
Distribution of cortical neurons projecting to the superior colliculus in macaque monkeys
Upload
royaninstituteCategory
view
1download
0
Cytotherapy, 2014; 16: 734e749
ORIGINAL PAPERS
Intra-renal arterial injection of autologous bone marrow mesenchymalstromal cells ameliorates cisplatin-induced acute kidney injury in arhesus Macaque mulatta monkey model
REZA MOGHADASALI1,2,3, MAHNAZ AZARNIA1, MOSTAFA HAJINASROLLAH2,HASSAN ARGHANI4, SEYED MAHDI NASSIRI5, MOHAMMAD MOLAZEM6,AHMAD VOSOUGH7, SOROUSH MOHITMAFI2,7, MOSTAFA NAJARASL2,ZAHRA AJDARI2, REZA SALMAN YAZDI8, MOHSEN BAGHERI8,HOSSEIN GHANAATI9, BEHROOZ RAFIEI9, YOUSOF GHEISARI10,HOSSEIN BAHARVAND2,3,11 & NASSER AGHDAMI2,3
1Department of Biology, Kharazmi University, Tehran, Iran, 2Department of Stem Cells and Developmental Biologyand 3Department of Regenerative Medicine at the Cell Science Research Center, Royan Institute for Stem Cell Biologyand Technology, ACECR, Tehran, Iran, 4Urology and Nephrology Research Center, Modarres Hospital, ShahidBeheshti University of Medical Sciences, Tehran, Iran, 5Department of Clinical Pathology and 6Department ofVeterinary Radiology, Faculty of Veterinary Medicine, University of Tehran, Tehran, Iran, 7Department of ClinicalSciences, Faculty of Veterinary Medicine, Karaj Branch, Islamic Azad University, Karaj, Iran, 8Department ofAndrology at the Reproductive Biomedicine Center, Royan Institute for Reproductive Biomedicine, ACECR, Tehran,Iran, 9Advanced Diagnostic and Interventional Radiology Research Center (ADIR), Tehran University of MedicalSciences, Tehran, Iran, 10Department of Genetics and Molecular Biology, Isfahan University of Medical Sciences,Isfahan, Iran, and 11Department of Developmental Biology, University of Science and Culture, ACECR, Tehran, Iran
AbstractBackground. Clinically, acute kidney injury (AKI) is a potentially devastating condition for which no specific therapy im-proves efficacy of the repair process. Bone marrow mesenchymal stromal cells (BM-MSCs) are proven to be beneficial forthe renal repair process after AKI in different experimental rodent models, but their efficacy in large animals and humansremains unknown. This study aims to assess the effect of autologous rhesus Macaque mulatta monkey BM-MSC trans-plantation in cisplatin-induced AKI. Methods. We chose a model of AKI induced by intravenous administration of 5 mg/kgcisplatin. BM-MSCs were transplanted through intra-arterial injection. The animals were followed for survival, biochemistryanalysis and pathology. Results. Transplantation of 5 � 106 cells/kg ameliorated renal function during the first week, as shownby significantly lower serum creatinine and urea values and higher urine creatinine and urea clearance without hyponatremia,hyperkalemia, proteinuria and polyuria up to 84 d compared with the vehicle and control groups. The superparamagneticiron oxide nanoparticle-labeled cells were found in both the glomeruli and tubules. BM-MSCs markedly acceleratedFoxp3þ T-regulatory cells in response to cisplatin-induced damage, as revealed by higher numbers of Foxp3þ cells withinthe tubuli of these monkeys compared with cisplatin-treated monkeys in the control and vehicle groups. Conclusions. Thesedata demonstrate that BM-MSCs in this unique large-animal model of cisplatin-induced AKI exhibited recovery andprotective properties.
Key Words: acute kidney injury, bone marrow mesenchymal stromal cells, cisplatin, intra-renal arterial injection
Introduction
Acute kidney injury (AKI) in humans is a proteansyndrome of varying severity that primarily resultsfrom ischemic and/or toxic injury. AKI occurs in upto 5% of hospitalized patients (1) and is a major
Correspondence: Nasser Aghdami, MD, PhD, Department of Stem Cells and Dfor Stem Cell Biology and Technology, ACECR, Tehran, Iran. E-mail: Nasser.A
(Received 18 July 2013; accepted 8 January 2014)
ISSN 1465-3249 Copyright � 2014, International Society for Cellular Therapy. Phttp://dx.doi.org/10.1016/j.jcyt.2014.01.004
cause of morbidity and mortality (2). Administrationof cisplatin, an anticancer agent, is accompanied by ahigh incidence of nephrotoxicity and renal dysfunc-tion. Currently, most renal-protective approachesfor AKI are only partially beneficial, and patients
evelopmental Biology at the Cell Science Research Center, Royan [email protected]
ublished by Elsevier Inc. All rights reserved.

BM-MSCs ameliorate AKI in monkey 735
either remain on dialysis or progress to chronic kid-ney disease (3).
Recent studies have tested stem cellebased ther-apy, which provides the exciting prospect of apowerful treatment to repair acute organ damageby virtue of the unique stem cell tropism and pro-regenerative capacity (4). Transplantation of humanbone marrow mesenchymal stromal cells (BM-MSCs) in cisplatin-induced AKI has been shown toimprove disease symptoms (5,6), decrease cisplatinnephrotoxicity in vitro and in vivo and enhancesurvival of mice with AKI (7).
Most studies have been conducted on AKImodels that used BM-MSCs from rodents. There-fore, we believe that the development of alternativelarge-animal models would benefit the evaluation ofstem cell therapeutic potential in the kidneys.
We developed a model of cisplatin-inducednephrotoxic injury in the adult rhesus Macacamulatta monkey that presented with a typical patternof AKI injury that closely mimicked AKI in humans.Furthermore, we have shown that transplanted BM-MSCs can engraft at the site of injury and amelioratethe outcome of AKI.
Methods
Cell culture and characterization of BM-MSCs
Bone marrow aspirates (3e5 mL) were collectedfrom the monkeys’ iliac crests after animals wereanesthetized with intramuscular injections of keta-mine (10 mg/kg) and xylazine (0.4 mg/kg). Weisolated mononuclear cells (MNCs) with the useof lymphodex (gravity: 1.077e1.080; Inno-TrainDiagnostik, Kronberg, Germany). MNCs werewashed and seeded at 1 � 106 cells/cm2 in MinimumEssential Medium Eagle Alpha Modification (LifeTechnologies, Grand island, NY, USA) mediumthat contained 10% fetal bovine serum (FBS;Hyclone, Logan, UT, USA), 1% L-glutamine (LifeTechnologies) and 1% penicillin-streptomycin (pen/strep) liquid (Life Technologies). Mesenchymal po-tential of MSCs was confirmed by adipogenic, oste-ogenic and chondrogenic differentiation.
We used the fibroblast-like colony-forming unitassay to evaluate primary cells grown on a tissue cul-ture dish. Total BM-MNCswere plated at a density of105e106 cells/cm2. After 14 d, the capability of BM-MSCs to form fibroblast-like colonies was assessed.Contrast-phase microscopy after crystal violet stain-ing was used for colony formation evaluation.
The expression of membranous antigen on BM-MSCs was analyzed by a FACSCalibur argon lasercytometer (BD Pharmingen, San Jose, CA, USA)and WinMDI 2.9 software. BM-MSCs were
incubated with antibody (1:20 dilution) for 30 min at4�C. Binding of antibodies to CD73, CD44, CD29,CD34, CD11b, CD14 (BD Pharmingen), CD105(Inc, Minneapolis, MN, USA), CD3 (Miltenyi Bio-tec GmbH, Bergisch Gladbach, Germany), CD19(eBioscience, San Diego, CA, USA) conjugated tophycoerythrin, CD45 and CD31 (BD Pharmingen)and CD90 (Dako, Glostrup, Denmark) conjugatedto fluoresceine isothiocyanate were analyzed bymeans of flow cytometry.
For karyotyping, BM-MSCs from the primarypassages were exposed to phytohemagglutinin (LifeTechnologies) for 15e17 h and then were treated withcolcemid (Life Technologies), detached with 0.125mL trypsin (Life Technologies) and treated with 0.075mol/L KCl in a 37�C water bath. Cells were fixed andspread onto slides. Air-dried slides were stained withfreshly prepared 10% Giemsa stain in a Gurr buffer(Invitrogen, Carlsbad, CA, USA) for 30 min.
Characterization and in vitro recognition of labeled cellswith superparamagnetic iron oxideeprotamine sulfatecomplex
Nanomag-DeSPIO nanoparticles (79e00e201,Micromod Partikeltechnologie GmbH, Rostoc,Germany) were used for contrast-enhanced magneticresonance (MR) nephrography. These nanoparticleswere dextran-coated iron oxide particles that had amean diameter of 20 nm. The solid content of thenanoparticles was 10 mg/mL and their Fe concentra-tion was 2.4 mg/mL. We conducted MR phantomstudies with the use of an aqueous solution of super-paramagnetic iron oxideeprotamine sulfate (SPIO)compounds in 500-mL plastic tubes with an ironconcentration that ranged from 0e200 mg Fe/mL.
