
IL-2 immunotherapy in chronically SIV-infected Rhesus Macaques
Upload
meduniwienCategory
view
1download
0
lable at ScienceDirect
Neuropharmacology 59 (2010) 612e618
Contents lists avai
Neuropharmacology
journal homepage: www.elsevier .com/locate/neuropharm
Anxiolytic-like effects of 8-acetylene imidazobenzodiazepinesin a rhesus monkey conflict procedure
Bradford D. Fischer a,*, Stephanie C. Licata a, Rahul V. Edwankar b, Zhi-Jian Wang b, Shengming Huang b,Xiaohui He b, Jianming Yu b, Hao Zhou b, Edward M. Johnson Jr. b, James M. Cook b, Roman Furtmüller c,Joachim Ramerstorfer c, Werner Sieghart c, Bryan L. Roth d, Samarpan Majumder d, James K. Rowlett a
aHarvard Medical School, New England Primate Research Center, One Pine Hill Drive, P.O. Box 9102, Southborough, Massachusetts 01772-9102, USAbUniversity of Wisconsin-Milwaukee, Department of Chemistry and Biochemistry, Milwaukee, Wisconsin, USAcMedical University of Vienna, Center for Brain Research, Vienna, AustriadUniversity of North Carolina at Chapel Hill, Department of Pharmacology, Chapel Hill, North Carolina, USA
a r t i c l e i n f o
Article history:Received 26 May 2010Received in revised form19 July 2010Accepted 10 August 2010
Keywords:BenzodiazepineGABAAnxietyAnxiolyticRhesus monkeyConflict
* Corresponding author. Tel.: þ1 508 624 8146; faxE-mail address: [email protected]
0028-3908/$ e see front matter � 2010 Elsevier Ltd.doi:10.1016/j.neuropharm.2010.08.011
a b s t r a c t
Conflict procedures can be used to study the receptor mechanisms underlying the anxiolytic effects ofbenzodiazepines and other GABAA receptor modulators. In the present study, we first determined theefficacy and binding affinity of the benzodiazepine diazepam and recently synthesized GABAA receptormodulators JY-XHe-053, XHe-II-053, HZ-166, SH-053-20F-S-CH3 and SH-053-20F-R-CH3 at GABAA
receptors containing a1, a2, a3 and a5 subunits. Results from these studies suggest that each compounddisplayed lower efficacy at GABAA receptors containing a1 subunits and varying degrees of efficacy andaffinity at GABAA receptors containing a2, a3 and a5 subunits. Next, we assessed their anxiolytic effectsusing a rhesus monkey conflict procedure in which behavior was maintained under a fixed-ratioschedule of food delivery in the absence (non-suppressed responding) and presence (suppressedresponding) of response-contingent electric shock. Relatively non-selective compounds, such as diaz-epam and JY-XHe-053 produced characteristic increases in rates of suppressed responding at low tointermediate doses and decreased the average rates of non-suppressed responding at higher doses. XHe-II-053 and HZ-166 also produced increases in suppressed responding at low to intermediate doses, butwere ineffective at decreasing rates of non-suppressed responding, consistent with their relatively lowefficacy at GABAA receptors containing a1 and a5 subunits. In contrast, SH-053-20F-S-CH3 and SH-053-20F-R-CH3 produced only partial increases in suppressed responding and were ineffective on non-suppressed responding, consistent with their profiles as partial agonists at GABAA receptors containinga2, a3 and a5 subunits. These behavioral effects suggest that the anxiolytic and rate-reducing effects ofGABAA receptor positive modulators are dependent on their relative efficacy and affinity at differentGABAA receptor subtypes.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Benzodiazepine-type drugs act as positive allosteric modulatorsof g-aminobutyric acid type A (GABAA) receptors, and are highlyefficacious agents for the treatment of anxiety-related disorders.The therapeutic use of benzodiazepine-type drugs is constrained,however, by other characteristic effects such as daytime drowsi-ness, impairment of motor coordination, memory deficits, andreinforcing effects that may contribute to their abuse (Griffiths andWeerts, 1997; Nutt, 2005). Research during the past two decades
: þ1 508 624 8197.u (B.D. Fischer).
All rights reserved.
has revealed the existence of multiple subtypes of the GABAAreceptor (e.g., Pritchett et al., 1989; Rudolph et al., 2001; Olsen andSieghart, 2008). Subsequent reports have postulated that thediverse behavioral effects of benzodiazepine-type drugsmay reflectactions at different subtypes of GABAA receptors (Rudolph et al.,1999; McKernan et al., 2000; Löw et al., 2000; Rowlett et al.,2005). Therefore, it may be possible to dissociate the clinicallyadvantageous and unwanted side-effects of these compounds.
GABAA receptors are pentameric proteins composed of severalsubunits that form a GABA-gated chloride channel. The majority ofGABAA receptors are composed of a, b, and g subunits and benzo-diazepines bind predominantly to a site on the native GABAAreceptor that is located at the interface of the g2 subunit and one ofthe a1, a2, a3, or a5 subunits. Benzodiazepines generally are

B.D. Fischer et al. / Neuropharmacology 59 (2010) 612e618 613
inactive at corresponding a4- and a6-subunit containing receptors.Approximately 90% of the GABAA receptors in the brain that possessa benzodiazepine binding site contain a1, a2, and a3 subunits(McKernan andWhiting, 1996), and GABAA receptors containing a1subunits (a1GABAA receptors) have been implicated in the sedativeeffects of benzodiazepines, whereas GABAA receptors containing a2and a3 subunits (a2GABAA and a3GABAA receptors) have beenimplicated in the anxiolytic effects of benzodiazepines (McKernanet al., 2000; Löw et al., 2000; Rowlett et al., 2005; Dixon et al.,2008). GABAA receptors containing a5 subunits (a5GABAA recep-tors), in contrast, are a relatively minor population that may playa role in memory processes, but not anxiolysis or motor effects(Collinson et al., 2002; Crestani et al., 2002; Atack et al., 2006, butsee Savi�c et al., 2008). Therefore, novel benzodiazepine-like drugsthat have pharmacological selectivity for a2GABAA and/or a3GABAAreceptors and low receptor activity at a1GABAA and a5GABAAreceptors may be particularly useful as anxiolytics lacking sedativeand amnestic side effects.
