
Rhesus Macaques Simian Immunodeficiency Virus Infection of ...
17
of February 7, 2022. This information is current as Rhesus Macaques Simian Immunodeficiency Virus Infection of Activation in the Colon during Chronic Axis Is a Potential Mediator of Immune Acetyl p65 - SIRT1 - Dysregulated miR-34a Xavier Alvarez Mahesh Mohan, Vinay Kumar, Andrew A. Lackner and ol.1401447 http://www.jimmunol.org/content/early/2014/11/29/jimmun published online 1 December 2014 J Immunol Material Supplementary 7.DCSupplemental http://www.jimmunol.org/content/suppl/2014/11/29/jimmunol.140144 average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2014 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on February 7, 2022 http://www.jimmunol.org/ Downloaded from by guest on February 7, 2022 http://www.jimmunol.org/ Downloaded from
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of Rhesus Macaques Simian Immunodeficiency Virus Infection of ...

of February 7, 2022.This information is current as Rhesus Macaques
Simian Immunodeficiency Virus Infection of Activation in the Colon during ChronicAxis Is a Potential Mediator of Immune
Acetyl p65−SIRT1−Dysregulated miR-34a
Xavier AlvarezMahesh Mohan, Vinay Kumar, Andrew A. Lackner and
ol.1401447http://www.jimmunol.org/content/early/2014/11/29/jimmun
published online 1 December 2014J Immunol
MaterialSupplementary
7.DCSupplementalhttp://www.jimmunol.org/content/suppl/2014/11/29/jimmunol.140144
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2014 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on February 7, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on February 7, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Dysregulated miR-34a–SIRT1–Acetyl p65 Axis Is a PotentialMediator of Immune Activation in the Colon during ChronicSimian Immunodeficiency Virus Infection of RhesusMacaques
Mahesh Mohan, Vinay Kumar, Andrew A. Lackner, and Xavier Alvarez
Persistent gastrointestinal inflammation, a hallmark of progressive HIV/SIV infection, causes disruption of the gastrointestinal
epithelial barrier, microbial translocation, and generalized immune activation/inflammation driving AIDS progression. Apart from
protein regulators, recent studies strongly suggest critical roles for microRNAs (miRNAs) in regulating and managing certain
aspects of the inflammatory process. To examine their immunoregulatory role, we profiled miRNA expression in the colon from
12 chronic SIV-infected and 4 control macaques. After applying multiple comparisons correction, 10 (3 upregulated and 7
downregulated) miRNAs showed differential expression. Most notably, miR-34a showed significant upregulation in both epithelial
and lamina propria leukocyte (LPL) compartments. Intense gH2A.X expression in colonic epithelium and LPLs confirmed the
contribution of DNA damage response in driving miR-34a upregulation. SIRT1 mRNA and protein decreased significantly in both
colonic epithelium and LPLs. Luciferase reporter assays validated rhesus macaque SIRT1 as a direct miR-34a target. Decreased
SIRT1 expression was associated with constitutively enhanced expression of the transcriptionally active form of the p65 (acety-
lated on lysine 310) subunit of NF-kB exclusively in the LPL compartment. The intensity and number of acetylated p65+ cells was
markedly elevated in LPLs of chronically SIV-infected macaques compared with uninfected controls and localized to increased
numbers of IgA+ and IgG+ plasma cells. These findings provide new insights into the potential role of the miR-34a–SIRT1–p65 axis
in causing hyperactivation of the intestinal B cell system. Our results point to a possible mechanism where the normal immuno-
suppressive function of SIRT1 is inhibited by elevated miR-34a expression resulting in constitutive activation of acetylated p65
(lysine 310). The Journal of Immunology, 2015, 194: 000–000.
Regardless of the route of transmission, mucosal tissues,particularly the gastrointestinal (GI) tract, are targeted byHIV/SIV, leading to rapid, severe, and sustained depletion
of CD4+ T cells in HIV-infected individuals and SIV-infectedrhesus macaques (1–5). As disease progresses, GI complicationssuch as anorexia, weight loss, and diarrhea become frequent andare being reported in patients despite the extensive use of highlyactive antiretroviral therapy (6). Histologically, GI disease ischaracterized by infiltration of the lamina propria by T cells,plasma cells, macrophages, and morphologic changes such asvillus blunting and crypt hyperplasia. An emerging feature ofHIV/SIV pathogenesis is the markedly elevated levels of micro-
bial translocation that occur in the later stages of infection (7, 8).This phenomenon has been proposed to play a key role in driv-ing localized and systemic immune activation, which is a well-recognized correlate of HIV/SIV disease progression. The mech-anisms leading to increased microbial translocation in AIDSpatients remain largely unknown. However, the “leaky gut syn-drome” is a preferred hypothesis, wherein luminal bacteria and/ortheir products enter the intestinal lamina propria through a dis-rupted epithelial barrier and pass via the portal blood into thesystemic circulation.Viral replication and CD4+ T cell depletion in the lamina
propria leukocyte (LPL) compartment is associated with elevatedexpression of proinflammatory genes and reduced expression ofgenes involved in maintenance of epithelial barrier, repair, anddigestive and metabolic functions (9–12). Furthermore, focusedlongitudinal examination of individual mucosal compartments hasrevealed deeper insights into the molecular pathological eventsoccurring in the intestinal LPL and epithelial compartments (13,14). Whereas inflammation and immune activation–related genesshowed marked changes in the LPL compartment, genes regu-lating enterocyte maturation, differentiation, and epithelial barrierfunction such as Wnt-TCF7L2, Notch signaling proteins, adherensjunction, hemidesmosomes, and desmosomes were found to besignificantly dysregulated in the epithelial compartment followingSIV infection (13, 14). Overall, these studies demonstrated con-siderable alterations in enterocyte structure and function thatcould facilitate microbial translocation.Although multiple mechanisms involving transcription factors,
chromatin modifications, and others such as histone modificationsare known to regulate gene expression, one important mechanism
Division of Comparative Pathology, Tulane National Primate Research Center,Covington, LA 70433
Received for publication June 5, 2014. Accepted for publication November 2, 2014.
This work was supported by National Institutes of Health Grants R01DK083929 (toM.M.), AI084793, and OD011104 (formerly RR00164).
The TLDA miRNA data in this article has been submitted to the Gene ExpressionOmnibus (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE60773) under ac-cession number GSE60773.
Address correspondence and reprint requests to Dr. Mohan Mahesh, Tulane NationalPrimate Research Center, 18703 Three Rivers Road, Covington, LA 70433. E-mailaddress: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: DDR, dsDNA damage response; Del, deleted;DSS, dextran sulfate sodium; GI, gastrointestinal; IBD, inflammatory bowel disease;IF, immunofluorescence; LNA, locked nucleic acid; LPL, lamina propria leukocyte;miRNA, microRNA; qRT-PCR, quantitative RT-PCR; ROI, region of interest; TLDA,TaqMan low-density array; UTR, untranslated region; WT, wild-type.
Copyright� 2014 by The American Association of Immunologists, Inc. 0022-1767/14/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1401447
Published December 1, 2014, doi:10.4049/jimmunol.1401447 by guest on February 7, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

mediated by small regulatory RNAs called microRNAs (miRNAs)has gained a lot of attention recently (15). miRNAs are ∼21–23 ntin length and have been described to impact almost all cellularprocesses by repressing gene expression at the posttranscriptionallevel (15). A growing body of evidence indicates that HIV in-fection is characterized by dysregulated miRNA expression (16),including direct targeting and crippling of the miRNA biosyn-thesis machinery by HIV (17). Recent studies performed in SIV-infected rhesus macaques also demonstrated dysregulated miRNAexpression in plasma (18), brain (19), and monocyte derivedmacrophages (20). We recently reported altered miRNA expres-sion in the intestine during acute SIV infection (21). More spe-cifically, we identified miR-190b to be significantly upregulated asearly as 7 d after SIV infection, and its expression remained ele-vated throughout SIV infection. Additional studies also suggestedthat miR-190b could influence disease pathogenesis by directlybinding to the 39 untranslated region (UTR) and regulating theexpression of MMTR6, a phosphatidylinositol 1,3-bisphosphatepreviously shown to inhibit T cell and macrophage activation. Inthe present study, we performed miRNA profiling in colon duringchronic SIV infection and detected marked changes in the ex-pression of miRNAs linked to inflammation, cell cycle arrest, andsenescence. Among these, the expression of miR-34a, a miRNAshown to regulate apoptosis, cell cycle control, and senescence(22–24), was markedly increased in both colonic epithelial andLPL compartments. Furthermore, increased miR-34a expressionwas associated with reduced expression of SIRT1, a longevity-inducing anti-inflammatory, anti-stress, and anti-aging class IIIhistone deacetylase in the colonic epithelium and LPLs. Mostnoticeably, reduced SIRT1 expression in the LPL compartmentwas accompanied by enhanced expression of acetylated p65 (ly-sine 310), suggesting hyperactivation of the NF-kB signalingpathway. Furthermore, enhanced constitutive expression of acet-ylated p65 (lysine 310) was localized to lamina propria plasmacells. The results of our study indicate that the miR-34a–SIRT1–acetyl p65 axis represents a potential molecular pathologicalmechanism that could drive chronic hyperactivation of the intes-tinal B cell system, leading to their dysfunction.
Materials and MethodsAnimal ethics statement
All experiments using rhesus macaques were approved by the TulaneUniversity Institutional Animal Care and Use Committee (protocol no.3574). The Tulane National Primate Research Center is an Association forAssessment and Accreditation of Laboratory Animal Care International–accredited facility (no. 000594). The National Institutes of Health Office ofLaboratory Animal Welfare assurance number for the Tulane NationalPrimate Research Center is A3071-01. All clinical procedures, includingadministration of anesthesia and analgesics, were carried out under thedirection of a laboratory animal veterinarian. Animals were anesthetizedwith ketamine hydrochloride for blood collection procedures. Intestinalresections were performed by laboratory animal veterinarians. Animalswere preanesthetized with ketamine hydrochloride, acepromazine, andglycopyrolate, intubated, and maintained on a mixture of isoflurane andoxygen. Buprenorphine was given intraoperatively and postoperatively foranalgesia. All possible measures are taken to minimize discomfort of allthe animals used in this study. Tulane University complies with NationalInstitutes of Health policy on animal welfare, the Animal Welfare Act, andall other applicable federal, state, and local laws.
Animals and tissue collection
Colon tissues were collected from a total of 61 Indian-origin rhesusmacaques, including 45 animals infected with pathogenic strains of SIV thatuse CCR5 in vivo and 16 uninfected macaques. All macaques were infectedwith SIVmac251. Animal IDs, duration of infection, route of infection,plasma and colonic viral loads, and intestinal histopathology in all SIV-infected animals are provided in Table I. For i.v. inoculation, a viraldose of 100 fifty percent tissue culture infective dose TCID50 was used.
However, for intravaginal and intrarectal inoculation, a higher viral dose of500 and 1000 fifty percent tissue culture infective dose, respectively, wasused. Twelve chronic SIV-infected animals (Table I) and four controlanimals (Table I) were used for TaqMan low-density array (TLDA)miRNA profiling. An additional 45 animals comprising 5 animals each at7–10 and 13–14 d postinfection, 7 each at 21 and 90 d postinfection, 9chronic SIV-infected, and 12 control macaques (Table I) were included forquantitative RT-PCR (qRT-PCR) confirmation, in situ hybridization, andimmunofluorescence studies. Data regarding viral inoculum, duration ofinfection, and plasma and intestinal viral loads for the 24 animals at 7–10,13–14, 21, and 90 d postinfection have been previously reported (21). Inthe control group, colon resection segments (∼5 cm long) were collectedfrom three macaques (HF54-Pre, HR42-Pre, HD08-Pre) before SIV in-fection (Table I).
All tissues were collected at necropsy in RNAlater (Ambion, Austin, TX) fortotal RNA extraction and qRT-PCR. For histopathologic evaluation, colontissues were fixed in 10% neutral buffered formalin, embedded in paraffin,sectioned at 7 mm, and stained with H&E for analysis. Sections were examinedin a blinded fashion, and histological signs of inflammation were scoredsemiquantitatively on a scale of 0–3 as follows: 0, within normal limits; 1,mild; 2, moderate; 3, severe. Additionally, the presence of crypt dilatation/cryptitis, amyloidosis, and lymphoid hyperplasia were also recorded (Table I).Mild changes such as lymphoid hyperplasia were assigned a score of 1.
Global miRNA profiling using TLDAs
For TLDA studies ∼350 ng total RNA from intact colon tissue was firstreverse transcribed, preamplified, and loaded onto TLDA cards and pro-cessed according to the manufacturer’s recommended protocol describedpreviously (21).