SPIO at a concentration of 200 mg Fe/mL andprotamine sulfate (ProS; 90 mg/mL) were put into amixing tube that contained serum-free medium. ProSwas prepared as a fresh stock solution of 10 mg/mL atthe time of use. For attached cells, SPIO-ProS com-plexes were applied directly to the cells followed by theaddition of an equal volumeof the serum-freemediumto the cells, which resulted in a final concentration of100 mg SPIO and 45 mg ProS per milliliter of medium.The cells were incubated for 2 h at 37�C. After incu-bation, we added 10% FBS, 1% L-glutamine and 1%pen/strep to the cells, which were then incubated for48 h.The iron-labeled cells were stainedwith Prussianblue dye (Accustain Iron Stain Kit, Sigma-Aldrich,Steinheim, Germany). For the transmission electronmicroscopic (TEM) studies, the SPIO-labeled cellpellet (1.5e2� 106) was fixed in 2.5% glutaraldehydein 0.1 mol/L phosphate-buffered saline (PBS) (pH7.4) for 2 h. After washing with PBS, cells were post-fixed with 1% osmium tetroxide for 1.5 h, again

736 R. Moghadasali et al.
washed in PBS, dehydrated in an acetone series,embedded in epoxy resin and processed for TEM.Unstained thin sections were evaluated for endocy-tosis of the SPIO nanoparticles. SPIO-labeled cells(1e5 � 106) were used for in vitro cellular MR mea-surement. An agar matrix was used as a suitableenvironment to measure the SPIO-labeled BM-MSCs. The cells were analyzed with the use of anti-bodies CD34, CD45, CD44, CD73, CD90 andCD105 (1:20 dilution) for 30 min at 4�C. Multi-potency differentiation potential of BM-MSCs afterexposure to theSPIO-ProS complexwas confirmedbyinduction of osteogenic and adipogenic differentia-tion.Cell viability and sensitivity to SPIO (100mg/mL)was determined by a standard spectrophotometric 3-(4, 5-dimethylthiazole- 2-yl)-2, 5 diphenyltetrazoliumbromide (MTT; Sigma-Aldrich, M5655) assay.
AKI monkey model
We used healthy rhesusMmulattamonkeys that were1.5e2.5 years old, with body weights of 3e4 kg.Monkeys were gifted from the Primate ResearchCenter at Royan Institute. All animal care, experi-mental and surgical processes and postoperativeeuthanasia were performed in strict accordance withthe ethical principles of theNIHGuide for the Care andUse of Laboratory Animals and the approval of theInstitutional Review Board and Institutional EthicalCommittee at Royan Institute (No. EC.89.1061). Allefforts were made to minimize the number of animalsused. Animals were housed one per cage and providedfree access to food and water throughout the study;they were assessed for tuberculosis, simian immuno-deficiency virus, herpes A and B viruses, hepatitis Aand B viruses and intestinal parasites. A preliminaryexperiment was performed to characterize the extentof tubular cell injury in the AKI monkey model byintravenous injection of cisplatin (Mylan, Athens,Greece) at two different doses, 5mg/kg (n¼ 4, male: 2and female: 2) and 7mg/kg (n¼ 4,male: 3 and female:1). We selected the 5 mg/kg dose for AKI modeling.Renal function, histopathological scores, percentageof fibrosis and hyaline casts were assessed within 4d after cisplatin administration. Treated animals werefollowed for 12 months. Ultrasonographic monitoringand analysis was performed for echogenicity (hyper-echoic renal cortex), cortico-medullary differentiationreduction and size changes (hypertrophic).
Experimental design
To test the effect of the human BM-MSC injection,we divided the male animals into the following threegroups. In group 1 (n ¼ 4) or the control group,animals at day 0 were only exposed to cisplatin.
Group 2 (n ¼ 3) was considered to be the vehiclegroup. At day 0, these animals were exposed tocisplatin followed by an intra-renal arterial injectionof normal saline injection 4 d after cisplatin admin-istration. Group 3 (n ¼ 3) was considered the celltransplantation (Tx) group. At day 0, the animalswere injected with cisplatin, followed by an intra-renal arterial injection of BM-MSCs 4 d later. Anormal group (n ¼ 3) at day 0 was considered to bethe control group of the experiment. This controlgroup had no exposure to cisplatin, no injury and noMSC graft. We used the ultrasonography guide toassist with performing the vehicle and cell injectionsthrough the renal artery.
MR imaging and in vivo cell tracking
MR images (or MR imaging, MRI) of the recipients’kidneys were obtained before injection of the BM-MSCsandat 1 and24hafter transplantation.MRIwasperformed with the use of a 1.5-T MRI system (GESigna Echospeed LX 1.5T MRI [Medical ImagingResources, Inc, Ann Arbor, MI, USA]). Mean signalintensities in the kidneys were evaluated by two radi-ologists experienced in MRI analysis, both of whomwere blinded to the study groups. Signal-to-noise ratioof the outer zone of the renal cortex on T2*-weightedMRI was calculated by dividing signal intensity by thebackground noise. To study intra-renal localization ofBM-MSCs, cells were labeled with SPIO and a PKH-26 red fluorescence cell linker. The PKH-26 cellmembrane embedding dye (Sigma-Aldrich) was usedto incorporate the dye into BM-MSC membranes.Briefly, after SPIO transfection, cells were suspendedin thediluentC thatwas provided,mixedwith an equalvolume of solution of PHK-26 cell linker and incu-bated for 4 min. PKH-26 became excited at a wave-length of 551 nm and emitted at a wavelength of 567nm. Labeling was stopped with an equal volume ofFBS in PBS for 1 min. Cells were washed once withcomplete medium and once with PBS. Prussian bluestaining and fluorescent co-labeling with PKH-26 wasperformed to localize the iron particles in SPIO-labeled MSCs or tissues that received BM-MSCs.After Prussian blue staining was performed, tissuesamples were rapidly dehydrated through gradedalcohol series and xylene andwere thenmounted. Thetissue samples were observed under fluorescence mi-croscopy and analyzed for the presence of PKH-26epositive cells (Olympus IX71; Tokyo, Japan).
Evaluation of renal function
Renal function was followed in all groups at days 4, 7,10, 14, 21, 28 and 84 after cisplatin injection. Sampleswere obtained from the monkey before and after AKI

BM-MSCs ameliorate AKI in monkey 737
and after cell injection over a period of 84 d. Hema-tologic, serum and urine biochemical markers weremeasured with the use of a Hitachi 717 ChemistryAnalyzer (RocheDiagnostics, Indianapolis, IN,USA).
Histopathological analyses and qualitative andquantitative evaluations of renal morphology
Histopathological analyses were performed at 4 and28 d after cisplatin administration. Tissues werewashed twice with PBS and were then fixed with 4%paraformaldehyde for 24 h at 4�C. The samples werethen dehydrated through a series of graded alcoholsolutions and xylol and were then embedded inparaffin. The paraffin-embedded specimens weresectioned into 5-mm thicknesses, placed on poly-l-lysine-coated glass slides and placed in an oven at atemperature of 60�C for 12 h, after which they weredeparaffinized and dewaxed in xylene, stained withhematoxylin and eosin and observed by means oflight microscopy. To assess for the presence oftubular injury, hematoxylin and eosinestained sec-tions were examined and scored for injury asdescribed previously (8). Briefly, 100 tubules werescored for the presence of injury for each kidneysection. A score from 0e3 was assigned for tubulestructures as follows: normal tubule with no lesion(score ¼ 0); tubular cell swelling with brush borderloss and nuclear condensation with loss of no morethan one third of the tubule nuclei (score ¼ 1); morethan one third and less than two thirds of the tubularprofile showing nuclear loss (score ¼ 2); and greaterthan two thirds of the tubular profile showing nuclearloss (score ¼ 3). The total score was calculated byadding all 100 scores to attain a maximum score of300. The numbers of hyaline casts were counted afterexamination of 20 cortical fields for each section.Three coded slides from each kidney were examinedin a blinded manner. To study tissue fibrosis,collagen was stained with Masson’s trichrome stain.To quantitatively assess fibrosis, the percentage ofcortex affected by fibrosis that included interstitialcortical fibrosis, areas of tubular loss with minimalfibrosis and periglomerular fibrosis was estimated tothe nearest 5%. For each section, the measurement offibrosis was repeated twice. Renal tissue slides werestained with periodic acid Schiff reagent and exam-ined by means of light microscopy. To study formaldifferentiation, we stained the samples with he-matoxylin and eosin (adipogenic), alizarin red (oste-ogenic) and toluidine blue stain (chondrogenic).
