
Geophagy amongst rhesus macaques on Cayo Santiago, Puerto Rico

A Test of the Cross-Generational Transmission of Grooming
Preferences in Macaques
Gabriele Schino*, Filippo Aureli�, Raffaella Ventura� & Alfonso Troisi§
*Istituto di Scienze e Tecnologie della Cognizione, Consiglio Nazionale delleRicerche, Rome, Italy; �School of Biological and Earth Sciences, Liverpool John
Moores University, UK; �School of Social and Health Sciences, Division ofPsychology, University of Abertay Dundee, UK; §Cattedra di Psichiatria, Univer-
sita Tor Vergata, Rome, Italy
Abstract
In this study we compared the grooming networks of macaque mothers andadult daughters in order to test whether primate social preferences may betransmitted from one generation to the next. We studied four social groupsbelonging to three different species (Macaca fascicularis, M. fuscata andM. sylvanus). We found no evidence for the transmission of individual degreesof kin bias or of preference for high-ranking individuals (transmission of �rules ofchoice�). Only in one of four social groups were idiosyncratic interindividualpreferences of mothers significantly similar to those of their adult daughters.Overall, the results of this study question the generality of the intergenerationaltransmission of social preferences in macaques. It is suggested that species-specificdegrees of preferences for kin and for high-ranking individuals should be selectedfor and strategically adjusted by individual females in relation to socialcircumstances.
Corresponding author: G. Schino, Via Lucilio 36, 00136 Roma, Italy.E-mail: [email protected]
Introduction
Primate social preferences reflect the need to establish social relationshipswith valuable partners (Wrangham 1983; Harcourt 1992). However, whileprimate social strategies and social preferences have been studied extensively,the study of their acquisition and learning during ontogeny has been largelyneglected (see Coussi-Korbel 1994 for a notable exception). Recently, both deWaal (1996) and Berman & Kapsalis (1999) reported that the affiliative relations(as measured by approaches and time spent in proximity) developed by immaturerhesus macaques (Macaca mulatta) can be constrained by their mother’s
Ethology 110, 137—146 (2004)
� 2004 Blackwell Verlag, BerlinISSN 0179–1613
U. S. Copyright Clearance Center Code Statement: 0179-1613/2004/11002–137/$15.00/0 www.blackwell-synergy.com

behaviour, so that the social networks of mothers and offspring come to show asignificant degree of similarity. These authors argued that cross-generationaltransmission of social tendencies is at the basis of the observed similarities in thesocial networks of mothers and offspring. However, several aspects of thishypothesized intergenerational transmission of social preferences remain to beclarified. First, it needs to be demonstrated that the observed effects extendbeyond the juvenile years into the adult life of offspring (when direct influence ofmothers is less pronounced). Secondly, these effects have to be confirmed in otherspecies and using other behavioural measures of affiliation. Thirdly, the putativemechanism(s) of transmission have to be further examined (Berman & Kapsalis1999).
In investigating cross-generational similarities in social networks one has alsoto distinguish between two different possibilities: the transmission of �rules ofchoice� (such as �prefer kin to a specific degree�), and the transmission ofidiosyncratic interindividual preferences (such as �prefer individual X overindividual Y�). In several primate species, social preferences are thought to begoverned by two main sources of attraction: an attraction to kin and an attractionto high rank (Seyfarth 1977; Schino 2001). Thus, mothers may be affecting theiroffspring social choices in terms of the degree of preference shown towards kinand towards high-ranking individuals. To evaluate the possibility of a cross-generational transmission of such rules of choice one should compare the degreeof preference shown towards kin (and towards high-ranking individuals) inmothers and offspring. Primates, however, may show interindividual preferencesthat are independent of the effects of kinship and social rank (in other words,relatedness and social rank do not explain 100% of the variance in the frequencyof affiliative interactions; see for example Schino 2001). Once the effects of thesetwo main sources of attraction are removed, idiosyncratic interindividualpreferences remain, and there is thus the possibility that these are indeedtransmitted from one generation to the next.
