
Dormant phages communicate via arbitrium to control exit ...
Upload
independentCategory
view
1download
0
Brain & Language 126 (2013) 181–187
Contents lists available at SciVerse ScienceDirect
Brain & Language
journal homepage: www.elsevier .com/locate /b&l
Tonkean macaques communicate with their right hand
0093-934X/$ - see front matter � 2013 Elsevier Inc. All rights reserved.http://dx.doi.org/10.1016/j.bandl.2013.05.004
⇑ Corresponding author. Address: Centre de Primatologie de l’Université deStrasbourg, Fort Foch, 67207 Niederhausbergen, France. Fax: +33 388137879.
E-mail addresses: [email protected] (H. Meunier), [email protected] (J. Fizet),[email protected] (J. Vauclair).
Hélène Meunier a,b,⇑, Jonas Fizet a,b, Jacques Vauclair b
a Primatology Centre of Strasbourg University, Fort Foch, 67207 Niederhausbergen, Franceb Centre for Research on the Psychology of Cognition, Language and Emotion, Aix-Marseille University, 29 Avenue Robert Schuman, 13621 Aix-en-Provence Cedex 1, France
a r t i c l e i n f o
Article history:Accepted 7 May 2013
Keywords:QHP taskHemispheric specializationGraspingPointingCommunicative gesturesMacaca tonkeanaLanguage origins
a b s t r a c t
There are two conflicting hypotheses to explain the origins of language. Vocal origin theory states thatlanguage results from the gradual evolution of animals’ vocal communication, but gestural origin theoryconsiders that language evolved from gestures, with the initial left-hemispheric control of manual ges-tures gradually encompassing vocalizations. To contribute to this debate, we investigated functionalhemispheric specialization related to hand biases when grasping or showing an object through manualgesture in Tonkean macaques. The results of this study, the first quantitative study on Tonkean macaques’handedness, showed a remarkable convergence of the Tonkean macaques’ handedness patterns withthose of baboons and human infants, with hand preferences for manual communicative gestures signif-icantly favoring the use of the right hand. Our findings support the hypothesis that left hemispheric lat-eralization for language is derived from a gestural communication system that was present in thecommon ancestor of macaques, baboons and humans.
� 2013 Elsevier Inc. All rights reserved.
1. Introduction
Human speech constitutes a particular communication systemin the animal kingdom. Our language is characterized by the flex-ible acquisition of shared conventions, and is cognitively very com-plex, requiring specific abilities such as intentionality or empathy(Pinker, 1998). It allows us to express an infinity of semantic nuan-ces and to transcend the spatiotemporal context in which speechtakes place (Kirtchuk, 2000). Our language is also the most obviousexpression of hemispheric specialization and has often been linkedto the fact that humans are mainly right-handed for many actions(Annett, 1985). It has been shown, for example, that humans pre-dominantly use their right hand for manual movements when theyare talking (Kimura, 1973) and that the degree of right-hand asym-metry for manual communication (e.g., pointing) increases duringthe development of speech in young children (e.g., Blake, O’Rourke,& Borzellino, 1994; Vauclair & Imbault, 2009).
The origins and lateralization of language have been debated formore than a century, but remain extremely controversial (e.g.,Christiansen & Kirby, 2003). Up to the 1980s, language was com-monly assumed to be unique to humans, suddenly emerging withHomo sapiens about 100,000 years ago. This theory has since fallenby the wayside and current theories include continuity betweenhumans and other animals and more ancestral origins (e.g., Fitch,
2010). Researchers therefore tend to adopt a phylogenetic ap-proach, and in this context nonhuman primates would appear tobe an ideal model for clarifying hypotheses on language origins.There are two main hypotheses: the vocal origin hypothesis statesthat language results from the gradual evolution of animals’ vocalcommunication (Lemasson, Gautier, & Hausberger, 2003; Lemas-son, Hausberger, & Zuberbuhler, 2005; Snowdon, 2002; Zuberbuh-ler, 2005), whereas the gestural origin hypothesis considers thatlanguage evolved from gestures (Corballis, 2002; Hewes, 1973;Vauclair, 2004). According to the latter, the left hemisphere ini-tially controlled manual gestures, and later came to encompassvocalizations, too. The close proximity between gestures and lan-guage raises the question of what role manual motor and commu-nicative functions played in the evolution of communicativesystems in humans (Gentilucci & Dalla Volta, 2008; Gentilucci, Dal-la Volta, & Gianelli, 2008). Observations of the gestural communi-cation systems of nonhuman primates have highlighted theirnumerous similarities with human language. Several species ofnonhuman primates spontaneously use their hands to communi-cate with their conspecifics in a variety of social contexts. Thisbehavior has mostly been studied in great apes (e.g., chimpanzees:Liebal, Call, & Tomasello, 2004; Pollick & De Waal, 2007; gorillas:Pika, Liebal, & Tomasello, 2003; bonobos: Pika, Liebal, & Tomasello,2005), while it also seems to be present in Old World monkeys(e.g., mandrills: Laidre, 2011; baboons: Kummer, 1968; Meguerdit-chian & Vauclair, 2006) and New World monkeys (e.g., squirrelmonkeys: Anderson, Kuwahata, & Fujita, 2007; capuchins: Mitchell& Anderson, 1997). Maestripieri (2005) found that communicativegestures varied between different species of macaques and most of

182 H. Meunier et al. / Brain & Language 126 (2013) 181–187
these gestures appear in dominance and submission contexts(Maestripieri, 2005). Unlike vocalizations, these communicativegestures are directed towards a particular recipient and requirean appropriate behavioral response from that recipient. Thesetwo characteristics suggest an intentional use of the gesture bythe sender (Arbib, Liebal, & Pika, 2008).