Transmission electron microscopy
Specimens were fixed with 2.5% glutaraldehyde in0.1 mol/L PBS (pH 7.4) for 2 h. After being washed
with PBS, they were post-fixed with 1% osmiumtetroxide for 1.5 h, again washed in PBS, dehydratedin an acetone series and embedded in epoxy resin.After resin polymerization, sections of approximately50 nm were cut and double-stained with uranyl ac-etate and lead citrate. Electron micrographs wereperformed with the use of a Zeiss EM 900 TEM(München, Germany).
Immunohistochemical staining
We used immunohistochemical staining to assess theFoxp3 protein in the renal tissue sections. Briefly,after deparaffin and rehydration, buffered formalin-fixed sections were treated with antigen retrieval(2H2O/CaCl2 and trypsin) for 20 min at 37�C, fol-lowed by permeabilization with 0.5% Triton X-100for 10 min at room temperature. Sections wereblocked with 10% normal goat serum and were thenincubated with mouse phycoerythrin anti-humanFoxp3 antibody (cross-reactivity with rhesus mon-key; 1:50; clone 259D, Biolegend, San Diego, CA,USA) for 2 h at 37�C. The immune complex Foxp3and anti-Foxp3 antibody on the tissue sections weredetected by fluorescence microscopy (OlympusIX71).
Statistical analysis
Values are given as mean � standard deviation.Analysis was performed with the use of GraphPadPrism 5.02 (Graphpad Software Inc, San Diego, CA,USA). Differences between the experimental groupswere tested by means of one-way analysis of variancewith Dunnett’s or Bonferroni’s multiple comparisontest or two-way repeated measurements. The inde-pendent-samples t tests were performed to identifysignificant differences between hematologic, serumand urine biochemical values of the male and femalemonkeys. A two-sided value of P < 0.05 wasconsidered significantly different.
Results
Characterization of rhesus monkey BM-MSCs
Phase-contrast microscopy of the BM-MSC culturedemonstrated a low heterogeneous population ofcells composed predominately of long, spindle-sha-ped cells and some smaller cells (Figure 1A). Mul-tipotentiality of BM-MSCs was demonstrated bydifferentiation into adipocytes, osteocytes andchondrocytes by staining with oil red, alizarin redand toluidine blue, respectively (Figure 1A). Thefibroblast-like colony-forming unit assay was used toascertain the colony-forming potential of BM-MSCs

Figure 1. Characterization of rhesus monkey BM-MSCs. Morphological features and differentiation capacity of in vitroeexpanded BM-MSCs obtained from bone marrow aspirate are shown. (A) Expanded nonhuman primate BM-MSCs demonstrate the spindle-shapedfibroblastic morphology of BM-MSCs at 14 d of culturing the marrow cells. To verify the capability of BM-MSCs to differentiate intoadipogenic, osteogenic and chondrogenic lineages, three groups of cells were separately stained with either oil red, alizarin red or toluidineblue (original magnification �10, scale bar: 500 mm). (B) A single cell expanded and formed colony-forming unit fibroblasts in culture, asshown by crystal violet staining. (C) To karyotype BM-MSCs cells, metaphase spreads of BM-MSCs were prepared. The chromosomes werearranged and analyzed by the G-banding method. According to the data, BM-MSCs maintained normal diploid karyotypes in culture. (D, E)Immunophenotypic characterization of BM-MSCs was performed by flow cytometry in both the primary culture and the third passage.Histogram analysis of cell surface markers and quantitative analysis showed no significant difference in BM-MSCs between the primaryculture and the third passage. The results indicate that BM-MSCs were not contaminated by hematopoietic cell lineages. Data are presentedas the mean of percentages of positive cells � standard deviation.
738 R. Moghadasali et al.
and stained with crystal violet dye (Figure 1B). Thefrequency of BM-MSCs (colonies per total platednumber of BM-MNCs) ranged from 1 in 1 � 104 to1 � 105. Cytogenetic data at the primary culture andthird passage revealed that BM-MSCs maintainedthe normal chromosome number of 42 chromo-somes (21 pairs), with no detectable rearrangements(Figure 1C).
As shown in Figure 1D,E, phenotype analysisrevealed that the primary cells were negative formarkers such as CD3, CD11b, CD14, CD19, CD31,CD34 and CD45 (leukocyte common antigen);strongly positive for CD29 (97.8% � 2.8%), CD44(71.9% � 16.8%) and CD73 (88.1% � 7.7%); andslightly positive for CD90 (Thy-1; 66.4% � 22.3%)
and CD105 (51.6% � 32.0%). We observed no sig-nificant difference in BM-MSCs between the primaryculture and third passage (Figure 1E and Supple-mentary Figure 1).
SPIO did not alter labeled BM-MSC characteristics andfunction
SPIO-labeled BM-MSCs also differentiated intoosteogenic and adipogenic lineages (SupplementaryFigure 2A). Electron microscopy showed that dark-colored SPIO crystals were located in cytoplasmicvacuoles at �85,000 magnification (SupplementaryFigure 2A). As shown in Supplementary Figure 2B,at 2 d after the cells were labeled with 100 mg/mL

Figure 2. Serum biochemical changes and survival rate of monkeys after injections of cisplatin (5 and 7 mg/kg). (A) Survival rates are shownfor monkeys at the 5 mg/kg (n ¼ 4, male ¼ 2, female ¼ 2) and 7 mg/kg (n ¼ 4, male ¼ 3, female ¼ 1) doses of cisplatin that were assessedwithin 12 months. (B) Before and 4 d after cisplatin injection, we collected serum that was analyzed for creatinine, urea, BUN to creatinine,ion Naþ and Kþ levels. Data are mean � standard deviation. AP < 0.001, BP < 0.01 and CP < 0.05 as compared after 7 mg/kg and 5 mg/kgcisplatin injections to normal; aP < 0.001, bP < 0.01 and cP < 0.05 compared after 7 mg/kg with 5 mg/kg cisplatin administration in 4 d;*P < 0.001 as compared with survival rate after 7 mg/kg with 5 mg/kg cisplatin administration and normal (without cisplatin) during12 months.
BM-MSCs ameliorate AKI in monkey 739
SPIO, we observed no statistically significant decreasein cell viability for labeled (99.1% � 24.1%) com-pared with control cells (non-labeled BM-MSCs;100.0% � 20.6%; P < 0.05). Phenotype analysis ofSPIO-labeled BM-MSCs demonstrated that thesecells were negative for hematopoietic stem cellmarkers CD34 and CD45 but were positive forCD44 (93.5% � 8.1%), CD73 (99.4% � 0.4%),CD90 (55.5% � 20.9%) and CD105 (74.0% �18.7%). We observed no significant difference inexpression of surface markers on BM-MSCs beforeand after labeling with SPIO (SupplementaryFigure 2C).
AKI is efficiently induced in the rhesus monkey bycisplatin
Monkeys that received 7 mg/kg of cisplatin died onmonths 1 (n ¼ 2), 3 (n ¼ 1) and 10 (n ¼ 1) aftercisplatin administration. However, there was mod-erate renal damage and lower mortality rate afteradministration of 5 mg/kg cisplatin during 1 year(Figure 2A).
Urine, serum and blood of 95 male and femalehealthy monkeys were analyzed to show their de-mographic status before cisplatin treatment (Supple-mentary Table I). Laboratory analyses of themonkeys4 d after injections of cisplatin demonstrated thatcreatinine and urea of the treated monkeys increasedsignificantly in comparison with normal animals[7 mg/kg cisplatin (creatinine: 8.4 � 3.6 mg/dL; urea:152.7 � 44.7 mg/dL) and 5 mg/kg (creatinine: 2.0 �1.9 mg/dL; urea: 58.4 � 41.7 mg/dL; Figure 2B)].
At the 7 mg/kg concentration, the renal tubulesshowed evidence of cytoplasmic vacuolization, loss ofbrush border, flattening and epithelial cell detach-ment in addition to luminal cell debris and nuclearfragmentation (Figure 3A). The most damagedtubules displayed areas of obvious denudation of thetubular basement membranes. Prominent hyalinecasts were present in the tubular lumina. Intravenousinjection of 5 mg/kg cisplatin induced moderatekidney failure (pathological score: 138.4 � 50.8),whereas 7 mg/kg cisplatin induced strong kidneyfailure (pathological score: 220.8 � 16.9; Figure 3B).Kidneys from monkeys treated with 7 mg/kg cisplatinshowed more severe structural changes that includedinflammation and fibrosis (Figure 3B).
