
Endocranial suture closure in rhesus macaques (Macaca mulatta)
12
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 80:417-428 (1989) Endocranial Suture Closure in Rhesus Macaques (Macaca mulaita) DEAN FALK, LYLE KONIGSBERG, R. CRISS HELMKAMP, JAMES CHEVERUD, MICHAEL VANNIER, AND CHARLES HILDEBOLT Department of Anthropology, State University of New York, Albany, New York 12222 (D.F.), Departments of Anthropology (L.K.,J.C.) and Cell Biology and Anatomy (J.C.), Northwestern University, Evanston, Illinois 60208; Department of Sociology and Anthropology, Purdue University, West Lafayette, Indiana 47907 (R.C.H.); Mallinckrodt Institute of Radiology, Washington University School of Medicine, St. Louis, Missouri 63110 (M.V., C.H.) KEY WORDS Endocasts, Petalias Age determination, Asymmetries, Cranium, ABSTRACT Endocasts from skulls of 330 rhesus monkeys (Mucaca mu- Zatta) of known age are scored for closure of nine bilateral and three unilateral sutures or segments of sutures. A variety of tests reveals a strong relationship between age and stages of suture closure, although increasingly broad confi- dence intervals prevent sutures from being very useful for precisely aging older macaques. The order in which endosutures begin to close, as well as that in which closure is finally achieved, is determined for macaques, and these sequences compared to those for endosutures of humans (Todd and Lyon, 1924). The basilar suture is the earliest to close, while the masto-occipital and rostral and caudal squamosal sutures achieve closure quite late in both species. On the other hand, humans and macaques differ in their schedules for the sphenofrontal suture and in the initiation of closure for the rostral portion of the squamosal suture. Two sutures close significantly sooner on the right than on the left side (the rostral squamosal and masto-occipital) and asymme- try favoring closure of the right lateral lambdoid suture also approaches significance at the 0.05 level. No sutures close significantly sooner on the left side. It is suggested that macaque sutures may close from the inside out, that endosutures are more sensitive than ectosutures for detecting sequences in which cranial sutures begin to close, and that directional asymmetries in suture closure of macaques may be related to minor asymmetries in braid skull shape (petalias). The use of cranial suture closure to age human skeletal material has had a check- ered history. Numerous workers have raised serious questions about using suture closure as an aging method (Singer, 1953; Brooks, 1955; McKern and Stewart, 1957). However, Meindl and Lovejoy (1985) have recently shown that a combination of segments of ectocranial sutures can contribute valuable information to age estimates. In this report, we explore the relationship between closure of endocranial sutures and age in rhesus monkeys (Macaca mulatta) and compare our results with those for both endocranial (Todd and Lyon, 1924) and ectocranial (Meindl and Lovejoy, 1985) sutures of humans. The order of suture closure is compared in the two species. Because Zivanovic (1983) has noted a considerable amount of hemispheric asym- metry in suture closure among humans, we tested for directional asymmetry in suture closure in rhesus monkeys. Our findings may shed light on the development of cranial petalias in human (LeMay, 1977; Galaburda et al., 1978) and nonhuman (LeMay et al., 1982) primates. Received October 19, 1988; revision accepted November 14, 1988. @ 1989 ALAN R. LISS, INC
Transcript of Endocranial suture closure in rhesus macaques (Macaca mulatta)

AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 80:417-428 (1989)
Endocranial Suture Closure in Rhesus Macaques (Macaca mulaita)
DEAN FALK, LYLE KONIGSBERG, R. CRISS HELMKAMP, JAMES CHEVERUD, MICHAEL VANNIER, AND CHARLES HILDEBOLT Department of Anthropology, State University of New York, Albany, New York 12222 (D.F.), Departments of Anthropology (L.K., J.C.) and Cell Biology and Anatomy (J.C.), Northwestern University, Evanston, Illinois 60208; Department of Sociology and Anthropology, Purdue University, West Lafayette, Indiana 47907 (R.C.H.); Mallinckrodt Institute of Radiology, Washington University School of Medicine, St. Louis, Missouri 63110 (M.V., C.H.)
KEY WORDS Endocasts, Petalias
Age determination, Asymmetries, Cranium,
ABSTRACT Endocasts from skulls of 330 rhesus monkeys (Mucaca mu- Zatta) of known age are scored for closure of nine bilateral and three unilateral sutures or segments of sutures. A variety of tests reveals a strong relationship between age and stages of suture closure, although increasingly broad confi- dence intervals prevent sutures from being very useful for precisely aging older macaques. The order in which endosutures begin to close, as well as that in which closure is finally achieved, is determined for macaques, and these sequences compared to those for endosutures of humans (Todd and Lyon, 1924). The basilar suture is the earliest to close, while the masto-occipital and rostral and caudal squamosal sutures achieve closure quite late in both species. On the other hand, humans and macaques differ in their schedules for the sphenofrontal suture and in the initiation of closure for the rostral portion of the squamosal suture. Two sutures close significantly sooner on the right than on the left side (the rostral squamosal and masto-occipital) and asymme- try favoring closure of the right lateral lambdoid suture also approaches significance at the 0.05 level. No sutures close significantly sooner on the left side. It is suggested that macaque sutures may close from the inside out, that endosutures are more sensitive than ectosutures for detecting sequences in which cranial sutures begin to close, and that directional asymmetries in suture closure of macaques may be related to minor asymmetries in braid skull shape (petalias).
The use of cranial suture closure to age human skeletal material has had a check- ered history. Numerous workers have raised serious questions about using suture closure as an aging method (Singer, 1953; Brooks, 1955; McKern and Stewart, 1957). However, Meindl and Lovejoy (1985) have recently shown that a combination of segments of ectocranial sutures can contribute valuable information to age estimates. In this report, we explore the relationship between closure of endocranial sutures and age in rhesus monkeys (Macaca mulatta) and compare our results with those for both endocranial (Todd and Lyon, 1924) and ectocranial (Meindl and
Lovejoy, 1985) sutures of humans. The order of suture closure is compared in the two species. Because Zivanovic (1983) has noted a considerable amount of hemispheric asym- metry in suture closure among humans, we tested for directional asymmetry in suture closure in rhesus monkeys. Our findings may shed light on the development of cranial petalias in human (LeMay, 1977; Galaburda et al., 1978) and nonhuman (LeMay et al., 1982) primates.
Received October 19, 1988; revision accepted November 14, 1988.
@ 1989 ALAN R. LISS, INC

418 D. FALK ET AL.
MATERIALS AND METHODS
Latex endocranial casts (endocasts) were prepared by D.F. from 330 skulls of rhesus macaques from Cay0 Santiago. In some anal- yses, sample sizes are reduced from 330 because of missing data. Samples from this same collection have been used to develop aging criteria based on pubic symphysis de- velopment (Rawlins, 1975) and dental erup- tion and epiphyseal fusion (Cheverud, 1981). Animals’ ages at death were known to within at most one year, with most ages at death known to within one week (Sade et al., 1985; Busse, personal communication). Endocasts, whose preparation is described in detail else- where (Radinsky, 1968; Falk, 1978), repro- duce features imprinted on the endocranial surface, including clear representations of sutures. Each endocast was dipped in polyvi- nylacetate for reinforcement, and then sprayed with metallic gold paint to enhance visibility of surface features.
Closure data were collected by RCH for nine bilateral sutures or segments of sutures including the masto-occipital, sphenofron- tal, sphenotemporal, medial coronal, lateral coronal, medial lambdoid, lateral lambdoid, rostral squamosal, and caudal squamosal (called the parieto-occipital by some authors). Three unilateral sutures (or seg- ments) were also scored: the basilar, rostral sagittal, and caudal sagittal (Fig. 1). Most of these features were reproduced on the 330 endocasts, although a few sutures were un- observable on some endocasts.