Quantitative real-time TaqMan stem loop miRNA and SYBRGreen RT-PCR
Expression of miR-34a and SIRT1 mRNAwas further investigated in bothcolonic epithelium and LPL compartments using qRT-PCR as previouslydescribed (21). For SIRT1 qRT-PCR assays, ∼1 mg total RNA was firstreverse transcribed using the SuperScript. III first-strand synthesis systemfor RT-PCR kit following the manufacturer’s protocol. Each qRT-PCRreaction (20 ml) contained the following: 23 Power SYBR Green mastermix without uracil-N-glycosylase (12.5 ml); SIRT1 forward, 59-AAGATGAGCCTGATATTCCAGAGAGAGCTG-39, reverse, 59-GCCTCTTGATCAT-CTCCATCAGTCCCAAA-39; GAPDH forward, 59-CAAGAGAGGCATT-CTCACCCTGAA-39, reverse, 59-TGGTGCCAGA TCTTCTCCATGTC-39;and b-actin forward, 59-CAACAGCCTCAAGATCGTCAGCAA-39, reverse59-GAGTCCTTCCACGATACCAAAGTTGTC-39 (200 nM); and cDNA (4ml).
Colonic epithelial and lamina propria cell isolation
To determine the mucosal compartment contributing to miR-34a upregu-lation, we separated the intestinal epithelial cells from the underlying LPLsand fibrovascular stroma as described previously (21).
In situ hybridization and immunofluorescence
In situ hybridization for miR-34a was performed using locked nucleicacid (LNA)-modified DNA probes (Exiqon, Vedbaek, Denmark). Briefly,7-mm formalin-fixed, paraffin-embedded tissue sections were first depar-afinized, rehydrated in descending series of ethanol, and pretreated in amicrowave with citrate buffer (Ag unmasking solution; Vector Labo-ratories, Burlingame, CA) for 20 min at high power according to themanufacturer’s instructions. Thereafter, sections were thoroughly washed,placed in a humidified chamber, and hybridized overnight at 50˚C with60 nM digoxigenin-labeled LNA-modified miR-34a DNA probes (5DigN/ACAACCAGCTAAGACACTGCCA/3Dig_N/, RNA melting temperatureof 85˚C) or scrambled probe (5DigN/GTGTAACACGTCTATACGCCCA/3Dig_N/, RNA melting temperature of 87˚C) in 13 miRNA in situ hy-bridization buffer (Exiqon). After hybridization, slides were washed with53 SSC (SSC buffer), 13 SSC, 0.23 SSC, and blocked with Dakoprotein-free blocker (Dako Laboratories) for 1 h. Fab fragments of ananti–digoxigenin Ab conjugated with alkaline phosphatase (Roche Diag-nostics, Penzberg, Germany) (1 in 250) were used to detect digoxigenin-labeled probes. Positive signals were detected by incubating the sectionswith permanent red (Dako Laboratories) for 7 min. Controls includedmatched tissues hybridized with LNA-modified scrambled probes.
Immunofluorescence (IF) for detecting gH2A.X (9F3, 1 in 250) (Abcam),acetyl p65 (lysine 310; 1 in 100) (Genetex), SIRT1 (E104, 19A7AB4, 1F3)(Abcam), SIRT1 (Bioss), SIRT1 (LS-B8356, 1 in 62.5) (LifeSpan Bio-sciences), and p21 (12D1, 1 in 50) (Cell Signaling Technology) was done asdescribed previously (10). Immunophenotyping of acetylated p65+ cells was
2 miR-34a UPREGULATION IN COLON DURING CHRONIC SIV INFECTION
by guest on February 7, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
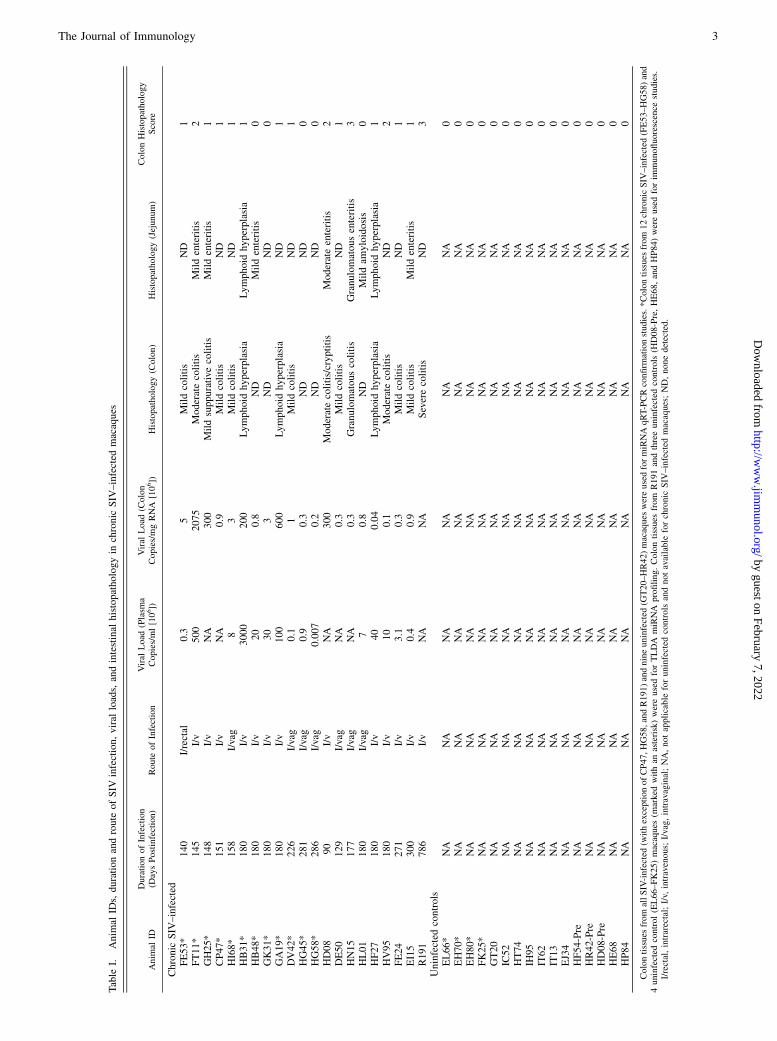
Table
I.Anim
alID
s,durationandroute
ofSIV
infection,viral
loads,andintestinal
histopathologyin
chronic
SIV
–infected
macaques
Anim
alID
DurationofInfection
(DaysPostinfection)
Route
ofInfection
Viral
Load
(Plasm
aCopies/ml[106])
Viral
Load
(Colon
Copies/mgRNA
[106])
Histopathology(Colon)
Histopathology(Jejunum)
ColonHistopathology
Score
Chronic
SIV
–infected
FE53*
140
I/rectal
0.3
5Mildcolitis
ND
1FT11*
145
I/v
500
2075
Moderate
colitis
Mildenteritis
2GH25*
148
I/v
NA
300
Mildsuppurative
colitis
Mildenteritis
1CP47*
151
I/v
NA
0.9
Mildcolitis
ND
1HI68*
158
I/vag
83
Mildcolitis
ND
1HB31*
180
I/v
3000
200
Lymphoidhyperplasia
Lymphoid
hyperplasia
1HB48*
180
I/v
20
0.8
ND
Mildenteritis
0GK31*
180
I/v
30
3ND
ND
0GA19*
180
I/v
100
600
Lymphoidhyperplasia
ND
1DV42*
226
I/vag
0.1
1Mildcolitis
ND
1HG45*
281
I/vag
0.9
0.3
ND
ND
0HG58*
286
I/vag
0.007
0.2
ND
ND
0HD08
90
I/v
NA
300
Moderatecolitis/cryptitis
Moderateenteritis
2DE50
129
I/vag
NA
0.3
Mildcolitis
ND
1HN15
177
I/vag
NA
0.3
Granulomatous
colitis
Granulomatous
enteritis
3HL01
180
I/vag
70.8
ND
Mildam
yloidosis
0HF27
180
I/v
40
0.04
Lymphoidhyperplasia
Lymphoid
hyperplasia
1HV95
180
I/v
10
0.1
Moderate
colitis
ND
2FE24
271
I/v
3.1
0.3
Mildcolitis
ND
1EI15
300
I/v
0.4
0.9
Mildcolitis
Mildenteritis
1R191
786
I/v
NA
NA
Severecolitis
ND
3Uninfected
controls
EL66*
NA
NA
NA
NA
NA
NA
0EH70*
NA
NA
NA
NA
NA
NA
0EH80*
NA
NA
NA
NA
NA
NA
0FK25*
NA
NA
NA
NA
NA
NA
0GT20
NA
NA
NA
NA
NA
NA
0IC52
NA
NA
NA
NA
NA
NA
0HT74
NA
NA
NA
NA
NA
NA
0IH
95NA
NA
NA
NA
NA
NA
0IT62
NA
NA
NA
NA
NA
NA
0IT13
NA
NA
NA
NA
NA
NA
0EJ34
NA
NA
NA
NA
NA
NA
0HF54-Pre
NA
NA
NA
NA
NA
NA
0HR42-Pre
NA
NA
NA
NA
NA
NA
0HD08-Pre
NA
NA
NA
NA
NA
NA
0HE68
NA
NA
NA
NA
NA
NA
0HP84
NA
NA
NA
NA
NA
NA
0
Colontissues
from
allSIV-infected
(withexceptionofCP47,HG58,andR191)andnineuninfected
(GT20–HR42)macaques
wereusedformiRNAqRT-PCRconfirm
ationstudies.*Colontissues
from
12chronicSIV
–infected
(FE53–HG58)and
4uninfected
control(EL66–FK25)macaques
(marked
withan
asterisk)wereusedforTLDA
miRNA
profiling.Colontissues
from
R191andthreeuninfected
controls
(HD08-Pre,HE68,andHP84)wereusedforim
munofluorescence
studies.
I/rectal,intrarectal;I/v,
intravenous;
I/vag,intravaginal;NA,notapplicable
foruninfected
controls
andnotavailable
forchronic
SIV
–infected
macaques;ND,nonedetected.
The Journal of Immunology 3
by guest on February 7, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

done using CD68 (1 in 20) (Dako, Glostrup, Denmark), CD163 (1 in 50)(Serotec, Raleigh, NC), CD3 (1 in 20) (Dako), anti-rhesus IgA (1 in 250),IgG (1 in 250) (Both Abs from Research Diagnostics, Flanders, NJ), andappropriate Alexa Fluor–conjugated secondary Abs.
Quantitative image analysis
Quantitation of cells and regions of interest (ROI) labeled by LNA-modifiedmiR-34a in situ hybridization probes was performed using Volocity 5.5 soft-ware (PerkinElmer, Waltham, MA) after capturing images on a Leica confocalmicroscope. Several ROI were hand drawn on the epithelial and LPL regionsin the images from colon. The data were first graphed and then analyzedusing the Mann–Whitney U test employing the Prism v5 software (GraphPadsoftware). A p value ,0.05 was considered statistically significant.
Cloning of 39 UTR of SIRT1 mRNA and Dual-Glo luciferasereporter gene assay
The 39 UTR of the rhesus SIRT1 mRNA contains two predicted miR-34abinding sites (TargetScan 6.2), that is, site 1, 59-UUCCACAAGUAUUA-AACUGCCAA-39 (nt 891–897), and site 2, 59-CCAGCUAGGACCAU-UACUGCCAG-39 (nt 1434–1440) (Table II). Accordingly, a G-blockdsDNA sequence (∼125 bases long) containing both miR-34a bindingsites along with ∼30 bases flanking the 59 and 39 ends of each site wassynthesized (Integrated DNA Technologies, Coralville, IA) for cloninginto the pmirGLO dual luciferase vector (Promega, Madison, WI). Asecond G-block sequence with the exact same flanking UTR sequences butwith both miR-34a binding sites (total 14 nt from both sites) deleted wasalso synthesized to serve as a negative control. Both oligonucleotidesequences were synthesized with a PmeI site on the 59 and XbaI site on the39 end for directional cloning. The pmirGLO vector was first cut with PmeIand XbaI restriction enzymes, gel purified, and ligated with either the wild-type (WT) sequence containing miR-34a binding sites (SIRT1-WT) or thedeleted (Del) sequence (SIRT1-Del). HEK293 cells were plated at a den-sity of 5 3 104 cells per well of a 96-well plate. At 60% confluence, cellswere cotransfected with ∼100 ng SIRT1-WT or SIRT1-Del UTR miRNAluciferase reporter vector and 100 nM miR-34a or negative control mimic(59-CCACAACCUCCUAGAAAGAGUAGA-39) using the DharmaFECTDuo transfection reagent (Thermo Fisher Scientific). In separate wells,cells were also transfected with pmirGLO vector (Promega) as a nor-malization control. After 72 h, the Dual-Glo luciferase assay was per-formed according to the manufacturer’s recommended protocol usingthe BioTek H4 Synergy plate reader (BioTek, Winooski, VT). Thenormalized firefly/renilla ratio was calculated to determine the relativereporter activity. Experiments were performed in six replicates andrepeated twice.
Quantitation of mucosal viral loads
Total RNA samples from all SIV-infected animals were subjected toa quantitative real-time TaqMan two-step RT-PCR analyses to determine theviral load in colon samples. Briefly, primers and probes specific to the SIVlong terminal repeat sequence were designed and used in the TaqMan real-time RT-PCR assay. Probes were conjugated with a fluorescent reporter dye(FAM) at the 59 end and a quencher dye at the 39 end. Fluorescence signalwas detected using the ABI Prism 7900 HT Fast Real-Time PCR System(Life Technologies). Data were captured and analyzed with SequenceDetection software v2.4 (Life Technologies). Viral copy number was de-termined by plotting CT values obtained from the colon and jejunumsamples against a standard curve (y = 23.384x + 39.029) (r2 = 0.998)generated with in vitro–transcribed RNA representing known viral copynumbers.