Of the two papers published thus far on the intergenerational transmission ofsocial preferences one (Berman & Kapsalis 1999) was clearly concerned only withrules of choice, while the other (de Waal 1996) focused on idiosyncraticinterindividual preferences. de Waal (1996), however, compared the socialpreferences of mothers among themselves with the social preferences of daughtersamong themselves. He thus restricted his analysis to the intergenerationaltransmission of intrageneration preferences. These two studies compared thesocial preferences of mothers (i.e. of adult females) with that of their offspringduring their infancy or adolescence and measured social preferences on thebasis of proxemic behaviour (i.e. non-agonistic approaches or interindividualproximity).
In this paper we investigate the above two possibilities of cross-generationaltransmission of social preferences by comparing grooming networks of mothersand of their adult daughters in macaques. Grooming is the most commonaffiliative behaviour of macaques and is considered a good index of affiliativerelationships (Oki & Maeda 1973). Our aim was to test the generality of the
138 G. Schino, F. Aureli, R. Ventura & A. Troisi

cross-generational transmission of social preferences by analysing the behaviourof three different species of macaques and focusing on social grooming, instead ofproximity, as a measure of interindividual affiliation. We observed fully adultsubjects in order to test whether the cross-generational influences reported forinfants and juveniles could be extended into the adult life of monkeys.
Methods
Subjects and Housing
Subjects of this study were four captive groups of macaques, two of long-tailed (Macaca fascicularis), one of Japanese (M. fuscata), and one of Barbary(M. sylvanus) macaques.
The long-tailed macaque groups were housed in indoor–outdoor enclosuresat the Laboratory of Comparative Physiology of the University of Utrecht, TheNetherlands. The two groups were established in 1971 and 1972, respectively, andmaternal kinship has been recorded since then. More details of the group historyand housing condition can be found in Aureli et al. (1995, 1997). At the time ofobservations, group A consisted of 26 animals and group B of 57 with nine and 24adult females (3.5-yr old or older and sexually active), respectively. Only the adultfemales were the subjects of the present study.
The Japanese macaque group was housed in a 700 m2 outdoor enclosure inthe Rome zoo (Bioparco), Italy. It was captured as a whole social group inTakasakiyama, Japan, in 1977, and has been under continuous observation at theRome zoo until 1999. Maternal genealogy is thus known for all animals born after1977. Details of the group history and housing condition can be found in Schinoet al. (1995). At the time of the observations the group comprised 71 animalsincluding 22 adult females (5-yr old or older, the typical reproductive years forJapanese macaques) that were the subjects of the observations.
The Barbary macaque group lived in a 5000 m2 outdoor grassy enclosure atthe Apenheul Zoo, the Netherlands. Information on maternal kinship has beenkept since the group formation in 1982. Details of the group history and housingcondition can be found in Aureli et al. (1997). At the time of the observations thegroup numbered 31 individuals. The 12 adult females (3.5-yr old or older andsexually active) were the subjects of the present study.
Behavioural Observations
Observations on the long-tailed macaques were conducted between Nov.1986 and Feb. 1987. The occurrences of allogrooming and the identity of groomerand gromee were collected in instantaneous group scans (Altmann 1974) carriedout every half an hour during 319 and 340 hours of observations in group A andB, respectively.
Observations on the Japanese macaques were carried out between Apr. andNov. 1996. The 22 female subjects were the focus of nine to 12 60-min observation
139Grooming in Macaques

sessions using the focal animal sampling technique (Altmann 1974). A total of 263hours of observation were made. The timing and duration of every episode ofallogrooming were recorded, along with the identity of the interactants.
Data on the Barbary macaques were collected between Dec. 1989 and Apr.1990. Data on allogrooming done and received by the adult females were gatheredduring 493 instantaneous group scans carried out every half an hour.
Within each group, a dominance hierarchy was assessed on the basis of thedirection of dyadic agonistic interactions.
Both the Japanese and Barbary macaques were observed outside of themating season (the long-tailed macaque is a non-seasonal breeder). Althoughobservations on the four different groups were conducted by different observersand with different data collection procedures (all providing estimates of the timespent grooming), observations within each group were consistent in bothobservers and procedures. Formal interobserver reliability tests were not carriedout, as allogrooming is a conspicuous and easily defined behaviour, so that majorbiases appear unlikely.
