru
sR
597
602
2435
mdashmdash
mdashmdash
mdashmdash
Su
per
ior
fro
nta
lsu
lcu
sL
213
000
16
452
243
6223
80
078
543
226
22
51mdash
mdashmdash
mdashS
up
erio
rfr
on
tal
sulc
us
R18
20
002
592
302
356
220
009
74
7028
053
mdashmdash
mdashmdash
Infe
rio
rfr
on
tal
gy
rus
L92
003
65
002
516
26mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nta
lg
yru
sR
304
lt00
01
64
054
12
11
536
00
03
60
752
12
25
238
00
78
48
050
12
25
Infe
rio
ro
ccip
ital
gy
rus
L152
00
05
48
12
57
260
210
894
lt00
01
60
32
52
264
211
591
00
02
50
32
50
266
211
Infe
rio
ro
ccip
ital
gy
rus
R186
00
02
67
754
260
213
2277
lt00
01
68
256
258
211
1161
lt00
01
48
750
258
217
Occ
ipit
alp
ole
B21
80
001
484
29
293
2mdash
mdashmdash
mdashmdash
mdashmdash
mdashS
cen
e-re
cog
nit
ion
(Ob
ject
)M
ed
ial
fro
nta
lco
rtex
B478
lt00
01
51
92
351
47
2215
lt00
01
59
62
856
39
314
00
32
46
42
10
56
39
Mid
dle
tem
po
ral
gy
rus
L490
lt00
01
59
42
60
212
225
1661
lt00
01
67
72
62
214
219
162
lt00
01
42
92
60
24
225
Mid
dle
tem
po
ral
gy
rus
R304
lt00
01
61
560
212
216
1017
lt00
01
56
960
26
223
15
lt00
02
29
662
24
223
Orb
ito
-fro
nta
lco
rtex
B95
001
45
040
512
2222
70
049
506
044
219
mdashmdash
mdashmdash
An
gu
lar
gy
rus
L22
70
001
530
251
254
29mdash
mdashmdash
mdashmdash
mdashmdash
mdashA
ng
ula
rg
yru
sR
142
000
64
9857
260
38mdash
mdashmdash
mdashmdash
mdashmdash
mdashR
SC
p
ost
ci
ng
ula
teco
rtex
B47
4lt
000
14
903
242
35mdash
mdashmdash
mdashmdash
mdashmdash
mdash
T-c
on
tras
tsco
mp
ared
each
con
dit
ion
ver
sus
the
mea
no
fth
eo
ther
two
con
dit
ion
s(e
g
T(s
ho
rt1
lon
g)
2gt
(S1
O)
2)
and
was
incl
usi
vel
ym
ask
edw
ith
the
acti
vat
ion
for
the
crit
ical
con
dit
ion
ver
sus
each
of
the
two
oth
erco
nd
itio
ns
(eg
T
(sh
ort
1lo
ng
)2gt
S2
and
T(s
ho
rt1
lon
g)
2gt
O
2)
InE
xp2
we
con
du
cted
two
sep
arat
ese
tso
fco
ntr
asts
atth
eg
rou
p-l
evel
F
irst
t-
con
tras
tsw
ere
con
du
cted
usi
ng
all
six
con
dit
ion
sto
com
par
eea
chco
nd
itio
nv
ersu
sth
em
ean
of
the
oth
ertw
oco
nd
itio
ns
(eg
S (
sam
e1
flip
ped
)gt
[T(e
asy
1h
ard
)1
O(o
ld1
new
)]
2)
see
colu
mn
sin
the
mid
dle
Se
con
d
for
Spat
ial
and
Ob
ject
con
dit
ion
sw
ew
ere
inte
rest
edin
acti
vit
yel
icit
edb
yth
ere
spo
nse
sas
soci
ated
wit
hldquos
een
rdquotr
ials
on
ly(e
g
S sa-
megt
[T(e
asy
1h
ard
)2
1O
old
]2)
se
eri
gh
tmo
stco
lum
ns
and
also
Sup
po
rtin
gIn
form
atio
nT
able
1B
rain
reg
ion
sh
igh
lig
hte
din
bo
ldco
rres
po
nd
toth
eta
sk-s
pec
ific
effe
cts
con
sist
entl
yo
bse
rved
acro
ssal
lex
per
imen
ts
anal
yse
sth
ese
reg
ion
sar
eal
sod
isp
lay
edin
Fig
ure
s2ndash
4P
-val
ues
are
corr
ecte
dfo
rm
ult
iple
com
par
iso
ns
exce
pt
for
the
rig
ht
ang
ula
rg
yru
sin
Exp
1an
dth
em
idd
lete
mp
ora
lg
yri
inE
xp2
Seen
-on
lyth
atar
ere
po
rted
ata
low
eru
nco
rrec
ted
thre
sho
ld(i
nit
alic
s)
Po
st
po
ster
ior
xy
zco
ord
inat
esin
the
stan
dar
dM
NI
spac
eo
fth
eac
tiv
atio
np
eak
sin
the
clu
ster
sk
nu
mb
ero
fv
oxe
lsin
each
clu
ster
R
SC
retr
osp
len
ial
cort
ex
L
R
B
left
ri
gh
th
emis
ph
ere
bil
ater
al
r Neural Correlates of Immediate Episodic Retrieval r
r 2503 r
by comparing Tshort vs Tlong trials In previous studiesusing LTM protocols we found that temporal distance fur-ther modulated activity in the precuneus [Kwok et al2012 2014 see also St Jacques et al 2008] Here this con-trast did not reveal any significant effect This is in con-trast to the significant behavioral cost of retrieving Tshort
vs Tlong temporal distances (see behavioral results andFig 1B plot on the left) that was analogous to our previ-ous LTM protocol Similar behavioral patterns have beenobserved in many other short-term studies [Hacker 1980Konishi et al 2002 Milner et al 1991 Muter 1979] yet toour knowledge there does not seem to exists a clear con-
sensus on the neural correlates subserving the temporaldistance effect separating two temporally very nearbyeventsitems
Exp 2 In Exp 2 we replicated the effect of the temporalretrieval task in the precuneus and found a fully signifi-cant effect in the right angular gyrus (see Fig 2 clusters inredmagenta) In addition in Exp 2 there was a significantactivation of the right superior frontal cortex and the rightinferior frontopolar cortex that were not found in Exp 1(Table I) The involvement of the precuneus and the rightangular gyrus was further confirmed using only trials
Figure 2
Clusters of brain activation and signal plots for the scene-
chronology judgment (Temporal task) Clusters of activation and
signal plots for (A) the precuneus and (B) the right angular
gyrus These regions activated selectively during the temporal
tasks in Exp 1 (in blue) Exp 2 (in red overlap between Exp 1
and 2 shown in magenta) The activation selective for the Tem-
poral task was computed with the contrast T(short1long)
2gt (S 1 O)2 inclusively masked with T(short1long)2gt S2 and
with T(short1long)2gtO2 The involvement of the precuneus and
the right angular gyrus was further confirmed using only ldquoseen
oldrdquo images (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 40 voxels Effect sizes are mean adjusted (sum to
zero) and expressed in arbitrary units (au 6 90 CI) LR left
right PreC precuneus AnG angular gyrus
r Kwok et al r
r 2504 r
including seen images (cf ldquoExp 2 Seen- onlyrdquo Table I)The signal plots on Figure 2 (panels on the right) show theactivity of the precuneus and the right angular gyrus forall the conditions of Exp 2 Both in the precuneus and theright angular gyrus the average activity in the temporaltask [T(easy1hard)2] were larger than the Ssame and theOold conditions Notably Exp 2 included the presentationof a single image and that the participants were asked tojudge the absolute temporal position of the probe ratherthan the relative temporal order Despite this importantchange of how temporal memory was probed the findingof analogous patterns of activation across both experi-ments highlights the link between immediate retrieval oftemporal information and activation of the precuneus andangular gyrus
Scene-layout judgment (Spatial task)
Exp 1 The contrast concerning the retrieval of the spatiallayout revealed activation in the superior parietal cortexbilaterally with the cluster extending to the intraparietalsulcus and the supramarginal gyrus This contrast alsoshowed activation of the superior frontal sulci the inferiorfrontal gyri plus the inferior occipital cortex (see Table Iand Fig 3 clusters displayed in bluemagenta signalsplots on the left side)
Exp 2 The same comparison in Exp 2 replicated theresults of Exp 1 showing activation in the superior parie-tal cortex bilaterally the superior frontal sulci the inferiorand dorsal occipital cortices and the right inferior frontalgyrus (Fig 3 clusters in redmagenta) When we consid-ered only the conditions including the seenold images(ldquoExp 2 Seen- onlyrdquo) we replicated the effects in the supe-rior parietal gyri the inferior occipital gyri and the rightinferior frontal gyrus (see Table I and Fig 3 signal plotson the right) The activation in the intraparietal sulci andthe superior frontal sulci did not reach statistical signifi-cance Inspection of the signal plots indicated that in theseareas there was less activation in the Ssame than Sflipped tri-als (cf bar 3 vs bar 4 signal plots of the superior fontalsulcus Fig 3A plot on the right) Nonetheless the activityin the Ssame condition was larger than those in the T andO conditions (cf bar 3 vs bars 1ndash2 and 5ndash6)
Scene-recognition judgment (Object task)
Exp 1 The Object task was associated with the activationof the medial frontal cortex (MFC) and the orbitofrontalcortex (but the latter did not fully replicate in Exp 2 seeTable I and Fig 4 clusters bluemagenta and signal plotson the left) together with bilateral activation in the middletemporal gyri the retrosplenialposterior cingulate gyrusand a portion of the angular gyrus The same contrast alsorevealed an activation of the hippocampus bilaterally Theactivation of the hippocampus was just below statisticalsignificance (P-FWE-corr 5 0052 Z 5 458 at x y z 5 2126 219 and P-FWE-corr 5 0077 Z 5 458 at x y z 5 224
29 225 considering the whole brain as the volume ofinterest) and it is reported here because the same effectwas previously found in the LTM dataset [Kwok et al2012 see also Discussion Section]
Exp 2 An almost identical pattern of activation wasfound in Exp 2 when we considered all the conditionsO(old1new)gt (T(easy1hard) 1 S(same1flipped))2 Significanteffects were found in the middle temporal gyri and alarge cluster including the medial frontal gyrus extendingventrally into the orbital-frontal cortex (Fig 4 clusters inredmagenta) However when we considered only the tri-als including seenold-images [Ooldgt (T(easy1hard)2 1 Ssame)2] the Object task was found to significantlyactivate only the MFC (Table I rightmost columns) indi-cating that this region was not affected by incidentalencoding
In summary across all three tasks the brain activationwas found to be similar across Exp 1 and Exp 2 Thescene-chronology Temporal task activated the precuneusand the right angular gyrus the scene-layout Spatial taskwas associated with activation of the posteriorsuperiorparietal cortex and the right inferior frontal gyrus whilethe scene-recognition Object task activated the MFC andthe middle temporal gurus (see Table I areas highlightedin bold) The control analysis of Exp 2 that consideredonly trials including seenold images confirmed thesetask-specific effects indicating that the pattern of dissocia-tionmdashand in particular for the Spatial and Object tasksmdashwas not merely driven by the unseennew images duringthe retrieval phase
Functional Neuroimaging Correspondence with
An Analogous LTM Dataset
We directly compared the task-specific dissociationobserved in the current dataset (Exp 1) with our previousLTM study that included the same task-specific retrievaltasks but after a much longer retention delay [ie 24 hbetween encoding and retrieval Kwok et al 2012] Com-mon across the two data sets the conjunction analysesrevealed an activation cluster in the precuneus for theTemporal task (Fig 5A and Table II) and the dorsal fron-toparietal network plus right inferior frontal gyrus andright occipital cortex for the Spatial task (Fig 5B and TableII) These formal tests show that the Temporal and Spatialtask-specific dissociation observed here for immediateretrieval matched well with the patterns observed duringretrieval from LTM By contrast for the Object task over-lapping effects in the MFC and in the hippocampus weredetected only at a more lenient uncorrected threshold ofP-uncorr 5 0001 In the hippocampus the lack of a fullysignificant ldquocommonrdquo effect across datasets was due to thefact that in the current Exp 1 the contrast comparing theObject task versus the two other tasks did not survive cor-rection for multiple comparisons considering the wholebrain as the volume of interest (P-corr 5 0052 and 0077
r Neural Correlates of Immediate Episodic Retrieval r
r 2505 r
Figure 3
Clusters of brain activation and signal plots for the Scene-layout
judgment (Spatial task) Clusters of activation and signal plots for
(A) the superior frontal cortex and the superior parietal cortex
bilaterally and (B) the right inferior occipital cortex and the right
inferior frontal cortex These regions activated for the Spatial task
in Exp 1 (blue) and in Exp 2 (red overlap between Exp 1 and Exp 2
in magenta) The activation selective for the Spatial task was com-
puted with the contrast Sgt [T(short1long)2 1 O]2 inclusively
masked with SgtT(short1long)2 and with SgtO The effects in the
superior parietal gyri the inferior occipital gyri and the right inferior
frontal gyrus were replicated when considering only sameold
images in Exp 2 (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 80 voxels Effect sizes are mean adjusted (sum to zero)
and are expressed in arbitrary units (au 6 90 CI) LR leftright
SFS superior frontal sulcus SPG superior parietal gyrus IFG infe-
rior frontal gyrus IOG inferior occipital gyrus
in the left and right hemispheres respectively cf above)Indeed the formal test assessing differences between thetwo datasets (ie ldquoTask 3 Experimentrdquo interactions seeSupporting Information Table 2) did not reveal any signifi-cant difference between LTM and immediate retrieval inthe hippocampus consistent with this region beinginvolved in the scene-recognition judgment irrespective ofthe retention delay (see also Discussion Section)
DISCUSSION
We have identified distinct sets of brain areas thatshowed task-specific activation during immediate retrievalfollowing the encoding of dynamic cinematic material Theprecuneus and the right angular gyrus were activatedwhile retrieving the temporal order of events (scene-chro-nology judgment) the superior parietal cortex wasrecruited during spatial related recall (scene-layout
Figure 4
Clusters of brain activation and signal plots for the Scene-
recognition judgment (Object task) Clusters of activation and
signal plots for (A) the MFC and (B) the MTG These regions
activated for the Object task in Exp 1 (blue) and in Exp 2 (red
overlap in magenta) The activation selective for the Object task
was computed with the contrast Ogt [T(short1long)2 1 S]2
inclusively masked with OgtT(short1long)2 and with Ogt S The
signal plots refer to the left hemisphere but analogous patterns
of activation were found in the right hemisphere (Table I) In
Exp 2 the task-specific effect of the Object task in the MFC
was replicated when considering only the sameold images
(Table I rightmost columns) Inspection of the signal plot for the
left MTG revealed less activity in the Oold than Onew trials (plots
on the right bars 5 and 6) although the Oold trials showed
numerically larger parameter estimates than the T and S condi-
tions (see p-uncorr reported in italics in Table I rightmost col-
umns) Activation clusters were estimated and displayed at P-
unc 5 0001 thresholded at a minimum of 80 voxels Effect sizes
are mean adjusted (sum to zero) and expressed in arbitrary
units (au 6 90 CI) LR leftright MTG middle temporal
gyrus MFC medial frontal cortex
r Neural Correlates of Immediate Episodic Retrieval r
r 2507 r
judgment) whereas activity in the MFC and the middletemporal gyrus (MTG) reflected processes associated withscene-recognition judgments These task-specific effectsoccurred independently of whether subjects were asked tojudge seenold or unseennew stimuli (cf ldquoSeen-onlyimagesrdquo contrasts in Exp 2) and independently ofretrieval success and general task difficulty (see Support-ing Information) With respect to the hypothesized corre-spondence between immediate retrieval and retrieval fromLTM we found that these task-specific dissociationsare independent of the informationrsquos mnemonic history(24 h vs 2 s retention ie differing by 43200 times inmagnitude)
One important qualifier for the following discussion isthat memory retrieval in all our tasks depended minimallyon memory buffers compared to other simpler itemsdis-plays that are typically used in ldquostandardrdquo short-termworking memory tasks [Shallice and Cooper 2011 Shiffrin1973] In each trial the number of items presented during
encoding was clearly well beyond the 4 6 1 items stipulatedby classical STM models [Cowan 2001] While memories ofsome materials (eg words spatial positions simple visualdisplays) are supported by limited-capacity buffers [Badde-ley and Hitch 1974] memory for other categories [egunknown faces and scenes see Shiffrin 1973] will tap intoLTM when the task requires the encoding of two or moreitems [Shallice and Cooper 2011] For this reason werestrict our discussion on the tripartite task-specific dissoci-ations with reference to highly complex naturalisticmaterial
Precuneus and Inferior Parietal Cortex Support
Temporal Order Recollection
The scene-chronology judgment task activated the pre-cuneus in both Exp 1 and 2 despite the considerablechanges in the trial structure for probing the retrieval oftemporal information (ie relative temporal order betweentwo probes in Exp 1 vs absolute temporal position of asingle probe in Exp 2) The chronology-judgment taskrequired retrieving the event sequences of many complex
Figure 5
Task-specific retrieval activation common in studies after imme-
diate delays and a longer delay Activation for the LTM study
[24 h Kwok et al 2012] is depicted in green and activation
for the current immediate retrieval study (data from current
Exp 1) depicted in red Activation clusters for both studies were
estimated and displayed at P-unc 5 0001
TABLE II Common memory retrieval activation
ImMem LTM common effects
Cluster Voxel
k P-corr Z x y z
Scene-chronology (Temporal)Precuneus B 112 0011 472 3 257 47Angular gyrus R 98 lt 0001 400 54 254 17
Scene-layout (Spatial)Superior parietal gyrus L 206 0001 595 221 269 56Supramarginal gyrus L 129 0008 346 260 236 44Intraparietal sulcus L 487 236 245 41Superior parietal gyrus R 651 lt 0001 520 24 263 59Dorsal occipital cortex R 372 30 275 29Intraparietal sulcus R 520 42 239 47Supramarginal gyrus R 482 57 233 53Superior frontal sulcus L 100 0022 482 224 3 56Superior frontal sulcus R 145 0004 481 30 0 53Inferior frontal gyrus R 95 0027 475 54 9 26Inferior occipital gyrus R 98 0024 515 54 257 210
Task-specific activation after immediate delays (current Exp 1ImMem) and a previous study that used a long delay betweenencoding and retrieval [24 h LTM Kwok et al 2012] Statisticalthreshold were set at P-FWE-corr 5 005 (cluster-level) consider-ing the whole brain as the volume of interest Note that theldquobetween-studiesrdquo overlap activation in the right angular gyruswas found at a lower uncorrected threshold (in italics) but impor-tantly within this area no differences or interactions betweenImMem and LTM studies were found (see differential effectsbetween the two studies in Supporting Information Table 2) LRB leftright hemispherebilateral
r Kwok et al r
r 2508 r
stimuli (scenes) that entail high-level conceptualhierarchi-cal relationships between them [Swallow et al 2011 Zackset al 2001] The precuneus was previously found to acti-vate in our LTM protocol that used the same temporalorder judgment [Kwok et al 2012 and see also results ofthe conjunction analyses] The current finding of precunealactivation using very different (much shorter) delays fitswell with the view that these processes are not strictlyassociated with retrieval following long retention delayssuggesting that this medial parietal region is implicatedduring temporal order retrieval irrespective of delay (Fig5A and Table II) Specifically we link the effect of chrono-logical retrieval in the precuneus with processes involvingan active reconstruction of past events [see also St Jacqueset al 2008] and possibly memory-related imagery[Fletcher et al 1995 Ishai et al 2000] These processeswould characterize temporal order judgments using com-plex naturalistic material but not order-judgment withsimpler stimulus material which previous studies haveassociated with other brain regions [eg middleprefron-tal areas Suzuki et al 2002]
Together with the precuneus temporal-related judgmentalso gave rise to activation of the right angular gyrus(AG) One possible interpretation relates the inferior parie-tal cortex to the engagement of attentional processes[Majerus et al 2007 2010] The scene-chronology judg-ment of Exp 2 provided us with a further qualifier to thiskind of temporal-related attention Although the Temporaltask in Exp 2 did not require any explicit temporal orderjudgment (only classifying a scene into either the first orsecond half of the movie) an accurate decision still neces-sitated an evaluation of the target scene with many othersegments of the movie This operation is fundamentallydifferent from the evaluation implicated in the scene-layout judgment which required decisions on comparingbetween a single scene and its own correspondentencoded snapshot The latter involves visuospatial analysisof the configuration within scenes The AG may subservedifferent types of attention among which one of them islinked with memory search operations [see Majerus et al2010 2006 also showing coactivation of these two regionsduring order retrieval of simple stimuli after short delays]
Superior Parietal Cortex Evaluates Spatial
Details Against Memory Representation
The second pattern consistently found across experi-ments (Exp 1 and Exp 2 see Fig 3) and across studieswith different retention delays (conjunction betweenimmediate retrieval and LTM studies see Fig 5B andTable II) was the activation of the superior parietal lobule(SPL) during the scene-layout judgment Previous studiesshowed sustained activation in the parietal cortex duringworking memory delays [Courtney et al 1998] and thatactivity there correlates with memory load [Todd andMaroi 2004] Here the amount of information contained
in the clips was equated in the three tasks especially inExp 2 where we counterbalanced the assignment of mov-ies across tasks The subjects also could not anticipatewhat specific type of information would be tested atretrieval and needed to keep rehearsing different aspectsof the clips Because of this we deem unlikely that theactivation of SPL that here was specific for the scene-layout tasks reflected general rehearsal processes [Magenet al 2009] andor updating operations [Bledowski et al2009 Leung et al 2007]
Instead we suggest a role for the SPL in mediating aninterface between the internal memory representation andexternal information It has been proposed that the supe-rior parietal cortex is engaged in allocating top-downattention to orient to aspects of the available visual inputand on information retrieved from memory [Ishai et al2002 Lepsien and Nobre 2007 Summerfield et al 2006]In line with this we found SPL activation when the sub-jects had to make decisions by comparing between theirmemory of the encoded clips and the scene stimuli pre-sented at retrieval This procedure might act on somevisuospatial storage which is held by the lateral occipitalcortex and mediated by the intraparietal sulcus (Fig 3B)The immediate memory task here was analogous to theSpatial task of our previous LTM study [Kwok et al2012] which also required matchingcomparing the avail-able external visual input and internal information storedin memory [see also Nobre et al 2004] This interfaceinvolves a range of top-down processes such as monitor-ing and verification [Cabeza et al 2008 Ciaramelli et al2008] which are independent of the probe imagersquos mne-monic status as shown by the equated effect sizes forldquosamerdquo and ldquoflippedrdquo images conditions (see signals plotsin Fig 3 and also ldquoRetrieval successrdquo analyses of Exp 2 inSupporting Information)
Recognition Processes in the Medial Frontal
Cortex and the Temporal Cortex
The third retrieval task involved scene-recognition judg-ments and was associated with the activation of the MFCand the MTG Moreover in Exp 1 we found a statisticaltrend in the hippocampus bilaterally an area that we pre-viously associated with scene-recognition judgment fromLTM (see next section about ldquoPossible correspondencesbetween retrieval from short- and long-term memoryrdquo)
The MFC was engaged both Exp 1 and 2 but was notactivated in the other two tasks thus a broad prefrontalcortexworking memory correspondence [Tsuchida andFellows 2009] seems insufficient to explain this task-specific effect The scene-recognition task in Exp 2 taxedboth recognition of ldquooldrdquo scenes and correct rejection ofldquonewrdquo scenes suggesting that the MFC here did not dis-tinguish between the correct rejection of new information(unseennew trials) and the recognition of old imagesOne candidate cognitive process that is common to both
r Neural Correlates of Immediate Episodic Retrieval r
r 2509 r
types of processes is feeling-of-knowing (FOK) FOKdepend on the cue specification content retrieval andappraisal of the retrieved memory fragments but are inde-pendent of the ultimate determination of content appropri-ateness [Schnyer et al 2005] Using imaging data duringsentence completion recognition Schnyer et al [2005]modeled a FOK-related directional path The path impli-cated the temporal cortex and the hippocampus andengaged the ventro-MFC at the end which is consistentwith our findings for the Object task implicating the MFCthe middle temporal cortex and at a lower statisticalthreshold the hippocampus
By contrast the activity in the MTG was larger forunseennew images than for seenold memory probes(Fig 4B) This suggests that new encoding may have con-tributed to the activation of the MTG during the retrievalphase of the Object task in Exp 1 The scene-recognitiontask required a lower precision in terms of retrievingdetails about the encoded material and could be solved bysimple recognition [Wixted and Squire 2010] Thus alter-natively larger activation for ldquonew vs oldrdquo stimuli mayrelate to some reduced activation associated with familiar-ity processes on old trials [see Henson et al 1999 whoreported reduced familiarity related activation in middletemporal gyri bilaterally and Henson and Rugg 2003 forreview] However we should point out that in Exp 2 theunseen images were extracted from edited-out parts of theoriginal clips and therefore contained the same objectcharacter depicted in the target images Therefore wedeem more likely that the relational association betweenobjects rather than their presenceabsence in the sceneshelped subjects to distinguish targets from foils We thussuggest that the activation of the MTG reflects processesutilizing the relational details among objects embedded inthe probe images for reaching the recognition decision[Diana et al 2007 Finke et al 2008 Hannula et al 2006Olson et al 2006] Future studies should consider theinclusion of some correlative behavioral measures to indexfamiliarity processes to elucidate further this issue More-over while the Temporal and Spatial tasks required mak-ing relative comparisons about aspects of the encodedvideo-clip the scene-recognition judgment merely requiredjudging whether the image presented at retrieval was pres-ent (or not) within the clip This may entail different proc-essing dynamics in the Object task compared with theother two tasks (cf also differences in retrieval times) Thelatter could be addressed using more subtle experimentalmanipulations for the scene-recognition judgment forexample by using digitally-modified foil images where asingle object in the scene has been modifiedremoved
Possible Correspondences Between Retrieval
from Short- and Long-Term Memory
The overlap between our current study and the previousLTM study using analogous tasks and materials lends
weight to the notion of scale-invariance in memory mecha-nisms [Brown et al 2007] as have been previously dem-onstrated in free recall tasks where the rate of item recallwas unvarying across recall span (day week year) andeven across from the past to the prospective future [May-lor et al 2001] Those previous behavioral findings implythat scale-invariance is a form of self-similarity in that theholistic pattern can exist at multiple levels of magnification[and even across different memory systems eg Maylor2002] Here our data affirmed the importance of the poste-rior parietal cortex in both immediate and long-termretrieval [Berryhill et al 2011 2007] and further character-ized its involvement in dealing with the spatial relationsof objects within scenes [Bricolo et al 2000 Buiatti et al2011] Our data also demonstrated such short-long-termcorrespondence exists in the precuneusangular gyrus fortemporal order retrieval (cf scene-chronology judgmenttask) In contrast the bilateral hippocampus and MFC acti-vations did not reach full statistical significance in theImMem LTM conjunction analysis At first sight thismay indicate that the scene-recognition task tapped intodistinct recognition-based mechanisms as a function ofretention delay However we deem this unlikely becausethe direct comparison between the two datasets did notshow any significant effect in these regions (see Support-ing Information Table 2) Indeed a more targeted test ask-ing the question of whether the hippocampus clusterspreviously found in LTM activated also in the currentimmediate retrieval experiments revealed significanteffects in both immediate retrieval experiments here (Exp1 x y z 5 224 29 225 P-FWE-corrlt 0001 Z 5 458 x yz 5 21 29 222 P-FWE-corr 5 0001 Z 5 451 and in Exp2 x y z 5 224 214 221 P-FWE-corr 5 0004 Z 5 396using the LTM clusters for small volume correction) Theseadditional tests point to a more general issue of sensitivitythat future studies may address using different methodo-logical approaches such as multivariate methods that canbe more sensitive than the standard univariate analysesused here Moreover the former would enable capturingtask-specific effects that are expressed as patterns of dis-tributed activation rather than local changes of activity asreported here [eg Jimura and Poldrack 2012]
Task Difficulty and Retrieval Success
We used several control analyses to show that the task-specific patterns of activation and the correspondencebetween the current data and our previous LTM study donot merely reflect some effect related to general task diffi-culty (see Supporting Information) First areas involved inoverall task difficulty should show progressively greateractivation with progressively more difficult tasks In Exp 1this would entail SgtTgtO However the patterns in theprecuneus and the angular gyrus (with TgtOgt S Fig 2A)and the MFC (Ogt SgtT Fig 4A) did not fit with simplepredictions based on overall task difficulty We also
r Kwok et al r
r 2510 r
assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r

conditions ldquoT(short1long)2gt (S 1 O)2rdquo ldquoSgt (T(short1long)2 1 O)2rdquo and ldquoOgt (T(short1long)2 1 S)2rdquo Sphericity-correction was applied to all group-level ANOVAs toaccount for any nonindependent error term for repeatedmeasures and any difference in error variance across con-ditions [Friston et al 2002] The P-values were correctedfor multiple comparisons using a cluster-level threshold ofP-FWE-corr 5 005 considering the whole brain as the vol-ume of interest The cluster size was estimated using aninitial voxel-level threshold of P-unc 5 0001 To ensurethe specificity of any task effect the main differential con-trast was inclusively masked with 2 additional effectsThese corresponded to the activation for the critical taskvs each of the two other tasks for example the Temporalcontrast ldquoT(short1long)gt (S 1 O)rdquo was inclusively maskedwith ldquoT(short1long)2gt Srdquo and ldquoT(short1long)2gtOrdquo Forthese additional and nonindependent masking contraststhe threshold was set to P-unc 5 005 Together with themain contrasts that compared the three retrieval taskswithin this ANOVA we also tested for the effect ofldquotemporal distancerdquo (TshortgtTlong)
Using the same first-level models we ran a secondwithin-subject ANOVA that now also included the averageresponse times (RT) for each condition per subject as acovariate of no interest (Supporting Information Table 1ldquoControl for task difficultyrdquo) The aim of this was to reas-sess the pattern of task-specific dissociation after havingaccounted for any differences in task difficulty (Fig 1B)This RT-controlled analysis was not independent of themain analysis and the results are reported in the Support-ing Information
Experiment 2
The aim of Exp 2 was to account for the possible influ-ence of ldquounseennewrdquo images that were presented as foilsduring the retrieval phase of the Spatial and Object tasksin Exp 1 (eg incidental encoding during retrieval) InExp 2 a single probe image was presented during theretrieval phase of the trial (Fig 1A Experiment 2) Thisprocedure imposed a new constraint on the Temporaltask which now could not include the factor of ldquotemporaldistancerdquo Instead we taxed temporal retrieval by askingparticipants to judge whether the single probe image wasextracted from the first or second half of the clip Asexpected the participants were faster and more accurate tojudge probes presented either near the beginning or theend of the clip compared with probes extracted fromaround the middle of the clip Thus we now divided thetemporal trials into Teasy and Thard accounting for thisbehavioral effect
For Exp 2 we performed one single set of first-levelmodels and a single second-level group-analysis The first-level models included the encoding-regressor (Movie) and6 retrieval-related regressors Teasy (probe extracted fromthe 1st or 4th quartile) Thard (probe extracted from the2nd or 3rd quartile) Ssame Sflipped Oold and Onew plus
one regressor modeling all the error trials (see SupportingInformation Table 1)
The group level analysis consisted of a within-subjectsANOVA that included the 6 conditions of interest TeasyThard Ssame Sflipped Oold and Onew Within this ANOVAwe first tested for the task-specific effects by comparingeach task vs the mean of the other two tasks In this initialanalysis we averaged the Teasy and Thard conditions ofthe Temporal task as well as seen and unseen images ofthe Spatial and Object tasks for example[T(easy1hard)gt (S(same1flipped) 1 O(old1new))2] The statisticalthresholds used here were the same as Exp 1 statisticalmaps were corrected for multiple comparisons using acluster-level threshold of P-FWE-corr 5 005 (whole-brain)The cluster size was estimated using an initial voxel-levelthreshold of P-unc 5 0001 Again all comparisons wereinclusively masked with the effect of the main task vseach of the two other tasks (see Exp 1 above for details)
Within the same ANOVA a second set of contrasts soughtto replicate the task-specific effects but now using onlyseenold images (Supporting Information Table 1 ldquoSeen-onlyimagesrdquo) hence excluding any possible confound associatedwith the presentation of unseen images during retrieval Weused the following contrasts [T(easy1hard)gt (Ssame 1 Oold)][Ssamegt (T(easy1hard)2 1 Oold)2] and [Ooldgt (T(easy1hard)2 1 Ssame)2] The statistical thresholds and masking proce-dure were identical to Exp 1 and Exp 2 initial analysis
In Exp 2 we also performed control tests to rule out thepossibility that our main findings of task-specific dissocia-tions merely reflected differences in task difficulty (Sup-porting Information Table 1 ldquoTask difficultyrdquo) This wasaddressed by comparing conditions that behaviorallyresulted in difficult vs easy retrieval which included thecorresponding ldquohardrdquo versus ldquoeasyrdquo trials for the Tempo-ral task as well as by trials including ldquoflippedrdquo versusldquosamerdquo images in the Spatial task (Fig 1B) The results ofthese additional analyses are reported in the SupportingInformation
Conjunction analyses Between immediate