Fig. 1. Left lateral view of endocast from rhesus monkey skull illustrating the sutures scored in this study. Numbers correspond to sutures listed in Table 3: 1, medial coronal; 2, lateral coronal; 3, rostral sagittal; 4, caudal sagittal; 5, medial lambdoid; 6, lateral lambdoid; 7, masto-occipital; 8, basilar; 9, sphenofrontal; 10, sphe- notemporal; 11, rostral squamosal; 12 caudal squamo- sal.
Suture closure was rated according to a modification of Fredric’s traditional scale after Krogman (1978). Sutures were scored as open (0) if there was sufficient separation of the adjacent cranial bones to permit liquid latex to flow through to the ectocranial sur- face along the entire length of the suture; closing (1) if the interdigitation of adjacent bones prevented passage of the liquid latex along a portion of the suture; closed (2) if interdigitation was complete along the en- tire suture thus blocking the flow of any latex to the ectocranial surface, and at least a portion of the course of the endocranial su- ture margin was still evident from its im- pressions on the inner table; and obliterated (3) if all endocranial traces of the suture had been eliminated by fusion of the endocranial margins.
Intraobserver error was measured by re- peated scoring of all variables on 30 en- docasts. Repeatability for individual sutures (segments) ranged from 90 to loo%, the av- erage being 93.5%. These repeatability scores are in keeping with those determined for single variables in other suture studies (e.g., Meindl and Lovejoy, 1985). We exam- ined the level of asymmetry in macaque suture closure to determine whether bilat- eral sutures can be combined or must be treated separately in an analysis of age re- gression. Although Zivanovic (1983) did not distinguish between fluctuating and direc- tional asymmetry in suture closure, we make this distinction here, as it is central to the question of combining bilateral sutures. Di- rectional asymmetry occures when a trait is typically more developed on one particular side than the other. Fluctuating asymmetry occurs when a trait differs from side to side, with neither side typically showing more development. Even under high levels of fluc- tuating asymmetry it may be possible to substitute an antimere for its partner, but with directional asymmetry an antimere will give biased age estimates using a standard developed on the opposite side.
To examine the lefvright correlations for closure of bilateral sutures we used a com- mon measure from an ordinal contingency table. The lefuright correlations express the relationship between sides, and are de- creased by any fluctuating asymmetry. The ordinal contingency tables are formed by cross tabulating observations on the 330 casts for a left and right suture across the four ordered classes of closure (0, 1, 2, 3). Goodman and Kruskal’s (1954) gamma ( y )

ENDOSUTURES OF MACAQUES 419
can then be used to measure the association between the left and right side. Gamma is defined as:
where C is the number of concordances and D the number of discordances across all pairs of individuals for the left and right suture. As an example of a concordance, if individual i has a score of 1 on the right suture and 2 on the left, then this individual’s ranking on the two sutures will be concordant if compared to an individual j with 0 on the right and 1 on the left. In other words, the rankings are concordant because individual i outranks individual j for each side. In a discordance, the sides rank inversely; for example, an individual scored as 2,l will be discordant with an individual scored as 1,2. Gamma varies from 1 (perfect concordance) to -1 (perfect discordance). “C” and “D’ can be easily calculated from the contingency table as:
C = C C nij nkl i < k j < l
(2) D = C C nij nk l
i < k j > l
where nij and nkl are the cell frequency counts for row i crossed with column j and row k crossed with column 1, respectively. Note that gamma ignores ties. For example, an individual scored as 1,2 will not be counted in the numerator or denominator of gamma if compared to an individual scored as 1,3, because these individuals are tied on the first suture. The significance of gamma can be assessed by dividing C - D by its standard error to produce an approximate z-score. The standard error of C - D is given as (Agresti 1984:180):
where pi+ and p+j are the marginal propor- tions across j and 1, respectively, and where n is the total sample size.
To examine directional asymmetry we used delta (A), a measure of marginal homo- geneity across the contingency table that preserves the ordinal character of suture closure. (One tests for correlations between
right and left sides with gamma, but uses delta to test for significant differences be- tween sides, i.e. sides may be correlated but still be significantly different.) Delta is de- fined as:
A = C C Pi+P+j - C C Pi+P+j (4) i < j i > j
and can be assessed for significance using a z-statistic given in Agresti (1983, 1984:207). Because this z-statistic is only asymptoti- cally normally distributed, we also calcu- lated a probability by randomly reassigning left and right sides, calculating delta, and building a Monte Carlo distribution by re- peated trials (using 999 runs).
With the existence of only a limited amount of fluctuating and directional asym- metry in suture closure (see Results), bilat- eral scorings were collapsed into single vari- ables. Consequently, for each of the nine bilateral sutures, we used the observable side if the other side was unobservable, and randomly selected a side for each individual if both sides were observable. This produced observations on 12 sutures per individual which we analyzed for interrelationships be- tween age and suture closure.
Since initial inspection of scatter plots of age (in months) against suture closure scores illustrated that age increases geometrically with respect to suture closure, logarithmic age was regressed on suture closure in order to derive predictive equations for chronolog- ical age based on suture closure state. Loga- rithmic scale ages correct for the problem of heteroscedasticity, where there is increasing age variation across increasing suture clo- sure states. Using stepwise regression we eliminated the less informative sutures to construct a more parsimonious regression equation. Rather than report the stepwise multiple regression of log age on suture clo- sure, we chose to average the retained su- tures (analogous to Meindl and Lovejoy’s (1985) composite suture scores) and present a simple regression of log age on average suture closure. These results were tested with a jackknife regression analysis (Efron, 1982). Because we are primarily interested in presenting a method for aging older indi- viduals (where dental eruption and epiphy- seal union are not useful), we recalculated the regression using only those animals who were 6 years or older at time of death.
In addition to calculating regressions of log age on suture closure, we examined the
420 D. FALK ET AL.
order in which sutures begin to close and (separately) the order in which closure is achieved. Our sequences for endocranial su- tures of macaques were compared with those for humans (Todd and Lyon, 19241, as well as the initiation and achievement sequences of ectosutures for humans (Meindl and Love- joy, 1985).
We also examined order of suture closure without separating initiation and closure se- quences by forming a matrix that contained counts for the number of times across all endocasts where the closure score for suture i exceeded the state for suture j. We then ordered the rows and columns of the matrix so that elements would increase off the main diagonal in the upper triangle and decrease off the diagonal in the lower triangle. This ordering was determined both by visual in- spection and by forming all possible permu- tations for various submatrices out of the 12 by 12 matrix. Additionally, at this time we tested for sex effects (difference in male/ female intercepts) and sex by suture interac-
tions (differences in malejfemale slopes of age against suture closure) to determine whether there are sex differences in suture closure timing. As sex was not found to be an important factor, we have not included the results of these tests here.
RESULTS
Table 1 contains the summary statistics for righaeft correlations between bilateral sutures. The correlations between the left and right sides are extremely high, suggest- ing that there is relatively little fluctuating asymmetry in suture closure. All correla- tions exceed 0.90 (note that gamma is con- strained between - 1 and + 1) and are highly significant (P < 0.01).
