
Grooming and consort partner selection in a troop of Japanese monkeys ( Macaca fuscata )

PRIMATES, 41 (1): 63-77, January 2000 63
The Development of Behavioural Sex Differences in Infant Rhesus Macaques (Macaca mulatta)
GILLIAN R. BROWN and ALAN E DIXSON University of Cambridge
ABSTRACT. Studies of infant rhesus macaques have generally reported sex differences in the frequency of expression of some behaviour patterns, such as rough-and-tumble play and socio-sexual mounting. In contrast, sex differences in other behaviour patterns, such as proximity to the mother, have been less con- sistantly reported. Using data on the behavioural development of infant rhesus macaques living in captive social groups, we have attempted to provide further evidence for, or against, sex differences in behaviour and to investigate the possible influence of maternal rank and parity on sex differences in infant behaviour and mother-infant interactions. The behaviour of 14 male and 20 female infants and their mothers was studied during the first six months of life, including measures of play behaviour, socio-sexual mounting, and mother-infant interactions. Our data reveal that, on average, male infants exhibited more rough-and- tumble play and mounting than female infants, and also exhibited stationary play, chasing play, and initi- ated play more frequently than females. Such sex differences appear to be robust in macaques and have been reported in a variety of housing conditions. Male and female infants did not differ in the amount of time spent at particular distances from their mothers, and mothers were not found to behave differently towards sons and daughters, using measures of restraint, rejection, and grooming. These results are in con- trast to previous studies on singly-housed mother-infant pairs but similar to those on free-ranging popula- tions. Mothers did behave differently towards their infants depending upon the mother's rank and previous number of offspring. These maternal characteristics may have significant consequences for the behavioural development of both male and female infant primates.
Key Words: Sex differences; Infant development; Rhesus macaques.
INTRODUCTION
Sex differences in primate behaviour are reported to emerge at a very early age. During the first year of life, male and female primates have been found to differ in the frequency with which they exhibit certain behaviour patterns. For example, male infants exhibit rough-and- tumble play more frequently than female infants in a variety of primate species living in free- ranging groups, including patas monkeys (Erythrocebus patas: ROWELL & CHISM, 1986), rhesus macaques (Macaca mulatta: SYMONS, 1978), Japanese macaques (Macacafuscata: NAKAMICHI, 1989), and olive baboons (Papio anubis: OWENS, 1975). Also, males less than ! yr of age have been reported to exhibit socio-sexual mounting more frequently than female infants in several species of macaques and baboons (e.g. ROSENBLUM & NADLER, 1971; OWENS, 1973; HANBY & BROWN, 1974; DIXSON, 1977).
Three main factors are involved in the development of sexually dimorphic behaviour pat- terns, such as rough-and-tumble play and mounting, in infant primates. Firstly, exposure of the developing brain to testicular hormones during foetal development masculinizes the neural sub- strates which govern the expression of sexually dimorphic patterns (e.g. in the rhesus monkey: Gou & MCEWEN, 1980). By contrast, the postnatal ' surge ' of testosterone, which occurs during the first few months of infancy in male rhesus monkeys and some other primates, has no effect

64 G.R. BROWN & A. E DIXSON
upon the development of sociosexual and playful behaviour patterns (WALLEN et al., 1995; NEVlSON et al., 1997; BROWN & DIXSON, 1999).
Secondly, the physical and social environment may affect the expression of sexually dimor- phic behaviour patterns during infancy and the development of the mother-infant relationship in captivity. Studies on singly-housed, captive pairs of macaques have reported male infants to be more active and restrained by their mothers less frequently than female infants, while female infants were rejected less and groomed more frequently than males (JENSEN et al., 1968; MITCHELL & BRANDT, 1970; MITCHELL, 1979; NAKAMICHI et al., 1990). In contrast, observations of free-ranging Japanese macaques and baboons have not replicated such findings (ALTMANN, 1980; NICOLSON, 1987; TANAKA, 1989). The housing of mother-infant pairs in single cages greatly restricts their physical environment and may not provide an accurate view of infant behavioural development. The presence of other group members has been shown to influence behaviour in young captive rhesus macaques. Male infant rhesus macaques raised in mixed-sex peer groups exhibit higher levels of rough-and-tumble play and mounting behaviour than male infants reared in social groups with only male peers (GOLDFOOT & WALLEN, 1978). However, sex differences in play and mounting behaviour appear to be relatively robust, at least in rhesus macaques, as they have been reported in a variety of different physical and social environments (reviewed by WALLEN, 1996).
Thirdly, characteristics of the mother, the presence of kin, and parameters of the local envi- ronment, such as foraging demands and predation risks, may also play a role (e.g. BERMAN et al., 1997; DE JONGE et al., 1981; FAmBANKS, 1988; HAUSER & FAIRBANKS, 1988; HOOLEY & SIMPSON, 1983; LEE, 1984a, b; SCHINO et al., 1995; TANAKA, 1989). In primates, the social rank and parity of the mother have traditionally been the factors most frequently investigated in stud- ies of infant behavioural development (BERMAN, 1984). Yet, few studies have investigated how the sex of the infant and the rank and parity of the mother combine to influence infant behav- ioural development. Are sex differences in infant behaviour perhaps more pronounced between sons and daughters of low-ranking mothers? In terms of maternal behaviour, do mothers of dif- fering rank or parity behave differently towards sons and daughters? How the development of sex differences in infant behaviour may be related to the mother's rank and parity, and how dif- ferent mothers may behave differently towards sons and daughters, deserves further investiga- tion.
The aim of this project was to investigate the occurrence of sex differences in infant behav- iour and to investigate the possible influence of maternal rank and parity on the expression of sex differences in behaviour and on the mother-infant relationship. The behaviour of captive infant rhesus macaques and their mothers living in small, matrilinially-based social groups of stable composition was studied during the first six months of infant life. Data on play, sociosex- ual behaviour and mother-infant interactions are reported.
METHODS
SUBJECTS AND HOUSING
This study was carried out on 34 infant rhesus macaques (20 females and 14 males) born dur- ing 1994 and 1995 at the Sub-Department of Animal Behaviour, Madingley (University of Cambridge, U.K.). The housing and maintenance of this well-established colony have been pre- viously described in detail (SIMPSON et al., 1989). The macaques were housed in six social groups, each usually consisting of one adult male and three to five adult females with their off-

Sex Differences in Infant Behaviour
Table 1. Numbers of mothers within each category of rank and parity.
65
Rank Parity
14 mothers with sons 8 high or medium 2 primiparous 6 multiparous
6 low I primiparous 5 multiparous
20 mothers with daughters 11 high or medium 3 primiparous 8 multiparous
9 low 2 primiparous 7 multiparous
spring. Group compositions were kept as constant as possible during the 2-year period of the study and have been described elsewhere (NEvlSON et al., 1997). Each group had an outdoor enclosure and constant access to a heated indoor room. The monkeys were fed with monkey chow between 08:30 and 09:30 and with fruit between 12:30 and 13:00. Water was continu- ously available.