The preclinical assessment of the anxiolytic effects of drugs canbe accomplished objectively and quantitatively with operant-basedconflict procedures. In these procedures, positively reinforcedbehavior is suppressed by response-contingent administration ofa noxious stimulus (e.g., mild electric shock; for review, see Millan,2003). Drugs with anxiolytic effects produce characteristicincreases in the rates of responding that are suppressed byresponse-contingent delivery of shock (e.g., Geller and Seifter,1960; Cook and Davidson, 1973; Kleven and Koek, 1999; Rowlettet al., 2006), and a particular strength of conflict procedures istheir predictive validity with respect to therapeutic effects inhumans. In this regard, strong positive correlations between thepotency of benzodiazepines to engender anti-conflict effects and tobe clinically effective in humans have been demonstrated in ratsand pigeons (Cook and Davidson,1973; Kleven and Koek,1999), andmore recently in rhesus monkeys (Rowlett et al., 2006). A distinctadvantage in using rhesus monkeys in a conflict procedure arisesfrom their close genetic similarity to humans.
In the present study, we first assessed the efficacy and bindingaffinity of the conventional benzodiazepine diazepam and therecently synthesized and structurally related positive GABAAreceptor modulators JY-XHe-053, XHe-II-053 and HZ-166. Theefficacy and binding affinity of the 4-methyl-JY-XHe-053
Fig. 1. Chemical structures of JY-XHe-053, XHe-II-053,
stereoisomers SH-053-20F-S-CH3 and SH-053-20F-R-CH3 was alsoassessed. Each of the novel GABAA receptor modulators demon-strated lower efficacy at a1GABAA receptors relative to diazepam atthe drug concentrations that can be reached under our experi-mental conditions. Further, each compound demonstrated varyingdegrees of either binding selectivity or relative efficacy fora2GABAA, a3GABAA and a5GABAA receptor subtypes. The primaryaim of the present study was to help elucidate the role of thesereceptor subtypes in the anxiolytic effects of benzodiazepines.Therefore, we used a rhesus monkey conflict procedure to assessthe anti-conflict and rate-reducing effects of these drugs. Based onstudies suggesting that benzodiazepine action at a1GABAA recep-tors contributes to their sedative effects, whereas a2GABAA anda3GABAA receptors are important for benzodiazepine-inducedanxiolysis, our hypothesis was that the novel GABAA receptormodulators would have anti-conflict effects similar to diazepambut would not be effective in disrupting rates of non-suppressedresponding.
2. Materials and methods
2.1. Drugs
JY-XHe-053 (8-ethynyl-6-(2-fluorophenyl)-4H-2,5,10b-triaza-benzo[e]azulene-3-carboxylic acid ethyl ester), XHe-II-053 (8-ethynyl-6-phenyl-4H-2,5,10b-triaza-benzo[e]azulene-3-carboxylic acid ethyl ester), HZ-166 (8-ethynyl-6-(20-pyridine)-4H-2,5,10b-triaza-benzo[e]azulene-3-carboxylic acid ethyl ester), SH-053-S-CH3-20F (the S-enan-tiomer of 4-methyl-JY-XHe-053; (S)-8-ethynyl-6-(2-fluoro-phenyl)-4-methyl-4H-2,5,10b-triaza-benzo[e]azulene-3-carboxylic acid ethyl ester) and SH-053-R-CH3-20F(the R-enantiomer of 4-methyl-JY-XHe-053; (R)-8-ethynyl-6-(2-fluoro-phenyl)-4-methyl-4H-2,5,10b-triaza-benzo[e]azulene-3-carboxylic acid ethyl ester) (Cook et al.,2009; Fig. 1) were synthesized at the Department of Chemistry and Biochemistry,University of Wisconsin-Milwaukee. JY-XHe-053, XHe-II-053, HZ-166, SH-053-20F-S-CH3 andSH-053-20F-R-CH3havea similar pharmacokinetic profile anddurationof action(Rivas et al., 2009; Cook et al., 2010). Diazepam was purchased from Tocris-Cookson(Ellisville, MO, USA). All drugs were dissolved in 20% ethanol, 60% propylene glycol, and20% sterile water. If necessary, the pH of a solutionwas adjusted to 7.0 with 1 N HCl.
2.2. Competition binding assays
Competition binding assays were performed in a total volume of 0.5 mL at 4 �Cfor 1 hour using [3H]flunitrazepam as the radiolabelled ligand. A total of 6 mg ofcloned human GABAA receptor DNA containing desired a subtype along with b2 andg2 subunits were used for transfecting HEK 293 T cell line using Fugene 6 (RocheDiagnostic) transfecting reagent. Cells were harvested 48 h after transfection,washed with TriseHCl buffer (pH 7.0) and Tris Acetate buffer (pH 7.4) and resulting
HZ-166, SH-053-20F-S-CH3 and SH-053-20F-R-CH3.