Table 2 contains the summary statistics for directional asymmetry of suture closure. Negative delta values indicate that the right side sutures are in a more advanced state of closure than are the left side. Alternatively, a positive asymmetry score indicates that the left side suture is more advanced. The
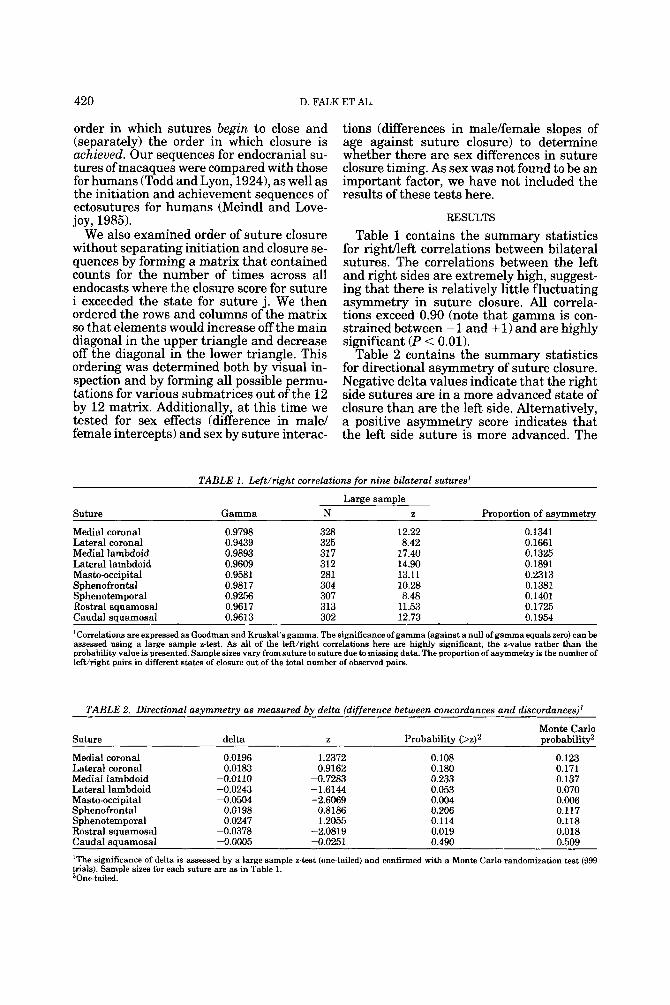
TABLE 1. Left/right correlations for nine bilateral sutures'
Large sample Suture Gamma N z Proportion of asymmetry
Medial coronal 0.9798 328 12.22 0.1341 Lateral coronal 0.9439 325 8.42 0.1661 Medial lambdoid 0.9893 31 7 17.40 0.1325 Lateral lambdoid 0.9609 312 14.90 0.1891 Masto-occipital 0.9581 281 13.11 0.2313 Sphenofrontal 0.9817 304 10.28 0.1381 Sphenotemporal 0.9256 307 8.48 0.1401 Rostral squamosal 0.9617 313 11.53 0.1725 Caudal squamosal 0.9613 302 12.73 0.1954
'Correlations are expressed as Goodman and Kruskal's gamma. The significance of gamma (against a null of gamma equals zero) can be assessed using a large sample z-test. As all of the lefthight correlations here are highly significant, the z-value rather than the probability value is presented. Sample sizes vary from suture to suture due to missing data. The proportion of asymmetry is the number of lefthight pairs in different states of closure out of the total number of observed pairs.
TABLE 2. Directional asymmetry as measured by delta (difference between concordances and discordances)'
Suture delta z Monte Carlo
Probabilitv (>zP Drobabilitv2
Medial coronal 0.0196 1.2372 0.108 0.123 Lateral coronal 0.0183 0.9162 0.180 0.171 Medial lambdoid -0.0110 -0.7283 0.233 0.137 Lateral lambdoid -0.0243 -1.6144 0.053 0.070 Masto-occipital -0.0504 -2.6069 0.004 0.006 Sphenofrontal 0.0198 0.8186 0.206 0.117 Sphenotemporal 0.0247 1.2055 0.114 0.118 Rostral squamosal -0.0378 -2.0819 0.019 0.018 Caudal squamosal -0.0005 -0.0251 0.490 0.509
'The significance of delta is assessed by a large sample z-test (one-tailed) and confirmed with a Monte Carlo randomization test (999 trials). Sample sizes for each suture are as in Table 1. 'One-tailed.

ENDOSUTURES OF MACAQUES 42 1
large sample z-score test indicates that there is a significant amount of directional asym- metry (at P < .05, one-tailed) for the masto- occipital and rostral squamosal sutures, al- though even for these sutures the level of asymmetry is small. Closure of the lateral lambdoid suture is nearly significantly asymmetrical at the 0.05 level (P = 0.053). These findings are substantiated with the Monte Carlo randomization test. For all three sutures the right sides are in a more advanced state of closure than are the left.
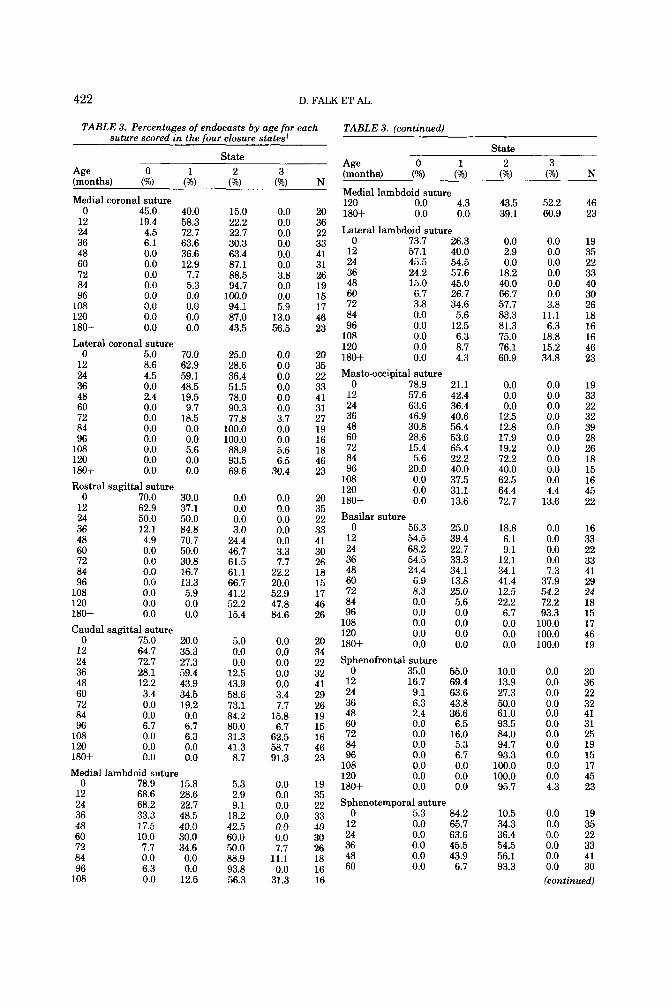
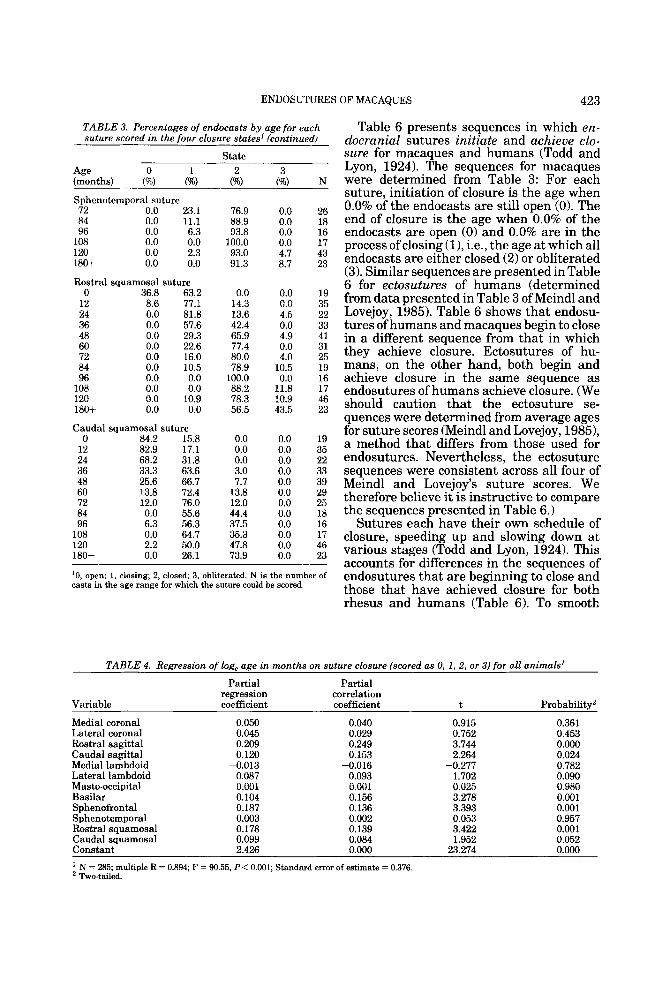
To summarize the progression of suture closure with age, Table 3 provides the per- centage of endocasts for each suture within each state of closure across yearly age inter- vals up to age 10 years. Percentages included for ages 10-15 years (120-180 months) are lumped in the entries for 120 months. All sutures follow an age progression of closure, with the speed of closure varying between sutures. Additionally, it can be determined from Table 3 that while some sutures always reach obliteration in older animals (e.g., the basilar suture), other sutures persist in even the oldest age category (e.g., the caudal squamosal).
