
Production and Characterization of Murine IgA Monoclonal Antibodies to the Surface Antigens of...
14
VIROLOGY 225, 97–110 (1996) ARTICLE NO. 0578 Production and Characterization of Murine IgA Monoclonal Antibodies to the Surface Antigens of Rhesus Rotavirus ANNA MARIA GIAMMARIOLI,* ERICH R. MACKOW,† LUCIA FIORE,‡ HARRY B. GREENBERG,§ and FRANCO M. RUGGERI* ,1 *Laboratorio di Ultrastrutture and ‡Laboratorio di Virologia, Istituto Superiore di Sanita ` , Viale Regina Elena 299, 00161 Rome, Italy; †Department of Medicine, State University of New York at Stony Brook, Stony Brook, New York 11794; and §Department of Medicine, Microbiology and Immunology, Stanford University School of Medicine, Stanford, California 94305, and Palo Alto Veterans Affairs Medical Center, Palo Alto, California 94304 Received June 10, 1996; accepted August 30, 1996 Rotavirus is the single most important cause of severe diarrhea in humans and is diffuse in most animal species worldwide, and an understanding of the antigenic properties of the virus is essential to the design of rational vaccine strategies. To better understand the localization of viral epitopes involved in antibody-mediated neutralization of virus infectivity, we have orally immunized mice with live rhesus rotavirus (RRV) and generated a panel of hybridoma cell clones secreting IgA class monoclonal antibodies. A total of 12 neutralizing IgA MAbs to VP4 and VP7 proteins were studied for their epitope specificity and topographical relationships by hemagglutination-inhibition assays, neutralization assays, and competitive-binding assays with previously mapped MAbs. In addition, neutralization-escape virus mutants were selected and gene segments for each variant were cloned and sequenced. Two IgA MAbs were found to be directed to the antigenic region A of the VP7 protein at amino acid 94, and 10 MAbs were directed at the VP8 trypsin cleavage fragment of VP4. Five of the VP4-specific MAbs identified the same neutralization epitope on the RRV VP8 protein, not previously associated with RRV neutralization. All neutralization-escape variants selected by this antibody group contained mutations at amino acids 132 – 135 of VP4. One IgA MAb selected for a mutation at amino acid 190 of VP4, and the corresponding viral mutant failed to agglutinate erythrocytes. This MAb mapped to an epitope recognized by 2 additional IgA MAbs. These results suggest that oral immunization of mice with RRV elicits an IgA immune response which is predominantly directed toward antigenic determi- nants on the VP8 portion of VP4. As a consequence, the route of immunization may alter immunodominant neutralization responses elicited to rotavirus. q 1996 Academic Press, Inc. INTRODUCTION antigenic neutralization specificity of the VP7 protein. More recently, the role of the VP4 spike protein in neu- Rotavirus is a leading cause of acute dehydrating tralization and the existence of multiple antigenic diarrhea particularly among young children in both de- forms of VP4 have been presented (Matsui et al., 1989; veloping and industrialized countries (Kapikian and Taniguchi et al., 1989). This, together with the indepen- Chanock, 1990). Infection is also widespread among dent segregation of the genes for VP7 and VP4 neutral- animals, representing a threat in husbandry particu- ization proteins (Gorziglia et al., 1990; Hoshino et al., larly for cattle and swines (Saif et al., 1994). The high 1985), has led to the adoption of a binarian nomencla- morbidity and mortality associated with human rotavi- ture for defining rotavirus strains using both the G rus infection make it necessary to develop a safe and (VP7) and the P (VP4) specificities (Nagesha and effective vaccine. Since the discovery of rotavirus Holmes, 1991). many studies have defined the protective role of anti- VP4 is an 85-kDa protein encoded by gene segment bodies against rotavirus infection and the existence of 4 which is present on the virion as spikes extending from neutralization epitopes on both outer capsid proteins, the VP7 matrix of the outer capsid (Prasad et al., 1988). VP4 and VP7 (Kalica et al., 1981; Greenberg et al., The VP4 protein is responsible for the hemagglutination 1983a,b; Hoshino et al., 1985; Taniguchi et al., 1985; property shared by several rotavirus strains (Mackow et Offit and Blavat, 1986a). VP7 is a 42-kDa glycoprotein al., 1989). Trypsin treatment of complete viral particles encoded by gene segments 7–9, and represents the results in cleavage of VP4 into VP8 (28 kDa) and VP5 major outer capsid structural protein and the primary (60 kDa), components which remain associated with the neutralization antigen of the virus (Bastardo et al., virion and are required for the enhancement of rotavirus 1981). Original rotavirus serotypes were based on the infectivity (Estes et al., 1981). In a recent study, we have shown that neutralizing monoclonal antibodies (MAbs) to the VP8 peptide efficiently prevent binding of rhesus 1 To whom correspondence and reprint requests should be ad- dressed. Fax: /39(6)49387140; E-mail: [email protected]. rotavirus (RRV) to fetal monkey kidney MA104 cells in 97 0042-6822/96 $18.00 Copyright q 1996 by Academic Press, Inc. All rights of reproduction in any form reserved.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Production and Characterization of Murine IgA Monoclonal Antibodies to the Surface Antigens of...