These animals have been previously used in a study of the role of testosterone in behavioural development (NEvlSON et al., 1997; BROWN & DIXSON, 1999). In summary, half of the male infants were treated with a GnRH agonist, and half of the female infants with testosterone, dur- ing the first six months of life. The remaining infants were not hormonally-manipulated. Due to the lack of effects of postnatal testosterone manipulation on the development of behaviour, we feel confident in reporting on the behavioural development of all these animals. Careful com- parison of the behaviour of hormonally-manipulated and control infants did nor reveal any sig- nificant alterations in the behavioural development of male or female infants or changes in their interactions with their mothers (BRowN & DIXSON, 1999).
In this macaque colony, adult females are regularly categorized as either low-, medium-, or high-ranking (depending upon whether they fall into the top, middle, or bottom third of all females within their group when placed in linear rank order), based upon observations of aggressive and submissive interactions during food provisioning. Only a small number of high- ranking mothers gave birth during the study period, as several had received surgery to prevent conception (hysterectomies) following problems with previous offspring. Thus, for all analyses, data for infants of high-ranking mothers have been combined with the data for infants of medium-ranking mothers. Preliminary investigation of the data did not reveal any significant differences in any behavioural measures between infants of high-ranking and medium-ranking mothers. Thus, observational data were available from 19 high- or medium-ranking mothers to be compared with data from 15 low-ranking mothers (Table 1). Females that had not previously been pregnant, or had had no previous offspring survive beyond a week after birth, were described as primiparous while those that had had a previous surviving offspring were described as multiparous. Data from those mothers that gave birth during both years (N=9) were treated independently.
COLLECTION OF BEHAVIOURAL DATA
Behavioural data were collected on the 34 infants in their social groups during the whole of their first six months of life, using both continuous recording and instantaneous sampling meth- ods (MARTIN & BATESON, 1986). Four focal watches, each lasting half an hour, were carried out on each infant each week. A total of 1768 hr of behavioural data were collected throughout the study, comprised of 52 hr of data collection per infant. Focal watches were carried out between

66 G.R. BROWN & A. E "DIxsoN
09:30 and 12:30 and between 13:00 and 17:30, to control for diurnal variations in behaviour and to avoid the disruptive effects of feeding periods. Focal watches were divided equally between morning sessions and afternoon sessions. Due to the large amount of data collected, several observers were required. The reliability of data collection between observers was regularly checked and showed high correlation (values of COHEN'S kappa ranging from 0.70 to 0.80; BAKEMAN & GOTrMAN, 1986). The behavioural definitions are based on those of previous stud- ies (H1NDE et al., 1964; HINDE & SPENCER-BOOTH, 1967).
Play behaviour: All occurrences of play behaviour during the focal watch were recorded as single events by continuous observation. A bout was defined as having ended if a 5-sec pause in the activity was noted. Three categories of social play behaviour involving the focal infant and another individual were defined: (1) stationary play: gentle movements by both individuals in a stationary position; (2) chase play: non-aggressive, vigorous chasing; and (3) rough-and-tumble play: non-aggressive, energetic movements such as wrestling, grabbing, and holding down. The number of play initiations by the focal infant, and the number of play initiations received by the infant, were also recorded. A play initiation was defined as an individual pulling an open- mouthed play-face, giving a gentle slap or grapple, or running up to, touching, and then running away from another individual. The total amount of play behaviour was obtained by adding together the occurrences of stationary play, chasing play, and rough-and-tumble play events observed during the focal watch.
Socio-sexnal behaviour: The total number of mounts (orientation of the pelvis towards the hindquarters of another animal) and presents (raising of the tail and orientation of hindquarters towards another animal) exhibited by focal infants were recorded using continuous observation. The occurrence of pelvic thrusting movements and of the double-foot clasp mounting posture was also noted.
Mother-infant proximity: The distance between the focal infant and its mother was recorded continuously during each focal watch. Four distances were defined: (1) infant and mother in ventro-ventral contact; (2) infant and mother in any other body contact, other than ventro-ven- tral contact; (3) infant and mother within arm's reach (<60cm) of each other; and (4) infant and mother at greater than arm 's reach from each other. The total amount of time spent in body con- tact with the mother was calculated by adding together the amount of time spent in ventro-ven- tral contact and in any other form of body contact. At 60-sec intervals, the infant was recorded as either being alone or having at least one other group member within arm's reach.
Mother-infant interactions: The total number of times that the mother restrained her infant, by holding on to it as it attempted to move away from her, and lifted her infant into ventro-ventral contact, usually following a disturbance, were recorded using continuous observation. The total number of times that mothers groomed their infants, and the total duration of such grooming bouts, were also recorded.
STATISTICAL ANALYSES
For those behaviour patterns recorded as single events, the data were used to calculate mean frequencies per hour for each of the 6 monthly age periods separately. For those behaviours recorded at 60-sec intervals, the proportion of scans spent in each behavioural state were calcu- lated for each month. For the data on mother-infant proximity, the proportion of time spent by

Sex Differences in Infant Behaviour 67
the infant at each of the distances from the mother was calculated for each month. The main sta- tistical test used was a repeated measures analysis of variance (ANOVA), using infant age in months as the repeated measure (ZAR, 1974). Any interactions revealed by these analyses of variance were further investigated using simple effect tests. In order to control the familywise
error rate, where simple effects were carried out, we divided alpha by the number of compar- isons. Those data sets which did not conform to a normal distribution, even following a square
root or arcsin transformation (MARTIn & BATESON, 1986; ZAR, 1974), were analysed using the nonparametric Mann-Whitney U-test (SIEGEL & CASTELLAN, 1988).
RESULTS
For all of the ANOVAs that revealed significant effects or interactions, the results are shown
in Table 2 and described in more detail below.
PLAY BEHAVIOUR
Male infants engaged in significantly more stationary play than female infants (Fig. la). An interaction was found between the sex of the infant and the frequency of play at different ages, which resulted from male infants exhibiting significantly higher frequencies of stationary play than female infants during months 3 and 4 [simple effects: F(1,130)=24.3 and 9.4 respectively, p<0 .006 for both]. Also, infants of low-ranking mothers exhibited higher average frequencies of stationary play over the six month period (3.4+0.2 bouts per hour, me a n+ S EM) than infants of high-/medium-ranking mothers (2.9+0.2 bouts per hour). Infants of primiparous and multi- parous mothers did not differ in their total amounts of stationary play, but an interaction was
Table 2, ANOVA table for all significant effects and interactions.