Table 4 presents the regression coeffi- cients and summary statistics for a regres- sion of log age (in months) on all 12 sutures for the 285 monkeys who had completely observable sutures. The highly significant (P < 0.001) multiple correlation of 0.894 indi- cates that there is a strong relationship be- tween age and stages of suture closure. The standard error of 0.376 for this regression is on a log scale and thus increases with in- creasing age. Additionally, the confidence interval for age is asymmetrical around the estimated age. For example, at an estimated age of one year (12 months), 8.2 months is one standard error below the estimated age, while 17.5 months is one standard error above. At an estimated age of 10 years, 6.9 years is one standard error below the esti- mated age, while 14.6 years is one standard error above. An additional point from Table 4 is that the sutures are not of uniform impor- tance in age prediction. As the partial regres- sion coefficients (and associated t-tests) in- dicate, some sutures add little to the determination of age.
Although we could present the multiple stepwise regression, it would be impossible to graph the confidence interval. Likewise, because the confidence interval must be cal- culated using the covariance matrix between the sutures, it is unlikely that researchers
would be willing to make such calculations on a case-by-case basis. In any event, the difference between the R2s from the stepwise multiple regression and from the simple re- gression of age on average suture closure was trivial.
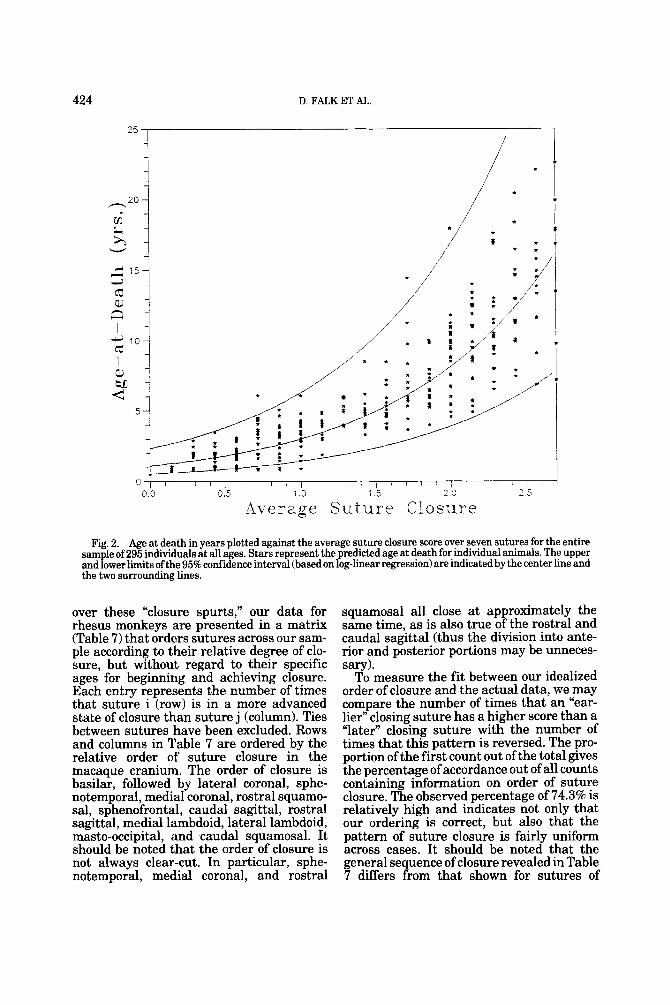
Figure 2 shows the exponential curve and the 95% confidence intervals for prediction of individual cases for the average of seven sutures (segments), including the rostral sagittal, caudal sagittal, lateral lambdoid, basilar, sphenofrontal, rostral squamosal, and caudal squamosal. The equation for Fig- ure 2, based on 295 cases, is log, age = 0.992 (average suture closure) + 2.595. There is a strong and highly significant relationship between log, age and average suture closure (R2 = 0.791, with a standard error of the estimate equal to 0.374 and an increasing uncertainty in age prediction with advanc- ing suture closure. This conclusion was sup- ported by a jacknife regression analysis. The latter analysis showed that the age esti- mates are generally unbiased.
For those animals who were six years or older a t the time of death, stepwise regres- sion was used to select five sutures to aver- age, including the medial coronal, caudal sagittal, basilar, sphenotemporal, and cau- dal squamosal sutures. This analysis, based on 130 individuals, resulted in the equation: log, age = 0.726 (average suture closure) + 3.423 (R2 = 0.46; SE of estimate = 0.252). Although as one would expect, there is less heteroscedasticity in the data, the confi- dence interval remained quite broad (see Fig. 3). Even so, estimates of older animals should be made using this second equation due to its smaller standard error of the esti- mates.
The durations of closure for specific endo- cranial sutures are listed for rhesus mon- keys and humans in Table 5. The data for rhesus monkeys were determined from Ta- ble 3; those for humans were taken from Todd and Lyon (1924). It is noteworthy that the sphenofrontal suture takes over ten times as long to close in humans as it does in rhesus monkeys. (It is difficult to compare the masto-occipital because it is not known when this suture reaches closure in macaques.) There is a long duration of clo- sure in the sphenofrontal suture of humans (see Table 6). The rostral squamosal of hu- mans, on the other hand, begins to close much later in life than it does in rhesus monkeys, in whom rostral and caudal por- tions have distinct schedules (Table 6).