VIROLOGY 225, 97–110 (1996)ARTICLE NO. 0578
Production and Characterization of Murine IgA Monoclonal Antibodies to theSurface Antigens of Rhesus Rotavirus
ANNA MARIA GIAMMARIOLI,* ERICH R. MACKOW,† LUCIA FIORE,‡ HARRY B. GREENBERG,§and FRANCO M. RUGGERI*,1
*Laboratorio di Ultrastrutture and ‡Laboratorio di Virologia, Istituto Superiore di Sanita, Viale Regina Elena 299, 00161 Rome, Italy; †Departmentof Medicine, State University of New York at Stony Brook, Stony Brook, New York 11794; and §Department of Medicine, Microbiology and
Immunology, Stanford University School of Medicine, Stanford, California 94305, and Palo Alto VeteransAffairs Medical Center, Palo Alto, California 94304
Received June 10, 1996; accepted August 30, 1996
Rotavirus is the single most important cause of severe diarrhea in humans and is diffuse in most animal species worldwide,and an understanding of the antigenic properties of the virus is essential to the design of rational vaccine strategies. Tobetter understand the localization of viral epitopes involved in antibody-mediated neutralization of virus infectivity, we haveorally immunized mice with live rhesus rotavirus (RRV) and generated a panel of hybridoma cell clones secreting IgA classmonoclonal antibodies. A total of 12 neutralizing IgA MAbs to VP4 and VP7 proteins were studied for their epitope specificityand topographical relationships by hemagglutination-inhibition assays, neutralization assays, and competitive-binding assayswith previously mapped MAbs. In addition, neutralization-escape virus mutants were selected and gene segments for eachvariant were cloned and sequenced. Two IgA MAbs were found to be directed to the antigenic region A of the VP7 proteinat amino acid 94, and 10 MAbs were directed at the VP8 trypsin cleavage fragment of VP4. Five of the VP4-specific MAbsidentified the same neutralization epitope on the RRV VP8 protein, not previously associated with RRV neutralization. Allneutralization-escape variants selected by this antibody group contained mutations at amino acids 132–135 of VP4. OneIgA MAb selected for a mutation at amino acid 190 of VP4, and the corresponding viral mutant failed to agglutinateerythrocytes. This MAb mapped to an epitope recognized by 2 additional IgA MAbs. These results suggest that oralimmunization of mice with RRV elicits an IgA immune response which is predominantly directed toward antigenic determi-nants on the VP8 portion of VP4. As a consequence, the route of immunization may alter immunodominant neutralizationresponses elicited to rotavirus. q 1996 Academic Press, Inc.