Data analysis and data availability
TLDA-SDS–run files from 12 SIV-infected and 4 control animals were loadedonto Relative Quantification Manager software v1.2.1 and analyzed usingautomatic baseline settings and manual threshold of 0.2. One (EL66) of thefour control samples was selected as a calibrator and set to 1. The remaining15 samples were expressed as an n-fold difference relative to the calibratorsample. The results from the Relative Quantification Manager analysis con-taining five columns (well, sample, detector, task, and CT values) were savedas a tab-delimited text file, imported, and analyzed using the DataAssist v3.01software (Life Technologies) employing the global normalization method, asthis method has been proposed to be more sensitive and accurate for ana-lyzing high-throughput TaqMan miRNA qRT-PCR than endogenous controls(25, 26). The CT upper limit was set to 33, meaning that all miRNA detectorswith a CT value $33 were excluded. Multiple comparisons correction forTLDA miRNA profiling (simultaneously applied to all 768 miRNA targets oncards A and B) was performed using the Benjamini–Hochberg method. Ap value,0.05 was considered significant. TLDA miRNA data were depositedwith Gene Expression Omnibus (accession no. GSE60773, http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE60773).
For miR-34a and SIRT1 qRT-PCR studies, one uninfected control macaquewith the highest (for miR-34a) or lowest (for SIRT1) DDCT value served asthe calibrator/reference and was assigned a value of 1. All differentiallyexpressed miRNAs or mRNAs in SIV-infected and other control macaquesare shown as an n-fold difference relative to this macaque. This approach waspreferred over calculating miRNA/mRNA fold change using an average of allcontrol animal DCT values mainly to facilitate graphing the control samplesso that variation within the control group can be displayed. Individual miR-34a and SIRT1 mRNA qRT-PCR data were analyzed by a nonparametricWilcoxon rank sum test for independent samples using RealTime StatMinersoftware (Integromics). A Spearman nonparametric one-tailed correlationanalysis was performed to determine the degree of association between coloninflammation scores/tissue viral loads and miR-34a fold expression. Firefly/renilla ratios were analyzed using one-way ANOVA, and post hoc compar-isons were performed using a Tukey multiple comparisons test.
ResultsMucosal and plasma viral loads, CD4+ T cell counts, andintestinal histopathology
All chronic SIV-infected macaques obtained at necropsy hadsubstantial viral loads in the colon that ranged from 0.04 3 106 to2075 3 106 copies/mg total RNA with a median of 0.9 3 106
copies/mg total RNA (Table I). Similar to colonic viral loads,plasma viral loads in chronic SIV-infected macaques ranged from0.3 3 106 to 5000 3 106 copies/ml with a median of 15 3 106
copies/ml. Plasma viral loads were not available for 6 animals(GH25, CP47, HD08, DE50, HN15, and R191).All chronic SIV-infected animals with the exception of four
(HF27, HB48, GK31, and HG58) progressed to AIDS. CD4+ T celldata were only available for eight chronic SIV-infected macaquesand, as shown in Supplemental Fig. 1, all eight had significantmucosal CD4+ T cell depletion compared with uninfected controlmacaques. Histologic evaluation of H&E-stained sections of colon(Table I) from all SIV-infected macaques revealed the presence ofmild to severe colitis including other intestinal lesions such ascrypt abscess/cryptitis (Table I). In contrast to the colon, only 9 of
Table II. Schematic representation of SIRT1 39 UTR depicting predicted binding site for miR-34a.
GeneGenBank
Accession No.Site
Conservation
BindingSites on39 UTR Target Site Sequence
PredictionAlgorithm
SIRT1(proximal site)
NM_001142498 Human/chimp/macaque/
orangutan/mouse
888–894 5’ . . .UUCCACAAGUAUUAAACUGCCAA. . .3’||||||
3’ UGUUGGUCGAUUCUGUGACGGU 5’
TargetScan (38)
SIRT1(distal site)
NM_001142498 Human/chimp/macaque/
orangutan/mouse
1428–1434 5’ . . .CCAGCUAGGACCAUUACUGCCAG. . .3’||||||| ||||||
3’UGUUGGUCGAUUCUG----UGACGGU 5’
TargetScan (38)
For alignment of the SIRT1 sequence with miR-34a, the top strand shows SIRT1 mRNA and the bottom sequence shows miR-34a.
4 miR-34a UPREGULATION IN COLON DURING CHRONIC SIV INFECTION
by guest on February 7, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

Table
III.
Changes
inmiRNA
expressionin
colonduringchronic
SIV
infection
miRNA
ID
Chronic
SIV
(n=12)
Uninfected
Controls
(n=4)
HB31
HB48
GK31
GA19
GH25
FT11
DV42
CP47
HI68
FE53
HG45
HG58
EL66
EH70
EH80
FK25
FoldChange
Adjusted
pValue
hsa-m
iR-181-a*
24.69
23.93
24.36
24.71
24.12
24.72
24.90
25.04
25.62
24.53
24.18
24.98
23.91
23.99
23.72
24.73
21.5
0.0027
hsa-m
iR-628-5p5p
25.93
25.04
25.40
25.51
24.78
24.97
25.08
25.09
25.21
24.99
24.17
25.18
24.10
24.37
24.12
25.10
21.6
0.0027
hsa-m
iR-378
25.03
24.87
25.34
25.27
24.65
25.70
25.27
25.76
26.11
24.71
25.07
25.46
24.93
24.87
24.95
25.83
21.2
0.0174
hsa-m
iR-424
25.92
25.16
25.54
25.32
25.13
25.27
25.97
25.92
26.52
25.18
26.09
25.92
26.07
26.06
25.84
26.94
1.4
0.0174
hsa-m
iR-425*
25.22
24.29
24.98
25.06
25.19
25.41
25.44
25.38
25.91
24.68
24.96
25.13
24.44
24.57
24.24
24.97
21.5
0.0174
hsa-m
iR-454*
29.52
28.81
29.09
29.78
29.25
29.79
29.77
29.31
29.51
29.21
29.02
29.39
28.23
28.60
28.42
28.97
21.8
0.0174
hsa-m
iR-223*
25.99
24.89
25.66
25.48
25.57
24.65
25.35
26.13
26.65
26.33
26.53
26.75
26.54
27.23
26.67
28.97
2.7
0.019
hsa-m
iR-24-2*
25.36
24.79
25.31
24.98
25.04
24.75
24.92
25.58
25.33
24.98
24.65
25.22
24.51
24.78
24.40
25.18
21.3
0.019
hsa-m
iR-520c-3p
32.61
29.79
32.98
31.59
31.06
31.12
31.94
32.16
31.29
31.80
31.35
31.20
30.51
30.43
30.33
31.20
21.8
0.019
hsa-m
iR-34a
21.44
20.92
20.74
20.39
19.43
19.67
20.32
21.32
21.14
20.82
19.96
21.01
21.76
21.97
20.97
22.16
2.2
0.0306
hsa-m
iR-34a*
24.17
23.40
23.32
22.79
22.21
23.14
23.90
24.83
24.38
24.37
23.53
24.67
24.99
25.56
24.60
25.11
2.6
0.0515
hsa-let-7b
18.06
17.44
17.62
17.68
17.00
17.77
17.98
18.00
18.51
17.24
17.51
17.87
17.98
18.21
17.72
18.68
1.3
0.0578
hsa-m
iR-193b*
27.03
26.74
26.67
26.42
25.97
26.56
27.79
28.64
28.21
28.00
26.18
28.62
27.80
28.45
28.01
29.03
2.3
0.0589
hsa-m
iR-146b-5p
16.11
15.98
16.81
15.35
15.29
14.79
15.72
16.44
16.67
16.20
16.47
16.59
17.21
17.05
16.23
18.16
2.1
0.0719
hsa-m
iR-30d
20.05
20.23
20.70
20.38
20.08
20.48
20.40
21.02
20.71
19.94
19.81
20.76
19.73
19.90
19.86
20.68
21.3
0.0758
hsa-m
iR-744*
26.22
25.74
25.97
26.25
25.91
27.28
26.71
26.78
27.15
25.89
25.96
26.54
25.72
25.56
25.53
26.51
21.5
0.0796
hsa-m
iR-18a
20.89
20.28
20.64
20.25
20.09
20.69
21.24
20.44
20.70
21.05
21.14
21.00
20.87
21.18
20.79
21.97
1.4
0.0825
hsa-m
iR-21
15.76
15.79
16.05
14.98
14.68
14.42
15.07
15.45
15.93
15.35
15.60
16.25
15.91
15.98
15.70
16.66
1.6
0.0825
hsa-m
iR-422a
24.87
24.01
24.74
24.78
24.01
24.30
23.57
24.44
24.92
23.44
23.89
23.80
23.43
23.23
22.88
24.08
21.8
0.0825
hsa-m
iR-656
26.98
25.27
26.07
26.86
26.39
25.95
25.89
25.98
26.35
25.02
25.63
25.58
24.64
24.96
24.64
26.02
21.9
0.0831
hsa-m
iR-22*
23.08
22.78
23.40
23.33
23.07
22.48
23.64
23.54
23.13
23.15
22.37
23.33
23.32
23.39
23.42
24.53
1.4
0.0845
hsa-m
iR-30c
15.71
14.74
14.86
14.83
14.55
14.76
14.88
15.15
15.57
14.49
14.55
14.92
14.51
14.55
14.17
15.17
21.3
0.0947
hsa-m
iR-129-3p
26.03
26.55
25.28
25.31
26.12
25.96
26.07
25.54
26.79
24.73
24.34
24.95
24.48
25.00
24.19
25.34
21.7
0.095
Thetable
show
sraw
CTandfold
changeforalldifferentially
expressed
(adjusted
p,
0.05,abovethedotted
line)
andmiRNAsthat
show
edatendency
tobestatisticallysignificant(p
,0.1,below
thedotted
line)
afterapplyingmultiple
comparisonscorrection(Benjamini–Hochbergadjusted
pvalues
forfalsediscovery
rate)in
thecolonof12chronic
SIV
–infected
macaques.
The Journal of Immunology 5
by guest on February 7, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

the 21 chronic SIV-infected macaques had mild to moderate en-teritis (Table I). Opportunistic pathogens such as Mycobacteriumavium intracellulare were not detected in 20 of 21 chronic SIV-infected macaques.
Chronic SIV infection of the GI tract/immune system ischaracterized by marked alterations in the expression ofinflammation-associated miRNAs
We performed global miRNA profiling of colon tissue from 12chronically SIV-infected and 4 uninfected control macaques usingthe TLDA cards to determine whether miRNA expression wasaltered in the GI tract/immune system during chronic SIV infection.After applying multiple comparisons correction, 10 miRNAs (3upregulated and 7 downregulated) were found to be statisticallysignificant (adjusted p , 0.05) and differentially expressed fol-lowing analysis using DataAssist software version 3.01 (Table III).Among the three upregulated miRNAs (cards A and B), the
expression of miR-223* and miR-34a was .2-fold, and miR-424showed an ∼1.4-fold increase in colon of chronic SIV-infectedmacaques (Table III). The expression of seven miRNAs de-creased from 1.2- to 1.8-fold (Table III). At least 13 other miRNAsthat comprised three previously well-characterized inflammation-associated miRNAs [let-7b (p = 0.0578), miR-146b-5p (p =0.0719), and miR-21 (p = 0.0825)] showed a tendency to be sta-tistically significant (p , 0.1) (Table III). When multiple com-parisons correction was not applied, 68 (31 upregulated and 37downregulated) miRNAs were found to be differentially expressed(Supplemental Table I). Among the upregulated miRNAs wasmiR-190b (3.2-fold increase), which we recently reported to besignificantly upregulated throughout the course of SIV infectionprimarily in response to viral infection/replication and notimmune/inflammatory response accompanying viral replication(21). These findings show that chronic SIV infection is charac-terized by marked changes in the expression of multipleinflammation-related miRNAs in the colon and that the profile isdominated by downregulated miRNAs.
miR-34a is an inflammation-associated miRNA, and itsexpression is significantly elevated in both colonic epithelialand LPL compartments of chronic SIV-infected macaques
Among the differentially expressed miRNAs, we decided to focuson miR-34a because it is a highly conserved miRNA previouslylinked to inflammation, apoptosis, and cellular senescence/cellcycle arrest (22–24). Further, miR-34a has been directly linkedto “inflammaging,” an aging-related state characterized by sys-temic chronic inflammation proposed to promote premature agingof HIV-infected patients (27). Furthermore, miR-34a has also beenassociated with the induction of cellular senescence in normalcolonic epithelial cells in response to treatment with irinotecan, atopoisomerase I–inhibiting anticancer drug (28). Finally, miR-34awas the most abundant (lowest CT) among the 10 differentiallyexpressed miRNAs whose expression was significantly upregu-lated in the colon of chronic SIV-infected macaques (Table III).Because miR-34a expression was significantly elevated in intact
colons of chronic SIV-infected macaques when using the TLDAcards, we next quantified its expression using a miR-34a–specificqRT-PCR assay separately in the colonic epithelial and LPLcompartments to identify the mucosal compartment contributingto its upregulation. The protocol used for intestinal cell isolationin the present study yields epithelial cells with ∼85–90% puritywith minimal contamination with intraepithelial lymphocytes (29).Similar purity has also been obtained for LPLs with minimumcontamination with epithelial cells (29). The LPLs isolated consist
FIGURE 1. qRT-PCR quantification of miR-34a in colon during
chronic (A and B) and acute (C) SIV infection. In chronically SIV-
infected macaques significant elevation of miR-34a expression was
found in both epithelial (A) and LPL (B) compartments of the colon.