TEM analysis showed that high-dose cisplatin(7 mg/kg) induced severe nephrotoxicity character-ized ultrastructurally by swelling and blunting of footprocesses and generation of cell junctions betweenpodocyte foot processes, nucleolar segregation andcytoplasmic blebbing with focal loss of microvilli.Cytoplasmic vacuoles with dense materials werefound in the proximal tubules. Cisplatin, by virtue ofits low molecular weight and uncharged character, isfreely filtered at the glomerulus and almostcompletely recovered in the urine. This process re-sults in an uptake of the drug by renal tubular cellsthat reaches its highest concentrations in the prox-imal tubular cells of the inner cortices and outermedullae (S3 segment) (9). These are the major sitesof cisplatin-induced renal damage; however, cisplatincan also injure the distal tubule and collecting duct ina dose-dependent manner (10,11). Therefore, both
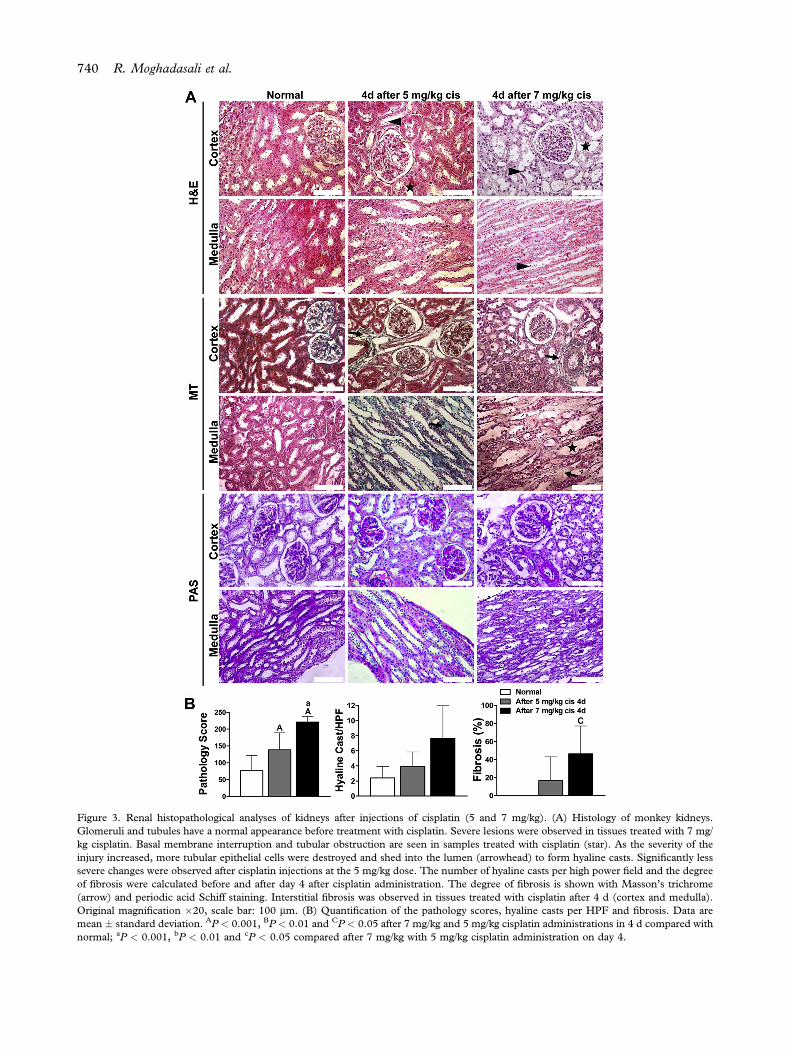
Figure 3. Renal histopathological analyses of kidneys after injections of cisplatin (5 and 7 mg/kg). (A) Histology of monkey kidneys.Glomeruli and tubules have a normal appearance before treatment with cisplatin. Severe lesions were observed in tissues treated with 7 mg/kg cisplatin. Basal membrane interruption and tubular obstruction are seen in samples treated with cisplatin (star). As the severity of theinjury increased, more tubular epithelial cells were destroyed and shed into the lumen (arrowhead) to form hyaline casts. Significantly lesssevere changes were observed after cisplatin injections at the 5 mg/kg dose. The number of hyaline casts per high power field and the degreeof fibrosis were calculated before and after day 4 after cisplatin administration. The degree of fibrosis is shown with Masson’s trichrome(arrow) and periodic acid Schiff staining. Interstitial fibrosis was observed in tissues treated with cisplatin after 4 d (cortex and medulla).Original magnification �20, scale bar: 100 mm. (B) Quantification of the pathology scores, hyaline casts per HPF and fibrosis. Data aremean � standard deviation. AP < 0.001, BP < 0.01 and CP < 0.05 after 7 mg/kg and 5 mg/kg cisplatin administrations in 4 d compared withnormal; aP < 0.001, bP < 0.01 and cP < 0.05 compared after 7 mg/kg with 5 mg/kg cisplatin administration on day 4.
740 R. Moghadasali et al.

BM-MSCs ameliorate AKI in monkey 741
proximal and distal tubules have shown frequenttubular necrosis. Significantly less severe changeswere observed after administration of 5 mg/kgcisplatin. In the proximal tubules, there was a lesssevere decrease in microvilli, although apical bleb-bing was noticeable, with large numbers of densebodies in the lumen of proximal tubules (Supple-mentary Figure 3). Sonographic analysis indicated that5 mg/kg of cisplatin (after 4 d) caused a hyperechoicrenal cortex and reduction in cortico-medullary dif-ferentiation but no change in size (SupplementaryFigure 4). We selected the 5 mg/kg cisplatin dosebecause of the high mortality rate seen with the 7mg/kg dose. Additionally, only male monkeys werechosen to avoid sex-related differences in the AKImodel (12).
BM-MSCs efficiently implanted in the injured area of thekidney after cisplatin treatment
Figure 4A shows a schematic overview of theexperiment and analysis of different stages. MRIsignal intensities were obtained from each image ofSPIO at 0e200 (0, 10, 25, 50, 100, 150 and 200) mgFe/mL concentrations through the use of an MRIsystem (GE Signa; LX Echospeed) (Figure 4B). MRIsignals decreased gradually from 0e200 mg Fe/mLconcentrations. Fe-labeled cells at the 5 � 106 con-centration emitted a dark signal that was sensitive inthe T2*-weighted image. This signal intensity wasnot observed in unlabeled control cells (Figure 4C).We did not detect any clinical evidence for immunereaction after injection of autologous BM-MSCs. Bymeans of MRI, we observed transplanted cells in therenal cortex; their presence was demonstrated bydistinct regional signal intensity loss induced by thesusceptibility effects of iron oxide particles. Thedistribution of the signal intensity loss was solelycortical and confirmed by in vivo images(Figure 4D). To verify the MRI results, kidney sliceswere stained with Prussian blue dye, which showedthe presence of SPIO-labeled BM-MSCs in the renaltissue (Figure 4E). These cells were located withinthe kidney glomeruli in the area of decreased signalintensity. Although the majority of BM-MSCslocalized in the glomeruli, some were observed in theinterstitial tissue and tubules. Co-labeling withfluorescence (PKH-26) increased the sensitivity fordetection of the labeled cells. PKH-26, a fluorescentdye, efficiently labeled BM-MSCs after transfectionwith SPIO. Iron oxide can be detected by Prussianblue staining, whereas PKH-26 is detectable byfluorescence. Therefore, dual labeling with iron ox-ide and PKH-26 enabled us to double-check cellhoming in vivo. The Prussian blue dye obscuredfluorescent emissions because during its fixation
process, the fluorescent dye leaked from the cell (13).Therefore, we observed SPIO-positive cells in blackunder fluorescence microscopy (SupplementaryFigure 5).