The study of manual communicative gestures and their asym-metries, providing information on underlying hemispheric special-ization, in nonhuman primates, thus constitutes an idealframework for clarifying the hypothesis of the gestural origin oflanguage and its cerebral lateralization (Corballis, 2002; Vauclair,2004). However, most studies of lateralization in nonhuman pri-mates have focused on noncommunicative motor actions. MacNei-lage, Studdert-Kennedy, and Lindblom (1987) for exampleproposed a specialization of the left hand for visually guided move-ments and of the right hand for more complex and/or bimanualmovements. But Papademetriou, Sheu, and Michel’s (2005) meta-analysis of primate hand preferences for reaching underlined thecurrent impossibility of forming empirically based theories fromwhich to generate worthwhile hypotheses about the evolution ofhuman brain–behavior relations. There is a clear lack of consis-tency between studies, and the considerable variability betweenfindings could be due mainly to the variability (e.g., in complexity)of the tasks used to assess hand preference (Fagot & Vauclair,1991). Tasks vary in terms not only of the reaching patterns thatare assessed but also of the individuals’ postures (e.g., sitting ormoving). McGrew and Marchant (1997) noted that tasks testingprimate handedness often involved reaching from an upright pos-ture, which is a highly unusual posture for nonhuman primates.The current literature highlights the need to collect more handed-ness data using similar, or at least comparable, tasks administeredto relatively large samples of individuals in the field and in captiv-ity. The biology of the species must, of course, be taken into ac-count before defining the degree of difficulty or complexity of agiven task. For example, a hanging task is easier for arboreal spe-cies than for terrestrial ones (Blois-Heulin, Bernard, & Bec, 2007;Blois-Heulin, Guitton, Nedellec-Bienvenue, Ropars, & Vallet, 2006).
Bishop, Ross, Daniels, and Bright (1996) were the first to pro-pose a standard method for quantifying handedness in humansto ensure uniformity in a field where failure to replicate is a majorproblem. They proposed a behavioral measure for quantifying theconsistency of hand preference (the so-called QHP task). This taskfosters the use of both hands by varying the spatial position of theobject to be reached for in relation to the body midline, the ideabeing to determine where, in the preferred hand’s contralateralspace, the individual shifts to using the non-preferred ipsilateralhand. The stronger our preference for our right hand, the furtherwe are likely to go into its contralateral space before switchingto our left hand, and viceversa. Meunier, Blois-Heulin, and Vauclair(2011) successfully adapted this task to Olive baboons (Papio anu-bis) to assess their handedness for a simple reaching task. However,it has been demonstrated that manual asymmetry is not always areliable cue for inferring hemispheric language specialization(Knecht et al., 2000a; Knecht et al., 2000b). One can assume thatthe study of hand preferences for communicative gestures (versusnoncommunicative gestures such as simple reaching) constitutes amore relevant approach to considering lateralization and languageorigin (Meguerditchian & Vauclair, 2009). The first studies to inves-tigate the lateralization of such behaviors in nonhuman primatesshowed that (i) great apes as well as baboons have a comparablepopulation-level bias in favor of their right hand when communi-cating with a human partner through manual gesture (chimpan-zees: Hopkins & Cantero, 2003; Hopkins & Wesley, 2002) orwhen threatening a conspecific or a human through hand-slapping(Olive baboons: Meguerditchian & Vauclair, 2006) and (ii) for bothchimpanzees and baboons, hand preference strength is signifi-
cantly greater for communicative tasks than for noncommunica-tive ones (Hopkins et al., 2005; Vauclair, Meguerditchian, &Hopkins, 2005), especially complex coordinated bimanual tasks(i.e., the tube task; see Hopkins, 1995, for more details; Meguerdit-chian, Molesti, & Vauclair, 2011).
One advantage of the standardized apparatus designed byBishop et al. (1996) and adapted to nonhuman primates by Meu-nier et al. (2011) is that it can be used in other tasks besides simplereaching, such as communication tasks involving pointing andmanual gesture (human infants: Jacquet, Esseily, Rider, & Fagard,2012; Olive baboons: Meunier, Vauclair, & Fagard, 2012). In thepresent study, we used this apparatus to explore handedness pat-terns for a communicative gesture with Tonkean macaques (Maca-ca tonkeana). In addition to this manual gesture, we also recordedhand biases for grasping in several spatial positions. Assuming thatlanguage, and thus its associated left-hemispheric control, does in-deed have its roots in gestural communication, we predicted thatthe Tonkean macaques would exhibit patterns of handedness com-parable to those of human infants and baboons. Furthermore, thesepatterns would differ according to the type of task, with the maca-ques being more right-hand/left-hemisphere specialized whenthey communicated through manual gesture than when they sim-ply grasped objects. The present study was the first of its kind totest macaques with the same setup as for both human infantsand baboons, and on the same communicative task.
2. Methods
All the experiments were carried out in accordance with thePrinciples of Laboratory Animal Care, and with CNRS guidelineson animal care.
2.1. Subjects
Data were collected from February to June 2011 at the Primatol-ogy Centre of Strasbourg University, France. The subjects were 13captive Tonkean macaques (M. tonkeana) including 5 adult males,4 adult females, 3 subadult males and 1 subadult female. All thesubjects lived in the same social group of 22 individuals and werehoused in a one-acre wooded area at the primatology center. Theyhad free access from the outdoor area to an indoor shelter. The ma-caques were fed with commercial primate pellets twice a day, freshfruits and vegetables once a day, and seeds three times a week.Water was available ad libitum, except during testing.
