
Resuscitation by positive pressure ventilation and Trishydroxymethylaminomethane of rhesus monkeys...
12
The Journal of PEDIATRICS 807 Resuscitation by positive pressure ventilation and Tris-hydroxymetbylaminomethane of rhesus monkeys asphyxiated at birth Rhesus monkeys delivered by cesarean section were asphyxiated under controlled environmental conditions and then resuscitated by positive pressure ventilation with oxygen. In one hall of the monkeys an infusion of Tris and glucose was given into the umbilical vein during resuscitation to restore the arterial pH rapidly to normal values. This reduced the time required to establish spontaneous breathing and increased the initial oxygen uptake. There was a linear relationship between duration o[ asphyxia and the return o[ respiratory [unction: for each minute a#er the last gasp that resuscitation was de[erred, subsequent onset o[ gasping was delayed about 2 minutes and spontaneous breathing about 4 minutes. Nearly one half o[ the asphyxiated animals developed respiratory di~cuIties similar to those seen in human in[ants suffering #ore the respiratory distress syndrome. K. Adams0ns, Jr., ~ R. Behrman, G. S. Dawes, ~ L. S. James, ~r and C. Koford OXFORD~ ENGLAND P R E V I O U S experiments with mature fe- tal lambs and monkeys indicated that in- fusion of alkali and glucose during the course of asphyxia lasting 10 to 15 minutes prolonged survivaP -3 and reduced or pre- From the Columbia-Presbyterian Medical Center, New York, United States; the Nu~eld Institute for Medical Research, University o[ Ox[ord, England; and the Laboratory of PerinataI Physiology, National Institute for Neurological Diseases and Blindness, San Juan, Puerto Rico. e*Obstetrics and Gynecology, University of Puerto Rico and WeUeome Senior Research Fellow. x~Depar(ment of Physiology, University of Puerto Rico. Address, the Nu~eld Institute for Medical Research, 43 Woodstock Road, Oxford, England. .... Department of Pediatrics, University of Puerto Rico and Investigator, New York City Health Research Council under Contract 1. 148. vented permanent brain injury. 4 The time between the beginning of resuscitation by positive pressure ventilation and the onset of gasping and spontaneous breathing was much reduced by the infusion. This could have been due either to the maintenance of the arterial pit and glycolysis during as- phyxia, or to the fact that the arterial pH was within normal limits when resuscitation was begun. The present experiments were designed to determine whether rapid in- fusion of alkali and glucose at the onset of ventilation would assist recovery. The previous experiments suggested that there was a definite relation between the
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Resuscitation by positive pressure ventilation and Trishydroxymethylaminomethane of rhesus monkeys...

T h e Journal of P E D I A T R I C S 8 0 7
Resuscitation by positive pressure ventilation
and Tris-hydroxymetbylaminomethane of
rhesus monkeys asphyxiated at birth
Rhesus monkeys delivered by cesarean section were asphyxiated under controlled
environmental conditions and then resuscitated by positive pressure ventilation with
oxygen. In one hall of the monkeys an infusion of Tris and glucose was given into
the umbilical vein during resuscitation to restore the arterial pH rapidly to normal
values. This reduced the time required to establish spontaneous breathing and
increased the initial oxygen uptake. There was a linear relationship between duration
o[ asphyxia and the return o[ respiratory [unction: for each minute a#er the last
gasp that resuscitation was de[erred, subsequent onset o[ gasping was delayed about
2 minutes and spontaneous breathing about 4 minutes. Nearly one half o[ the
asphyxiated animals developed respiratory di~cuIties similar to those seen in human
in[ants suffering #ore the respiratory distress syndrome.
K. Adams0ns, Jr., ~ R. Behrman, G. S. Dawes, ~ L. S. James, ~r and C. Koford
OXFORD~ E N G L A N D
P R E V I O U S experiments with mature fe -
t a l lambs and monkeys indicated that in- fusion of alkali and glucose during the course of asphyxia lasting 10 to 15 minutes prolonged survivaP -3 and reduced or pre-
From the Columbia-Presbyterian Medical Center, New York, United States; the Nu~eld Institute for Medical Research, University o[ Ox[ord, England; and the Laboratory of PerinataI Physiology, National Institute for Neurological Diseases and Blindness, San Juan, Puerto Rico. e*Obstetrics and Gynecology, University of Puerto Rico and WeUeome Senior Research Fellow. x~Depar(ment of Physiology, University of Puerto Rico. Address, the Nu~e ld Institute for Medical Research, 43 Woodstock Road, Oxford, England. ... . Department of Pediatrics, University of Puerto
Rico and Investigator, New York City Health Research Council under Contract 1. 148.
vented permanent brain injury. 4 The time between the beginning of resuscitation by positive pressure ventilation and the onset of gasping and spontaneous breathing was much reduced by the infusion. This could have been due either to the maintenance of the arterial p i t and glycolysis during as- phyxia, or to the fact that the arterial pH was within normal limits when resuscitation was begun. The present experiments were designed to determine whether rapid in- fusion of alkali and glucose at the onset of ventilation would assist recovery.
The previous experiments suggested that there was a definite relation between the

8 0 8 Adamsons et al. December 1964
"•7"---..•OLUM E ENDOTRACHEAL I~ RECORDER
f
J , HEAT , V;Z;T PUMP ] EXCHANGER ]--LLIME F - -
Fig. 1. Schematic diagram of apparatus for mea- surement of 02 consumption during positive pres- sure ventilation or spontaneous breathing. When the bulb is compressed to raise the pressure in the closed circuit, and hence to inflate the lungs, a relay closes off the tube connecting the volume recorder to the circuit. The apparatus is filled with 90 to 95 per cent 02 initially, and the vol- ume recorder is refilled at fixed time intervals.