422 D. FALK ET AL.
TABLE 3. Percentages of endocasts by age for each suture scored in the four closure states'
TABLE 3. (continued)
State 2
(%)
State 2
(%)
Age 0 1 (months) (%) (96)
Medial lambdoid suture 120 0.0 4.3 180+ 0.0 0.0 Lateral lambdoid suture
0 73.7 26.3 12 57.1 40.0 24 45.5 54.5 36 24.2 57.6 48 15.0 45.0 60 6.7 26.7 72 3.8 34.6 84 0.0 5.6 96 0.0 12.5
108 0.0 6.3 120 0.0 8.7 180f 0.0 4.3 Masto-occipital suture
0 78.9 21.1 12 57.6 42.4 24 63.6 36.4 36 46.9 40.6 48 30.8 56.4 60 28.6 53.6 72 15.4 65.4 84 5.6 22.2 96 20.0 40.0
108 0.0 37.5 120 0.0 31.1 180f 0.0 13.6 Basilar suture
0 56.3 25.0 12 54.5 39.4 24 68.2 22.7 36 54.5 33.3 48 24.4 34.1 60 6.9 13.8 72 8.3 25.0 84 0.0 5.6 96 0.0 0.0
108 0.0 0.0 120 0.0 0.0 180+ 0.0 0.0 Sphenofrontal suture
0 35.0 55.0 12 16.7 69.4 24 9.1 63.6 36 6.3 43.8 48 2.4 36.6 60 0.0 6.5 72 0.0 16.0 84 0.0 5.3 96 0.0 6.7
108 0.0 0.0 120 0.0 0.0 180f 0.0 0.0 Sphenotemporal suture
0 5.3 84.2 12 0.0 65.7 24 0.0 63.6 36 0.0 45.5 48 0.0 43.9 60 0.0 6.7
Age 0 1 (months) (%) (%)
Medial coronal suture 0 45.0 40.0
12 19.4 58.3 24 4.5 72.7 36 6.1 63.6 48 0.0 36.6 60 0.0 12.9 72 0.0 7.7 84 0.0 5.3 96 0.0 0.0
108 0.0 0.0 120 0.0 0.0 180f 0.0 0.0 Lateral coronal suture
0 5.0 70.0 12 8.6 62.9 24 4.5 59.1 36 0.0 48.5 48 2.4 19.5 60 0.0 9.7 72 0.0 18.5 84 0.0 0.0 96 0.0 0.0
108 0.0 5.6 120 0.0 0.0 180f 0.0 0.0 Rostra1 sagittal suture
0 70.0 30.0 12 62.9 37.1 24 50.0 50.0 36 12.1 84.8 48 4.9 70.7 60 0.0 50.0 72 0.0 30.8 84 0.0 16.7 96 0.0 13.3
108 0.0 5.9 120 0.0 0.0 180f 0.0 0.0 Caudal sagittal suture
0 75.0 20.0 12 64.7 35.3 24 72.7 27.3 36 28.1 59.4 48 12.2 43.9 60 3.4 34.5 72 0.0 19.2 84 0.0 0.0 96 6.7 6.7
108 0.0 6.3 120 0.0 0.0 180f 0.0 0.0 Medial lambdoid suture
0 78.9 15.8 12 68.6 28.6 24 68.2 22.7 36 33.3 48.5 48 17.5 40.0 60 10.0 30.0 72 7.7 34.6 84 0.0 0.0 96 6.3 0.0
108 0.0 12.5
43.5 39.1
52.2 46 60.9 23 15.0 0.0
0.0 0.0 0.0 0.0 0.0 3.8 0.0 0.0 5.9
13.0 56.5
20 36 22 33 41 31 26 19
22.2 22.7 30.3 63.4
0.0 2.9 0.0
18.2 40.0 66.7 57.7 83.3 81.3 75.0 76.1 60.9
0.0 19 0.0 35 0.0 22 0.0 33 0.0 40 0.0 30 3.8 26
11.1 18 6.3 16
18.8 16
87.1 88.5 94.7 100.0 94.1 87.0
~~
15 17 46 23 43.5
15.2 46 34.8 23 25.0
28.6 36.4 51.5 78.0 90.3 77.8
100.0 100.0 88.9 93.5 69.6
0.0 0.0 0.0 0.0 0.0 0.0 3.7 0.0 0.0 5.6 6.5
30.4
20 35 22
0.0 0.0 0.0
12.5 12.8 17.9 19.2 72.2 40.0
0.0 19 0.0 33 0.0 22 0.0 32 0.0 39 0.0 28 0.0 26 0.0 18 0.0 15 0.0 16 4.4 45
13.6 22
33 41 31 27 19 16 18 46 23
62.5 64.4 72.7 0.0
0.0 0.0 3.0
24.4 46.7 61.5 61.1 66.7 41.2 52.2 15.4
0.0 0.0 0.0 0.0 0.0
20 35 22
18.8 6.1 9.1
12.1 34.1 41.4 12.5 22.2 6.7 0.0 0.0 0.0
0.0 16 0.0 33 0.0 22 0.0 33 7.3 41
37.9 29 54.2 24 72.2 18 93.3 15
100.0 17 100.0 46 100.0 19
33 41 30 26 18 15 17 46 26
3.3 7.7
22.2 20.0 52.9 47.8 84.6
5.0 0.0 0.0
0.0 0.0 0.0 0.0
20 34 22 32 41
10.0 0.0 20 0.0 36 0.0 22 0.0 32 0.0 41 0.0 31 0.0 25 0.0 19 0.0 15 0.0 17 0.0 45 4.3 23
12.5 43.9
...
13.9 27.3 50.0 61.0 93.5
0.0 3.4 7.7
15.8
58.6 73.1 84.2
29 26 19 15 16 46 23
80.0 31.3 41.3 8.7
6.7 62.5 58.7 91.3
84.0 94.7 93.3
100.0 100.0 95.7 5.3
2.9 9.1
18.2 42.5 60.0 50.0 88.9 93.8 56.3
0.0 0.0 0.0
19 35 22
10.5 34.3 36.4 54.5 56.1 93.3
0.0 19 0.0 35 0.0 22 0.0 33 0.0 41 0.0 30 (continued)
0.0 0.0 0.0 7.7
11.1 0.0
31.3
33 40 30 26 18 16 16
423 ENDOSUTURES OF MACAQUES
TABLE 3. Percentages of endocasts by age for each suture scored in the four closure states' (continued)
State Age 0 1 2 3 (months) (W) (W) (W) (%) N Sphenotemporal suture 72 0.0 23.1 76.9 0.0 26 84 0.0 11.1 88.9 0.0 18 96 0.0 6.3 93.8 0.0 16
108 0.0 0.0 100.0 0.0 17 120 0.0 2.3 93.0 4.7 43 180+ 0.0 0.0 91.3 8.7 23
Rostral squamosal suture 0 36.8 63.2
12 8.6 77.1 24 0.0 81.8 36 0.0 57.6 48 0.0 29.3 60 0.0 22.6 72 0.0 16.0 84 0.0 10.5 96 0.0 0.0
108 0.0 0.0 120 0.0 10.9 180+ 0.0 0.0
0.0 14.3 13.6 42.4 65.9 77.4 80.0 78.9
100.0 88.2 78.3 56.5
0.0 19 0.0 35 4.5 22 0.0 33 4.9 41 0.0 31 4.0 25
10.5 19 0.0 16
11.8 17 10.9 46 43.5 23
Caudal squamosal suture 0 84.2 15.8 0.0 0.0 19
12 82.9 17.1 0.0 0.0 35 24 68.2 31.8 0.0 0.0 22 36 33.3 63.6 3.0 0.0 33 48 25.6 66.7 7.7 0.0 39 60 13.8 72.4 13.8 0.0 29 72 12.0 76.0 12.0 0.0 25 84 0.0 55.6 44.4 0.0 18 96 6.3 56.3 37.5 0.0 16
108 0.0 64.7 35.3 0.0 17 120 2.2 50.0 47.8 0.0 46 180+ 0.0 26.1 73.9 0.0 23
'0, open; 1, closing; 2, closed; 3, obliterated. N is the number of casts in the age range for which the suture could be scored.