Effect or interaction Description F value (d:f) Stationary play Sex M >F F ( 1,26) = 14.4*
Sex• M>F (months 3,4) F (5,130)=4.2" Rank x Age L>H/M F (1,26) =6.5' Parityx Age MP>PP (month 4) F (1,130)=2.5"
Chasing play Sex M>F F (1,26) =6.5* Sex• Age M>F (month 4) F (5,130)=2.5" Rank L>H/M F (1,26)=4.4"
Rough-and-tumble play Sex M>F F (1,26) = 12.0** SexxAge M>F (months 3-6) F (5,130)=5.2"*
Total play Sex M>F F ( 1,26)= 18.9** Sex• M>F (months 3-6) F (5,130)=5.5"* Rank L>H/M F ( 1,26) =6.2"
Play initiation Sex x Age M>F (months 3,4,6) F (5,130)=2.8* Receives play initiation Sex M>F F (1,26)=5.2"
Sexx Age M>F (months 2,3) F (5,130)=2.5" RankxAge H/M>L (month 6) F ( I, 130)=3.2" Parity• MP>PP (month 4) F (5,130)=2.6"
Ventro-ventral contact Parity PP>MP F (1,26) = 8.7* Body contact Parity PP>MP F (I ,26)=3.8(*)
Sex x Rank F>M (low-ranking) F (I,26)=3.6(*) Out of arm's reach Parity MP>PP F ( 1,26)=5.0* Maternal rejection Rank x Age H/M>L (month 2) F (5,130) = 3.3* Maternal lift Parity PP>MP F ( 1,26)= 18.3**
M: Males; F: females; L: low-ranking; H/M: high-/medium-ranking; MP: multiparous; PP: primiparous. * p<0.05; �9 * p<0.01; (*) p<0.1.

Frequency (per hour)
Frequency (per hour)
a) Stationary play
t I I 1 2 "
1 0 "
8' 6 4 2 0 1 2 3 4 5 6
c) Rough-and-tumble play
! |
1 2 3 4 5 6
b) Chasing play d) Total play
I I 2 0 " t
~ 15'
10
5
0 1 2 3 4 5 6
1 2 3 4 5 6
Month Month
68 G.R. BROWN & A. E DIXSON
Fig. 1. Mean hourly frequency with which male infants (solid circles, N=I4) and female infants (open circles, N=20) exhibited: a) stationary play; b) chasing play; c) rough-and-tumble play; d) the total fre- quency of play during the first six months of life. Means and SEMs: * p<0.05 (ANOVA); t p<0.006 (simple effects).
found between parity and age. During the fourth month of life, infants of multiparous mothers exhibit stationary play (5.0+0.4 bouts per hour) more frequently than infants of primiparous mothers [2.7 +0.7 bouts per hour; simple effects: F(i, 130) = 13.4, p<0.006].
Male infants exhibited higher frequencies of chasing play than female infants (Fig. lb). An interaction between infant sex and age resulted from male infants exhibiting chasing play more frequently than female infants during month 4 [simple effects: F(I, 130)= 14.6, p<0.006]. Also, infants of low-ranking mothers exhibited higher average frequencies of chasing play over the six month period (2.4+0.3 bouts per hour) than infants of high-/medium-ranking mothers (2.1 +0.2 bouts per hour).
Male infants exhibited significantly higher frequencies of rough-and-tumble play than female infants (Fig. lc). An interaction between infant sex and age revealed that this sex difference was significant from month 3 onwards [simple effects: F(1,130)=16.5, 35.8, 29.0, and 40.0 for months 3, 4, 5, and 6 respectively, p<0.006 for all]. No interactions were found between the frequency of rough-and-tumble play and the rank or parity of the mother.
Male infants exhibited significantly higher total amounts of play than female infants (Fig. ld). An interaction between infant sex and the total frequency of play at different ages revealed that this difference was significant from month 3 onwards [simple effects: F(1,130)=32.1, 32.1, 21.6, and 18.7 for months 3, 4, 5, and 6 respectively, p<0.006 for all]. Infants of low-ranking mothers exhibited slightly higher average frequencies of play over the six month period (6.2+0.6 bouts per hour) than infants of medium-/high-ranking mothers (5.5_0.3 bouts per hour).
Sex Differences in Infant Behaviour 69
Frequency (per hour)
a) Focal infant initiates play
t
1 2 3 4 5 6
7
6
5
Frequency 4 (per hour) 3
2
1
0
b) Focal infant receives play
I t
1 2 3 4 5 6
Month
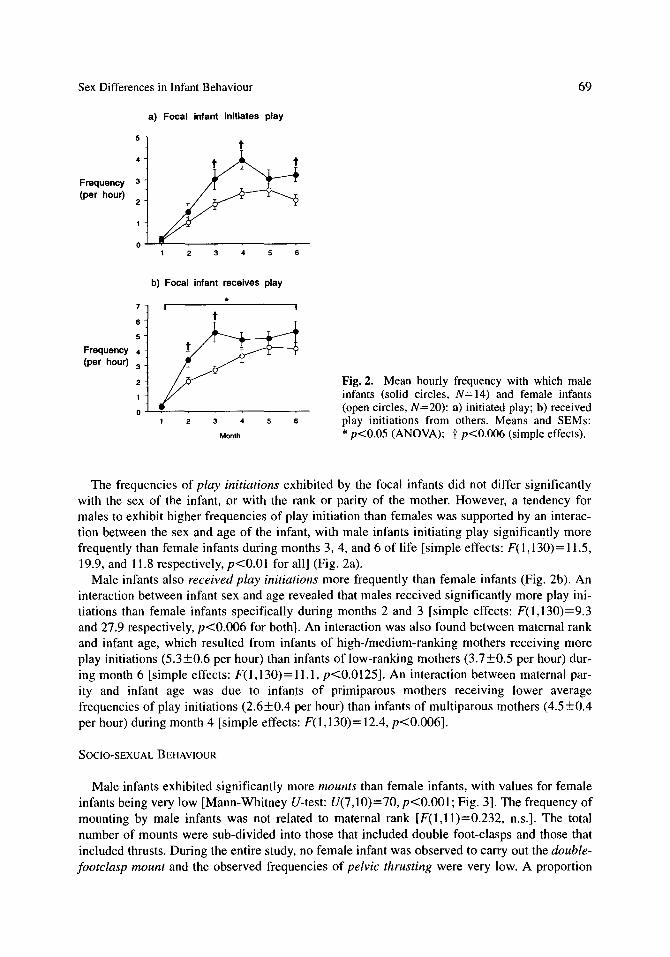
Fig. 2. Mean hourly frequency with which male infants (solid circles, N=I4) and female infants (open circles, N=20): a) initiated play; b) received play initiations from others. Means and SEMs: * p<0.05 (ANOVA); t p<0.006 (simple effects).
The frequencies of play initiations exhibited by the focal infants did not differ significantly with the sex of the infant, or with the rank or parity of the mother. However, a tendency for males to exhibit higher frequencies of play initiation than females was supported by an interac- tion between the sex and age of the infant, with male infants initiating play significantly more frequently than female infants during months 3, 4, and 6 of life [simple effects: F(1,130)= 11.5, 19.9, and 11.8 respectively, p<0.01 for all] (Fig. 2a).