MM.HG
160 / ~
P CO 2
0 - 7.4-
7.2
ZO
6.8
PH
1 ASPHYXIA
I I I E r 0 I0 20 40 60
MINUTES
Fig. 2. Changes in arterial blood pH and PCO2 during asphyxia and recovery in 12 newly de- livered rhesus monkeys, resuscitated by positive pressure ventilation alone (Q) or combined with a rapid intravenous infusion of Tris and glucose (O). The vertical bars indicate the S.E. of the means,
duration of asphyxia and the interval f rom the start of resuscitation until the establish- ment of rhythmical breathing, and also that
asphyxia might increase the incidence of respiratory distress, Further experiments have been undertaken to examine these questions.
M A T E R I A L A N D M E T H O D S
Observations were made on rhesus mon- keys delivered by cesarean section. The mothers were either f rom the cage colony in San Juan (and the date of conception was known) or from the free-ranging colony on Cayo Santiago, an inbred community established in 1938 with monkeys imported from IndiaP Pregnant monkeys on Cayo Santiago which were thought to be near term were caught and examined in order to estimate fetal weight. The biparietal di- ameter, which is about 5 cm. at term, was measured through the abdominal wall. The size of the fetal parts and the engagement of the head in the pelvis also proved useful indications of size and maturi ty. If the fetus was judged to be premature, the mother was kept in a cage until fetal size was con- sidered adequate. All the fetuses were single- tons and weighed 310 to 590 grams. They were well developed and their physiologic behavior (heart rate and blood pressure, arterial pH, PCO2, lactate and pyruvate concentrations, and time to last gasp during asphyxia) was like that of the monkeys of 139 to 159 days' gestation studied previ- ouslyY, a
The uterus was exposed under local anes- thesia, and the fetus was delivered and asphyxiated by slipping a rubber bag filled with saline solution over the head and tying the umbilical cord. The infant was then placed at once on a table whose environ- mental temperature was 30 ~ C. All the "in- fants were in good condition on delivery, struggling and making strong respiratory efforts. Blood samples were wi thdrawn from the placental end of the umbilical arteries. Earlier experiments had shown good agree- ment between the pH, PCO2, and lactate concentrations measured in such samples and those from a femoral ar tery before de- livery of the fetus. Needle electrodes for recording the electrocardiogram were in-
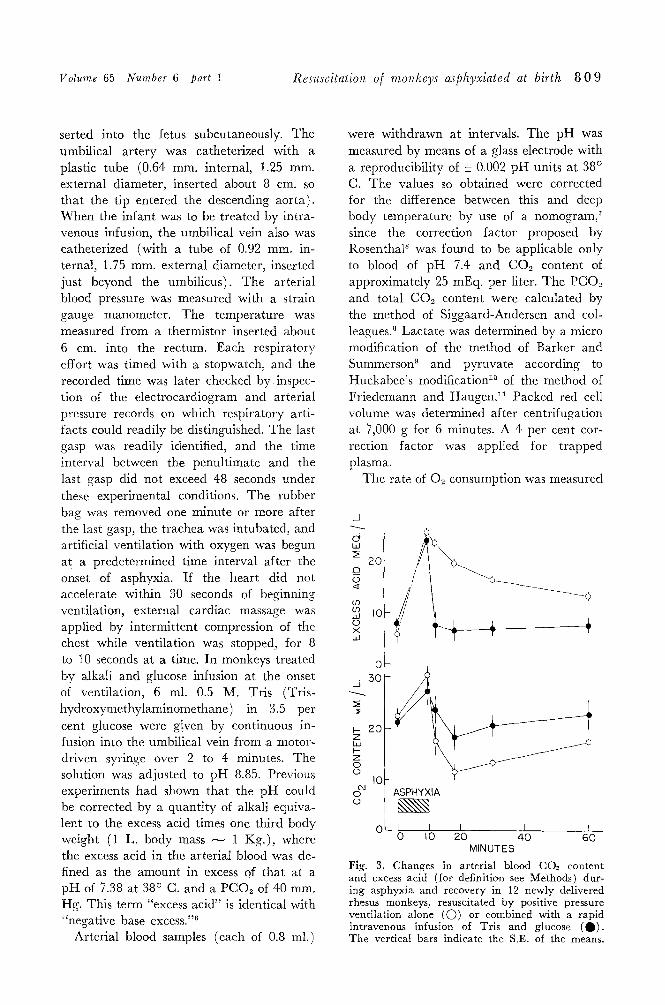
Volume 65 Number 6 part 1 Resuscitation o[ monkeys asphyxiated at birth 8 0 9
serted into the fetus subcutaneously. The umbilical artery was catheterized with a plastic tube (0.64 mm. internal, 1.25 mm. external diameter, inserted about 8 cm. so that the tip entered the descending aorta). When the infant was to be treated by intra- venous infusion, the umbilical vein also was catheterized (with a tube of 0.92 mm. in- ternal, 1.75 mm. external diameter, inserted just beyond the umbilicus). The arterial blood pressure was measured with a strain gauge manometer. The temperature was measured from a thermistor inserted about 6 cm. into the rectum. Each respiratory effort was timed with a stopwatch, and the recorded time was later checked b y inspec- tion of the electrocardiogram and arterial pressure records on which respiratory arti- facts could readily be distinguished. The last gasp was readily identified, and the time interval between the penultimate and the last gasp did not exceed 48 seconds under these experimental conditions. The rubber bag was removed one minute or more after the last gasp, the trachea was intubated, and artificial ventilation with oxygen was begun at a predetermined time interval after the onset of asphyxia. If the heart did not accelerate within 30 seconds of beginning ventilation, external cardiac massage was applied by intermittent compression of the chest while ventilation was stopped, for 8 to 10 seconds at a time. In monkeys treated by alkali and glucose infusion at the onset of ventilation, 6 ml. 0.5 M. Tris (Tris- hydroxymethylaminomethane) in 3.5 per cent glucose were given by continuous in- fusion into the umbilical vein from a motor- driven syringe over 2 to 4 minutes. The solution was adjusted to pH 8.85. Previous experiments had shown that the pH could be corrected by a quantity of alkali equiva- lent to the excess acid times one third body weight (1 L. body mass ~ 1 Kg.), where the excess acid in the arterial blood was de- fined as the amount in excess of that at a pH of 7.38 at 38 ~ C. and a PCO~ of 40 mm. Hg. This term "excess acid" is identical with "negative base excess. ''6
Arterial blood samples (each of 0.8 ml.)