memory and LTM
To compare the current findings about task-specificretrieval after short delays with previous data using analo-gous retrieval tasks but a much longer retention delay[24 h Kwok et al 2012] we performed ldquobetween-studiesrdquo analyses Please note that the two paradigms dif-fered not only in terms of the retention delay (which dif-fered by a factor of 43200) but also in the presenceamount of interleaving events between encoding andretrieval In the LTM version participants were exposed to24 h of new episodic experiences following encodingwhich might cause retroactive interference on the alreadyacquired information [Wohldmann et al 2008] whereassuch retroactive interference was absent in the currentstudy on immediate retrieval The video clips were alsoshorter by a factor of about 250 compared to the 42-min
r Neural Correlates of Immediate Episodic Retrieval r
r 2501 r
episode used in the LTM paradigm Because of these differ-ences our analyses focused on commonalities rather thandifferences between LTM and immediate retrieval Theseanalyses considered the current Exp 1 which included thesame retrieval tasks as the LTM experiment Separately foreach of the three tasks we computed the relevant contrasts(eg Tgt (S1O)2) in the two studies and entered these inthree separate two samples t-tests For the identification ofcommon activation across studies we performed null-conjunction analyses (eg [Tgt (S1O)2]LTM [Tgt (S1O)2]ImMem) Statistical threshold were set at P-FWE-corr 5 005 cluster-level corrected at the whole brain level(cluster size estimated and displayed at P-unc 5 0001) con-sistent with all the analyses above For completeness thecontrasts testing for any differences between studies arereported in Supporting Information Table 2
Additional control analyses Retrieval success
Finally we asked to what extent any task-specific effectsidentified in our main analyses depended on the success-ful retrieval of stored information in the clips For this wecarried out additional analyses testing for retrieval per-formance in Exp 1 (ldquoTask 3 Accuracyrdquo in T and S tasks)and for accurate performance on seenold vs unseennewstimuli in Exp 2 (ldquoTask 3 Seen-unseenrdquo in S and O tasks)Please see Supporting Information for details of these anal-yses and the corresponding results
RESULTS
Behavioral Results
Experiment 1
Subjects performed significantly above chance in allthree tasks (all Plt 0001) The percentage of correct accu-racy and response times (RT) were analyzed usingrepeated measures ANOVAs with three levels correspond-ing to the TSO tasks This showed a main effect of con-ditions both for accuracy [F(2 32) 5 8142 Plt 0001] andRT [F(232) 5 3123 Plt 0001] Post hoc t-tests revealedthat subjects performed significantly better and faster inthe Object than Spatial task [Accuracy t(16) 5 1311Plt 0001 RT t(16) 5 652 Plt 0001] and Object than Tem-poral task [Accuracy t(16) 5 979 Plt 0001 RT t(16) 5
671 Plt 0001] Subjects were also more accurate on Tem-poral trials than on Spatial trials [Accuracy t(16) 5 348Plt 0001 but no difference in RT Pgt 01] For the Tempo-ral task we also examined the effect of temporal distanceexpecting that the retrieval would be more difficult whenthe two probes were temporally close (Tshortlt 19 s) com-pared with temporally far (Tlonggt 19 s) during encodingIndeed t-tests showed that subjects responded less accu-rately and slower in short than in long distance trials[Accuracy t(16) 5 391 Plt 0005 RT t(16) 5 315 Plt 001](Fig 1B plot on the left)
Experiment 2
In Exp 2 subjects also performed better than chance inall three tasks (all Plt 0001) Repeated measures ANOVAsshowed main effects of tasks both for accuracy [F(232) 5 919 Plt 0005] and RT [F(232) 5 1766 Plt 0001]Post hoc t-tests revealed that subjects performed withhigher accuracy and shorter RT in the Object comparedwith the Spatial task [Accuracy t(16) 5 406 Plt 0001 RTt(16) 5 532 Plt 0001] and in the Object compared withthe Temporal task [Accuracy t(16) 5 277 Plt 005 RTt(16) 5 383 Plt 0005] The difference in accuracy betweenthe Temporal and Spatial tasks approached significance(Plt 009) while there was no RT difference between Tem-poral and Spatial tasks (Pgt 01) Additional t-tests directlycompared different trial types within each task For theTemporal task subjects performed more accurately andfaster in the ldquoeasyrdquo than ldquohardrdquo trials [Accuracyt(16) 5 657 Plt 0001 RT t(16) 5 527 Plt 0001)] For theSpatial task they were more accurate in recognizingldquosamerdquo than correctly rejecting ldquoflippedrdquo images [Accu-racy t(16) 5 534 Plt 0001 RT Pgt 01] For the Objecttask subjects were faster to respond to unseen Onew
images compared to seen Oold images [RT t(16) 5 294Plt 001] without any difference in accuracy (Pgt 01) (Fig1B plot on the right)
Functional Neuroimaging Task-Specific Retrieval
In both Exp 1 and 2 we assessed brain activity specificto each of the three retrieval tasks (Temporal Spatial andObject) by comparing each task versus the mean of theother two (ldquoTask specificityrdquo contrasts see SupportingInformation Table 1) In Exp 2 we also carried out addi-tional nonindependent control tests using only trials con-taining seenold images (ldquoSeen-only imagesrdquo contrasts)The latter served to ensure that any task-specific dissocia-tion did not arise from the presentation of unseennewimages during the retrieval phase of the Spatial and Objecttasks (ie flipped- and new-image foils)
Scene-chronology judgment (Temporal task)
Exp 1 The comparison of the Temporal task with theother two retrieval conditions revealed significant activa-tion in the right precuneus bilateral caudate and bilateralcuneus (see Table I and Fig 2A clusters in bluemagenta) The signal plots in Figure 2 (panels on the left)show that the precuneus and the right angular gyrus wereactivated more by the Temporal task (average of Tshort andTlong) than by the Spatial and Object tasks Note that theeffect in the right angular gyrus was detected only atuncorrected threshold (P-unclt 0001) and it is reportedhere because it was fully significant in Exp 2 (see below)and as well as in the LTM dataset [Kwok et al 2012]
Together with the main comparison for the Temporaltask in Exp 1 we assessed the effect of temporal distance
r Kwok et al r
r 2502 r
TA
BL
EI
Task
-sp
ecif
icre
trie
val
acti
vati
on
Ex
p
1E
xp
2
Ex
p
2S
een
-on
ly
Clu
ster
Vo
xel
Clu
ster
Vo
xel
Clu
ster
Vo
xel
KP
-co
rr
Zx
yz
kP
-co
rr
Zx
yz
KP
-co
rr
Zx
yz
Sce
ne-
chro
no
log
y(T
emp
ora
l)P
recu
neu
sB
258
lt00
01
71
06
254
47
2666
lt00
01
51
74
272
43
1835
lt00
01
40
06
270
41
Po
st
cin
gu
late
cort
exB
mdashmdash
mdashmdash
564
02
3027
535
62
3631
Cau
dat
eB
148
000
54
302
122
1514
mdashmdash
mdashmdash
mdashmdash
mdashmdash
Cu
neu
sL
820
052
522
215
260
17mdash
mdashmdash
mdashmdash
mdashmdash
mdashC
un
eus
R88
004
25
2118
257
20mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nto
po
lar
cort
exR
mdashmdash
mdashmdash
874
lt0
001
547
2860
970
60
001
575
2860
7S
up
erio
rfr
on
tal
cort
exR
mdashmdash
mdashmdash
685
000
15
3526
2455
889
000
15
6026
2853
An
gu
lar
gy
rus
R43
lt00
01
46
345
269
29
457
00
07
48
444
266
45
509
00
04
45
844
268
43
Sce
ne-
lay
ou
t(S
pat
ial)
Su
peri
or
pari
eta
lg
yru
sL
1142
lt00
01
69
02
21
266
59
2530
lt00
01
66
72
18
268
59
377
00
16
49
92
18
266
59
Do
rsal
occ
ipit
alco
rtex
L5
112
272
759
521
228
280
9mdash
mdashmdash
mdashS
up
ram
arg
inal
gy
rus
L6
212
602
3338
mdashmdash
mdashmdash
mdashmdash
Intr
apar
ieta
lsu
lcu
sL
577
242
239
415
562
362
4445
mdashmdash
mdashmdash
Su
peri
or
pari
eta
lg
yru
sR
1314
lt00
01
67
221
263
59
3026
lt00
01
75
516
266
59
1262
lt00
01
47
426
268
47
Do
rsal
occ
ipit
alco
rtex
R4
1230
293
237
4034
278
294
1030
274
39In
trap
arie
tal
sulc
us
R6
2536
239
4433
40
026
548
402
4045
mdashmdash
mdashmdash
Su
pra
mar
gin
alg
yru
sR
597
602
2435
mdashmdash
mdashmdash
mdashmdash
Su
per
ior
fro
nta
lsu
lcu
sL
213
000
16
452
243
6223
80
078
543
226
22
51mdash
mdashmdash
mdashS
up
erio
rfr
on
tal
sulc
us
R18
20
002
592
302
356
220
009
74
7028
053
mdashmdash
mdashmdash
Infe
rio
rfr
on
tal
gy
rus
L92
003
65
002
516
26mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nta
lg
yru
sR
304
lt00
01
64
054
12
11
536
00
03
60
752
12
25
238
00
78
48
050
12
25
Infe
rio
ro
ccip
ital
gy
rus
L152
00
05
48
12
57
260
210
894
lt00
01
60
32
52
264
211
591
00
02
50
32
50
266
211
Infe
rio
ro
ccip
ital
gy
rus
R186
00
02
67
754
260
213
2277
lt00
01
68
256
258
211
1161
lt00
01
48
750
258
217
Occ
ipit
alp
ole
B21
80
001
484
29
293
2mdash
mdashmdash
mdashmdash
mdashmdash
mdashS
cen
e-re
cog
nit
ion
(Ob
ject
)M
ed
ial
fro
nta
lco
rtex
B478
lt00
01
51
92
351
47
2215
lt00
01
59
62
856
39
314
00
32
46
42
10
56
39
Mid
dle
tem
po
ral
gy
rus
L490
lt00
01
59
42
60
212
225
1661
lt00
01
67
72
62
214
219
162
lt00
01
42
92
60
24
225
Mid
dle
tem
po
ral
gy
rus
R304
lt00
01
61
560
212
216
1017
lt00
01
56
960
26
223
15
lt00
02
29
662
24
223
Orb
ito
-fro
nta
lco
rtex
B95
001
45
040
512
2222
70
049
506
044
219
mdashmdash
mdashmdash
An
gu
lar
gy
rus
L22
70
001
530
251
254
29mdash
mdashmdash
mdashmdash
mdashmdash
mdashA
ng
ula
rg
yru
sR
142
000
64
9857
260
38mdash
mdashmdash
mdashmdash
mdashmdash
mdashR
SC
p
ost
ci
ng
ula
teco
rtex
B47
4lt
000
14
903
242
35mdash
mdashmdash
mdashmdash
mdashmdash
mdash
T-c
on
tras
tsco
mp
ared
each
con
dit
ion
ver
sus
the
mea
no
fth
eo
ther
two
con
dit
ion
s(e
g
T(s
ho
rt1
lon
g)
2gt
(S1
O)
2)
and
was
incl
usi
vel
ym
ask
edw
ith
the
acti
vat
ion
for
the
crit
ical
con
dit
ion
ver
sus
each
of
the
two
oth
erco
nd
itio
ns
(eg
T
(sh
ort
1lo
ng
)2gt
S2
and
T(s
ho
rt1
lon
g)
2gt
O
2)
InE
xp2
we
con
du
cted
two
sep
arat
ese
tso
fco
ntr
asts
atth
eg
rou
p-l
evel
F
irst
t-
con
tras
tsw
ere
con
du
cted
usi
ng
all
six
con
dit
ion
sto
com
par
eea
chco
nd
itio
nv
ersu
sth
em
ean
of
the
oth
ertw
oco
nd
itio
ns
(eg
S (
sam
e1
flip
ped
)gt
[T(e
asy
1h
ard
)1
O(o
ld1
new
)]
2)
see
colu
mn
sin
the
mid
dle
Se
con
d
for
Spat
ial
and
Ob
ject
con
dit
ion
sw
ew
ere
inte
rest
edin
acti
vit
yel
icit
edb
yth
ere
spo
nse
sas
soci
ated
wit
hldquos
een
rdquotr
ials
on
ly(e
g
S sa-
megt
[T(e
asy
1h
ard
)2
1O
old
]2)
se
eri
gh
tmo
stco
lum
ns
and
also
Sup
po
rtin
gIn
form
atio
nT
able
1B
rain
reg
ion
sh
igh
lig
hte
din
bo
ldco
rres
po
nd
toth
eta
sk-s
pec
ific
effe
cts
con
sist
entl
yo
bse
rved
acro
ssal
lex
per
imen
ts
anal
yse
sth
ese
reg
ion
sar
eal
sod
isp
lay
edin
Fig
ure
s2ndash
4P
-val
ues
are
corr
ecte
dfo
rm
ult
iple
com
par
iso
ns
exce
pt
for
the
rig
ht
ang
ula
rg
yru
sin
Exp
1an
dth
em
idd
lete
mp
ora
lg
yri
inE
xp2
Seen
-on
lyth
atar
ere
po
rted
ata
low
eru
nco
rrec
ted
thre
sho
ld(i
nit
alic
s)
Po
st
po
ster
ior
xy
zco
ord
inat
esin
the
stan
dar
dM
NI
spac
eo
fth
eac
tiv
atio
np
eak
sin
the
clu
ster
sk
nu
mb
ero
fv
oxe
lsin
each
clu
ster
R
SC
retr
osp
len
ial
cort
ex
L
R
B
left
ri
gh
th
emis
ph
ere
bil
ater
al
r Neural Correlates of Immediate Episodic Retrieval r
r 2503 r
by comparing Tshort vs Tlong trials In previous studiesusing LTM protocols we found that temporal distance fur-ther modulated activity in the precuneus [Kwok et al2012 2014 see also St Jacques et al 2008] Here this con-trast did not reveal any significant effect This is in con-trast to the significant behavioral cost of retrieving Tshort
vs Tlong temporal distances (see behavioral results andFig 1B plot on the left) that was analogous to our previ-ous LTM protocol Similar behavioral patterns have beenobserved in many other short-term studies [Hacker 1980Konishi et al 2002 Milner et al 1991 Muter 1979] yet toour knowledge there does not seem to exists a clear con-
sensus on the neural correlates subserving the temporaldistance effect separating two temporally very nearbyeventsitems
Exp 2 In Exp 2 we replicated the effect of the temporalretrieval task in the precuneus and found a fully signifi-cant effect in the right angular gyrus (see Fig 2 clusters inredmagenta) In addition in Exp 2 there was a significantactivation of the right superior frontal cortex and the rightinferior frontopolar cortex that were not found in Exp 1(Table I) The involvement of the precuneus and the rightangular gyrus was further confirmed using only trials
Figure 2
Clusters of brain activation and signal plots for the scene-
chronology judgment (Temporal task) Clusters of activation and
signal plots for (A) the precuneus and (B) the right angular
gyrus These regions activated selectively during the temporal
tasks in Exp 1 (in blue) Exp 2 (in red overlap between Exp 1
and 2 shown in magenta) The activation selective for the Tem-
poral task was computed with the contrast T(short1long)
2gt (S 1 O)2 inclusively masked with T(short1long)2gt S2 and
with T(short1long)2gtO2 The involvement of the precuneus and
the right angular gyrus was further confirmed using only ldquoseen
oldrdquo images (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 40 voxels Effect sizes are mean adjusted (sum to
zero) and expressed in arbitrary units (au 6 90 CI) LR left
right PreC precuneus AnG angular gyrus
r Kwok et al r
r 2504 r
including seen images (cf ldquoExp 2 Seen- onlyrdquo Table I)The signal plots on Figure 2 (panels on the right) show theactivity of the precuneus and the right angular gyrus forall the conditions of Exp 2 Both in the precuneus and theright angular gyrus the average activity in the temporaltask [T(easy1hard)2] were larger than the Ssame and theOold conditions Notably Exp 2 included the presentationof a single image and that the participants were asked tojudge the absolute temporal position of the probe ratherthan the relative temporal order Despite this importantchange of how temporal memory was probed the findingof analogous patterns of activation across both experi-ments highlights the link between immediate retrieval oftemporal information and activation of the precuneus andangular gyrus
Scene-layout judgment (Spatial task)
Exp 1 The contrast concerning the retrieval of the spatiallayout revealed activation in the superior parietal cortexbilaterally with the cluster extending to the intraparietalsulcus and the supramarginal gyrus This contrast alsoshowed activation of the superior frontal sulci the inferiorfrontal gyri plus the inferior occipital cortex (see Table Iand Fig 3 clusters displayed in bluemagenta signalsplots on the left side)
Exp 2 The same comparison in Exp 2 replicated theresults of Exp 1 showing activation in the superior parie-tal cortex bilaterally the superior frontal sulci the inferiorand dorsal occipital cortices and the right inferior frontalgyrus (Fig 3 clusters in redmagenta) When we consid-ered only the conditions including the seenold images(ldquoExp 2 Seen- onlyrdquo) we replicated the effects in the supe-rior parietal gyri the inferior occipital gyri and the rightinferior frontal gyrus (see Table I and Fig 3 signal plotson the right) The activation in the intraparietal sulci andthe superior frontal sulci did not reach statistical signifi-cance Inspection of the signal plots indicated that in theseareas there was less activation in the Ssame than Sflipped tri-als (cf bar 3 vs bar 4 signal plots of the superior fontalsulcus Fig 3A plot on the right) Nonetheless the activityin the Ssame condition was larger than those in the T andO conditions (cf bar 3 vs bars 1ndash2 and 5ndash6)
Scene-recognition judgment (Object task)
Exp 1 The Object task was associated with the activationof the medial frontal cortex (MFC) and the orbitofrontalcortex (but the latter did not fully replicate in Exp 2 seeTable I and Fig 4 clusters bluemagenta and signal plotson the left) together with bilateral activation in the middletemporal gyri the retrosplenialposterior cingulate gyrusand a portion of the angular gyrus The same contrast alsorevealed an activation of the hippocampus bilaterally Theactivation of the hippocampus was just below statisticalsignificance (P-FWE-corr 5 0052 Z 5 458 at x y z 5 2126 219 and P-FWE-corr 5 0077 Z 5 458 at x y z 5 224
29 225 considering the whole brain as the volume ofinterest) and it is reported here because the same effectwas previously found in the LTM dataset [Kwok et al2012 see also Discussion Section]
Exp 2 An almost identical pattern of activation wasfound in Exp 2 when we considered all the conditionsO(old1new)gt (T(easy1hard) 1 S(same1flipped))2 Significanteffects were found in the middle temporal gyri and alarge cluster including the medial frontal gyrus extendingventrally into the orbital-frontal cortex (Fig 4 clusters inredmagenta) However when we considered only the tri-als including seenold-images [Ooldgt (T(easy1hard)2 1 Ssame)2] the Object task was found to significantlyactivate only the MFC (Table I rightmost columns) indi-cating that this region was not affected by incidentalencoding
In summary across all three tasks the brain activationwas found to be similar across Exp 1 and Exp 2 Thescene-chronology Temporal task activated the precuneusand the right angular gyrus the scene-layout Spatial taskwas associated with activation of the posteriorsuperiorparietal cortex and the right inferior frontal gyrus whilethe scene-recognition Object task activated the MFC andthe middle temporal gurus (see Table I areas highlightedin bold) The control analysis of Exp 2 that consideredonly trials including seenold images confirmed thesetask-specific effects indicating that the pattern of dissocia-tionmdashand in particular for the Spatial and Object tasksmdashwas not merely driven by the unseennew images duringthe retrieval phase
Functional Neuroimaging Correspondence with
An Analogous LTM Dataset
We directly compared the task-specific dissociationobserved in the current dataset (Exp 1) with our previousLTM study that included the same task-specific retrievaltasks but after a much longer retention delay [ie 24 hbetween encoding and retrieval Kwok et al 2012] Com-mon across the two data sets the conjunction analysesrevealed an activation cluster in the precuneus for theTemporal task (Fig 5A and Table II) and the dorsal fron-toparietal network plus right inferior frontal gyrus andright occipital cortex for the Spatial task (Fig 5B and TableII) These formal tests show that the Temporal and Spatialtask-specific dissociation observed here for immediateretrieval matched well with the patterns observed duringretrieval from LTM By contrast for the Object task over-lapping effects in the MFC and in the hippocampus weredetected only at a more lenient uncorrected threshold ofP-uncorr 5 0001 In the hippocampus the lack of a fullysignificant ldquocommonrdquo effect across datasets was due to thefact that in the current Exp 1 the contrast comparing theObject task versus the two other tasks did not survive cor-rection for multiple comparisons considering the wholebrain as the volume of interest (P-corr 5 0052 and 0077
r Neural Correlates of Immediate Episodic Retrieval r
r 2505 r
Figure 3
Clusters of brain activation and signal plots for the Scene-layout
judgment (Spatial task) Clusters of activation and signal plots for
(A) the superior frontal cortex and the superior parietal cortex
bilaterally and (B) the right inferior occipital cortex and the right
inferior frontal cortex These regions activated for the Spatial task
in Exp 1 (blue) and in Exp 2 (red overlap between Exp 1 and Exp 2
in magenta) The activation selective for the Spatial task was com-
puted with the contrast Sgt [T(short1long)2 1 O]2 inclusively
masked with SgtT(short1long)2 and with SgtO The effects in the
superior parietal gyri the inferior occipital gyri and the right inferior
frontal gyrus were replicated when considering only sameold
images in Exp 2 (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 80 voxels Effect sizes are mean adjusted (sum to zero)
and are expressed in arbitrary units (au 6 90 CI) LR leftright
SFS superior frontal sulcus SPG superior parietal gyrus IFG infe-
rior frontal gyrus IOG inferior occipital gyrus
in the left and right hemispheres respectively cf above)Indeed the formal test assessing differences between thetwo datasets (ie ldquoTask 3 Experimentrdquo interactions seeSupporting Information Table 2) did not reveal any signifi-cant difference between LTM and immediate retrieval inthe hippocampus consistent with this region beinginvolved in the scene-recognition judgment irrespective ofthe retention delay (see also Discussion Section)
DISCUSSION
We have identified distinct sets of brain areas thatshowed task-specific activation during immediate retrievalfollowing the encoding of dynamic cinematic material Theprecuneus and the right angular gyrus were activatedwhile retrieving the temporal order of events (scene-chro-nology judgment) the superior parietal cortex wasrecruited during spatial related recall (scene-layout
Figure 4
Clusters of brain activation and signal plots for the Scene-
recognition judgment (Object task) Clusters of activation and
signal plots for (A) the MFC and (B) the MTG These regions
activated for the Object task in Exp 1 (blue) and in Exp 2 (red
overlap in magenta) The activation selective for the Object task
was computed with the contrast Ogt [T(short1long)2 1 S]2
inclusively masked with OgtT(short1long)2 and with Ogt S The
signal plots refer to the left hemisphere but analogous patterns
of activation were found in the right hemisphere (Table I) In
Exp 2 the task-specific effect of the Object task in the MFC
was replicated when considering only the sameold images
(Table I rightmost columns) Inspection of the signal plot for the
left MTG revealed less activity in the Oold than Onew trials (plots
on the right bars 5 and 6) although the Oold trials showed
numerically larger parameter estimates than the T and S condi-
tions (see p-uncorr reported in italics in Table I rightmost col-
umns) Activation clusters were estimated and displayed at P-
unc 5 0001 thresholded at a minimum of 80 voxels Effect sizes
are mean adjusted (sum to zero) and expressed in arbitrary
units (au 6 90 CI) LR leftright MTG middle temporal
gyrus MFC medial frontal cortex
r Neural Correlates of Immediate Episodic Retrieval r
r 2507 r
judgment) whereas activity in the MFC and the middletemporal gyrus (MTG) reflected processes associated withscene-recognition judgments These task-specific effectsoccurred independently of whether subjects were asked tojudge seenold or unseennew stimuli (cf ldquoSeen-onlyimagesrdquo contrasts in Exp 2) and independently ofretrieval success and general task difficulty (see Support-ing Information) With respect to the hypothesized corre-spondence between immediate retrieval and retrieval fromLTM we found that these task-specific dissociationsare independent of the informationrsquos mnemonic history(24 h vs 2 s retention ie differing by 43200 times inmagnitude)
One important qualifier for the following discussion isthat memory retrieval in all our tasks depended minimallyon memory buffers compared to other simpler itemsdis-plays that are typically used in ldquostandardrdquo short-termworking memory tasks [Shallice and Cooper 2011 Shiffrin1973] In each trial the number of items presented during
encoding was clearly well beyond the 4 6 1 items stipulatedby classical STM models [Cowan 2001] While memories ofsome materials (eg words spatial positions simple visualdisplays) are supported by limited-capacity buffers [Badde-ley and Hitch 1974] memory for other categories [egunknown faces and scenes see Shiffrin 1973] will tap intoLTM when the task requires the encoding of two or moreitems [Shallice and Cooper 2011] For this reason werestrict our discussion on the tripartite task-specific dissoci-ations with reference to highly complex naturalisticmaterial
Precuneus and Inferior Parietal Cortex Support
Temporal Order Recollection
The scene-chronology judgment task activated the pre-cuneus in both Exp 1 and 2 despite the considerablechanges in the trial structure for probing the retrieval oftemporal information (ie relative temporal order betweentwo probes in Exp 1 vs absolute temporal position of asingle probe in Exp 2) The chronology-judgment taskrequired retrieving the event sequences of many complex
Figure 5
Task-specific retrieval activation common in studies after imme-
diate delays and a longer delay Activation for the LTM study
[24 h Kwok et al 2012] is depicted in green and activation
for the current immediate retrieval study (data from current
Exp 1) depicted in red Activation clusters for both studies were
estimated and displayed at P-unc 5 0001
TABLE II Common memory retrieval activation
ImMem LTM common effects
Cluster Voxel
k P-corr Z x y z
Scene-chronology (Temporal)Precuneus B 112 0011 472 3 257 47Angular gyrus R 98 lt 0001 400 54 254 17
Scene-layout (Spatial)Superior parietal gyrus L 206 0001 595 221 269 56Supramarginal gyrus L 129 0008 346 260 236 44Intraparietal sulcus L 487 236 245 41Superior parietal gyrus R 651 lt 0001 520 24 263 59Dorsal occipital cortex R 372 30 275 29Intraparietal sulcus R 520 42 239 47Supramarginal gyrus R 482 57 233 53Superior frontal sulcus L 100 0022 482 224 3 56Superior frontal sulcus R 145 0004 481 30 0 53Inferior frontal gyrus R 95 0027 475 54 9 26Inferior occipital gyrus R 98 0024 515 54 257 210
Task-specific activation after immediate delays (current Exp 1ImMem) and a previous study that used a long delay betweenencoding and retrieval [24 h LTM Kwok et al 2012] Statisticalthreshold were set at P-FWE-corr 5 005 (cluster-level) consider-ing the whole brain as the volume of interest Note that theldquobetween-studiesrdquo overlap activation in the right angular gyruswas found at a lower uncorrected threshold (in italics) but impor-tantly within this area no differences or interactions betweenImMem and LTM studies were found (see differential effectsbetween the two studies in Supporting Information Table 2) LRB leftright hemispherebilateral
r Kwok et al r
r 2508 r
stimuli (scenes) that entail high-level conceptualhierarchi-cal relationships between them [Swallow et al 2011 Zackset al 2001] The precuneus was previously found to acti-vate in our LTM protocol that used the same temporalorder judgment [Kwok et al 2012 and see also results ofthe conjunction analyses] The current finding of precunealactivation using very different (much shorter) delays fitswell with the view that these processes are not strictlyassociated with retrieval following long retention delayssuggesting that this medial parietal region is implicatedduring temporal order retrieval irrespective of delay (Fig5A and Table II) Specifically we link the effect of chrono-logical retrieval in the precuneus with processes involvingan active reconstruction of past events [see also St Jacqueset al 2008] and possibly memory-related imagery[Fletcher et al 1995 Ishai et al 2000] These processeswould characterize temporal order judgments using com-plex naturalistic material but not order-judgment withsimpler stimulus material which previous studies haveassociated with other brain regions [eg middleprefron-tal areas Suzuki et al 2002]
Together with the precuneus temporal-related judgmentalso gave rise to activation of the right angular gyrus(AG) One possible interpretation relates the inferior parie-tal cortex to the engagement of attentional processes[Majerus et al 2007 2010] The scene-chronology judg-ment of Exp 2 provided us with a further qualifier to thiskind of temporal-related attention Although the Temporaltask in Exp 2 did not require any explicit temporal orderjudgment (only classifying a scene into either the first orsecond half of the movie) an accurate decision still neces-sitated an evaluation of the target scene with many othersegments of the movie This operation is fundamentallydifferent from the evaluation implicated in the scene-layout judgment which required decisions on comparingbetween a single scene and its own correspondentencoded snapshot The latter involves visuospatial analysisof the configuration within scenes The AG may subservedifferent types of attention among which one of them islinked with memory search operations [see Majerus et al2010 2006 also showing coactivation of these two regionsduring order retrieval of simple stimuli after short delays]
Superior Parietal Cortex Evaluates Spatial
Details Against Memory Representation
The second pattern consistently found across experi-ments (Exp 1 and Exp 2 see Fig 3) and across studieswith different retention delays (conjunction betweenimmediate retrieval and LTM studies see Fig 5B andTable II) was the activation of the superior parietal lobule(SPL) during the scene-layout judgment Previous studiesshowed sustained activation in the parietal cortex duringworking memory delays [Courtney et al 1998] and thatactivity there correlates with memory load [Todd andMaroi 2004] Here the amount of information contained
in the clips was equated in the three tasks especially inExp 2 where we counterbalanced the assignment of mov-ies across tasks The subjects also could not anticipatewhat specific type of information would be tested atretrieval and needed to keep rehearsing different aspectsof the clips Because of this we deem unlikely that theactivation of SPL that here was specific for the scene-layout tasks reflected general rehearsal processes [Magenet al 2009] andor updating operations [Bledowski et al2009 Leung et al 2007]
Instead we suggest a role for the SPL in mediating aninterface between the internal memory representation andexternal information It has been proposed that the supe-rior parietal cortex is engaged in allocating top-downattention to orient to aspects of the available visual inputand on information retrieved from memory [Ishai et al2002 Lepsien and Nobre 2007 Summerfield et al 2006]In line with this we found SPL activation when the sub-jects had to make decisions by comparing between theirmemory of the encoded clips and the scene stimuli pre-sented at retrieval This procedure might act on somevisuospatial storage which is held by the lateral occipitalcortex and mediated by the intraparietal sulcus (Fig 3B)The immediate memory task here was analogous to theSpatial task of our previous LTM study [Kwok et al2012] which also required matchingcomparing the avail-able external visual input and internal information storedin memory [see also Nobre et al 2004] This interfaceinvolves a range of top-down processes such as monitor-ing and verification [Cabeza et al 2008 Ciaramelli et al2008] which are independent of the probe imagersquos mne-monic status as shown by the equated effect sizes forldquosamerdquo and ldquoflippedrdquo images conditions (see signals plotsin Fig 3 and also ldquoRetrieval successrdquo analyses of Exp 2 inSupporting Information)
Recognition Processes in the Medial Frontal
Cortex and the Temporal Cortex
The third retrieval task involved scene-recognition judg-ments and was associated with the activation of the MFCand the MTG Moreover in Exp 1 we found a statisticaltrend in the hippocampus bilaterally an area that we pre-viously associated with scene-recognition judgment fromLTM (see next section about ldquoPossible correspondencesbetween retrieval from short- and long-term memoryrdquo)
The MFC was engaged both Exp 1 and 2 but was notactivated in the other two tasks thus a broad prefrontalcortexworking memory correspondence [Tsuchida andFellows 2009] seems insufficient to explain this task-specific effect The scene-recognition task in Exp 2 taxedboth recognition of ldquooldrdquo scenes and correct rejection ofldquonewrdquo scenes suggesting that the MFC here did not dis-tinguish between the correct rejection of new information(unseennew trials) and the recognition of old imagesOne candidate cognitive process that is common to both
r Neural Correlates of Immediate Episodic Retrieval r
r 2509 r
types of processes is feeling-of-knowing (FOK) FOKdepend on the cue specification content retrieval andappraisal of the retrieved memory fragments but are inde-pendent of the ultimate determination of content appropri-ateness [Schnyer et al 2005] Using imaging data duringsentence completion recognition Schnyer et al [2005]modeled a FOK-related directional path The path impli-cated the temporal cortex and the hippocampus andengaged the ventro-MFC at the end which is consistentwith our findings for the Object task implicating the MFCthe middle temporal cortex and at a lower statisticalthreshold the hippocampus
By contrast the activity in the MTG was larger forunseennew images than for seenold memory probes(Fig 4B) This suggests that new encoding may have con-tributed to the activation of the MTG during the retrievalphase of the Object task in Exp 1 The scene-recognitiontask required a lower precision in terms of retrievingdetails about the encoded material and could be solved bysimple recognition [Wixted and Squire 2010] Thus alter-natively larger activation for ldquonew vs oldrdquo stimuli mayrelate to some reduced activation associated with familiar-ity processes on old trials [see Henson et al 1999 whoreported reduced familiarity related activation in middletemporal gyri bilaterally and Henson and Rugg 2003 forreview] However we should point out that in Exp 2 theunseen images were extracted from edited-out parts of theoriginal clips and therefore contained the same objectcharacter depicted in the target images Therefore wedeem more likely that the relational association betweenobjects rather than their presenceabsence in the sceneshelped subjects to distinguish targets from foils We thussuggest that the activation of the MTG reflects processesutilizing the relational details among objects embedded inthe probe images for reaching the recognition decision[Diana et al 2007 Finke et al 2008 Hannula et al 2006Olson et al 2006] Future studies should consider theinclusion of some correlative behavioral measures to indexfamiliarity processes to elucidate further this issue More-over while the Temporal and Spatial tasks required mak-ing relative comparisons about aspects of the encodedvideo-clip the scene-recognition judgment merely requiredjudging whether the image presented at retrieval was pres-ent (or not) within the clip This may entail different proc-essing dynamics in the Object task compared with theother two tasks (cf also differences in retrieval times) Thelatter could be addressed using more subtle experimentalmanipulations for the scene-recognition judgment forexample by using digitally-modified foil images where asingle object in the scene has been modifiedremoved
Possible Correspondences Between Retrieval
from Short- and Long-Term Memory
The overlap between our current study and the previousLTM study using analogous tasks and materials lends