Male infants also received play initiations more frequently than female infants (Fig. 2b). An interaction between infant sex and age revealed that males received significantly more play ini- tiations than female infants specifically during months 2 and 3 [simple effects: F(1,130)=9.3 and 27.9 respectively, p<0.006 for both]. An interaction was also found between maternal rank and infant age, which resulted from infants of high-/medium-ranking mothers receiving more play initiations (5.3+0.6 per hour) than infants of low-ranking mothers (3.7+0.5 per hour) dur- ing month 6 [simple effects: F(I, 130) = 11.1, p<0.0125]. An interaction between maternal par- ity and infant age was due to infants of primiparous mothers receiving lower average frequencies of play initiations (2.6+0.4 per hour) than infants of multiparous mothers (4.5+0.4 per hour) during month 4 [simple effects: F(I,130)= 12.4, p<0.006].
SOC10-SEXUAL BEHAVIOUR
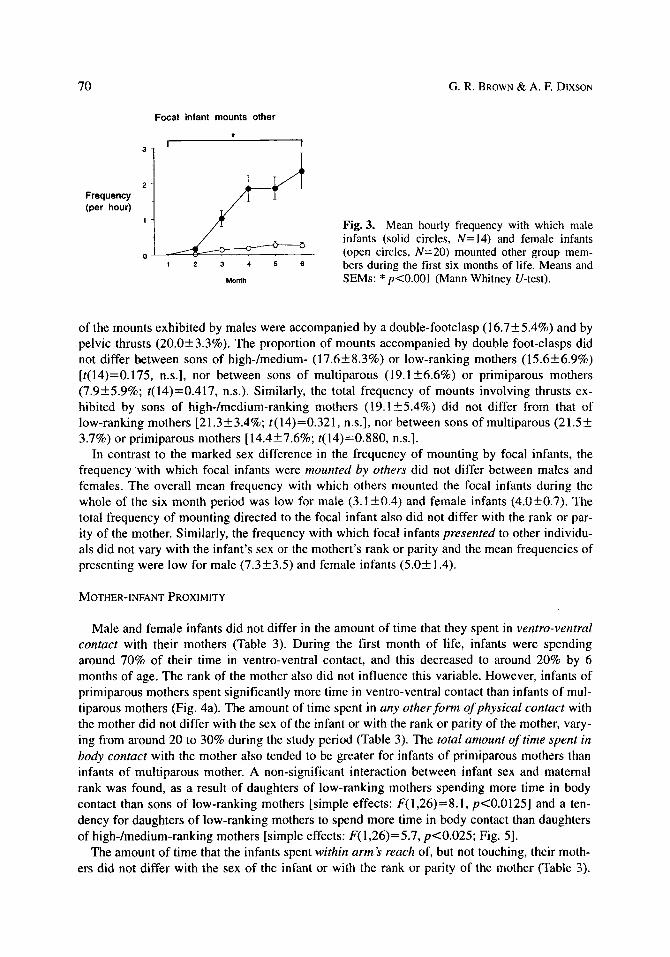
Male infants exhibited significantly more mounts than female infants, with values for female infants being very low [Mann-Whitney U-test: U(7,10)=70, p<0.001; Fig. 3]. The frequency of mounting by male infants was not related to maternal rank [F(1,11)=0.232, n.s.]. The total number of mounts were sub-divided into those that included double foot-clasps and those that included thrusts. During the entire study, no female infant was observed to carry out the double- footclasp mount and the observed frequencies of pelvic thrusting were very low. A proportion
70 G.R. BROWN & A. E DIXSON
2 Frequency (per hour)
1
Foca l infant mounts other
I I
1 2 3 4 5 6
Month
Fig. 3. Mean hourly frequency with which male infants (solid circles, N=I4) and female infants (open circles, N=20) mounted other group mem- bers during the first six months of life. Means and SEMs: * p<0.001 (Mann Whitney U-test).
of the mounts exhibited by males were accompanied by a double-footclasp (16.7+5.4%) and by .pelvic thrusts (20.0+3.3%). The proportion of mounts accompanied by double foot-clasps did not differ between sons of high-/medium- (17.6+8.3%) or low-ranking mothers (15.6+6.9%) [t(14)=0.175, n.s.], nor between sons of multiparous (19.1+6.6%) or primiparous mothers (7.9+5.9%; t(14)=0.417, n.s.). Similarly, the total frequency of mounts involving thrusts ex- hibited by sons of high-/medium-ranking mothers (19.1+5.4%) did not differ from that of low-ranking mothers [21.3+3.4%; t(14)=0.321, n.s.], nor between sons of multiparous (21.5+ 3.7%) or primiparous mothers [14.4_+7.6%; t(14)=0.880, n.s.].
In contrast to the marked sex difference in the frequency of mounting by focal infants, the frequency with which focal infants were mounted by others did not differ between males and females. The overall mean frequency with which others mounted the focal infants during the whole of the six month period was low for male (3.1__+0.4) and female infants (4.0+0.7). The total frequency of mounting directed to the focal infant also did not differ with the rank or par- ity of the mother. Similarly, the frequency with which focal infants presented to other individu- als did not vary with the infant's sex or the mothert's rank or parity and the mean frequencies of presenting were low for male (7.3+3.5) and female infants (5.0+1.4).