were withdrawn at intervals. The pH was measured by means of a glass electrode with a reproducibility of + 0.002 pH units at 38 ~ C. The values so obtained were corrected for the difference between this and deep body temperature by use of a nomogram, 7 since the correction factor proposed by Rosenthal s was found to be applicable only to blood of pH 7.4 and CO2 content of approximately 25 mEq. per liter. The PCOu and total CO2 content were calculated by the method of Siggaard-Andersen and col- leagues2 Lactate was determined by a micro modification of the method of Barker and Summerson 9 and pyruvate according to Huckabee's modification 1~ of the method of Friedemann and Haugen. 11 Packed red cell volume was determined after centrifugation at 7,000 g for 6 minutes. A 4 per cent cor- rection factor was applied for trapped plasma.
The rate of 02 consumption was measured
.d
( 3 Lfl
s 0
o3
o3 I0 hl (_3 X I,I
20( �84
0
~z 2o
(.3 I 0 -
o ASPHYXIA
0 _ 1 I I I I 0 I0 20 40 60
MINUTES
Fig. 3. Changes in arterial blood CO~ content and excess acid (for definition see Methods) dur- ing asphyxia and recovery in 12 newly delivered rhesus monkeys, resuscitated by positive pressure ventilation alone (O) or combined with a rapid intravenous infusion of Tris and glucose (0) . The vertical bars indicate the S.E. of the means.
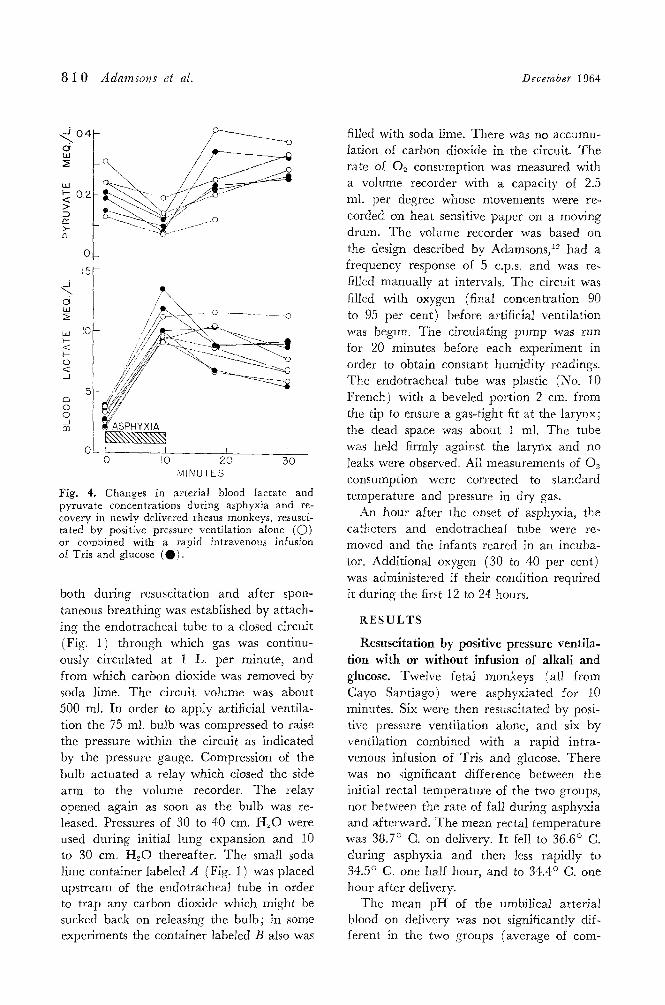
8 1 0 Adamsons et al. December 1964
0 . 4
a txl
w
~- 02 >.
gr" >- a .
o
~5 < d tad :2
W l 0 b--
t-- O
.-3
5 EB O O
tin
', I f I o IO 20 5o
MINUTES
Fig. 4. Changes in arterial blood lactate and pyruvate concentrations during asphyxia and re- covery in newly delivered rhesus monkeys, resusci- tated by positive pressure ventilation alone (O) or combined with a rapid intravenous infusion of Tris and glucose (0) .
both during resuscitation and after spon- taneous breathing was established by attach- ing the endotracheal tube to a closed circuit (Fig. 1) through which gas was continu- ously circulated at 1 L. per minute, and from which carbon dioxide was removed by soda lime. The circuit volume was about 500 ml. In order to apply artificial ventila- tion the 75 ml. bulb was compressed to raise the pressure within the circuit as indicated by the pressure gauge. Compression of the bulb actuated a relay which closed the side arm to the volume recorder. The relay opened again as soon as the bulb was re- leased. Pressures of 30 to 40 cm. H20 were used during initial lung expansion and 10 to 30 cm. H~O thereafter. The small soda lime container labeled A (Fig. 1) was placed upstream of the endotracheal tube in order to trap any carbon dioxide which might be sucked back on releasing the bulb; in some experiments the container labeled B also was
filled with soda lime. There was no accumu- lation of carbon dioxide in the circuit. The rate of 02 consumption was measured with a volume recorder with a capacity of 2.5 ml. per degree whose movements were re- corded on heat sensitive paper on a moving drum. The volume recorder was based on the design described by Adamsons, 12 had a frequency response of 5 c.p.s, and was re- filled manually at intervals. The circuit was filled with oxygen (final concentration 90 to 95 per cent) before artificial ventilation was begun. The circulating pump was run for 20 minutes before each experiment in order to obtain constant humidity readings. The endotracheaI tube was plastic (No. 10 French) with a beveled portion 2 cm. from the tip to ensure a gas-tight fit at the larynx; the dead space was about 1 ml. The tube was held firmly against the larynx and no leaks were observed. All measurements of 02 consumption were corrected to standard temperature and pressure in dry gas.