3. Experimental procedure
The experimental procedure was an adaptation of Bishop’s QHPtask, originally designed by Bishop et al. (1996) to evaluate the de-gree of hand preference in human children. These authors origi-nally placed seven picture cards, all 30� apart one from another,within the children’s reach. Children stood in front of the templateat the center of the baseline and were asked to pick up a specific,named card and place it in a box located directly in front of them.The experimenter recorded the hand used to pick up each card. Thecard order was random but the sequence of positions was the samefor all participants (Bishop et al., 1996). We adapted this apparatusto nonhuman primates for two tasks that differed only in their nat-ure: a noncommunicative, simple reaching task and a task involv-ing a communicative gesture.
The apparatus (see Fig. 1), composed of an experimental table(for the reaching task) and five platforms (for the communicativetask), was fixed to the wire mesh of a cage, next to the outdoorenclosure. To adjust the horizontal position of the subject, a con-crete block was placed inside the cage, perpendicular to the wire

Fig. 1. Schema of the experimental setup. Points on the smaller semicirclerepresent the seven positions used for the simple reaching task and points on thelarger semicircle represent the five positions used for the communicative task.
H. Meunier et al. / Brain & Language 126 (2013) 181–187 183
mesh and 25 cm below the level of the table, in its median plane.When a subject sat down on the concrete block, it therefore facedthe apparatus and was in line with the middle of the experimentaltable. A 10 � 60 cm hole in the wire mesh allowed the subject tomove its arms freely over the table. For both tasks, the subject’s ini-tial working position was seated in front of the apparatus with itssagittal plane in line with that of the table. Both hands were freeand placed on the table, symmetrically in relation to the centerof the table. Between two trials, the experimenter waited for thesubject to sit down again and resume the initial working position.The subjects came of their own free accord and were free to stopthe experiment whenever they wanted.
For the grasping task, seven positions were marked out in asemicircle on the experimental table, each separated by 30� andwithin reach of each subject’s hand (see Fig. 1). Positions 1, 2 and3 were located to the left of the subject, positions 5, 6 and 7 tothe right of the subject, and position 4 in the center. The distancebetween the positions and the subject was adjusted to take thesubject arm’s length into account: 20 cm for subadults and adultfemales, 25 cm for adult males.
Once the subject was in the initial working position, the exper-imenter placed a raisin on one of the seven positions. The experi-menter then recorded which hand was used to grasp the fooditem. As in Bishop’s QHP task, the position order of the baits wasrandom, but the presentation order was the same for all subjects.The use of the experimenter’s left or right hand to place each raisinon one of the seven positions on the experimental table was alsorandomized.
We recorded 4556 trials, taking all the positions and all the sub-jects into account. This number of trials corresponded to the num-ber of trials per subject for each of the seven positions (range: 20–60; M = 50; SE = 3.76).
For the communicative task, before the data recording phase,subjects underwent a preliminary training phase to ensure thatthey could all show the baited container with their right and lefthand. This training phase is necessary because showing an objectusing a manual gesture is not part of the natural behavioral reper-toire of this species. In the training protocol, the hand used by theexperimenter and the positions of the raisin were randomized. Incontrast to the grasping task, positions 1 and 7 of Bishop’s QHP taskwere eliminated, as they were too close to the subject. Thus, theapparatus featured five positions, each 80 cm from the subject(nonreachable distance). These positions were extensions of the ra-dii of positions 2–6 of the grasping task (see Fig. 1). Opaque con-
tainers were placed on the platforms at each position so that thebait could be concealed and thereby trigger communication be-tween the subject and the experimenter.
This task was performed by six subjects, which were trained toproduce manual communicative gestures via the following proce-dure. First, when the subject was correctly seated, a reachable rai-sin was presented to it on the experimental table such that thesubject could easily grasp it. This action was repeated severaltimes, the raisin being gradually moved away until it was in anambiguous situation for the animal, just out of reaching distance.At this stage, when the subject attempted to grasp the raisin, theexperimenter gave it directly to it. Next, the raisin was placed80 cm away from the subject, namely at a distance beyond the sub-ject’s reach. Finally, containers were placed in every position sothat the raisin was out of sight. The training phase ended whenthe subject was able to point to the out-of-reach baited containerwith its right and its left hand, without trying to grasp it. This tech-nique had already been used successfully with baboons (see Meu-nier et al., 2012). A manual communicative gesture was deemed tobe valid if the subject pointed with its whole hand, implying anarm extension towards the container, with the hand extended, fin-gers abducted (spread slightly apart) and the palm either held ver-tically or pronated to some degree (i.e. facing down, Leavens,Hopkins, & Bard, 1996).
Two experimenters performed the task. First, Experimenter 1hid the raisin in one of the five positions and went away. Experi-menter 2 was present, but standing at the back, from where hecould not make out which container was baited. In this way, we as-sure that (i) Experimenter 2 did not know where the food was hid-den and relied on information from the subject (via its manualgestures) to know which container to lift in order to reward it,and (ii) the monkeys has to request the food, and so, communicate,and that they are not just producing frustrated reaching responses.Experimenter 2 then positioned himself so that he was facing themacaque, which had to communicate the information to him byshowing through a manual gesture one of the containers. If the ma-caque showed the right container, Experimenter 2 gave the raisinto it and recorded the hand used for showing. If the macaqueshowed the wrong container, the food was removed and a new trialcommenced. Experimenter 1 waited until the subject was posi-tioned in the correct working position described above beforestarting the next trial.