Table 6 presents sequences in which en- docranial sutures initiate and achieve clo- sure for macaques and humans (Todd and Lyon, 1924). The sequences for macaques were determined from Table 3: For each suture, initiation of closure is the age when 0.0% of the endocasts are still open (0). The end of closure is the age when 0.0% of the endocasts are open (0) and 0.0% are in the process of closing (11, i.e., the age at which all endocasts are either closed (2) or obliterated (3). Similar sequences are presented in Table 6 for ectosutures of humans (determined from data presented in Table 3 of Meindl and Lovejoy, 1985). Table 6 shows that endosu- tures of humans and macaques begin to close in a different sequence from that in which they achieve closure. Ectosutures of hu- mans, on the other hand, both begin and achieve closure in the same sequence as endosutures of humans achieve closure. (We should caution that the ectosuture se- quences were determined from average ages for suture scores (Meindl and Lovejoy, 19851, a method that differs from those used for endosutures. Nevertheless, the ectosuture sequences were consistent across all four of Meindl and Lovejoy's suture scores. We therefore believe it is instructive to compare the sequences presented in Table 6.)
Sutures each have their own schedule of closure, speeding up and slowing down at various stages (Todd and Lyon, 1924). This accounts for differences in the sequences of endosutures that are beginning to close and those that have achieved closure for both rhesus and humans (Table 6). To smooth
TABLE 4. Regression of log, age in months on suture closure (scored as 0, 1,2, or 3) for all animals'
Partial Partial regression correlation
Variable coefficient coefficient t Probability2
Medial coronal Lateral coronal Rostral sagittal Caudal sagittal Medial lambdoid Lateral lambdoid Masto-occipital Basilar Sphenofrontal Sphenotemporal Rostral squamosal Caudal squamosal Constant
0.050 0.045 0.209 0.120
-0.013 0.087 0.001 0.104 0.187 0.003 0.178 0.099 2.426
0.040 0.029 0.249 0.153
-0.016 0.093 0.001 0.156 0.136 0.002 0.139 0.084 0.000
0.915 0.752 3.744 2.264
-0.277 1.702 0.025 3.278 3.393 0.053 3.422 1.952
23.274
0.361 0.453 0.000 0.024 0.782 0.090 0.980 0.001 0.001 0.957 0.001 0.052 0.000
' N = 285; multiple R = 0.894; F = 90.55, P < 0.001; Standard error of estimate = 0.376 Two-tailed.
424 D. FALK ET AL.
I 25 ,
/ = :
: / i i i -
O , ’ , ’ , 1 1 1 I I 1 I I l l ’ ~ l l l ~ ~ l l
0 9 0 5 1 0 1 5 2 0 25
Average Suture Closure
Fig. 2. Age at death in years plotted against the average suture closure score over seven sutures for the entire Sam le of 295 individuals at all ages. Stars represent the predicted age at death for individual animals. The upper and Power limits of the 95% confidence interval (based on loglinear regression) are indicatedby the center line and the two surrounding lines.
over these “closure spurts,” our data for rhesus monkeys are presented in a matrix (Table 7) that orders sutures across our sam- ple according to their relative degree of clo- sure, but without regard to their specific ages for beginning and achieving closure. Each entry represents the number of times that suture i (row) is in a more advanced state of closure than suture j (column). Ties between sutures have been excluded. Rows and columns in Table 7 are ordered by the relative order of suture closure in the macaque cranium. The order of closure is basilar, followed by lateral coronal, sphe- notemporal, medial coronal, rostral squamo- sal, sphenofrontal, caudal sagittal, rostral sagittal, medial lambdoid, lateral lambdoid, masto-occipital, and caudal squamosal. It should be noted that the order of closure is not always clear-cut. In particular, sphe- notemporal, medial coronal, and rostral
squamosal all close at approximately the same time, as is also true of the rostral and caudal sagittal (thus the division into ante- rior and posterior portions may be unneces- sary).
To measure the fit between our idealized order of closure and the actual data, we may compare the number of times that an “ear- lier” closing suture has a higher score than a ’later” closing suture with the number of times that this pattern is reversed. The pro- portion of the first count out of the total gives the percentage of accordance out of all counts containing information on order of suture closure. The observed percentage of 74.3% is relatively high and indicates not only that our ordering is correct, but also that the pattern of suture closure is fair1 uniform across cases. It should be note B that the general sequence of closure revealed in Table 7 differs from that shown for sutures of
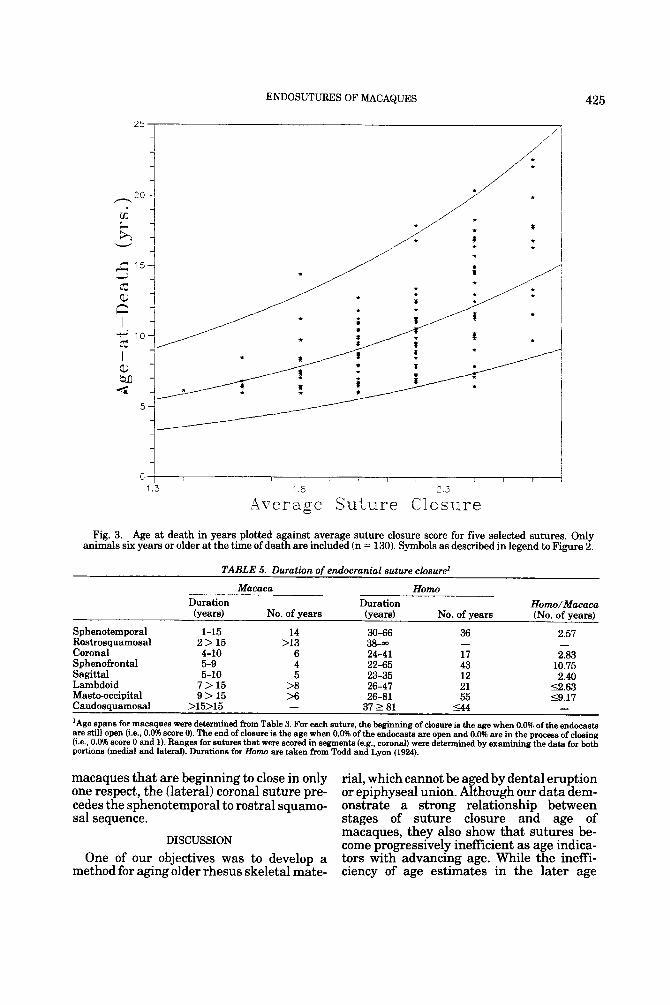
ENDOSUTURES OF MACAQUES 425
0 , I I 1 I I I I I 1
1 3 ' 6 2 3
-4verage Suture Closure
Fig. 3. Age at death in years plotted against average suture closure score for five selected sutures. Only animals six years or older at the time of death are included (n = 130). Symbols as described in legend to Figure 2.