MOTHER-INFANT PROXIMITY
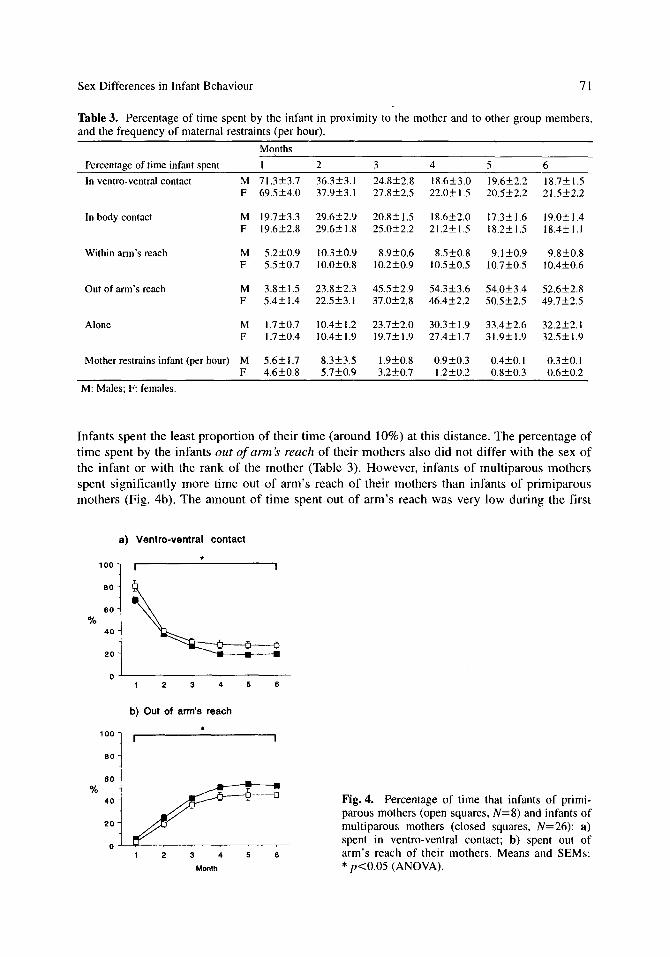
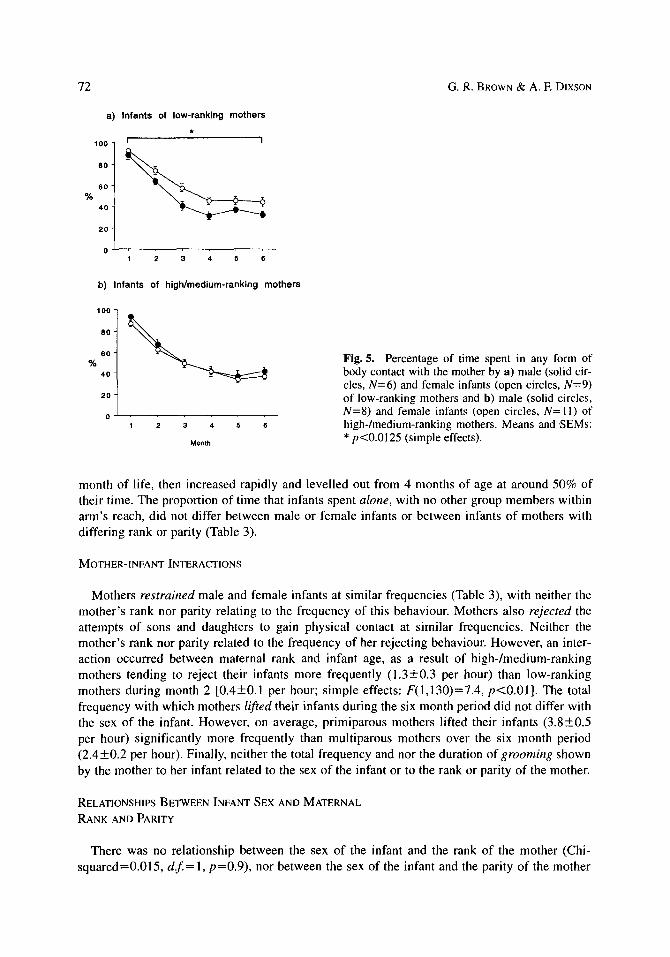
Male and female infants did not differ in the amount of time that they spent in ventro-ventral contact with their mothers (Table 3). During the first month of life, infants were spending around 70% of their time in ventro-ventral contact, and this decreased to around 20% by 6 months of age. The rank of the mother also did not influence this variable. However, infants of primiparous mothers spent significantly more time in ventro-ventral contact than infants of mul- tiparous mothers (Fig. 4a). The amount of time spent in any other form of physical contact with the mother did not differ with the sex of the infant or with the rank or parity of the mother, vary- ing from around 20 to 30% during the study period (Table 3). The total amount of time spent in body contact with the mother also tended to be greater for infants of primiparous mothers than infants of multiparous mother. A non-significant interaction between infant sex and maternal rank was found, as a result of daughters of low-ranking mothers spending more time in body contact than sons of low-ranking mothers [simple effects: F(1,26)=8.1, p<0.0125] and a ten- dency for daughters of low-ranking mothers to spend more time in body contact than daughters of high-/medium-ranking mothers [simple effects: F(1,26)=5.7, p<0.025; Fig. 5].
The amount of time that the infants spent within arm's reach of, but not touching, their moth- ers did not differ with the sex of the infant or with the rank or parity of the mother (Table 3).
Sex Differences in Infant Behaviour 71
Table 3. Percentage of time spent by the infant in proximity to the mother and to other group members, and the frequency of maternal restraints (per hour).
Months
Percentage of time infant spent I 2 3 4 5 6
In ventro-ventral contact M 71.3+3.7 36.3+3.1 24.8+2.8 18.6+3.0 19.6-+2.2 18.7-+1.5 F 69.5+4.0 37.9+3.1 27.8-+2.5 22.0_+1.5 20.5_+2.2 21.5+2.2
In body contact M 19.7+3.3 29.6+2.9 20.8+ 1.5 18.6+2.0 17.3+ 1.6 19.0+ 1.4 F 19.6+2.8 29.6+1.8 25.0+2.2 21.2+1.5 18.2+1.5 18.4+1.1
Within arm's reach M 5.2+0.9 10.3+0.9 8.9+0.6 8.5+0.8 9.1+0.9 9.8+0.8 F 5.5+0.7 10.0+0.8 10.2+0.9 10.5+0.5 10.7+0.5 10.4+0.6
Out of arm's reach M 3.8___ 1.5 23.8+2.3 45.5+2.9 54.3+3.6 54.0+3.4 52.6_+2.8 F 5.4+1.4 22.5+3�9 37.0-+2.8 46.4-+2.2 50.5+2.5 49.7+2.5
Alone M 1.7+0.7 10.4+ 1.2 23.7-+2.0 30.3-+ 1.9 33.4+2.6 32.2+2.1 F 1.7+0.4 10.4+1.9 19.7+1.9 27.4-+1.7 31.9+1.9 32.5+1.9
Mother restrains infant (per hour) M 5.6-+ 1.7 8.3+3.5 1.9+0.8 0.9+0.3 0.4-+0.1 0.3+0.1 F 4.6+0.8 5.7+0.9 3 .2-+0.7 1.2-+0.2 0.8+0.3 0.6+0.2
M: Males; F: females�9
Infants spent the least proport ion o f their t ime (around 10%) at this distance. The percentage o f
t ime spent by the infants out of arm's reach of their mothers also did not differ with the sex o f
the infant or with the rank of the mother (Table 3). However , infants o f mult iparous mothers
spent s ignif icantly more t ime out o f a rm's reach o f their mothers than infants o f pr imiparous
mothers (Fig. 4b). The amount o f t ime spent out o f a rm ' s reach was very low during the first
a)
1 O0
8O 2
6O %
4 O
2O
0
1 O0
80
60 %
4O
2O
0
Ventro-ventral contact
i
m - - - m
1 2 3 4
b) Out of arm's reach
5 6
! I
1 2 3 4 5 6
Month
Fig. 4. Percentage of time that infants of primi- parous mothers (open squares, N=8) and infants of multiparous mothers (closed squares, N=26): a) spent in ventro-ventral contact; b) spent out of arm's reach of their mothers�9 Means and SEMs: * p<0.05 (ANOVA).