We tested six macaques, including three adult males, one adultfemale and two subadult males. We recorded 854 trials, taking allthe positions and all the subjects into account. This number of tri-als corresponded to the number of trials per subject for each of thefive positions (range: 20–30; M = 28.5; SE = 1.53).
3.1. Data analysis
Z-scores were calculated for each subject and for each positionused in the two tasks, on the basis of the total number of reachesmade with each hand using the following formula:z-score ¼ ½R� ðRþ LÞ=2�=
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi½ðRþ LÞ=4�
p(where R represents the total
number of reaches using the right hand and L the total number ofreaches using the left hand). These scores allowed us to categorizemacaques as left-handed (z < �1.96), right-handed (z > 1.96) orambidextrous (�1.96 < z < 1.96) for each position.
We tested the effect of item position on hand preferences ineach task with a generalized linear mixed model (GLMM) for bino-mial data. Data set including individual data from the six individ-uals having performed both tasks were analyzed by fittingGLMM’s for binomial data. Both factors task (reaching versus ges-ture) and position (2–6) and their interactions were included asfixed effects and a random intercept was considered to deal withthe replications among the individuals. Model selection was based

Fig. 2. Number of left-handed, right-handed or individuals with no hand preferencefor each position and each task. (a) reaching task and (b) manual gesture task.
184 H. Meunier et al. / Brain & Language 126 (2013) 181–187
on the lowest AIC. We assessed the p-values of factors’ effect byusing likelihood ratio tests. Post-hoc multiple comparisons testsusing Tukey correction were conducted for comparing each pairof positions.
Results were considered to be significant when the p-value wasbelow 0.05.
4. Results
4.1. Individual hand preferences for each task and position
Regarding the six individuals having performed both tasks, andaccording to their individual z-scores (Table 1), the number of left-handers, right-handers and individuals with no preference variesacross the position for both tasks (see Fig. 2). In the reaching task,all the individuals are significantly left-handed for the positions ontheir left side (positions 1, 2 and 3) and right-handed for the posi-tions on their right (positions 5, 6 and 7). In the central position,one individual presents no preference and the five others areright-handed (Fig. 2a). Concerning the manual gesture task, handpreferences also depend on positions of the item to be shownbut the global pattern is less symmetrical: on their left side, threeindividuals are left-handed, two have no preference and one isright handed in position 2, whereas in position 3, one individualis still left-handed, two individuals have no preference and threeindividuals are right-handed. In the central position and the rightpositions, all the individuals are right-handed (Fig. 2b).
4.2. Effect of item position on hand preference
Regarding the reaching task, item position is a significant factor(LRT = 3660.5, Df = 6, p-value < 0.001). Multiple comparisons usingTukey correction reveal that all the positions differ significantlyfrom each other (p < 0.05) except for pairs of positions 1–2 and6–7 (see Fig. 3).
Regarding the communicative task, in positions 5 and 6, all ourindividuals exclusively used their right hand. Because of the nullvariance in those positions, it was not possible to test the effectof item position on manual gesture data with a GLMM includingall our data. Qualitatively, one can note that, as in the reachingtask, the more the item to be shown is on the right, the more ourindividuals are right-handed.
4.3. Effect of the communicative nature of the task on hand preference
We tested the effect of task on our data with a GLMM for bino-mial data with task, position and their interaction as fixed effectsand a random intercept to deal with the replications. This model
Table 1Individual z-score for each individual and each task (z-score > 1.96: significant right-hand phand preference).
Individuals Reaching
Position 1 Position 2 Position 3 Position 4 Position 5 Positio
Patsy �7.49 �7.75 �5.42 6.45 7.75 7.75Shan �7.23 �7.75 �5.42 5.16 7.23 7.49Ulysse �7.23 �5.94 �3.36 6.20 7.75 7.75Uruk �7.49 �7.49 �6.71 1.29 6.71 7.75Vischnu �7.75 �7.49 �6.97 2.07 7.23 7.49Wallace �6.41 �6.30 �4.91 3.18 5.20 6.43Ulrich �7.75 �7.75 �7.23 �2.32 4.13 5.94Lady �5.92 �5.75 �1.52 5.33 5.49 5.40Gaetan �7.75 �7.75 �6.97 �4.39 4.65 6.71Jeanne �5.29 �5.00 �5.00 1.80 4.31 5.20Néreis �5.48 �5.11 �2.56 1.83 4.75 5.48Vega �7.75 �7.75 �7.23 �4.13 2.84 6.71Walt �3.64 �3.13 �0.23 2.06 4.26 4.69
indicates that the interaction task/position does not affect signifi-cantly our data (LRT = 1.4669; Df = 4; p = 0.8325), i.e. task effectis the same whatever the position and vice versa, position effectis the same whatever the task. Then, we tested the same data witha GLMM with binomial taking into account the task and the posi-tion but not the interaction as fixed effects. This second model re-veals a lowest AIC than the first one does. This model showed thattask significantly influences hand preferences, subjects using sig-nificantly more their right hand for the communicative task thanfor the grasping task (LRT = 313.34; Df = 1; p < 0.0001, see alsoFig. 4).