An hour after the onset of asphyxia, the catheters and endotracheat tube were re- moved and the infants reared in an incuba- tor. Additional oxygen (30 to 40 per cent) was administered if their condition required it during the first 12 to 24 hours,
R E S U L T S
Resuscitation by positive pressure ventila- tion with or without infusion of alkali and glucose. Twelve fetaI monkeys (all from Cayo Santiago) were asphyxiated for 10 minutes. Six were then resuscitated by posi- tive pressure ventilation alone, and six by ventilation combined with a rapid intra- venous infusion of Tris and glucose. There was no significant difference between the initial rectal temperature of the two groups, nor between the rate of fall during asphyxia and afterward. The mean rectal temperature was 38.7 ~ C. on delivery. It fell to 36.6 ~ C. during asphyxia and then less rapidly to 34.5 ~ C. one half hour, and to 34.4 ~ C. one hour after delivery.
The mean pH of the umbilical arterial blood on delivery was not significantly dif- ferent in the two groups (average of com-

Volume 65 Number 6 part 1 Resuscitation o[ monkeys asphyxiated at birth 8 1 1
bined means 7.30). During asphyxia the arterial p H fell below 6.8 on the average (Fig. 2). On resuscitation it rose more rapidly and to a higher vMue in monkeys which received an infusion of Tris and glu- cose ( Q ) . The arterial PCO2 rose from a mean of 43 mm. Hg to more than 150 mm. Hg during asphyxia (Fig. 2). On ventila- tion it fell rapidly, by about 100 mm. Hg within 2 minutes, in all the monkeys.
The CO2 content of the arterial blood rose during asphyxia (Fig. 3). On ventila- tion it fell very rapidly in monkeys which did not receive an infusion of Tris and glu- cose ((D); it fell significantly less rapidly in those which did ( ~ ) . During asphyxia the arterial excess acid (see Methods for defini- tion) rose at a rate of nearly 2 mEq. per liter per minute. On resuscitation by posi- tive pressure ventilation alone the excess acid fell slowly (Fig. 3, C)). In monkeys which also received an infusion of Tris and glucose the excess acid was rapidly neutralized (Fig.
3, 0 ) . The blood lactate and pyruvate concen-
trations were measured in some of the mon- keys (Fig. 4). The mean umbilical arterial lactate concentration on delivery was 3.1 mEq. per liter. I t rose rapidly during asphyxia and fell only slowly on resuscita- tion. The blood pyruvate concentration fell during asphyxia and rose quickly on re- suscitation. There was no significant differ- ence between the changes in the monkeys which received an infusion and those which did not. During asphyxia the lactate-pyru- vate ratio rose from a mean of 19 to 125; after resuscitation it fell rapidly. I t reached a value of 39 at one half hour and 34 at one hour after delivery.
The hematocrit increased from 41 to 44 per cent during asphyxia. In monkeys re- suscitated by ventilation alone the hemato- crit returned to the initial level within 10 minutes. In those which were given an in- fusion the mean hematocrit fell to 34 per cent in 2 minutes and to 32 per cent after 6 minutes. I t rose to 38 per cent by one half hour after delivery, and to 40 per cent by one hour.
The heart rate and mean arterial pres- sure in fetal monkeys before asphyxia are 180 to 200 beats per minute and 40 to 45 ram. Hg respectively. 2, 8 Both fell during asphyxia (Fig. 5). On resuscitation there was a prompt rise in heart rate and blood pressure in 6 of the 12 monkeys. In the other 6 (of which 4 received artificial venti- lation alone and the other 2 an infusion also) the heart did not accelerate within 30 seconds, external cardiac massage was ap- plied, and caused a prompt response (Fig. 6, upper section). On the average the ar- terial pressure rose more rapidly in the monkeys which were given an infusion as well as artificial ventilation (Fig. 5, Q ) , but the difference was not significant.
The mean time to the last gasp during asphyxia was not significantly different in the two groups of monkeys. The combined mean was 8.17 + 0.27 minutes (S.E.). After resuscitation the mean times to the first gasp were 2.60 +_ 0.46 in monkeys resusci- tated by ventilation alone and 1.83 + 0.32 minutes in those which were given an in-
BEATS 2O(
W I--
<~ 15( rr"
t-- n~ I00
W :]:
5O
MM. H( 60
w rr"
03 03 w 4-0 I:1:: Q._
J <I:
E: 20 w I--
<~
0
/MIN.
ASPHYXIA ~\\\\\\\\\\\\\\\\\\\\~ I I I I ~ _
0 5 IO 15 20 MINUTES
Fig. 5. Changes in arterial blood pressure and heart rate during asphyxia and recovery in 12 newly delivered rhesus monkeys, resuscitated by positive pressure ventilation alone ((D) or com- bined with a rapid intravenous infusion of Tris and glucose (Q). The vertical bars indicate the S.E. of the means.