Data were analyzed using a nonparametric Wilcoxon rank sum test.
The error bars represent SE of mean fold change within each group.
**p , 0.01 compared with uninfected controls. In contrast, miR-34a
expression in the colons of rhesus macaques at 7–10, 13–14, 21, and
90 d after SIV infection (C) did not differ from uninfected control
macaques. Data analysis using a nonparametric Kruskal–Wallis test
revealed no differences among the different groups.
6 miR-34a UPREGULATION IN COLON DURING CHRONIC SIV INFECTION
by guest on February 7, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

mostly of lymphocytes (50–60%) but also contain substantialnumbers of plasma cells and macrophages and smaller numbers ofneutrophils, dendritic cells, and eosinophils. Interestingly, miR-34a expression was significantly elevated in both epithelium(∼3.8-fold increase; p = 0.0057) (Fig. 1A) and LPL compartments(∼2.7-fold increase; p = 0.0051) (Fig. 1B).We then sought to determine the timing of miR-34a upregulation
during the course of SIV infection by quantifying miR-34a ex-pression in colon samples collected from 24 additional macaquesat 7–10 (n = 5), 13–14 (n = 5), 21 (n = 7), and 90 (n = 7) dpostinfection. CD4+ T cell status and plasma and intestinal viralloads for all 24 animals have been previously reported (21). Asshown in Fig. 1C, no significant increase in miR-34a expressionwas observed at any of the time points compared with unin-fected controls. This suggests that miR-34a upregulation is tiedmore tightly with inflammation, which is minimal during acuteinfection than viral load, which is very high acutely, particu-larly at 10–14 (peak viral load) d post infection. Additionally,miR-34a fold change in both colonic epithelium and LPL com-partments showed very good positive correlation with tissue viralloads and inflammation scores (Fig. 2). Interestingly, the corre-lation was higher in the LPL (Fig. 2C, 2D) compared with theepithelial compartment (Fig. 2A, 2B). These data suggested thatmiR-34a upregulation had a strong association with inflammation
and that both epithelial (Fig. 1A) and LPL (Fig. 1B) compartmentscontribute to its elevated expression detected in intact colon tissue(Table III).
Overall, upregulation of the inflammation-associated miR-34a inboth epithelial and LPL compartments suggests that it could po-tentially exert distinct pathophysiologic effects in both mucosalcompartments through epigenetic regulation of gene expression.
Colonic crypt epithelial cells of chronic SIV-infected macaquesstrongly express the dsDNA damage response sensor proteingH2A.X
Recent studies show that miR-34a transcription is elevated in re-sponse to irradiation or oxidative stress (inflammation)–inducedDNA damage (30, 31). Consequently, we verified the presence ofdsDNA damage response (DDR) in the colon by determininggH2A.X expression. Both dsDNA breaks and telomere uncappingcan induce a DDR, resulting in the activation and recruitment ofATM/ATR proteins to the site of damage (32), which then phos-phorylates histone H2A.X proteins on Ser139 to facilitate the focalassembly of checkpoint and DNA repair proteins to the DNAdamage site (32). Consistent with these findings, enhanced phos-pho-gH2A.X (Ser139) expression was detected in colonic cryptepithelial cells of chronic SIV-infected macaques (Fig. 3A, 3B), anindicator of active DDR signaling. More strikingly, epithelial cells
FIGURE 2. Correlation of miR-34a expression in colonic epithelial (A and B) and LPL (C and D) compartments with tissue viral loads and inflammation
scores. miR-34a fold change showed positive statistical correlation with tissue viral load and inflammation scores (p, 0.05) in both colonic epithelium and LPLs.
The Journal of Immunology 7
by guest on February 7, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

present along the entire circumference of the crypts showed in-tense nuclear staining for phospho-gH2A.X (open circles inFig. 3A, 3B). In contrast, phospho-gH2A.X staining was eitherextremely weak or completely absent in the crypt epithelial cellsof the control macaque (Fig. 3C). Additionally, elevated phospho-gH2A.X expression was also detected in LPLs (Fig. 3A–C).Interestingly, persistent DDR manifested by phospho-gH2A.X
accumulation at DNA damage foci is also considered an initiatingsignal for cellular senescence/aging (33). In this context, elevatedmiR-34a expression in renal and endothelial cells was welldemonstrated to promote cellular senescence (23, 24). Accord-ingly, we used a combination of phosho-gH2A.X and the prolif-eration marker Ki67 (32) where senescent cells should stainnegative for Ki67 and positive for phospho-gH2A.X. Surprisingly,most of the colonic crypt epithelial cells of chronic SIV-infectedmacaques strongly expressed both phospho-gH2A.X (Fig. 3A, 3B)and Ki67 (Fig. 3D–F), suggesting that these cells, althoughshowing clear evidence of DDR, did not undergo senescence andretained the capacity to proliferate. Additionally, the absence ofcell cycle arrest/cellular senescence was further confirmed basedon the expression pattern of cyclin-dependent kinase inhibitorp21, a cell cycle arrest/cellular senescence–promoting protein thatunlike Ki67 stained differentiated enterocytes near the lumen butnot the crypt epithelial cells (Supplemental Fig. 2). Quantitativeimage analysis showed significantly increased expression ofphospho-gH2A.X in both colonic epithelial (p, 0.0001) and LPL(p = 0.0034) compartments (Fig. 4A). Interestingly, between thetwo mucosal compartments, phospho-gH2A.X expression in theepithelium showed better statistical significance compared withthe LPL compartment (Fig. 4A). Collectively, the increased ex-pression of phospho-gH2A.X expression in colonic crypt epithe-
lial cells of chronic SIV-infected macaques provides compellingevidence of oxidative stress-mediated DDR secondary to inflam-mation and represents a potential mechanism driving miR-34aupregulation (34, 35). Nevertheless, the positive Ki67 stainingsuggests that unlike inflammatory bowel disease (IBD) patients(36), colonic crypt epithelial cells in chronic SIV-infected mac-aques have not been subjected to oxidative stress that is longenough (persistent DDR) to induce changes consistent with cel-lular senescence.
Enhanced miR-34a expression in the colonic mucosa leads tomarked downregulation of SIRT1 mRNA and proteinexpression
To better understand the biological significance of miR-34a up-regulation, we focused on SIRT1, a predicted and validated targetof miR-34a (34). SIRT1 is a NAD+-dependent class III histone/protein deacetylase known to deacetylate transcription factors,enzymes, DNA binding histone proteins, and such, and in doing soregulates critical cellular processes such as inflammation, DNArepair, cell senescence/aging, cellular metabolism, and stress re-sponse (35). Furthermore, decreased expression of SIRT1 hasbeen detected in several chronic inflammatory conditions suchas chronic obstructive pulmonary disease, atherosclerosis, obesity,and dextran sulfate sodium (DSS)–induced colitis (35, 37). Mostimportantly, SIRT1 inhibits cellular senescence/aging and alsofunctions as a negative regulator of inflammation and immuneactivation by deacetylating the p65 subunit of NF-kB at lysine 310(35, 37).The 39 UTR of the rhesus macaque SIRT1 mRNA has two
predicted miR-34a binding sites (38) (Table II). Although humanSIRT1 mRNA has been validated as a direct target of miR-34a
FIGURE 3. Colonic epithelial cells of chronic SIV-infected macaques strongly expressed the DDR sensor protein phospho-gH2A.X. (A and B) White
circles in (A) and (B) demarcate the periphery of a crypt containing multiple phospho-gH2A.X+ nuclei. Arrowheads (A and B) point to phospho-gH2A.X+
crypt epithelial nuclei. In contrast, colonic epithelial cells from the normal uninfected control macaque showed extremely low to undetectable phospho-
gH2A.X expression (C). White circles in (C) demarcate the periphery of four crypts that showed extremely weak to no phospho-gH2A.X expression in the
nuclei. Long thin arrows (C) point to phospho-gH2A.X+ LPLs. Most of the colonic epithelial cells from both SIV-infected (D and E) and control macaques
(F) also stained positive for Ki67, a cell proliferation marker suggesting that these cells did not become senescent. Epithelial and LPL phospho-gH2A.X+
(A–C) staining cells are shown in green (Alexa Fluor 488). Epithelial and LPL Ki67+ (D–F) staining cells are shown in red (Alexa Fluor 568). (D)–(F)
involve double labels with gH2A.X (green) and Ki67 (red). The individual channels (green for phospho-gH2A.X and red for Ki67) and gray for differential
interference contrast to reveal tissue architecture appear on the left with a larger merged image on the right (D–F). Colocalization of green (phospho-gH2A.X)
and red (Ki67) appear light yellow. Original magnification 340 (all panels).
8 miR-34a UPREGULATION IN COLON DURING CHRONIC SIV INFECTION
by guest on February 7, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
(34), it is not known whether mml-miR-34a can bind the 39 UTRof rhesus macaque SIRT1 mRNA and repress its expression. Be-cause mRNA destabilization has been proposed as a predominantmechanism by which miRNAs repress gene expression (39), wenext quantified SIRT1 mRNA separately in colonic epithelium andLPLs. Interestingly, SIRT1 mRNA expression significantly de-creased by ∼2.4- and ∼1.8-fold in colonic epithelium (p = 0.0118)and LPLs (p = 0.015), respectively (Fig. 4B, 4C). Furthermore,cotransfection of SIRT1-WT UTR vector with miR-34a mimicresulted in significant reduction (∼30%) in firefly/renilla ratios(p , 0.05)). In contrast, cotransfection of SIRT1-Del UTR vectorwith miR-34a restored the firefly/renilla ratios to the level ob-served with unmanipulated pmirGLO vector (Fig. 4D). Interest-ingly, cotransfection of SIRT1-WT UTR vector with a negativecontrol mimic did not reduce firefly/renilla ratios compared withwells transfected with miR-34a mimic (p, 0.05) (Fig. 4D). These
results clearly show that mml-miR-34a can physically interactwith the 39 UTR of rhesus macaque SIRT1 mRNA and regulate itsexpression.We next performed IF on formalin-fixed paraffin-embedded
colon sections using anti-SIRT1–specific Abs to check whetherchanges in mRNA expression were paralleled at the protein level.After evaluating five anti-human SIRT1-specific Abs, one cross-reacted with the rhesus macaque for IF studies. As shown inFig. 5, SIRT1 protein expression was detected in the colonic ep-ithelium and LPLs of chronic SIV-infected (Fig. 5A, 5B) andcontrol macaques (Fig. 5D, 5E). Furthermore, SIRT1 protein lo-calized to both cytoplasm and nuclei of epithelial and LPLs(Fig. 5C). Consistent with decreased SIRT1 mRNA expression,image analysis confirmed significantly (p , 0.05) reduced SIRT1protein expression in the epithelial and LPL compartments of bothchronic SIV-infected compared with uninfected control macaques
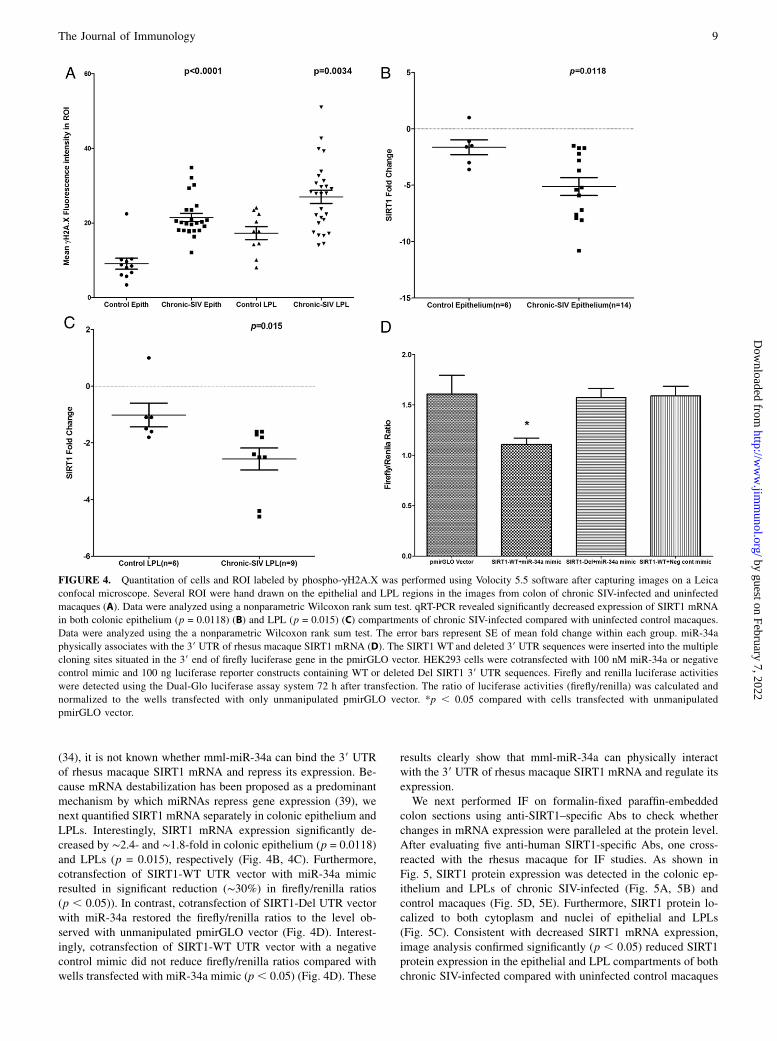
FIGURE 4. Quantitation of cells and ROI labeled by phospho-gH2A.X was performed using Volocity 5.5 software after capturing images on a Leica
confocal microscope. Several ROI were hand drawn on the epithelial and LPL regions in the images from colon of chronic SIV-infected and uninfected
macaques (A). Data were analyzed using a nonparametric Wilcoxon rank sum test. qRT-PCR revealed significantly decreased expression of SIRT1 mRNA
in both colonic epithelium (p = 0.0118) (B) and LPL (p = 0.015) (C) compartments of chronic SIV-infected compared with uninfected control macaques.