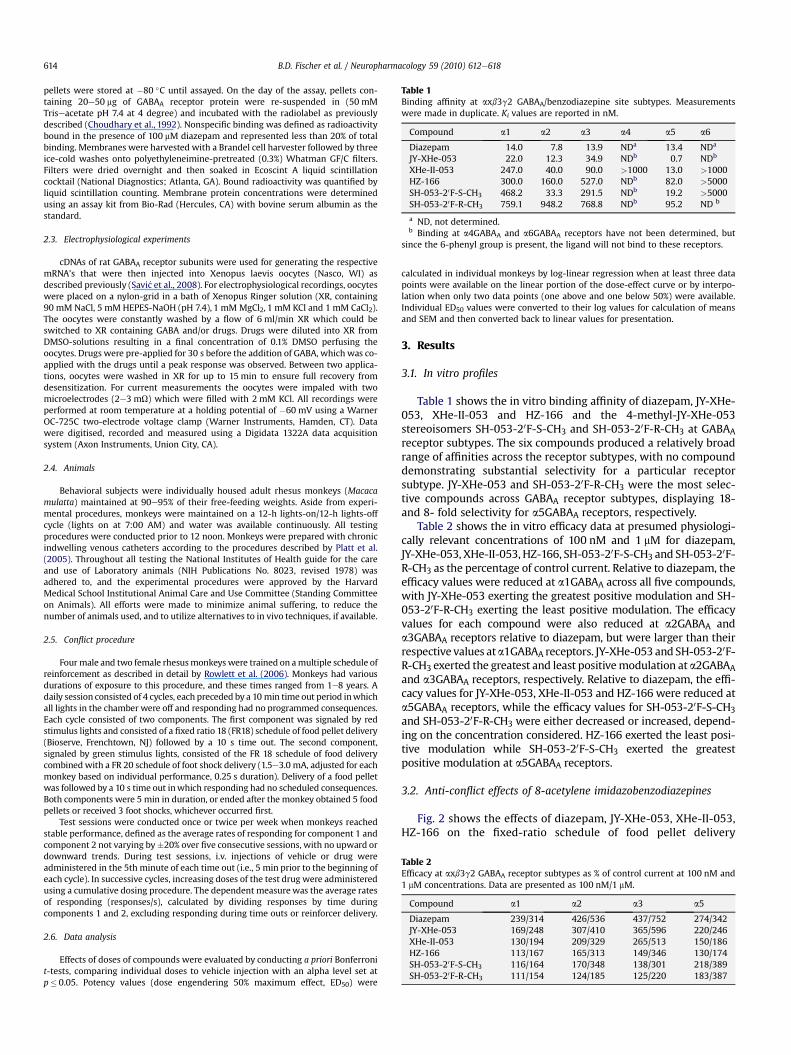
Table 1Binding affinity at axb3g2 GABAA/benzodiazepine site subtypes. Measurementswere made in duplicate. Ki values are reported in nM.
Compound a1 a2 a3 a4 a5 a6
Diazepam 14.0 7.8 13.9 NDa 13.4 NDa
JY-XHe-053 22.0 12.3 34.9 NDb 0.7 NDb
XHe-II-053 247.0 40.0 90.0 >1000 13.0 >1000HZ-166 300.0 160.0 527.0 NDb 82.0 >5000SH-053-20F-S-CH3 468.2 33.3 291.5 NDb 19.2 >5000SH-053-20F-R-CH3 759.1 948.2 768.8 NDb 95.2 ND b
a ND, not determined.b Binding at a4GABAA and a6GABAA receptors have not been determined, but
since the 6-phenyl group is present, the ligand will not bind to these receptors.
Table 2Efficacy at axb3g2 GABAA receptor subtypes as % of control current at 100 nM and1 mM concentrations. Data are presented as 100 nM/1 mM.
Compound a1 a2 a3 a5
Diazepam 239/314 426/536 437/752 274/342JY-XHe-053 169/248 307/410 365/596 220/246XHe-II-053 130/194 209/329 265/513 150/186HZ-166 113/167 165/313 149/346 130/174SH-053-20F-S-CH3 116/164 170/348 138/301 218/389SH-053-20F-R-CH3 111/154 124/185 125/220 183/387
B.D. Fischer et al. / Neuropharmacology 59 (2010) 612e618614
pellets were stored at �80 �C until assayed. On the day of the assay, pellets con-taining 20e50 mg of GABAA receptor protein were re-suspended in (50 mMTriseacetate pH 7.4 at 4 degree) and incubated with the radiolabel as previouslydescribed (Choudhary et al., 1992). Nonspecific binding was defined as radioactivitybound in the presence of 100 mM diazepam and represented less than 20% of totalbinding. Membranes were harvested with a Brandel cell harvester followed by threeice-cold washes onto polyethyleneimine-pretreated (0.3%) Whatman GF/C filters.Filters were dried overnight and then soaked in Ecoscint A liquid scintillationcocktail (National Diagnostics; Atlanta, GA). Bound radioactivity was quantified byliquid scintillation counting. Membrane protein concentrations were determinedusing an assay kit from Bio-Rad (Hercules, CA) with bovine serum albumin as thestandard.
2.3. Electrophysiological experiments
cDNAs of rat GABAA receptor subunits were used for generating the respectivemRNA’s that were then injected into Xenopus laevis oocytes (Nasco, WI) asdescribed previously (Savi�c et al., 2008). For electrophysiological recordings, oocyteswere placed on a nylon-grid in a bath of Xenopus Ringer solution (XR, containing90 mM NaCl, 5 mM HEPES-NaOH (pH 7.4), 1 mM MgCl2, 1 mM KCl and 1 mM CaCl2).The oocytes were constantly washed by a flow of 6 ml/min XR which could beswitched to XR containing GABA and/or drugs. Drugs were diluted into XR fromDMSO-solutions resulting in a final concentration of 0.1% DMSO perfusing theoocytes. Drugs were pre-applied for 30 s before the addition of GABA, which was co-applied with the drugs until a peak response was observed. Between two applica-tions, oocytes were washed in XR for up to 15 min to ensure full recovery fromdesensitization. For current measurements the oocytes were impaled with twomicroelectrodes (2e3 mU) which were filled with 2 mM KCl. All recordings wereperformed at room temperature at a holding potential of �60 mV using a WarnerOC-725C two-electrode voltage clamp (Warner Instruments, Hamden, CT). Datawere digitised, recorded and measured using a Digidata 1322A data acquisitionsystem (Axon Instruments, Union City, CA).
2.4. Animals
Behavioral subjects were individually housed adult rhesus monkeys (Macacamulatta) maintained at 90e95% of their free-feeding weights. Aside from experi-mental procedures, monkeys were maintained on a 12-h lights-on/12-h lights-offcycle (lights on at 7:00 AM) and water was available continuously. All testingprocedures were conducted prior to 12 noon. Monkeys were prepared with chronicindwelling venous catheters according to the procedures described by Platt et al.(2005). Throughout all testing the National Institutes of Health guide for the careand use of Laboratory animals (NIH Publications No. 8023, revised 1978) wasadhered to, and the experimental procedures were approved by the HarvardMedical School Institutional Animal Care and Use Committee (Standing Committeeon Animals). All efforts were made to minimize animal suffering, to reduce thenumber of animals used, and to utilize alternatives to in vivo techniques, if available.