BM-MSCs attenuated renal dysfunction in the cisplatin-treated rhesus monkey
In the control group, onemonkey died at 8months; inthe vehicle group, one monkey died at 6 months. Incontrast, no deaths occurred after cell transplantationduring 9 months (Figure 5A). No significant differ-ence existed in renal functional analyses between thevehicle and control groups (Figure 5B). Thus, wecompared the results between the cell Tx and vehiclegroups. In this study, intravenous injection of 5 mg/kgcisplatin induced increases in serum creatinine andurea at day 4, which confirmed the presence of AKI.Injection of 5 � 106 MSCs/kg ameliorated renalfunction, as reflected by a significantly lower serumcreatinine level at day 7 in the cell Tx group (0.9� 0.3mg/dL) versus the vehicle (1.8 � 1.6 mg/dL, P <0.001) group. This result stabilized from days 10(0.9 � 0.1 mg/dL cell Tx vs 1.7 � 1.0 vehicle, P <0.001), 14 (0.9 � 0.0 mg/dL cell Tx vs 1.4 � 0.8vehicle, P < 0.001), 21 (0.8 � 0.0 mg/dL cell Tx vs1.3 � 0.7 vehicle, P < 0.01) and 28 (0.8 � 0.1 mg/dLcell Tx vs 1.3� 0.6 vehicle, P< 0.01) to 84 (0.8� 0.2mg/dL cell Tx vs 1.0 � 0.5 vehicle). We observedsignificantly lower urea values on day 7 in the cell Txgroup (35.5 � 10.6 mg/dL) versus vehicle group(58.7 � 48.5, P< 0.001). This result declined on day10 in the cell Tx group (29.3 � 8.1 mg/dL) versusvehicle group (50.0 � 33.3, P < 0.05) and stabilizedfrom day 14 in the cell Tx group (25.0 � 3.6 mg/dL)compared with the vehicle group (46.0 � 30.3, P <0.01) up to day 84 (19.7 � 1.5 mg/dL cell Tx vs34.2 � 12.7 vehicle). In the cell Tx group, weobserved no hyponatremia or hyperkalemia; however,the vehicle group had decreased Naþ ions that peakedon day 4 (137.5� 3.5 mEq/L) and stabilized by day 7(137.5� 3.5 mEq/L). These ions increased on day 10(142.3 � 5.9 mEq/L) and stabilized by day 84(144.0 � 4.6 mEq/L). There was an increase in Kþ
ions, which peaked on day 14 (5.3 � 0.4 mEq/L),declined on day 21 (4.8 � 0.0 mEq/L) and stabilizedon day 28 (5.0 � 1.0 mEq/L). We observed valueshigher than baseline at day 84 (5.4 � 0.7 mEq/L).Creatinine clearance decreased after cisplatin injec-tion, then peaked on day 4 in the cell Tx group (34.1�9.2 mL/24 h) versus vehicle group (34.0 � 35.4 mL/24 h). There was a higher creatinine clearance levelafter the cell injection that increased on day 7 in thecell Tx group (62.8� 29.6mL/24 h) versus the vehiclegroup (35.6 � 24.5 mL/24 h) that stabilized on day10 in the cell Tx group (64.7 � 12.7 mL/24 h) versus

Figure 4. Schematic overview of experimental set-up and in vitro and in vivo cell tracking by MRI. (A) Schematic overview ofexperimental and analysis stages. (B) In vitro cellular MRI and evaluation of iron labeling of MSCs by cellular MRI. Comparative MRphantom study of cell-free SPIO solution at different doses (0e200 mg Fe/mL) show the relation between signal intensity and ironconcentration on T2*-weighted images. Coronal sequence image of phantom shows marked decrease in signal intensity with increasingmagnetic nanoparticle concentration. (C) Experimental MRI of phantom of labeled BM-MSCs with 100 mg Fe/mL placed in 1%phantom agar in plastic wells to select sensitive concentration of SPIO-labeled cells for MRI. Axial and coronal sequence images ofphantom shows marked decrease in signal intensity with increasing cell numbers. From left: 5 � 106, 3 � 106, 2 � 106 and 1 � 106
unlabeled BM-MSCs and agar. (D) Axial and coronal T2*-weighted MRI of the monkeys’ kidneys obtained before, 2 h and 24 h afterinjection with labeled BM-MSCs. After BM-MSC injection, MRI showed a mild signal intensity decrease around the renal pelvis(arrows). Signal intensity loss in the artery zone of the renal cortex (arrows) as detected on T2*-weighted MRI of the kidneys obtained2 h after injection with labeled BM-MSCs. At 24 h after labeled BM-MSCs were transplanted, a decrease in signal intensity (whitearrows) in the cortical and outer zone is still visible on the T2*-weighted MRI. (E) Micrograph of histological specimen from neph-rotoxic kidneys obtained at 24 h, 3 d and 28 d after SPIO-labeled BM-MSC injection. Arrows indicate Prussian blue stainepositiveparticles in glomeruli and tubules of the renal cortex. Most cells positive for Prussian blue staining (arrow) are located in the glomeruliin the outer zone of the renal cortex (original magnification �40 and �20, scale bar: 50 mm).
742 R. Moghadasali et al.

Figure 5. Effect of BM-MSCs on renal function and survival rate of monkeys after cell transplantation. (A) We assessed monkey survivalrates within 9 months in the different groups. (B) Dynamics of serum creatinine, urea, BUN to creatinine, Naþ and Kþ ions, creatinineclearance, urea clearance, urine volume and urine protein excretion levels in monkeys with AKI after intra-renal arterial injections of normalsaline (vehicle group) and BM-MSCs (cell Tx group) compared with the cisplatin-alone group (control) during 84 d of follow-up. Values aremean � standard deviation. AP < 0.001, BP < 0.01 and CP < 0.05 compared between the cell Tx and control groups; aP < 0.001, bP < 0.01and cP < 0.05 compared between the cell Tx and vehicle groups; *P < 0.001 between the control with vehicle and cell Tx groups during9 months. #P < 0.01 compared between the vehicle and cell Tx groups during 9 months.
BM-MSCs ameliorate AKI in monkey 743

744 R. Moghadasali et al.
the vehicle group (44.0 � 11.4 mL/24 h). Thiscontinued until day 84 in the cell Tx group (69.0 �16.9 mL/24 h) compared with the vehicle group (60.1� 18.8 mL/24 h). There was a decrease in ureaclearance after cisplatin administration that peaked onday 4 in the cell Tx group (13.9 � 7.3 mL/24 h)compared with the vehicle group (19.0 � 15.8 mL/24h). In the cell Tx group, urea clearance increased onday 7 in this group (31.2 � 16.9 mL/24 h) comparedwith the vehicle group (16.3 � 9.8 mL/24 h). Thisincrease was also observed on day 28 in the cell Txgroup (48.6� 14.2 mL/24 h) versus the vehicle group(20.8 � 10.9 mL/24 h). We observed a significantdifference in urea clearance on day 84 in the cell Txgroup (53.9 � 49.0 mL/24 h) compared with thevehicle group (34.1 � 22.8, P <0.01). Proteinuria, asprotein/creatinine, increased on day 4 in the cell Txgroup (0.7 � 0.4 mg/24 h) compared with the vehiclegroup (0.6 � 0.4 mg/24 h). However, in the cell Txgroup, protein excretion declined on day 7 to 0.3� 0.3mg/24 h compared with 1.0 � 0.5 mg/24 h for thevehicle group. This decline was significantly lower onday 28 for the cell Tx group (0.3 � 0.2 mg/24 h)compared with the vehicle group (1.6 � 1.2 mg/24 h,P < 0.001) and was 0.3 � 0.3 mg/24 h for the cell Txgroup compared with 0.9� 0.6mg/24 h for the vehiclegroup on day 84. Diuresis as determined by the vol-ume of urine over a 24-h period induced a decrease inall groups, which peaked on day 4 for the cell Tx group(110.3 � 77.0 mL/24 h) compared with the vehiclegroup (106.0 � 50.3 mL/24 h). This value increased,as evidenced by polyuria on day 14 in the vehicle group(332.0 � 92.9 mL/24 h) but not in the cell Tx group(167.0 � 69.6 mL/24 h). Urine volume graduallyincreased by day 28 in the cell Tx group (318.5� 26.2mL/24 h) versus the vehicle group (255.5 � 85.0 mL/24 h) compared with normal values and stabilized onday 84 at 307.7 � 27.3 mL/24 h for the cell Tx groupcompared with 342.6 � 83.5 mL/24 h for the vehiclegroup (Figure 5). Overall, levels of creatinine, urea,Kþ ion and protein/creatinine were higher than base-line, whereas creatinine clearance and urea clearancewere lower than baseline in the vehicle group over 84d. No significant difference existed between urine pHand specific gravity in monkeys with AKI in all groups(Supplementary Table II).
BM-MSCs preserved globular structure but did notregenerate tubular damage after cisplatin treatment
By light microscopy, we detected significant changesin the kidneys 4 d after cisplatin (5 mg/kg) injec-tions (Figure 6 and Supplementary Table III). Ina study of proximal tubules, we observed the lossof brush border, cytoplasmic vacuolization, nuclearfragmentation and segmentation, loss of the epithelial
cells, apoptosis and necrosis of the epithelial cells, inaddition to the release of cell debris in the lumen.Distal and proximal tubules were remarkable forpathological changes. Tubules that sustained the mostinjuries displayed areas of apparent denudation of thetubular basementmembranes (Figure 6A). Pathologicscores for tubular damage peaked on day 4 at 147.9 �9.2 for the cell Tx group versus 161.5 � 18.5 for thevehicle group. Hyaline casts were visible as prominentin tubular lumens in the cell Tx group (6.4 � 0.9)compared with the vehicle group (6.5 � 4.9;Figure 6B). On day 28, tubular changes were stillpresent; however, there was partial recovery observedin the three groups (Figure 6B). Unlike the beneficialeffects on renal function, BM-MSCs did not have aremarkable effect on renal pathology over 28 d, asevidenced by the pathologic score of 150.5 � 18.3 inthe cell Tx group versus 150.0 � 2.4 for the vehiclegroup and hyaline casts per high power field value of6.2� 0.6 for the cell Tx group versus 5.6� 4.0 for thevehicle group. Moreover, the intra-renal trans-plantation of BM-MSCs in our study did not result inany maldifferentiation to chondrogenic, osteogenicand adipogenic lineages (Supplementary Figure 6).