Data were analyzed using the a nonparametric Wilcoxon rank sum test. The error bars represent SE of mean fold change within each group. miR-34a
physically associates with the 39 UTR of rhesus macaque SIRT1 mRNA (D). The SIRT1 WT and deleted 39 UTR sequences were inserted into the multiple
cloning sites situated in the 39 end of firefly luciferase gene in the pmirGLO vector. HEK293 cells were cotransfected with 100 nM miR-34a or negative
control mimic and 100 ng luciferase reporter constructs containing WT or deleted Del SIRT1 39 UTR sequences. Firefly and renilla luciferase activities
were detected using the Dual-Glo luciferase assay system 72 h after transfection. The ratio of luciferase activities (firefly/renilla) was calculated and
normalized to the wells transfected with only unmanipulated pmirGLO vector. *p , 0.05 compared with cells transfected with unmanipulated
pmirGLO vector.
The Journal of Immunology 9
by guest on February 7, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
(Fig. 5F). Based on the results of the luciferase assay (Fig. 3C),decreased SIRT1 mRNA and protein expression in the colonicepithelium and LPL may be directly attributed to miR-34a bindingand inhibition of target gene expression.
miR-34a–mediated downregulation of SIRT1 was associatedwith constitutive expression of acetylated p65 (lysine 310) inlamina propria plasma cells
Because SIRT1 is known to deacetylate the p65 subunit of NF-kBon lysine 310 (35), we hypothesized that miR-34a–mediateddownregulation of SIRT1 will result in constitutive expression ofthe transcriptionally active form of p65 (acetylated on lysine 310)in the colon. In agreement with our hypothesis, IF studies revealedintense expression of acetylated p65 exclusively in the LPLcompartment of chronic SIV-infected macaques (Fig. 6A, 6B).The inset image in Fig. 6A and 6B shows a magnified view ofacetylated p65+ cells in the colonic lamina propria. This Abdetects p65 only when acetylated on lysine 310 and has been usedpreviously for immunohistochemistry studies (40). Additionally,the Ab recognizing the nonacetylated peptide has been removedby chromatography using nonacetylated peptide sequences. Incontrast to chronic SIV-infected macaques, occasional acetylatedp65 (lysine 310)+ cells were detected in the colonic lamina propriaof the control macaque (Fig. 6C). We next immunophenotypedacetyl p65+ cells using markers specific to macrophages (CD68,
CD163) and T lymphocytes (CD3). Surprisingly, none of themarkers colocalized with acetylated p65+, suggesting that neithermacrophages nor T lymphocytes were contributing to the elevatedacetylated p65 expression (Fig. 6D, 6F). Finally, using anti-rhesusIgA and IgG Abs we successfully identified Ab-producing plasmacells as the primary cell type contributing to enhanced acetylatedp65 (lysine 310) expression (Fig. 7A, 7B). IgA+/acetylated p65+
and IgG+/acetylated p65+ cells were rare in the control macaque(Fig. 7C, 7D). Furthermore, image analysis revealed significantlyincreased numbers of IgA+/acetylated p65+ and IgG+/acetylatedp65+ cells in the colonic lamina propria of chronic SIV-infectedmacaques compared with uninfected control macaques (Fig. 8A–D). Image analysis provided additional evidence that plasma cellsin the colonic lamina propria of chronic SIV-infected macaquescarry a higher percentage of IgG+/acetylated p65+ cells comparedwith control macaques (Fig. 8B, 8D). In contrast, there werevery few IgG+/acetylated p65+ cells in the three control mac-aques (Fig. 8B, 8D). Similar to the colon, IgA+/acetylated p65+
cells were also detected in the jejunum but only in macaquesthat showed histological signs of enteritis (HD08) (Table I)(data not shown). IgA+/acetylated p65+ cells were extremelyrare in the jejunum of a chronically SIV-infected macaque(GA19) that did not show histological evidence of enteritis(Table I) (data not shown). Although we could not detectacetylated p65 in T cells and macrophages, it is possible that the
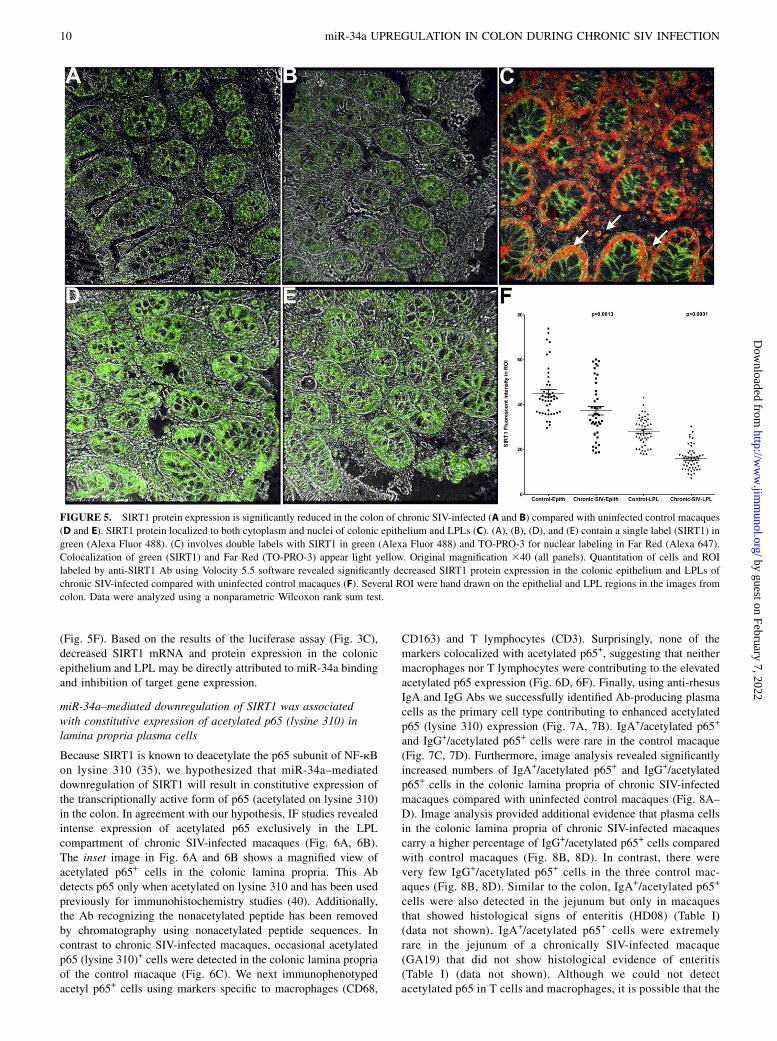
FIGURE 5. SIRT1 protein expression is significantly reduced in the colon of chronic SIV-infected (A and B) compared with uninfected control macaques
(D and E). SIRT1 protein localized to both cytoplasm and nuclei of colonic epithelium and LPLs (C). (A), (B), (D), and (E) contain a single label (SIRT1) in
green (Alexa Fluor 488). (C) involves double labels with SIRT1 in green (Alexa Fluor 488) and TO-PRO-3 for nuclear labeling in Far Red (Alexa 647).
Colocalization of green (SIRT1) and Far Red (TO-PRO-3) appear light yellow. Original magnification 340 (all panels). Quantitation of cells and ROI
labeled by anti-SIRT1 Ab using Volocity 5.5 software revealed significantly decreased SIRT1 protein expression in the colonic epithelium and LPLs of
chronic SIV-infected compared with uninfected control macaques (F). Several ROI were hand drawn on the epithelial and LPL regions in the images from
colon. Data were analyzed using a nonparametric Wilcoxon rank sum test.
10 miR-34a UPREGULATION IN COLON DURING CHRONIC SIV INFECTION
by guest on February 7, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

transcriptionally active protein could be present at levels belowthe detection limit of IF.
In situ hybridization detects strong miR-34a expression in thecolonic epithelium and lamina propria plasma cells
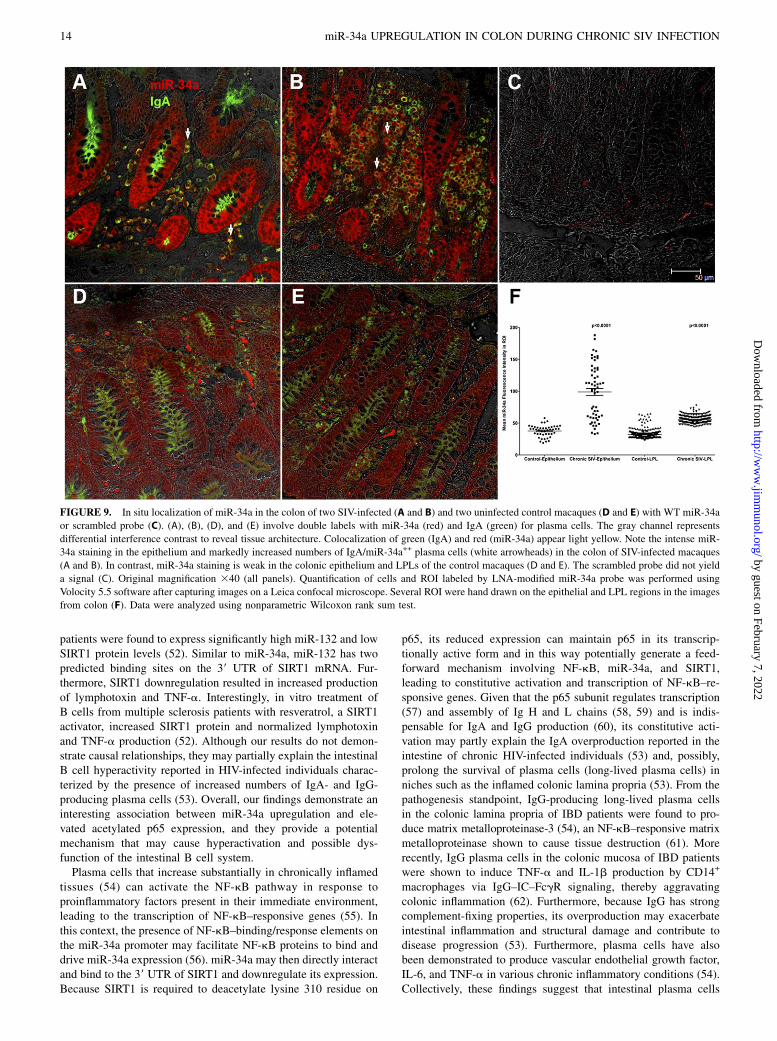
Because acetylated p65 expression was detected only in plasmacells, we next determined whether these cells also expressedmiR-34a to directly link expression of constitutive acetylatedp65 to elevated miR-34a expression. As shown in Fig. 9A and 9B,after 7 min incubation with permanent red substrate (Dako),miR-34a expression was strongly detected in colonic epitheliumand LPLs of both chronic SIV-infected macaques. In contrast,miR-34a staining was very weak in uninfected control macaques(Fig. 9D, 9E) examined in the same experiment. Furthermore, thesection stained with the scrambled probe did not yield any signal(Fig. 9C). All images were captured using a Leica confocal mi-croscope employing the same laser strength (red and green). Thesefindings are strongly supported by quantitative image analysisshowing significantly elevated miR-34a signal intensity in colonicepithelial and LPL compartments of chronic SIV-infected mac-aques compared with controls (Fig. 9F). Double labeling withmiR-34a and acetylated p65 was attempted without success, as notall Abs work well on tissue sections previously subjected to in situhybridization. The in situ hybridization results were in perfectagreement with the qRT-PCR data showing elevated miR-34aexpression in colonic epithelium and LPLs of chronic SIV-infected macaques (Fig. 1A, 1B). Collectively, the intense miR-34a expression in IgA+ plasma cells (Fig. 9A, 9B) suggests thatenhanced miR-34a expression may promote constitutive activationof acetylated p65 (lysine 310), possibly through SIRT1 down-regulation.