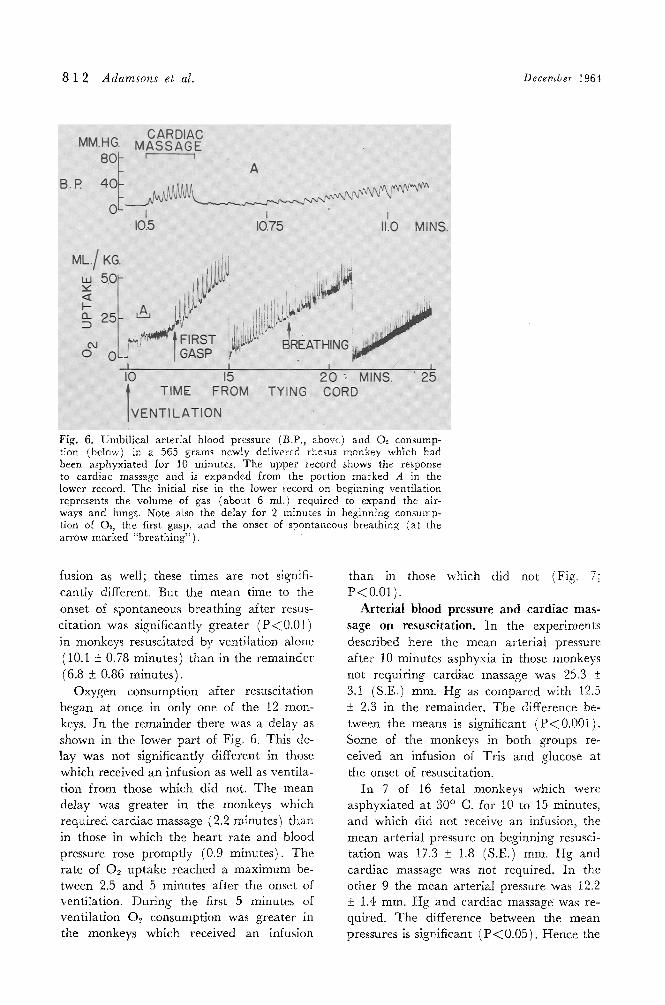
8 1 2 Adamsons et al. December 1964
CARDIAC M M . H G . M A S S A G E
80b r ~ . _
B . R 4 ' t , " ,~
' J t 1 E 10.5 IQ75 I1.0 MINS
ML/KG.
h- a . 25
(M O .rq
, , ,,ditl " t JI,,t~l:
L TIME FROM TYlNG CORD
VENTILATION
Fig. 6. Umbilical arterial blood pressure (B.P., above) and O~. consump- t{on (below) in a 565 grams newly delivered rhesus monkey which had been asphyxiated for 10 minutes. The upper record shows the response to cardiac massage and is expanded from the portion marked A in the lower record. The initial rise in the lower record on beginning ventilation represents the volume of gas (about 6 ml.) required to expand the air- ways and lungs. Note aim the delay for 2 minutes in beginning consump- tion of O~, the first gasp, and the onset of spontaneous breathing (at the arrow marked "breathing").
fusion as well; these times are not signifi- cant ly different. But the mean time to the
onset of spontaneous b rea th ing af ter resus- c i ta t ion was significantly grea te r ( P < 0 . 0 1 )
in monkeys resuscitated by vent i la t ion alone (10.I • 0.78 minutes) than in the r ema inde r
(6.8 + 0.86 minutes) .
Oxygen consumption after resuscitation began at once in only one of the 12 mon- keys. I n the remainder there was a delay as shown in the lower pa r t of Fig. 6. This de- lay was not significantly different in those
which received an infusion as well as vent i la- t ion f rom those which did not. T h e mean de lay was grea ter in the monkeys which requi red ca rd iac massage (2.2 minutes) than in those in which the hea r t ra te and blood pressure rose p rompt ly (0.9 minutes) . T h e ra te of 02 up take reached a m a x i m u m be- tween 2.5 and 5 minutes after the onset of ventilation. Dur ing the first 5 minutes of vent i la t ion O2 consumpt ion was grea te r in the monkeys which received an infusion
than in those which did not {Fig. 7: P<0.01).
Arte r i a l blood pressure and cardZac mas- sage on resuscitat ion. In the exper iments
described here the mean ar ter ia l pressure af ter 10 minutes asphyxia in those monkeys
not requi r ing ca rd iac massage was 25,3 +_ 3. I (S.E.) mm. H g as compared with 12.5 _+ 2.3 in the remainder . T h e difference be- tween the means is significant ( P < 0 . 0 0 1 ) . Some of the monkeys in both groups re- ceived an infusion of Tr is and glucose at the onset of resuscitation.
I n 7 of 16 fetal monkeys which were asphyxia ted at 30 ~ C. for 10 to 15 minutes, and which did not receive an infusion, the mean ar te r ia l pressure on beginning resusci- ta t ion was 17.3 • 1.8 (S.E.) mm. H g and card iac massage was not required. In the o ther 9 the mean ar te r ia l pressure was 12.2 • 1.4 ram. H g and card iac massage was re- quired. The difference between the mean pressures is s ignif icant ( P < 0.05). Hence the

Volume 65 Number 6 part 1 Resuscitation o[ monkeys asphyxiated at birth 8 1 3
need to apply cardiac massage was also related to the arterial pressure on resuscita- tion in the absence of any infusion.
The relation between duration of asphyxia and recovery of respiratory activity after resuscitation. In 16 fetal monkeys asphyxi- ated for 10, 12.5, or 15 minutes (which were not given an infusion of alkali and glucose) the mean time from the onset of asphyxia until the last gasp was 8.42 + 0.24 (S.E.) minutes. Six were ventilated with 02 after 10 minutes and 5 each after 12.5 and 15 minutes asphyxia. They were given car- diac massage if required, as described, but received no other treatment. In Fig. 8 are shown the means ( O ) and S.E. (vertical lines) of the observations in each category. There is an approximately linear relation- ship between the duration of asphyxia and the onset of gasping and of rhythmic spon- taneous breathing. The individual measure- ments were used to calculate the regression lines. The time to last gasp is not signifi- cantly different from the intercept on the abscissa of the regression line relating time to first gasp (after beginning resuscitation) to the duration of asphyxia (Fig. 8, broken line). For each minute of delay from the time of the last gasp to the onset of arti-
ficial ventilation, the time to the first gasp was increased by about 2 minutes. The re- gression line relating time to spontaneous breathing (after beginning resuscitation) to the duration of asphyxia (Fig. 8, solid line) intercepts the abscissa at 7.76 minutes, which is less than the time to last gasp. Hence one may conclude that had artificial ventilation been started at the last gasp, gasping would have continued but spontaneous breathing would not have been established for an~ other 3 or 4 minutes even under the most favorable conditions. For each additional minute delay in beginning resuscitation, the start of rhythmical breathing would be de- layed by more than 4 minutes on the aver- age.