Electron microscopy analysis of all cisplatin-treated monkeys showed cytoplasmic vacuolization,loss of brush border, apical blebbing, phag-olysosomes and enlarged mitochondria with irregularcrests (Supplementary Figure 7). In contrast, thekidneys from monkeys that received BM-MSCs onday 28 showed substantially less severe ultrastruc-tural damage. Significantly, areas of cell detachmentwere either absent or detectable in only a few scat-tered tubules. Glomerular structure was better pre-served in the cell Tx group compared with that in thecontrol or vehicle groups. Proximal epithelial cells inthe control and vehicle kidneys were dark andshrunken, with condensed nuclei and cytoplasmicvacuoles, many of which contained dense materialsand apical blebbing. Hypertrophied proximal as wellas distal epithelial cells with dividing mitochondriawere also observed. Ultrastructural analyses of theproximal and distal tubules in the cell Tx group wereremarkable for approximately normal morphologies,with slender microvilli in the proximal tubules andno prominent hypertrophic changes (SupplementaryFigure 7).
BM-MSCs increased Foxp3þ T-regulatory cellinfiltration in cisplatin-induced damaged kidneys
As shown in Figure 7, Foxp3-expressing T-regula-tory (Treg) cells were observed in the cell Tx groupafter cell transplantation at day 28. However, asvisualized by periodic acid Schiff staining, there wasan increase in interstitial fibrosis and matrix. There

Figure 6. Effect of BM-MSCs on renal histopathology. (A) Renal injury characterization and morphological changes after intra-renalarterial injection of BM-MSCs. Histopathological changes, number of hyaline casts per high power field and the degree of fibrosis in themonkeys’ kidneys were compared 4 and 28 d after injection of 5 mg/kg cisplatin in the control, vehicle and cell Tx groups. The degree offibrosis is visualized by Masson’s trichrome staining (arrow). Interstitial fibrosis was observed in tissues treated with cisplatin (5 mg/kg) after4 and 28 d (original magnification �20, scale bar: 100 mm). (B) Histopathological analysis, hyaline casts and fibrosis scores were notsignificantly different between the control, vehicle and cell Tx groups at 4 and 28 d after cisplatin (5 mg/kg) injection. Data are mean �standard deviation.
BM-MSCs ameliorate AKI in monkey 745
were no Foxp3þ Treg cells visualized after cisplatininjection on day 4 and in the control and vehiclegroups after 28 d.
Discussion
Initially, we generated a cisplatin-induced AKImodel in the monkey. According to histological andfunctional assessments, 7 mg/kg cisplatin adminis-tration was considered to be the lethal dose and 5mg/kg induced AKI. The pattern of lesions observedin our 5 mg/kg cisplatin-induced AKI monkey modelappeared to better mimic human nephropathy (14).
In this condition, we assessed the ability of trans-planted MSCs to ameliorate AKI as early anddecrease side effects of cisplatin as long term up to84 d.
Moreover, we have shown that the intra-renalarterial transplantation of BM-MSCs amelioratedrenal function and improved tubular damage in thismodel, as detected by biochemical but not histolog-ical analyses. Our results agreed with several reportsin rodent models that have shown which MSCsimproved the outcome of AKI (15,16). Most studiesreported the beneficial effect of MSC infusion onrenal function during the early period after AKI

Figure 7. Presence of Foxp3þ regulatory T (Treg) cells after cell transplantation. Kidney tissues obtained from days 4 and 28 after cisplatininjection in the control, vehicle and cell Tx groups were fixed in 10% formalin and embedded in paraffin. Foxp3 protein in 5-mm-thicksections was identified by mouse anti-human Foxp3 antibody. Histopathological hallmarks of nephropathy are shown in periodic acidSchiffestained sections. Red (arrow) indicates positive staining of Foxp3; blue (arrow), nucleus (Hoechst stain) (original magnification �20,scale bar: 100 mm).
746 R. Moghadasali et al.
induced as ischemia-reperfusion injury (15,17,18)and nephrotoxines such as cisplatin (7,19). Howeversome studies (20e22) have failed to demonstrate anybeneficial effect. The beneficial effect of BM-MSCshas been also observed for AKI in a rat model (23).
We have chosen the intra-renal arterial route,which was successfully used by other researchers(17,24e26) to more efficiently deliver cells to thetargeted organ. Because the amount of stem cellsinjected might be critical, in the current study weinjected approximately 5� 106MSCs/kg bodyweight,
which was the lowest cell number (5 � 106) deemedsensitive for MRI tracking (Figure 4C).
Despite biochemical improvement after celltransplantation, we have not observed any significanthistological improvement. Evidence exists thatshows, in some cases such as septic-induced kidneyinjury, biological indices cannot be immediately fol-lowed by histological findings, even after treatment(27). Liu et al. (28,29) have shown that injections ofBM-MSC are helpful for AKI repair, as evidenced bya significant decrease in blood urea nitrogen (BUN)

BM-MSCs ameliorate AKI in monkey 747
level without any significant changes in acute tubularnecrosis scoring. An erythropoietin injection com-bined with MSC injection can cause a sharp decreasein BUN and creatinine levels as well as in acutetubular necrosis scoring. On the other hand, cisplatinis a toxic drug with high mortality rates (3). Aftercisplatin administration, we have observed increasedhistopathological scores and high levels of necrosis.Hence, it was presumed that the beneficial effects ofBM-MSCs on the remaining live cells were acompensatory. It was demonstrated that animals whoreceived MSCs 7 d after ischemia-reperfusion injuryshowed substantial resolution of necrotic injury inthe larger areas; however, tubular injury was stillevident in significant regions (15).
The mechanism(s) of improvement by MSCtransplantation in a cisplatin-induced AKI model isunclear. However, three primary mechanisms couldexplain the therapeutic effects of MSCs in the AKImodel. First, MSCs could generate soluble factorsthat facilitate kidney recovery in response to acutedamage. We observed BM-MSCs in the glomeruliand tubules after cell transplantation. Moreover,numerous studies have reported engraftment ofvarious MSCs in acute renal failure models(5,15,21,30e36). It was also demonstrated that onlya transient recruitment of MSCs in renal vasculaturewithout a direct incorporation within the regenerat-ing tubules could improve the injured kidney modelthrough survival of the kidney cells as the result oflimited apoptosis and enhanced proliferation of theendogenous tubular epithelial cells (18,20,24,37,38).Bruno et al. (39) have suggested that MSC-derivedexosomes might activate a proliferative program inthe surviving tubular cells after an injury.
The contribution of paracrine effects has beenfurther supported by a recent study reporting thatinfusion of BM-MSCs reduced the severity ofcisplatin-induced AKI in an adult rodent modelwithout cell integration into the kidney (40). Wehave also described that cytokines secreted by MSCsstimulate renal tubule cell regeneration after neph-rotoxicity in vitro (41).
The second mechanism by which MSCs couldpromote kidney repair is through inducing Tregcells. Growing evidence supports that inflammationcontributes to AKI (42,43). It appears that afterkidney ischemia-reperfusion injury, Treg cells facili-tated the recovery process through suppression ofinnate immune responses (44,45). Additionally,several anti-inflammatory components have thecapability to attenuate kidney injury in animalmodels of ischemia/reperfusion-induced AKI. It hasbeen demonstrated that the beneficial effect ofFTY720, an immunosuppressive agent, in solid or-gan transplantation was associated with a higher
frequency of CD4þ CD25þ Tregs and an upregu-lation of Foxp3 messenger RNA expression in thespleen and kidneys (46). The transcription factorFoxp3 is expressed by CD4þCD25þ cells and pro-grams their development and function (47e50). Weobserved that Foxp3þ Treg cells infiltrated into therenal tissue in the cell Tx group by day 28 aftertransplantation. A protective effect of Foxp3þ Tregcells on cisplatin-induced nephrotoxicity in mice(51,52) has been shown, which suggested that thepresence of Foxp3-expressing Treg cells inside therenal tissues assists with nephrotoxicity recoverythrough its regulatory function. We also could notdetect Foxp3þ Treg cells in renal tissue aftercisplatin administration at day 4 in all groups; rather,a number of Foxp3þ Treg cells were observed at day28 in the Tx group. This might be related to theirrecruitment by transplanted MSCs, and this result inattenuating ischemic AKI (44,53). MSCs have alsobeen shown to induce Tregs both in vitro (54e57)and in vivo (58e62).
Moreover, cell fusion with host kidney cells hasbeen reported to be another mechanism for the ther-apeutic effects of MSCs in a kidney injury. This waspreviously demonstrated by fusion of transplantedhematopoietic stem cells and hepatocytes (63,64).However, this event appeared to be restricted to amouse model of human liver failure (fumar-ylacetoacetate hydrolase deficiency) with its high se-lective pressure in these reports. Numerous studieshave since demonstrated that hematopoietic stem cellsand MSCs improved liver function without cellularfusion in a variety of different animal models (65e68).