2.5. Conflict procedure
Fourmale and two female rhesusmonkeys were trained on amultiple schedule ofreinforcement as described in detail by Rowlett et al. (2006). Monkeys had variousdurations of exposure to this procedure, and these times ranged from 1e8 years. Adaily session consisted of 4 cycles, each preceded by a 10 min time out period inwhichall lights in the chamber were off and responding had no programmed consequences.Each cycle consisted of two components. The first component was signaled by redstimulus lights and consisted of a fixed ratio 18 (FR18) schedule of food pellet delivery(Bioserve, Frenchtown, NJ) followed by a 10 s time out. The second component,signaled by green stimulus lights, consisted of the FR 18 schedule of food deliverycombinedwith a FR 20 schedule of foot shock delivery (1.5e3.0 mA, adjusted for eachmonkey based on individual performance, 0.25 s duration). Delivery of a food pelletwas followed by a 10 s time out inwhich responding had no scheduled consequences.Both components were 5 min in duration, or ended after the monkey obtained 5 foodpellets or received 3 foot shocks, whichever occurred first.
Test sessions were conducted once or twice per week when monkeys reachedstable performance, defined as the average rates of responding for component 1 andcomponent 2 not varying by�20% over five consecutive sessions, with no upward ordownward trends. During test sessions, i.v. injections of vehicle or drug wereadministered in the 5th minute of each time out (i.e., 5 min prior to the beginning ofeach cycle). In successive cycles, increasing doses of the test drug were administeredusing a cumulative dosing procedure. The dependent measurewas the average ratesof responding (responses/s), calculated by dividing responses by time duringcomponents 1 and 2, excluding responding during time outs or reinforcer delivery.
2.6. Data analysis
Effects of doses of compounds were evaluated by conducting a priori Bonferronit-tests, comparing individual doses to vehicle injection with an alpha level set atp� 0.05. Potency values (dose engendering 50% maximum effect, ED50) were
calculated in individual monkeys by log-linear regression when at least three datapoints were available on the linear portion of the dose-effect curve or by interpo-lation when only two data points (one above and one below 50%) were available.Individual ED50 values were converted to their log values for calculation of meansand SEM and then converted back to linear values for presentation.
3. Results
3.1. In vitro profiles
Table 1 shows the in vitro binding affinity of diazepam, JY-XHe-053, XHe-II-053 and HZ-166 and the 4-methyl-JY-XHe-053stereoisomers SH-053-20F-S-CH3 and SH-053-20F-R-CH3 at GABAAreceptor subtypes. The six compounds produced a relatively broadrange of affinities across the receptor subtypes, with no compounddemonstrating substantial selectivity for a particular receptorsubtype. JY-XHe-053 and SH-053-20F-R-CH3 were the most selec-tive compounds across GABAA receptor subtypes, displaying 18-and 8- fold selectivity for a5GABAA receptors, respectively.
Table 2 shows the in vitro efficacy data at presumed physiologi-cally relevant concentrations of 100 nM and 1 mM for diazepam,JY-XHe-053, XHe-II-053, HZ-166, SH-053-20F-S-CH3 and SH-053-20F-R-CH3 as the percentage of control current. Relative to diazepam, theefficacy values were reduced at a1GABAA across all five compounds,with JY-XHe-053 exerting the greatest positive modulation and SH-053-20F-R-CH3 exerting the least positive modulation. The efficacyvalues for each compound were also reduced at a2GABAA anda3GABAA receptors relative to diazepam, but were larger than theirrespective values ata1GABAA receptors. JY-XHe-053 and SH-053-20F-R-CH3 exerted the greatest and least positivemodulation at a2GABAAand a3GABAA receptors, respectively. Relative to diazepam, the effi-cacy values for JY-XHe-053, XHe-II-053 and HZ-166 were reduced ata5GABAA receptors, while the efficacy values for SH-053-20F-S-CH3and SH-053-20F-R-CH3 were either decreased or increased, depend-ing on the concentration considered. HZ-166 exerted the least posi-tive modulation while SH-053-20F-S-CH3 exerted the greatestpositive modulation at a5GABAA receptors.