5. Discussion
This study has yielded several original results. It is the first tohave collected information on hand preferences of several Tonkeanmacaques using tasks of varying nature, i.e., communicative or not(but see Ueno and Fujita (1998) study describing hand preference
reference; 1.96 > z-score > �1.96: no hand preference; z-score < �1.96: significant left-
Manual gesture
n 6 Position 7 Position 2 Position 3 Position 4 Position 5 Position 6
7.75 4.75 5.48 5.48 5.48 5.487.75 �5.48 �4.02 5.11 5.48 5.487.75 �4.02 �1.10 4.75 5.48 5.487.75 �1.10 5.11 5.48 5.48 5.487.75 �4.75 1.83 5.11 5.48 5.486.86 1.15 2.98 4.47 4.69 4.587.23 – – – – –6.00 – – – – –7.75 – – – – –5.00 – – – – –5.48 – – – – –7.75 – – – – –4.12 – – – – –
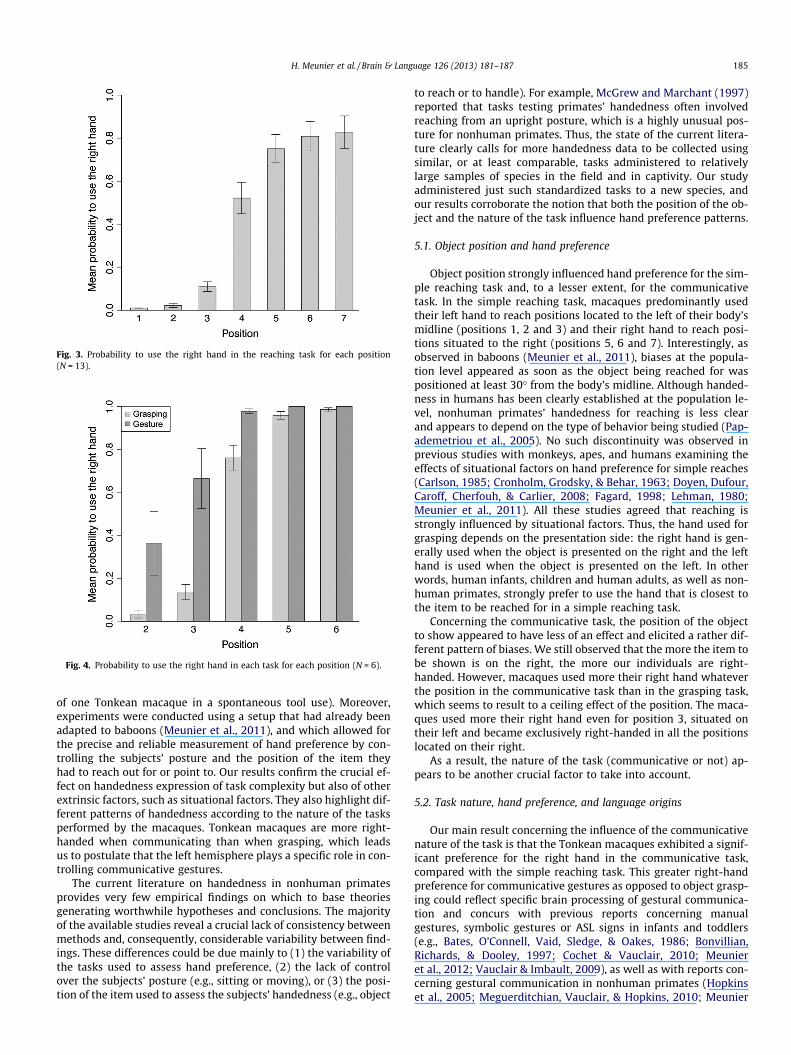
Fig. 3. Probability to use the right hand in the reaching task for each position(N = 13).
Fig. 4. Probability to use the right hand in each task for each position (N = 6).
H. Meunier et al. / Brain & Language 126 (2013) 181–187 185
of one Tonkean macaque in a spontaneous tool use). Moreover,experiments were conducted using a setup that had already beenadapted to baboons (Meunier et al., 2011), and which allowed forthe precise and reliable measurement of hand preference by con-trolling the subjects’ posture and the position of the item theyhad to reach out for or point to. Our results confirm the crucial ef-fect on handedness expression of task complexity but also of otherextrinsic factors, such as situational factors. They also highlight dif-ferent patterns of handedness according to the nature of the tasksperformed by the macaques. Tonkean macaques are more right-handed when communicating than when grasping, which leadsus to postulate that the left hemisphere plays a specific role in con-trolling communicative gestures.
The current literature on handedness in nonhuman primatesprovides very few empirical findings on which to base theoriesgenerating worthwhile hypotheses and conclusions. The majorityof the available studies reveal a crucial lack of consistency betweenmethods and, consequently, considerable variability between find-ings. These differences could be due mainly to (1) the variability ofthe tasks used to assess hand preference, (2) the lack of controlover the subjects’ posture (e.g., sitting or moving), or (3) the posi-tion of the item used to assess the subjects’ handedness (e.g., object
to reach or to handle). For example, McGrew and Marchant (1997)reported that tasks testing primates’ handedness often involvedreaching from an upright posture, which is a highly unusual pos-ture for nonhuman primates. Thus, the state of the current litera-ture clearly calls for more handedness data to be collected usingsimilar, or at least comparable, tasks administered to relativelylarge samples of species in the field and in captivity. Our studyadministered just such standardized tasks to a new species, andour results corroborate the notion that both the position of the ob-ject and the nature of the task influence hand preference patterns.