72 G.R. BROWN & A. F. DIXSON
100 '
80"
6 0 " %
4 0
20'
0
a) Infants of low-ranking mothers
I I
1 2 3 4 5 6
b) Infants of high/medium-ranking mothers
1 O0 �9
80"
6 0 " %
4 0
2 0 "
0 1 2 3 4 5 6
Month
Fig. 5. Percentage of time spent in any form of body contact with the mother by a) male (solid cir- cles, N=6) and female infants (open circles, N=9) of low-ranking mothers and b) male (solid circles, N=8) and female infants (open circles, N=ll) of high-/medium-ranking mothers. Means and SEMs: * p<0.0125 (simple effects).
month of life, then increased rapidly and levelled out from 4 months of age at around 50% of their time. The proportion of time that infants spent alone, with no other group members within arm's reach, did not differ between male or female infants or between infants of mothers with differing rank or parity (Table 3).
MOTHER-INFANT INTERACTIONS
Mothers restrained male and female infants at similar frequencies (Table 3), with neither the mother's rank nor parity relating to the frequency of this behaviour. Mothers also rejected the attempts of sons and daughters to gain physical contact at similar frequencies. Neither the mother's rank nor parity related to the frequency of her rejecting behaviour. However, an inter- action occurred between maternal rank and infant age, as a result of high-/medium-ranking mothers tending to reject their infants more frequently (1.3+0.3 per hour) than low-ranking mothers during month 2 [0.4+0.1 per hour; simple effects: F(1,130)=7.4, p<0.01]. The total frequency with which mothers lifted their infants during the six month period did not differ with the sex of the infant. However, on average, primiparous mothers lifted their infants (3.8+0.5 per hour) significantly more frequently than multiparous mothers over the six month period (2.4+0.2 per hour). Finally, neither the total frequency and nor the duration of grooming shown by the mother to her infant related to the sex of the infant or to the rank or parity of the mother.
RELATIONSHIPS BETWEEN INFANT SEX AND MATERNAL
RANK AND PARITY
There was no relationship between the sex of the infant and the rank of the mother (Chi- squared =0.015, d.f. = 1, p=0.9), nor between the sex of the infant and the parity of the mother

Sex Differences in Infant Behaviour 73
(Chi-squared=0.058, d.f.= 1, p=0.8). Also, there was no relationship between the rank and par- ity of the mother (Chi-squared=0.186, d.f.= 1, p=0.7). While mothers of different rank did not differ in age [Mann-Whitney U(15,19)=127, n.s.], primiparous mothers were significantly younger on average (4.6+0.5 years, mean and SEM) than multiparous mothers [9.6+0.4 years; U(8,26) = 5.5, p<0.0001 ].
DISCUSSION
The results of this study indicate that male and female infant rhesus macaques differ in the frequencies with which they exhibit play and mounting behaviour during the first six months of life, in line with previous studies. However, other behaviours, such as the amount of time spent in proximity with the mother, were not found to differ for male and female infants. The strength of our results lies in the relatively large sample size used, the amount of detailed behavioural data collected and the social setting in which the animals were housed. The sex of the infant may be a much less important factor in certain aspects of behavioural development than previ- ously thought. Our data also provide further evidence that the social rank and the parity of the mother affect mother-infant interactions.
While previous studies have reported that male infants play more frequently than female infants in several species of primates, we have provided a detailed study of the development of play throughout early infancy. Stationary play was the first of the play categories to emerge; this occurred during the second month of life. Chasing play and rough-and-tumble play followed, from around 3 months of age. Female infants had lower average frequencies of play than male infants for all categories and not solely for the high energy types of play. By 5 and 6 months of age, females were playing at a frequency of around 10 bouts per hour, and exhibited roughly equal amounts of stationary, chasing and rough-and-tumble play. Male infants were playing at a frequency of around 15 bouts per hour by 5 and 6 months of age, with rough-and-tumble play being most frequently exhibited.
Male infants also initiated play with other individuals more frequently than female infants, especially during the third, fourth, and sixth months of life. While male infants were between 2 and 3 months of age, however, the play partner was more likely to be the one to initiate play. Other group members initiated play more frequently with male than female infants during this time period. The sex and age of those group members initiating play bouts with infants could be further investigated. The number of older siblings could influence the play behaviour of infants; during the fourth month of life, infants of multiparous mothers received more play initiations, and exhibited higher frequencies of stationary play, than infants of primiparous mothers. A pre- vious study similarly reported that sons of multiparous rhesus macaque mothers exhibit more play with peers than sons of primiparous mothers (SEAu 1966).
Infants of low-ranking mothers had slightly higher average frequencies of stationary play, chasing play, and total amounts of play than infants of high-/medium-ranking mothers. Infants of high-/medium-ranking mothers received similar amounts of play initiations from other indi- viduals as infants of low-ranking mothers during the first five months of life, and higher fre- quencies of play initiations during the sixth month; this suggests that infants of high- /medium-ranking mothers did not lack willing play partners. Indeed, a study on free-ranging baboons has reported that juvenile females initiate play more frequently with infants from higher-ranking matrilines (chacma baboons, Papio cynocephalus ursinus: CHENEu 1978). In contrast to our findings, a previous study on rhesus macaques suggested that l-yr old infants from high-ranking matrilines played more frequently than infants from low-ranking matrilines,

74 G.R. BROWN & A. E DIXSON
although this relationship was reversed for 3 yr olds (GARD & MEIER, 1977). Most studies of play during the juvenile period have reported that offspring from high-ranking matrilines play more frequently than those from low-ranking matrilines (vervets: FEDIGAN, 1972; rhesus macaques: TARTABIN1 & D1ENSKE, 1979). However, a more recent study of yellow baboons reported that daughters of low-ranking mothers played more than daughters of high-ranking mothers (WASSER & WASSER, 1995). Further studies to clarify rank effects on play behaviour and the play behaviour of individual animals would be of interest.
Male infants also exhibited higher frequencies of mounting behaviour than female infants during the first six months of life, in line with previous studies (GOLDFOOT & WALLEN, 1978; HINDE & SPENCER-BOOTH, 1967). The frequency with which male infants exhibited pelvic thrusting and the double-foot clasp position did not relate to maternal rank or parity, although individual males differed greatly in the extent to which they exhibited these additional patterns. The frequency with which infants exhibited presents to, and were mounted by, other individuals did not differ between any of the groups of infants.
Our results show that male and female infants did not differ in the amount of time spent in proximity to the mother and the mothers did not differ in the frequency with which they restrained, rejected, or groomed male or female infants. As reviewed in the Introduction, previ- ous studies supporting sex differences in these behaviour patterns were mostly carried out on singly-housed mother-infant pairs, while detailed studies of free-ranging populations generally did not demonstrate such differences. While the captive conditions used in our study may have influenced behavioural development, a comparative study of rhesus macaques in our captive environment and in semi-free ranging groups suggests that mother-infant interactions are simi- lar in the two settings (BERMAN, 1980).