Data Analysis
Data analysis focused on the behaviour of mothers and their adult daughters.There were five and 13 mother-daughter dyads in the long-tailed macaque groupsA and B, respectively, 14 dyads in the Japanese macaque group, and five dyads inthe Barbary macaque group.
We first compared the grooming matrices of mothers and daughters usingrow-wise matrix correlations (the Kr test: Hemelrijk 1990a,b; de Vries 1993). Thetest of significance was based on 10 000 permutations. matman software was used(Noldus Information Technology 1998).
In order to calculate the degree of kin preference shown by each subject werelied on the method proposed by Berman & Kapsalis (1999). For each subject weregressed the grooming directed to each other group member on the groupmember’s degree of relatedness (r) to the subject through maternal lines. Degreeof relatedness to individuals belonging to different matrilines was assumed to bezero. The slope of the regression (b) was used as a measure of the subject’s degreeof kin preference. The degree of preference for high-ranking individuals wasestimated using a similar method, except that grooming was regressed on therecipient’s social rank. Correlations between the mothers� and the daughters�degree of preference for kin and for high-ranking individuals were calculatedusing the Kendall non-parametric correlation coefficient (s).
In order to evaluate the relation betweenmothers� and daughters� idiosyncraticpreferences we relied again on row-wise matrix correlations. First, we calculated foreach individual the multiple regression of grooming on the recipients� degree ofrelatedness and social rank. We then used the residuals of such multiple regressionsas a measure of the subject’s individual preferences once the effects of kinshipand dominance rank had been removed. We constructed matrices containingthe residuals of the above regressions of each individual (with mothers and
140 G. Schino, F. Aureli, R. Ventura & A. Troisi

corresponding daughters being in the same order in their respective matrix), andcorrelated the mothers� matrix with the daughters� matrix using the Kr test.
In order to allow comparison with the results of de Waal (1996), we alsocompared the intrageneration preferences of mothers and daughters. In otherwords, we tested whether daughters tended to prefer the daughters of theirmother’s preferred partners. For each social group we derived from the originalmatrix of grooming distribution a submatrix including only the mothers and asubmatrix including only the daughters (again with mothers and daughters beingin the same order in their respective matrix). The two submatrices were thencorrelated using the Kr test. In our sample some of the mothers had more thanone daughter present in the group. As the Kr test requires the two matrices to becorrelated to be of the same size, we randomly selected one daughter (the first inalphabetical order) for each mother for this analysis. This procedure reducedsample size and thus allowed only data from the two largest groups to be properlyanalysed (n ¼ 8 and n ¼ 7 in the long-tailed macaque group B and in theJapanese macaque group, respectively).
Overall, we used non-parametric statistics whenever possible. The onlyexceptions are the analyses of the slopes and residuals of regressions (see above),which are by necessity based on parametric techniques. Thus, parametric methodswere only used to obtain individual estimates of the degrees of preference for kinand for high-ranking individuals and of idiosyncratic preferences. Inferentialstatistics (and thus p-values) were always based on non-parametric techniques. Allprobabilities are two-tailed. Significance was set at p < 0.05.
Results
Overall Similarity of Grooming Distribution
The grooming matrices of mothers and daughters were significantly similarin three of four groups (long-tailed macaque group A: srow-wise ¼ 0.495,p < 0.02; long-tailed macaque group B: srow-wise ¼ 0.060, ns; Japanese macaques:srow-wise ¼ 0.176, p < 0.002; Barbary macaques: srow-wise ¼ 0.374, p < 0.01).