Asphyxia and respiratory distress. Respir- atory distress and pulmonary edema were encountered frequently during the recovery period. In monkeys which recovered without respiratory distress the respiratory rate rose to 60 to 100 per minute and the accessory muscles of respiration were used very little. The first sign of impending difficulty was retraction of the lower end of the sternum during inspiration, within 5 to 10 minutes of the establishment of spontaneous breath- ing. During the next 30 to 60 minutes
ML./K G. MIN.
z I0 ~ T CONTROL
o
Z O O
#
I0 20 50 IO 20 50 MINS.
TIME FROM TYING CORD
Fig. 7. Changes in 02 consumption in 12 rhesus monkeys asphyxiated for 10 minutes after delivery. Six received an infusion of alkali and glucose ("infused") at the onset of ventilation (indicated by the arrows), while six were ventilated alone ("control"). The vertical bars indicate the S.E. of the means. The mean 02 consumption of the infused groups during the first 5 minutes' resuscitation was sig- nificantly greater than that of the control group (P < 0.01).

8 1 4 Adamsons et aI. December 1964
MINS. ITIME TO z 50 / I BREATHING o I -
V- 20 z taJ
> , TO FIRST / ;GASP rr I 0 / / / h
hi
I--" I
0 5 " I 0 15 MINS.
DURATION OF A S P H Y X I A
Fig. 8. Time from beginning positiye pressure ventilation with O5 until the first gasp (---) and until the establishment of rhythmic breathing (--) in newly delivered rhesus monkeys asphyxi- ated for 10, 12.5, or 15 minutes at 30 ~ C. The vertical bars indicate the S.E. of the means in each group of 5 or 6 monkeys. None received any infusion.
respiratory efforts became increasingly forceful, with deep recession of the sternum and lower costal margins, grunting expira- tion, and pallor. The respiratory rate rose to 110 to 160 per minute. As the condition grew worse, if the monkey was not given high 02 mixtures to breathe, the respiratory rate decreased, and there were cyanotic at- tacks followed by gasping or irregular periodic breathing. In 3 monkeys that weighed less than 400 grams and in one of 520 grams frank pulmonary edema de- veioped (as evidenced by the appearance of pink froth in the endotracheal tube).
The rate of 02 consumption, measured in a neutral thermal environment at 35 ~ C., was almost always less in monkeys with respiratory distress, in spite of the vigorous movements of the respiratory muscles, than in asymptomatic animals after resuscitation from asphyxia. It ranged from 4.5 to 8.0 ml. per kilogram-minute in 13 monkeys with respiratory distress while breathing air as compared with normal values of 8 to 12 ml. per kilogram-minute. When the 02 con- tent of the inspired gas was raised, 02 con- sumption increased, and this proved a useful
quantitative measure of therapeutic effect. Oxygen consumption always rose to normal values before the signs of respiratory distress subsided. Recovery, when it occurred, took place over a period of a few hours to 2�89 days. In those that died, the lungs were collapsed and airless at autopsy. Microscopic examination revealed hyaline membranes similar to those seen in human infants (Fig. 9). No difficulty was experienced in the initial expansion of the lungs of any of the animals, whether or not they subseqeuntly developed respiratory distress, and in no instance has positive pressure ventilation caused a pneumothorax in 68 monkeys of 132 to 162 days' gestation (term is approxi- mately 168 days).
Table I shows that the incidence of pul- monary complications after asphyxia on de- livery was greater in the monkeys with lower birth weight. Of 13 monkeys weighing more than 500 grams 5 developed respira- tory distress and/or pulmonary edema and all recovered. Of 13 monkeys weighing less than 400 grams 9 developed respiratory complications and 7 died within 24 hours. This incidence may be compared with that after delivery by cesarean section without asphyxia in the same laboratory (Windle, personal communication). Of 20 monkeys weighing 400 grams or more, none de- veloped respiratory distress; of 7 weighing less than 400 grams, 2 very immature mon- keys developed respiratory distress. It may, therefore, be concluded that asphyxia in monkeys greatly increases the incidence of this condition.
Infusion of alkali (Tris at pH 8.85, sodium carbonate or sodium bicarbonate) and glu- cose during asphyxia or on resuscitation did not prevent the development of respiratory distress, the incidence in the treated and control groups being similar. The numbers are, however, small (Table II) . Of 15 monkeys which received no infusion, 11 re- quired cardiac massage as well as artificial ventilation: of these eleven, 5 developed respiratory distress and 6 did not. The num- ber of untreated monkeys was insufficient to indicate whether the incidence of pulmonary

Volume 65 Number 6 part 1 Resuscitation o/ monkeys asphyxiated at birth 8 1 5
�9 complications increased as the duration of asphyxia was extended from 10 to 12.5 or 15 minutes.
D I S C U S S I O N
Before considering the effect of an in- fusion of alkali and glucose, in addition to ventilation, on the course of recovery after asphyxia, we would like to emphasize the quantitative relation between the duration of asphyxia and the rate of recovery. In fetal rhesus monkeys which are resuscitated before the last gasp (i.e., after less than 8.4 minutes asphyxia on the average) per- manent cerebral damage is either absent or trivial. In monkeys resuscitated after as- phyxia for 12.5 minutes the damage is widespread and sometimes accompanied by overt signs of neurologic disorder? The evi- dence reported in the present experiments shows that each minute which is allowed to elapse after the last gasp, before be- ginning resuscitation, entails a large and linearly related increase in the time re- quired to establish breathing even under the most favorable conditions, when positive pressure ventilation is given with 100 per cent 02 and recovery of the circulation is assisted by cardiac massage when necessary. I t is evident that even a small prolongation of asphyxia leads to rapidly increasing im- pairment of the mechanism of breathing.