INTRODUCTION antigenic neutralization specificity of the VP7 protein.More recently, the role of the VP4 spike protein in neu-
Rotavirus is a leading cause of acute dehydrating tralization and the existence of multiple antigenicdiarrhea particularly among young children in both de- forms of VP4 have been presented (Matsui et al., 1989;veloping and industrialized countries (Kapikian and Taniguchi et al., 1989). This, together with the indepen-Chanock, 1990). Infection is also widespread among dent segregation of the genes for VP7 and VP4 neutral-animals, representing a threat in husbandry particu- ization proteins (Gorziglia et al., 1990; Hoshino et al.,larly for cattle and swines (Saif et al., 1994). The high 1985), has led to the adoption of a binarian nomencla-morbidity and mortality associated with human rotavi- ture for defining rotavirus strains using both the Grus infection make it necessary to develop a safe and (VP7) and the P (VP4) specificities (Nagesha andeffective vaccine. Since the discovery of rotavirus Holmes, 1991).many studies have defined the protective role of anti- VP4 is an 85-kDa protein encoded by gene segmentbodies against rotavirus infection and the existence of 4 which is present on the virion as spikes extending fromneutralization epitopes on both outer capsid proteins, the VP7 matrix of the outer capsid (Prasad et al., 1988).VP4 and VP7 (Kalica et al., 1981; Greenberg et al., The VP4 protein is responsible for the hemagglutination1983a,b; Hoshino et al., 1985; Taniguchi et al., 1985; property shared by several rotavirus strains (Mackow etOffit and Blavat, 1986a). VP7 is a 42-kDa glycoprotein al., 1989). Trypsin treatment of complete viral particlesencoded by gene segments 7 – 9, and represents the results in cleavage of VP4 into VP8 (28 kDa) and VP5major outer capsid structural protein and the primary (60 kDa), components which remain associated with theneutralization antigen of the virus (Bastardo et al., virion and are required for the enhancement of rotavirus1981). Original rotavirus serotypes were based on the infectivity (Estes et al., 1981). In a recent study, we have
shown that neutralizing monoclonal antibodies (MAbs)to the VP8 peptide efficiently prevent binding of rhesus1 To whom correspondence and reprint requests should be ad-
dressed. Fax: /39(6)49387140; E-mail: [email protected]. rotavirus (RRV) to fetal monkey kidney MA104 cells in
970042-6822/96 $18.00Copyright q 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.
AID VY 8224 / 6a21$$$341 10-14-96 13:36:12 vira AP: Virology

98 GIAMMARIOLI ET AL.
culture (Ruggeri and Greenberg, 1991). Although this MATERIALS AND METHODSfinding suggests that VP8 is responsible for viral attach-
Cells and virusment to cellular receptors, the amino acids accountingfor this function are still unknown. MA104 cells were grown in Medium 199 supple-
mented with 7% fetal bovine serum (FBS) at 377 in a 5%Most information on the antigenic response to rotavi-rus is derived from parenterally immunized mice by CO2 atmosphere. The virus used in all experiments was
the rhesus rotavirus strain RRV. Production of virusstudying the elicited hyperimmune sera and monoclonalantibodies (IgG, IgM). Studies of parenterally immunized stocks was carried out in MA104 cell monolayers in 175-
cm2 flasks infected at a m.o.i. of 0.05. Virus was activatedmice emphasize the immunogenicity and immunodomi-nance of VP7 neutralization epitopes (Kalica et al., 1981; with 5 mg/ml trypsin (type IX; Sigma Chemical Co., St.