3.2. Anti-conflict effects of 8-acetylene imidazobenzodiazepines
Fig. 2 shows the effects of diazepam, JY-XHe-053, XHe-II-053,HZ-166 on the fixed-ratio schedule of food pellet delivery
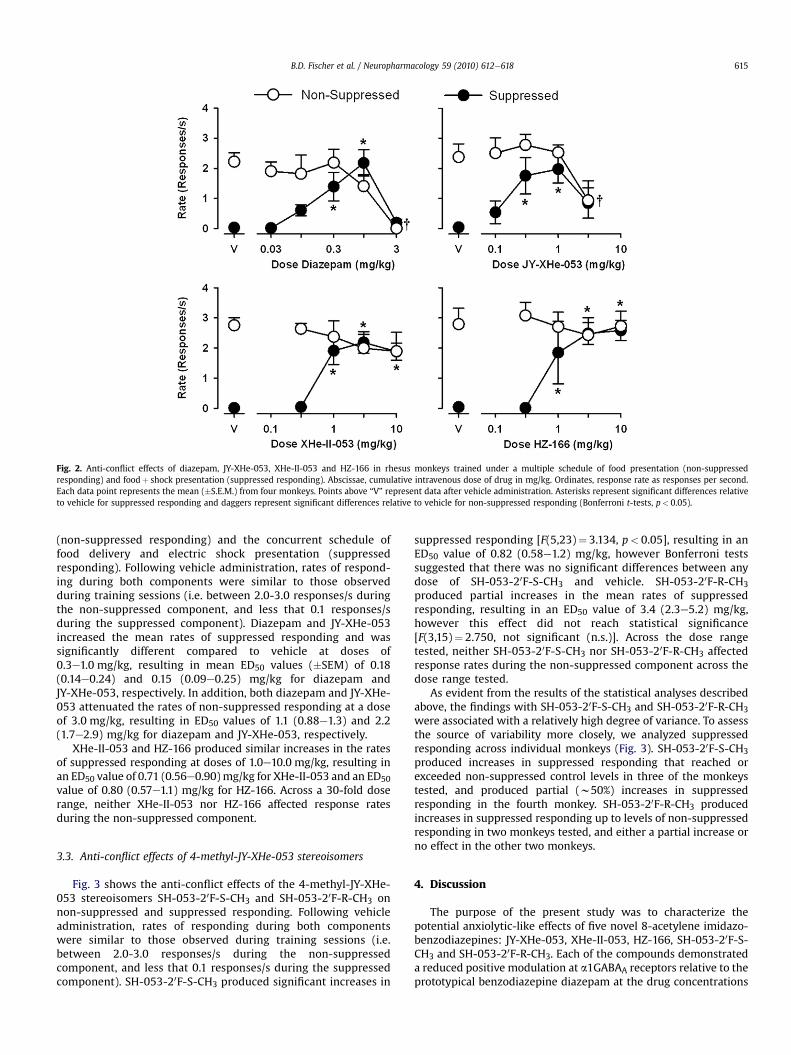
Fig. 2. Anti-conflict effects of diazepam, JY-XHe-053, XHe-II-053 and HZ-166 in rhesus monkeys trained under a multiple schedule of food presentation (non-suppressedresponding) and foodþ shock presentation (suppressed responding). Abscissae, cumulative intravenous dose of drug in mg/kg. Ordinates, response rate as responses per second.Each data point represents the mean (�S.E.M.) from four monkeys. Points above “V” represent data after vehicle administration. Asterisks represent significant differences relativeto vehicle for suppressed responding and daggers represent significant differences relative to vehicle for non-suppressed responding (Bonferroni t-tests, p< 0.05).
B.D. Fischer et al. / Neuropharmacology 59 (2010) 612e618 615
(non-suppressed responding) and the concurrent schedule offood delivery and electric shock presentation (suppressedresponding). Following vehicle administration, rates of respond-ing during both components were similar to those observedduring training sessions (i.e. between 2.0-3.0 responses/s duringthe non-suppressed component, and less that 0.1 responses/sduring the suppressed component). Diazepam and JY-XHe-053increased the mean rates of suppressed responding and wassignificantly different compared to vehicle at doses of0.3e1.0 mg/kg, resulting in mean ED50 values (�SEM) of 0.18(0.14e0.24) and 0.15 (0.09e0.25) mg/kg for diazepam andJY-XHe-053, respectively. In addition, both diazepam and JY-XHe-053 attenuated the rates of non-suppressed responding at a doseof 3.0 mg/kg, resulting in ED50 values of 1.1 (0.88e1.3) and 2.2(1.7e2.9) mg/kg for diazepam and JY-XHe-053, respectively.
XHe-II-053 and HZ-166 produced similar increases in the ratesof suppressed responding at doses of 1.0e10.0 mg/kg, resulting inan ED50 value of 0.71 (0.56e0.90) mg/kg for XHe-II-053 and an ED50value of 0.80 (0.57e1.1) mg/kg for HZ-166. Across a 30-fold doserange, neither XHe-II-053 nor HZ-166 affected response ratesduring the non-suppressed component.
3.3. Anti-conflict effects of 4-methyl-JY-XHe-053 stereoisomers
Fig. 3 shows the anti-conflict effects of the 4-methyl-JY-XHe-053 stereoisomers SH-053-20F-S-CH3 and SH-053-20F-R-CH3 onnon-suppressed and suppressed responding. Following vehicleadministration, rates of responding during both componentswere similar to those observed during training sessions (i.e.between 2.0-3.0 responses/s during the non-suppressedcomponent, and less that 0.1 responses/s during the suppressedcomponent). SH-053-20F-S-CH3 produced significant increases in
suppressed responding [F(5,23)¼ 3.134, p< 0.05], resulting in anED50 value of 0.82 (0.58e1.2) mg/kg, however Bonferroni testssuggested that there was no significant differences between anydose of SH-053-20F-S-CH3 and vehicle. SH-053-20F-R-CH3produced partial increases in the mean rates of suppressedresponding, resulting in an ED50 value of 3.4 (2.3e5.2) mg/kg,however this effect did not reach statistical significance[F(3,15)¼ 2.750, not significant (n.s.)]. Across the dose rangetested, neither SH-053-20F-S-CH3 nor SH-053-20F-R-CH3 affectedresponse rates during the non-suppressed component across thedose range tested.
As evident from the results of the statistical analyses describedabove, the findings with SH-053-20F-S-CH3 and SH-053-20F-R-CH3were associated with a relatively high degree of variance. To assessthe source of variability more closely, we analyzed suppressedresponding across individual monkeys (Fig. 3). SH-053-20F-S-CH3produced increases in suppressed responding that reached orexceeded non-suppressed control levels in three of the monkeystested, and produced partial (w50%) increases in suppressedresponding in the fourth monkey. SH-053-20F-R-CH3 producedincreases in suppressed responding up to levels of non-suppressedresponding in two monkeys tested, and either a partial increase orno effect in the other two monkeys.
4. Discussion
The purpose of the present study was to characterize thepotential anxiolytic-like effects of five novel 8-acetylene imidazo-benzodiazepines: JY-XHe-053, XHe-II-053, HZ-166, SH-053-20F-S-CH3 and SH-053-20F-R-CH3. Each of the compounds demonstrateda reduced positive modulation at a1GABAA receptors relative to theprototypical benzodiazepine diazepam at the drug concentrations

Fig. 3. Anti-conflict effects of SH-053-20F-S-CH3 and SH-053-20F-R-CH3 in rhesus monkeys trained under a multiple schedule of food presentation (non-suppressed responding) andfoodþ shock presentation (suppressed responding). Abscissae, cumulative intravenous dose of drug in mg/kg. Ordinates, response rate as responses per second. Top panels, eachdata point represents the mean (�S.E.M.) from four monkeys. Points above “V” represent data after vehicle administration. Bottom panels, data points represent the rate of sup-pressed responding in individual monkeys. Dashed lines represent rates of non-suppressed responding after vehicle administration.