5.1. Object position and hand preference
Object position strongly influenced hand preference for the sim-ple reaching task and, to a lesser extent, for the communicativetask. In the simple reaching task, macaques predominantly usedtheir left hand to reach positions located to the left of their body’smidline (positions 1, 2 and 3) and their right hand to reach posi-tions situated to the right (positions 5, 6 and 7). Interestingly, asobserved in baboons (Meunier et al., 2011), biases at the popula-tion level appeared as soon as the object being reached for waspositioned at least 30� from the body’s midline. Although handed-ness in humans has been clearly established at the population le-vel, nonhuman primates’ handedness for reaching is less clearand appears to depend on the type of behavior being studied (Pap-ademetriou et al., 2005). No such discontinuity was observed inprevious studies with monkeys, apes, and humans examining theeffects of situational factors on hand preference for simple reaches(Carlson, 1985; Cronholm, Grodsky, & Behar, 1963; Doyen, Dufour,Caroff, Cherfouh, & Carlier, 2008; Fagard, 1998; Lehman, 1980;Meunier et al., 2011). All these studies agreed that reaching isstrongly influenced by situational factors. Thus, the hand used forgrasping depends on the presentation side: the right hand is gen-erally used when the object is presented on the right and the lefthand is used when the object is presented on the left. In otherwords, human infants, children and human adults, as well as non-human primates, strongly prefer to use the hand that is closest tothe item to be reached for in a simple reaching task.
Concerning the communicative task, the position of the objectto show appeared to have less of an effect and elicited a rather dif-ferent pattern of biases. We still observed that the more the item tobe shown is on the right, the more our individuals are right-handed. However, macaques used more their right hand whateverthe position in the communicative task than in the grasping task,which seems to result to a ceiling effect of the position. The maca-ques used more their right hand even for position 3, situated ontheir left and became exclusively right-handed in all the positionslocated on their right.
As a result, the nature of the task (communicative or not) ap-pears to be another crucial factor to take into account.
5.2. Task nature, hand preference, and language origins
Our main result concerning the influence of the communicativenature of the task is that the Tonkean macaques exhibited a signif-icant preference for the right hand in the communicative task,compared with the simple reaching task. This greater right-handpreference for communicative gestures as opposed to object grasp-ing could reflect specific brain processing of gestural communica-tion and concurs with previous reports concerning manualgestures, symbolic gestures or ASL signs in infants and toddlers(e.g., Bates, O’Connell, Vaid, Sledge, & Oakes, 1986; Bonvillian,Richards, & Dooley, 1997; Cochet & Vauclair, 2010; Meunieret al., 2012; Vauclair & Imbault, 2009), as well as with reports con-cerning gestural communication in nonhuman primates (Hopkinset al., 2005; Meguerditchian, Vauclair, & Hopkins, 2010; Meunier

186 H. Meunier et al. / Brain & Language 126 (2013) 181–187
et al., 2012). Moreover, chimpanzees’ right-handedness for food-begging gestures (Taglialatela, Cantalupo, & Hopkins, 2006; Tagli-alatela, Russell, Schaeffer, & Hopkins, 2008) and for clapping, i.e.,a communicative signal (Meguerditchian, Gardner, Schapiro, &Hopkins, 2012) is associated with morphological left asymmetriesin the homologue of Broca’s area (inferior frontal gyrus).
6. Conclusions
These findings support the hypothesis that language and itsleft-hemispheric control may be derived from a gestural communi-cation system that was present in the common ancestor of maca-ques, baboons, chimpanzees and humans. Vocalizations, albeitlatecomers in the evolution of intentional communication, mayhave gradually invaded this gestural system in the course of evolu-tion (Corballis, 2010), eventually leading to the dominance of thevocal modality (speech) in humans.
Acknowledgments
This research was supported by a French National ResearchAgency (ANR) Grant, reference ANR-08-BLAN-0011_01. Theauthors are grateful to Nicolas Herrenschmidt and all his teamfor permission to conduct the study at the Primate Centre of Stras-bourg University, France. The authors also sincerely thank Char-lotte Canteloup and Laurence Böes for helping with the datacollection, Jean-Marc Woock, Adrien Panter and Steve Lapp fortheir logistical aid, all the caretakers for their helpful assistancethroughout the study and to Elizabeth Portier for correcting theEnglish. Thanks are extended to Nicolas Poulin from CeStatS forstatistical assistance.
References
Anderson, J. R., Kuwahata, H., & Fujita, K. (2007). Gaze alternation during ‘‘pointing’’by squirrel monkeys (Saimiri sciureus)? Animal Cognition, 10, 267–271.
Annett, M. (1985). Left, right, hand and brain: The right shift theory. Hillsdale, NJ:Erlbaum.
Arbib, M. A., Liebal, K., & Pika, S. (2008). Primate vocalization, gesture, and theevolution of human language. Current Anthropology, 49, 1053–1076.
Bates, E., O’Connell, B., Vaid, J., Sledge, P., & Oakes, L. (1986). Language and handpreference in early development. Developmental Neuropsychology, 2, 1–15.
Bishop, D. V. M., Ross, V. A., Daniels, M. S., & Bright, P. (1996). The measurement ofhand preference: A validation study comparing three groups of right-handers.British Journal of Psychology, 87, 269–285.
Blake, J., O’Rourke, P., & Borzellino, G. (1994). Form and function in the developmentof pointing and reaching gestures. Infant Behavior and Development, 17,195–203.