Louis, MO) at 377 for 1 hr before infection, and was grownGreenberg et al., 1983a; Gerna et al., 1984; Lazdins et al.,1985; Sabara et al., 1985). However, increasing evidence in serum-free medium containing 0.5 mg/ml trypsin. When
approximately 80% of the cells showed cytopathic effect,suggests that in both natural and experimental infectionsVP4 may be a more effective antigen than VP7 (Shaw et monolayers were freeze–thawed thrice and fluorocarbon
extracted.al., 1987; Svensson et al., 1987; Ward et al., 1993). Also,rotavirus is targeted to mature cells of the gut mucosa
Virus purificationduring infection and, as a result, immunity is thought tobe primarily provided by a secretory IgA antibody re- RRV-infected cell lysates were ultracentrifuged atsponse. For several viruses, resistance to infection 35,000 rpm in a Beckman Ti45 rotor for 2 hr at 47. Theseems to correlate with the presence of specific IgA pellet was resuspended in 10 mM Tris, 100 mM NaCl,antibody in mucosal secretions (Mills et al., 1971; Liew 2 mM CaCl2 , pH 7.4 (TNC), and layered onto a 25%et al., 1984). Renegar and Small (1991) have recently sucrose cushion in TNC. After centrifugation in a Beck-demonstrated that immunity to influenza virus could be man SW 41 at 35,000 rpm for 2 hr at 47, the virus pelletabrogated by intranasal application of anti-IgA antibodies was resuspended in 1 to 2 ml of TNC, and either usedbut not anti-IgG or -IgM antibodies consistent with IgA directly for immunization of mice or subjected to furtherbeing the major mediator of viral neutralization. Secretory purification. In the latter case, the virus suspensionIgA is a polymeric immunoglobulin which is actively was made up to 6 ml in TNC containing CsCl at atransported through the mucosal epithelia by a mecha- concentration corresponding to an average density ofnism which is not available to monomeric immunoglobu- 1.37 g/ml. After centrifugation at 33,000 rpm in an SWlins like IgG (Solari and Kraehenbhul, 1985). Transport 41 rotor for 20 hr at 127, the bands corresponding tois mediated by the polymeric immunoglobulin receptor both triple-shelled (TLP) and double-shelled (DLP) viral(pIgR) which is largely expressed on the mucous mem- particles were diluted with TNC and centrifugedbrane epithelial cells. The complex of pIgR and dimeric through a sucrose cushion as described before. Puri-IgA (dIgA) is endocytosed and transported through vesic- fied TLP and DLP particles were resuspended in 1 mlular compartments to the apical surface of cells (transcy- of TNC and stored at 47 before use.tosed), where proteolysis of pIgR results in release ofdIgA bound to the ectoplasmic domain of pIgR (known Immunization of miceas secretory component). It has been suggested that
Balb/c mice were supplied by Charles River Labora-transcytosed IgA antibodies may interfere with viral repli-tories. Both adult (6 weeks) female mice and 6-day-oldcation by binding to newly synthesized viral proteinspups from rotavirus serum-antibody-negative dams werewithin infected cells, thus providing a novel mechanismorally primed with 100 ml of a semipurified RRV prepara-of mucosal intracellular virus neutralization (Burns et al.,tion. After 3 weeks, mice were further fed with RRV in1996; Kaetzel et al., 1991; Mazanek et al., 1992).the presence of 5 to 10 mg of purified cholera toxin (kindlyIn this study, we have addressed the secretory IgAdonated by A. Gallina, Rome) using sodium bicarbonateimmune response to oral rotavirus administration. We(6%) to neutralize the acidic pH of the stomach. Approxi-have defined the neutralization specificity of IgA classmately 12 days after boosting, serum samples weremonoclonal antibodies elicited in response to oral RRVtaken from immunized animals and tested for the pres-immunization of mice. Our results suggest that oral rota-ence of neutralizing (Nt) and hemagglutination-inhibitingvirus immunization elicits a strong neutralizing IgA mono-(HI) antibody to RRV. Most mice had an Nt or HI antibodyclonal antibody response to immunodominant domainstiter ranging from 1:800 to 1:3200 after the second boost.on the VP8 portion of RRV VP4. These results indicate