In conclusion, our report shows that intra-renalarterial injection of autologous BM-MSCs amelio-rates cisplatin-induced AKI in a rhesus M mulattamonkey model. This provides a tool to confirm re-sults obtained in murine experimental studies and aninvaluable tool for evaluating cell transplantationprotocols before their use in clinical studies. Furtherstudies are necessary to definitively elucidate themechanism of improvement in the AKI scenario.
Acknowledgments
This work was supported by a grant from the CharityAssociation for Health Research and Royan Institute(Tehran, Iran). We thank Dr Abbas Piryaei, NajmehSadat Masoudi, Ehsan Janzamin, Fazel SahraneshinSamani, Javad Firouzi and Pardis Khosravani fortechnical assistance and critical comments.
Disclosure of interests: The authors have nocommercial, proprietary, or financial interest in theproducts or companies described in this article.

748 R. Moghadasali et al.
References
1. Nash K, Hafeez A, Hou S. Hospital-acquired renal insuffi-ciency. Am J Kidney Dis. 2002;39:930e6.
2. Schrier RW, Wang W, Poole B, Mitra A. Acute renal failure:definitions, diagnosis, pathogenesis, and therapy. J ClinInvest. 2004;114:5e14.
3. Pabla N, Dong Z. Cisplatin nephrotoxicity: mechanisms andrenoprotective strategies. Kidney Int. 2008;73:994e1007.
4. Aejaz HM, Aleem AK, Parveen N, Khaja MN, Narusu ML,Habibullah CM. Stem cell therapy-present status. TransplantProc. 2007;39:694e9.
5. Morigi M, Imberti B, Zoja C, Corna D, Tomasoni S,Abbate M, et al. Mesenchymal stem cells are renotropic,helping to repair the kidney and improve function in acuterenal failure. J Am Soc Nephrol. 2004;15:1794e804.
6. Morigi M, Introna M, Imberti B, Corna D, Abbate M,Rota C, et al. Human bone marrow mesenchymal stem cellsaccelerate recovery of acute renal injury and prolong survivalin mice. Stem Cells. 2008;26:2075e82.
7. Eliopoulos N, Zhao J, Bouchentouf M, Forner K, Birman E,Yuan S, et al. Human marrow-derived mesenchymal stromalcells decrease cisplatin renotoxicity in vitro and in vivo andenhance survival of mice post-intraperitoneal injection. Am JPhysiol Renal Physiol. 2010;299:F1288e98.
8. Gheisari Y, Nassiri SM, Arefian E, Ahmadbeigi N,Azadmanesh K, Jamali M, et al. Severely damaged kidneyspossess multipotent renoprotective stem cells. Cytotherapy.2010;12:303e12.
9. Kuhlmann MK, Burkhardt G, Kohler H. Insights into po-tential cellular mechanisms of cisplatin nephrotoxicity andtheir clinical application. Nephrol Dial Transplant. 1997;12:2478e80.
10. Arany I, Safirstein RL. Cisplatin nephrotoxicity. SeminNephrol. 2003;23:460e4.
11. Taguchi T, Nazneen A, Abid MR, Razzaque MS. Cisplatin-associated nephrotoxicity and pathological events. ContribNephrol. 2005;148:107e21.
12. Wei Q, Wang MH, Dong Z. Differential gender differences inischemic and nephrotoxic acute renal failure. Am J Nephrol.2005;25:491e9.
13. Flexman JA, Minoshima S, Kim Y, Cross DJ. Magneto-optical labeling of fetal neural stem cells for in vivo MRItracking. Conf Proc IEEE Eng Med Biol Soc. 2006;1:5631e4.
14. Rosen S, Heyman SN. Difficulties in understanding human“acute tubular necrosis”: limited data and flawed animalmodels. Kidney Int. 2001;60:1220e4.
15. Kale S, Karihaloo A, Clark PR, Kashgarian M, Krause DS,Cantley LG. Bone marrow stem cells contribute to repair ofthe ischemically injured renal tubule. J Clin Invest. 2003;112:42e9.
16. Reinders ME, Fibbe WE, Rabelink TJ. Multipotent mesen-chymal stromal cell therapy in renal disease and kidneytransplantation. Nephrol Dial Transplant. 2010;25:17e24.
17. Togel F, Weiss K, Yang Y, Hu Z, Zhang P, Westenfelder C.Vasculotropic, paracrine actions of infused mesenchymal stemcells are important to the recovery from acute kidney injury.Am J Physiol Renal Physiol. 2007;292:F1626e35.
18. Togel F, Hu Z, Weiss K, Isaac J, Lange C, Westenfelder C.Administered mesenchymal stem cells protect againstischemic acute renal failure through differentiation-indepen-dent mechanisms. Am J Physiol Renal Physiol. 2005;289:F31e42.
19. Yuan L, Wu MJ, Sun HY, Xiong J, Zhang Y, Liu CY, et al.VEGF-modified human embryonic mesenchymal stem cellimplantation enhances protection against cisplatin-induced
acute kidney injury. Am J Physiol Renal Physiol. 2011;300:F207e18.
20. Duffield JS, Park KM, Hsiao LL, Kelley VR, Scadden DT,Ichimura T, et al. Restoration of tubular epithelial cells duringrepair of the postischemic kidney occurs independently of bonemarrow-derived stem cells. J Clin Invest. 2005;115:1743e55.
21. Lin F, Moran A, Igarashi P. Intrarenal cells, not bonemarrow-derived cells, are the major source for regeneration inpostischemic kidney. J Clin Invest. 2005;115:1756e64.
22. Lin F, Cordes K, Li L, Hood L, Couser WG, Shankland SJ,et al. Hematopoietic stem cells contribute to the regenerationof renal tubules after renal ischemia-reperfusion injury inmice. J Am Soc Nephrol. 2003;14:1188e99.
23. Togel F, Cohen A, Zhang P, Yang Y, Hu Z, Westenfelder C.Autologous and allogeneic marrow stromal cells are safe andeffective for the treatment of acute kidney injury. Stem CellsDev. 2009;18:475e85.
24. Lange C, Togel F, Ittrich H, Clayton F, Nolte-Ernsting C,Zander AR, et al. Administered mesenchymal stem cellsenhance recovery from ischemia/reperfusion-induced acuterenal failure in rats. Kidney Int. 2005;68:1613e7.
25. Kunter U, Rong S, Djuric Z, Boor P, Muller-Newen G,Yu D, et al. Transplanted mesenchymal stem cells accelerateglomerular healing in experimental glomerulonephritis. J AmSoc Nephrol. 2006;17:2202e12.
26. Behr L, Hekmati M, Lucchini A, Houcinet K, Faussat AM,Borenstein N, et al. Evaluation of the effect of autologousmesenchymal stem cell injection in a large-animal model ofbilateral kidney ischaemia reperfusion injury. Cell Prolif.2009;42:284e97.
27. Langenberg C, Gobe G, Hood S, May CN, Bellomo R. Renalhistopathology during experimental septic acute kidney injuryand recovery. Crit Care Med. 2014;42:e58e67.
28. Liu N, Tian J, Cheng J, Zhang J. Effect of erythropoietin onthe migration of bone marrow-derived mesenchymal stemcells to the acute kidney injury microenvironment. Exp CellRes. 2013;319:2019e27.
29. Liu N, Han G, Cheng J, Huang J, Tian J. Erythropoietinpromotes the repair effect of acute kidney injury by bone-marrow mesenchymal stem cells transplantation. Exp BiolMed (Maywood). 2013;238:678e86.
30. Fang TC, Alison MR, Cook HT, Jeffery R, Wright NA,Poulsom R. Proliferation of bone marrow-derived cells con-tributes to regeneration after folic acid-induced acute tubularinjury. J Am Soc Nephrol. 2005;16:1723e32.
31. Broekema M, Harmsen MC, Koerts JA, Petersen AH, vanLuyn MJ, Navis G, et al. Determinants of tubular bonemarrow-derived cell engraftment after renal ischemia/reper-fusion in rats. Kidney Int. 2005;68:2572e81.
32. Imasawa T, Utsunomiya Y, Kawamura T, Zhong Y,Nagasawa R, Okabe M, et al. The potential of bone marrow-derived cells to differentiate to glomerular mesangial cells.J Am Soc Nephrol. 2001;12:1401e9.
33. Behr L, Hekmati M, Fromont G, Borenstein N, Noel LH,Lelievre-Pegorier M, et al. Intra renal arterial injection ofautologous mesenchymal stem cells in an ovine model in thepostischemic kidney. Nephron Physiol. 2007;107:p65e76.