TABLE 5. Duration of endocranial suture closure'
Macaca Homo HomdMacaca
(years) No. of years (years) No. of years (No. of years) Duration Duration
Sphenotemporal Rostrosquamosal Coronal Sphenofrontal Sagittal Lambdoid Masto-occipital Caudosquamosal
1-15 2>15 4-10 5-9 5-10 7 > 15 9 > 15
>15>15
14 30-66 36 2.57
6 24-41 17 2.83 4 22-65 43 10.75 5 23-35 12 2.40
>8 26-47 21 12.63 >6 26-81 55 59.17 - 37 2 81 5 4 4 -
- >I3 38- -
'Age spans for macaques were determined from Table 3. For each suture, the beginning of closure is the age when 0.0% of the endocasta are still open (i.e., 0.0% score 0). The end of closure is the age when 0.0% of the endocasta are open and 0.0% are in the process of closing (i.e., 0.0% score 0 and 1). Ranges for sutures that were scored in segments (e.g., coronal) were determined by examining the data for both portions (medial and lateral). Durations for Homo are taken from Todd and Lyon (1924).
macaques that are beginning to close in only rial, which cannot be aged by dental eruption one respect, the (lateral) coronal suture pre- or epiphyseal union. Although our data dem- cedes the sphenotemporal to rostra1 squamo- onstrate a strong relationship between sal sequence. stages of suture closure and age of
macaques, they also show that sutures be- come progressively inefficient as age indica- tors with advancing age. While the ineffi- ciency of age estimates in the later age
DISCUSSION
One of our objectives was to develop a method for aging older rhesus skeletal mate-

426 D. FALK ET AL.
TABLE 6. Order of suture closure in humans and macaques’
Seauence
Macaca endosutures Initiation sptem-r/sq-cor-(spfro sag)-lb-masoc-c/sq Closure spfro-(cor sag)-sptem-(lb masoc r/sq c/sq)
Initiation spfro-sag-cor-(lb masoc)-sptem-c/sq-r/sq Closure sag - cor- lb - spfro -sptem -masoc- c/sq -r/sq
Initiation sag-cor-lb-spfro-sptem Closure sag-cor-lb-spfro-sptem
Homo endosutures
Homo ectosutures
’Sequences in which cranial sutures begin to close (initiation) and those in which they achieve closure. Endosuture sequences for macaques and humans (Todd and Lyon, 1924) determined from Table 5. The sequences for human ectosutures determined from Meindl and Lovejoy’s (1985) Table 3 (counting their point 7, pterion, a s the lateral coronal) remained the same across all four of their suture scores. Although the methods vaned by which authors determined and quantified ages for suture closure, the overall sequences seem fairly comparable. Abbreviations: c/sq, caudal squamosal; cor, coronal; lb, lambdoid; masoc, masto-occipital; r/sq, rostra1 squamosal; sag, sagittal; spfro, sphenofrontal; sptem, sphenotemporal. Sutures contained in parentheses closed at the same time.
TABLE 7. Matrix of relationshim between suture closure states’
8
50 60 43
6 55 7 29
12 26
2
138 0
33 36 41 24 65 59 45 24 9 3
10
140 44 0
49 47 29 72 65 59 28 7 0
1
134 54 55 0
52 41 55 47 48 29 10 3
11
136 61 54 54
0 45 68 59 50 28 5
9
150 57 51 57 60 0
78 69 63 39 12 6
4
113 109 105 91
101 89 0
31 42 50 26 24
3
121 123 114 98
107 97 39 0
53 56 31 18
__
__
5
134 121 119 112 112 108 70 78 0
45 31 23
6 7
155 181 121 186 112 175 109 179 109 171 100 161 95 149 95 156 57 131 0 118
17 0 16 52
12
196 222 210 209 220 203 176 171 165 148 81 0
-
- ’Each entry represents the number of times that suture i (row) is in a more advanced state of closure that suture j (column). The rows and columns are ordered from earliest to latest closing sutures, with numbering of sutures following that given in Table 3.
intervals is disturbing, it is important to note that this is not a problem unique to suture closure. Rather, all age indicators become progressively inefficient with advancing age due to the divergence between individuals with different biological aging rates. Biolog- ical aging rates may vary due to genetic and environmental factors such as nutrition. Be- cause of the broad confidence intervals asso- ciated with our analyses, we must conclude that cranial sutures (endocasts) are not very useful for precisely aging older macaques. However, cranial suture closure can be used with caution as an age estimator in macaques.
Rawlins (1975) investigated age-related changes in the pubic symphysis of subadult and adult macaques using a subset of the sample employed here. He found symphy- seal morphology to be of little use in aging adult females because of extensive remodel- ing accompanying parturition. Limited esti- mates of age could be made in adult males using the extent of symphyseal fusion. How- ever, Rawlins (1975) noted the serious need
for alternative aging methods €or adult pri- mates. While endocranial sutural fusion cannot, by itself, provide very precise age estimates for adult macaque skulls (see Figs. 2, 3), it is the best currently available method.
From a comparison of Meindl and Love- joy’s (1985) Table 3 with our Table 5 , it appears that human endosutures begin clos- ing at ages that are five or more years younger than the average ages at which their ectocranial portions are reported to still be open. Thus cranial sutures of humans may close from the inside out, as suggested by Acsadi and Nemeskeri (1970:116): “ossifi- cation of sutures begins inwardly and spreads outwardly.”
Meindle and Lovejoy (1985) claim that ectocranial sutures are superior to endocra- nial sutures for estimating age in humans because the mean deviations they obtained in ectocranial scoring of combined vault su- tures for various ages are approximately half of the mean deviations found by Acsadi and Nemeskeri (1970) for their combined endo-

ENDOSUTURES OF MACAQUES 427
cranial vault sutures. However, the former authors combined seven small lengths of suture (“sites”) from the lambdoid, sagittal, and coronal sutures in their vault calcula- tions whereas the latter workers used the overall degree of suture closure for coronal, sagittal, and lambdoid sutures (i.e., three variables) in their calculations. Thus, it is not clear that the variables used to represent vault sutures in the two studies differ only in regard to their externallinternal locations on the skull. Hence, mean deviations from these two studies do not appear to be comparable and therefore should not be used to draw conclusions about which type of suture is the best predictor of age.
As shown in Table 6, endosutures of hu- mans may be more sensitive than ectosu- tures for detecting details of the initiation of suture closure. That is, ectosutures that are beginning to close reflect the closing se- quence of endosutures, but fail t o reveal the actual sequence in which cranial sutures begin to close ke. , the initiation sequence for endosutures). For instance, if one relied only on ectosutures, the important role of the sphenofrontal suture in human cranial growth would not be detected.
The pattern of endosuture closure in the macaque can be compared to that for hu- mans to determine whether a general pri- mate pattern exists (Table 6). There are some broad similarities in pattern between the two species, with macaques and humans being more similar in the sequences in which their cranial sutures achieve closure, than they are in the sequences in which their sutures begin to close. For example, the basi- lar suture is the earliest closing suture for both humans (Krogman, 1978) and macaques, while the masto-occipital and ros- tral and caudal squamosal sutures (and to an extent, the sphenotemporal) achieve closure quite late in both species.
Humans and macaques differ dramati- cally in their schedules for the sphenofrontal endosuture. The sphenofrontal suture of hu- mans is the first (of those listed in Table 6) to begin closing, but the fourth to achieve clo- sure. This accounts for the fact that it takes over ten times as long to close in humans as it does to close in rhesus monkeys (Table 5). It is tempting to speculate that this difference may be related to expansion of the frontal lobes and/or temporal poles in Homo relative to Macaca.
Although the squamosal endosuture is the last to close in both humans and macaques, its rostral portion begins to close early only
in the latter species. This difference is diffi- cult to interpret. It may simply be that the rostral squamosal is “riding along” with the sphenotemporal (its neighbor) in the initia- tion sequence of macaques. It is interesting to note that sulci from occipital regions of the cerebral cortex generally do not reproduce well on endocasts from macaques (or other monkeys) (Falk, 1978). Poor reproducibility of occipital sulci may be related to the late initiation and closure schedules for lamb- doid, masto-occipital, and caudal squamosal sutures for macaques. (For reasons that are not entirely clear, endocasts prepared from human skulls reproduce overall poorer sul- cal patterns than do endocasts from monkey skulls.)