weight to the notion of scale-invariance in memory mecha-nisms [Brown et al 2007] as have been previously dem-onstrated in free recall tasks where the rate of item recallwas unvarying across recall span (day week year) andeven across from the past to the prospective future [May-lor et al 2001] Those previous behavioral findings implythat scale-invariance is a form of self-similarity in that theholistic pattern can exist at multiple levels of magnification[and even across different memory systems eg Maylor2002] Here our data affirmed the importance of the poste-rior parietal cortex in both immediate and long-termretrieval [Berryhill et al 2011 2007] and further character-ized its involvement in dealing with the spatial relationsof objects within scenes [Bricolo et al 2000 Buiatti et al2011] Our data also demonstrated such short-long-termcorrespondence exists in the precuneusangular gyrus fortemporal order retrieval (cf scene-chronology judgmenttask) In contrast the bilateral hippocampus and MFC acti-vations did not reach full statistical significance in theImMem LTM conjunction analysis At first sight thismay indicate that the scene-recognition task tapped intodistinct recognition-based mechanisms as a function ofretention delay However we deem this unlikely becausethe direct comparison between the two datasets did notshow any significant effect in these regions (see Support-ing Information Table 2) Indeed a more targeted test ask-ing the question of whether the hippocampus clusterspreviously found in LTM activated also in the currentimmediate retrieval experiments revealed significanteffects in both immediate retrieval experiments here (Exp1 x y z 5 224 29 225 P-FWE-corrlt 0001 Z 5 458 x yz 5 21 29 222 P-FWE-corr 5 0001 Z 5 451 and in Exp2 x y z 5 224 214 221 P-FWE-corr 5 0004 Z 5 396using the LTM clusters for small volume correction) Theseadditional tests point to a more general issue of sensitivitythat future studies may address using different methodo-logical approaches such as multivariate methods that canbe more sensitive than the standard univariate analysesused here Moreover the former would enable capturingtask-specific effects that are expressed as patterns of dis-tributed activation rather than local changes of activity asreported here [eg Jimura and Poldrack 2012]
Task Difficulty and Retrieval Success
We used several control analyses to show that the task-specific patterns of activation and the correspondencebetween the current data and our previous LTM study donot merely reflect some effect related to general task diffi-culty (see Supporting Information) First areas involved inoverall task difficulty should show progressively greateractivation with progressively more difficult tasks In Exp 1this would entail SgtTgtO However the patterns in theprecuneus and the angular gyrus (with TgtOgt S Fig 2A)and the MFC (Ogt SgtT Fig 4A) did not fit with simplepredictions based on overall task difficulty We also
r Kwok et al r
r 2510 r
assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r

episode used in the LTM paradigm Because of these differ-ences our analyses focused on commonalities rather thandifferences between LTM and immediate retrieval Theseanalyses considered the current Exp 1 which included thesame retrieval tasks as the LTM experiment Separately foreach of the three tasks we computed the relevant contrasts(eg Tgt (S1O)2) in the two studies and entered these inthree separate two samples t-tests For the identification ofcommon activation across studies we performed null-conjunction analyses (eg [Tgt (S1O)2]LTM [Tgt (S1O)2]ImMem) Statistical threshold were set at P-FWE-corr 5 005 cluster-level corrected at the whole brain level(cluster size estimated and displayed at P-unc 5 0001) con-sistent with all the analyses above For completeness thecontrasts testing for any differences between studies arereported in Supporting Information Table 2
Additional control analyses Retrieval success
Finally we asked to what extent any task-specific effectsidentified in our main analyses depended on the success-ful retrieval of stored information in the clips For this wecarried out additional analyses testing for retrieval per-formance in Exp 1 (ldquoTask 3 Accuracyrdquo in T and S tasks)and for accurate performance on seenold vs unseennewstimuli in Exp 2 (ldquoTask 3 Seen-unseenrdquo in S and O tasks)Please see Supporting Information for details of these anal-yses and the corresponding results
RESULTS
Behavioral Results
Experiment 1
Subjects performed significantly above chance in allthree tasks (all Plt 0001) The percentage of correct accu-racy and response times (RT) were analyzed usingrepeated measures ANOVAs with three levels correspond-ing to the TSO tasks This showed a main effect of con-ditions both for accuracy [F(2 32) 5 8142 Plt 0001] andRT [F(232) 5 3123 Plt 0001] Post hoc t-tests revealedthat subjects performed significantly better and faster inthe Object than Spatial task [Accuracy t(16) 5 1311Plt 0001 RT t(16) 5 652 Plt 0001] and Object than Tem-poral task [Accuracy t(16) 5 979 Plt 0001 RT t(16) 5
671 Plt 0001] Subjects were also more accurate on Tem-poral trials than on Spatial trials [Accuracy t(16) 5 348Plt 0001 but no difference in RT Pgt 01] For the Tempo-ral task we also examined the effect of temporal distanceexpecting that the retrieval would be more difficult whenthe two probes were temporally close (Tshortlt 19 s) com-pared with temporally far (Tlonggt 19 s) during encodingIndeed t-tests showed that subjects responded less accu-rately and slower in short than in long distance trials[Accuracy t(16) 5 391 Plt 0005 RT t(16) 5 315 Plt 001](Fig 1B plot on the left)
Experiment 2
In Exp 2 subjects also performed better than chance inall three tasks (all Plt 0001) Repeated measures ANOVAsshowed main effects of tasks both for accuracy [F(232) 5 919 Plt 0005] and RT [F(232) 5 1766 Plt 0001]Post hoc t-tests revealed that subjects performed withhigher accuracy and shorter RT in the Object comparedwith the Spatial task [Accuracy t(16) 5 406 Plt 0001 RTt(16) 5 532 Plt 0001] and in the Object compared withthe Temporal task [Accuracy t(16) 5 277 Plt 005 RTt(16) 5 383 Plt 0005] The difference in accuracy betweenthe Temporal and Spatial tasks approached significance(Plt 009) while there was no RT difference between Tem-poral and Spatial tasks (Pgt 01) Additional t-tests directlycompared different trial types within each task For theTemporal task subjects performed more accurately andfaster in the ldquoeasyrdquo than ldquohardrdquo trials [Accuracyt(16) 5 657 Plt 0001 RT t(16) 5 527 Plt 0001)] For theSpatial task they were more accurate in recognizingldquosamerdquo than correctly rejecting ldquoflippedrdquo images [Accu-racy t(16) 5 534 Plt 0001 RT Pgt 01] For the Objecttask subjects were faster to respond to unseen Onew
images compared to seen Oold images [RT t(16) 5 294Plt 001] without any difference in accuracy (Pgt 01) (Fig1B plot on the right)
Functional Neuroimaging Task-Specific Retrieval
In both Exp 1 and 2 we assessed brain activity specificto each of the three retrieval tasks (Temporal Spatial andObject) by comparing each task versus the mean of theother two (ldquoTask specificityrdquo contrasts see SupportingInformation Table 1) In Exp 2 we also carried out addi-tional nonindependent control tests using only trials con-taining seenold images (ldquoSeen-only imagesrdquo contrasts)The latter served to ensure that any task-specific dissocia-tion did not arise from the presentation of unseennewimages during the retrieval phase of the Spatial and Objecttasks (ie flipped- and new-image foils)
Scene-chronology judgment (Temporal task)
Exp 1 The comparison of the Temporal task with theother two retrieval conditions revealed significant activa-tion in the right precuneus bilateral caudate and bilateralcuneus (see Table I and Fig 2A clusters in bluemagenta) The signal plots in Figure 2 (panels on the left)show that the precuneus and the right angular gyrus wereactivated more by the Temporal task (average of Tshort andTlong) than by the Spatial and Object tasks Note that theeffect in the right angular gyrus was detected only atuncorrected threshold (P-unclt 0001) and it is reportedhere because it was fully significant in Exp 2 (see below)and as well as in the LTM dataset [Kwok et al 2012]
Together with the main comparison for the Temporaltask in Exp 1 we assessed the effect of temporal distance
r Kwok et al r
r 2502 r
TA
BL
EI
Task
-sp
ecif
icre
trie
val
acti
vati
on
Ex
p
1E
xp
2
Ex
p
2S
een
-on
ly
Clu
ster
Vo
xel
Clu
ster
Vo
xel
Clu
ster
Vo
xel
KP
-co
rr
Zx
yz
kP
-co
rr
Zx
yz
KP
-co
rr
Zx
yz
Sce
ne-
chro
no
log
y(T
emp
ora
l)P
recu
neu
sB
258
lt00
01
71
06
254
47
2666
lt00
01
51
74
272
43
1835
lt00
01
40
06
270
41
Po
st
cin
gu
late
cort
exB
mdashmdash
mdashmdash
564
02
3027
535
62
3631
Cau
dat
eB
148
000
54
302
122
1514
mdashmdash
mdashmdash
mdashmdash
mdashmdash
Cu
neu
sL
820
052
522
215
260
17mdash
mdashmdash
mdashmdash
mdashmdash
mdashC
un
eus
R88
004
25
2118
257
20mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nto
po
lar
cort
exR
mdashmdash
mdashmdash
874
lt0
001
547
2860
970
60
001
575
2860
7S
up
erio
rfr
on
tal
cort
exR
mdashmdash
mdashmdash
685
000
15
3526
2455
889
000
15
6026
2853
An
gu
lar
gy
rus
R43
lt00
01
46
345
269
29
457
00
07
48
444
266
45
509
00
04
45
844
268
43
Sce
ne-
lay
ou
t(S
pat
ial)
Su
peri
or
pari
eta
lg
yru
sL
1142
lt00
01
69
02
21
266
59
2530
lt00
01
66
72
18
268
59
377
00
16
49
92
18
266
59
Do
rsal
occ
ipit
alco
rtex
L5
112
272
759
521
228
280
9mdash
mdashmdash
mdashS
up
ram
arg
inal
gy
rus
L6
212
602
3338
mdashmdash
mdashmdash
mdashmdash
Intr
apar
ieta
lsu
lcu
sL
577
242
239
415
562
362
4445
mdashmdash
mdashmdash
Su
peri
or
pari
eta
lg
yru
sR
1314
lt00
01
67
221
263
59
3026
lt00
01
75
516
266
59
1262
lt00
01
47
426
268
47
Do
rsal
occ
ipit
alco
rtex
R4
1230
293
237
4034
278
294
1030
274
39In
trap
arie
tal
sulc
us
R6
2536
239
4433
40
026
548
402
4045
mdashmdash
mdashmdash
Su
pra
mar
gin
alg
yru
sR
597
602
2435
mdashmdash
mdashmdash
mdashmdash
Su
per
ior
fro
nta
lsu
lcu
sL
213
000
16
452
243
6223
80
078
543
226
22
51mdash
mdashmdash
mdashS
up
erio
rfr
on
tal
sulc
us
R18
20
002
592
302
356
220
009
74
7028
053
mdashmdash
mdashmdash
Infe
rio
rfr
on
tal
gy
rus
L92
003
65
002
516
26mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nta
lg
yru
sR
304
lt00
01
64
054
12
11
536
00
03
60
752
12
25
238
00
78
48
050
12
25
Infe
rio
ro
ccip
ital
gy
rus
L152
00
05
48
12
57
260
210
894
lt00
01
60
32
52
264
211
591
00
02
50
32
50
266
211
Infe
rio
ro
ccip
ital
gy
rus
R186
00
02
67
754
260
213
2277
lt00
01
68
256
258
211
1161
lt00
01
48
750
258
217
Occ
ipit
alp
ole
B21
80
001
484
29
293
2mdash
mdashmdash
mdashmdash
mdashmdash
mdashS
cen
e-re
cog
nit
ion
(Ob
ject
)M
ed
ial
fro
nta
lco
rtex
B478
lt00
01
51
92
351
47
2215
lt00
01
59
62
856
39
314
00
32
46
42
10
56
39
Mid
dle
tem
po
ral
gy
rus
L490
lt00
01
59
42
60
212
225
1661
lt00
01
67
72
62
214
219
162
lt00
01
42
92
60
24
225
Mid
dle
tem
po
ral
gy
rus
R304
lt00
01
61
560
212
216
1017
lt00
01
56
960
26
223
15
lt00
02
29
662
24
223
Orb
ito
-fro
nta
lco
rtex
B95
001
45
040
512
2222
70
049
506
044
219
mdashmdash
mdashmdash
An
gu
lar
gy
rus
L22
70
001
530
251
254
29mdash
mdashmdash
mdashmdash
mdashmdash
mdashA
ng
ula
rg
yru
sR
142
000
64
9857
260
38mdash
mdashmdash
mdashmdash
mdashmdash
mdashR
SC
p
ost
ci
ng
ula
teco
rtex
B47
4lt
000
14
903
242
35mdash
mdashmdash
mdashmdash
mdashmdash
mdash
T-c
on
tras
tsco
mp
ared
each
con
dit
ion
ver
sus
the
mea
no
fth
eo
ther
two
con
dit
ion
s(e
g
T(s
ho
rt1
lon
g)
2gt
(S1
O)
2)
and
was
incl
usi
vel
ym
ask
edw
ith
the
acti
vat
ion
for
the
crit
ical
con
dit
ion
ver
sus
each
of
the
two
oth
erco
nd
itio
ns
(eg
T
(sh
ort
1lo
ng
)2gt
S2
and
T(s
ho
rt1
lon
g)
2gt
O
2)
InE
xp2
we
con
du
cted
two
sep
arat
ese
tso
fco
ntr
asts
atth
eg
rou
p-l
evel
F
irst
t-
con
tras
tsw
ere
con
du
cted
usi
ng
all
six
con
dit
ion
sto
com
par
eea
chco
nd
itio
nv
ersu
sth
em
ean
of
the
oth
ertw
oco
nd
itio
ns
(eg
S (
sam
e1
flip
ped
)gt
[T(e
asy
1h
ard
)1
O(o
ld1
new
)]
2)
see
colu
mn
sin
the
mid
dle
Se
con
d
for
Spat
ial
and
Ob
ject
con
dit
ion
sw
ew
ere
inte
rest
edin
acti
vit
yel
icit
edb
yth
ere
spo
nse
sas
soci
ated
wit
hldquos
een
rdquotr
ials
on
ly(e
g
S sa-
megt
[T(e
asy
1h
ard
)2
1O
old
]2)
se
eri
gh
tmo
stco
lum
ns
and
also
Sup
po
rtin
gIn
form
atio
nT
able
1B
rain
reg
ion
sh
igh
lig
hte
din
bo
ldco
rres
po
nd
toth
eta
sk-s
pec
ific
effe
cts
con
sist
entl
yo
bse
rved
acro
ssal
lex
per
imen
ts
anal
yse
sth
ese
reg
ion
sar
eal
sod
isp
lay
edin
Fig
ure
s2ndash
4P
-val
ues
are
corr
ecte
dfo
rm
ult
iple
com
par
iso
ns
exce
pt
for
the
rig
ht
ang
ula
rg
yru
sin
Exp
1an
dth
em
idd
lete
mp
ora
lg
yri
inE
xp2
Seen
-on
lyth
atar
ere
po
rted
ata
low
eru
nco
rrec
ted
thre
sho
ld(i
nit
alic
s)
Po
st
po
ster
ior
xy
zco
ord
inat
esin
the
stan
dar
dM
NI
spac
eo
fth
eac
tiv
atio
np
eak
sin
the
clu
ster
sk
nu
mb
ero
fv
oxe
lsin
each
clu
ster
R
SC
retr
osp
len
ial
cort
ex
L
R
B
left
ri
gh
th
emis
ph
ere
bil
ater
al
r Neural Correlates of Immediate Episodic Retrieval r
r 2503 r
by comparing Tshort vs Tlong trials In previous studiesusing LTM protocols we found that temporal distance fur-ther modulated activity in the precuneus [Kwok et al2012 2014 see also St Jacques et al 2008] Here this con-trast did not reveal any significant effect This is in con-trast to the significant behavioral cost of retrieving Tshort
vs Tlong temporal distances (see behavioral results andFig 1B plot on the left) that was analogous to our previ-ous LTM protocol Similar behavioral patterns have beenobserved in many other short-term studies [Hacker 1980Konishi et al 2002 Milner et al 1991 Muter 1979] yet toour knowledge there does not seem to exists a clear con-
sensus on the neural correlates subserving the temporaldistance effect separating two temporally very nearbyeventsitems
Exp 2 In Exp 2 we replicated the effect of the temporalretrieval task in the precuneus and found a fully signifi-cant effect in the right angular gyrus (see Fig 2 clusters inredmagenta) In addition in Exp 2 there was a significantactivation of the right superior frontal cortex and the rightinferior frontopolar cortex that were not found in Exp 1(Table I) The involvement of the precuneus and the rightangular gyrus was further confirmed using only trials
Figure 2
Clusters of brain activation and signal plots for the scene-
chronology judgment (Temporal task) Clusters of activation and
signal plots for (A) the precuneus and (B) the right angular
gyrus These regions activated selectively during the temporal
tasks in Exp 1 (in blue) Exp 2 (in red overlap between Exp 1
and 2 shown in magenta) The activation selective for the Tem-
poral task was computed with the contrast T(short1long)
2gt (S 1 O)2 inclusively masked with T(short1long)2gt S2 and
with T(short1long)2gtO2 The involvement of the precuneus and
the right angular gyrus was further confirmed using only ldquoseen
oldrdquo images (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 40 voxels Effect sizes are mean adjusted (sum to
zero) and expressed in arbitrary units (au 6 90 CI) LR left
right PreC precuneus AnG angular gyrus
r Kwok et al r
r 2504 r
including seen images (cf ldquoExp 2 Seen- onlyrdquo Table I)The signal plots on Figure 2 (panels on the right) show theactivity of the precuneus and the right angular gyrus forall the conditions of Exp 2 Both in the precuneus and theright angular gyrus the average activity in the temporaltask [T(easy1hard)2] were larger than the Ssame and theOold conditions Notably Exp 2 included the presentationof a single image and that the participants were asked tojudge the absolute temporal position of the probe ratherthan the relative temporal order Despite this importantchange of how temporal memory was probed the findingof analogous patterns of activation across both experi-ments highlights the link between immediate retrieval oftemporal information and activation of the precuneus andangular gyrus
Scene-layout judgment (Spatial task)
Exp 1 The contrast concerning the retrieval of the spatiallayout revealed activation in the superior parietal cortexbilaterally with the cluster extending to the intraparietalsulcus and the supramarginal gyrus This contrast alsoshowed activation of the superior frontal sulci the inferiorfrontal gyri plus the inferior occipital cortex (see Table Iand Fig 3 clusters displayed in bluemagenta signalsplots on the left side)
Exp 2 The same comparison in Exp 2 replicated theresults of Exp 1 showing activation in the superior parie-tal cortex bilaterally the superior frontal sulci the inferiorand dorsal occipital cortices and the right inferior frontalgyrus (Fig 3 clusters in redmagenta) When we consid-ered only the conditions including the seenold images(ldquoExp 2 Seen- onlyrdquo) we replicated the effects in the supe-rior parietal gyri the inferior occipital gyri and the rightinferior frontal gyrus (see Table I and Fig 3 signal plotson the right) The activation in the intraparietal sulci andthe superior frontal sulci did not reach statistical signifi-cance Inspection of the signal plots indicated that in theseareas there was less activation in the Ssame than Sflipped tri-als (cf bar 3 vs bar 4 signal plots of the superior fontalsulcus Fig 3A plot on the right) Nonetheless the activityin the Ssame condition was larger than those in the T andO conditions (cf bar 3 vs bars 1ndash2 and 5ndash6)
Scene-recognition judgment (Object task)
Exp 1 The Object task was associated with the activationof the medial frontal cortex (MFC) and the orbitofrontalcortex (but the latter did not fully replicate in Exp 2 seeTable I and Fig 4 clusters bluemagenta and signal plotson the left) together with bilateral activation in the middletemporal gyri the retrosplenialposterior cingulate gyrusand a portion of the angular gyrus The same contrast alsorevealed an activation of the hippocampus bilaterally Theactivation of the hippocampus was just below statisticalsignificance (P-FWE-corr 5 0052 Z 5 458 at x y z 5 2126 219 and P-FWE-corr 5 0077 Z 5 458 at x y z 5 224
29 225 considering the whole brain as the volume ofinterest) and it is reported here because the same effectwas previously found in the LTM dataset [Kwok et al2012 see also Discussion Section]
Exp 2 An almost identical pattern of activation wasfound in Exp 2 when we considered all the conditionsO(old1new)gt (T(easy1hard) 1 S(same1flipped))2 Significanteffects were found in the middle temporal gyri and alarge cluster including the medial frontal gyrus extendingventrally into the orbital-frontal cortex (Fig 4 clusters inredmagenta) However when we considered only the tri-als including seenold-images [Ooldgt (T(easy1hard)2 1 Ssame)2] the Object task was found to significantlyactivate only the MFC (Table I rightmost columns) indi-cating that this region was not affected by incidentalencoding
In summary across all three tasks the brain activationwas found to be similar across Exp 1 and Exp 2 Thescene-chronology Temporal task activated the precuneusand the right angular gyrus the scene-layout Spatial taskwas associated with activation of the posteriorsuperiorparietal cortex and the right inferior frontal gyrus whilethe scene-recognition Object task activated the MFC andthe middle temporal gurus (see Table I areas highlightedin bold) The control analysis of Exp 2 that consideredonly trials including seenold images confirmed thesetask-specific effects indicating that the pattern of dissocia-tionmdashand in particular for the Spatial and Object tasksmdashwas not merely driven by the unseennew images duringthe retrieval phase
Functional Neuroimaging Correspondence with
An Analogous LTM Dataset
We directly compared the task-specific dissociationobserved in the current dataset (Exp 1) with our previousLTM study that included the same task-specific retrievaltasks but after a much longer retention delay [ie 24 hbetween encoding and retrieval Kwok et al 2012] Com-mon across the two data sets the conjunction analysesrevealed an activation cluster in the precuneus for theTemporal task (Fig 5A and Table II) and the dorsal fron-toparietal network plus right inferior frontal gyrus andright occipital cortex for the Spatial task (Fig 5B and TableII) These formal tests show that the Temporal and Spatialtask-specific dissociation observed here for immediateretrieval matched well with the patterns observed duringretrieval from LTM By contrast for the Object task over-lapping effects in the MFC and in the hippocampus weredetected only at a more lenient uncorrected threshold ofP-uncorr 5 0001 In the hippocampus the lack of a fullysignificant ldquocommonrdquo effect across datasets was due to thefact that in the current Exp 1 the contrast comparing theObject task versus the two other tasks did not survive cor-rection for multiple comparisons considering the wholebrain as the volume of interest (P-corr 5 0052 and 0077
r Neural Correlates of Immediate Episodic Retrieval r
r 2505 r
Figure 3
Clusters of brain activation and signal plots for the Scene-layout
judgment (Spatial task) Clusters of activation and signal plots for
(A) the superior frontal cortex and the superior parietal cortex
bilaterally and (B) the right inferior occipital cortex and the right
inferior frontal cortex These regions activated for the Spatial task
in Exp 1 (blue) and in Exp 2 (red overlap between Exp 1 and Exp 2
in magenta) The activation selective for the Spatial task was com-
puted with the contrast Sgt [T(short1long)2 1 O]2 inclusively
masked with SgtT(short1long)2 and with SgtO The effects in the
superior parietal gyri the inferior occipital gyri and the right inferior
frontal gyrus were replicated when considering only sameold
images in Exp 2 (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 80 voxels Effect sizes are mean adjusted (sum to zero)
and are expressed in arbitrary units (au 6 90 CI) LR leftright
SFS superior frontal sulcus SPG superior parietal gyrus IFG infe-
rior frontal gyrus IOG inferior occipital gyrus
in the left and right hemispheres respectively cf above)Indeed the formal test assessing differences between thetwo datasets (ie ldquoTask 3 Experimentrdquo interactions seeSupporting Information Table 2) did not reveal any signifi-cant difference between LTM and immediate retrieval inthe hippocampus consistent with this region beinginvolved in the scene-recognition judgment irrespective ofthe retention delay (see also Discussion Section)
DISCUSSION
We have identified distinct sets of brain areas thatshowed task-specific activation during immediate retrievalfollowing the encoding of dynamic cinematic material Theprecuneus and the right angular gyrus were activatedwhile retrieving the temporal order of events (scene-chro-nology judgment) the superior parietal cortex wasrecruited during spatial related recall (scene-layout
Figure 4
Clusters of brain activation and signal plots for the Scene-
recognition judgment (Object task) Clusters of activation and
signal plots for (A) the MFC and (B) the MTG These regions
activated for the Object task in Exp 1 (blue) and in Exp 2 (red
overlap in magenta) The activation selective for the Object task
was computed with the contrast Ogt [T(short1long)2 1 S]2
inclusively masked with OgtT(short1long)2 and with Ogt S The
signal plots refer to the left hemisphere but analogous patterns
of activation were found in the right hemisphere (Table I) In
Exp 2 the task-specific effect of the Object task in the MFC
was replicated when considering only the sameold images
(Table I rightmost columns) Inspection of the signal plot for the
left MTG revealed less activity in the Oold than Onew trials (plots
on the right bars 5 and 6) although the Oold trials showed
numerically larger parameter estimates than the T and S condi-
tions (see p-uncorr reported in italics in Table I rightmost col-
umns) Activation clusters were estimated and displayed at P-
unc 5 0001 thresholded at a minimum of 80 voxels Effect sizes
are mean adjusted (sum to zero) and expressed in arbitrary
units (au 6 90 CI) LR leftright MTG middle temporal
gyrus MFC medial frontal cortex
r Neural Correlates of Immediate Episodic Retrieval r
r 2507 r
judgment) whereas activity in the MFC and the middletemporal gyrus (MTG) reflected processes associated withscene-recognition judgments These task-specific effectsoccurred independently of whether subjects were asked tojudge seenold or unseennew stimuli (cf ldquoSeen-onlyimagesrdquo contrasts in Exp 2) and independently ofretrieval success and general task difficulty (see Support-ing Information) With respect to the hypothesized corre-spondence between immediate retrieval and retrieval fromLTM we found that these task-specific dissociationsare independent of the informationrsquos mnemonic history(24 h vs 2 s retention ie differing by 43200 times inmagnitude)
One important qualifier for the following discussion isthat memory retrieval in all our tasks depended minimallyon memory buffers compared to other simpler itemsdis-plays that are typically used in ldquostandardrdquo short-termworking memory tasks [Shallice and Cooper 2011 Shiffrin1973] In each trial the number of items presented during
encoding was clearly well beyond the 4 6 1 items stipulatedby classical STM models [Cowan 2001] While memories ofsome materials (eg words spatial positions simple visualdisplays) are supported by limited-capacity buffers [Badde-ley and Hitch 1974] memory for other categories [egunknown faces and scenes see Shiffrin 1973] will tap intoLTM when the task requires the encoding of two or moreitems [Shallice and Cooper 2011] For this reason werestrict our discussion on the tripartite task-specific dissoci-ations with reference to highly complex naturalisticmaterial
Precuneus and Inferior Parietal Cortex Support
Temporal Order Recollection
The scene-chronology judgment task activated the pre-cuneus in both Exp 1 and 2 despite the considerablechanges in the trial structure for probing the retrieval oftemporal information (ie relative temporal order betweentwo probes in Exp 1 vs absolute temporal position of asingle probe in Exp 2) The chronology-judgment taskrequired retrieving the event sequences of many complex
Figure 5
Task-specific retrieval activation common in studies after imme-
diate delays and a longer delay Activation for the LTM study
[24 h Kwok et al 2012] is depicted in green and activation
for the current immediate retrieval study (data from current
Exp 1) depicted in red Activation clusters for both studies were
estimated and displayed at P-unc 5 0001
TABLE II Common memory retrieval activation
ImMem LTM common effects
Cluster Voxel
k P-corr Z x y z
Scene-chronology (Temporal)Precuneus B 112 0011 472 3 257 47Angular gyrus R 98 lt 0001 400 54 254 17
Scene-layout (Spatial)Superior parietal gyrus L 206 0001 595 221 269 56Supramarginal gyrus L 129 0008 346 260 236 44Intraparietal sulcus L 487 236 245 41Superior parietal gyrus R 651 lt 0001 520 24 263 59Dorsal occipital cortex R 372 30 275 29Intraparietal sulcus R 520 42 239 47Supramarginal gyrus R 482 57 233 53Superior frontal sulcus L 100 0022 482 224 3 56Superior frontal sulcus R 145 0004 481 30 0 53Inferior frontal gyrus R 95 0027 475 54 9 26Inferior occipital gyrus R 98 0024 515 54 257 210
Task-specific activation after immediate delays (current Exp 1ImMem) and a previous study that used a long delay betweenencoding and retrieval [24 h LTM Kwok et al 2012] Statisticalthreshold were set at P-FWE-corr 5 005 (cluster-level) consider-ing the whole brain as the volume of interest Note that theldquobetween-studiesrdquo overlap activation in the right angular gyruswas found at a lower uncorrected threshold (in italics) but impor-tantly within this area no differences or interactions betweenImMem and LTM studies were found (see differential effectsbetween the two studies in Supporting Information Table 2) LRB leftright hemispherebilateral
r Kwok et al r
r 2508 r
stimuli (scenes) that entail high-level conceptualhierarchi-cal relationships between them [Swallow et al 2011 Zackset al 2001] The precuneus was previously found to acti-vate in our LTM protocol that used the same temporalorder judgment [Kwok et al 2012 and see also results ofthe conjunction analyses] The current finding of precunealactivation using very different (much shorter) delays fitswell with the view that these processes are not strictlyassociated with retrieval following long retention delayssuggesting that this medial parietal region is implicatedduring temporal order retrieval irrespective of delay (Fig5A and Table II) Specifically we link the effect of chrono-logical retrieval in the precuneus with processes involvingan active reconstruction of past events [see also St Jacqueset al 2008] and possibly memory-related imagery[Fletcher et al 1995 Ishai et al 2000] These processeswould characterize temporal order judgments using com-plex naturalistic material but not order-judgment withsimpler stimulus material which previous studies haveassociated with other brain regions [eg middleprefron-tal areas Suzuki et al 2002]
Together with the precuneus temporal-related judgmentalso gave rise to activation of the right angular gyrus(AG) One possible interpretation relates the inferior parie-tal cortex to the engagement of attentional processes[Majerus et al 2007 2010] The scene-chronology judg-ment of Exp 2 provided us with a further qualifier to thiskind of temporal-related attention Although the Temporaltask in Exp 2 did not require any explicit temporal orderjudgment (only classifying a scene into either the first orsecond half of the movie) an accurate decision still neces-sitated an evaluation of the target scene with many othersegments of the movie This operation is fundamentallydifferent from the evaluation implicated in the scene-layout judgment which required decisions on comparingbetween a single scene and its own correspondentencoded snapshot The latter involves visuospatial analysisof the configuration within scenes The AG may subservedifferent types of attention among which one of them islinked with memory search operations [see Majerus et al2010 2006 also showing coactivation of these two regionsduring order retrieval of simple stimuli after short delays]
Superior Parietal Cortex Evaluates Spatial
Details Against Memory Representation
The second pattern consistently found across experi-ments (Exp 1 and Exp 2 see Fig 3) and across studieswith different retention delays (conjunction betweenimmediate retrieval and LTM studies see Fig 5B andTable II) was the activation of the superior parietal lobule(SPL) during the scene-layout judgment Previous studiesshowed sustained activation in the parietal cortex duringworking memory delays [Courtney et al 1998] and thatactivity there correlates with memory load [Todd andMaroi 2004] Here the amount of information contained
in the clips was equated in the three tasks especially inExp 2 where we counterbalanced the assignment of mov-ies across tasks The subjects also could not anticipatewhat specific type of information would be tested atretrieval and needed to keep rehearsing different aspectsof the clips Because of this we deem unlikely that theactivation of SPL that here was specific for the scene-layout tasks reflected general rehearsal processes [Magenet al 2009] andor updating operations [Bledowski et al2009 Leung et al 2007]
Instead we suggest a role for the SPL in mediating aninterface between the internal memory representation andexternal information It has been proposed that the supe-rior parietal cortex is engaged in allocating top-downattention to orient to aspects of the available visual inputand on information retrieved from memory [Ishai et al2002 Lepsien and Nobre 2007 Summerfield et al 2006]In line with this we found SPL activation when the sub-jects had to make decisions by comparing between theirmemory of the encoded clips and the scene stimuli pre-sented at retrieval This procedure might act on somevisuospatial storage which is held by the lateral occipitalcortex and mediated by the intraparietal sulcus (Fig 3B)The immediate memory task here was analogous to theSpatial task of our previous LTM study [Kwok et al2012] which also required matchingcomparing the avail-able external visual input and internal information storedin memory [see also Nobre et al 2004] This interfaceinvolves a range of top-down processes such as monitor-ing and verification [Cabeza et al 2008 Ciaramelli et al2008] which are independent of the probe imagersquos mne-monic status as shown by the equated effect sizes forldquosamerdquo and ldquoflippedrdquo images conditions (see signals plotsin Fig 3 and also ldquoRetrieval successrdquo analyses of Exp 2 inSupporting Information)
Recognition Processes in the Medial Frontal
Cortex and the Temporal Cortex
The third retrieval task involved scene-recognition judg-ments and was associated with the activation of the MFCand the MTG Moreover in Exp 1 we found a statisticaltrend in the hippocampus bilaterally an area that we pre-viously associated with scene-recognition judgment fromLTM (see next section about ldquoPossible correspondencesbetween retrieval from short- and long-term memoryrdquo)
The MFC was engaged both Exp 1 and 2 but was notactivated in the other two tasks thus a broad prefrontalcortexworking memory correspondence [Tsuchida andFellows 2009] seems insufficient to explain this task-specific effect The scene-recognition task in Exp 2 taxedboth recognition of ldquooldrdquo scenes and correct rejection ofldquonewrdquo scenes suggesting that the MFC here did not dis-tinguish between the correct rejection of new information(unseennew trials) and the recognition of old imagesOne candidate cognitive process that is common to both
r Neural Correlates of Immediate Episodic Retrieval r
r 2509 r
types of processes is feeling-of-knowing (FOK) FOKdepend on the cue specification content retrieval andappraisal of the retrieved memory fragments but are inde-pendent of the ultimate determination of content appropri-ateness [Schnyer et al 2005] Using imaging data duringsentence completion recognition Schnyer et al [2005]modeled a FOK-related directional path The path impli-cated the temporal cortex and the hippocampus andengaged the ventro-MFC at the end which is consistentwith our findings for the Object task implicating the MFCthe middle temporal cortex and at a lower statisticalthreshold the hippocampus