DiscussionGI inflammation, a major manifestation of HIV/SIV infection thatis necessary for pathogen clearance, can, however, result instructural and functional damage when not properly regulated thatcan promote microbial translocation and systemic immune acti-vation (1–4). In addition to proinflammatory cytokines and tran-scription factors, recent studies have proposed critical roles formiRNAs in controlling and managing certain aspects of the in-flammatory process (41). Moreover, the recent finding that miR-NAs primarily regulate gene expression by destabilizing anddegrading mRNA targets (39) suggests that the previously re-ported alterations in intestinal gene expression profiles by us (13,14) and others (11, 12) could be directly attributed to the regu-latory functions of miRNAs. Therefore, the next logical step is toprofile, identify, and characterize the specific miRNAs that exertthese regulatory effects. To this end, we have reported miRNAdysregulation in colon during acute SIV infection (21). Morespecifically, we identified miR-190b to be significantly upregu-lated in both colon and jejunum at all stages of SIV infectionprimarily in response to viral replication. In the present study, weagain focused on the colon, as it was found to be more severelyimpacted than the jejunum/small intestine during chronic SIVinfection (Table I). Furthermore, and more importantly, theincreased bacterial concentration in the colon (1012 bacterialorganisms/ml contents) (42) makes it a major source of bacteria/microbes contributing to microbial translocation during chronicHIV/SIV infection. In the present study, we report the identifica-tion of miR-34a, a miRNA previously linked to chronic inflam-mation, cell cycle arrest, and senescence (22–24, 27, 28), to besignificantly dysregulated in both colonic epithelial and LPLcompartments. Furthermore, our findings suggest that the miR-
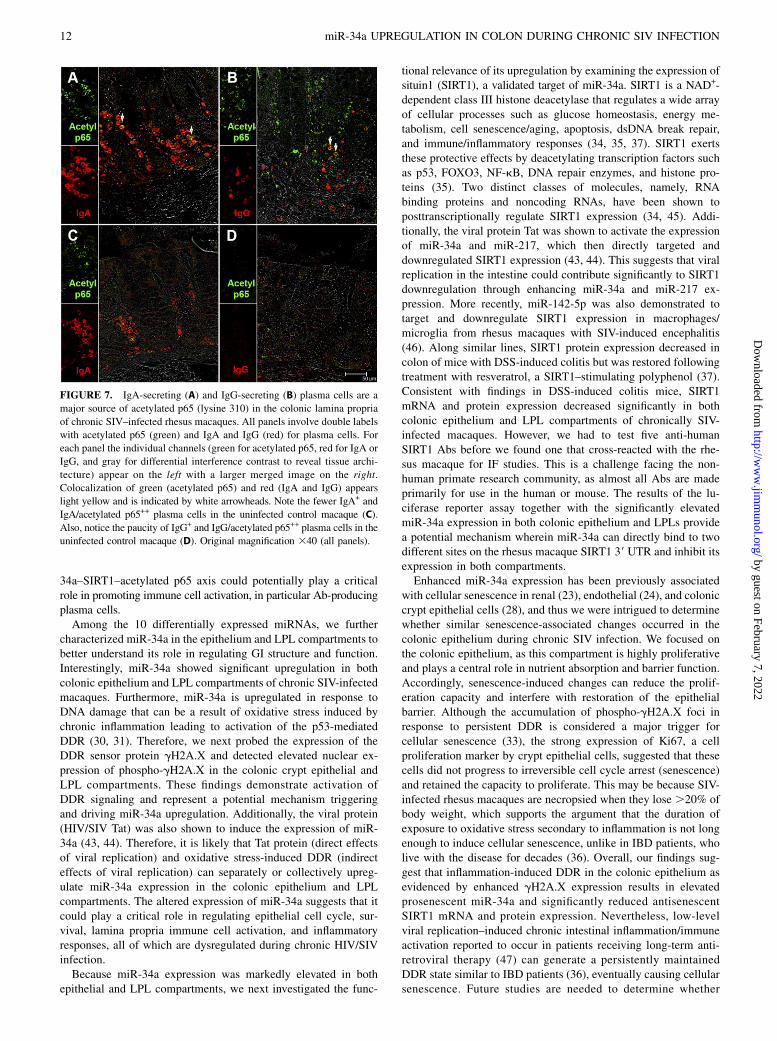
FIGURE 6. Presence of increased numbers of acetylated p65 (lysine 310)–expressing cells in the colonic lamina propria of two chronic SIV–infected
rhesus macaques (A and B). The inset image shows a close-up view of acetylated p65+ cells in the colonic lamina propria. Note the fewer acetylated p65+
cells in the uninfected control macaque (C). Acetylated p65+ staining cells are shown in green (Alexa Fluor 488). Lymphocytes [CD3 (D)] and macrophages
[CD68 (E), CD163 (F)] are not the predominant cellular source of acetylated p65 (lysine 310). All panels involve double labels with acetylated p65 in green
and CD3 for lymphocytes and CD68/CD163 for macrophages in red. For each panel the individual channels (green for acetylated p65 and red for CD3,
CD68, and CD163) and gray for differential interference contrast to reveal tissue architecture appear on the left with a larger merged image on the right.
Note that acetylatedp65+ cells do not colocalize with CD3, CD68, and CD163. Original magnification 340 (all panels).
The Journal of Immunology 11
by guest on February 7, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
34a–SIRT1–acetylated p65 axis could potentially play a criticalrole in promoting immune cell activation, in particular Ab-producingplasma cells.Among the 10 differentially expressed miRNAs, we further
characterized miR-34a in the epithelium and LPL compartments tobetter understand its role in regulating GI structure and function.Interestingly, miR-34a showed significant upregulation in bothcolonic epithelium and LPL compartments of chronic SIV-infectedmacaques. Furthermore, miR-34a is upregulated in response toDNA damage that can be a result of oxidative stress induced bychronic inflammation leading to activation of the p53-mediatedDDR (30, 31). Therefore, we next probed the expression of theDDR sensor protein gH2A.X and detected elevated nuclear ex-pression of phospho-gH2A.X in the colonic crypt epithelial andLPL compartments. These findings demonstrate activation ofDDR signaling and represent a potential mechanism triggeringand driving miR-34a upregulation. Additionally, the viral protein(HIV/SIV Tat) was also shown to induce the expression of miR-34a (43, 44). Therefore, it is likely that Tat protein (direct effectsof viral replication) and oxidative stress-induced DDR (indirecteffects of viral replication) can separately or collectively upreg-ulate miR-34a expression in the colonic epithelium and LPLcompartments. The altered expression of miR-34a suggests that itcould play a critical role in regulating epithelial cell cycle, sur-vival, lamina propria immune cell activation, and inflammatoryresponses, all of which are dysregulated during chronic HIV/SIVinfection.Because miR-34a expression was markedly elevated in both
epithelial and LPL compartments, we next investigated the func-
tional relevance of its upregulation by examining the expression ofsituin1 (SIRT1), a validated target of miR-34a. SIRT1 is a NAD+-dependent class III histone deacetylase that regulates a wide arrayof cellular processes such as glucose homeostasis, energy me-tabolism, cell senescence/aging, apoptosis, dsDNA break repair,and immune/inflammatory responses (34, 35, 37). SIRT1 exertsthese protective effects by deacetylating transcription factors suchas p53, FOXO3, NF-kB, DNA repair enzymes, and histone pro-teins (35). Two distinct classes of molecules, namely, RNAbinding proteins and noncoding RNAs, have been shown toposttranscriptionally regulate SIRT1 expression (34, 45). Addi-tionally, the viral protein Tat was shown to activate the expressionof miR-34a and miR-217, which then directly targeted anddownregulated SIRT1 expression (43, 44). This suggests that viralreplication in the intestine could contribute significantly to SIRT1downregulation through enhancing miR-34a and miR-217 ex-pression. More recently, miR-142-5p was also demonstrated totarget and downregulate SIRT1 expression in macrophages/microglia from rhesus macaques with SIV-induced encephalitis(46). Along similar lines, SIRT1 protein expression decreased incolon of mice with DSS-induced colitis but was restored followingtreatment with resveratrol, a SIRT1–stimulating polyphenol (37).Consistent with findings in DSS-induced colitis mice, SIRT1mRNA and protein expression decreased significantly in bothcolonic epithelium and LPL compartments of chronically SIV-infected macaques. However, we had to test five anti-humanSIRT1 Abs before we found one that cross-reacted with the rhe-sus macaque for IF studies. This is a challenge facing the non-human primate research community, as almost all Abs are madeprimarily for use in the human or mouse. The results of the lu-ciferase reporter assay together with the significantly elevatedmiR-34a expression in both colonic epithelium and LPLs providea potential mechanism wherein miR-34a can directly bind to twodifferent sites on the rhesus macaque SIRT1 39 UTR and inhibit itsexpression in both compartments.Enhanced miR-34a expression has been previously associated
with cellular senescence in renal (23), endothelial (24), and coloniccrypt epithelial cells (28), and thus we were intrigued to determinewhether similar senescence-associated changes occurred in thecolonic epithelium during chronic SIV infection. We focused onthe colonic epithelium, as this compartment is highly proliferativeand plays a central role in nutrient absorption and barrier function.Accordingly, senescence-induced changes can reduce the prolif-eration capacity and interfere with restoration of the epithelialbarrier. Although the accumulation of phospho-gH2A.X foci inresponse to persistent DDR is considered a major trigger forcellular senescence (33), the strong expression of Ki67, a cellproliferation marker by crypt epithelial cells, suggested that thesecells did not progress to irreversible cell cycle arrest (senescence)and retained the capacity to proliferate. This may be because SIV-infected rhesus macaques are necropsied when they lose .20% ofbody weight, which supports the argument that the duration ofexposure to oxidative stress secondary to inflammation is not longenough to induce cellular senescence, unlike in IBD patients, wholive with the disease for decades (36). Overall, our findings sug-gest that inflammation-induced DDR in the colonic epithelium asevidenced by enhanced gH2A.X expression results in elevatedprosenescent miR-34a and significantly reduced antisenescentSIRT1 mRNA and protein expression. Nevertheless, low-levelviral replication–induced chronic intestinal inflammation/immuneactivation reported to occur in patients receiving long-term anti-retroviral therapy (47) can generate a persistently maintainedDDR state similar to IBD patients (36), eventually causing cellularsenescence. Future studies are needed to determine whether
FIGURE 7. IgA-secreting (A) and IgG-secreting (B) plasma cells are a
major source of acetylated p65 (lysine 310) in the colonic lamina propria
of chronic SIV–infected rhesus macaques. All panels involve double labels
with acetylated p65 (green) and IgA and IgG (red) for plasma cells. For
each panel the individual channels (green for acetylated p65, red for IgA or
IgG, and gray for differential interference contrast to reveal tissue archi-
tecture) appear on the left with a larger merged image on the right.
Colocalization of green (acetylated p65) and red (IgA and IgG) appears
light yellow and is indicated by white arrowheads. Note the fewer IgA+ and
IgA/acetylated p65++ plasma cells in the uninfected control macaque (C).
Also, notice the paucity of IgG+ and IgG/acetylated p65++ plasma cells in the
uninfected control macaque (D). Original magnification 340 (all panels).
12 miR-34a UPREGULATION IN COLON DURING CHRONIC SIV INFECTION
by guest on February 7, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

senescence-associated changes occur in long term antiretroviraltherapy–treated individuals, as they can cause accelerated agingof the intestine, leading to epithelial barrier dysfunction, microbialtranslocation, and AIDS progression.In the LPL compartment, miR-34a expression increased and
SIRT1 expression decreased significantly. Because the LPL com-partment is enriched for immune cells and primarily involved inmediating immune/inflammatory responses, we focused on theRelA/p65 subunit of NF-kB, a transcription factor central to im-mune and inflammatory responses (48). Most notably, posttrans-lational modifications such as phosphorylation, acetylation, andSUMOylation are necessary for modulating NF-kB–mediatedgene expression (48). Among the different modifications, acety-lation of the p65 subunit catalyzed by p300/CBP and p300/CBP-associated factor can occur on seven distinct lysine residues (ly-sine 122, 123, 218, 221, 310, 314, and 315) that can either activateor inhibit NF-kB signaling depending on which lysine residue isacetylated (48). Among these, acetylation of lysine 310 is ofsignificant interest because it is crucial for full activation of NF-kB transcriptional potential (48). Equally important is the findingthat SIRT1 (48) and HDAC3 (48) are two major enzymes that candeacetylate lysine 310 on p65, thereby inhibiting its transcrip-tional activity. Additionally, Tat-mediated miR-34a upregulationinhibited SIRT1 expression in TZM-bl cells, resulting in enhancedacetylated p65 (lysine 310) expression (43). Furthermore, Tat hasalso been shown to directly bind to the deacetylase domain ofSIRT1 and suppress its ability to deacetylate lysine 310 on the p65subunit of NF-kB, resulting in enhanced acetylated p65 activity
(49). Therefore, we probed the downstream effects of miR-34a–mediated SIRT1 downregulation in the LPL compartment by de-termining the expression of acetylated p65 (lysine 310). Using IFstudies, acetylated p65 (lysine 310) expression was found to bemarkedly increased solely in the LPL compartment of chronicSIV-infected macaques. Interestingly, acetylated p65+ cells werenegative for CD68, CD163, and CD3. Subsequently, using anti–rhesus macaque IgG and IgA Abs, we successfully localizedacetylated p65 (lysine 310) to plasma cells. Furthermore, mark-edly increased numbers of IgA+/acetyl p65+ and IgG+/acetyl p65+
cells were detected in the colons of chronic SIV-infected mac-aques. These findings were not surprising, as DDR signalingevidenced by strong phospho-gH2A.X in other lamina propriacells such as lymphocytes and macrophages also activates theJAK-STAT3 pathway for efficient dsDNA repair (50). We previ-ously demonstrated constitutive activation of the phosphorylatedform of STAT3 (tyrosine 705) in CD3+ lymphocytes and CD68+
macrophages (10). These results, together with our previouslypublished findings (10), suggest that whereas plasma cells activatep65 through acetylation of lysine 310, lymphocytes and macro-phages can constitutively activate STAT3 by phosphorylationof tyrosine 705 in response to oxidative stress–induced DDR.Moreover, using in situ hybridization we successfully localizedmiR-34a expression to the colonic epithelium and plasma cells inthe lamina propria. These interesting findings are consistent witha previous study that identified miR-34a to be significantly up-regulated following B cell activation in vitro with 1 mM CpG (51).More recently, activated B cells isolated from multiple sclerosis
FIGURE 8. Quantification of the number of IgA/acetylated p65++ (A) and IgG/acetylated p65++ (B) in the lamina propria of three chronic SIV–infected
and three control rhesus macaques. Positive cells were counted at an original magnification of340 in 10 high-power fields (HPF) (one HPF = 0.2827 mm2).