Our finding that rhesus monkeys lack sex- ual dimorphism in timing of suture closure mirrors numerous reports concerning cra- nial sutures of humans (Todd and Lyon, 1924; Meindl and Lovejoy, 1985; Acsadi and Nemeskeri, 1970). It may be that this is the general primate pattern, or that sexual di- morphism in closure of some sutures is too subtle to have been detected in this study.
It is important to note that each cast was scored at one sitting. Consequently, the right and left side observations were not made independently, with the result that error may have been introduced to the determina- tion of fluctuating asymmetry for this sam- ple. The proportion of endocasts showing asymmetrical closure is generally lower than the figures reported by Zivanovic (1983) for humans. It is, however, difficult to interpret these results, as Zivanovic does not state whether bilateral sutures were scored inde- pendently within his sample. Additionally, proportion of asymmetrical cases is not a very telling measure, as it does not take into account the ordinal nature of suture closure. For example, a suture scored as 0 on the left and 3 on the right is considered in the same category with a suture scored 0 on the left and 1 on the right by Zivanovic (19831, but not here.
With this in mind, one ofthe most interest- ing findings of this study is that there is a small but statistically significant directional asymmetry in closure of the masto-occipital and rostral squamosal sutures of macaques, with right sides preceding the left. (Asym- metry favoring closure of the right lateral lambdoid suture also approaches signifi- cance at the 0.05 level.) Although these su- tures do display significant asymmetry in closure, we still chose to collapse all bilateral suture scores into single sutures. We sug-

428 D. FALK ET AL.
gest, however, that both sides of a cranium should be scored to determine whether the sides are symmetrical.
Because the caudal squamosal does not begin to close in rhesus monkeys until after 15 years (Table 51, we are unable to deter- mine whether or not closure in this portion of the squamosal is also asymmetrical. This would not be surprising however, since the caudal squamosal forms a complex with the other three sutures.
Asymmetry in premature closure of the lambdoid suture occurs infrequently as a clinical condition (lambdoid craniosynosto- sis) in humans (Furuya et al., 1984; Hinton et al., 1984; Muakkassa et al., 1984). Inter- estingly, more cases of lambdoid cranio- synostosis involve early fusion of the right than the left lambdoid suture and are associ- ated with “flattening of the occipital bone on the affected side, compensatory bulging of the anterior ipsilateral portion of the calva- ria.. . .” (Furuya et al., 1984:63). Thus, shape features that characterize the more prevalent right lambdoid craniosynostosis are associated with right frontal and left occipital projections (relative to their coun- terparts). Less dramatic and nonpathologi- cal braidskull asymmetries called “petalias” occur in similar directions in human popula- tions and are statistically associated with right-handedness (LeMay, 1977; Galaburda et al., 1978; LeMay et al., 1982). Shape as- symetries that are less striking but in a similar direction to the right frontal and left occipital petalias that are modal for human skulldbrains have been documented for en- docasts of Old World monkeys (LeMay et al., 1982). Although we are not suggesting that the asymmetries we report for suture closure in macaques are examples of craniosynosto- sis, our findings do suggest the possibility that cranial petalias of primates (including humans) may occur in conjunction with asymmetrical suture closure. If so, asymme- tries of the skull may be due to differences in timing of developmental events on the two sides.
ACKNOWLEDGMENTS We thank Jim Neely for commenting on
the manuscript, and the University of Puerto Rico is acknowledged for providing free ac- cess to the Cay0 Santiago skeletal collection. This research is supported by Public Health Service grant 7 R01 NS24904.
LITERATURE CITED
Acsadi G, and Nemeskeri J (1970) History of Human Life
Span and Mortality. Budapest: Hungarian Academic Society.
Agresti A (1983) Testing marginal homogeneity for ordi- nal categorical variables. Biometrics 39:505-510.
Agresti A (1984) Analysis of Ordinal Categorical Data. New York John Wiley and Sons.
Brooks ST (1955) Skeletal age at death: The reliability of cranial and pubic age indicators. Am. J. Phys. Anthro-
Cheverud J (1981) Epiphyseal fusion and dental erup- tion in Macaca mulatta. Am. J . Phys. Anthropol.
Efron B (1982) The Jackknife, the Bootstrap and Other Resampling Plans. Philadelphia: The Society for In- dustrial and Applied Mathematics.
Falk D (1978) External neuroanatomy of Old World monkeys (Cercopithecoidea). Contrib. Primatol.
Furuya Y, Edwards MSB, Alpers CE, Tress BM, Norman D, and Ousterhout DK (1984) Computerized tomogra- phy of cranial sutures. J. Neurosurg. 61:59-70.
Galaburda AM, LeMay M, Kemper TL, and Geschwind N (1978) Right-left asymmetries in the brain. Science 199:852-856.
Goodman LA, and Kruskal WH (1954) Measures of association for cross-classification. J. Am. Statist. As- SOC. 49:73%764.
Hinton DR, Becker LE, Muakkassa KF, and Hoffman HJ (1984) Lambdoid synostosis Part 1: The lambdoid suture: Normal development and pathology of “synos- tosis.” J. Neurosurg. 61:333-339.
Krogman WM (1978) The Human Skeleton in Forensic Medicine. Springfield, IL: Charles C. Thomas.
LeMay M (1977) Asymmetries of the skull and handed- ness. Phrenology revisited. J. Neurol. Sci. 32:243-253.
LeMay M, Billig MS, and Geschwind N (1982) Asymme- tries of the brains and skulls of nonhuman primates. In E Armstrong and D Falk (eds.): Primate Brain Evolution: Methods and Concepts. New York: Plenum,
McKern SW, and Stewart TD (1957) Skeletal Age Changes in Young American Males: Analysed from the Standpoint of Age Identification. Natick, MA: Head- quarters Q.M. Res. and Dev. Command, Tech. Rep.,
Meindl RS, and Lovejoy CO (1985) Ectocranial suture closure: A revised method for the determination of skeletal age at death based on the lateral-anterior sutures. Am. J. Phys. Anthropol. 68:5746.
Muakkassa KF, Hoffman HJ, Hinton DR, Hendrick EB, Humphreys RP, and Ash J (1984) Lambdoid synosto- sis Part 2: Review of cases managed at The Hospital for Sick Children, 1972-1982. J. Neurosurg. 61:340-347.
Radinsky LB (1968) A new approach to mammalian cranial analysis, illustrated by examples of prosimian primates. J . Morphol. 124:167-180.
Rawlins R (1975) Age changes in the pubic symphysis of Macaca mulatta. Am. J. Phys. Anthropol. 42:477-488.
Sade DS, Chepko-Sade BD, Schneider JM, Roberts FS, and Richtsmeier JT (1985) Basic Demographic Obser- vations on Free-Ranging Rhesus Monkeys. New Ha- ven, CT: Human Relations Area Files.
Singer R (1953) Estimation of age from cranial suture closure. J. Forensic Med. 1:52-59.
Todd TW, and Lyon J r DW (1924) Endocranial suture closure. Part I. Adult males of white stock. Am. J. Phys. Anthropol. 7:325-384.
Zivanovic S (1983) A note on the effect of asymmetry in suture closure in mature human skulls. Am. J. Phys. Anthropol. 60:431435.
pol. 13:567-589.
56:157-168.
15:l-95.
pp. 263-271.
EP-45.




