When alkali and glucose were infused during the course of asphyxia there was a significant reduction in the time from be- ginning resuscitation until the first gasp, and the time required to ' establish spontaneous breathing was decreased by 65 per cent 2 or 72 per cent 3 on the average. In the present experiments, in which alkali and glucose were infused over the first 2 to 4 minutes after beginning ventilation, the time to the first gasp was not significantly reduced, which is hardly surprising as it was only 2.5 minutes in monkeys which did not receive any infusion. The time required to establish spontaneous breathing was decreased by 33 per cent.
I t is evident that asphyxia does not end as soon as ventilation is begun. Even when
the heart rate and blood pressure rise swiftly the rate of 02 uptake does not at once increase to the normal level. During this period restoration of the arterial p H to normal would make more energy avail- able by glycolysis, and enable the brain and myocardium to recover more rapidly. Other experiments have shown that rapid intra- venous infusion of alkali and glucose after the last gasp, in the absence of ventilation, causes a rapid increase in blood pressure and gasping begins (Daniel, Dawes, James, and Ross, unpublished). Also, in the presence of adequate oxygenation, a fall in arterial pH is known to reduce the pressor response to epinephrine 13,1~ and myocardial con- tractility. 15
There are other possible means by which
Fig. 9. Section of lung stained with hematoxylin and eosin from rhesus monkey, 151 days gesta- tional age, asphyxiated for 15 minutes at birth, which developed respiratory distress and died 26 hours after delivery. Note the presence of-ex- tensive atelectasis and hyaline membranes. (Origi- nal magnification x350.)

8 1 6 Adamsons et al. December 1964
Table I. Incidence of respiratory distress and/or pulmonary edema in monkeys asphyxiated on delivery for 10 to 15 minutes
Weight Total (grams) No.
)500 13 400 to 500 17
~400 13
No. with No. pulmonary dying in
complications 24 hours
5 0 7 2 9 7
Table II. Incidence of pulmonary complications in monkeys asphyxiated on delivery for 10 to 15 minutes and either untreated or given alkali and glucose
Given alkali and glucose ~
Weight During During re- (grams) Untreated asphyxia suscitation
) 5 0 0 3/6 0/2 2/5 400 to 500 3/6 4/11
%400 2/3 2/5 5/5 "~Either 0.5M Tris, 0 .7M sodimn carbonate or 0.6M so-
dimn bicarbonate, each with 3 to 7.5 per cent glucose.
infusion of alkali and glucose might facilitate recovery. Thus, observations on both mature and premature fetal lambs have shown that acute asphyxia causes intense pulmonary vasoconstriction~6, ~r as also does a ri~e in arterial PCO2 in both unventilated is and ventilated lungs. 19 A rapid elevation in arterial pH, with a consequent fall in PCO2, may contribute to pulmonary vasodilatation during resuscitation. The effect of pH on the hemoglobin-oxygen dissociation curve (the Bohr effect) could be a subsidiary fac- tor. As the pig is increased the 02 affinity of hemoglobin is increased. At a PO2 of 30 mm. Hg and pH 6.9 the blood of a fetal monkey is 48 per cent saturated, whereas at pig 7.4 it is 83 per cent saturated. 2~ This may partly explain the very rapid increase in 02 uptake in those monkeys which were given Tris on resuscitation. The absolute rise in O., uptake above the average level of about 8 ml. per kilogram-minute was 15 ml. per kilogram, which would be almost sufficient to saturate the circulating blood completely.
The similarity between the condition of
respiratory distress as seen in monkeys (and in lambs), delivered by cesarean section and asphyxiated, and that observed in the naturally occurring disease of the human infant has already been remarked upon?' 2 Respiratory distress has occurred in 21 of 43 monkeys, an incidence of almost 50 per cent. The incidence was significantly greater in monkeys of lower birth weight (Table I). All the monkeys, whether asphyxiated for 10, 12.5, or 15 minutes, were in acute cir- culatory failure when resuscitation was be- gun, and most (including all those which received no infusion) had ceased gasping many minutes previously. Nevertheless, in 11 monkeys which received no infusion and which required cardiac massage as well as artificial ventilation, respiratory distress was observed on recovery in only 5. It was not seen in 2 monkeys whose hearts had stopped beating, one for more than 4 minutes. Hence, respiratory distress is not an invariable con- sequence of extreme acute asphyxia on de- livery. The evidence indicates that asphyxia at birth could be a causal factor, in view of the low incidence in animals which were not asphyxiated at birth. Restoration of arterial pH either during or after asphyxia did not prevent its development.
The object of the original research was to determine whether it was possible to pro- long survival during asphyxia at birth by infusion of alkali and glucose. When this proved possible both as to the cardiovascular and respiratory systems, in monkeys as well as lambs, it seemed worth while to explore the therapeutic possibilities. Sodium carbon- ate proved to be too alkaline, sodium bi- carbonate was relatively ineffective in the volumes which might reasonably be given to newborn monkeys, but Tris was effective and safe in the doses used. 3 Adequate venti- lation restored the arterial pH and PCO._, almost as rapidly as infusion of alkali, but alkaline infusion combined with ventilation established spontaneous rhythmic breathing sooner.
Tris has now been given as a rapid in- travenous infusion, in doses of 5.0 to 7.7 raM. per kilogram to 16 newly delivered

Volume 65 Number 6 part 1 Resuscitation of monkeys asphyxiated at birth 8 1 7
monkeys weighing 310 to 570 grams, during asphyxia or on resuscitation. I t did not appear to cause respiratory depression in these doses, unlike unasphyxiated adult ani- mals? 1 I t might be wise, in practice, not to try to correct the arterial p H fully by infusion of Tris. In all but one monkey respiratory function, as judged by return of gasping and the onset of spontaneous rhyth- mic breathing, was achieved in a shorter period of time in treated animals than in those resuscitated by ventilation alone. In the one animal in which the onset of spon- taneous breathing was delayed, the circula- tion had collapsed exceptionally early dur- ing asphyxia.
The asphyxia to which these monkeys have been subjected was both acute and prolonged. Sufficient experimental experi- ence has now been accumulated to justify a cautious clinical trial of this method of t reatment in severely asphyxiated h u m a n in- fants, combined with artificial ventilation and, if necessary, cardiac massage.