Each datum point represents the number of cells in a single HPF; the error bars represent the mean values. Average IgA/acetylated p65++ (C) and IgG/
acetylated p65++ (D) in the colonic lamina propria of three SIV-infected and three uninfected control macaques. All slides were analyzed by two inves-
tigators in a blinded manner. Data were analyzed using a Wilcoxon rank sum test. ***p , 0.0001 compared with uninfected controls.
The Journal of Immunology 13
by guest on February 7, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
patients were found to express significantly high miR-132 and lowSIRT1 protein levels (52). Similar to miR-34a, miR-132 has twopredicted binding sites on the 39 UTR of SIRT1 mRNA. Fur-thermore, SIRT1 downregulation resulted in increased productionof lymphotoxin and TNF-a. Interestingly, in vitro treatment ofB cells from multiple sclerosis patients with resveratrol, a SIRT1activator, increased SIRT1 protein and normalized lymphotoxinand TNF-a production (52). Although our results do not demon-strate causal relationships, they may partially explain the intestinalB cell hyperactivity reported in HIV-infected individuals charac-terized by the presence of increased numbers of IgA- and IgG-producing plasma cells (53). Overall, our findings demonstrate aninteresting association between miR-34a upregulation and ele-vated acetylated p65 expression, and they provide a potentialmechanism that may cause hyperactivation and possible dys-function of the intestinal B cell system.Plasma cells that increase substantially in chronically inflamed
tissues (54) can activate the NF-kB pathway in response toproinflammatory factors present in their immediate environment,leading to the transcription of NF-kB–responsive genes (55). Inthis context, the presence of NF-kB–binding/response elements onthe miR-34a promoter may facilitate NF-kB proteins to bind anddrive miR-34a expression (56). miR-34a may then directly interactand bind to the 39 UTR of SIRT1 and downregulate its expression.Because SIRT1 is required to deacetylate lysine 310 residue on
p65, its reduced expression can maintain p65 in its transcrip-tionally active form and in this way potentially generate a feed-forward mechanism involving NF-kB, miR-34a, and SIRT1,leading to constitutive activation and transcription of NF-kB–re-sponsive genes. Given that the p65 subunit regulates transcription(57) and assembly of Ig H and L chains (58, 59) and is indis-pensable for IgA and IgG production (60), its constitutive acti-vation may partly explain the IgA overproduction reported in theintestine of chronic HIV-infected individuals (53) and, possibly,prolong the survival of plasma cells (long-lived plasma cells) inniches such as the inflamed colonic lamina propria (53). From thepathogenesis standpoint, IgG-producing long-lived plasma cellsin the colonic lamina propria of IBD patients were found to pro-duce matrix metalloproteinase-3 (54), an NF-kB–responsive matrixmetalloproteinase shown to cause tissue destruction (61). Morerecently, IgG plasma cells in the colonic mucosa of IBD patientswere shown to induce TNF-a and IL-1b production by CD14+
macrophages via IgG–IC–FcgR signaling, thereby aggravatingcolonic inflammation (62). Furthermore, because IgG has strongcomplement-fixing properties, its overproduction may exacerbateintestinal inflammation and structural damage and contribute todisease progression (53). Furthermore, plasma cells have alsobeen demonstrated to produce vascular endothelial growth factor,IL-6, and TNF-a in various chronic inflammatory conditions (54).Collectively, these findings suggest that intestinal plasma cells
FIGURE 9. In situ localization of miR-34a in the colon of two SIV-infected (A and B) and two uninfected control macaques (D and E) with WT miR-34a
or scrambled probe (C). (A), (B), (D), and (E) involve double labels with miR-34a (red) and IgA (green) for plasma cells. The gray channel represents
differential interference contrast to reveal tissue architecture. Colocalization of green (IgA) and red (miR-34a) appear light yellow. Note the intense miR-
34a staining in the epithelium and markedly increased numbers of IgA/miR-34a++ plasma cells (white arrowheads) in the colon of SIV-infected macaques
(A and B). In contrast, miR-34a staining is weak in the colonic epithelium and LPLs of the control macaques (D and E). The scrambled probe did not yield
a signal (C). Original magnification 340 (all panels). Quantification of cells and ROI labeled by LNA-modified miR-34a probe was performed using
Volocity 5.5 software after capturing images on a Leica confocal microscope. Several ROI were hand drawn on the epithelial and LPL regions in the images
from colon (F). Data were analyzed using nonparametric Wilcoxon rank sum test.
14 miR-34a UPREGULATION IN COLON DURING CHRONIC SIV INFECTION
by guest on February 7, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

may have broader functions in modulating inflammatory respon-ses that can potentially contribute to epithelial barrier disruption,and the miR-34a–SIRT1-mediated constitutive NF-kB activationmay considerably assist in these functions.In summary, our results demonstrate marked miRNA dysregu-
lation in the intestine during chronic SIV infection. Whereas miR-34a was significantly upregulated in both colonic epithelium andLPL compartments, SIRT1 mRNA and protein expression sig-nificantly decreased in both compartments. Elevated miR-34a andreduced SIRT1 expression were associated with constitutive ex-pression of acetylated p65 in colonic IgA+ and IgG+ plasma cells.The findings suggest a possible role for the miR-34a–SIRT1–p65axis in causing hyperactivity and dysfunction of the mucosalB cell system. The recent demonstration that miR-34a expres-sion is elevated following in vitro B cell activation using CpGmolecules (51) and its ability to regulate B cell development/differentiation by modulating Foxp1 expression (63) strengthensthe possibility. Apart from SIRT1, miR-34a has also been shownto regulate the expression of several cell cycle regulators (NMYC,CCND1, CDK4, CDK6, MET) (22), and future studies involvingcell sorting protocols and high-throughput proteomics are neededto identify and validate additional targets (∼640 predicted) tounderstand the full functional significance of its upregulation inthe colon during HIV/SIV infection. Additionally, in vivo miR-34amodulation studies are needed to determine whether a causal re-lationship exists between miR-34a upegulation and constitutiveacetylated p65 expression. Nevertheless, importantly note thatdirect in vivo miRNA inhibition using antagomirs is currently notfeasible owing to lack of an efficient in vivo delivery system thatcan selectively target specific segments of the intestine. However,the recent demonstration that a 12 mo supplementation of resver-atrol, an SIRT1-activating anti-inflammatory polyphenol present ingrape extracts, modulated miR-34a expression in PBMCs of type 2diabetes and hypertensive patients (64) is very encouraging, as ithighlights an alternative approach preferably in combination withantiretroviral drugs to modulate miR-34a expression and chronicpersistent inflammation.
AcknowledgmentsWe thank Ronald S. Veazey, Maurice Duplantis, Yun Te Lin, Faith
R. Schiro, Cecily C. Midkiff, Christopher Monjure, and Coty Tatum for
technical assistance.
DisclosuresThe authors have no financial conflicts of interest.
References1. Veazey, R. S., M. DeMaria, L. V. Chalifoux, D. E. Shvetz, D. R. Pauley,
H. L. Knight, M. Rosenzweig, R. P. Johnson, R. C. Desrosiers, andA. A. Lackner. 1998. Gastrointestinal tract as a major site of CD4+ T cell de-pletion and viral replication in SIV infection. Science 280: 427–431.
2. Smit-McBride, Z., J. J. Mattapallil, M. McChesney, D. Ferrick, and S. Dandekar.1998. Gastrointestinal T lymphocytes retain high potential for cytokine responsesbut have severe CD4+ T-cell depletion at all stages of simian immunodeficiencyvirus infection compared to peripheral lymphocytes. J. Virol. 72: 6646–6656.
3. Mattapallil, J. J., D. C. Douek, B. Hill, Y. Nishimura, M. Martin, andM. Roederer. 2005. Massive infection and loss of memory CD4+ T cells inmultiple tissues during acute SIV infection. Nature 434: 1093–1097.
4. Brenchley, J. M., T. W. Schacker, L. E. Ruff, D. A. Price, J. H. Taylor,G. J. Beilman, P. L. Nguyen, A. Khoruts, M. Larson, A. T. Haase, andD. C. Douek. 2004. CD4+ T cell depletion during all stages of HIV diseaseoccurs predominantly in the gastrointestinal tract. J. Exp. Med. 200: 749–759.
5. Mehandru, S., M. A. Poles, K. Tenner-Racz, A. Horowitz, A. Hurley, C. Hogan,D. Boden, P. Racz, and M. Markowitz. 2004. Primary HIV-1 infection is asso-ciated with preferential depletion of CD4+ T lymphocytes from effector sites inthe gastrointestinal tract. J. Exp. Med. 200: 761–770.
6. MacArthur, R. D., and H. L. DuPont. 2012. Etiology and pharmacologic man-agement of noninfectious diarrhea in HIV-infected individuals in the highlyactive antiretroviral therapy era. Clin. Infect. Dis. 55: 860–867.
7. Klatt, N. R., N. T. Funderburg, and J. M. Brenchley. 2013. Microbial translo-cation, immune activation, and HIV disease. Trends Microbiol. 21: 6–13.
8. Marchetti, G., C. Tincati, and G. Silvestri. 2013. Microbial translocation in thepathogenesis of HIV infection and AIDS. Clin. Microbiol. Rev. 26: 2–18.
9. McGowan, I., J. Elliott, M. Fuerst, P. Taing, J. Boscardin, M. Poles, and P. Anton.2004. Increased HIV-1 mucosal replication is associated with generalized mu-cosal cytokine activation. J. Acquir. Immune Defic. Syndr. 37: 1228–1236.
10. Mohan, M., P. P. Aye, J. T. Borda, X. Alvarez, and A. A. Lackner. 2007. Gas-trointestinal disease in simian immunodeficiency virus-infected rhesus macaquesis characterized by proinflammatory dysregulation of the interleukin-6-Januskinase/signal transducer and activator of transcription3 pathway. Am. J.Pathol. 171: 1952–1965.
11. George, M. D., S. Sankaran, E. Reay, A. C. Gelli, and S. Dandekar. 2003. High-throughput gene expression profiling indicates dysregulation of intestinal cellcycle mediators and growth factors during primary simian immunodeficiencyvirus infection. Virology 312: 84–94.
12. Sankaran, S., M. D. George, E. Reay, M. Guadalupe, J. Flamm, T. Prindiville,and S. Dandekar. 2008. Rapid onset of intestinal epithelial barrier dysfunction inprimary human immunodeficiency virus infection is driven by an imbalancebetween immune response and mucosal repair and regeneration. J. Virol. 82:538–545.
13. Mohan, M., D. Kaushal, P. P. Aye, X. Alvarez, R. S. Veazey, and A. A. Lackner.2012. Focused examination of the intestinal lamina propria yields greater mo-lecular insight into mechanisms underlying SIV induced immune dysfunction.PLoS ONE 7: e34561.
14. Mohan, M., D. Kaushal, P. P. Aye, X. Alvarez, R. S. Veazey, and A. A. Lackner.2013. Focused examination of the intestinal epithelium reveals transcriptionalsignatures consistent with disturbances in enterocyte maturation and differenti-ation during the course of SIV infection. PLoS ONE 8: e60122.
15. Ambros, V. 2004. The functions of animal microRNAs. Nature 431: 350–355.16. Swaminathan, S., D. D. Murray, and A. D. Kelleher. 2013. miRNAs and HIV:
unforeseen determinants of host-pathogen interaction. Immunol. Rev. 254: 265–280.
17. Triboulet, R., B. Mari, Y. L. Lin, C. Chable-Bessia, Y. Bennasser, K. Lebrigand,B. Cardinaud, T. Maurin, P. Barbry, V. Baillat, et al. 2007. Suppression ofmicroRNA-silencing pathway by HIV-1 during virus replication. Science 315:1579–1582.