By contrast the activity in the MTG was larger forunseennew images than for seenold memory probes(Fig 4B) This suggests that new encoding may have con-tributed to the activation of the MTG during the retrievalphase of the Object task in Exp 1 The scene-recognitiontask required a lower precision in terms of retrievingdetails about the encoded material and could be solved bysimple recognition [Wixted and Squire 2010] Thus alter-natively larger activation for ldquonew vs oldrdquo stimuli mayrelate to some reduced activation associated with familiar-ity processes on old trials [see Henson et al 1999 whoreported reduced familiarity related activation in middletemporal gyri bilaterally and Henson and Rugg 2003 forreview] However we should point out that in Exp 2 theunseen images were extracted from edited-out parts of theoriginal clips and therefore contained the same objectcharacter depicted in the target images Therefore wedeem more likely that the relational association betweenobjects rather than their presenceabsence in the sceneshelped subjects to distinguish targets from foils We thussuggest that the activation of the MTG reflects processesutilizing the relational details among objects embedded inthe probe images for reaching the recognition decision[Diana et al 2007 Finke et al 2008 Hannula et al 2006Olson et al 2006] Future studies should consider theinclusion of some correlative behavioral measures to indexfamiliarity processes to elucidate further this issue More-over while the Temporal and Spatial tasks required mak-ing relative comparisons about aspects of the encodedvideo-clip the scene-recognition judgment merely requiredjudging whether the image presented at retrieval was pres-ent (or not) within the clip This may entail different proc-essing dynamics in the Object task compared with theother two tasks (cf also differences in retrieval times) Thelatter could be addressed using more subtle experimentalmanipulations for the scene-recognition judgment forexample by using digitally-modified foil images where asingle object in the scene has been modifiedremoved
Possible Correspondences Between Retrieval
from Short- and Long-Term Memory
The overlap between our current study and the previousLTM study using analogous tasks and materials lends
weight to the notion of scale-invariance in memory mecha-nisms [Brown et al 2007] as have been previously dem-onstrated in free recall tasks where the rate of item recallwas unvarying across recall span (day week year) andeven across from the past to the prospective future [May-lor et al 2001] Those previous behavioral findings implythat scale-invariance is a form of self-similarity in that theholistic pattern can exist at multiple levels of magnification[and even across different memory systems eg Maylor2002] Here our data affirmed the importance of the poste-rior parietal cortex in both immediate and long-termretrieval [Berryhill et al 2011 2007] and further character-ized its involvement in dealing with the spatial relationsof objects within scenes [Bricolo et al 2000 Buiatti et al2011] Our data also demonstrated such short-long-termcorrespondence exists in the precuneusangular gyrus fortemporal order retrieval (cf scene-chronology judgmenttask) In contrast the bilateral hippocampus and MFC acti-vations did not reach full statistical significance in theImMem LTM conjunction analysis At first sight thismay indicate that the scene-recognition task tapped intodistinct recognition-based mechanisms as a function ofretention delay However we deem this unlikely becausethe direct comparison between the two datasets did notshow any significant effect in these regions (see Support-ing Information Table 2) Indeed a more targeted test ask-ing the question of whether the hippocampus clusterspreviously found in LTM activated also in the currentimmediate retrieval experiments revealed significanteffects in both immediate retrieval experiments here (Exp1 x y z 5 224 29 225 P-FWE-corrlt 0001 Z 5 458 x yz 5 21 29 222 P-FWE-corr 5 0001 Z 5 451 and in Exp2 x y z 5 224 214 221 P-FWE-corr 5 0004 Z 5 396using the LTM clusters for small volume correction) Theseadditional tests point to a more general issue of sensitivitythat future studies may address using different methodo-logical approaches such as multivariate methods that canbe more sensitive than the standard univariate analysesused here Moreover the former would enable capturingtask-specific effects that are expressed as patterns of dis-tributed activation rather than local changes of activity asreported here [eg Jimura and Poldrack 2012]
Task Difficulty and Retrieval Success
We used several control analyses to show that the task-specific patterns of activation and the correspondencebetween the current data and our previous LTM study donot merely reflect some effect related to general task diffi-culty (see Supporting Information) First areas involved inoverall task difficulty should show progressively greateractivation with progressively more difficult tasks In Exp 1this would entail SgtTgtO However the patterns in theprecuneus and the angular gyrus (with TgtOgt S Fig 2A)and the MFC (Ogt SgtT Fig 4A) did not fit with simplepredictions based on overall task difficulty We also
r Kwok et al r
r 2510 r
assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r
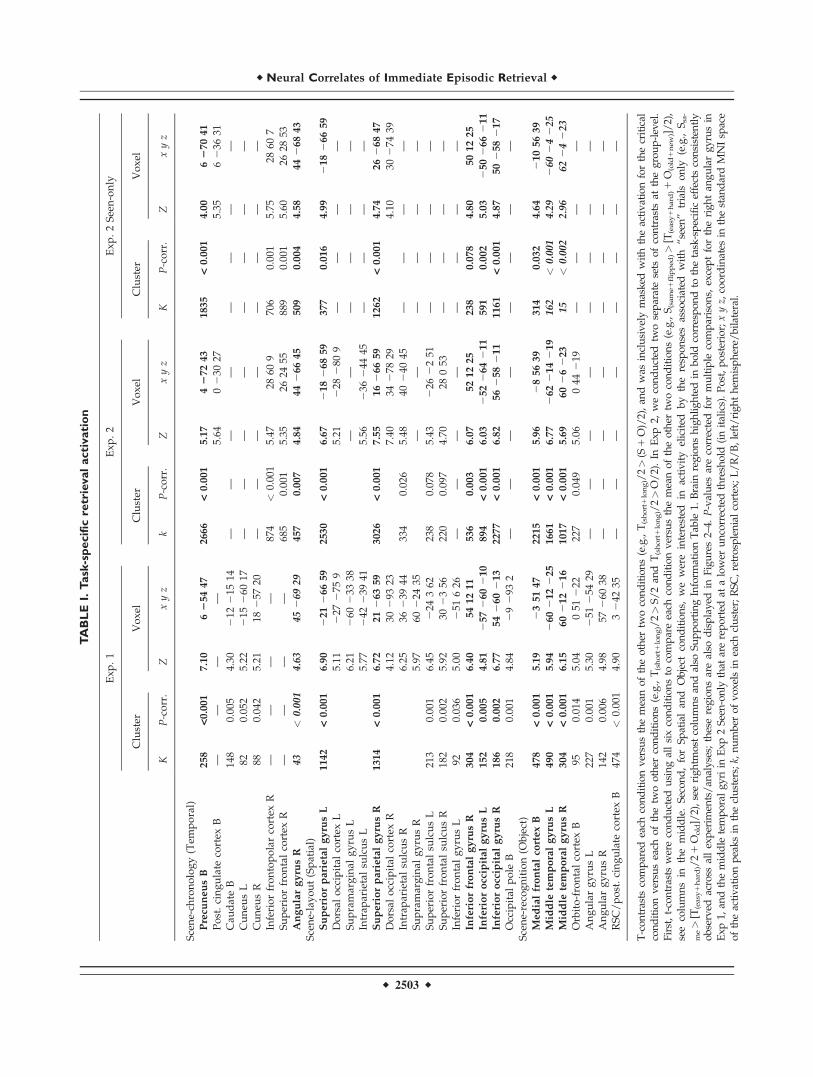
TA
BL
EI
Task
-sp
ecif
icre
trie
val
acti
vati
on
Ex
p
1E
xp
2
Ex
p
2S
een
-on
ly
Clu
ster
Vo
xel
Clu
ster
Vo
xel
Clu
ster
Vo
xel
KP
-co
rr
Zx
yz
kP
-co
rr
Zx
yz
KP
-co
rr
Zx
yz
Sce
ne-
chro
no
log
y(T
emp
ora
l)P
recu
neu
sB
258
lt00
01
71
06
254
47
2666
lt00
01
51
74
272
43
1835
lt00
01
40
06
270
41
Po
st
cin
gu
late
cort
exB
mdashmdash
mdashmdash
564
02
3027
535
62
3631
Cau
dat
eB
148
000
54
302
122
1514
mdashmdash
mdashmdash
mdashmdash
mdashmdash
Cu
neu
sL
820
052
522
215
260
17mdash
mdashmdash
mdashmdash
mdashmdash
mdashC
un
eus
R88
004
25
2118
257
20mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nto
po
lar
cort
exR
mdashmdash
mdashmdash
874
lt0
001
547
2860
970
60
001
575
2860
7S
up
erio
rfr
on
tal
cort
exR
mdashmdash
mdashmdash
685
000
15
3526
2455
889
000
15
6026
2853
An
gu
lar
gy
rus
R43
lt00
01
46
345
269
29
457
00
07
48
444
266
45
509
00
04
45
844
268
43
Sce
ne-
lay
ou
t(S
pat
ial)
Su
peri
or
pari
eta
lg
yru
sL
1142
lt00
01
69
02
21
266
59
2530
lt00
01
66
72
18
268
59
377
00
16
49
92
18
266
59
Do
rsal
occ
ipit
alco
rtex
L5
112
272
759
521
228
280
9mdash
mdashmdash
mdashS
up
ram
arg
inal
gy
rus
L6
212
602
3338
mdashmdash
mdashmdash
mdashmdash
Intr
apar
ieta
lsu
lcu
sL
577
242
239
415
562
362
4445
mdashmdash
mdashmdash
Su
peri
or
pari
eta
lg
yru
sR
1314
lt00
01
67
221
263
59
3026
lt00
01
75
516
266
59
1262
lt00
01
47
426
268
47
Do
rsal
occ
ipit
alco
rtex
R4
1230
293
237
4034
278
294
1030
274
39In
trap
arie
tal
sulc
us
R6
2536
239
4433
40
026
548
402
4045
mdashmdash
mdashmdash
Su
pra
mar
gin
alg
yru
sR
597
602
2435
mdashmdash
mdashmdash
mdashmdash
Su
per
ior
fro
nta
lsu
lcu
sL
213
000
16
452
243
6223
80
078
543
226
22
51mdash
mdashmdash
mdashS
up
erio
rfr
on
tal
sulc
us
R18
20
002
592
302
356
220
009
74
7028
053
mdashmdash
mdashmdash
Infe
rio
rfr
on
tal
gy
rus
L92
003
65
002
516
26mdash
mdashmdash
mdashmdash
mdashmdash
mdashIn
feri
or
fro
nta
lg
yru
sR
304
lt00
01
64
054
12
11
536
00
03
60
752
12
25
238
00
78
48
050
12
25
Infe
rio
ro
ccip
ital
gy
rus
L152
00
05
48
12
57
260
210
894
lt00
01
60
32
52
264
211
591
00
02
50
32
50
266
211
Infe
rio
ro
ccip
ital
gy
rus
R186
00
02
67
754
260
213
2277
lt00
01
68
256
258
211
1161
lt00
01
48
750
258
217
Occ
ipit
alp
ole
B21
80
001
484
29
293
2mdash
mdashmdash
mdashmdash
mdashmdash
mdashS
cen
e-re
cog
nit
ion
(Ob
ject
)M
ed
ial
fro
nta
lco
rtex
B478
lt00
01
51
92
351
47
2215
lt00
01
59
62
856
39
314
00
32
46
42
10
56
39
Mid
dle
tem
po
ral
gy
rus
L490
lt00
01
59
42
60
212
225
1661
lt00
01
67
72
62
214
219
162
lt00
01
42
92
60
24
225
Mid
dle
tem
po
ral
gy
rus
R304
lt00
01
61
560
212
216
1017
lt00
01
56
960
26
223
15
lt00
02
29
662
24
223
Orb
ito
-fro
nta
lco
rtex
B95
001
45
040
512
2222
70
049
506
044
219
mdashmdash
mdashmdash
An
gu
lar
gy
rus
L22
70
001
530
251
254
29mdash
mdashmdash
mdashmdash
mdashmdash
mdashA
ng
ula
rg
yru
sR
142
000
64
9857
260
38mdash
mdashmdash
mdashmdash
mdashmdash
mdashR
SC
p
ost
ci
ng
ula
teco
rtex
B47
4lt
000
14
903
242
35mdash
mdashmdash
mdashmdash
mdashmdash
mdash
T-c
on
tras
tsco
mp
ared
each
con
dit
ion
ver
sus
the
mea
no
fth
eo
ther
two
con
dit
ion
s(e
g
T(s
ho
rt1
lon
g)
2gt
(S1
O)
2)
and
was
incl
usi
vel
ym
ask
edw
ith
the
acti
vat
ion
for
the
crit
ical
con
dit
ion
ver
sus
each
of
the
two
oth
erco
nd
itio
ns
(eg
T
(sh
ort
1lo
ng
)2gt
S2
and
T(s
ho
rt1
lon
g)
2gt
O
2)
InE
xp2
we
con
du
cted
two
sep
arat
ese
tso
fco
ntr
asts
atth
eg
rou
p-l
evel
F
irst
t-
con
tras
tsw
ere
con
du
cted
usi
ng
all
six
con
dit
ion
sto
com
par
eea
chco
nd
itio
nv
ersu
sth
em
ean
of
the
oth
ertw
oco
nd
itio
ns
(eg
S (
sam
e1
flip
ped
)gt
[T(e
asy
1h
ard
)1
O(o
ld1
new
)]
2)
see
colu
mn
sin
the
mid
dle
Se
con
d
for
Spat
ial
and
Ob
ject
con
dit
ion
sw
ew
ere
inte
rest
edin
acti
vit
yel
icit
edb
yth
ere
spo
nse
sas
soci
ated
wit
hldquos
een
rdquotr
ials
on
ly(e
g
S sa-
megt
[T(e
asy
1h
ard
)2
1O
old
]2)
se
eri
gh
tmo
stco
lum
ns
and
also
Sup
po
rtin
gIn
form
atio
nT
able
1B
rain
reg
ion
sh
igh
lig
hte
din
bo
ldco
rres
po
nd
toth
eta
sk-s
pec
ific
effe
cts
con
sist
entl
yo
bse
rved
acro
ssal
lex
per
imen
ts
anal
yse
sth
ese
reg
ion
sar
eal
sod
isp
lay
edin
Fig
ure
s2ndash
4P
-val
ues
are
corr
ecte
dfo
rm
ult
iple
com
par
iso
ns
exce
pt
for
the
rig
ht
ang
ula
rg
yru
sin
Exp
1an
dth
em
idd
lete
mp
ora
lg
yri
inE
xp2
Seen
-on
lyth
atar
ere
po
rted
ata
low
eru
nco
rrec
ted
thre
sho
ld(i
nit
alic
s)
Po
st
po
ster
ior
xy
zco
ord
inat
esin
the
stan
dar
dM
NI
spac
eo
fth
eac
tiv
atio
np
eak
sin
the
clu
ster
sk
nu
mb
ero
fv
oxe
lsin
each
clu
ster
R
SC
retr
osp
len
ial
cort
ex
L
R
B
left
ri
gh
th
emis
ph
ere
bil
ater
al
r Neural Correlates of Immediate Episodic Retrieval r
r 2503 r
by comparing Tshort vs Tlong trials In previous studiesusing LTM protocols we found that temporal distance fur-ther modulated activity in the precuneus [Kwok et al2012 2014 see also St Jacques et al 2008] Here this con-trast did not reveal any significant effect This is in con-trast to the significant behavioral cost of retrieving Tshort
vs Tlong temporal distances (see behavioral results andFig 1B plot on the left) that was analogous to our previ-ous LTM protocol Similar behavioral patterns have beenobserved in many other short-term studies [Hacker 1980Konishi et al 2002 Milner et al 1991 Muter 1979] yet toour knowledge there does not seem to exists a clear con-
sensus on the neural correlates subserving the temporaldistance effect separating two temporally very nearbyeventsitems
Exp 2 In Exp 2 we replicated the effect of the temporalretrieval task in the precuneus and found a fully signifi-cant effect in the right angular gyrus (see Fig 2 clusters inredmagenta) In addition in Exp 2 there was a significantactivation of the right superior frontal cortex and the rightinferior frontopolar cortex that were not found in Exp 1(Table I) The involvement of the precuneus and the rightangular gyrus was further confirmed using only trials
Figure 2
Clusters of brain activation and signal plots for the scene-
chronology judgment (Temporal task) Clusters of activation and
signal plots for (A) the precuneus and (B) the right angular
gyrus These regions activated selectively during the temporal
tasks in Exp 1 (in blue) Exp 2 (in red overlap between Exp 1
and 2 shown in magenta) The activation selective for the Tem-
poral task was computed with the contrast T(short1long)
2gt (S 1 O)2 inclusively masked with T(short1long)2gt S2 and
with T(short1long)2gtO2 The involvement of the precuneus and
the right angular gyrus was further confirmed using only ldquoseen
oldrdquo images (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 40 voxels Effect sizes are mean adjusted (sum to
zero) and expressed in arbitrary units (au 6 90 CI) LR left
right PreC precuneus AnG angular gyrus
r Kwok et al r
r 2504 r
including seen images (cf ldquoExp 2 Seen- onlyrdquo Table I)The signal plots on Figure 2 (panels on the right) show theactivity of the precuneus and the right angular gyrus forall the conditions of Exp 2 Both in the precuneus and theright angular gyrus the average activity in the temporaltask [T(easy1hard)2] were larger than the Ssame and theOold conditions Notably Exp 2 included the presentationof a single image and that the participants were asked tojudge the absolute temporal position of the probe ratherthan the relative temporal order Despite this importantchange of how temporal memory was probed the findingof analogous patterns of activation across both experi-ments highlights the link between immediate retrieval oftemporal information and activation of the precuneus andangular gyrus
Scene-layout judgment (Spatial task)
Exp 1 The contrast concerning the retrieval of the spatiallayout revealed activation in the superior parietal cortexbilaterally with the cluster extending to the intraparietalsulcus and the supramarginal gyrus This contrast alsoshowed activation of the superior frontal sulci the inferiorfrontal gyri plus the inferior occipital cortex (see Table Iand Fig 3 clusters displayed in bluemagenta signalsplots on the left side)
Exp 2 The same comparison in Exp 2 replicated theresults of Exp 1 showing activation in the superior parie-tal cortex bilaterally the superior frontal sulci the inferiorand dorsal occipital cortices and the right inferior frontalgyrus (Fig 3 clusters in redmagenta) When we consid-ered only the conditions including the seenold images(ldquoExp 2 Seen- onlyrdquo) we replicated the effects in the supe-rior parietal gyri the inferior occipital gyri and the rightinferior frontal gyrus (see Table I and Fig 3 signal plotson the right) The activation in the intraparietal sulci andthe superior frontal sulci did not reach statistical signifi-cance Inspection of the signal plots indicated that in theseareas there was less activation in the Ssame than Sflipped tri-als (cf bar 3 vs bar 4 signal plots of the superior fontalsulcus Fig 3A plot on the right) Nonetheless the activityin the Ssame condition was larger than those in the T andO conditions (cf bar 3 vs bars 1ndash2 and 5ndash6)
Scene-recognition judgment (Object task)
Exp 1 The Object task was associated with the activationof the medial frontal cortex (MFC) and the orbitofrontalcortex (but the latter did not fully replicate in Exp 2 seeTable I and Fig 4 clusters bluemagenta and signal plotson the left) together with bilateral activation in the middletemporal gyri the retrosplenialposterior cingulate gyrusand a portion of the angular gyrus The same contrast alsorevealed an activation of the hippocampus bilaterally Theactivation of the hippocampus was just below statisticalsignificance (P-FWE-corr 5 0052 Z 5 458 at x y z 5 2126 219 and P-FWE-corr 5 0077 Z 5 458 at x y z 5 224
29 225 considering the whole brain as the volume ofinterest) and it is reported here because the same effectwas previously found in the LTM dataset [Kwok et al2012 see also Discussion Section]
Exp 2 An almost identical pattern of activation wasfound in Exp 2 when we considered all the conditionsO(old1new)gt (T(easy1hard) 1 S(same1flipped))2 Significanteffects were found in the middle temporal gyri and alarge cluster including the medial frontal gyrus extendingventrally into the orbital-frontal cortex (Fig 4 clusters inredmagenta) However when we considered only the tri-als including seenold-images [Ooldgt (T(easy1hard)2 1 Ssame)2] the Object task was found to significantlyactivate only the MFC (Table I rightmost columns) indi-cating that this region was not affected by incidentalencoding
In summary across all three tasks the brain activationwas found to be similar across Exp 1 and Exp 2 Thescene-chronology Temporal task activated the precuneusand the right angular gyrus the scene-layout Spatial taskwas associated with activation of the posteriorsuperiorparietal cortex and the right inferior frontal gyrus whilethe scene-recognition Object task activated the MFC andthe middle temporal gurus (see Table I areas highlightedin bold) The control analysis of Exp 2 that consideredonly trials including seenold images confirmed thesetask-specific effects indicating that the pattern of dissocia-tionmdashand in particular for the Spatial and Object tasksmdashwas not merely driven by the unseennew images duringthe retrieval phase
Functional Neuroimaging Correspondence with
An Analogous LTM Dataset
We directly compared the task-specific dissociationobserved in the current dataset (Exp 1) with our previousLTM study that included the same task-specific retrievaltasks but after a much longer retention delay [ie 24 hbetween encoding and retrieval Kwok et al 2012] Com-mon across the two data sets the conjunction analysesrevealed an activation cluster in the precuneus for theTemporal task (Fig 5A and Table II) and the dorsal fron-toparietal network plus right inferior frontal gyrus andright occipital cortex for the Spatial task (Fig 5B and TableII) These formal tests show that the Temporal and Spatialtask-specific dissociation observed here for immediateretrieval matched well with the patterns observed duringretrieval from LTM By contrast for the Object task over-lapping effects in the MFC and in the hippocampus weredetected only at a more lenient uncorrected threshold ofP-uncorr 5 0001 In the hippocampus the lack of a fullysignificant ldquocommonrdquo effect across datasets was due to thefact that in the current Exp 1 the contrast comparing theObject task versus the two other tasks did not survive cor-rection for multiple comparisons considering the wholebrain as the volume of interest (P-corr 5 0052 and 0077
r Neural Correlates of Immediate Episodic Retrieval r
r 2505 r
Figure 3
Clusters of brain activation and signal plots for the Scene-layout
judgment (Spatial task) Clusters of activation and signal plots for
(A) the superior frontal cortex and the superior parietal cortex
bilaterally and (B) the right inferior occipital cortex and the right
inferior frontal cortex These regions activated for the Spatial task
in Exp 1 (blue) and in Exp 2 (red overlap between Exp 1 and Exp 2
in magenta) The activation selective for the Spatial task was com-
puted with the contrast Sgt [T(short1long)2 1 O]2 inclusively
masked with SgtT(short1long)2 and with SgtO The effects in the
superior parietal gyri the inferior occipital gyri and the right inferior
frontal gyrus were replicated when considering only sameold
images in Exp 2 (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 80 voxels Effect sizes are mean adjusted (sum to zero)
and are expressed in arbitrary units (au 6 90 CI) LR leftright
SFS superior frontal sulcus SPG superior parietal gyrus IFG infe-
rior frontal gyrus IOG inferior occipital gyrus
in the left and right hemispheres respectively cf above)Indeed the formal test assessing differences between thetwo datasets (ie ldquoTask 3 Experimentrdquo interactions seeSupporting Information Table 2) did not reveal any signifi-cant difference between LTM and immediate retrieval inthe hippocampus consistent with this region beinginvolved in the scene-recognition judgment irrespective ofthe retention delay (see also Discussion Section)
DISCUSSION
We have identified distinct sets of brain areas thatshowed task-specific activation during immediate retrievalfollowing the encoding of dynamic cinematic material Theprecuneus and the right angular gyrus were activatedwhile retrieving the temporal order of events (scene-chro-nology judgment) the superior parietal cortex wasrecruited during spatial related recall (scene-layout
Figure 4
Clusters of brain activation and signal plots for the Scene-
recognition judgment (Object task) Clusters of activation and
signal plots for (A) the MFC and (B) the MTG These regions
activated for the Object task in Exp 1 (blue) and in Exp 2 (red
overlap in magenta) The activation selective for the Object task
was computed with the contrast Ogt [T(short1long)2 1 S]2
inclusively masked with OgtT(short1long)2 and with Ogt S The
signal plots refer to the left hemisphere but analogous patterns
of activation were found in the right hemisphere (Table I) In
Exp 2 the task-specific effect of the Object task in the MFC
was replicated when considering only the sameold images
(Table I rightmost columns) Inspection of the signal plot for the
left MTG revealed less activity in the Oold than Onew trials (plots
on the right bars 5 and 6) although the Oold trials showed
numerically larger parameter estimates than the T and S condi-
tions (see p-uncorr reported in italics in Table I rightmost col-
umns) Activation clusters were estimated and displayed at P-
unc 5 0001 thresholded at a minimum of 80 voxels Effect sizes
are mean adjusted (sum to zero) and expressed in arbitrary
units (au 6 90 CI) LR leftright MTG middle temporal
gyrus MFC medial frontal cortex
r Neural Correlates of Immediate Episodic Retrieval r
r 2507 r
judgment) whereas activity in the MFC and the middletemporal gyrus (MTG) reflected processes associated withscene-recognition judgments These task-specific effectsoccurred independently of whether subjects were asked tojudge seenold or unseennew stimuli (cf ldquoSeen-onlyimagesrdquo contrasts in Exp 2) and independently ofretrieval success and general task difficulty (see Support-ing Information) With respect to the hypothesized corre-spondence between immediate retrieval and retrieval fromLTM we found that these task-specific dissociationsare independent of the informationrsquos mnemonic history(24 h vs 2 s retention ie differing by 43200 times inmagnitude)
One important qualifier for the following discussion isthat memory retrieval in all our tasks depended minimallyon memory buffers compared to other simpler itemsdis-plays that are typically used in ldquostandardrdquo short-termworking memory tasks [Shallice and Cooper 2011 Shiffrin1973] In each trial the number of items presented during
encoding was clearly well beyond the 4 6 1 items stipulatedby classical STM models [Cowan 2001] While memories ofsome materials (eg words spatial positions simple visualdisplays) are supported by limited-capacity buffers [Badde-ley and Hitch 1974] memory for other categories [egunknown faces and scenes see Shiffrin 1973] will tap intoLTM when the task requires the encoding of two or moreitems [Shallice and Cooper 2011] For this reason werestrict our discussion on the tripartite task-specific dissoci-ations with reference to highly complex naturalisticmaterial
Precuneus and Inferior Parietal Cortex Support
Temporal Order Recollection
The scene-chronology judgment task activated the pre-cuneus in both Exp 1 and 2 despite the considerablechanges in the trial structure for probing the retrieval oftemporal information (ie relative temporal order betweentwo probes in Exp 1 vs absolute temporal position of asingle probe in Exp 2) The chronology-judgment taskrequired retrieving the event sequences of many complex
Figure 5
Task-specific retrieval activation common in studies after imme-
diate delays and a longer delay Activation for the LTM study
[24 h Kwok et al 2012] is depicted in green and activation
for the current immediate retrieval study (data from current
Exp 1) depicted in red Activation clusters for both studies were
estimated and displayed at P-unc 5 0001
TABLE II Common memory retrieval activation
ImMem LTM common effects
Cluster Voxel
k P-corr Z x y z
Scene-chronology (Temporal)Precuneus B 112 0011 472 3 257 47Angular gyrus R 98 lt 0001 400 54 254 17
Scene-layout (Spatial)Superior parietal gyrus L 206 0001 595 221 269 56Supramarginal gyrus L 129 0008 346 260 236 44Intraparietal sulcus L 487 236 245 41Superior parietal gyrus R 651 lt 0001 520 24 263 59Dorsal occipital cortex R 372 30 275 29Intraparietal sulcus R 520 42 239 47Supramarginal gyrus R 482 57 233 53Superior frontal sulcus L 100 0022 482 224 3 56Superior frontal sulcus R 145 0004 481 30 0 53Inferior frontal gyrus R 95 0027 475 54 9 26Inferior occipital gyrus R 98 0024 515 54 257 210
Task-specific activation after immediate delays (current Exp 1ImMem) and a previous study that used a long delay betweenencoding and retrieval [24 h LTM Kwok et al 2012] Statisticalthreshold were set at P-FWE-corr 5 005 (cluster-level) consider-ing the whole brain as the volume of interest Note that theldquobetween-studiesrdquo overlap activation in the right angular gyruswas found at a lower uncorrected threshold (in italics) but impor-tantly within this area no differences or interactions betweenImMem and LTM studies were found (see differential effectsbetween the two studies in Supporting Information Table 2) LRB leftright hemispherebilateral
r Kwok et al r
r 2508 r
stimuli (scenes) that entail high-level conceptualhierarchi-cal relationships between them [Swallow et al 2011 Zackset al 2001] The precuneus was previously found to acti-vate in our LTM protocol that used the same temporalorder judgment [Kwok et al 2012 and see also results ofthe conjunction analyses] The current finding of precunealactivation using very different (much shorter) delays fitswell with the view that these processes are not strictlyassociated with retrieval following long retention delayssuggesting that this medial parietal region is implicatedduring temporal order retrieval irrespective of delay (Fig5A and Table II) Specifically we link the effect of chrono-logical retrieval in the precuneus with processes involvingan active reconstruction of past events [see also St Jacqueset al 2008] and possibly memory-related imagery[Fletcher et al 1995 Ishai et al 2000] These processeswould characterize temporal order judgments using com-plex naturalistic material but not order-judgment withsimpler stimulus material which previous studies haveassociated with other brain regions [eg middleprefron-tal areas Suzuki et al 2002]
Together with the precuneus temporal-related judgmentalso gave rise to activation of the right angular gyrus(AG) One possible interpretation relates the inferior parie-tal cortex to the engagement of attentional processes[Majerus et al 2007 2010] The scene-chronology judg-ment of Exp 2 provided us with a further qualifier to thiskind of temporal-related attention Although the Temporaltask in Exp 2 did not require any explicit temporal orderjudgment (only classifying a scene into either the first orsecond half of the movie) an accurate decision still neces-sitated an evaluation of the target scene with many othersegments of the movie This operation is fundamentallydifferent from the evaluation implicated in the scene-layout judgment which required decisions on comparingbetween a single scene and its own correspondentencoded snapshot The latter involves visuospatial analysisof the configuration within scenes The AG may subservedifferent types of attention among which one of them islinked with memory search operations [see Majerus et al2010 2006 also showing coactivation of these two regionsduring order retrieval of simple stimuli after short delays]
Superior Parietal Cortex Evaluates Spatial
Details Against Memory Representation
The second pattern consistently found across experi-ments (Exp 1 and Exp 2 see Fig 3) and across studieswith different retention delays (conjunction betweenimmediate retrieval and LTM studies see Fig 5B andTable II) was the activation of the superior parietal lobule(SPL) during the scene-layout judgment Previous studiesshowed sustained activation in the parietal cortex duringworking memory delays [Courtney et al 1998] and thatactivity there correlates with memory load [Todd andMaroi 2004] Here the amount of information contained
in the clips was equated in the three tasks especially inExp 2 where we counterbalanced the assignment of mov-ies across tasks The subjects also could not anticipatewhat specific type of information would be tested atretrieval and needed to keep rehearsing different aspectsof the clips Because of this we deem unlikely that theactivation of SPL that here was specific for the scene-layout tasks reflected general rehearsal processes [Magenet al 2009] andor updating operations [Bledowski et al2009 Leung et al 2007]
Instead we suggest a role for the SPL in mediating aninterface between the internal memory representation andexternal information It has been proposed that the supe-rior parietal cortex is engaged in allocating top-downattention to orient to aspects of the available visual inputand on information retrieved from memory [Ishai et al2002 Lepsien and Nobre 2007 Summerfield et al 2006]In line with this we found SPL activation when the sub-jects had to make decisions by comparing between theirmemory of the encoded clips and the scene stimuli pre-sented at retrieval This procedure might act on somevisuospatial storage which is held by the lateral occipitalcortex and mediated by the intraparietal sulcus (Fig 3B)The immediate memory task here was analogous to theSpatial task of our previous LTM study [Kwok et al2012] which also required matchingcomparing the avail-able external visual input and internal information storedin memory [see also Nobre et al 2004] This interfaceinvolves a range of top-down processes such as monitor-ing and verification [Cabeza et al 2008 Ciaramelli et al2008] which are independent of the probe imagersquos mne-monic status as shown by the equated effect sizes forldquosamerdquo and ldquoflippedrdquo images conditions (see signals plotsin Fig 3 and also ldquoRetrieval successrdquo analyses of Exp 2 inSupporting Information)
Recognition Processes in the Medial Frontal
Cortex and the Temporal Cortex
The third retrieval task involved scene-recognition judg-ments and was associated with the activation of the MFCand the MTG Moreover in Exp 1 we found a statisticaltrend in the hippocampus bilaterally an area that we pre-viously associated with scene-recognition judgment fromLTM (see next section about ldquoPossible correspondencesbetween retrieval from short- and long-term memoryrdquo)
The MFC was engaged both Exp 1 and 2 but was notactivated in the other two tasks thus a broad prefrontalcortexworking memory correspondence [Tsuchida andFellows 2009] seems insufficient to explain this task-specific effect The scene-recognition task in Exp 2 taxedboth recognition of ldquooldrdquo scenes and correct rejection ofldquonewrdquo scenes suggesting that the MFC here did not dis-tinguish between the correct rejection of new information(unseennew trials) and the recognition of old imagesOne candidate cognitive process that is common to both
r Neural Correlates of Immediate Episodic Retrieval r
r 2509 r
types of processes is feeling-of-knowing (FOK) FOKdepend on the cue specification content retrieval andappraisal of the retrieved memory fragments but are inde-pendent of the ultimate determination of content appropri-ateness [Schnyer et al 2005] Using imaging data duringsentence completion recognition Schnyer et al [2005]modeled a FOK-related directional path The path impli-cated the temporal cortex and the hippocampus andengaged the ventro-MFC at the end which is consistentwith our findings for the Object task implicating the MFCthe middle temporal cortex and at a lower statisticalthreshold the hippocampus
By contrast the activity in the MTG was larger forunseennew images than for seenold memory probes(Fig 4B) This suggests that new encoding may have con-tributed to the activation of the MTG during the retrievalphase of the Object task in Exp 1 The scene-recognitiontask required a lower precision in terms of retrievingdetails about the encoded material and could be solved bysimple recognition [Wixted and Squire 2010] Thus alter-natively larger activation for ldquonew vs oldrdquo stimuli mayrelate to some reduced activation associated with familiar-ity processes on old trials [see Henson et al 1999 whoreported reduced familiarity related activation in middletemporal gyri bilaterally and Henson and Rugg 2003 forreview] However we should point out that in Exp 2 theunseen images were extracted from edited-out parts of theoriginal clips and therefore contained the same objectcharacter depicted in the target images Therefore wedeem more likely that the relational association betweenobjects rather than their presenceabsence in the sceneshelped subjects to distinguish targets from foils We thussuggest that the activation of the MTG reflects processesutilizing the relational details among objects embedded inthe probe images for reaching the recognition decision[Diana et al 2007 Finke et al 2008 Hannula et al 2006Olson et al 2006] Future studies should consider theinclusion of some correlative behavioral measures to indexfamiliarity processes to elucidate further this issue More-over while the Temporal and Spatial tasks required mak-ing relative comparisons about aspects of the encodedvideo-clip the scene-recognition judgment merely requiredjudging whether the image presented at retrieval was pres-ent (or not) within the clip This may entail different proc-essing dynamics in the Object task compared with theother two tasks (cf also differences in retrieval times) Thelatter could be addressed using more subtle experimentalmanipulations for the scene-recognition judgment forexample by using digitally-modified foil images where asingle object in the scene has been modifiedremoved
Possible Correspondences Between Retrieval
from Short- and Long-Term Memory
The overlap between our current study and the previousLTM study using analogous tasks and materials lends
weight to the notion of scale-invariance in memory mecha-nisms [Brown et al 2007] as have been previously dem-onstrated in free recall tasks where the rate of item recallwas unvarying across recall span (day week year) andeven across from the past to the prospective future [May-lor et al 2001] Those previous behavioral findings implythat scale-invariance is a form of self-similarity in that theholistic pattern can exist at multiple levels of magnification[and even across different memory systems eg Maylor2002] Here our data affirmed the importance of the poste-rior parietal cortex in both immediate and long-termretrieval [Berryhill et al 2011 2007] and further character-ized its involvement in dealing with the spatial relationsof objects within scenes [Bricolo et al 2000 Buiatti et al2011] Our data also demonstrated such short-long-termcorrespondence exists in the precuneusangular gyrus fortemporal order retrieval (cf scene-chronology judgmenttask) In contrast the bilateral hippocampus and MFC acti-vations did not reach full statistical significance in theImMem LTM conjunction analysis At first sight thismay indicate that the scene-recognition task tapped intodistinct recognition-based mechanisms as a function ofretention delay However we deem this unlikely becausethe direct comparison between the two datasets did notshow any significant effect in these regions (see Support-ing Information Table 2) Indeed a more targeted test ask-ing the question of whether the hippocampus clusterspreviously found in LTM activated also in the currentimmediate retrieval experiments revealed significanteffects in both immediate retrieval experiments here (Exp1 x y z 5 224 29 225 P-FWE-corrlt 0001 Z 5 458 x yz 5 21 29 222 P-FWE-corr 5 0001 Z 5 451 and in Exp2 x y z 5 224 214 221 P-FWE-corr 5 0004 Z 5 396using the LTM clusters for small volume correction) Theseadditional tests point to a more general issue of sensitivitythat future studies may address using different methodo-logical approaches such as multivariate methods that canbe more sensitive than the standard univariate analysesused here Moreover the former would enable capturingtask-specific effects that are expressed as patterns of dis-tributed activation rather than local changes of activity asreported here [eg Jimura and Poldrack 2012]