B.D. Fischer et al. / Neuropharmacology 59 (2010) 612e618616
that can be reached under our experimental conditions. Potentia-tion of GABA at a2GABAA and a3GABAA receptors was also lowerrelative to diazepam; however the efficacy value at these receptorsubtypes was greater than the a1GABAA efficacy value across eachof the compounds. Additionally, each compound produced varyingdegrees of potentiation at a5GABAA receptors, with SH-053-20F-S-CH3 and SH-053-20F-R-CH3, the active isomers of 4-methyl-JY-XHe-053, exceeding those produced by diazepam.
Behavioral procedures that assess the effects of drugs on experi-mentally-induced conflict are used often to assess the potentialanxiolytic effects of these drugs in humans (Geller and Seifter, 1960;Spealman, 1979; Kleven and Koek, 1999; Rowlett et al., 2006). Theanti-conflict effects of a series of conventional benzodiazepines withnonselective efficacy across a1GABAA, a2GABAA, a3GABAA, anda5GABAA receptors subtypes has been described (Rowlett et al.,2006). In the present study, diazepam produced behavioral effectsthat were consistent with those of previously reported findings: itengendered a robust anti-conflict effect at low to intermediate dosesthat was both graded and dose-dependent. Further, this effectoccurred at doses similar to those that would be predicted based onrelative potencies for benzodiazepines that are effective in the clinic(Rowlett et al., 2006). Together, these observations provide furthersupport for the use of the rhesus monkey conflict procedure in pre-dicting the anxiolytic effects of drugs in humans.
The main finding from these experiments is that, similar todiazepam, thenovel compounds JY-XHe-053,XHe-II-053andHZ-166produced increases in positively reinforced behavior that was sup-pressed by response-contingent electric shock. In contrast, SH-053-20F-S-CH3 and SH-053-20F-R-CH3 produced increases in suppressedresponding in some monkeys and not others, with the average datafor SH-053-20F-R-CH3not reaching statistical significance.Among thenovel compounds studied, only JY-XHe-053 produced diazepam-likereductions in rates of non-suppressed responding. These findingssuggest that the anxiolytic and rate-reducing effects of benzodiaze-pines and other GABAA receptor positive modulators are dependenton their relative efficacy and affinity at GABAA receptor subtypes.
Previous studies from our laboratory also have used the rhesusmonkey conflict procedure to assess the anxiolytic effects of otherGABAA receptor positivemodulators with either selective affinity orselective efficacy for GABAA receptor subtypes (Licata et al., 2005;Rowlett et al., 2005, 2006). Results from these studies provideevidence for a differential role of GABAA receptors in the anxiolyticeffects of benzodiazepines. As an example, L-838,417, a drug withfunctional selectivity and partial agonist activity at a2GABAA,a3GABAA, and a5GABAA receptors, produced an anti-conflict effectsimilar to conventional non-selective benzodiazepines (Rowlettet al., 2005). A similar result was observed when SL651498,a drug with high intrinsic efficacy at a2GABAA and a3GABAA

B.D. Fischer et al. / Neuropharmacology 59 (2010) 612e618 617
receptors, was assessed in the conflict procedure (Licata et al.,2005). Together with data suggesting that drugs selective fora1GABAA receptors (e.g. zolpidem, zaleplon) are only marginallyeffective in this procedure (Rowlett et al., 2005, 2006), theseexperiments have supported a key role for a2GABAA and a3GABAAreceptors, but not a1GABAA receptors, in benzodiazepine-inducedanxiolysis. Subsequent studies with TPA023 (a drug with partialagonist properties at a2GABAA and a3GABAA receptors) in otherrodent and primate models of anxiety have supported thishypothesis (Atack et al., 2006).
In the present study, the in vitro electrophysiology experimentssuggest that XHe-II-053 and HZ-166 have high intrinsic efficacy ata2GABAA and a3GABAA receptor subtypes relative to their efficacyat a1GABAA and a5GABAA receptor subtypes.When XHe-II-053 andHZ-166 were assessed in the conflict procedure, each produced ananti-conflict effect that was quantitatively similar to diazepam.Together, these data provide further evidence that compoundswithselective efficacy at a2GABAA and a3GABAA receptors can produceanxiolytic effects in primates. Over the dose range tested, XHe-II-053 and HZ-166 produced an anti-conflict effect without producingdiazepam-like alterations in non-suppressed responding. The lackof response rate-suppressing effects of HZ-166 and XHe-II-053 issimilar to our previous results with L-838,417 and SL651498(Rowlett et al., 2005; Licata et al., 2005), and it is noteworthy thateach compound (XHe-II-053, HZ-166, L-838,417 and SL651498) hasreduced efficacy at a1GABAA receptors relative to diazepam. Thesefindings support the idea that the a1GABAA receptor subtype maybe involved in the response rate-reducing effects of benzodiaze-pines at doses greater than those that produce anxiolysis.
Similar to the pharmacological profile of XHe-II-053 and HZ-166, the electrophysiological experiments suggest that JY-XHe-053has preferential activity at a2GABAA and a3GABAA receptorsubtypes. In agreement with the hypothesis that a2GABAA and/ora3GABAA receptors mediate the anxiolytic effects of benzodiaze-pines, JY-XHe-053 also produced a robust anti-conflict effect.Unlike XHe-II-053 and HZ-166 however, JY-XHe-053 also producedsignificant reductions in response rates at the highest dose tested.This finding was unexpected, considering the relatively lowintrinsic efficacy of JY-XHe-053 at a1GABAA receptors. However, itis interesting to note that, relative to XHe-II-053 and HZ-166, JY-XHe-053 has both greater efficacy at and greater affinity fora1GABAA receptors. These observations support the hypothesis thata2GABAA and a3GABAA receptors mediate the anxiolytic effects ofbenzodiazepines, and also raise the possibility that the subtledifferences at a1GABAA receptors between JY-XHe-053 and theother compounds may be sufficient for rate-reducing effects.