that the route of immunization is a key factor in the evalu-Fusion
ation of mucosal neutralizing antibody responses andthat the enhancement of immune responses to secretory Mice showing a seroconversion to RRV received a
final oral boost along with an intravenous injection ofIgA-specific neutralization domains needs to be consid-ered in rational vaccine development. virus 4 days before fusion. Animals were sacrified, and
AID VY 8224 / 6a21$$$342 10-14-96 13:36:12 vira AP: Virology

99IgA MONOCLONAL ANTIBODIES TO ROTAVIRUS SURFACE ANTIGENS
the small bowel, the mesenteric lymph nodes, and the Enzyme-linked immunosorbent assayspleen were resected. Preparation of spleens for fusion For analysis of hybridoma supernatants, microtiterwas performed as previously described by Greenberg ELISA plates were incubated overnight with 50 ml/welland co-workers (1983b). The gut was thoroughly of purified DS-RRV diluted 1:100 in PBS, and blocked withwashed using sterile medium (DMEM) supplemented either 1% gelatin or 5% FBS in PBS for 1 hr at 377. Wellswith 200 mg/ml gentamicin and 5 mg/ml Fungizone, and were incubated with a 1:10 dilution of hybridoma cellthe Peyer’s patches were dissected with a fine scalpel supernatants in PBS containing 5% bovine serum albu-and homogenized with a dounce homogenizer together min (BSA) for 2 hr at 377. After washing, plates werewith mesenteric lymph nodes. The resulting cell sus- incubated with a 1:2000 dilution of an anti-mouse IgA (a-pension was centrifuged through an FBS cushion at chain) antibody conjugated with alkaline phosphatase1000 rpm for 8 min in a Beckman T-J6 centrifuge. Lym- (Sigma) for 1 hr at 377. Duplicate plates were examinedphocytes were counted using a vital stain, and the con- for IgG and IgM MAbs, and secondary incubation wascentration of cells was adjusted to about 109/ml. Sepa- effected with a 1:2000 dilution of anti-mouse IgG (H/L)rate fusions were carried out simultaneously for spleen antibody (Sigma). Reaction was then developed with p-cells and cells from regional lymphoid tissues, using the nitrophenol phosphate (Sigma 104) in 10 mM diethano-same procedure (Greenberg et al., 1983b). Hybridoma lamine, pH 10. After 1-hr incubation at 377, optical densi-supernatants were screened using an HI assay with ties were measured at a wavelength of 405 nm using ahuman group O erythrocytes, a focus-reduction micro- Bio-rad ELISA reader. Absorbance values equal to orneutralization test (FRN), and an enzyme-linked immu- higher than fivefold the background were considered tonosorbent assay (ELISA) using purified TLP-RRV as cap- be positive. In particular assays, ELISA plates wereture antigen and an alkaline phosphatase-conjugated coated with DLP-RRV or with a 1:100 dilution of baculovi-anti-mouse IgA (a-chain) or IgG (H/L) secondary anti- rus-expressed RRV VP8 (Fiore et al., 1991) or VP5(1) (L.body (see below). Selected cultures were cloned by Fiore, unpublished).limiting dilution using a mouse thymocyte feeder layer,and MAbs were amplified in mouse ascites. Confirma- Western blottingtion of MAb isotype was performed by ELISA using a
Semipurified RRV preparations were subjected tomouse antibody isotyping kit (Pierce, Rockford, IL).SDS–PAGE under denaturing conditions, and viral pro-tein bands were transferred to nitrocellulose paper using