Task Difficulty and Retrieval Success
We used several control analyses to show that the task-specific patterns of activation and the correspondencebetween the current data and our previous LTM study donot merely reflect some effect related to general task diffi-culty (see Supporting Information) First areas involved inoverall task difficulty should show progressively greateractivation with progressively more difficult tasks In Exp 1this would entail SgtTgtO However the patterns in theprecuneus and the angular gyrus (with TgtOgt S Fig 2A)and the MFC (Ogt SgtT Fig 4A) did not fit with simplepredictions based on overall task difficulty We also
r Kwok et al r
r 2510 r
assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r

by comparing Tshort vs Tlong trials In previous studiesusing LTM protocols we found that temporal distance fur-ther modulated activity in the precuneus [Kwok et al2012 2014 see also St Jacques et al 2008] Here this con-trast did not reveal any significant effect This is in con-trast to the significant behavioral cost of retrieving Tshort
vs Tlong temporal distances (see behavioral results andFig 1B plot on the left) that was analogous to our previ-ous LTM protocol Similar behavioral patterns have beenobserved in many other short-term studies [Hacker 1980Konishi et al 2002 Milner et al 1991 Muter 1979] yet toour knowledge there does not seem to exists a clear con-
sensus on the neural correlates subserving the temporaldistance effect separating two temporally very nearbyeventsitems
Exp 2 In Exp 2 we replicated the effect of the temporalretrieval task in the precuneus and found a fully signifi-cant effect in the right angular gyrus (see Fig 2 clusters inredmagenta) In addition in Exp 2 there was a significantactivation of the right superior frontal cortex and the rightinferior frontopolar cortex that were not found in Exp 1(Table I) The involvement of the precuneus and the rightangular gyrus was further confirmed using only trials
Figure 2
Clusters of brain activation and signal plots for the scene-
chronology judgment (Temporal task) Clusters of activation and
signal plots for (A) the precuneus and (B) the right angular
gyrus These regions activated selectively during the temporal
tasks in Exp 1 (in blue) Exp 2 (in red overlap between Exp 1
and 2 shown in magenta) The activation selective for the Tem-
poral task was computed with the contrast T(short1long)
2gt (S 1 O)2 inclusively masked with T(short1long)2gt S2 and
with T(short1long)2gtO2 The involvement of the precuneus and
the right angular gyrus was further confirmed using only ldquoseen
oldrdquo images (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 40 voxels Effect sizes are mean adjusted (sum to
zero) and expressed in arbitrary units (au 6 90 CI) LR left
right PreC precuneus AnG angular gyrus
r Kwok et al r
r 2504 r
including seen images (cf ldquoExp 2 Seen- onlyrdquo Table I)The signal plots on Figure 2 (panels on the right) show theactivity of the precuneus and the right angular gyrus forall the conditions of Exp 2 Both in the precuneus and theright angular gyrus the average activity in the temporaltask [T(easy1hard)2] were larger than the Ssame and theOold conditions Notably Exp 2 included the presentationof a single image and that the participants were asked tojudge the absolute temporal position of the probe ratherthan the relative temporal order Despite this importantchange of how temporal memory was probed the findingof analogous patterns of activation across both experi-ments highlights the link between immediate retrieval oftemporal information and activation of the precuneus andangular gyrus
Scene-layout judgment (Spatial task)
Exp 1 The contrast concerning the retrieval of the spatiallayout revealed activation in the superior parietal cortexbilaterally with the cluster extending to the intraparietalsulcus and the supramarginal gyrus This contrast alsoshowed activation of the superior frontal sulci the inferiorfrontal gyri plus the inferior occipital cortex (see Table Iand Fig 3 clusters displayed in bluemagenta signalsplots on the left side)
Exp 2 The same comparison in Exp 2 replicated theresults of Exp 1 showing activation in the superior parie-tal cortex bilaterally the superior frontal sulci the inferiorand dorsal occipital cortices and the right inferior frontalgyrus (Fig 3 clusters in redmagenta) When we consid-ered only the conditions including the seenold images(ldquoExp 2 Seen- onlyrdquo) we replicated the effects in the supe-rior parietal gyri the inferior occipital gyri and the rightinferior frontal gyrus (see Table I and Fig 3 signal plotson the right) The activation in the intraparietal sulci andthe superior frontal sulci did not reach statistical signifi-cance Inspection of the signal plots indicated that in theseareas there was less activation in the Ssame than Sflipped tri-als (cf bar 3 vs bar 4 signal plots of the superior fontalsulcus Fig 3A plot on the right) Nonetheless the activityin the Ssame condition was larger than those in the T andO conditions (cf bar 3 vs bars 1ndash2 and 5ndash6)
Scene-recognition judgment (Object task)
Exp 1 The Object task was associated with the activationof the medial frontal cortex (MFC) and the orbitofrontalcortex (but the latter did not fully replicate in Exp 2 seeTable I and Fig 4 clusters bluemagenta and signal plotson the left) together with bilateral activation in the middletemporal gyri the retrosplenialposterior cingulate gyrusand a portion of the angular gyrus The same contrast alsorevealed an activation of the hippocampus bilaterally Theactivation of the hippocampus was just below statisticalsignificance (P-FWE-corr 5 0052 Z 5 458 at x y z 5 2126 219 and P-FWE-corr 5 0077 Z 5 458 at x y z 5 224
29 225 considering the whole brain as the volume ofinterest) and it is reported here because the same effectwas previously found in the LTM dataset [Kwok et al2012 see also Discussion Section]
Exp 2 An almost identical pattern of activation wasfound in Exp 2 when we considered all the conditionsO(old1new)gt (T(easy1hard) 1 S(same1flipped))2 Significanteffects were found in the middle temporal gyri and alarge cluster including the medial frontal gyrus extendingventrally into the orbital-frontal cortex (Fig 4 clusters inredmagenta) However when we considered only the tri-als including seenold-images [Ooldgt (T(easy1hard)2 1 Ssame)2] the Object task was found to significantlyactivate only the MFC (Table I rightmost columns) indi-cating that this region was not affected by incidentalencoding
In summary across all three tasks the brain activationwas found to be similar across Exp 1 and Exp 2 Thescene-chronology Temporal task activated the precuneusand the right angular gyrus the scene-layout Spatial taskwas associated with activation of the posteriorsuperiorparietal cortex and the right inferior frontal gyrus whilethe scene-recognition Object task activated the MFC andthe middle temporal gurus (see Table I areas highlightedin bold) The control analysis of Exp 2 that consideredonly trials including seenold images confirmed thesetask-specific effects indicating that the pattern of dissocia-tionmdashand in particular for the Spatial and Object tasksmdashwas not merely driven by the unseennew images duringthe retrieval phase
Functional Neuroimaging Correspondence with
An Analogous LTM Dataset
We directly compared the task-specific dissociationobserved in the current dataset (Exp 1) with our previousLTM study that included the same task-specific retrievaltasks but after a much longer retention delay [ie 24 hbetween encoding and retrieval Kwok et al 2012] Com-mon across the two data sets the conjunction analysesrevealed an activation cluster in the precuneus for theTemporal task (Fig 5A and Table II) and the dorsal fron-toparietal network plus right inferior frontal gyrus andright occipital cortex for the Spatial task (Fig 5B and TableII) These formal tests show that the Temporal and Spatialtask-specific dissociation observed here for immediateretrieval matched well with the patterns observed duringretrieval from LTM By contrast for the Object task over-lapping effects in the MFC and in the hippocampus weredetected only at a more lenient uncorrected threshold ofP-uncorr 5 0001 In the hippocampus the lack of a fullysignificant ldquocommonrdquo effect across datasets was due to thefact that in the current Exp 1 the contrast comparing theObject task versus the two other tasks did not survive cor-rection for multiple comparisons considering the wholebrain as the volume of interest (P-corr 5 0052 and 0077
r Neural Correlates of Immediate Episodic Retrieval r
r 2505 r
Figure 3
Clusters of brain activation and signal plots for the Scene-layout
judgment (Spatial task) Clusters of activation and signal plots for
(A) the superior frontal cortex and the superior parietal cortex
bilaterally and (B) the right inferior occipital cortex and the right
inferior frontal cortex These regions activated for the Spatial task
in Exp 1 (blue) and in Exp 2 (red overlap between Exp 1 and Exp 2
in magenta) The activation selective for the Spatial task was com-
puted with the contrast Sgt [T(short1long)2 1 O]2 inclusively
masked with SgtT(short1long)2 and with SgtO The effects in the
superior parietal gyri the inferior occipital gyri and the right inferior
frontal gyrus were replicated when considering only sameold
images in Exp 2 (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 80 voxels Effect sizes are mean adjusted (sum to zero)
and are expressed in arbitrary units (au 6 90 CI) LR leftright
SFS superior frontal sulcus SPG superior parietal gyrus IFG infe-
rior frontal gyrus IOG inferior occipital gyrus
in the left and right hemispheres respectively cf above)Indeed the formal test assessing differences between thetwo datasets (ie ldquoTask 3 Experimentrdquo interactions seeSupporting Information Table 2) did not reveal any signifi-cant difference between LTM and immediate retrieval inthe hippocampus consistent with this region beinginvolved in the scene-recognition judgment irrespective ofthe retention delay (see also Discussion Section)
DISCUSSION
We have identified distinct sets of brain areas thatshowed task-specific activation during immediate retrievalfollowing the encoding of dynamic cinematic material Theprecuneus and the right angular gyrus were activatedwhile retrieving the temporal order of events (scene-chro-nology judgment) the superior parietal cortex wasrecruited during spatial related recall (scene-layout
Figure 4
Clusters of brain activation and signal plots for the Scene-
recognition judgment (Object task) Clusters of activation and
signal plots for (A) the MFC and (B) the MTG These regions
activated for the Object task in Exp 1 (blue) and in Exp 2 (red
overlap in magenta) The activation selective for the Object task
was computed with the contrast Ogt [T(short1long)2 1 S]2
inclusively masked with OgtT(short1long)2 and with Ogt S The
signal plots refer to the left hemisphere but analogous patterns
of activation were found in the right hemisphere (Table I) In
Exp 2 the task-specific effect of the Object task in the MFC
was replicated when considering only the sameold images
(Table I rightmost columns) Inspection of the signal plot for the
left MTG revealed less activity in the Oold than Onew trials (plots
on the right bars 5 and 6) although the Oold trials showed
numerically larger parameter estimates than the T and S condi-
tions (see p-uncorr reported in italics in Table I rightmost col-
umns) Activation clusters were estimated and displayed at P-
unc 5 0001 thresholded at a minimum of 80 voxels Effect sizes
are mean adjusted (sum to zero) and expressed in arbitrary
units (au 6 90 CI) LR leftright MTG middle temporal
gyrus MFC medial frontal cortex
r Neural Correlates of Immediate Episodic Retrieval r
r 2507 r
judgment) whereas activity in the MFC and the middletemporal gyrus (MTG) reflected processes associated withscene-recognition judgments These task-specific effectsoccurred independently of whether subjects were asked tojudge seenold or unseennew stimuli (cf ldquoSeen-onlyimagesrdquo contrasts in Exp 2) and independently ofretrieval success and general task difficulty (see Support-ing Information) With respect to the hypothesized corre-spondence between immediate retrieval and retrieval fromLTM we found that these task-specific dissociationsare independent of the informationrsquos mnemonic history(24 h vs 2 s retention ie differing by 43200 times inmagnitude)
One important qualifier for the following discussion isthat memory retrieval in all our tasks depended minimallyon memory buffers compared to other simpler itemsdis-plays that are typically used in ldquostandardrdquo short-termworking memory tasks [Shallice and Cooper 2011 Shiffrin1973] In each trial the number of items presented during
encoding was clearly well beyond the 4 6 1 items stipulatedby classical STM models [Cowan 2001] While memories ofsome materials (eg words spatial positions simple visualdisplays) are supported by limited-capacity buffers [Badde-ley and Hitch 1974] memory for other categories [egunknown faces and scenes see Shiffrin 1973] will tap intoLTM when the task requires the encoding of two or moreitems [Shallice and Cooper 2011] For this reason werestrict our discussion on the tripartite task-specific dissoci-ations with reference to highly complex naturalisticmaterial
Precuneus and Inferior Parietal Cortex Support
Temporal Order Recollection
The scene-chronology judgment task activated the pre-cuneus in both Exp 1 and 2 despite the considerablechanges in the trial structure for probing the retrieval oftemporal information (ie relative temporal order betweentwo probes in Exp 1 vs absolute temporal position of asingle probe in Exp 2) The chronology-judgment taskrequired retrieving the event sequences of many complex
Figure 5
Task-specific retrieval activation common in studies after imme-
diate delays and a longer delay Activation for the LTM study
[24 h Kwok et al 2012] is depicted in green and activation
for the current immediate retrieval study (data from current
Exp 1) depicted in red Activation clusters for both studies were
estimated and displayed at P-unc 5 0001
TABLE II Common memory retrieval activation
ImMem LTM common effects
Cluster Voxel
k P-corr Z x y z
Scene-chronology (Temporal)Precuneus B 112 0011 472 3 257 47Angular gyrus R 98 lt 0001 400 54 254 17
Scene-layout (Spatial)Superior parietal gyrus L 206 0001 595 221 269 56Supramarginal gyrus L 129 0008 346 260 236 44Intraparietal sulcus L 487 236 245 41Superior parietal gyrus R 651 lt 0001 520 24 263 59Dorsal occipital cortex R 372 30 275 29Intraparietal sulcus R 520 42 239 47Supramarginal gyrus R 482 57 233 53Superior frontal sulcus L 100 0022 482 224 3 56Superior frontal sulcus R 145 0004 481 30 0 53Inferior frontal gyrus R 95 0027 475 54 9 26Inferior occipital gyrus R 98 0024 515 54 257 210
Task-specific activation after immediate delays (current Exp 1ImMem) and a previous study that used a long delay betweenencoding and retrieval [24 h LTM Kwok et al 2012] Statisticalthreshold were set at P-FWE-corr 5 005 (cluster-level) consider-ing the whole brain as the volume of interest Note that theldquobetween-studiesrdquo overlap activation in the right angular gyruswas found at a lower uncorrected threshold (in italics) but impor-tantly within this area no differences or interactions betweenImMem and LTM studies were found (see differential effectsbetween the two studies in Supporting Information Table 2) LRB leftright hemispherebilateral
r Kwok et al r
r 2508 r
stimuli (scenes) that entail high-level conceptualhierarchi-cal relationships between them [Swallow et al 2011 Zackset al 2001] The precuneus was previously found to acti-vate in our LTM protocol that used the same temporalorder judgment [Kwok et al 2012 and see also results ofthe conjunction analyses] The current finding of precunealactivation using very different (much shorter) delays fitswell with the view that these processes are not strictlyassociated with retrieval following long retention delayssuggesting that this medial parietal region is implicatedduring temporal order retrieval irrespective of delay (Fig5A and Table II) Specifically we link the effect of chrono-logical retrieval in the precuneus with processes involvingan active reconstruction of past events [see also St Jacqueset al 2008] and possibly memory-related imagery[Fletcher et al 1995 Ishai et al 2000] These processeswould characterize temporal order judgments using com-plex naturalistic material but not order-judgment withsimpler stimulus material which previous studies haveassociated with other brain regions [eg middleprefron-tal areas Suzuki et al 2002]
Together with the precuneus temporal-related judgmentalso gave rise to activation of the right angular gyrus(AG) One possible interpretation relates the inferior parie-tal cortex to the engagement of attentional processes[Majerus et al 2007 2010] The scene-chronology judg-ment of Exp 2 provided us with a further qualifier to thiskind of temporal-related attention Although the Temporaltask in Exp 2 did not require any explicit temporal orderjudgment (only classifying a scene into either the first orsecond half of the movie) an accurate decision still neces-sitated an evaluation of the target scene with many othersegments of the movie This operation is fundamentallydifferent from the evaluation implicated in the scene-layout judgment which required decisions on comparingbetween a single scene and its own correspondentencoded snapshot The latter involves visuospatial analysisof the configuration within scenes The AG may subservedifferent types of attention among which one of them islinked with memory search operations [see Majerus et al2010 2006 also showing coactivation of these two regionsduring order retrieval of simple stimuli after short delays]
Superior Parietal Cortex Evaluates Spatial
Details Against Memory Representation
The second pattern consistently found across experi-ments (Exp 1 and Exp 2 see Fig 3) and across studieswith different retention delays (conjunction betweenimmediate retrieval and LTM studies see Fig 5B andTable II) was the activation of the superior parietal lobule(SPL) during the scene-layout judgment Previous studiesshowed sustained activation in the parietal cortex duringworking memory delays [Courtney et al 1998] and thatactivity there correlates with memory load [Todd andMaroi 2004] Here the amount of information contained
in the clips was equated in the three tasks especially inExp 2 where we counterbalanced the assignment of mov-ies across tasks The subjects also could not anticipatewhat specific type of information would be tested atretrieval and needed to keep rehearsing different aspectsof the clips Because of this we deem unlikely that theactivation of SPL that here was specific for the scene-layout tasks reflected general rehearsal processes [Magenet al 2009] andor updating operations [Bledowski et al2009 Leung et al 2007]
Instead we suggest a role for the SPL in mediating aninterface between the internal memory representation andexternal information It has been proposed that the supe-rior parietal cortex is engaged in allocating top-downattention to orient to aspects of the available visual inputand on information retrieved from memory [Ishai et al2002 Lepsien and Nobre 2007 Summerfield et al 2006]In line with this we found SPL activation when the sub-jects had to make decisions by comparing between theirmemory of the encoded clips and the scene stimuli pre-sented at retrieval This procedure might act on somevisuospatial storage which is held by the lateral occipitalcortex and mediated by the intraparietal sulcus (Fig 3B)The immediate memory task here was analogous to theSpatial task of our previous LTM study [Kwok et al2012] which also required matchingcomparing the avail-able external visual input and internal information storedin memory [see also Nobre et al 2004] This interfaceinvolves a range of top-down processes such as monitor-ing and verification [Cabeza et al 2008 Ciaramelli et al2008] which are independent of the probe imagersquos mne-monic status as shown by the equated effect sizes forldquosamerdquo and ldquoflippedrdquo images conditions (see signals plotsin Fig 3 and also ldquoRetrieval successrdquo analyses of Exp 2 inSupporting Information)
Recognition Processes in the Medial Frontal
Cortex and the Temporal Cortex
The third retrieval task involved scene-recognition judg-ments and was associated with the activation of the MFCand the MTG Moreover in Exp 1 we found a statisticaltrend in the hippocampus bilaterally an area that we pre-viously associated with scene-recognition judgment fromLTM (see next section about ldquoPossible correspondencesbetween retrieval from short- and long-term memoryrdquo)
The MFC was engaged both Exp 1 and 2 but was notactivated in the other two tasks thus a broad prefrontalcortexworking memory correspondence [Tsuchida andFellows 2009] seems insufficient to explain this task-specific effect The scene-recognition task in Exp 2 taxedboth recognition of ldquooldrdquo scenes and correct rejection ofldquonewrdquo scenes suggesting that the MFC here did not dis-tinguish between the correct rejection of new information(unseennew trials) and the recognition of old imagesOne candidate cognitive process that is common to both
r Neural Correlates of Immediate Episodic Retrieval r
r 2509 r
types of processes is feeling-of-knowing (FOK) FOKdepend on the cue specification content retrieval andappraisal of the retrieved memory fragments but are inde-pendent of the ultimate determination of content appropri-ateness [Schnyer et al 2005] Using imaging data duringsentence completion recognition Schnyer et al [2005]modeled a FOK-related directional path The path impli-cated the temporal cortex and the hippocampus andengaged the ventro-MFC at the end which is consistentwith our findings for the Object task implicating the MFCthe middle temporal cortex and at a lower statisticalthreshold the hippocampus
By contrast the activity in the MTG was larger forunseennew images than for seenold memory probes(Fig 4B) This suggests that new encoding may have con-tributed to the activation of the MTG during the retrievalphase of the Object task in Exp 1 The scene-recognitiontask required a lower precision in terms of retrievingdetails about the encoded material and could be solved bysimple recognition [Wixted and Squire 2010] Thus alter-natively larger activation for ldquonew vs oldrdquo stimuli mayrelate to some reduced activation associated with familiar-ity processes on old trials [see Henson et al 1999 whoreported reduced familiarity related activation in middletemporal gyri bilaterally and Henson and Rugg 2003 forreview] However we should point out that in Exp 2 theunseen images were extracted from edited-out parts of theoriginal clips and therefore contained the same objectcharacter depicted in the target images Therefore wedeem more likely that the relational association betweenobjects rather than their presenceabsence in the sceneshelped subjects to distinguish targets from foils We thussuggest that the activation of the MTG reflects processesutilizing the relational details among objects embedded inthe probe images for reaching the recognition decision[Diana et al 2007 Finke et al 2008 Hannula et al 2006Olson et al 2006] Future studies should consider theinclusion of some correlative behavioral measures to indexfamiliarity processes to elucidate further this issue More-over while the Temporal and Spatial tasks required mak-ing relative comparisons about aspects of the encodedvideo-clip the scene-recognition judgment merely requiredjudging whether the image presented at retrieval was pres-ent (or not) within the clip This may entail different proc-essing dynamics in the Object task compared with theother two tasks (cf also differences in retrieval times) Thelatter could be addressed using more subtle experimentalmanipulations for the scene-recognition judgment forexample by using digitally-modified foil images where asingle object in the scene has been modifiedremoved
Possible Correspondences Between Retrieval
from Short- and Long-Term Memory
The overlap between our current study and the previousLTM study using analogous tasks and materials lends
weight to the notion of scale-invariance in memory mecha-nisms [Brown et al 2007] as have been previously dem-onstrated in free recall tasks where the rate of item recallwas unvarying across recall span (day week year) andeven across from the past to the prospective future [May-lor et al 2001] Those previous behavioral findings implythat scale-invariance is a form of self-similarity in that theholistic pattern can exist at multiple levels of magnification[and even across different memory systems eg Maylor2002] Here our data affirmed the importance of the poste-rior parietal cortex in both immediate and long-termretrieval [Berryhill et al 2011 2007] and further character-ized its involvement in dealing with the spatial relationsof objects within scenes [Bricolo et al 2000 Buiatti et al2011] Our data also demonstrated such short-long-termcorrespondence exists in the precuneusangular gyrus fortemporal order retrieval (cf scene-chronology judgmenttask) In contrast the bilateral hippocampus and MFC acti-vations did not reach full statistical significance in theImMem LTM conjunction analysis At first sight thismay indicate that the scene-recognition task tapped intodistinct recognition-based mechanisms as a function ofretention delay However we deem this unlikely becausethe direct comparison between the two datasets did notshow any significant effect in these regions (see Support-ing Information Table 2) Indeed a more targeted test ask-ing the question of whether the hippocampus clusterspreviously found in LTM activated also in the currentimmediate retrieval experiments revealed significanteffects in both immediate retrieval experiments here (Exp1 x y z 5 224 29 225 P-FWE-corrlt 0001 Z 5 458 x yz 5 21 29 222 P-FWE-corr 5 0001 Z 5 451 and in Exp2 x y z 5 224 214 221 P-FWE-corr 5 0004 Z 5 396using the LTM clusters for small volume correction) Theseadditional tests point to a more general issue of sensitivitythat future studies may address using different methodo-logical approaches such as multivariate methods that canbe more sensitive than the standard univariate analysesused here Moreover the former would enable capturingtask-specific effects that are expressed as patterns of dis-tributed activation rather than local changes of activity asreported here [eg Jimura and Poldrack 2012]
Task Difficulty and Retrieval Success
We used several control analyses to show that the task-specific patterns of activation and the correspondencebetween the current data and our previous LTM study donot merely reflect some effect related to general task diffi-culty (see Supporting Information) First areas involved inoverall task difficulty should show progressively greateractivation with progressively more difficult tasks In Exp 1this would entail SgtTgtO However the patterns in theprecuneus and the angular gyrus (with TgtOgt S Fig 2A)and the MFC (Ogt SgtT Fig 4A) did not fit with simplepredictions based on overall task difficulty We also
r Kwok et al r
r 2510 r
assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r

including seen images (cf ldquoExp 2 Seen- onlyrdquo Table I)The signal plots on Figure 2 (panels on the right) show theactivity of the precuneus and the right angular gyrus forall the conditions of Exp 2 Both in the precuneus and theright angular gyrus the average activity in the temporaltask [T(easy1hard)2] were larger than the Ssame and theOold conditions Notably Exp 2 included the presentationof a single image and that the participants were asked tojudge the absolute temporal position of the probe ratherthan the relative temporal order Despite this importantchange of how temporal memory was probed the findingof analogous patterns of activation across both experi-ments highlights the link between immediate retrieval oftemporal information and activation of the precuneus andangular gyrus
Scene-layout judgment (Spatial task)
Exp 1 The contrast concerning the retrieval of the spatiallayout revealed activation in the superior parietal cortexbilaterally with the cluster extending to the intraparietalsulcus and the supramarginal gyrus This contrast alsoshowed activation of the superior frontal sulci the inferiorfrontal gyri plus the inferior occipital cortex (see Table Iand Fig 3 clusters displayed in bluemagenta signalsplots on the left side)
Exp 2 The same comparison in Exp 2 replicated theresults of Exp 1 showing activation in the superior parie-tal cortex bilaterally the superior frontal sulci the inferiorand dorsal occipital cortices and the right inferior frontalgyrus (Fig 3 clusters in redmagenta) When we consid-ered only the conditions including the seenold images(ldquoExp 2 Seen- onlyrdquo) we replicated the effects in the supe-rior parietal gyri the inferior occipital gyri and the rightinferior frontal gyrus (see Table I and Fig 3 signal plotson the right) The activation in the intraparietal sulci andthe superior frontal sulci did not reach statistical signifi-cance Inspection of the signal plots indicated that in theseareas there was less activation in the Ssame than Sflipped tri-als (cf bar 3 vs bar 4 signal plots of the superior fontalsulcus Fig 3A plot on the right) Nonetheless the activityin the Ssame condition was larger than those in the T andO conditions (cf bar 3 vs bars 1ndash2 and 5ndash6)
Scene-recognition judgment (Object task)
Exp 1 The Object task was associated with the activationof the medial frontal cortex (MFC) and the orbitofrontalcortex (but the latter did not fully replicate in Exp 2 seeTable I and Fig 4 clusters bluemagenta and signal plotson the left) together with bilateral activation in the middletemporal gyri the retrosplenialposterior cingulate gyrusand a portion of the angular gyrus The same contrast alsorevealed an activation of the hippocampus bilaterally Theactivation of the hippocampus was just below statisticalsignificance (P-FWE-corr 5 0052 Z 5 458 at x y z 5 2126 219 and P-FWE-corr 5 0077 Z 5 458 at x y z 5 224
29 225 considering the whole brain as the volume ofinterest) and it is reported here because the same effectwas previously found in the LTM dataset [Kwok et al2012 see also Discussion Section]
Exp 2 An almost identical pattern of activation wasfound in Exp 2 when we considered all the conditionsO(old1new)gt (T(easy1hard) 1 S(same1flipped))2 Significanteffects were found in the middle temporal gyri and alarge cluster including the medial frontal gyrus extendingventrally into the orbital-frontal cortex (Fig 4 clusters inredmagenta) However when we considered only the tri-als including seenold-images [Ooldgt (T(easy1hard)2 1 Ssame)2] the Object task was found to significantlyactivate only the MFC (Table I rightmost columns) indi-cating that this region was not affected by incidentalencoding
In summary across all three tasks the brain activationwas found to be similar across Exp 1 and Exp 2 Thescene-chronology Temporal task activated the precuneusand the right angular gyrus the scene-layout Spatial taskwas associated with activation of the posteriorsuperiorparietal cortex and the right inferior frontal gyrus whilethe scene-recognition Object task activated the MFC andthe middle temporal gurus (see Table I areas highlightedin bold) The control analysis of Exp 2 that consideredonly trials including seenold images confirmed thesetask-specific effects indicating that the pattern of dissocia-tionmdashand in particular for the Spatial and Object tasksmdashwas not merely driven by the unseennew images duringthe retrieval phase
Functional Neuroimaging Correspondence with
An Analogous LTM Dataset
We directly compared the task-specific dissociationobserved in the current dataset (Exp 1) with our previousLTM study that included the same task-specific retrievaltasks but after a much longer retention delay [ie 24 hbetween encoding and retrieval Kwok et al 2012] Com-mon across the two data sets the conjunction analysesrevealed an activation cluster in the precuneus for theTemporal task (Fig 5A and Table II) and the dorsal fron-toparietal network plus right inferior frontal gyrus andright occipital cortex for the Spatial task (Fig 5B and TableII) These formal tests show that the Temporal and Spatialtask-specific dissociation observed here for immediateretrieval matched well with the patterns observed duringretrieval from LTM By contrast for the Object task over-lapping effects in the MFC and in the hippocampus weredetected only at a more lenient uncorrected threshold ofP-uncorr 5 0001 In the hippocampus the lack of a fullysignificant ldquocommonrdquo effect across datasets was due to thefact that in the current Exp 1 the contrast comparing theObject task versus the two other tasks did not survive cor-rection for multiple comparisons considering the wholebrain as the volume of interest (P-corr 5 0052 and 0077
r Neural Correlates of Immediate Episodic Retrieval r
r 2505 r
Figure 3
Clusters of brain activation and signal plots for the Scene-layout
judgment (Spatial task) Clusters of activation and signal plots for
(A) the superior frontal cortex and the superior parietal cortex
bilaterally and (B) the right inferior occipital cortex and the right
inferior frontal cortex These regions activated for the Spatial task
in Exp 1 (blue) and in Exp 2 (red overlap between Exp 1 and Exp 2
in magenta) The activation selective for the Spatial task was com-
puted with the contrast Sgt [T(short1long)2 1 O]2 inclusively
masked with SgtT(short1long)2 and with SgtO The effects in the
superior parietal gyri the inferior occipital gyri and the right inferior
frontal gyrus were replicated when considering only sameold
images in Exp 2 (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 80 voxels Effect sizes are mean adjusted (sum to zero)
and are expressed in arbitrary units (au 6 90 CI) LR leftright
SFS superior frontal sulcus SPG superior parietal gyrus IFG infe-
rior frontal gyrus IOG inferior occipital gyrus