Blois-Heulin, C., Bernard, V., & Bec, P. (2007). Postural effect on manual laterality indifferent tasks in captive grey-cheeked mangabey (Lophocebus albigena). Journalof Comparative Psychology, 121, 205–213.
Blois-Heulin, C., Guitton, J. S., Nedellec-Bienvenue, D., Ropars, L., & Vallet, E. (2006).Hand preference in unimanual and bimanual tasks and postural effect onmanual laterality in captive red-capped mangabeys (Cercocebus torquatustorquatus). American Journal of Primatology, 68, 429–444.
Bonvillian, J. D., Richards, H. C., & Dooley, T. T. (1997). Early sign languageacquisition and the development of hand preference in young children. Brainand Language, 58, 1–22.
Carlson, D. F. (1985). Development of the infants’ hand preference for visuallydirected reaching: Preliminary report of a longitudinal study. Infant MentalHealth Journal, 6, 158–174.
Christiansen, M. H., & Kirby, S. (2003). Language evolution: Consensus andcontroversies. Trends in Cognitive Sciences, 7, 300–307.
Cochet, H., & Vauclair, J. (2010). Pointing gestures produced by toddlers from 15 to30 months: Different functions, hand shapes and laterality patterns. InfantBehavior and Development, 33, 432–442.
Corballis, M. C. (2002). From hand to mouth. The origins of language. Princeton, NJ:Princeton University Press.
Corballis, M. C. (2010). Mirror neurons and the evolution of language. Brain andLanguage, 112, 25–35.
Cronholm, J. M., Grodsky, M., & Behar, I. (1963). Situational factors in the lateralpreference in rhesus monkeys. Journal of Genetic Psychology, 103, 167–174.
Doyen, A.-L., Dufour, T., Caroff, X., Cherfouh, A., & Carlier, M. (2008). Handpreference and hand performance: Cross-sectional developmental trends andfamily resemblance in degree of laterality. Laterality, 13, 179–197.
Fagard, J. (1998). Changes in grasping skills and the emergence of bimanualcoordination during the first year of life. In K. J. Connolly (Ed.), The psychology ofthe hand (pp. 123–143). London: Mac Keith Press.
Fagot, J., & Vauclair, J. (1991). Manual laterality in nonhuman-primates: Adistinction between handedness and manual specialization. PsychologicalBulletin, 109, 76–89.
Fitch, W. T. (2010). The evolution of language. London: Cambridge University Press.Gentilucci, M., & Dalla Volta, R. D. (2008). Spoken language and arm gestures are
controlled by the same motor control system. Quarterly Journal of ExperimentalPsychology, 61, 944–957.
Gentilucci, M., Dalla Volta, R. D., & Gianelli, C. (2008). When the hands speak. Journalof Physiology-Paris, 102, 21–30.
Hewes, G. W. (1973). Primate communication and gestural origin of language.Current Anthropology, 14, 5–24.
Hopkins, W. D. (1995). Hand preferences for a coordinated bimanual task in 110chimpanzees (Pan troglodytes): Cross-sectional analysis. Journal of ComparativePsychology, 109, 291–297.
Hopkins, W. D., & Cantero, M. (2003). From hand to mouth in the evolution oflanguage: The influence of vocal behavior on lateralized hand use in manualgestures by chimpanzees (Pan troglodytes). Developmental Science, 6, 55–61.
Hopkins, W. D., Russell, J., Freeman, H., Buehler, N., Reynolds, E., & Schapiro, S. J.(2005). The distribution and development of handedness for manual gestures incaptive chimpanzees (Pan troglodytes). Psychological Science, 16, 487–493.
Hopkins, W. D., & Wesley, M. J. (2002). Gestural communication in chimpanzees(Pan troglodytes): The influence of experimenter position on gesture type andhand preference. Laterality, 7, 19–30.
Jacquet, A.-Y., Esseily, R., Rider, D., & Fagard, J. (2012). Handedness for graspingobjects and declarative pointing: A longitudinal study. DevelopmentalPsychobiology, 54, 36–46. http://dx.doi.org/10.1002/dev.20572.
Kimura, D. (1973). Manual activity during speaking: I. Right-handers.Neuropsychologia, 11, 45–50.
Kirtchuk, P. I. (2000). Some cognitive and typological semantic remarks on thelanguage of 4QMMT. In T. Muraoka & J. F. Elwolde (Eds.), Diggers at the well(pp. 131–136). Pa Leiden: E.J. Brill.
Knecht, S., Deppe, M., Drager, B., Bobe, L., Lohmann, H., Ringelstein, E. B., et al.(2000a). Language lateralization in healthy right-handers. Brain, 123, 74–81.
Knecht, S., Drager, B., Deppe, M., Bobe, L., Lohmann, H., Floel, A., et al. (2000b).Handedness and hemispheric language dominance in healthy humans. Brain,123, 2512–2518.
Kummer, H. (1968). Social organization of hamadryas baboons – A field study.Chicago: University of Chicago Press.
Laidre, M. E. (2011). Meaningfull gesture in monkeys? Investigating whethermandrills create social culture. PloS One, 6, e14610.
Leavens, D. A., Hopkins, W. D., & Bard, K. A. (1996). Indexical and referentialpointing in chimpanzees (Pan troglodytes). Journal of Comparative Psychology,110, 346–353.
Lehman, R. A. W. (1980). Distribution and changes in strength of hand preference ofcynomolgus monkeys. Brain Behavior and Evolution, 17, 209–217.
Lemasson, A., Gautier, J. P., & Hausberger, M. (2003). Vocal similarities and socialbonds in Campbell’s monkey (Cercopithecus campbelli). Comptes RendusBiologies, 326, 1185–1193.