Radioimmunoprecipitation (RIPA) a semidry apparatus (Bio-Rad) according to the manufac-turer’s instructions. After blocking with 5% skim milk inPreparation of 35S-labeled RRV-infected MA104 cellPBS for 1 hr at 47, blots were incubated with a 1:100lysates was performed as previously describeddilution of ascites fluids in PBS–1% BSA for 3 hr at 47.(Greenberg et al., 1983b). A conventional immunoprecipi-Secondary incubation was carried out with anti-mousetation assay using immobilized protein A could not beIgA or IgG or anti-guinea pig IgG antibodies conjugatedused because of the lack of reactivity of our IgA MAbswith alkaline phosphatase. After washing, detection ofwith protein A. Therefore, we have adopted a methodbands was performed by incubation with bromochloroin-similar to that formerly described by Burns and co-work-dolyl phosphate/nitroblue tetrazolium in 100 mM Tris, 100ers (1988). Wells of a high-binding ELISA plate (Immuno-mM NaCl, 5 mM MgCl2 (pH 9.5).lon 2; Dynatech Laboratories, Inc., Chantilly VA) were
antibody coated by overnight incubation at 47 with a 1:100 Hemagglutination-inhibition assaydilution of ascites fluid in phosphate-buffered saline
HI assay was performed as previously described (Kal-(PBS). After washing with PBS containing 0.5% Tween 20ica et al., 1983). Briefly, 25 ml of a 1:10 dilution of hybrid-(PBS/T), the plates were blocked with 1% gelatin (Bio-oma cell supernatants for screening of fusions or serialRad Laboratories, Richmond, CA) in PBS for 1 hr at 47. Intwofold dilutions of selected MAbs in PBS/0.5% BSA werethe case of guinea pig or mouse control sera, to increasemixed with equal volumes of RRV suspension containingantibody binding the wells were precoated with soluble8 hemagglutinating units in 96-well plates. After 1-hr incu-protein A (10 mg/ml in PBS). Fifty microliters of radiola-bation at room temperature, 50 ml of human group Obeled lysate were then added to each well, and the plateserythrocytes at the concentration of 0.4% in PBS/BSAwere further incubated overnight at 47 on an orbitalwere added to each reaction. HI titers were calculated asshaker. After extensive washing with RIPA buffer (10 mMthe reciprocal of the highest antibody dilution preventingTris–HCl, 100 mM NaCl, 600 mM KCl, 0.2 mM EDTA, 1%hemagglutination.Triton X-100; pH 7.6), wells were incubated with 70 ml of
electrophoresis sample buffer (Laemmli, 1970) for 1 minEpitope competition ELISAon a heated metal block. SDS–polyacrylamide gel elec-
trophoresis (PAGE) and autoradiography were performed This assay was used to determine the ability of theIgA MAbs to compete with IgG MAbs of known epitopeas previously described (Greenberg et al., 1983b).
AID VY 8224 / 6a21$$$342 10-14-96 13:36:12 vira AP: Virology

100 GIAMMARIOLI ET AL.
specificity for binding to RRV. TLP-RRV-coated ELISA sence of antibody. Neutralization was considered signifi-cant when there was a 60% or greater reduction in plaqueplates were incubated with serial twofold dilution (50 ml/
well) of either ascites fluid containing IgA MAbs or puri- number.fied IgG or IgM preparations (Shaw et al., 1986; Mackow
Production of RRV antigenic variantset al., 1988a,b) in PBS/BSA for 1 hr at 377. Twenty-fivemicroliters of an appropriate dilution of each challenge Neutralization-resistant RRV variants were selected inIgG and IgM or IgA antibody was then added, and incuba- MA104 cell monolayers by direct plaquing of 107 to 108
tion was continued for 45 min. After rinsing with PBS/T, PFU of RRV previously neutralized with each single IgAan alkaline phosphatase-conjugated anti-mouse IgG (g- MAb and including the antibody in the overlay mediumchain), IgM (m-chain), or IgA (a-chain) diluted to 1:2000 during growth, as described in detail by Shaw and co-was added, respectively. Plates were incubated for 1 workers (1986). In some cases, a preliminary amplifica-hr at 377 and washed, and reaction was developed as tion of virus in liquid medium in the presence of antibodydescribed above. Antigen, detecting antibody, and chal- was needed prior to plaquing under an agarose-con-lenge antibody concentrations were all adjusted to give taining overlay. Single well-developed viral plaques werean OD405 of 0.8–1.2 in the absence of competition. After