in the left and right hemispheres respectively cf above)Indeed the formal test assessing differences between thetwo datasets (ie ldquoTask 3 Experimentrdquo interactions seeSupporting Information Table 2) did not reveal any signifi-cant difference between LTM and immediate retrieval inthe hippocampus consistent with this region beinginvolved in the scene-recognition judgment irrespective ofthe retention delay (see also Discussion Section)
DISCUSSION
We have identified distinct sets of brain areas thatshowed task-specific activation during immediate retrievalfollowing the encoding of dynamic cinematic material Theprecuneus and the right angular gyrus were activatedwhile retrieving the temporal order of events (scene-chro-nology judgment) the superior parietal cortex wasrecruited during spatial related recall (scene-layout
Figure 4
Clusters of brain activation and signal plots for the Scene-
recognition judgment (Object task) Clusters of activation and
signal plots for (A) the MFC and (B) the MTG These regions
activated for the Object task in Exp 1 (blue) and in Exp 2 (red
overlap in magenta) The activation selective for the Object task
was computed with the contrast Ogt [T(short1long)2 1 S]2
inclusively masked with OgtT(short1long)2 and with Ogt S The
signal plots refer to the left hemisphere but analogous patterns
of activation were found in the right hemisphere (Table I) In
Exp 2 the task-specific effect of the Object task in the MFC
was replicated when considering only the sameold images
(Table I rightmost columns) Inspection of the signal plot for the
left MTG revealed less activity in the Oold than Onew trials (plots
on the right bars 5 and 6) although the Oold trials showed
numerically larger parameter estimates than the T and S condi-
tions (see p-uncorr reported in italics in Table I rightmost col-
umns) Activation clusters were estimated and displayed at P-
unc 5 0001 thresholded at a minimum of 80 voxels Effect sizes
are mean adjusted (sum to zero) and expressed in arbitrary
units (au 6 90 CI) LR leftright MTG middle temporal
gyrus MFC medial frontal cortex
r Neural Correlates of Immediate Episodic Retrieval r
r 2507 r
judgment) whereas activity in the MFC and the middletemporal gyrus (MTG) reflected processes associated withscene-recognition judgments These task-specific effectsoccurred independently of whether subjects were asked tojudge seenold or unseennew stimuli (cf ldquoSeen-onlyimagesrdquo contrasts in Exp 2) and independently ofretrieval success and general task difficulty (see Support-ing Information) With respect to the hypothesized corre-spondence between immediate retrieval and retrieval fromLTM we found that these task-specific dissociationsare independent of the informationrsquos mnemonic history(24 h vs 2 s retention ie differing by 43200 times inmagnitude)
One important qualifier for the following discussion isthat memory retrieval in all our tasks depended minimallyon memory buffers compared to other simpler itemsdis-plays that are typically used in ldquostandardrdquo short-termworking memory tasks [Shallice and Cooper 2011 Shiffrin1973] In each trial the number of items presented during
encoding was clearly well beyond the 4 6 1 items stipulatedby classical STM models [Cowan 2001] While memories ofsome materials (eg words spatial positions simple visualdisplays) are supported by limited-capacity buffers [Badde-ley and Hitch 1974] memory for other categories [egunknown faces and scenes see Shiffrin 1973] will tap intoLTM when the task requires the encoding of two or moreitems [Shallice and Cooper 2011] For this reason werestrict our discussion on the tripartite task-specific dissoci-ations with reference to highly complex naturalisticmaterial
Precuneus and Inferior Parietal Cortex Support
Temporal Order Recollection
The scene-chronology judgment task activated the pre-cuneus in both Exp 1 and 2 despite the considerablechanges in the trial structure for probing the retrieval oftemporal information (ie relative temporal order betweentwo probes in Exp 1 vs absolute temporal position of asingle probe in Exp 2) The chronology-judgment taskrequired retrieving the event sequences of many complex
Figure 5
Task-specific retrieval activation common in studies after imme-
diate delays and a longer delay Activation for the LTM study
[24 h Kwok et al 2012] is depicted in green and activation
for the current immediate retrieval study (data from current
Exp 1) depicted in red Activation clusters for both studies were
estimated and displayed at P-unc 5 0001
TABLE II Common memory retrieval activation
ImMem LTM common effects
Cluster Voxel
k P-corr Z x y z
Scene-chronology (Temporal)Precuneus B 112 0011 472 3 257 47Angular gyrus R 98 lt 0001 400 54 254 17
Scene-layout (Spatial)Superior parietal gyrus L 206 0001 595 221 269 56Supramarginal gyrus L 129 0008 346 260 236 44Intraparietal sulcus L 487 236 245 41Superior parietal gyrus R 651 lt 0001 520 24 263 59Dorsal occipital cortex R 372 30 275 29Intraparietal sulcus R 520 42 239 47Supramarginal gyrus R 482 57 233 53Superior frontal sulcus L 100 0022 482 224 3 56Superior frontal sulcus R 145 0004 481 30 0 53Inferior frontal gyrus R 95 0027 475 54 9 26Inferior occipital gyrus R 98 0024 515 54 257 210
Task-specific activation after immediate delays (current Exp 1ImMem) and a previous study that used a long delay betweenencoding and retrieval [24 h LTM Kwok et al 2012] Statisticalthreshold were set at P-FWE-corr 5 005 (cluster-level) consider-ing the whole brain as the volume of interest Note that theldquobetween-studiesrdquo overlap activation in the right angular gyruswas found at a lower uncorrected threshold (in italics) but impor-tantly within this area no differences or interactions betweenImMem and LTM studies were found (see differential effectsbetween the two studies in Supporting Information Table 2) LRB leftright hemispherebilateral
r Kwok et al r
r 2508 r
stimuli (scenes) that entail high-level conceptualhierarchi-cal relationships between them [Swallow et al 2011 Zackset al 2001] The precuneus was previously found to acti-vate in our LTM protocol that used the same temporalorder judgment [Kwok et al 2012 and see also results ofthe conjunction analyses] The current finding of precunealactivation using very different (much shorter) delays fitswell with the view that these processes are not strictlyassociated with retrieval following long retention delayssuggesting that this medial parietal region is implicatedduring temporal order retrieval irrespective of delay (Fig5A and Table II) Specifically we link the effect of chrono-logical retrieval in the precuneus with processes involvingan active reconstruction of past events [see also St Jacqueset al 2008] and possibly memory-related imagery[Fletcher et al 1995 Ishai et al 2000] These processeswould characterize temporal order judgments using com-plex naturalistic material but not order-judgment withsimpler stimulus material which previous studies haveassociated with other brain regions [eg middleprefron-tal areas Suzuki et al 2002]
Together with the precuneus temporal-related judgmentalso gave rise to activation of the right angular gyrus(AG) One possible interpretation relates the inferior parie-tal cortex to the engagement of attentional processes[Majerus et al 2007 2010] The scene-chronology judg-ment of Exp 2 provided us with a further qualifier to thiskind of temporal-related attention Although the Temporaltask in Exp 2 did not require any explicit temporal orderjudgment (only classifying a scene into either the first orsecond half of the movie) an accurate decision still neces-sitated an evaluation of the target scene with many othersegments of the movie This operation is fundamentallydifferent from the evaluation implicated in the scene-layout judgment which required decisions on comparingbetween a single scene and its own correspondentencoded snapshot The latter involves visuospatial analysisof the configuration within scenes The AG may subservedifferent types of attention among which one of them islinked with memory search operations [see Majerus et al2010 2006 also showing coactivation of these two regionsduring order retrieval of simple stimuli after short delays]
Superior Parietal Cortex Evaluates Spatial
Details Against Memory Representation
The second pattern consistently found across experi-ments (Exp 1 and Exp 2 see Fig 3) and across studieswith different retention delays (conjunction betweenimmediate retrieval and LTM studies see Fig 5B andTable II) was the activation of the superior parietal lobule(SPL) during the scene-layout judgment Previous studiesshowed sustained activation in the parietal cortex duringworking memory delays [Courtney et al 1998] and thatactivity there correlates with memory load [Todd andMaroi 2004] Here the amount of information contained
in the clips was equated in the three tasks especially inExp 2 where we counterbalanced the assignment of mov-ies across tasks The subjects also could not anticipatewhat specific type of information would be tested atretrieval and needed to keep rehearsing different aspectsof the clips Because of this we deem unlikely that theactivation of SPL that here was specific for the scene-layout tasks reflected general rehearsal processes [Magenet al 2009] andor updating operations [Bledowski et al2009 Leung et al 2007]
Instead we suggest a role for the SPL in mediating aninterface between the internal memory representation andexternal information It has been proposed that the supe-rior parietal cortex is engaged in allocating top-downattention to orient to aspects of the available visual inputand on information retrieved from memory [Ishai et al2002 Lepsien and Nobre 2007 Summerfield et al 2006]In line with this we found SPL activation when the sub-jects had to make decisions by comparing between theirmemory of the encoded clips and the scene stimuli pre-sented at retrieval This procedure might act on somevisuospatial storage which is held by the lateral occipitalcortex and mediated by the intraparietal sulcus (Fig 3B)The immediate memory task here was analogous to theSpatial task of our previous LTM study [Kwok et al2012] which also required matchingcomparing the avail-able external visual input and internal information storedin memory [see also Nobre et al 2004] This interfaceinvolves a range of top-down processes such as monitor-ing and verification [Cabeza et al 2008 Ciaramelli et al2008] which are independent of the probe imagersquos mne-monic status as shown by the equated effect sizes forldquosamerdquo and ldquoflippedrdquo images conditions (see signals plotsin Fig 3 and also ldquoRetrieval successrdquo analyses of Exp 2 inSupporting Information)
Recognition Processes in the Medial Frontal
Cortex and the Temporal Cortex
The third retrieval task involved scene-recognition judg-ments and was associated with the activation of the MFCand the MTG Moreover in Exp 1 we found a statisticaltrend in the hippocampus bilaterally an area that we pre-viously associated with scene-recognition judgment fromLTM (see next section about ldquoPossible correspondencesbetween retrieval from short- and long-term memoryrdquo)
The MFC was engaged both Exp 1 and 2 but was notactivated in the other two tasks thus a broad prefrontalcortexworking memory correspondence [Tsuchida andFellows 2009] seems insufficient to explain this task-specific effect The scene-recognition task in Exp 2 taxedboth recognition of ldquooldrdquo scenes and correct rejection ofldquonewrdquo scenes suggesting that the MFC here did not dis-tinguish between the correct rejection of new information(unseennew trials) and the recognition of old imagesOne candidate cognitive process that is common to both
r Neural Correlates of Immediate Episodic Retrieval r
r 2509 r
types of processes is feeling-of-knowing (FOK) FOKdepend on the cue specification content retrieval andappraisal of the retrieved memory fragments but are inde-pendent of the ultimate determination of content appropri-ateness [Schnyer et al 2005] Using imaging data duringsentence completion recognition Schnyer et al [2005]modeled a FOK-related directional path The path impli-cated the temporal cortex and the hippocampus andengaged the ventro-MFC at the end which is consistentwith our findings for the Object task implicating the MFCthe middle temporal cortex and at a lower statisticalthreshold the hippocampus
By contrast the activity in the MTG was larger forunseennew images than for seenold memory probes(Fig 4B) This suggests that new encoding may have con-tributed to the activation of the MTG during the retrievalphase of the Object task in Exp 1 The scene-recognitiontask required a lower precision in terms of retrievingdetails about the encoded material and could be solved bysimple recognition [Wixted and Squire 2010] Thus alter-natively larger activation for ldquonew vs oldrdquo stimuli mayrelate to some reduced activation associated with familiar-ity processes on old trials [see Henson et al 1999 whoreported reduced familiarity related activation in middletemporal gyri bilaterally and Henson and Rugg 2003 forreview] However we should point out that in Exp 2 theunseen images were extracted from edited-out parts of theoriginal clips and therefore contained the same objectcharacter depicted in the target images Therefore wedeem more likely that the relational association betweenobjects rather than their presenceabsence in the sceneshelped subjects to distinguish targets from foils We thussuggest that the activation of the MTG reflects processesutilizing the relational details among objects embedded inthe probe images for reaching the recognition decision[Diana et al 2007 Finke et al 2008 Hannula et al 2006Olson et al 2006] Future studies should consider theinclusion of some correlative behavioral measures to indexfamiliarity processes to elucidate further this issue More-over while the Temporal and Spatial tasks required mak-ing relative comparisons about aspects of the encodedvideo-clip the scene-recognition judgment merely requiredjudging whether the image presented at retrieval was pres-ent (or not) within the clip This may entail different proc-essing dynamics in the Object task compared with theother two tasks (cf also differences in retrieval times) Thelatter could be addressed using more subtle experimentalmanipulations for the scene-recognition judgment forexample by using digitally-modified foil images where asingle object in the scene has been modifiedremoved
Possible Correspondences Between Retrieval
from Short- and Long-Term Memory
The overlap between our current study and the previousLTM study using analogous tasks and materials lends
weight to the notion of scale-invariance in memory mecha-nisms [Brown et al 2007] as have been previously dem-onstrated in free recall tasks where the rate of item recallwas unvarying across recall span (day week year) andeven across from the past to the prospective future [May-lor et al 2001] Those previous behavioral findings implythat scale-invariance is a form of self-similarity in that theholistic pattern can exist at multiple levels of magnification[and even across different memory systems eg Maylor2002] Here our data affirmed the importance of the poste-rior parietal cortex in both immediate and long-termretrieval [Berryhill et al 2011 2007] and further character-ized its involvement in dealing with the spatial relationsof objects within scenes [Bricolo et al 2000 Buiatti et al2011] Our data also demonstrated such short-long-termcorrespondence exists in the precuneusangular gyrus fortemporal order retrieval (cf scene-chronology judgmenttask) In contrast the bilateral hippocampus and MFC acti-vations did not reach full statistical significance in theImMem LTM conjunction analysis At first sight thismay indicate that the scene-recognition task tapped intodistinct recognition-based mechanisms as a function ofretention delay However we deem this unlikely becausethe direct comparison between the two datasets did notshow any significant effect in these regions (see Support-ing Information Table 2) Indeed a more targeted test ask-ing the question of whether the hippocampus clusterspreviously found in LTM activated also in the currentimmediate retrieval experiments revealed significanteffects in both immediate retrieval experiments here (Exp1 x y z 5 224 29 225 P-FWE-corrlt 0001 Z 5 458 x yz 5 21 29 222 P-FWE-corr 5 0001 Z 5 451 and in Exp2 x y z 5 224 214 221 P-FWE-corr 5 0004 Z 5 396using the LTM clusters for small volume correction) Theseadditional tests point to a more general issue of sensitivitythat future studies may address using different methodo-logical approaches such as multivariate methods that canbe more sensitive than the standard univariate analysesused here Moreover the former would enable capturingtask-specific effects that are expressed as patterns of dis-tributed activation rather than local changes of activity asreported here [eg Jimura and Poldrack 2012]
Task Difficulty and Retrieval Success
We used several control analyses to show that the task-specific patterns of activation and the correspondencebetween the current data and our previous LTM study donot merely reflect some effect related to general task diffi-culty (see Supporting Information) First areas involved inoverall task difficulty should show progressively greateractivation with progressively more difficult tasks In Exp 1this would entail SgtTgtO However the patterns in theprecuneus and the angular gyrus (with TgtOgt S Fig 2A)and the MFC (Ogt SgtT Fig 4A) did not fit with simplepredictions based on overall task difficulty We also
r Kwok et al r
r 2510 r
assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r
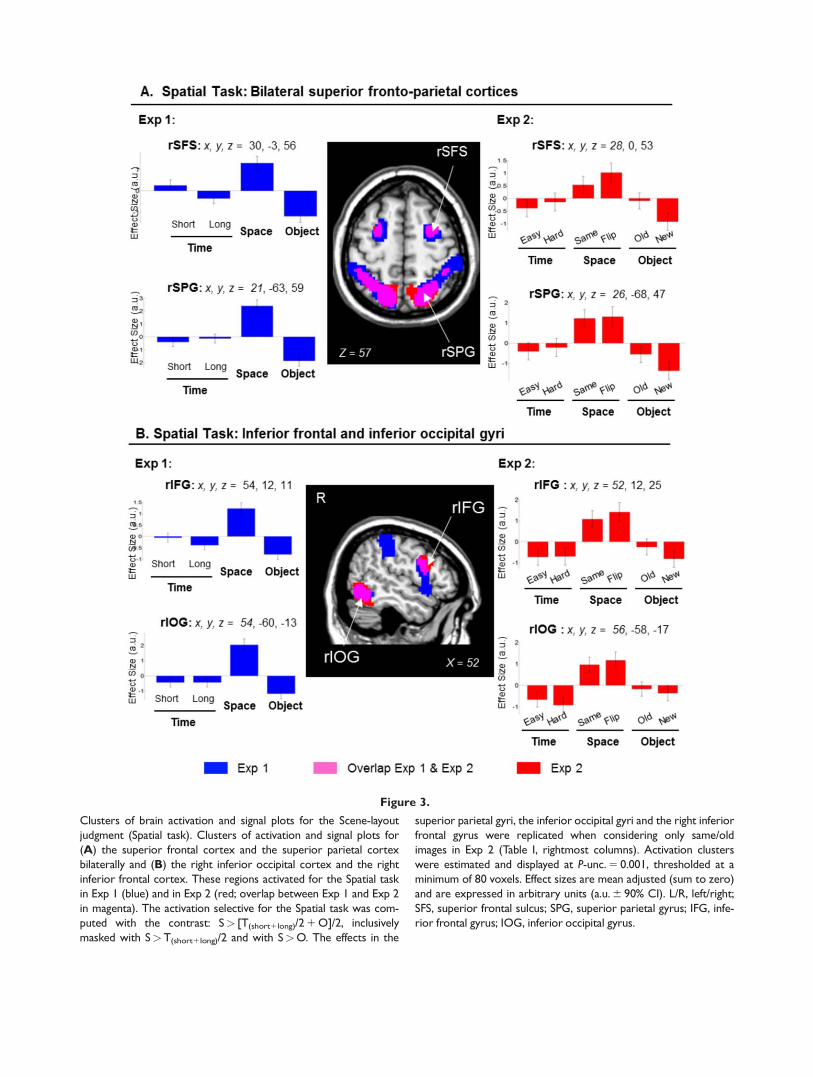
Figure 3
Clusters of brain activation and signal plots for the Scene-layout
judgment (Spatial task) Clusters of activation and signal plots for
(A) the superior frontal cortex and the superior parietal cortex
bilaterally and (B) the right inferior occipital cortex and the right
inferior frontal cortex These regions activated for the Spatial task
in Exp 1 (blue) and in Exp 2 (red overlap between Exp 1 and Exp 2
in magenta) The activation selective for the Spatial task was com-
puted with the contrast Sgt [T(short1long)2 1 O]2 inclusively
masked with SgtT(short1long)2 and with SgtO The effects in the
superior parietal gyri the inferior occipital gyri and the right inferior
frontal gyrus were replicated when considering only sameold
images in Exp 2 (Table I rightmost columns) Activation clusters
were estimated and displayed at P-unc 5 0001 thresholded at a
minimum of 80 voxels Effect sizes are mean adjusted (sum to zero)
and are expressed in arbitrary units (au 6 90 CI) LR leftright
SFS superior frontal sulcus SPG superior parietal gyrus IFG infe-
rior frontal gyrus IOG inferior occipital gyrus
in the left and right hemispheres respectively cf above)Indeed the formal test assessing differences between thetwo datasets (ie ldquoTask 3 Experimentrdquo interactions seeSupporting Information Table 2) did not reveal any signifi-cant difference between LTM and immediate retrieval inthe hippocampus consistent with this region beinginvolved in the scene-recognition judgment irrespective ofthe retention delay (see also Discussion Section)
DISCUSSION
We have identified distinct sets of brain areas thatshowed task-specific activation during immediate retrievalfollowing the encoding of dynamic cinematic material Theprecuneus and the right angular gyrus were activatedwhile retrieving the temporal order of events (scene-chro-nology judgment) the superior parietal cortex wasrecruited during spatial related recall (scene-layout
Figure 4
Clusters of brain activation and signal plots for the Scene-
recognition judgment (Object task) Clusters of activation and
signal plots for (A) the MFC and (B) the MTG These regions
activated for the Object task in Exp 1 (blue) and in Exp 2 (red
overlap in magenta) The activation selective for the Object task
was computed with the contrast Ogt [T(short1long)2 1 S]2
inclusively masked with OgtT(short1long)2 and with Ogt S The
signal plots refer to the left hemisphere but analogous patterns
of activation were found in the right hemisphere (Table I) In
Exp 2 the task-specific effect of the Object task in the MFC
was replicated when considering only the sameold images
(Table I rightmost columns) Inspection of the signal plot for the
left MTG revealed less activity in the Oold than Onew trials (plots
on the right bars 5 and 6) although the Oold trials showed
numerically larger parameter estimates than the T and S condi-
tions (see p-uncorr reported in italics in Table I rightmost col-
umns) Activation clusters were estimated and displayed at P-
unc 5 0001 thresholded at a minimum of 80 voxels Effect sizes
are mean adjusted (sum to zero) and expressed in arbitrary
units (au 6 90 CI) LR leftright MTG middle temporal
gyrus MFC medial frontal cortex
r Neural Correlates of Immediate Episodic Retrieval r
r 2507 r
judgment) whereas activity in the MFC and the middletemporal gyrus (MTG) reflected processes associated withscene-recognition judgments These task-specific effectsoccurred independently of whether subjects were asked tojudge seenold or unseennew stimuli (cf ldquoSeen-onlyimagesrdquo contrasts in Exp 2) and independently ofretrieval success and general task difficulty (see Support-ing Information) With respect to the hypothesized corre-spondence between immediate retrieval and retrieval fromLTM we found that these task-specific dissociationsare independent of the informationrsquos mnemonic history(24 h vs 2 s retention ie differing by 43200 times inmagnitude)
One important qualifier for the following discussion isthat memory retrieval in all our tasks depended minimallyon memory buffers compared to other simpler itemsdis-plays that are typically used in ldquostandardrdquo short-termworking memory tasks [Shallice and Cooper 2011 Shiffrin1973] In each trial the number of items presented during
encoding was clearly well beyond the 4 6 1 items stipulatedby classical STM models [Cowan 2001] While memories ofsome materials (eg words spatial positions simple visualdisplays) are supported by limited-capacity buffers [Badde-ley and Hitch 1974] memory for other categories [egunknown faces and scenes see Shiffrin 1973] will tap intoLTM when the task requires the encoding of two or moreitems [Shallice and Cooper 2011] For this reason werestrict our discussion on the tripartite task-specific dissoci-ations with reference to highly complex naturalisticmaterial
Precuneus and Inferior Parietal Cortex Support
Temporal Order Recollection
The scene-chronology judgment task activated the pre-cuneus in both Exp 1 and 2 despite the considerablechanges in the trial structure for probing the retrieval oftemporal information (ie relative temporal order betweentwo probes in Exp 1 vs absolute temporal position of asingle probe in Exp 2) The chronology-judgment taskrequired retrieving the event sequences of many complex
Figure 5
Task-specific retrieval activation common in studies after imme-
diate delays and a longer delay Activation for the LTM study
[24 h Kwok et al 2012] is depicted in green and activation
for the current immediate retrieval study (data from current
Exp 1) depicted in red Activation clusters for both studies were
estimated and displayed at P-unc 5 0001
TABLE II Common memory retrieval activation
ImMem LTM common effects
Cluster Voxel
k P-corr Z x y z
Scene-chronology (Temporal)Precuneus B 112 0011 472 3 257 47Angular gyrus R 98 lt 0001 400 54 254 17
Scene-layout (Spatial)Superior parietal gyrus L 206 0001 595 221 269 56Supramarginal gyrus L 129 0008 346 260 236 44Intraparietal sulcus L 487 236 245 41Superior parietal gyrus R 651 lt 0001 520 24 263 59Dorsal occipital cortex R 372 30 275 29Intraparietal sulcus R 520 42 239 47Supramarginal gyrus R 482 57 233 53Superior frontal sulcus L 100 0022 482 224 3 56Superior frontal sulcus R 145 0004 481 30 0 53Inferior frontal gyrus R 95 0027 475 54 9 26Inferior occipital gyrus R 98 0024 515 54 257 210
Task-specific activation after immediate delays (current Exp 1ImMem) and a previous study that used a long delay betweenencoding and retrieval [24 h LTM Kwok et al 2012] Statisticalthreshold were set at P-FWE-corr 5 005 (cluster-level) consider-ing the whole brain as the volume of interest Note that theldquobetween-studiesrdquo overlap activation in the right angular gyruswas found at a lower uncorrected threshold (in italics) but impor-tantly within this area no differences or interactions betweenImMem and LTM studies were found (see differential effectsbetween the two studies in Supporting Information Table 2) LRB leftright hemispherebilateral
r Kwok et al r
r 2508 r
stimuli (scenes) that entail high-level conceptualhierarchi-cal relationships between them [Swallow et al 2011 Zackset al 2001] The precuneus was previously found to acti-vate in our LTM protocol that used the same temporalorder judgment [Kwok et al 2012 and see also results ofthe conjunction analyses] The current finding of precunealactivation using very different (much shorter) delays fitswell with the view that these processes are not strictlyassociated with retrieval following long retention delayssuggesting that this medial parietal region is implicatedduring temporal order retrieval irrespective of delay (Fig5A and Table II) Specifically we link the effect of chrono-logical retrieval in the precuneus with processes involvingan active reconstruction of past events [see also St Jacqueset al 2008] and possibly memory-related imagery[Fletcher et al 1995 Ishai et al 2000] These processeswould characterize temporal order judgments using com-plex naturalistic material but not order-judgment withsimpler stimulus material which previous studies haveassociated with other brain regions [eg middleprefron-tal areas Suzuki et al 2002]
Together with the precuneus temporal-related judgmentalso gave rise to activation of the right angular gyrus(AG) One possible interpretation relates the inferior parie-tal cortex to the engagement of attentional processes[Majerus et al 2007 2010] The scene-chronology judg-ment of Exp 2 provided us with a further qualifier to thiskind of temporal-related attention Although the Temporaltask in Exp 2 did not require any explicit temporal orderjudgment (only classifying a scene into either the first orsecond half of the movie) an accurate decision still neces-sitated an evaluation of the target scene with many othersegments of the movie This operation is fundamentallydifferent from the evaluation implicated in the scene-layout judgment which required decisions on comparingbetween a single scene and its own correspondentencoded snapshot The latter involves visuospatial analysisof the configuration within scenes The AG may subservedifferent types of attention among which one of them islinked with memory search operations [see Majerus et al2010 2006 also showing coactivation of these two regionsduring order retrieval of simple stimuli after short delays]
Superior Parietal Cortex Evaluates Spatial
Details Against Memory Representation
The second pattern consistently found across experi-ments (Exp 1 and Exp 2 see Fig 3) and across studieswith different retention delays (conjunction betweenimmediate retrieval and LTM studies see Fig 5B andTable II) was the activation of the superior parietal lobule(SPL) during the scene-layout judgment Previous studiesshowed sustained activation in the parietal cortex duringworking memory delays [Courtney et al 1998] and thatactivity there correlates with memory load [Todd andMaroi 2004] Here the amount of information contained
in the clips was equated in the three tasks especially inExp 2 where we counterbalanced the assignment of mov-ies across tasks The subjects also could not anticipatewhat specific type of information would be tested atretrieval and needed to keep rehearsing different aspectsof the clips Because of this we deem unlikely that theactivation of SPL that here was specific for the scene-layout tasks reflected general rehearsal processes [Magenet al 2009] andor updating operations [Bledowski et al2009 Leung et al 2007]
Instead we suggest a role for the SPL in mediating aninterface between the internal memory representation andexternal information It has been proposed that the supe-rior parietal cortex is engaged in allocating top-downattention to orient to aspects of the available visual inputand on information retrieved from memory [Ishai et al2002 Lepsien and Nobre 2007 Summerfield et al 2006]In line with this we found SPL activation when the sub-jects had to make decisions by comparing between theirmemory of the encoded clips and the scene stimuli pre-sented at retrieval This procedure might act on somevisuospatial storage which is held by the lateral occipitalcortex and mediated by the intraparietal sulcus (Fig 3B)The immediate memory task here was analogous to theSpatial task of our previous LTM study [Kwok et al2012] which also required matchingcomparing the avail-able external visual input and internal information storedin memory [see also Nobre et al 2004] This interfaceinvolves a range of top-down processes such as monitor-ing and verification [Cabeza et al 2008 Ciaramelli et al2008] which are independent of the probe imagersquos mne-monic status as shown by the equated effect sizes forldquosamerdquo and ldquoflippedrdquo images conditions (see signals plotsin Fig 3 and also ldquoRetrieval successrdquo analyses of Exp 2 inSupporting Information)
Recognition Processes in the Medial Frontal
Cortex and the Temporal Cortex
The third retrieval task involved scene-recognition judg-ments and was associated with the activation of the MFCand the MTG Moreover in Exp 1 we found a statisticaltrend in the hippocampus bilaterally an area that we pre-viously associated with scene-recognition judgment fromLTM (see next section about ldquoPossible correspondencesbetween retrieval from short- and long-term memoryrdquo)
The MFC was engaged both Exp 1 and 2 but was notactivated in the other two tasks thus a broad prefrontalcortexworking memory correspondence [Tsuchida andFellows 2009] seems insufficient to explain this task-specific effect The scene-recognition task in Exp 2 taxedboth recognition of ldquooldrdquo scenes and correct rejection ofldquonewrdquo scenes suggesting that the MFC here did not dis-tinguish between the correct rejection of new information(unseennew trials) and the recognition of old imagesOne candidate cognitive process that is common to both
r Neural Correlates of Immediate Episodic Retrieval r
r 2509 r
types of processes is feeling-of-knowing (FOK) FOKdepend on the cue specification content retrieval andappraisal of the retrieved memory fragments but are inde-pendent of the ultimate determination of content appropri-ateness [Schnyer et al 2005] Using imaging data duringsentence completion recognition Schnyer et al [2005]modeled a FOK-related directional path The path impli-cated the temporal cortex and the hippocampus andengaged the ventro-MFC at the end which is consistentwith our findings for the Object task implicating the MFCthe middle temporal cortex and at a lower statisticalthreshold the hippocampus
By contrast the activity in the MTG was larger forunseennew images than for seenold memory probes(Fig 4B) This suggests that new encoding may have con-tributed to the activation of the MTG during the retrievalphase of the Object task in Exp 1 The scene-recognitiontask required a lower precision in terms of retrievingdetails about the encoded material and could be solved bysimple recognition [Wixted and Squire 2010] Thus alter-natively larger activation for ldquonew vs oldrdquo stimuli mayrelate to some reduced activation associated with familiar-ity processes on old trials [see Henson et al 1999 whoreported reduced familiarity related activation in middletemporal gyri bilaterally and Henson and Rugg 2003 forreview] However we should point out that in Exp 2 theunseen images were extracted from edited-out parts of theoriginal clips and therefore contained the same objectcharacter depicted in the target images Therefore wedeem more likely that the relational association betweenobjects rather than their presenceabsence in the sceneshelped subjects to distinguish targets from foils We thussuggest that the activation of the MTG reflects processesutilizing the relational details among objects embedded inthe probe images for reaching the recognition decision[Diana et al 2007 Finke et al 2008 Hannula et al 2006Olson et al 2006] Future studies should consider theinclusion of some correlative behavioral measures to indexfamiliarity processes to elucidate further this issue More-over while the Temporal and Spatial tasks required mak-ing relative comparisons about aspects of the encodedvideo-clip the scene-recognition judgment merely requiredjudging whether the image presented at retrieval was pres-ent (or not) within the clip This may entail different proc-essing dynamics in the Object task compared with theother two tasks (cf also differences in retrieval times) Thelatter could be addressed using more subtle experimentalmanipulations for the scene-recognition judgment forexample by using digitally-modified foil images where asingle object in the scene has been modifiedremoved
Possible Correspondences Between Retrieval
from Short- and Long-Term Memory
The overlap between our current study and the previousLTM study using analogous tasks and materials lends
weight to the notion of scale-invariance in memory mecha-nisms [Brown et al 2007] as have been previously dem-onstrated in free recall tasks where the rate of item recallwas unvarying across recall span (day week year) andeven across from the past to the prospective future [May-lor et al 2001] Those previous behavioral findings implythat scale-invariance is a form of self-similarity in that theholistic pattern can exist at multiple levels of magnification[and even across different memory systems eg Maylor2002] Here our data affirmed the importance of the poste-rior parietal cortex in both immediate and long-termretrieval [Berryhill et al 2011 2007] and further character-ized its involvement in dealing with the spatial relationsof objects within scenes [Bricolo et al 2000 Buiatti et al2011] Our data also demonstrated such short-long-termcorrespondence exists in the precuneusangular gyrus fortemporal order retrieval (cf scene-chronology judgmenttask) In contrast the bilateral hippocampus and MFC acti-vations did not reach full statistical significance in theImMem LTM conjunction analysis At first sight thismay indicate that the scene-recognition task tapped intodistinct recognition-based mechanisms as a function ofretention delay However we deem this unlikely becausethe direct comparison between the two datasets did notshow any significant effect in these regions (see Support-ing Information Table 2) Indeed a more targeted test ask-ing the question of whether the hippocampus clusterspreviously found in LTM activated also in the currentimmediate retrieval experiments revealed significanteffects in both immediate retrieval experiments here (Exp1 x y z 5 224 29 225 P-FWE-corrlt 0001 Z 5 458 x yz 5 21 29 222 P-FWE-corr 5 0001 Z 5 451 and in Exp2 x y z 5 224 214 221 P-FWE-corr 5 0004 Z 5 396using the LTM clusters for small volume correction) Theseadditional tests point to a more general issue of sensitivitythat future studies may address using different methodo-logical approaches such as multivariate methods that canbe more sensitive than the standard univariate analysesused here Moreover the former would enable capturingtask-specific effects that are expressed as patterns of dis-tributed activation rather than local changes of activity asreported here [eg Jimura and Poldrack 2012]