Transmission of Rules of Choice
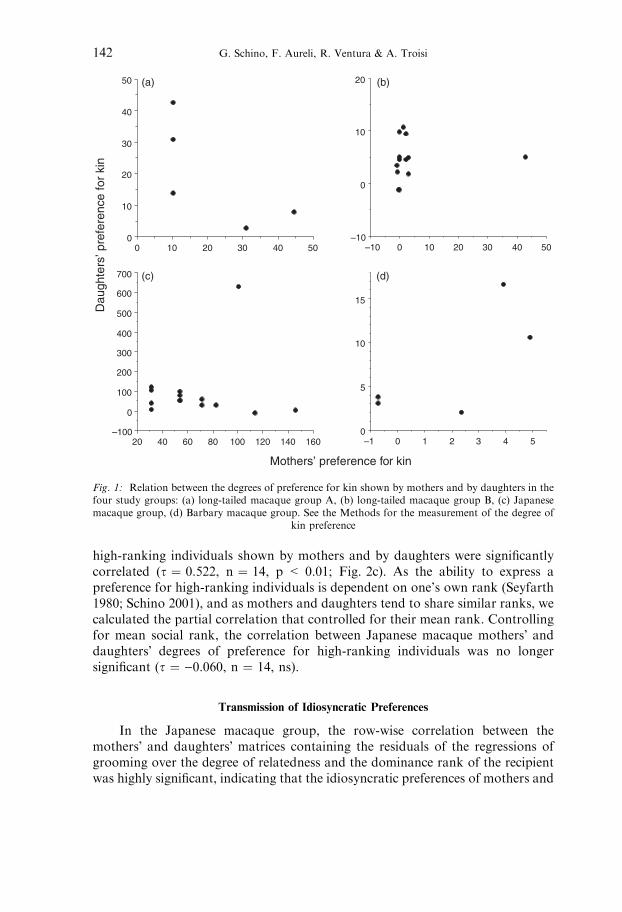
In none of the study groups the degrees of kin preference shown by mothersand by daughters were significantly correlated (long-tailed macaque group A:s ¼ )0.598, n ¼ 5, ns; long-tailed macaque group B: s ¼ 0.267, n ¼ 13, ns;Japanese macaques: s ¼ )0.309, n ¼ 14, ns; Barbary macaques: s ¼ 0.316,n ¼ 5, ns; Fig. 1).
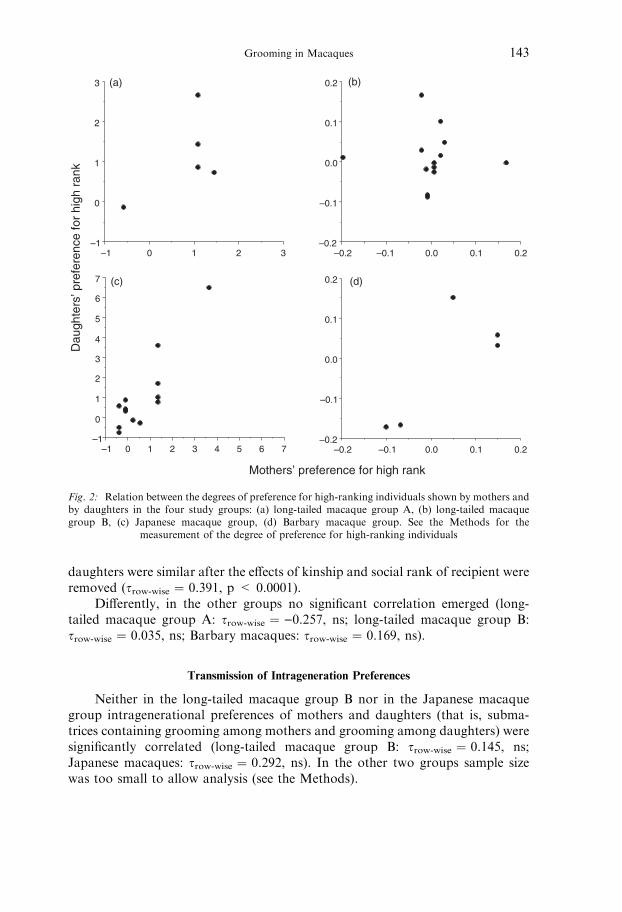
In three of the macaque groups under study correlations between themother’s and daughter’s degrees of preference for high-ranking individuals werenot significant (long-tailed macaque group A: s ¼ 0.120, n ¼ 5, ns; long-tailedmacaque group B: s ¼ 0.094, n ¼ 13, ns; Barbary macaques: s ¼ 0.527, n ¼ 5,ns; Fig. 2). Differently, in the Japanese macaque group degrees of preference for
141Grooming in Macaques
high-ranking individuals shown by mothers and by daughters were significantlycorrelated (s ¼ 0.522, n ¼ 14, p < 0.01; Fig. 2c). As the ability to express apreference for high-ranking individuals is dependent on one’s own rank (Seyfarth1980; Schino 2001), and as mothers and daughters tend to share similar ranks, wecalculated the partial correlation that controlled for their mean rank. Controllingfor mean social rank, the correlation between Japanese macaque mothers� anddaughters� degrees of preference for high-ranking individuals was no longersignificant (s ¼ )0.060, n ¼ 14, ns).