In addition to investigating whether mothers interacted differently with sons and daughters, we investigated whether the behaviour of the mother correlated with her social rank or parity. In group-living rhesus macaques and free-ranging yellow baboons, mothers from high-ranking matrilines have been reported to restrict their infants less and reject their infants more than mothers of low rank (rhesus macaques: MAESTRIPIERI, 1998; yellow baboons: ALTMANN, 1980; WASSER & WASSER, 1995). Previous studies have also reported an interaction between the sex of the infant and the rank of the mother; sons of high-ranking mothers were rejected more fre- quently than daughters of high-ranking mothers (rhesus macaques: SIMPSON, 1983), and sons of low-ranking mothers spent less time in ventro-ventral contact with their mothers than daughters of low-ranking mothers (WASSER & WASSER, 1995). In contrast, recent studies on Japanese macaques have found no effect of maternal rank on the development of mother-infant relation- ships (TANAKA, 1989; SCHINO et al., 1995). Our results agree with the suggestion that the rank of the mother may influence the rejection behaviour of mothers but not with respect to the sex of the infant. Mothers of high/medium rank rejected their infants more frequently than low- ranking mothers during the second month of life.
Our results do, however, reveal an interaction between infant sex and maternal rank in that daughters of low-ranking mothers spent more time in physical contact with their mothers than sons of low-ranking mothers, with no difference being found for daughters and sons of high- ranking mothers. If previous studies that have suggested that daughters spend more time with mothers during infancy than sons were biased in their sample of mother-infant pairs, this may explain the inconsistancy of this reported sex difference in mother-infant relationships. This finding highlights the need to study a large sample of mother-infant pairs when investigating infant development and to take into account characteristics of the mother.
Our findings also reveal that primiparous and multiparous mothers differed in some measure- ments of mother-infant interactions. Infants o f primiparous mothers spent significantly more

Sex Differences in Infant Behaviour 75
time in ventro-ventral contact and less time out of arm's reach of the mother for the whole of the 6-month period, in line with previous studies on captive, socially-living rhesus macaques (HINDE & SPENCER-BOOTH, 1967) and singly-housed mother-infant pairs of cynomolgus macaques (NAKAMICHI et al., 1996). The presence or absence of older siblings may be an impor- tant factor explaining differences in mother-infant interactions between multiparous and primi- parous mothers. Primiparous mothers in our study also lifted their infants significantly more frequently than multiparous mothers, usually when another group member approached the infant or when there was a disturbance or fight in the group. These data support the suggestion that primiparous mothers act more protectively towards their infants than multiparous mothers (rhesus macaques: MITCHELL & STEVENS, 1968; SLAY, 1966; pigtail macaques: MAESTRIPIER1, 1998; Japanese macaques: TANAKA, 1989), although no differences in the amount of restraint or rejection exhibited by multiparous or primiparous mothers were revealed in our study. The pre- vious finding, that primiparous mothers act more protectively towards their daughters than their sons (HOOLEY & SIMPSON, 1981), was not upheld.
In summary, whilst the occurrence of sex differences in play and mounting behaviour have been confirmed by our study, previous reports of sex differences in mother-infant relationships have not been replicated. The only measure of mother-infant interactions that differed with infant sex was the amount of time spent in proximity to the mother by sons and daughters of low-ranking mothers. However, regardless of infant sex, mothers of different maternal rank and parity were found to differ in certain aspects of mother-infant interactions, and these maternal characteristics may have an important influence on the behavioural development of infant primates.
Acknowledgements. We are grateful to the following people for assisting in the collection of behavioural data: CLAIRE NEVISON, DAWN EAGLE, KHIA ATKINS, FRAN MARTEL, and KIRSTEN OLUFSEN. The research programme and monkey colony was supported by the British Medical Research Council. The rhesus mon- keys were maintained by DAVID RAYMENT and JEFF NIGHTINGALE. We are grateful to two anonymous refer- ees for comments on the manuscript.
REFERENCES
ALTMANN, J. 1980. Baboon Mothers and Infants. Harvard Univ. Press, Cambridge, Massachusetts. BAKEMAN, R.; GOTrMAN, J. M. 1986. Observing Interaction: An Introduction to Sequential Analysis.
Cambridge Univ. Press, Cambridge. BERMAN, C. M. 1980. Mother-infant relationships among free-ranging rhesus monkeys on Cayo Santiago:
a comparison with captive pairs. Anim. Behav., 28: 860-873. BERMAN, C. M. 1984. Variation in mother-infant relationships: traditional and non-traditional factors. In:
Female Primates: Studies by Women Primatologists, SMALL, M. F. (ed.), Alan Liss, New York, pp. J7-36.
BERMAN, C. M.; RASMUSSEN, K. L. R.; SUOMI, S. J. 1997. Group size, infant development and social net- works in free-ranging rhesus monkeys. Anim. Behav., 53: 405-421.
BROWN, G. R.; DIXSON, A. E 1999. Investigation of the role of postnatal testosterone in the expression of sex differences in behaviour in infant rhesus macaques (Macaca mulatta). Horm. Behav., 35: 186-194.
CHENEY, D. L. 1978. The play partners of immature baboons. Anim. Behav., 26: 1038-1050. DE JONGE, G.; DIENSKE, H.; VAN LUXEMBURG, E.; RmBENS, L. 1981. How rhesus monkey infants budget
their time between mothers and peers. Anim. Behav., 29: 598-609. DIXSON, A. F. 1977. Observations on the displays, menstrual cycles and sexual behaviour of the 'black
ape' of Celebes (Macaca nigra). J. Zool. Lond., 182: 63-84. FAIRBANKS, L. A. 1988. Vervet monkey grandmothers: effects on mother-infant relationships. Behaviour,
104: 176-188.

76 G.R. BROWN & A. E DIXSON
FEDIGAN, L. 1972. Social and solitary play in a colony of vervet monkeys (Cercopithecus aethiops). Primates, 13: 347-364.
GARD, G. C.; MEIER, G. W. 1977. Social and contextual factors of play behaviour in sub-adult rhesus mon- keys. Primates, 18: 367-377.
GOLDFOOT, D. A.; WALLEN, K. 1978. Development of gender role behaviors in heterosexual and isosexual groups of infant rhesus monkeys. In: Recent Advances in Primatology, Vol. 1: Behavior, CHIVERS, D. J.; HERBERT, J. (eds.), Academic Press, London, pp. 155-159.