In contrast to the diazepam-like anti-conflict and rate-reducingeffects observed with JY-XHe-053, the two isomers of 4-methyl-JY-XHe-053, SH-053-20F-S-CH3 and SH-053-20F-R-CH3 producedrelatively weak increases in suppressed responding and failed toproduce significant decreases in non-suppressed responding. Thereasons for these differences are unclear. Based on the in vitro data,the primary differences between the two isomers and the parentcompound were that (1) the affinities of the isomers were reducedconsiderably relative to JY-XHe-053, (2) the efficacies of theisomers at the a5GABAA receptor were increased relative to that ofthe parent compound and (3) the efficacy of SH-053-20F-R-CH3 ata2GABAA and a3GABAA receptors at the achievable drug concen-trations was markedly less than that of the parent compound. Therelatively low potencies of the isomers may have necessitated veryhigh levels of compound for consistent effects, raising the possi-bility that high enough concentrations were not sufficient in brainto for consistent effects to occur. Additionally, it is interesting tonote that among the compounds tested, both SH-053-20F-S-CH3and SH-053-20F-R-CH3 have substantial efficacy and high affinity
for a5GABAA receptors. In fact, in contrast to the other compoundsand receptor subtypes, SH-053-20F-S-CH3 and SH-053-20F-R-CH3both exceed diazepam in efficacy at this subtype when assessed atthe physiologically relevant concentration of 1 mM. Therefore, theunexpected finding that SH-053-20F-R-CH3 lacked significant anti-conflict effects may result from its pharmacological action ata5GABAA receptors coupled with its low efficacy at a2GABAA anda3GABAA receptors. Further, the statistically significant but weakanti-conflict effects of SH-053-20F-S-CH3 may also be a conse-quence of its unique pharmacological profile at a2GABAA, a3GABAA,and a5GABAA receptors. Support for these hypotheses comes froma recent report implicating a5GABAA receptors in benzodiazepine-induced psychomotor effects (Savi�c et al., 2008), which the authorssuggest may serve to mask anxiolysis as measured in preclinicalprocedures.
The experiments described here provide further evidence thatthe anti-conflict effects of benzodiazepines in non-human primatesare likely mediated by different GABAA receptors that containdistinct a subunits. Additionally, the results from this study suggestthat it is possible to separate anxiolytic-like effects from effectsindicative of a general disruption of behavior. Our findings providefurther evidence that novel benzodiazepine-like drugs that havepharmacological selectivity for a2GABAA and/or a3GABAA recep-tors and low receptor activity at a1GABAA and a5GABAA receptorsmay be particularly useful as non-sedating anxiolytics. Also, theresults from our studies suggest that subtle differences in a1GABAAreceptor activation may be sufficient for the rate-reducing effects ofbenzodiazepine-like drugs. Finally, our findings raise the possibilitythat exceptional activity at a5GABAA receptors may blunt theanxiolytic-like effects of benzodiazepines, regardless of theirpharmacology at other receptor subtypes. Together, these obser-vations should provide an important framework for studying therole of different GABAA receptor subtypes in the behavioral effectsof benzodiazepine-type drugs, which in turn should help guideboth the current clinical use of benzodiazepines as well as thedevelopment of improved therapeutic agents for treating anxietydisorders.
Acknowledgments
This work was supported by USPHS grants DA11792, RR00168,and MH046851. The authors acknowledge the technical assistanceof Kristen Bano and Annemarie Duggan.
References
Atack, J.R., Wafford, K.A., Tye, S.J., Cook, S.M., Sohal, B., Pike, A., Sur, C., Melillo, D.,Bristow, L., Bromidge, F., Ragan, I., Kerby, J., Street, L., Carling, R., Castro, J.L.,Whiting, P.,Dawson,G.R.,McKernan,R.M., 2006. TPA023 [7-(1,1-Dimethylethyl)-6-(2-ethyl-2H-1,2,4-triazol-30ylmethoxy)-3-(2-fluorophenyl)-1,2,4-triazolo[4,3-b]pyridazine], an agonist selective for a2 and a3-containing GABAA receptors, isa nonsedating anxiolytic in rodents and primates. J. Pharmacol. Exp. Ther. 316,410e422.
Choudhary, M.S., Craigo, S., Roth, B.L., 1992. Identification of receptor domains thatmodify ligand binding to 5-hydroxytryptamine2 and 5-hydroxytryptamine1cserotonin receptors. Mol. Pharmacol. 42, 627e633.
Collinson, N., Kuenzi, F.M., Jarolimek, W., Maubach, K.A., Cothliff, R., Sur, C.,Smith, A., Otu, F.M., Howell, O., Atack, J.R., McKernan, R.M., Seabrook, G.R.,Dawson, G.R., Whiting, P.J., Rosahl, T.W., 2002. Enhanced learning and memoryand altered GABAergic synaptic transmission in mice lacking the alpha 5subunit of the GABAA receptor. J. Neurosci. 22, 5572e5580.
Cook, L., Davidson, A.B., 1973. Effects of behaviorally active drugs in a conflict-punishment procedure in rats. In: Garattini, S., Mussini, E., Randall, L.O. (Eds.),The Benzodiazepines. Raven, New York, pp. 327e345.
Cook, J.M., Zhou, H., Huang, S., Srirama Sarma, P.V.V., Zhang, C., 2009. Stereospecificanxiolytic and anticonvulsant agents with reduced muscle-relaxant, sedative-hypnotic and ataxic effects, US Patent 7,618,958 B2, Issued November 17, 2009.
Cook, J.M., Edwankar, R., Edwankar, C., Huang, S., Jain, H., Rivas, F., Yang, J., Zhou, H.,2010. Selective anticonvulsant agents and their uses, T08025US Utility, Serial #12/725,763, Filed March 17, 2010.

B.D. Fischer et al. / Neuropharmacology 59 (2010) 612e618618
Crestani, F., Keist, R., Fritschy, J.M., Benke, D., Vogt, K., Prut, L., Blüthmann, H.,Möhler, H., Rudolph, U., 2002. Trace fear conditioning involves hippocampalalpha5 GABA(A) receptors. Proc. Natl. Acad. Sci. U.S.A. 99, 8980e8985.