Transmission of Idiosyncratic Preferences
In the Japanese macaque group, the row-wise correlation between themothers� and daughters� matrices containing the residuals of the regressions ofgrooming over the degree of relatedness and the dominance rank of the recipientwas highly significant, indicating that the idiosyncratic preferences of mothers and
504030201000
10
20
30
40
50
50403020100–10–10
0
10
20
16014012010080604020–100
0
100
200
300
400
500
600
700
543210-1–10
5
10
15
Mothers’ preference for kin
Dau
ghte
rs’ p
refe
renc
e fo
r ki
n
(a) (b)
(c) (d)
Fig. 1: Relation between the degrees of preference for kin shown by mothers and by daughters in thefour study groups: (a) long-tailed macaque group A, (b) long-tailed macaque group B, (c) Japanesemacaque group, (d) Barbary macaque group. See the Methods for the measurement of the degree of
kin preference
142 G. Schino, F. Aureli, R. Ventura & A. Troisi
daughters were similar after the effects of kinship and social rank of recipient wereremoved (srow-wise ¼ 0.391, p < 0.0001).
Differently, in the other groups no significant correlation emerged (long-tailed macaque group A: srow-wise ¼ )0.257, ns; long-tailed macaque group B:srow-wise ¼ 0.035, ns; Barbary macaques: srow-wise ¼ 0.169, ns).
Transmission of Intrageneration Preferences
Neither in the long-tailed macaque group B nor in the Japanese macaquegroup intragenerational preferences of mothers and daughters (that is, subma-trices containing grooming among mothers and grooming among daughters) weresignificantly correlated (long-tailed macaque group B: srow-wise ¼ 0.145, ns;Japanese macaques: srow-wise ¼ 0.292, ns). In the other two groups sample sizewas too small to allow analysis (see the Methods).
3210–1–1
0
1
2
3
76543210–1–1
0
1
2
3
4
5
6
7
0.20.10.0–0.1–0.2–0.2
–0.1
0.0
0.1
0.2
0.20.10.0–0.1–0.2–0.2
–0.1
0.0
0.1
0.2(a) (b)
(c) (d)
Mothers’ preference for high rank
Dau
ghte
rs’ p
refe
renc
e fo
r hi
gh r
ank
Fig. 2: Relation between the degrees of preference for high-ranking individuals shown by mothers andby daughters in the four study groups: (a) long-tailed macaque group A, (b) long-tailed macaquegroup B, (c) Japanese macaque group, (d) Barbary macaque group. See the Methods for the
measurement of the degree of preference for high-ranking individuals
143Grooming in Macaques

Discussion
The results of this study provide little support for the hypothesis thatmacaques may transmit their grooming preferences from one generation to thenext. Although two of four of our study groups provided rather small samplesizes, the overall consistency of the results (despite methodological differences)make us relatively confident of their biological significance.
To properly investigate intergenerational transmission of grooming prefer-ences one should first distinguish between two different possibilities: thequantitative transmission of rules of choice and the transmission of idiosyncraticpreferences. Such distinction between these two possibilities had not beenpreviously recognized, but we believe future studies should take it into accountboth in their methods and in the discussion of their results.
The grooming distributions shown by mothers and daughters were signifi-cantly similar in three of four of the study groups. This result, however, is likelydue to the fact that both mothers and daughters preferred to groom their (mutual)kin, and both preferred high-ranking individuals. The question, however, is notwhether both mothers and daughters prefer kin, but whether they show similardegrees of preference. In this regard, we did not find any evidence favouring thequantitative transmission of rules of choice in any of the four macaque groupsunder study. Berman & Kapsalis (1999) observed that the degree of kin biasshown by 6-mo-old rhesus monkey infants was correlated to that of their motherand suggested that the perpetuation of similar degrees of kin bias acrossgenerations may depend on mothers and infants having similar experiences withmutual kin and non-kin. Our results, however, suggest that by the time offspringhave reached adulthood their social experiences may have diverged enough fromthat of their mothers so that no significant correlation between their and theirmothers� social preferences was left.