picked from plates, and subjected to at least two furthersubtraction of background (usually õ0.080 in wells ex- cycles of plaque purification before cell culture expan-posed to all reagents except the challenge antibody), sion in antibody-free medium. Identification of neutraliza-reduction of test OD values higher than 70% compared tion-escape RRV mutants was eventually confirmed byto control wells was considered indicative of a significant neutralization and HI tests. Virus variants were desig-competition between each pair of antibodies tested. nated with the same code of the selecting MAb preceded
by a ‘‘v’’.Focus reduction neutralization assays
Nucleotide sequencingAn immunoperoxidase staining technique for detectinginfected cells was used in a microneutralization assay Plus-stranded RNAs of RRV and neutralization-escapeas described previously (Shaw et al., 1985, 1986). Approx- mutants were synthesized in vitro using the endogenousimately 200 focus-forming units of RRV were mixed either RNA polymerase activity of DLP particles, as describedwith a 1:10 dilution of hybridoma cell supernatants for previously (Flores et al., 1982). After phenol extraction,screening of fusions or with serial dilutions of MAbs, and ssRNA was precipitated with 2 M LiCl in an ice baththe mixtures were incubated for 60 min at 377 prior to overnight, resuspended in H2O, and further precipitatedinoculation onto MA104 cells in 96-well plates. Infected in 70% ethanol in the presence of 0.3 M sodium acetatecells were fixed 18 hr later with cold methanol, and incu- at 0707. Genes 4 and 9 were sequenced directly frombated with a guinea pig hyperimmune antiserum to RRV mRNA by Sanger’s method essentially as previously de-(1:1000) followed by incubation with a peroxidase-conju- scribed (Sanger et al., 1977; Zagursky et al., 1985), usinggated antibody to guinea pig Ig (1:2000). Staining was avian myeloblastosis virus reverse-transcriptase (Seike-performed by reaction with aminoethylcarbazole in 50 gaku, Inc.) and a series of negative-strand oligonucleo-mM acetate buffer, pH 6. Neutralizing titers were ex- tide primers spanning the whole length of either gene atpressed as the inverse dilution resulting in a 60% or intervals of 200 to 250 nucleotides (Mackow et al.,higher reduction in the number of infected MA104 cells. 1988a,b). To verify sequence in regions of RNA with a
high degree of secondary structure, additional plus-Plaque reduction neutralization assay stranded primers were used to sequence cDNA synthe-
sized by reverse transcription of viral mRNAs and PCRTrypsin-activated RRV was diluted with 1% BSA inamplification. Specific DNA regions were sequenced di-M199 to give a concentration of approximately 108 PFU/rectly or following cloning into pCRscript (Stratagene,ml. Aliquots of the viral suspension were then reactedInc.) with Sequenase 2.0 (U.S. Biochemical Corp., Cleve-with each MAb through a wide range of ascites dilutions.land, Ohio).Neutralization was allowed to take place for 2 hr at 377
and overnight at 47; each reaction mixture was then seri-RESULTSally diluted and plated onto MA104 cells in 6-well plates
in duplicate. After 90-min adsorption at room tempera- Hybridoma screeningture, monolayers were washed three times and overlaidwith M199 containing 0.5 mg/ml trypsin and 0.55% aga- A total of 73 hybridoma cell culture supernatants from
three separate fusions were found to react with RRV inrose (SeaKem; FMC BioProducts, Rockland, ME). Afterincubation at 377 for 3 days in a CO2 atmosphere, plaques an ELISA assay specific for murine IgA antibody. Of
these, 33 supernatants also showed HI and neutralizingwere stained with neutral red and counted. Data wereexpressed as the percentage of nonneutralized virus activity at the initial screening. Twelve neutralizing and
6 nonneutralizing cultures were randomly chosen for fur-compared to control wells infected with virus in the ab-