34. Herrera MB, Bussolati B, Bruno S, Morando L, Mauriello-Romanazzi G, Sanavio F, et al. Exogenous mesenchymal stemcells localize to the kidney by means of CD44 following acutetubular injury. Kidney Int. 2007;72:430e41.
35. HerreraMB, Bussolati B, Bruno S, Fonsato V,Romanazzi GM,Camussi G. Mesenchymal stem cells contribute to the renalrepair of acute tubular epithelial injury. Int JMolMed. 2004;14:1035e41.
36. Togel F, Yang Y, Zhang P, Hu Z, Westenfelder C. Biolu-minescence imaging to monitor the in vivo distribution of

BM-MSCs ameliorate AKI in monkey 749
administered mesenchymal stem cells in acute kidney injury.Am J Physiol Renal Physiol. 2008;295:F315e21.
37. Humphreys BD, Bonventre JV. Mesenchymal stem cells inacute kidney injury. Annu Rev Med. 2008;59:311e25.
38. Humphreys BD, Valerius MT, Kobayashi A, Mugford JW,Soeung S, Duffield JS, et al. Intrinsic epithelial cells repair thekidney after injury. Cell Stem Cell. 2008;2:284e91.
39. Bruno S, Grange C, Deregibus MC, Calogero RA, Saviozzi S,Collino F, et al. Mesenchymal stem cell-derived microvesiclesprotect against acute tubular injury. J Am Soc Nephrol. 2009;20:1053e67.
40. Bi B, Schmitt R, Israilova M, Nishio H, Cantley LG. Stromalcells protect against acute tubular injury via an endocrine ef-fect. J Am Soc Nephrol. 2007;18:2486e96.
41. Moghadasali R, Mutsaers HA, Azarnia M, Aghdami N,Baharvand H, Torensma R, et al. Mesenchymal stem cell-conditioned medium accelerates regeneration of human renalproximal tubule epithelial cells after gentamicin toxicity. ExpToxicol Pathol. 2013;65:595e600.
42. Bonventre JV, Zuk A. Ischemic acute renal failure: an in-flammatory disease? Kidney Int. 2004;66:480e5.
43. Friedewald JJ, Rabb H. Inflammatory cells in ischemic acuterenal failure. Kidney Int. 2004;66:486e91.
44. Gandolfo MT, Jang HR, Bagnasco SM, Ko GJ, Agreda P,Satpute SR, et al. Foxp3þ regulatory T cells participate inrepair of ischemic acute kidney injury. Kidney Int. 2009;76:717e29.
45. Kinsey GR, Li L, Okusa MD. Inflammation in acute kidneyinjury. Nephron Exp Nephrol. 2008;109:e102e7.
46. Kim MG, Lee SY, Ko YS, Lee HY, Jo SK, Cho WY, et al.CD4þ CD25þ regulatory T cells partially mediate thebeneficial effects of FTY720, a sphingosine-1-phosphateanalogue, during ischaemia/reperfusion-induced acute kidneyinjury. Nephrol Dial Transplant. 2011;26:111e24.
47. Roncador G, Brown PJ, Maestre L, Hue S, Martinez-Torrecuadrada JL, Ling KL, et al. Analysis of FOXP3 proteinexpression in human CD4þCD25þ regulatory T cells at thesingle-cell level. Eur J Immunol. 2005;35:1681e91.
48. Fontenot JD, Gavin MA, Rudensky AY. Foxp3 programs thedevelopment and function of CD4þCD25þ regulatory Tcells. Nat Immunol. 2003;4:330e6.
49. O’Garra A, Vieira P. Twenty-first century Foxp3. NatImmunol. 2003;4:304e6.
50. Schubert LA, Jeffery E, Zhang Y, Ramsdell F, Ziegler SF.Scurfin (FOXP3) acts as a repressor of transcription andregulates T cell activation. J Biol Chem. 2001;276:37672e9.
51. Lee H, Nho D, Chung HS, Shin MK, Kim SH, Bae H.CD4þCD25þ regulatory T cells attenuate cisplatin-inducednephrotoxicity in mice. Kidney Int. 2010;78:1100e9.
52. Liu M, Chien CC, Burne-Taney M, Molls RR, Racusen LC,Colvin RB, et al. A pathophysiologic role for T lymphocytes inmurine acute cisplatin nephrotoxicity. J Am Soc Nephrol.2006;17:765e74.
53. Hu J, Zhang L, Wang N, Ding R, Cui S, Zhu F, et al.Mesenchymal stem cells attenuate ischemic acute kidneyinjury by inducing regulatory T cells through splenocyte in-teractions. Kidney Int. 2013;84:521e31.
54. Gonzalez-Rey E, Gonzalez MA, Varela N, O’Valle F, Her-nandez-Cortes P, Rico L, et al. Human adipose-derivedmesenchymal stem cells reduce inflammatory and T cell re-sponses and induce regulatory T cells in vitro in rheumatoidarthritis. Ann Rheum Dis. 2010;69:241e8.
55. Ye Z, Wang Y, Xie HY, Zheng SS. Immunosuppressive ef-fects of rat mesenchymal stem cells: involvement of
CD4þCD25þ regulatory T cells. Hepatobiliary Pancreat DisInt. 2008;7:608e14.
56. Selmani Z, Naji A, Zidi I, Favier B, Gaiffe E, Obert L, et al.Human leukocyte antigen-G5 secretion by humanmesenchymalstem cells is required to suppress T lymphocyte and natural killerfunction and to induce CD4þCD25highFOXP3þ regulatoryT cells. Stem Cells. 2008;26:212e22.
57. Prevosto C, Zancolli M, Canevali P, Zocchi MR, Poggi A.Generation of CD4þ or CD8þ regulatory T cells uponmesenchymal stem cell-lymphocyte interaction. Haematolog-ica. 2007;92:881e8.
58. Parekkadan B, Upadhyay R, Dunham J, Iwamoto Y,Mizoguchi E, Mizoguchi A, et al. Bone marrow stromal celltransplants prevent experimental enterocolitis and requirehost CD11bþ splenocytes. Gastroenterology. 2011;140:966e75.
59. Patel SA, Meyer JR, Greco SJ, Corcoran KE, Bryan M,Rameshwar P. Mesenchymal stem cells protect breast cancercells through regulatory T cells: role of mesenchymal stemcell-derived TGF-beta. J Immunol. 2010;184:5885e94.
60. Casiraghi F, Azzollini N, Cassis P, Imberti B, Morigi M,Cugini D, et al. Pretransplant infusion of mesenchymal stemcells prolongs the survival of a semiallogeneic heart transplantthrough the generation of regulatory T cells. J Immunol.2008;181:3933e46.
61. Madec AM, Mallone R, Afonso G, Abou Mrad E, Mesnier A,Eljaafari A, et al. Mesenchymal stem cells protect NOD micefrom diabetes by inducing regulatory T cells. Diabetologia.2009;52:1391e9.
62. Sun J, Han ZB, Liao W, Yang SG, Yang Z, Yu J, et al.Intrapulmonary delivery of human umbilical cord mesen-chymal stem cells attenuates acute lung injury by expandingCD4þCD25þ Forkhead Boxp3 (FOXP3)þ regulatory T cellsand balancing anti- and pro-inflammatory factors. CellPhysiol Biochem. 2011;27:587e96.
63. Wang X, Willenbring H, Akkari Y, Torimaru Y, Foster M, Al-Dhalimy M, et al. Cell fusion is the principal source of bone-marrow-derived hepatocytes. Nature. 2003;422:897e901.
64. Camargo FD, Finegold M, Goodell MA. Hematopoieticmyelomonocytic cells are the major source of hepatocytefusion partners. J Clin Invest. 2004;113:1266e70.
65. Kuo TK, Hung SP, Chuang CH, Chen CT, Shih YR,Fang SC, et al. Stem cell therapy for liver disease: parametersgoverning the success of using bone marrow mesenchymalstem cells. Gastroenterology. 2008;134:2111e21.
66. Aurich I, Mueller LP, Aurich H, Luetzkendorf J, Tisljar K,Dollinger MM, et al. Functional integration of hepatocytesderived from human mesenchymal stem cells into mouselivers. Gut. 2007;56:405e15.
67. Aurich H, Sgodda M, Kaltwasser P, Vetter M, Weise A,Liehr T, et al. Hepatocyte differentiation of mesenchymalstem cells from human adipose tissue in vitro promotes he-patic integration in vivo. Gut. 2009;58:570e81.
68. Jang YY, Collector MI, Baylin SB, Diehl AM, Sharkis SJ.Hematopoietic stem cells convert into liver cells within dayswithout fusion. Nat Cell Biol. 2004;6:532e9.
Supplementary data
Supplementary data related to this article can befound at http://dx.doi.org/10.1016/j.jcyt.2014.01.004.
Copyright © 2022 FDOKUMEN




