Rules of choice may be based on abstract concepts of kinship and socialrank. Macaques seem indeed to be able to form such abstract social concepts(Dasser 1988a,b), so that it seems unlikely that cognitive limitations constraint thepossibility of cross-generational transmission. On the other hand, from anevolutionary point of view it is difficult to hypothesize any adaptive advantage forthe cross-generational transmission of rules of choice. Social preferences thatcercopithecine females are able to express seem to derive from a compromisebetween attraction to valuable partners and competition for those partners(Seyfarth 1977; Schino 2001) with the added need to take into account possiblemarket effects (Barrett et al. 1999). Therefore, simply modelling one’s behaviouron that of one’s own mother is unlikely to have optimal consequences, at least forlow-ranking animals (whose mothers are most constrained in their social choices).It is more likely that a general propensity toward a certain degree of kin bias andattraction to high rank could be transmitted either genetically or socially (thusexplaining the overall similarity in the preferences of mothers and daughters),and individuals are then expected to strive for attaining such optimal degreein relation to the social competition they experience. The observation of
144 G. Schino, F. Aureli, R. Ventura & A. Troisi

species-specific degrees of kin bias (Aureli et al. 1997) is consistent with this view,while a role for phylogeny has also been suggested (Thierry et al. 2000).
Evidence for the transmission of idiosyncratic interindividual preferences wasmixed. Only in one of four social groups were idiosyncratic preferences of mothersand daughters significantly correlated (although two of the non-significant resultswere based on rather small sample sizes). It seems unlikely that variations inmethodology and housing conditions were at the basis of the observed differences,while the possible role of relatedness through paternal lines could not be explored(Smith et al. 2003).
It is possible that mother-daughter frequent proximity was at the basis of thesimilarity in their social preferences observed in Japanese macaques. Given thatdaughters spend much of their time in the company of their mothers, they arelikely to share proximity with any individual that happens to be a frequent socialpartner of their mothers. If such scenario is correct, the results we obtained wouldbe better described as mother–daughter shared preferences rather than mother–daughter transmission of preferences. To distinguish between these two compet-ing hypotheses one would need to experimentally remove mothers in order to testwhether the daughters� preferences persist in the absence of mothers and couldthus be correctly labelled as transmitted from one generation to the next. Sharedmother–daughter social preferences are expected to be more common in thosespecies (such as rhesus and Japanese macaques) showing the highest level of kinbias (Thierry 2000), as kin bias itself can facilitate the sharing of social preferences(Berman & Kapsalis 1999).
de Waal (1996) compared the intrageneration social preferences of mothersand of daughters (mother to mother vs. daughter to daughter preferences). Therationale for this analysis appears to assume that macaque social relationships arestrongly constrained by age differences, so that close bonds are established mostlyor only between same-age individuals. We also correlated the intragenerationpreferences of mothers and daughters, but no evidence for a similarity ofpreferences emerged.
In conclusion, the results of this study question the generality of theintergenerational transmission of social preferences in macaques. More long-termstudies of primate social preferences are needed to fully elucidate this phenomenon.
Acknowledgements
We thank the University of Utrecht, the Rome zoo (Bioparco) and the Apenheul zoo forallowing us to study their groups of macaques, Henk Westland for expert assistance, Carel van Schaik,Marjolijn Das, Karen Hiujsmans, Werner Groenhout, Marc Moonen, Desiree Verleur and KarinVocking for making data available, and Frans de Waal for his critical comments.
Literature Cited
Altmann, J. 1974: Observational study of behaviour: sampling methods. Behaviour, 49, 227—265.Aureli, F, van Panthaleon van Eck, C. J. & Veenema, H. C. 1995: Long-tailed macaques avoid conflicts
during short-term crowding. Aggress. Behav. 21, 113—122.