18. Witwer, K. W., S. L. Sarbanes, J. Liu, and J. E. Clements. 2011. A plasmamicroRNA signature of acute lentiviral infection: biomarkers of central nervoussystem disease. AIDS 25: 2057–2067.
19. Yelamanchili, S. V., A. D. Chaudhuri, L. N. Chen, H. Xiong, and H. S. Fox.2010. MicroRNA-21 dysregulates the expression of MEF2C in neurons inmonkey and human SIV/HIV neurological disease. Cell Death Dis. 1: e77.
20. Sisk, J. M., K. W. Witwer, P. M. Tarwater, and J. E. Clements. 2013. SIV rep-lication is directly downregulated by four antiviral miRNAs. Retrovirology 10:95.
21. Mohan, M., L. C. Chandra, W. Torben, P. P. Aye, X. Alvarez, and A. A. Lackner.2014. miR-190b is markedly upregulated in the intestine in response to SIVreplication and partly regulates myotubularin related protein-6 expression. J.Immunol. 193: 1301–1313.
22. Chen, F., and S. J. Hu. 2012. Effect of microRNA-34a in cell cycle, differenti-ation, and apoptosis: a review. J. Biochem. Mol. Toxicol. 26: 79–86.
23. Bai, X. Y., Y. Ma, R. Ding, B. Fu, S. Shi, and X. M. Chen. 2011. miR-335 andmiR-34a promote renal senescence by suppressing mitochondrial antioxidativeenzymes. J. Am. Soc. Nephrol. 22: 1252–1261.
24. Qin, B., H. Yang, and B. Xiao. 2012. Role of microRNAs in endothelial in-flammation and senescence. Mol. Biol. Rep. 39: 4509–4518.
25. Mestdagh, P., P. Van Vlierberghe, A. De Weer, D. Muth, F. Westermann,F. Speleman, and J. Vandesompele. 2009. A novel and universal method formicroRNA RT-qPCR data normalization. Genome Biol. 10: R64.
26. D’haene, B., P. Mestdagh, J. Hellemans, and J. Vandesompele. 2012. miRNAexpression profiling: from reference genes to global mean normalization.Methods Mol. Biol. 822: 261–272.
27. Rippo, M. R., F. Olivieri, V. Monsurro, F. Prattichizzo, M. C. Albertini, andA. D. Procopio. 2014. MitomiRs in human inflamm-aging: A hypothesis in-volving miR-181a, miR-34a and miR-146a. Exp. Gerontol. 56: 154–163.
28. Rudolf, E., S. John, and M. Cervinka. 2012. Irinotecan induces senescence andapoptosis in colonic cells in vitro. Toxicol. Lett. 214: 1–8.
29. Pan, D., A. Das, D. Liu, R. S. Veazey, and B. Pahar. 2012. Isolation and char-acterization of intestinal epithelial cells from normal and SIV-infected rhesusmacaques. PLoS ONE 7: e30247.
30. Kato, M., T. Paranjape, R. U. M€uller, S. Nallur, E. Gillespie, K. Keane,A. Esquela-Kerscher, J. B. Weidhaas, and F. J. Slack. 2009. The mir-34microRNA is required for the DNA damage response in vivo in C. elegans andin vitro in human breast cancer cells. Oncogene 28: 2419–2424.
31. Hu, H., and R. A. Gatti. 2011. MicroRNAs: new players in the DNA damageresponse. J. Mol. Cell Biol. 3: 151–158.
32. Lawless, C., C. Wang, D. Jurk, A. Merz, Tv. Zglinicki, and J. F. Passos. 2010.Quantitative assessment of markers for cell senescence. Exp. Gerontol. 45: 772–778.
33. Wang, C., D. Jurk, M. Maddick, G. Nelson, C. Martin-Ruiz, and T. vonZglinicki. 2009. DNA damage response and cellular senescence in tissues ofaging mice. Aging Cell 8: 311–323.
34. Yamakuchi, M., M. Ferlito, and C. J. Lowenstein. 2008. miR-34a repression ofSIRT1 regulates apoptosis. Proc. Natl. Acad. Sci. USA 105: 13421–13426.
35. Kauppinen, A., T. Suuronen, J. Ojala, K. Kaarniranta, and A. Salminen. 2013.Antagonistic crosstalk between NF-kB and SIRT1 in the regulation of inflam-mation and metabolic disorders. Cell. Signal. 25: 1939–1948.
The Journal of Immunology 15
by guest on February 7, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

36. Risques, R. A., L. A. Lai, T. A. Brentnall, L. Li, Z. Feng, J. Gallaher,M. T. Mandelson, J. D. Potter, M. P. Bronner, and P. S. Rabinovitch. 2008.Ulcerative colitis is a disease of accelerated colon aging: evidence from telomereattrition and DNA damage. Gastroenterology 135: 410–418.
37. Singh, U. P., N. P. Singh, B. Singh, L. J. Hofseth, R. L. Price, M. Nagarkatti, andP. S. Nagarkatti. 2010. Resveratrol (trans-3,5,49-trihydroxystilbene) induces si-lent mating type information regulation-1 and down-regulates nuclear tran-scription factor-kB activation to abrogate dextran sulfate sodium-induced colitis.J. Pharmacol. Exp. Ther. 332: 829–839.
38. Lewis, B. P., C. B. Burge, and D. P. Bartel. 2005. Conserved seed pairing, oftenflanked by adenosines, indicates that thousands of human genes are microRNAtargets. Cell 120: 15–20.
39. Guo, H., N. T. Ingolia, J. S. Weissman, and D. P. Bartel. 2010. MammalianmicroRNAs predominantly act to decrease target mRNA levels. Nature 466:835–840.
40. Su, B. H., Y. L. Tseng, G. S. Shieh, Y. C. Chen, Y. C. Shiang, P. Wu, K. J. Li,T. H. Yen, A. L. Shiau, and C. L. Wu. 2013. Prothymosin a overexpressioncontributes to the development of pulmonary emphysema. Nat. Commun. 4:1906.
41. O’Connell, R. M., D. S. Rao, and D. Baltimore. 2012. MicroRNA regulation ofinflammatory responses. Annu. Rev. Immunol. 30: 295–312.
42. Artis, D. 2008. Epithelial-cell recognition of commensal bacteria and mainte-nance of immune homeostasis in the gut. Nat. Rev. Immunol. 8: 411–420.
43. Zhang, H. S., X. Y. Chen, T. C. Wu, W. W. Sang, and Z. Ruan. 2012. MiR-34a isinvolved in Tat-induced HIV-1 long terminal repeat (LTR) transactivationthrough the SIRT1/NFkB pathway. FEBS Lett. 586: 4203–4207.
44. Chang, J. R., R. Mukerjee, A. Bagashev, L. Del Valle, T. Chabrashvili,B. J. Hawkins, J. J. He, and B. E. Sawaya. 2011. HIV-1 Tat protein promotesneuronal dysfunction through disruption of microRNAs. J. Biol. Chem. 286:41125–41134.
45. Yu, Z., D. Fan, B. Gui, L. Shi, C. Xuan, L. Shan, Q. Wang, Y. Shang, andY. Wang. 2012. Neurodegeneration-associated TDP-43 interacts with fragile Xmental retardation protein (FMRP)/Staufen (STAU1) and regulates SIRT1 ex-pression in neuronal cells. J. Biol. Chem. 287: 22560–22572.
46. Chaudhuri, A. D., S. V. Yelamanchili, M. C. Marcondes, and H. S. Fox. 2013.Up-regulation of microRNA-142 in simian immunodeficiency virus encephalitisleads to repression of sirtuin1. FASEB J. 27: 3720–3729.
47. Chun, T. W., D. C. Nickle, J. S. Justement, J. H. Meyers, G. Roby,C. W. Hallahan, S. Kottilil, S. Moir, J. M. Mican, J. I. Mullins, et al. 2008.Persistence of HIV in gut-associated lymphoid tissue despite long-term anti-retroviral therapy. J. Infect. Dis. 197: 714–720.
48. Huang, B., X. D. Yang, A. Lamb, and L. F. Chen. 2010. Posttranslationalmodifications of NF-kB: another layer of regulation for NF-kB signaling path-way. Cell. Signal. 22: 1282–1290.
49. Kwon, H. S., M. M. Brent, R. Getachew, P. Jayakumar, L. F. Chen, M. Schnolzer,M. W. McBurney, R. Marmorstein, W. C. Greene, and M. Ott. 2008. Humanimmunodeficiency virus type 1 Tat protein inhibits the SIRT1 deacetylase andinduces T cell hyperactivation. Cell Host Microbe 3: 158–167.
50. Barry, S. P., P. A. Townsend, R. A. Knight, T. M. Scarabelli, D. S. Latchman, andA. Stephanou. 2010. STAT3 modulates the DNA damage response pathway. Int.J. Exp. Pathol. 91: 506–514.
51. Li, S., H. F. Moffett, J. Lu, L. Werner, H. Zhang, J. Ritz, D. Neuberg,K. W. Wucherpfennig, J. R. Brown, and C. D. Novina. 2011. MicroRNA ex-pression profiling identifies activated B cell status in chronic lymphocytic leu-kemia cells. PLoS ONE 6: e16956.
52. Miyazaki, Y., R. Li, A. Rezk, H. Misirliyan, C. Moore, N. Farooqi, M. Solis,L. G. Goiry, O. de Faria Junior, V. D. Dang, et al; CIHR/MSSC New EmergingTeam Grant in Clinical Autoimmunity.; MSSRF Canadian B cells in MS Team.2014. A novel microRNA-132-surtuin-1 axis underlies aberrant B-cell cytokineregulation in patients with relapsing-remitting multiple sclerosis. PLoS ONE 9:e105421.
53. Nilssen, D. E., O. Øktedalen, and P. Brandtzaeg. 2004. Intestinal B cell hyper-activity in AIDS is controlled by highly active antiretroviral therapy. Gut 53:487–493.
54. Gordon, J. N., K. M. Pickard, A. Di Sabatino, J. D. Prothero, S. L. Pender,P. M. Goggin, and T. T. MacDonald. 2008. Matrix metalloproteinase-3 pro-duction by gut IgG plasma cells in chronic inflammatory bowel disease. Inflamm.Bowel Dis. 14: 195–203.
55. Staudt, L. M. 2010. Oncogenic activation of NF-kB. Cold Spring Harb. Perspect.Biol. 2: a000109.
56. Li, J., K. Wang, X. Chen, H. Meng, M. Song, Y. Wang, X. Xu, and Y. Bai. 2012.Transcriptional activation of microRNA-34a by NF-kB in human esophagealcancer cells. BMC Mol. Biol. 13: 4.
57. Lefranc, G., and M. P. Lefranc. 1990. Regulation of the immunoglobulin genetranscription. Biochimie 72: 7–17.
58. Oltz, E. M. 2001. Regulation of antigen receptor gene assembly in lymphocytes.Immunol. Res. 23: 121–133.
59. Durandy, A. 2002. Hyper-IgM syndromes: a model for studying the regulation ofclass switch recombination and somatic hypermutation generation. Biochem.Soc. Trans. 30: 815–818.
60. Doi, T. S., T. Takahashi, O. Taguchi, T. Azuma, and Y. Obata. 1997. NF-kBRelA-deficient lymphocytes: normal development of T cells and B cells, im-paired production of IgA and IgG1 and reduced proliferative responses. J. Exp.Med. 185: 953–961.
61. Roman-Blas, J. A., and S. A. Jimenez. 2006. NF-kB as a potential therapeutictarget in osteoarthritis and rheumatoid arthritis. Osteoarthritis Cartilage 14:839–848.
62. Uo, M., T. Hisamatsu, J. Miyoshi, D. Kaito, K. Yoneno, M. T. Kitazume,M. Mori, A. Sugita, K. Koganei, K. Matsuoka, et al. 2013. Mucosal CXCR4+
IgG plasma cells contribute to the pathogenesis of human ulcerative colitisthrough FcgR-mediated CD14 macrophage activation. Gut 62: 1734–1744.
63. Rao, D. S., R. M. O’Connell, A. A. Chaudhuri, Y. Garcia-Flores, T. L. Geiger,and D. Baltimore. 2010. MicroRNA-34a perturbs B lymphocyte development byrepressing the forkhead box transcription factor Foxp1. Immunity 33: 48–59.
64. Tome-Carneiro, J., M. Larrosa, M. J. Yanez-Gascon, A. Davalos, J. Gil-Zamorano, M. Gonzalvez, F. J. Garcıa-Almagro, J. A. Ruiz Ros, F. A. Tomas-Barberan, J. C. Espın, and M. T. Garcıa-Conesa. 2013. One-year supplementa-tion with a grape extract containing resveratrol modulates inflammatory-relatedmicroRNAs and cytokines expression in peripheral blood mononuclear cells oftype 2 diabetes and hypertensive patients with coronary artery disease. Phar-macol. Res. 72: 69–82.
16 miR-34a UPREGULATION IN COLON DURING CHRONIC SIV INFECTION
by guest on February 7, 2022http://w
ww
.jimm
unol.org/D
ownloaded from




