Task Difficulty and Retrieval Success
We used several control analyses to show that the task-specific patterns of activation and the correspondencebetween the current data and our previous LTM study donot merely reflect some effect related to general task diffi-culty (see Supporting Information) First areas involved inoverall task difficulty should show progressively greateractivation with progressively more difficult tasks In Exp 1this would entail SgtTgtO However the patterns in theprecuneus and the angular gyrus (with TgtOgt S Fig 2A)and the MFC (Ogt SgtT Fig 4A) did not fit with simplepredictions based on overall task difficulty We also
r Kwok et al r
r 2510 r
assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r

in the left and right hemispheres respectively cf above)Indeed the formal test assessing differences between thetwo datasets (ie ldquoTask 3 Experimentrdquo interactions seeSupporting Information Table 2) did not reveal any signifi-cant difference between LTM and immediate retrieval inthe hippocampus consistent with this region beinginvolved in the scene-recognition judgment irrespective ofthe retention delay (see also Discussion Section)
DISCUSSION
We have identified distinct sets of brain areas thatshowed task-specific activation during immediate retrievalfollowing the encoding of dynamic cinematic material Theprecuneus and the right angular gyrus were activatedwhile retrieving the temporal order of events (scene-chro-nology judgment) the superior parietal cortex wasrecruited during spatial related recall (scene-layout
Figure 4
Clusters of brain activation and signal plots for the Scene-
recognition judgment (Object task) Clusters of activation and
signal plots for (A) the MFC and (B) the MTG These regions
activated for the Object task in Exp 1 (blue) and in Exp 2 (red
overlap in magenta) The activation selective for the Object task
was computed with the contrast Ogt [T(short1long)2 1 S]2
inclusively masked with OgtT(short1long)2 and with Ogt S The
signal plots refer to the left hemisphere but analogous patterns
of activation were found in the right hemisphere (Table I) In
Exp 2 the task-specific effect of the Object task in the MFC
was replicated when considering only the sameold images
(Table I rightmost columns) Inspection of the signal plot for the
left MTG revealed less activity in the Oold than Onew trials (plots
on the right bars 5 and 6) although the Oold trials showed
numerically larger parameter estimates than the T and S condi-
tions (see p-uncorr reported in italics in Table I rightmost col-
umns) Activation clusters were estimated and displayed at P-
unc 5 0001 thresholded at a minimum of 80 voxels Effect sizes
are mean adjusted (sum to zero) and expressed in arbitrary
units (au 6 90 CI) LR leftright MTG middle temporal
gyrus MFC medial frontal cortex
r Neural Correlates of Immediate Episodic Retrieval r
r 2507 r
judgment) whereas activity in the MFC and the middletemporal gyrus (MTG) reflected processes associated withscene-recognition judgments These task-specific effectsoccurred independently of whether subjects were asked tojudge seenold or unseennew stimuli (cf ldquoSeen-onlyimagesrdquo contrasts in Exp 2) and independently ofretrieval success and general task difficulty (see Support-ing Information) With respect to the hypothesized corre-spondence between immediate retrieval and retrieval fromLTM we found that these task-specific dissociationsare independent of the informationrsquos mnemonic history(24 h vs 2 s retention ie differing by 43200 times inmagnitude)
One important qualifier for the following discussion isthat memory retrieval in all our tasks depended minimallyon memory buffers compared to other simpler itemsdis-plays that are typically used in ldquostandardrdquo short-termworking memory tasks [Shallice and Cooper 2011 Shiffrin1973] In each trial the number of items presented during
encoding was clearly well beyond the 4 6 1 items stipulatedby classical STM models [Cowan 2001] While memories ofsome materials (eg words spatial positions simple visualdisplays) are supported by limited-capacity buffers [Badde-ley and Hitch 1974] memory for other categories [egunknown faces and scenes see Shiffrin 1973] will tap intoLTM when the task requires the encoding of two or moreitems [Shallice and Cooper 2011] For this reason werestrict our discussion on the tripartite task-specific dissoci-ations with reference to highly complex naturalisticmaterial
Precuneus and Inferior Parietal Cortex Support
Temporal Order Recollection
The scene-chronology judgment task activated the pre-cuneus in both Exp 1 and 2 despite the considerablechanges in the trial structure for probing the retrieval oftemporal information (ie relative temporal order betweentwo probes in Exp 1 vs absolute temporal position of asingle probe in Exp 2) The chronology-judgment taskrequired retrieving the event sequences of many complex
Figure 5
Task-specific retrieval activation common in studies after imme-
diate delays and a longer delay Activation for the LTM study
[24 h Kwok et al 2012] is depicted in green and activation
for the current immediate retrieval study (data from current
Exp 1) depicted in red Activation clusters for both studies were
estimated and displayed at P-unc 5 0001
TABLE II Common memory retrieval activation
ImMem LTM common effects
Cluster Voxel
k P-corr Z x y z
Scene-chronology (Temporal)Precuneus B 112 0011 472 3 257 47Angular gyrus R 98 lt 0001 400 54 254 17
Scene-layout (Spatial)Superior parietal gyrus L 206 0001 595 221 269 56Supramarginal gyrus L 129 0008 346 260 236 44Intraparietal sulcus L 487 236 245 41Superior parietal gyrus R 651 lt 0001 520 24 263 59Dorsal occipital cortex R 372 30 275 29Intraparietal sulcus R 520 42 239 47Supramarginal gyrus R 482 57 233 53Superior frontal sulcus L 100 0022 482 224 3 56Superior frontal sulcus R 145 0004 481 30 0 53Inferior frontal gyrus R 95 0027 475 54 9 26Inferior occipital gyrus R 98 0024 515 54 257 210
Task-specific activation after immediate delays (current Exp 1ImMem) and a previous study that used a long delay betweenencoding and retrieval [24 h LTM Kwok et al 2012] Statisticalthreshold were set at P-FWE-corr 5 005 (cluster-level) consider-ing the whole brain as the volume of interest Note that theldquobetween-studiesrdquo overlap activation in the right angular gyruswas found at a lower uncorrected threshold (in italics) but impor-tantly within this area no differences or interactions betweenImMem and LTM studies were found (see differential effectsbetween the two studies in Supporting Information Table 2) LRB leftright hemispherebilateral
r Kwok et al r
r 2508 r
stimuli (scenes) that entail high-level conceptualhierarchi-cal relationships between them [Swallow et al 2011 Zackset al 2001] The precuneus was previously found to acti-vate in our LTM protocol that used the same temporalorder judgment [Kwok et al 2012 and see also results ofthe conjunction analyses] The current finding of precunealactivation using very different (much shorter) delays fitswell with the view that these processes are not strictlyassociated with retrieval following long retention delayssuggesting that this medial parietal region is implicatedduring temporal order retrieval irrespective of delay (Fig5A and Table II) Specifically we link the effect of chrono-logical retrieval in the precuneus with processes involvingan active reconstruction of past events [see also St Jacqueset al 2008] and possibly memory-related imagery[Fletcher et al 1995 Ishai et al 2000] These processeswould characterize temporal order judgments using com-plex naturalistic material but not order-judgment withsimpler stimulus material which previous studies haveassociated with other brain regions [eg middleprefron-tal areas Suzuki et al 2002]
Together with the precuneus temporal-related judgmentalso gave rise to activation of the right angular gyrus(AG) One possible interpretation relates the inferior parie-tal cortex to the engagement of attentional processes[Majerus et al 2007 2010] The scene-chronology judg-ment of Exp 2 provided us with a further qualifier to thiskind of temporal-related attention Although the Temporaltask in Exp 2 did not require any explicit temporal orderjudgment (only classifying a scene into either the first orsecond half of the movie) an accurate decision still neces-sitated an evaluation of the target scene with many othersegments of the movie This operation is fundamentallydifferent from the evaluation implicated in the scene-layout judgment which required decisions on comparingbetween a single scene and its own correspondentencoded snapshot The latter involves visuospatial analysisof the configuration within scenes The AG may subservedifferent types of attention among which one of them islinked with memory search operations [see Majerus et al2010 2006 also showing coactivation of these two regionsduring order retrieval of simple stimuli after short delays]
Superior Parietal Cortex Evaluates Spatial
Details Against Memory Representation
The second pattern consistently found across experi-ments (Exp 1 and Exp 2 see Fig 3) and across studieswith different retention delays (conjunction betweenimmediate retrieval and LTM studies see Fig 5B andTable II) was the activation of the superior parietal lobule(SPL) during the scene-layout judgment Previous studiesshowed sustained activation in the parietal cortex duringworking memory delays [Courtney et al 1998] and thatactivity there correlates with memory load [Todd andMaroi 2004] Here the amount of information contained
in the clips was equated in the three tasks especially inExp 2 where we counterbalanced the assignment of mov-ies across tasks The subjects also could not anticipatewhat specific type of information would be tested atretrieval and needed to keep rehearsing different aspectsof the clips Because of this we deem unlikely that theactivation of SPL that here was specific for the scene-layout tasks reflected general rehearsal processes [Magenet al 2009] andor updating operations [Bledowski et al2009 Leung et al 2007]
Instead we suggest a role for the SPL in mediating aninterface between the internal memory representation andexternal information It has been proposed that the supe-rior parietal cortex is engaged in allocating top-downattention to orient to aspects of the available visual inputand on information retrieved from memory [Ishai et al2002 Lepsien and Nobre 2007 Summerfield et al 2006]In line with this we found SPL activation when the sub-jects had to make decisions by comparing between theirmemory of the encoded clips and the scene stimuli pre-sented at retrieval This procedure might act on somevisuospatial storage which is held by the lateral occipitalcortex and mediated by the intraparietal sulcus (Fig 3B)The immediate memory task here was analogous to theSpatial task of our previous LTM study [Kwok et al2012] which also required matchingcomparing the avail-able external visual input and internal information storedin memory [see also Nobre et al 2004] This interfaceinvolves a range of top-down processes such as monitor-ing and verification [Cabeza et al 2008 Ciaramelli et al2008] which are independent of the probe imagersquos mne-monic status as shown by the equated effect sizes forldquosamerdquo and ldquoflippedrdquo images conditions (see signals plotsin Fig 3 and also ldquoRetrieval successrdquo analyses of Exp 2 inSupporting Information)
Recognition Processes in the Medial Frontal
Cortex and the Temporal Cortex
The third retrieval task involved scene-recognition judg-ments and was associated with the activation of the MFCand the MTG Moreover in Exp 1 we found a statisticaltrend in the hippocampus bilaterally an area that we pre-viously associated with scene-recognition judgment fromLTM (see next section about ldquoPossible correspondencesbetween retrieval from short- and long-term memoryrdquo)
The MFC was engaged both Exp 1 and 2 but was notactivated in the other two tasks thus a broad prefrontalcortexworking memory correspondence [Tsuchida andFellows 2009] seems insufficient to explain this task-specific effect The scene-recognition task in Exp 2 taxedboth recognition of ldquooldrdquo scenes and correct rejection ofldquonewrdquo scenes suggesting that the MFC here did not dis-tinguish between the correct rejection of new information(unseennew trials) and the recognition of old imagesOne candidate cognitive process that is common to both
r Neural Correlates of Immediate Episodic Retrieval r
r 2509 r
types of processes is feeling-of-knowing (FOK) FOKdepend on the cue specification content retrieval andappraisal of the retrieved memory fragments but are inde-pendent of the ultimate determination of content appropri-ateness [Schnyer et al 2005] Using imaging data duringsentence completion recognition Schnyer et al [2005]modeled a FOK-related directional path The path impli-cated the temporal cortex and the hippocampus andengaged the ventro-MFC at the end which is consistentwith our findings for the Object task implicating the MFCthe middle temporal cortex and at a lower statisticalthreshold the hippocampus
By contrast the activity in the MTG was larger forunseennew images than for seenold memory probes(Fig 4B) This suggests that new encoding may have con-tributed to the activation of the MTG during the retrievalphase of the Object task in Exp 1 The scene-recognitiontask required a lower precision in terms of retrievingdetails about the encoded material and could be solved bysimple recognition [Wixted and Squire 2010] Thus alter-natively larger activation for ldquonew vs oldrdquo stimuli mayrelate to some reduced activation associated with familiar-ity processes on old trials [see Henson et al 1999 whoreported reduced familiarity related activation in middletemporal gyri bilaterally and Henson and Rugg 2003 forreview] However we should point out that in Exp 2 theunseen images were extracted from edited-out parts of theoriginal clips and therefore contained the same objectcharacter depicted in the target images Therefore wedeem more likely that the relational association betweenobjects rather than their presenceabsence in the sceneshelped subjects to distinguish targets from foils We thussuggest that the activation of the MTG reflects processesutilizing the relational details among objects embedded inthe probe images for reaching the recognition decision[Diana et al 2007 Finke et al 2008 Hannula et al 2006Olson et al 2006] Future studies should consider theinclusion of some correlative behavioral measures to indexfamiliarity processes to elucidate further this issue More-over while the Temporal and Spatial tasks required mak-ing relative comparisons about aspects of the encodedvideo-clip the scene-recognition judgment merely requiredjudging whether the image presented at retrieval was pres-ent (or not) within the clip This may entail different proc-essing dynamics in the Object task compared with theother two tasks (cf also differences in retrieval times) Thelatter could be addressed using more subtle experimentalmanipulations for the scene-recognition judgment forexample by using digitally-modified foil images where asingle object in the scene has been modifiedremoved
Possible Correspondences Between Retrieval
from Short- and Long-Term Memory
The overlap between our current study and the previousLTM study using analogous tasks and materials lends
weight to the notion of scale-invariance in memory mecha-nisms [Brown et al 2007] as have been previously dem-onstrated in free recall tasks where the rate of item recallwas unvarying across recall span (day week year) andeven across from the past to the prospective future [May-lor et al 2001] Those previous behavioral findings implythat scale-invariance is a form of self-similarity in that theholistic pattern can exist at multiple levels of magnification[and even across different memory systems eg Maylor2002] Here our data affirmed the importance of the poste-rior parietal cortex in both immediate and long-termretrieval [Berryhill et al 2011 2007] and further character-ized its involvement in dealing with the spatial relationsof objects within scenes [Bricolo et al 2000 Buiatti et al2011] Our data also demonstrated such short-long-termcorrespondence exists in the precuneusangular gyrus fortemporal order retrieval (cf scene-chronology judgmenttask) In contrast the bilateral hippocampus and MFC acti-vations did not reach full statistical significance in theImMem LTM conjunction analysis At first sight thismay indicate that the scene-recognition task tapped intodistinct recognition-based mechanisms as a function ofretention delay However we deem this unlikely becausethe direct comparison between the two datasets did notshow any significant effect in these regions (see Support-ing Information Table 2) Indeed a more targeted test ask-ing the question of whether the hippocampus clusterspreviously found in LTM activated also in the currentimmediate retrieval experiments revealed significanteffects in both immediate retrieval experiments here (Exp1 x y z 5 224 29 225 P-FWE-corrlt 0001 Z 5 458 x yz 5 21 29 222 P-FWE-corr 5 0001 Z 5 451 and in Exp2 x y z 5 224 214 221 P-FWE-corr 5 0004 Z 5 396using the LTM clusters for small volume correction) Theseadditional tests point to a more general issue of sensitivitythat future studies may address using different methodo-logical approaches such as multivariate methods that canbe more sensitive than the standard univariate analysesused here Moreover the former would enable capturingtask-specific effects that are expressed as patterns of dis-tributed activation rather than local changes of activity asreported here [eg Jimura and Poldrack 2012]
Task Difficulty and Retrieval Success
We used several control analyses to show that the task-specific patterns of activation and the correspondencebetween the current data and our previous LTM study donot merely reflect some effect related to general task diffi-culty (see Supporting Information) First areas involved inoverall task difficulty should show progressively greateractivation with progressively more difficult tasks In Exp 1this would entail SgtTgtO However the patterns in theprecuneus and the angular gyrus (with TgtOgt S Fig 2A)and the MFC (Ogt SgtT Fig 4A) did not fit with simplepredictions based on overall task difficulty We also
r Kwok et al r
r 2510 r
assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r

judgment) whereas activity in the MFC and the middletemporal gyrus (MTG) reflected processes associated withscene-recognition judgments These task-specific effectsoccurred independently of whether subjects were asked tojudge seenold or unseennew stimuli (cf ldquoSeen-onlyimagesrdquo contrasts in Exp 2) and independently ofretrieval success and general task difficulty (see Support-ing Information) With respect to the hypothesized corre-spondence between immediate retrieval and retrieval fromLTM we found that these task-specific dissociationsare independent of the informationrsquos mnemonic history(24 h vs 2 s retention ie differing by 43200 times inmagnitude)
One important qualifier for the following discussion isthat memory retrieval in all our tasks depended minimallyon memory buffers compared to other simpler itemsdis-plays that are typically used in ldquostandardrdquo short-termworking memory tasks [Shallice and Cooper 2011 Shiffrin1973] In each trial the number of items presented during
encoding was clearly well beyond the 4 6 1 items stipulatedby classical STM models [Cowan 2001] While memories ofsome materials (eg words spatial positions simple visualdisplays) are supported by limited-capacity buffers [Badde-ley and Hitch 1974] memory for other categories [egunknown faces and scenes see Shiffrin 1973] will tap intoLTM when the task requires the encoding of two or moreitems [Shallice and Cooper 2011] For this reason werestrict our discussion on the tripartite task-specific dissoci-ations with reference to highly complex naturalisticmaterial
Precuneus and Inferior Parietal Cortex Support
Temporal Order Recollection
The scene-chronology judgment task activated the pre-cuneus in both Exp 1 and 2 despite the considerablechanges in the trial structure for probing the retrieval oftemporal information (ie relative temporal order betweentwo probes in Exp 1 vs absolute temporal position of asingle probe in Exp 2) The chronology-judgment taskrequired retrieving the event sequences of many complex
Figure 5
Task-specific retrieval activation common in studies after imme-
diate delays and a longer delay Activation for the LTM study
[24 h Kwok et al 2012] is depicted in green and activation
for the current immediate retrieval study (data from current
Exp 1) depicted in red Activation clusters for both studies were
estimated and displayed at P-unc 5 0001
TABLE II Common memory retrieval activation
ImMem LTM common effects
Cluster Voxel
k P-corr Z x y z
Scene-chronology (Temporal)Precuneus B 112 0011 472 3 257 47Angular gyrus R 98 lt 0001 400 54 254 17
Scene-layout (Spatial)Superior parietal gyrus L 206 0001 595 221 269 56Supramarginal gyrus L 129 0008 346 260 236 44Intraparietal sulcus L 487 236 245 41Superior parietal gyrus R 651 lt 0001 520 24 263 59Dorsal occipital cortex R 372 30 275 29Intraparietal sulcus R 520 42 239 47Supramarginal gyrus R 482 57 233 53Superior frontal sulcus L 100 0022 482 224 3 56Superior frontal sulcus R 145 0004 481 30 0 53Inferior frontal gyrus R 95 0027 475 54 9 26Inferior occipital gyrus R 98 0024 515 54 257 210
Task-specific activation after immediate delays (current Exp 1ImMem) and a previous study that used a long delay betweenencoding and retrieval [24 h LTM Kwok et al 2012] Statisticalthreshold were set at P-FWE-corr 5 005 (cluster-level) consider-ing the whole brain as the volume of interest Note that theldquobetween-studiesrdquo overlap activation in the right angular gyruswas found at a lower uncorrected threshold (in italics) but impor-tantly within this area no differences or interactions betweenImMem and LTM studies were found (see differential effectsbetween the two studies in Supporting Information Table 2) LRB leftright hemispherebilateral
r Kwok et al r
r 2508 r
stimuli (scenes) that entail high-level conceptualhierarchi-cal relationships between them [Swallow et al 2011 Zackset al 2001] The precuneus was previously found to acti-vate in our LTM protocol that used the same temporalorder judgment [Kwok et al 2012 and see also results ofthe conjunction analyses] The current finding of precunealactivation using very different (much shorter) delays fitswell with the view that these processes are not strictlyassociated with retrieval following long retention delayssuggesting that this medial parietal region is implicatedduring temporal order retrieval irrespective of delay (Fig5A and Table II) Specifically we link the effect of chrono-logical retrieval in the precuneus with processes involvingan active reconstruction of past events [see also St Jacqueset al 2008] and possibly memory-related imagery[Fletcher et al 1995 Ishai et al 2000] These processeswould characterize temporal order judgments using com-plex naturalistic material but not order-judgment withsimpler stimulus material which previous studies haveassociated with other brain regions [eg middleprefron-tal areas Suzuki et al 2002]
Together with the precuneus temporal-related judgmentalso gave rise to activation of the right angular gyrus(AG) One possible interpretation relates the inferior parie-tal cortex to the engagement of attentional processes[Majerus et al 2007 2010] The scene-chronology judg-ment of Exp 2 provided us with a further qualifier to thiskind of temporal-related attention Although the Temporaltask in Exp 2 did not require any explicit temporal orderjudgment (only classifying a scene into either the first orsecond half of the movie) an accurate decision still neces-sitated an evaluation of the target scene with many othersegments of the movie This operation is fundamentallydifferent from the evaluation implicated in the scene-layout judgment which required decisions on comparingbetween a single scene and its own correspondentencoded snapshot The latter involves visuospatial analysisof the configuration within scenes The AG may subservedifferent types of attention among which one of them islinked with memory search operations [see Majerus et al2010 2006 also showing coactivation of these two regionsduring order retrieval of simple stimuli after short delays]
Superior Parietal Cortex Evaluates Spatial
Details Against Memory Representation
The second pattern consistently found across experi-ments (Exp 1 and Exp 2 see Fig 3) and across studieswith different retention delays (conjunction betweenimmediate retrieval and LTM studies see Fig 5B andTable II) was the activation of the superior parietal lobule(SPL) during the scene-layout judgment Previous studiesshowed sustained activation in the parietal cortex duringworking memory delays [Courtney et al 1998] and thatactivity there correlates with memory load [Todd andMaroi 2004] Here the amount of information contained
in the clips was equated in the three tasks especially inExp 2 where we counterbalanced the assignment of mov-ies across tasks The subjects also could not anticipatewhat specific type of information would be tested atretrieval and needed to keep rehearsing different aspectsof the clips Because of this we deem unlikely that theactivation of SPL that here was specific for the scene-layout tasks reflected general rehearsal processes [Magenet al 2009] andor updating operations [Bledowski et al2009 Leung et al 2007]
Instead we suggest a role for the SPL in mediating aninterface between the internal memory representation andexternal information It has been proposed that the supe-rior parietal cortex is engaged in allocating top-downattention to orient to aspects of the available visual inputand on information retrieved from memory [Ishai et al2002 Lepsien and Nobre 2007 Summerfield et al 2006]In line with this we found SPL activation when the sub-jects had to make decisions by comparing between theirmemory of the encoded clips and the scene stimuli pre-sented at retrieval This procedure might act on somevisuospatial storage which is held by the lateral occipitalcortex and mediated by the intraparietal sulcus (Fig 3B)The immediate memory task here was analogous to theSpatial task of our previous LTM study [Kwok et al2012] which also required matchingcomparing the avail-able external visual input and internal information storedin memory [see also Nobre et al 2004] This interfaceinvolves a range of top-down processes such as monitor-ing and verification [Cabeza et al 2008 Ciaramelli et al2008] which are independent of the probe imagersquos mne-monic status as shown by the equated effect sizes forldquosamerdquo and ldquoflippedrdquo images conditions (see signals plotsin Fig 3 and also ldquoRetrieval successrdquo analyses of Exp 2 inSupporting Information)
Recognition Processes in the Medial Frontal
Cortex and the Temporal Cortex
The third retrieval task involved scene-recognition judg-ments and was associated with the activation of the MFCand the MTG Moreover in Exp 1 we found a statisticaltrend in the hippocampus bilaterally an area that we pre-viously associated with scene-recognition judgment fromLTM (see next section about ldquoPossible correspondencesbetween retrieval from short- and long-term memoryrdquo)
The MFC was engaged both Exp 1 and 2 but was notactivated in the other two tasks thus a broad prefrontalcortexworking memory correspondence [Tsuchida andFellows 2009] seems insufficient to explain this task-specific effect The scene-recognition task in Exp 2 taxedboth recognition of ldquooldrdquo scenes and correct rejection ofldquonewrdquo scenes suggesting that the MFC here did not dis-tinguish between the correct rejection of new information(unseennew trials) and the recognition of old imagesOne candidate cognitive process that is common to both
r Neural Correlates of Immediate Episodic Retrieval r
r 2509 r
types of processes is feeling-of-knowing (FOK) FOKdepend on the cue specification content retrieval andappraisal of the retrieved memory fragments but are inde-pendent of the ultimate determination of content appropri-ateness [Schnyer et al 2005] Using imaging data duringsentence completion recognition Schnyer et al [2005]modeled a FOK-related directional path The path impli-cated the temporal cortex and the hippocampus andengaged the ventro-MFC at the end which is consistentwith our findings for the Object task implicating the MFCthe middle temporal cortex and at a lower statisticalthreshold the hippocampus
By contrast the activity in the MTG was larger forunseennew images than for seenold memory probes(Fig 4B) This suggests that new encoding may have con-tributed to the activation of the MTG during the retrievalphase of the Object task in Exp 1 The scene-recognitiontask required a lower precision in terms of retrievingdetails about the encoded material and could be solved bysimple recognition [Wixted and Squire 2010] Thus alter-natively larger activation for ldquonew vs oldrdquo stimuli mayrelate to some reduced activation associated with familiar-ity processes on old trials [see Henson et al 1999 whoreported reduced familiarity related activation in middletemporal gyri bilaterally and Henson and Rugg 2003 forreview] However we should point out that in Exp 2 theunseen images were extracted from edited-out parts of theoriginal clips and therefore contained the same objectcharacter depicted in the target images Therefore wedeem more likely that the relational association betweenobjects rather than their presenceabsence in the sceneshelped subjects to distinguish targets from foils We thussuggest that the activation of the MTG reflects processesutilizing the relational details among objects embedded inthe probe images for reaching the recognition decision[Diana et al 2007 Finke et al 2008 Hannula et al 2006Olson et al 2006] Future studies should consider theinclusion of some correlative behavioral measures to indexfamiliarity processes to elucidate further this issue More-over while the Temporal and Spatial tasks required mak-ing relative comparisons about aspects of the encodedvideo-clip the scene-recognition judgment merely requiredjudging whether the image presented at retrieval was pres-ent (or not) within the clip This may entail different proc-essing dynamics in the Object task compared with theother two tasks (cf also differences in retrieval times) Thelatter could be addressed using more subtle experimentalmanipulations for the scene-recognition judgment forexample by using digitally-modified foil images where asingle object in the scene has been modifiedremoved
Possible Correspondences Between Retrieval
from Short- and Long-Term Memory
The overlap between our current study and the previousLTM study using analogous tasks and materials lends
weight to the notion of scale-invariance in memory mecha-nisms [Brown et al 2007] as have been previously dem-onstrated in free recall tasks where the rate of item recallwas unvarying across recall span (day week year) andeven across from the past to the prospective future [May-lor et al 2001] Those previous behavioral findings implythat scale-invariance is a form of self-similarity in that theholistic pattern can exist at multiple levels of magnification[and even across different memory systems eg Maylor2002] Here our data affirmed the importance of the poste-rior parietal cortex in both immediate and long-termretrieval [Berryhill et al 2011 2007] and further character-ized its involvement in dealing with the spatial relationsof objects within scenes [Bricolo et al 2000 Buiatti et al2011] Our data also demonstrated such short-long-termcorrespondence exists in the precuneusangular gyrus fortemporal order retrieval (cf scene-chronology judgmenttask) In contrast the bilateral hippocampus and MFC acti-vations did not reach full statistical significance in theImMem LTM conjunction analysis At first sight thismay indicate that the scene-recognition task tapped intodistinct recognition-based mechanisms as a function ofretention delay However we deem this unlikely becausethe direct comparison between the two datasets did notshow any significant effect in these regions (see Support-ing Information Table 2) Indeed a more targeted test ask-ing the question of whether the hippocampus clusterspreviously found in LTM activated also in the currentimmediate retrieval experiments revealed significanteffects in both immediate retrieval experiments here (Exp1 x y z 5 224 29 225 P-FWE-corrlt 0001 Z 5 458 x yz 5 21 29 222 P-FWE-corr 5 0001 Z 5 451 and in Exp2 x y z 5 224 214 221 P-FWE-corr 5 0004 Z 5 396using the LTM clusters for small volume correction) Theseadditional tests point to a more general issue of sensitivitythat future studies may address using different methodo-logical approaches such as multivariate methods that canbe more sensitive than the standard univariate analysesused here Moreover the former would enable capturingtask-specific effects that are expressed as patterns of dis-tributed activation rather than local changes of activity asreported here [eg Jimura and Poldrack 2012]
Task Difficulty and Retrieval Success
We used several control analyses to show that the task-specific patterns of activation and the correspondencebetween the current data and our previous LTM study donot merely reflect some effect related to general task diffi-culty (see Supporting Information) First areas involved inoverall task difficulty should show progressively greateractivation with progressively more difficult tasks In Exp 1this would entail SgtTgtO However the patterns in theprecuneus and the angular gyrus (with TgtOgt S Fig 2A)and the MFC (Ogt SgtT Fig 4A) did not fit with simplepredictions based on overall task difficulty We also
r Kwok et al r
r 2510 r
assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r

stimuli (scenes) that entail high-level conceptualhierarchi-cal relationships between them [Swallow et al 2011 Zackset al 2001] The precuneus was previously found to acti-vate in our LTM protocol that used the same temporalorder judgment [Kwok et al 2012 and see also results ofthe conjunction analyses] The current finding of precunealactivation using very different (much shorter) delays fitswell with the view that these processes are not strictlyassociated with retrieval following long retention delayssuggesting that this medial parietal region is implicatedduring temporal order retrieval irrespective of delay (Fig5A and Table II) Specifically we link the effect of chrono-logical retrieval in the precuneus with processes involvingan active reconstruction of past events [see also St Jacqueset al 2008] and possibly memory-related imagery[Fletcher et al 1995 Ishai et al 2000] These processeswould characterize temporal order judgments using com-plex naturalistic material but not order-judgment withsimpler stimulus material which previous studies haveassociated with other brain regions [eg middleprefron-tal areas Suzuki et al 2002]
Together with the precuneus temporal-related judgmentalso gave rise to activation of the right angular gyrus(AG) One possible interpretation relates the inferior parie-tal cortex to the engagement of attentional processes[Majerus et al 2007 2010] The scene-chronology judg-ment of Exp 2 provided us with a further qualifier to thiskind of temporal-related attention Although the Temporaltask in Exp 2 did not require any explicit temporal orderjudgment (only classifying a scene into either the first orsecond half of the movie) an accurate decision still neces-sitated an evaluation of the target scene with many othersegments of the movie This operation is fundamentallydifferent from the evaluation implicated in the scene-layout judgment which required decisions on comparingbetween a single scene and its own correspondentencoded snapshot The latter involves visuospatial analysisof the configuration within scenes The AG may subservedifferent types of attention among which one of them islinked with memory search operations [see Majerus et al2010 2006 also showing coactivation of these two regionsduring order retrieval of simple stimuli after short delays]
Superior Parietal Cortex Evaluates Spatial
Details Against Memory Representation
The second pattern consistently found across experi-ments (Exp 1 and Exp 2 see Fig 3) and across studieswith different retention delays (conjunction betweenimmediate retrieval and LTM studies see Fig 5B andTable II) was the activation of the superior parietal lobule(SPL) during the scene-layout judgment Previous studiesshowed sustained activation in the parietal cortex duringworking memory delays [Courtney et al 1998] and thatactivity there correlates with memory load [Todd andMaroi 2004] Here the amount of information contained
in the clips was equated in the three tasks especially inExp 2 where we counterbalanced the assignment of mov-ies across tasks The subjects also could not anticipatewhat specific type of information would be tested atretrieval and needed to keep rehearsing different aspectsof the clips Because of this we deem unlikely that theactivation of SPL that here was specific for the scene-layout tasks reflected general rehearsal processes [Magenet al 2009] andor updating operations [Bledowski et al2009 Leung et al 2007]
Instead we suggest a role for the SPL in mediating aninterface between the internal memory representation andexternal information It has been proposed that the supe-rior parietal cortex is engaged in allocating top-downattention to orient to aspects of the available visual inputand on information retrieved from memory [Ishai et al2002 Lepsien and Nobre 2007 Summerfield et al 2006]In line with this we found SPL activation when the sub-jects had to make decisions by comparing between theirmemory of the encoded clips and the scene stimuli pre-sented at retrieval This procedure might act on somevisuospatial storage which is held by the lateral occipitalcortex and mediated by the intraparietal sulcus (Fig 3B)The immediate memory task here was analogous to theSpatial task of our previous LTM study [Kwok et al2012] which also required matchingcomparing the avail-able external visual input and internal information storedin memory [see also Nobre et al 2004] This interfaceinvolves a range of top-down processes such as monitor-ing and verification [Cabeza et al 2008 Ciaramelli et al2008] which are independent of the probe imagersquos mne-monic status as shown by the equated effect sizes forldquosamerdquo and ldquoflippedrdquo images conditions (see signals plotsin Fig 3 and also ldquoRetrieval successrdquo analyses of Exp 2 inSupporting Information)
Recognition Processes in the Medial Frontal
Cortex and the Temporal Cortex
The third retrieval task involved scene-recognition judg-ments and was associated with the activation of the MFCand the MTG Moreover in Exp 1 we found a statisticaltrend in the hippocampus bilaterally an area that we pre-viously associated with scene-recognition judgment fromLTM (see next section about ldquoPossible correspondencesbetween retrieval from short- and long-term memoryrdquo)
The MFC was engaged both Exp 1 and 2 but was notactivated in the other two tasks thus a broad prefrontalcortexworking memory correspondence [Tsuchida andFellows 2009] seems insufficient to explain this task-specific effect The scene-recognition task in Exp 2 taxedboth recognition of ldquooldrdquo scenes and correct rejection ofldquonewrdquo scenes suggesting that the MFC here did not dis-tinguish between the correct rejection of new information(unseennew trials) and the recognition of old imagesOne candidate cognitive process that is common to both
r Neural Correlates of Immediate Episodic Retrieval r
r 2509 r
types of processes is feeling-of-knowing (FOK) FOKdepend on the cue specification content retrieval andappraisal of the retrieved memory fragments but are inde-pendent of the ultimate determination of content appropri-ateness [Schnyer et al 2005] Using imaging data duringsentence completion recognition Schnyer et al [2005]modeled a FOK-related directional path The path impli-cated the temporal cortex and the hippocampus andengaged the ventro-MFC at the end which is consistentwith our findings for the Object task implicating the MFCthe middle temporal cortex and at a lower statisticalthreshold the hippocampus
By contrast the activity in the MTG was larger forunseennew images than for seenold memory probes(Fig 4B) This suggests that new encoding may have con-tributed to the activation of the MTG during the retrievalphase of the Object task in Exp 1 The scene-recognitiontask required a lower precision in terms of retrievingdetails about the encoded material and could be solved bysimple recognition [Wixted and Squire 2010] Thus alter-natively larger activation for ldquonew vs oldrdquo stimuli mayrelate to some reduced activation associated with familiar-ity processes on old trials [see Henson et al 1999 whoreported reduced familiarity related activation in middletemporal gyri bilaterally and Henson and Rugg 2003 forreview] However we should point out that in Exp 2 theunseen images were extracted from edited-out parts of theoriginal clips and therefore contained the same objectcharacter depicted in the target images Therefore wedeem more likely that the relational association betweenobjects rather than their presenceabsence in the sceneshelped subjects to distinguish targets from foils We thussuggest that the activation of the MTG reflects processesutilizing the relational details among objects embedded inthe probe images for reaching the recognition decision[Diana et al 2007 Finke et al 2008 Hannula et al 2006Olson et al 2006] Future studies should consider theinclusion of some correlative behavioral measures to indexfamiliarity processes to elucidate further this issue More-over while the Temporal and Spatial tasks required mak-ing relative comparisons about aspects of the encodedvideo-clip the scene-recognition judgment merely requiredjudging whether the image presented at retrieval was pres-ent (or not) within the clip This may entail different proc-essing dynamics in the Object task compared with theother two tasks (cf also differences in retrieval times) Thelatter could be addressed using more subtle experimentalmanipulations for the scene-recognition judgment forexample by using digitally-modified foil images where asingle object in the scene has been modifiedremoved
Possible Correspondences Between Retrieval
from Short- and Long-Term Memory
The overlap between our current study and the previousLTM study using analogous tasks and materials lends
weight to the notion of scale-invariance in memory mecha-nisms [Brown et al 2007] as have been previously dem-onstrated in free recall tasks where the rate of item recallwas unvarying across recall span (day week year) andeven across from the past to the prospective future [May-lor et al 2001] Those previous behavioral findings implythat scale-invariance is a form of self-similarity in that theholistic pattern can exist at multiple levels of magnification[and even across different memory systems eg Maylor2002] Here our data affirmed the importance of the poste-rior parietal cortex in both immediate and long-termretrieval [Berryhill et al 2011 2007] and further character-ized its involvement in dealing with the spatial relationsof objects within scenes [Bricolo et al 2000 Buiatti et al2011] Our data also demonstrated such short-long-termcorrespondence exists in the precuneusangular gyrus fortemporal order retrieval (cf scene-chronology judgmenttask) In contrast the bilateral hippocampus and MFC acti-vations did not reach full statistical significance in theImMem LTM conjunction analysis At first sight thismay indicate that the scene-recognition task tapped intodistinct recognition-based mechanisms as a function ofretention delay However we deem this unlikely becausethe direct comparison between the two datasets did notshow any significant effect in these regions (see Support-ing Information Table 2) Indeed a more targeted test ask-ing the question of whether the hippocampus clusterspreviously found in LTM activated also in the currentimmediate retrieval experiments revealed significanteffects in both immediate retrieval experiments here (Exp1 x y z 5 224 29 225 P-FWE-corrlt 0001 Z 5 458 x yz 5 21 29 222 P-FWE-corr 5 0001 Z 5 451 and in Exp2 x y z 5 224 214 221 P-FWE-corr 5 0004 Z 5 396using the LTM clusters for small volume correction) Theseadditional tests point to a more general issue of sensitivitythat future studies may address using different methodo-logical approaches such as multivariate methods that canbe more sensitive than the standard univariate analysesused here Moreover the former would enable capturingtask-specific effects that are expressed as patterns of dis-tributed activation rather than local changes of activity asreported here [eg Jimura and Poldrack 2012]
Task Difficulty and Retrieval Success
We used several control analyses to show that the task-specific patterns of activation and the correspondencebetween the current data and our previous LTM study donot merely reflect some effect related to general task diffi-culty (see Supporting Information) First areas involved inoverall task difficulty should show progressively greateractivation with progressively more difficult tasks In Exp 1this would entail SgtTgtO However the patterns in theprecuneus and the angular gyrus (with TgtOgt S Fig 2A)and the MFC (Ogt SgtT Fig 4A) did not fit with simplepredictions based on overall task difficulty We also
r Kwok et al r
r 2510 r
assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r

types of processes is feeling-of-knowing (FOK) FOKdepend on the cue specification content retrieval andappraisal of the retrieved memory fragments but are inde-pendent of the ultimate determination of content appropri-ateness [Schnyer et al 2005] Using imaging data duringsentence completion recognition Schnyer et al [2005]modeled a FOK-related directional path The path impli-cated the temporal cortex and the hippocampus andengaged the ventro-MFC at the end which is consistentwith our findings for the Object task implicating the MFCthe middle temporal cortex and at a lower statisticalthreshold the hippocampus
By contrast the activity in the MTG was larger forunseennew images than for seenold memory probes(Fig 4B) This suggests that new encoding may have con-tributed to the activation of the MTG during the retrievalphase of the Object task in Exp 1 The scene-recognitiontask required a lower precision in terms of retrievingdetails about the encoded material and could be solved bysimple recognition [Wixted and Squire 2010] Thus alter-natively larger activation for ldquonew vs oldrdquo stimuli mayrelate to some reduced activation associated with familiar-ity processes on old trials [see Henson et al 1999 whoreported reduced familiarity related activation in middletemporal gyri bilaterally and Henson and Rugg 2003 forreview] However we should point out that in Exp 2 theunseen images were extracted from edited-out parts of theoriginal clips and therefore contained the same objectcharacter depicted in the target images Therefore wedeem more likely that the relational association betweenobjects rather than their presenceabsence in the sceneshelped subjects to distinguish targets from foils We thussuggest that the activation of the MTG reflects processesutilizing the relational details among objects embedded inthe probe images for reaching the recognition decision[Diana et al 2007 Finke et al 2008 Hannula et al 2006Olson et al 2006] Future studies should consider theinclusion of some correlative behavioral measures to indexfamiliarity processes to elucidate further this issue More-over while the Temporal and Spatial tasks required mak-ing relative comparisons about aspects of the encodedvideo-clip the scene-recognition judgment merely requiredjudging whether the image presented at retrieval was pres-ent (or not) within the clip This may entail different proc-essing dynamics in the Object task compared with theother two tasks (cf also differences in retrieval times) Thelatter could be addressed using more subtle experimentalmanipulations for the scene-recognition judgment forexample by using digitally-modified foil images where asingle object in the scene has been modifiedremoved
Possible Correspondences Between Retrieval
from Short- and Long-Term Memory
The overlap between our current study and the previousLTM study using analogous tasks and materials lends
weight to the notion of scale-invariance in memory mecha-nisms [Brown et al 2007] as have been previously dem-onstrated in free recall tasks where the rate of item recallwas unvarying across recall span (day week year) andeven across from the past to the prospective future [May-lor et al 2001] Those previous behavioral findings implythat scale-invariance is a form of self-similarity in that theholistic pattern can exist at multiple levels of magnification[and even across different memory systems eg Maylor2002] Here our data affirmed the importance of the poste-rior parietal cortex in both immediate and long-termretrieval [Berryhill et al 2011 2007] and further character-ized its involvement in dealing with the spatial relationsof objects within scenes [Bricolo et al 2000 Buiatti et al2011] Our data also demonstrated such short-long-termcorrespondence exists in the precuneusangular gyrus fortemporal order retrieval (cf scene-chronology judgmenttask) In contrast the bilateral hippocampus and MFC acti-vations did not reach full statistical significance in theImMem LTM conjunction analysis At first sight thismay indicate that the scene-recognition task tapped intodistinct recognition-based mechanisms as a function ofretention delay However we deem this unlikely becausethe direct comparison between the two datasets did notshow any significant effect in these regions (see Support-ing Information Table 2) Indeed a more targeted test ask-ing the question of whether the hippocampus clusterspreviously found in LTM activated also in the currentimmediate retrieval experiments revealed significanteffects in both immediate retrieval experiments here (Exp1 x y z 5 224 29 225 P-FWE-corrlt 0001 Z 5 458 x yz 5 21 29 222 P-FWE-corr 5 0001 Z 5 451 and in Exp2 x y z 5 224 214 221 P-FWE-corr 5 0004 Z 5 396using the LTM clusters for small volume correction) Theseadditional tests point to a more general issue of sensitivitythat future studies may address using different methodo-logical approaches such as multivariate methods that canbe more sensitive than the standard univariate analysesused here Moreover the former would enable capturingtask-specific effects that are expressed as patterns of dis-tributed activation rather than local changes of activity asreported here [eg Jimura and Poldrack 2012]
Task Difficulty and Retrieval Success
We used several control analyses to show that the task-specific patterns of activation and the correspondencebetween the current data and our previous LTM study donot merely reflect some effect related to general task diffi-culty (see Supporting Information) First areas involved inoverall task difficulty should show progressively greateractivation with progressively more difficult tasks In Exp 1this would entail SgtTgtO However the patterns in theprecuneus and the angular gyrus (with TgtOgt S Fig 2A)and the MFC (Ogt SgtT Fig 4A) did not fit with simplepredictions based on overall task difficulty We also
r Kwok et al r
r 2510 r
assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r

assessed the effect of difficulty using the Temporal andSpatial tasks in Exp 2 The contrast ldquohardgt easyrdquo Tempo-ral trials revealed one single activation cluster in the sup-plementary frontal eye field which has been associatedwith difficult decision making [Heekeren et al 2004]None of the critical task-specific regions (Figs 2ndash4) wasactivated by this ldquohardgt easyrdquo comparison Behaviorallythere were also differences between ldquoflipped vs samerdquoSpatial trials The corresponding contrast showed only oneactivation cluster in the left inferior frontal gyrus Thisregion was initially associated with the Spatial task in Exp1 but no corresponding effect was found in Exp 2 (seeTable I) For Exp 1 we additionally replicated the mainfindings of task-specific dissociation by entering RT as acovariate of no interest to account for task difficulty As awhole these results indicate that the task-specific dissocia-tion during immediate retrieval cannot be simply attrib-uted to task difficulty The same pattern was previouslydemonstrated with analogous control analyses of the LTMdataset [see p 2949 Kwok et al 2012] indicating that dif-ferences in overall task difficulty cannot account for thetask-specific correspondence between retrieval followingshort and long retention delays
Similar to the previous LTM study [Kwok et al 2012]also here the task-specific activation patterns were inde-pendent of retrieval success (see Supporting Information)This emphasizes the centrality of ldquoretrieval attemptsrdquo[Rugg and Wilding 2000] both for immediate retrievaland for retrieval after long delays in line with the pro-posal that there might exist time-scale invariance in theneural correlates of memory retrieval In Exp 1 we foundthat the task-specific activation associated with the scene-chronology (Temporal) and scene-layout (Spatial) taskswere common for correct and incorrect trials In Exp 2 thetask-specific effects for the scene-layout and the scene-recognition (Object) tasks were common for trials includ-ing oldseen pictures and trials including unseennewstimuli These results were in line with our analogousanalyses on the LTM dataset [p 2950 Kwok et al 2012]and indicate that the task-specific dissociation found bothfor immediate and long-delay retrieval should be relatedto the ldquoattemptrdquo to retrieve specific types of informationrather than to the successful recovery of stimulus contentfrom memory
Finally rather than advocating that retrieval from STMand LTM are ldquothe samerdquo here we sought to characterizethe specific demands in each task and map the corre-sponding neural correlates A limitation of the currentapproach using naturalistic material is that the complexityof the movie stimuli may have led the participants to usea mixture of different strategies during retrieval Forinstance the subjects could attempt to solve the scene-chronology task by remembering which objects occurredmore recently (ie recency judgment) or rely on non-spatial details within the scenes to identify the targetimage in the scene-layout task Nonetheless the findingsof analogous dissociations in both Exp 1 and Exp 2 [and
LTM study in Kwok et al 2012] which used considerablydifferent tasks to probe memory for the same whatwherewhen type of information suggest that these corre-spondences are largely independent on task characteristicsInstead each task might flexibly recruit a subset of a hostof ldquoatomicrdquo cognitive processes to subserve task specificfunctions [Van Snellenberg and Wager 2009] meetingretrieval demands across different retention intervals
CONCLUSION
Cast in a ldquowhat-where-whenrdquo framework we showedtask-specific functional dissociations during the immediateretrieval of information about complex and naturalisticmaterial The precuneus and the right angular gyrus wereactivated during retrieving the temporal order of eventsthe superior parietal cortex in spatial-related judgmentwhereas the MFC together with middle temporal areaswere activated during scene recognition By comparing thecurrent results involving short retention delays with a pre-vious LTM paradigm with a 24-h delay between encodingand retrieval we found delay-independent functional cor-respondence of the judgment specific patterns The find-ings can be ascribed to a subset of atomic processesguided by task-specific (whatwherewhen) and stimuli-specific (complex scenes) constraints that operate in ascale-free manner across a wide range of retention delays
ACKNOWLEDGEMENT
The authors thank Prof Tim Shallice for his substantialcontribution to the refinement of many ideas in this articleand Sara Pernigotti for editing the video clips
REFERENCES
Baddeley A Hitch GJ (1974) Working memory In Bower GHeditor The Psychology of Learning and Motivation Advancesin Research and Theory New York Academic Press pp 47ndash89
Berryhill ME Phuong L Picasso L Cabeza R Olson IR (2007)Parietal lobe and episodic memory Bilateral damage causesimpaired free recall of autobiographical memory J Neurosci2714415ndash14423
Berryhill ME Chein J Olson IR (2011) At the intersection ofattention and memory The mechanistic role of the posteriorparietal lobe in working memory Neuropsychologia 491306ndash1315
Bledowski C Rahm B Rowe JB (2009) What ldquoworksrdquo in workingmemory Separate systems for selection and updating of criti-cal information J Neurosci 2913735ndash13741
Bricolo E Shallice T Priftis K Meneghello F (2000) Selectivespace transformation deficit in a patient with spatial agnosiaNeurocase 6307ndash319
Brown GDA Neath I Chater N (2007) A temporal ratio model ofmemory Psychol Rev 114539ndash576
Buiatti T Mussoni A Toraldo A Skrap M Shallice T (2011) Twoqualitatively different impairments in making rotation opera-tions Cortex 47166ndash179
r Neural Correlates of Immediate Episodic Retrieval r
r 2511 r
Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r

Burgess N Maguire EA Spiers HJ OrsquoKeefe J (2001) A temporo-parietal and prefrontal network for retrieving the spatial con-text of lifelike events NeuroImage 14439ndash453
Cabeza R Mangels JA Nyberg L Habib R Houle S McIntosh ARTulving E (1997) Brain regions differentially involved in remem-bering what and when A PET study Neuron 19863ndash870
Cabeza R Ciaramelli E Olson IR Moscovitch M (2008) The parie-tal cortex and episodic memory An attentional account NatRev Neurosci 9613ndash625
Ciaramelli E Grady CL Moscovitch M (2008) Top-down andbottom-up attention to memory A hypothesis (AtoM) on therole of the posterior parietal cortex in memory retrieval Neu-ropsychologia 461828ndash1851
Courtney SM Petit L Maisog JM Ungerleider LG Haxby JV(1998) An area specialized for spatial working memory inhuman frontal cortex Science 2791347ndash1351
Cowan N (2001) The magical number 4 in short-term memory Areconsideration of mental storage capacity Behav Brain Sci 2487ndash114
Diana RA Yonelinas AP Ranganath C (2007) Imaging recollec-tion and familiarity in the medial temporal lobe A three-component model Trends Cogn Sci 11379ndash386
Ekstrom AD Bookheimer SY (2007) Spatial and temporal episodicmemory retrieval recruit dissociable functional networks in thehuman brain Learn Mem 14645ndash654
Ekstrom AD Copara MS Isham EA Wang W-C Yonelinas AP(2011) Dissociable networks involved in spatial and temporalorder source retrieval NeuroImage 561803ndash1813
Finke C Braun M Ostendorf F Lehmann T-N Hoffmann K-TKopp U Ploner CJ (2008) The human hippocampal formationmediates short-term memory of colourndashlocation associationsNeuropsychologia 46614ndash623
Fletcher PC Frith CD Baker SC Shallice T Frackowiak RSJDolan RJ (1995) The Mindrsquos EyemdashPrecuneus activation inmemory-related imagery NeuroImage 2195ndash200
Friston KJ Glaser DE Henson RN Kiebel S Phillips CAshburner J (2002) Classical and Bayesian inference in neuroi-maging Applications Neuroimage 16484ndash512
Fujii T Suzuki M Okuda J Ohtake H Tanji K Yamaguchi K ItohM Yamadori A (2004) Neural correlates of context memorywith real-world events NeuroImage 211596ndash1603
Furman O Mendelsohn A Dudai Y (2012) The episodic engramtransformed Time reduces retrieval-related brain activity butcorrelates it with memory accuracy Learn Mem 19575ndash587
Fuster JM (2006) The cognit A network model of cortical repre-sentation Int J Psychophysiol 60125ndash132
Fuster JM (2009) Cortex and memory Emergence of a new para-digm J Cogn Neurosci 212047ndash2072
Gaffan D (2002) Against memory systems Philos Trans R SocLond B Biol Sci 3571111ndash1121
Hacker MJ (1980) Speed and accuracy of recency judgments forevents in short-term memory J Exp Psychol Hum Learn Mem6651ndash675
Hannula DE Tranel D Cohen NJ (2006) The long and the shortof it Relational memory impairments in amnesia even at shortlags J Neurosci 268352ndash8359
Harrison A Jolicoeur P Marois R (2010) ldquoWhatrdquo and ldquoWhererdquo inthe intraparietal sulcus An fMRI study of object identity andlocation in visual short-term memory Cereb Cortex 202478ndash2485
Hayama HR Vilberg KL Rugg MD (2012) Overlap between theneural correlates of cued recall and source memory Evidence
for a generic recollection network J Cogn Neurosci 241127ndash1137
Hayes SM Ryan L Schnyer DM Nadel L (2004) An fMRI studyof episodic memory Retrieval of object spatial and temporalinformation Behav Neurosci 118885ndash896
Heekeren HR Marrett S Bandettini PA Ungerleider LG (2004) Ageneral mechanism for perceptual decision-making in thehuman brain Nature 431859ndash862
Henson R Rugg M (2003) Neural response suppression haemo-dynamic repetition effects and behavioural priming Neuro-psychologia 41263ndash270
Henson RNA Rugg MD Shallice T Josephs O Dolan RJ (1999)Recollection and familiarity in recognition memory An event-related functional magnetic resonance imaging studyJ Neurosci 193962ndash3972
Holland S Smulders T (2011) Do humans use episodic memoryto solve a What-Where-When memory task Anim Cogn 1495ndash102
Howard MW Kahana MJ (2002) A distributed representation oftemporal context J Math Psychol 46269ndash299
Howard MW Fotedar MS Datey AV Hasselmo ME (2005) Thetemporal context model in spatial navigation and relationallearning Toward a common explanation of medial temporallobe function across domains Psychol Rev 11275ndash116
Ishai A Ungerleider LG Haxby JV (2000) Distributed neural sys-tems for the generation of visual images Neuron 28979ndash990
Ishai A Haxby JV Ungerleider LG (2002) Visual imagery offamous faces Effects of memory and attention revealed byfMRI NeuroImage 171729ndash1741
Jimura K Poldrack RA (2012) Analyses of regional-average acti-vation and multivoxel pattern information tell complementarystories Neuropsychologia 50544ndash552
Jonides J Lacey SC Nee DE (2005) Processes of working memoryin mind and brain Curr Dir Psychol Sci 142ndash5
Jonides J Lewis RL Nee DE Lustig CA Berman MG Moore KS(2008) The mind and brain of short-term memory Annu RevPsychol 59193ndash224
Konishi S Uchida I Okuaki T Machida T Shirouzu I MiyashitaY (2002) Neural correlates of recency judgment J Neurosci 229549ndash9555
Kwok SC Shallice T Macaluso E (2012) Functional anatomy oftemporal organisation and domain-specificity of episodic mem-ory retrieval Neuropsychologia 502943ndash2955
Kwok SC Shallice T Macaluso E (2014) Set-relevance determinesthe impact of distractors on episodic memory retrieval J CognNeurosci 262070ndash2086
Lepsien J Nobre AC (2007) Attentional modulation of object rep-resentations in working memory Cereb Cortex 172072ndash2083
Leung H-C Oh H Ferri J Yi Y (2007) Load response functions inthe human spatial working memory circuit during locationmemory updating Neuroimage 35368
Magen H Emmanouil T-A McMains SA Kastner S Treisman A(2009) Attentional demands predict short-term memory loadresponse in posterior parietal cortex Neuropsychologia 471790ndash1798
Majerus S Poncelet M Van der Linden M Albouy G SalmonE Sterpenich V Vandewalle G Collette F Maquet P(2006) The left intraparietal sulcus and verbal short-termmemory Focus of attention or serial order Neuroimage32880ndash891
Majerus S Bastin C Poncelet M Van der Linden M Salmon ECollette F Maquet P (2007) Short-term memory and the left
r Kwok et al r
r 2512 r
intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r

intraparietal sulcus Focus of attention Further evidence froma face short-term memory paradigm Neuroimage 35357ndash363
Majerus S DrsquoArgembeau A Perez TM Belayachi S Van derLinden M Collette F Salmon E Seurinck R Fias W Maquet P(2010) The commonality of neural networks for verbal and vis-ual short-term memory J Cogn Neurosci 222570ndash2593
Maylor EA (2002) Serial position effects in semantic memoryReconstructing the order of verses of hymns Psychon Bull Rev9816ndash820
Maylor EA Chater N Brown GDA (2001) Scale invariance in theretrieval of retrospective and prospective memories PsychonBull Rev 8162ndash167
Milner B Corsi P Leonard G (1991) Frontal-lobe contribution torecency judgements Neuropsychologia 29601ndash618
Mohr HM Goebel R Linden DEJ (2006) Content- and task-specific dissociations of frontal activity during maintenanceand manipulation in visual working memory J Neurosci 264465ndash4471
Munk MHJ Linden DEJ Muckli L Lanfermann H Zanella FESinger W Goebel R (2002) Distributed cortical systems in vis-ual short-term memory revealed by event-related functionalmagnetic resonance imaging Cereb Cortex 12866ndash876
Muter P (1979) Response latencies in discriminations of recencyJ Exp Psychol Hum Learn Mem 5160ndash169
Nee DE Berman MG Moore KS Jonides J (2008) Neuroscientificevidence about the distinction between short- and long-termmemory Curr Dir Psychol Sci 17102ndash106
Nobre AC Coull JT Maquet P Frith CD Vandenberghe RMesulam MM (2004) Orienting attention to locations in per-ceptual versus mental representations J Cogn Neurosci 16363ndash373
Nyberg L McIntosh A R Cabeza R Habib R Houle S Tulving E(1996) General and specific brain regions involved in encodingand retrieval of events What where and when Proc NatlAcad Sci USA 9311280ndash11285
Olson IR Page K Moore KS Chatterjee A Verfaellie M (2006)Working memory for conjunctions relies on the medial tempo-ral lobe J Neurosci 264596ndash4601
Penny W Holmes A (2004) Random effects analysis In Fracko-wiak RS Friston KJ Frith CD Dolan RJ Price CJZeki S Ashburner J Penny W editors Human Brain Func-tion 2nd ed San Diego CA Elsevier p 843ndash851
Postle BR (2006) Working memory as an emergent property ofthe mind and brain Neuroscience 13923ndash38
Ranganath C Blumenfeld RS (2005) Doubts about double dissoci-ations between short- and long-term memory Trends Cogn Sci9374ndash380
Rugg MD Wilding EL (2000) Retrieval processing and episodicmemory Trends Cogn Sci 4108ndash115
Schnyer DM Nicholls L Verfaellie M (2005) The role of VMPC inmetamemorial judgments of content retrievability J Cogn Neu-rosci 17832ndash846
Shallice T Cooper RP (2011) The Organisation of Mind NewYork Oxford University Press
Shallice T Warrington EK (1970) Independent functioning ofverbal memory stores A neuropsychological study Q J ExpPsychol 22261ndash273
Shiffrin RM (1973) Visual free recall Science 180980ndash982Smith TJ Henderson JM (2008) Edit blindness The relationship
between attention and global change blindness in dynamicscenes J Eye Mov Res 21ndash17
St Jacques P Rubin DC LaBar KS Cabeza R (2008) The shortand long of it Neural correlates of temporal-order memory forautobiographical events J Cogn Neurosci 201327ndash1341
Summerfield JJ Lepsien J Gitelman DR Mesulam MM NobreAC (2006) Orienting attention based on long-term memoryexperience Neuron 49905ndash916
Suzuki M Fujii T Tsukiura T Okuda J Umetsu A Nagasaka TMugikura S Yanagawa I Takahashi S Yamadori A (2002)Neural basis of temporal context memory A functional MRIstudy NeuroImage 171790ndash1796
Swallow KM Barch DM Head D Maley CJ Holder D Zacks JM(2011) Changes in events alter how people remember recentinformation J Cogn Neurosci 231052ndash1064
Todd JJ Maroi R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex Nature 428751ndash754
Tsuchida A Fellows LK (2009) Lesion evidence that two distinctregions within prefrontal cortex are critical for n-back perform-ance in humans J Cogn Neurosci 212263ndash2275
Van Snellenberg JX Wager TD (2009) Cognitive and motivationalfunctions of the human prefrontal cortex In Christensen A-LBougakov D Goldberg E editors Luriarsquos Legacy in the 21stCentury New York Oxford University Press pp 30ndash61
Wixted JT Squire LR (2010) The role of the human hippocampusin familiarity-based and recollection-based recognition mem-ory Behav Brain Res 215197ndash208
Wohldmann EL Healy AF Bourne LE Jr (2008) A mental practicesuperiority effect Less retroactive interference and more trans-fer than physical practice J Exp Psychol Learn Mem Cogn 34823ndash833
Zacks JM Braver TS Sheridan MA Donaldson DI Snyder AZOllinger JM Buckner RL Raichle ME (2001) Human brainactivity time-locked to perceptual event boundaries Nat Neu-rosci 4651ndash655
r Neural Correlates of Immediate Episodic Retrieval r
r 2513 r
Copyright © 2022 FDOKUMEN




