145Grooming in Macaques

Aureli, F., Das, M. & Veenema, H. C. 1997: Differential kinship effect on reconciliation in three speciesof macaques (Macaca fascicularis, M. fuscata and M. sylvanus). J. Comp. Psychol. 111, 91—99.
Barrett, L. & Henzi, S. P., Weingrill, T., Lycett, J. E., Hill, R. A. 1999. Market forces predict groomingreciprocity in female baboons. Proc. R. Soc. Lond. B 266, 665—670.
Berman, C. M. & Kapsalis, E. 1999: Development of kin bias among rhesus monkeys: maternaltransmission or individual learning? Anim. Behav. 58, 883—894.
Coussi-Korbel, S. 1994: Learning to outwit a competitor in mangabeys (Cercocebus torquatustorquatus). J. Comp. Psychol. 108, 164—171.
Dasser, V. 1988a: A social concept in Java monkeys. Anim. Behav. 36, 225—230.Dasser, V. 1988b: Mapping social concepts in monkeys. In: Machiavellian Intelligence (Byrne, R.W. &
Whiten, A., eds). Clarendon Press, Oxford, pp. 85—93.Harcourt, A. H. 1992: Coalitions and alliances: are primates more complex than non-primates? In:
Coalitions and Alliances in Humans and Other Animals (Harcourt, A. H. & de Waal, F. B. M.,eds). Oxford Univ. Press, Oxford, pp. 445—471.
Hemelrijk, C. K. 1990a: Models of and test for reciprocity unidirectionality and other social interactionpatterns at a group level. Anim. Behav. 39, 1013—1029.
Hemelrijk, C. K. 1990b: A matrix partial correlation test used in investigations of reciprocity and othersocial interaction patterns at group level. J. Theor. Biol. 143, 405—420.
Noldus Information Technology. 1998: MatMan, Reference Manual, Version 1.0. Wageningen.Oki, J. & Maeda, Y. 1973: Grooming as a regulator of behaviour in Japanese macaques. In:
Behavioural Regulators of Behaviour in Primates (Carpenter, C. R., ed.). Bucknell Univ. Press,Lewisburg, pp. 149—163.
Schino, G. 2001: Grooming, competition and social rank among female primates: a meta-analysis.Anim. Behav. 62, 265—271.
Schino, G., D’Amato, F. R. & Troisi, A. 1995: Mother-infant relationships in Japanese macaques:sources of interindividual variation. Anim. Behav. 49, 151—158.
Seyfarth, R. M. 1977: A model of social grooming among adult female monkeys. J. Theor. Biol. 65,671—698.
Seyfarth, R. M. 1980: The distribution of grooming and related behaviours among adult female vervetmonkeys. Anim. Behav. 28, 798—813.
Smith, K., Alberts, S. A. & Altmann, J. 2003. Wild female baboons bias their social behaviour towardspaternal half-sisters. Proc. R. Soc. Lond. Biol. Sci. 270, 503—510.
Thierry, B. 2000: Covariation of conflict management patterns across macaque species. In: NaturalConflict Resolution (Aureli, F. & de Waal, F. B. M., eds). University of California Press,Berkeley, pp. 106—128.
Thierry, B., Iwaniuk, A. N. & Pellis, S. M. 2000: The influence of phylogeny on the social behaviour ofmacaques (Primates: Cercopithecidae: genus Macaca). Ethology 106, 713—728.
de Vries, H. 1993: The row-wise correlation between two proximity matrices and the partial row-wisecorrelation. Psychometrika 58, 53—69.
de Waal, F. B. M. 1996: Macaque social culture: development and perpetuation of affiliative networks.J. Comp. Psychol. 110, 147—154.
Wrangham, R. W. 1983: Social relationships in comparative perspective. In: Primate SocialRelationships (Hinde, R. A., ed.). Blackwell, Oxford, pp. 325—334.
Received: April 7, 2003
Initial acceptance: August 27, 2003
Final acceptance: October 30, 2003 (M. Taborsky)
146 G. Schino, F. Aureli, R. Ventura & A. Troisi
Copyright © 2022 FDOKUMEN




