Lemasson, A., Hausberger, M., & Zuberbuhler, K. (2005). Socially meaningful vocalplasticity in adult Campbell’s monkeys (Cercopithecus campbelli). Journal ofComparative Psychology, 119, 220–229.
Liebal, K., Call, J., & Tomasello, M. (2004). Use of gesture sequences in chimpanzees.American Journal of Primatology, 64, 377–396.
MacNeilage, P. F., Studdert-Kennedy, M. G., & Lindblom, B. (1987). Primatehandedness reconsidered. Behavioural and Brain Sciences, 10, 247–303.
Maestripieri, D. (2005). Gestural communication in three species of macaques(Macaca mulatta, M. nemestrina, M. arctoides). Use of signals in relation todominance and social context. Gesture, 5, 55–71.
McGrew, W. C., & Marchant, L. F. (1997). Using the tools at hand: Manual lateralityand elementary technology in Cebus spp. and Pan spp. International Journal ofPrimatology, 18, 787–810.
Meguerditchian, A., Gardner, M. J., Schapiro, S. J., & Hopkins, W. D. (2012). The soundof one hand clapping: handedness and perisylvian neural correlates of acommunicative gesture in chimpanzees. Proceeding of the Royal Society(Biology), 279, 1959–1966.
Meguerditchian, A., Molesti, S., & Vauclair, J. (2011). Right-handednesspredominance in 162 baboons for gestural communication: Consistencyacross time and groups. Behavioral Neuroscience, 125, 653–660.
Meguerditchian, A., & Vauclair, J. (2006). Baboons communicate with their righthand. Behavioural Brain Research, 171, 170–174.
Meguerditchian, A., & Vauclair, J. (2009). Contrast of hand preferences betweencommunicative gestures and non communicative actions in baboons:Implications for the origins of hemispheric specialization for language. Brainand Language, 108, 167–174.
Meguerditchian, A., Vauclair, J., & Hopkins, W. D. (2010). Captive chimpanzees usetheir right hand to communicate with each other: Implications for the origin ofthe cerebral substrate for language. Cortex, 46, 40–48.
Meunier, H., Blois-Heulin, C., & Vauclair, J. (2011). A new tool for measuring handpreference in non-human primates: Adaptation of Bishop’s Quantifying HandPreference task for Olive baboons. Behavioural Brain Research, 218, 1–7.
Meunier, H., Vauclair, J., & Fagard, J. (2012). Human infants and baboons show thesame pattern of handedness for a communicative gesture. PloS One, 7(3),e33959. http://dx.doi.org/10.1371/journal.pone.0033959.

H. Meunier et al. / Brain & Language 126 (2013) 181–187 187
Mitchell, R. W., & Anderson, J. R. (1997). Pointing, withholding information, anddeception in capuchin monkeys (Cebus apella). Journal of ComparativePsychology, 111, 351–361.
Papademetriou, E., Sheu, C. F., & Michel, G. F. (2005). A meta-analysis of primatehand preferences, particularly for reaching. Journal of Comparative Psychology,119, 33–48.
Pika, S., Liebal, K., & Tomasello, M. (2003). Gestural communication in younggorillas (Gorilla gorilla): Gestural repertoire, learning, and use. American Journalof Primatology, 60, 95–111.
Pika, S., Liebal, K., & Tomasello, M. (2005). Gestural communication in subadultbonobos (Pan paniscus): Repertoire and use. American Journal of Primatology, 65,39–61.
Pinker, S. (1998). Words and rules. Lingua, 106, 219–242.Pollick, A. F., & De Waal, F. B. M. (2007). Ape gestures and language evolution.
Proceedings of the National Academy of Sciences, 104, 8184–8189.Snowdon, C. T. (2002). From primate communication to human language. In F. B. M.
de Waal (Ed.), Tree of origin: What primate behavior can tell us about human socialevolution (pp. 193–227). Cambridge, MA: Harvard University Press.
Taglialatela, J. P., Cantalupo, C., & Hopkins, W. D. (2006). Gesture handedness predictsasymmetry in the chimpanzee inferior frontal gyrus. NeuroReport, 17, 923–927.
Taglialatela, J. P., Russell, J. L., Schaeffer, J. A., & Hopkins, W. D. (2008).Communicative signaling activates ‘Broca’s homolog in chimpanzees. CurrentBiology, 18(5), 343–348.
Ueno, Y., & Fujita, K. (1998). Spontaneous tool use by a tonkean macaque (Macacatonkeana). Folia Primatologica, 69, 318–324.
Vauclair, J. (2004). Lateralization of communicative signals in nonhuman primatesand the hypothesis of the gestural origin of language. Interaction Studies. SocialBehaviour and Communication in Biological and Artificial Systems, 5, 363–384.
Vauclair, J., & Imbault, J. (2009). Relationships between manual preferences forobject manipulation and pointing gestures in infants and toddlers.Developmental Science, 12, 1060–1069.
Vauclair, J., Meguerditchian, A., & Hopkins, W. D. (2005). Hand preferences forunimanual and coordinated bimanual tasks in baboons (Papio anubis). CognitiveBrain Research, 25, 210–216.
Zuberbuhler, K. (2005). The phylogenetic roots of language – Evidence from primatecommunication and cognition. Current Directions in Psychological Science, 14,126–130.
Copyright © 2022 FDOKUMEN




