Dixon, C.I., Rosahl, T.W., Stephens, D.N., 2008. Targeted deletion of the GABRA2 geneencoding alpha2-subunits of GABA(A) receptors facilitates performance ofa conditioned emotional response, and abolishes anxiolytic effects of benzo-diazepines and barbiturates. Pharmacol. Biochem. Behav. 90, 1e8.
Geller, I., Seifter, J., 1960. The effects of meprobamate, barbiturate, damphetamineand promazine on experimentally-induced conflict in the rat. Psychopharma-cologia 1, 482e492.
Griffiths, R.R., Weerts, E.M., 1997. Benzodiazepine self-administration in humansand laboratory animalseimplications for problems of long-term use and abuse.Psychopharmacology (Berl). 134, 1e37.
Kleven, M.S., Koek, W., 1999. Effects of different classes of partial benzodiazepineagonists on punished and unpunished responding in pigeons. Psychopharma-cology (Berl). 144, 405e410.
Licata, S.C., Platt, D.M., Cook, J.M., Sarma, P.V., Griebel, G., Rowlett, J.K., 2005. Contri-bution of GABAA receptor subtypes to the anxiolytic-like, motor, and discrimina-tive stimulus effects of benzodiazepines: studies with the functionally selectiveligand SL651498 [6-fluoro-9-methyl-2-phenyl-4-(pyrrolidin-1-yl-carbonyl)-2,9-dihydro-1H-pyridol[3,4-b]indol-1-one]. J. Pharmacol. Exp. Ther. 313, 1118e1125.
Löw, K., Crestani, F., Keist, R., Benke, D., Brünig, I., Benson, J.A., Fritschy, J.M.,Rülicke, T., Bluethmann, H., Möhler, H., Rudolph, U., 2000. Molecular andneuronal substrate for the selective attenuation of anxiety. Science 290,131e134.
McKernan, R.M., Whiting, P.J., 1996. Which GABAA-receptor subtypes really occur inthe brain? Trends Neurosci. 19, 139e143.
McKernan, R.M., Rosahl, T.W., Reynolds, D.S., Sur, C., Wafford, K.A., Atack, J.R.,Farrar, S., Myers, J., Cook, G., Ferris, P., Garrett, L., Bristow, L., Marshall, G.,Macaulay, A., Brown, N., Howell, O., Moore, K.W., Carling, R.W., Street, L.J.,Castro, J.L., Ragan, C.I., Dawson, G.R., Whiting, P.J., 2000. Sedative but notanxiolytic properties of benzodiazepines are mediated by the GABA(A) receptoralpha1 subtype. Nat. Neurosci. 3, 587e592.
Millan, M.J., 2003. The neurobiology and control of anxious states. Prog. Neurobiol.70, 83e244.
Nutt, D.J., 2005. Overview of diagnosis and drug treatments of anxiety disorders.CNS Spectr. 10, 49e56.
Olsen, R.W., Sieghart, W., 2008. International Union of Pharmacology. LXX. Subtypesof gamma-aminobutyric acid(A) receptors: classification on the basis of subunitcomposition, pharmacology, and function. Update. Pharmacol. Rev. 60, 243e260.
Platt, D.M., Carey, G.J., Spealman, R.D., 2005. Intravenous self-administration tech-niques in monkeys. In: Enna, S., Williams, M., Ferkany, J., Kenakin, T., Porsolt, R.,Sullivam, J. (Eds.), Current Protocols in Neuroscience. Wiley, New York Unit 9.21.
Pritchett, D.B., Lüddens, H., Seeburg, P.H., 1989. Type I and type II GABAA-benzo-diazepine receptors produced in transfected cells. Science 245, 1389e1392.
Rivas, F.M., Stables, J.P., Murphree, L., Edwankar, R.V., Edwankar, C.R., Huang, S.,Jain, H.D., Zhou, H., Majumder, S., Sankar, S., Roth, B.L., Ramerstorfer, J.,Furtmüller, R., Sieghart, W., Cook, J.M., 2009. Antiseizure activity of novelgamma-aminobutyric acid (A) receptor subtype-selective benzodiazepineanalogues in mice and rat models. J. Med. Chem. 52, 1795e1798.
Rowlett, J.K., Platt, D.M., Lelas, S., Atack, J.R., Dawson, G.R., 2005. Different GABAAreceptor subtypes mediate the anxiolytic, abuse-related, and motor effects ofbenzodiazepine-like drugs in primates. Proc. Natl. Acad. Sci. U.S.A. 102, 915e920.
Rowlett, J.K., Lelas, S., Tornatzky, W., Licata, S.C., 2006. Anti-conflict effects ofbenzodiazepines in rhesus monkeys: relationship with therapeutic doses inhumans and role of GABAA receptors. Psychopharmacology (Berl.) 184, 201e211.
Rudolph, U., Crestani, F., Benke, D., Brünig, I., Benson, J.A., Fritschy, J.M., Martin, J.R.,Bluethmann, H., Möhler, H., 1999. Benzodiazepine actions mediated by specificgamma-aminobutyric acid(A) receptor subtypes. Nature 401, 796e800.
Rudolph, U., Crestani, F., Möhler, H., 2001. GABA(A) receptor subtypes: dissectingtheir pharmacological functions. Trends Pharmacol. Sci. 22, 188e194.
Savi�c, M.M., Clayton, T., Furtmüller, R., Gavrilovi�c, I., Samardzi�c, J., Savi�c, S., Huck, S.,Sieghart, W., Cook, J.M., 2008. PWZ-029, a compound with moderate inverseagonist functional selectivity at GABA(A) receptors containing alpha5 subunits,improves passive, but not active, avoidance learning in rats. Brain Res. 1208,150e159.
Spealman, R.D., 1979. Comparison of drug effects on responding punished bypressurized air or electric shock delivery in squirrel monkeys: pentobarbital,chlordiazepoxide, d-amphetamine and cocaine. J. Pharmacol. Exp. Ther. 209,309e315.
Copyright © 2022 FDOKUMEN




