GoY, R. W.; MCEWEN, B. S. 1980. Sexual Differentiation of the Brain. MIT Press, Cambridge, Massachusetts. HANBY, J. P.; BROWN, C. E. 1974. The development of sociosexual behaviors in Japanese macaques
(Macacafuscata). Behaviour, 49:151 - 196. HAUSER, M. D.; FAIRBANKS, L. A. 1988. Mother-offspring conflict in vervet monkeys: variation in response
to ecological conditions. Anim. Behav., 36: 802-813. HINDE, R. m.; ROWELL, T. E.; SPENCER-BOOTH, Y. 1964. Behaviour of socially living rhesus monkeys in
their first six months. Proc. Zool. Soc. Lond., 143: 609-649. HINDE, R. A.; SPENCER-BOOTH, Y. 1967. The behaviour of socially living rhesus monkeys in their first two
and a half years. Anim. Behav., 15: 169-196. HOOLEY, J. M.; SIMPSON, M. J. m. 1981. A comparison of primiparous and multiparous mother-infant
dyads in Macaca mulatta. Primates, 22: 379-392. HOOLEY, J. M.; SIMPSON, M. J. A. 1983. Influence of siblings on the infant's relationship with the mother
and others. In: Primate Social Relationships: An Integrated Approach, HINDE, R. A. (ed.), Blackwell Scientific Publ., Oxford, pp. 139-141.
JENSEN, G. D.; BOBmTT, R. A.; GORDON, B. N. 1968. Sex differences in the development of independence of infant monkeys. Behaviour, 30: 1-14.
LEE, P. C. 1984a. Early infant development and maternal care in free-ranging vervet monkeys. Primates, 25: 36-47.
LEE, P. C. 1984b. Ecological constraints on the social development of vervet monkeys. Behaviour, 91: 245- 262.
MAESTRIPIERI, D. 1998. Social and demographic influences on mothering style in pigtail macaques. Ethology, 104: 379-385.
MARTIN, P.; BATESON, P. 1986. Measuring Behaviour: An Introductory Guide. Cambridge Univ. Press, Cambridge.
MITCHELL, G. 1979. Behavioural Sex Differences in Nonhuman Primates. Van Nostrand Reinhold, New York.
MITCHELL, G.; BRANDT, E. M. 1970. Behavioral differences related to experience of mother and sex of infant in the rhesus monkey. Develop. Psychol., 3: 149.
MITCHEL, G.; STEVENS, C. W. 1968. Primiparous and multiparous monkey mothers in a mildly stressful social situation: first three months. Develop. Psychobiol., 1: 280-286.
NAKAMICHI, M. 1989. Sex differences in social development during the first four years in a free-ranging group of Japanese monkeys, Macacafuscata. Anim. Behav., 38: 737-748.
NAKAMICHI, M.; CHO, E; MINAMI, T. 1990. Mother-infant interactions of wild-born, individually-caged cynomolgus monkeys (Macaca fascicularis) during the first 14 weeks of infant life. Primates, 31: 213-224.
NAKAMICHI, M.; MINAMI, T.; CHO, F. 1996. Comparison between wild-born mother-female infant interac- tions and laboratory-born mother-female infant interactions during the first 14 weeks after birth in individually caged cynomolgus macaques. Primates, 37:155-166.
NEVISON, C. M.; BROWN, G. R.; DIXSON, A. E 1997. Effects of altering testosterone in early infancy on social behaviour in captive yearling rhesus monkeys. Phsiol. Behav., 62: 1397-1403.
NICOLSON, N. A. 1987. Infants, mothers, and other females. In: Primate Societies, SMUTS, B. B.; CHENEY, D. L.; SEYFARTH, R. M.; WRANGHAM, R. W.; STRUHSAKER, T. T. (eds.), Univ. of Chicago Press, Chicago, pp. 330-342.
OWENS, N. W. 1973. The development of behaviour in free-living baboons, Papio anubis. Unpubl. Ph.D. thesis, Univ. of Cambridge, Cambridge.
OWENS, N. W. 1975. Social play behaviour in free-living baboons, Papio anubis. Anim. Behav., 23: 387-408.
ROSENBLUM, L. A.; NADLER, R. D. 1971. The ontogeny of sexual behavior in male bonnet macaques. In: Influence of Hormones on the Nervous System, FORD, D. H. (ed.), Karger, Basel, pp. 126-139.
ROWELL, T. E.; CHISM, J. 1986. The ontogeny of sex differences in the behavior of paras monkeys. Int. J. Primatol., 7:83 - 107.

Sex Diffcrences in Infant Behaviour 77
S(:HINO, G.; D'A.'vlATO, F. R.; TROtSt, A. 1995. Mother-infant relationships in Japanese macaques: sources of inter-individual variation. Anita. Behav., 49:151 - 158.
SE^Y, B. 1966. Maternal behavior in primiparous and multiparous rhesus monkcys. Folia Primatol., 4: 146- 168.
SIF.GEI., S.; C,~STt3LLAN, N. J. 1988. Nonparametric Stastisticsfor the Behavioural Sciences. McGraw-Hill, London.
SIMPSON, M. J. A. 1983. Effect of the sex of an infant on the mother-infant relationship and the mother's subsequent reproduction. In: Prhnate Social Relationships: An Integrated Approach, HINDE, R. A. (ed.), Blackwell, Oxlord, pp. 53 -57 .
SIMPSON, M. J. A.; GORE, M. A.; JANUS, M.; RAYMENT, E D. G. 1989. Prior cxpcrience of risk and individ- ual differences in enterprise shown by rhesus monkey infants in the second half of their first year. Primates, 30: 493-509.
SYMONS, D. 1978. Play and Aggression: A Study of Rhesus Monkeys. Columbia Univ. Press, New York. TaNaKA, I. 1989. Variability in the development of mothcr-infant relationships among free-ranging
Japanese macaques. Primates, 30: 477-491. TARTABINI, A.; DIENSKE, H. 1979. Social play and rank order in rhesus monkeys (Macaca mulatta). Behav.
Process., 4: 375-383. WALLLN, K. 1996. Nature needs nurture: the interaction of hormonal and social influences on the develop-
ment of behavioral sex differences in rhesus monkeys. Horm. Behav., 30: 364-378. WALLEN, K.; MAESTRIPIERI, D.; MANN, D. R. 1995. Effects of neonatal testicular suppression with a GnRH
antagonist on social behavior in group-living juvenile rhesus monkeys. Horm. Behav., 29: 322-337. WASSER, L. M.; WASSER, S. K. 1995. Environmental variation and dcvelopmental rate among tree ranging
yellow baboons (Papio cynocephalus). Amen J. Primatol., 35: 15-30. Z;',R, J. H. 1974. Biostastistical Analysis. Prentice-Hall, New Jersey.
- - Received: March 5, 1999; Accepted: September 13, 1999
Authors" Names and Address: GILI.IAN R. BROWN and ALAN F. DIXSON. Sub-Deparmsent o[Animal Behaviom: University of Cambridge. Madingle); Cambridge. CB3 8AA. England. e-mail: [email protected]

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Copyright © 2022 FDOKUMEN




