S U M M A R Y
1. Fetal rhesus monkeys were delivered by cesarean section under local anesthesia and were asphyxiated under controlled en- vironmental conditions. They were resusci- tated by artificial ventilation with 02 and by cardiac massage, if necessary.
2. Rapid intravenous infusion of alkali (0.5M Tris) with glucose on resuscitation reduced the time required to establish rhyth- mic breathing, and increased the immediate Oz uptake.
3. The effect of asphyxia for 10, 12.5, or 15 minutes on subsequent resuscitation was analyzed. The mean time f rom the onset of asphyxia until the last gasp was 8.42 minutes. For each minute after the last gasp, the interval f rom beginning resuscitation until the first gasp was more than 2 minutes, and until the establishment of rhythmical breathing it was more than 4 minutes.
4. Respiratory distress a n d / o r pu lmonary edema were observed in 2t of 43 monkeys asphyxiated for 10 to 15 minutes on de- livery. The incidence was higher in the
monkeys with the lower birth weights. In- fusion of alkali and glucose during asphyxia or on resuscitation did not prevent the de- velopment of respiratory distress.
This work was carried out with the aid of a grant from the United Cerebral Palsy Research and Educational Foundation for which we are most grateful. We also acknowledge the help of United States Public Health Service Grants A-4359 and RG-9069. We would like to thank the National Institute for Neurological Disease and Blindness and in particular Dr. W. F. Windle, for their hospitality in Playa Humacao, Puerto Rico, and the staff of the Ecology section of the laboratory of Perinatal Physiology for its help. We are also very grateful to Dr. Salha Daniel and Dr. Gillian Gandy for expert assistance.
R E F E R E N C E S
1. Dawes, G. S., Mott, J. C., and Stafford, A.: Prolongation of survival in the anoxic foetal lamb, J. Physiol. 153: 16, 1960.
2. Dawes, G. S., Jacobson, H. N., Mott, J. C., Shelley, H. J., and Stafford, A.: The treat- ment of asphyxiated, mature foetal lambs and rhesus monkeys with intravenous glucose and sodium carbonate, J. Physiol. 169: 167, 1963.
3. Adamsons, K., Behrman, R., Dawes, G. S., Dawkins, M. J. R., James, L. S., and Ross, B. B.: The treatment of acidosis with alkali and glucose during asphyxia in foetal rhesus monkeys, J. Physiol. 169: 679, 1963.
4. Dawes, G. S., Hibbard, E., and Windle, W. F.: The effect of alkali and glucose in- fusion on permanent brain damage in rhesus monkeys asphyxiated at birth, J. PEDIAT. 65: 801, 1964.
5. Carpenter, C. R.: Sexual behavior (~f free ranging rhesus monkeys (Macaea Mulatta), J. Comp. Psychol. 33: 113, 1942.
6. Siggaard-Andersen, O., Engle, K., Jorgensen, K., and Astrup, P.: A micro method for de- termination of pH, carbon dioxide tension, base excess and standard bicarbonate in capillary blood, Scandinav. J. Clin. & Lab. Invest. 12: 172, 1960.
7. Adamsons, K., Daniel, S. S., Gandy , G., and James, L. S.: The influence of temperature on blood pH of the human adult and new- born, J. Appl. Physiol. (in press).
8. Rosenthal, T. B.: The effect of temperature on the pH of blood and plasma in vitro, J. Biol. Chem. 173: 25, i948.
9. Barker, S. B., and Summerson, W. H.: The colorimetric determination of lactic acid in

8 1 8 Adamsons et al. December i964
biological material, J. Biol. Chem. 188: 535, 1941.
10. Huckabee, W. E.: Control of concentration gradients of pyruvate and lactate across cell membranes in blood, J. Appl. Physiol. 9: I63, 1956.
11. Friedemann, T. D., and Haugen, G. E.: Pyruvic acid. The determination of keto acids in blood and urine, J. Biol. Chem. 147: 415, 1943.
12. Adamsons, K.: Breathing and the thermal environment in young rabbits, J. Physiol. 149: 144, 1959.
13. Burget, G. E., and Visscher, M. B.: Variation of the pH of the blood and the response of the vascular system to adrenalin, Am. J. Physiol. 81: 113, 1927.
14. Wood, W. B., Manley, E. S., and Wood- bury, R. A.: The effects of CO~-induced respiratory acidosis in the depressor and pres- sor components of the dog's blood pressure response to epinephrine, J. Pharmacol. 139: 238, 1963.
15. Darby, T. D.: Effects of 2 amino-2-hy- droxymethyl-l, 3 propanediol during shock
and catecholamine administration, Ann. N. Y. Acad. Sc. 92: 674, 1961.
16. Dawes, G. S., and Mott, J. C.: The vascular tone of the foetal lung, J. Physiol. 164: 465, 1962.
17. Cassin, S., Dawes, G. S., and Ross, B. B.: Pulmonary blood flow and vascular resistance in immature foetal lambs, J. Physiol. 171: 80, 1964.
18. Cassin, S., Dawes, G. S., Mott, J. C., Ross, B. B., and Strang, L. B.: The vascular re- sistance of the foetal and newly-ventilated lung of the lamb, J. Physiol. 171: 61, 1964.
19. Cook, C. D., Drinker, R. A., Jacobson, H. N., Levison, H., and Strang, L. B.: Control of pulmonary blood flow in the foetal and newly born lamb, J. Physiol. 169: 10, 1963.
20. Behrman, R. E., Heller, C. J., Battaglia, F. C., and Hellegers, A. E.: A comparison of the oxygen affinity of maternal and fetal blood of the macaca mulatta, Quart. J. Exper. Physiol. 48: 258, 1963.
21. Nahas, G. G.: The pharmacology of Tris (hydroxymethyl) aminomethane (Tham), Pharmacol. Rev. 14: 447, 1962.




