AID VY 8224 / 6a21$$$342 10-14-96 13:36:12 vira AP: Virology

101IgA MONOCLONAL ANTIBODIES TO ROTAVIRUS SURFACE ANTIGENS
TABLE 1
Reactivity of IgA MAbs with Triple- and Double-Shelled RRV Particles and Baculovirus-Expressed RRV Proteins by ELISA
ELISA ODa values toMonoclonal RRV protein
antibody Ig class HI titer assigned TLP-RRV DLP-RRV VP8 VP5
MAb 1C4 IgA 51,200 VP8 0.9 0.2 2.0 0.1MAb 1E4 IgA 102,400 VP8 1.1 0.1 2.1 0.1MAb 2A10 IgA 102,400 VP8 0.6 0.0 1.5 0.0MAb 2B12 IgA 51,200 VP8 0.9 0.2 1.5 0.1MAb 3F12 IgA 204,800 VP8 0.5 0.2 1.3 0.2MAb 3G6 IgA 409,600 VP8 0.7 0.0 1.4 0.0MAb 4B6 IgA 102,400 VP8 0.6 0.0 1.9 0.2MAb 4B7 IgA 204,800 VP8 0.8 0.1 1.6 0.2MAb 4C2 IgA 204,800 VP8 0.6 0.0 1.4 0.1MAb 5E2 IgA 25,600 VP8 0.5 0.1 1.7 0.0MAb 1C3 IgA — VP5 0.9 0.2 0.1 1.7MAb 4G12 IgA 25,600 VP7 0.7 0.1 0.2 0.2MAb 5B9 IgA 25,600 VP7 0.8 0.2 0.2 0.2MAb 1D4 IgA — VP6 1.3 2.3 0.2 0.2MAb 2C5 IgA — VP6 1.9 2.2 0.3 0.2MAb 2F7 IgA — VP6 1.3 1.7 0.1 0.1MAb 2F8 IgA — VP6 0.8 1.5 0.1 0.1MAb 3H5 IgA — VP6 1.4 2.0 0.2 0.2MAb 2G4 IgG 25,600 VP5b 2.2 0.2 0.1 1.4MAb 7A12 IgG 51,200 VP8b 2.2 0.1 2.6 0.1MAb 159 IgG 25,600 VP7b 2.8 0.4 0.2 0.3Mouse, serum 6.400 ú3.0 ú3.0 ú3.0 2.6
a Significant optical densities are indicated in bold.b References on control MAbs are indicated under Materials and Methods.
ther study and cloned thrice, and MAbs were amplified Two further neutralizing MAbs (4G12 and 5B9) did notreact with VP4 subfragments; however, these antibodiesas murine ascites. All MAbs were confirmed to be IgA
by an ELISA assay using class- and subclass-specific strongly stained wells coated with TLP-RRV but not DLP-RRV. Although to a lesser extent than TLP-RRV, controlantisera to mouse immunoglobulins. Ascites fluids were
also analyzed by SDS–PAGE under nonreducing condi- IgG MAb 159 also stained DLP-RRV wells, which is prob-ably related to the higher sensitivity of the IgG–ELISAtions, and the gel was blotted onto nitrocellulose paper.
In all cases, staining with an alkaline phosphatase-conju- used. As DLPs differ from whole virions in that they lackboth outer capsid proteins VP4 and VP7, these findingsgated antiserum to mouse IgA revealed the presence of a
major protein band migrating with an apparent molecular suggested that MAbs 4G12 and 5B9 might recognizeVP7, present as a contaminant in the DLP-RRV gradientweight of 300,000 Da, and less evident bands at 150,000
and 60,000 Da (not shown). This indicates that most anti- fraction. The remaining 5 nonneutralizing MAbs (1D4,2C5, 2F7, 2F8, and 3H5) stained both TLP- and DLP-body in our preparations was dimeric.RRV-coated wells, but gave no reaction with any of theexpressed viral proteins.Determination of the specificity of MAbs for RRV
To confirm antigen specificity of IgA MAbs, ascitesproteinsfluids were used in a solid-phase immunoprecipitation
None of the selected IgA MAbs reacted with RRV pro- test as capture antibodies for radiolabeled viral proteinsteins in Western blots (not shown), suggesting that they from RRV-infected cell lysates. Since our IgAs did notrecognize conformation-dependent epitopes. For 11 bind protein A-activated beads, this assay was used asMAbs, identification of the protein specificity could be an alternative method to a conventional RIPA, which usesestablished by using baculovirus-expressed RRV VP4 immobilized protein A for immune complex capture. Re-subfragments VP8 and VP5(1) as capture antigens in an sults of this assay are shown in Fig. 1. Consistent withELISA assay (Table 1). Ten of the 12 neutralizing antibod- the above experiments, all MAbs reacting with VP8 ories efficiently stained wells coated with VP8, but did not VP5(1) in ELISA immunoprecipitated radiolabeled VP4.react with RRV VP5(1). Conversely, MAb 1C3 stained Because trypsin cleavage of VP4 does not occur insideVP5(1), but not VP8. This antibody did not inhibit RRV infected cells, neither VP8 nor VP5 bands can be shown
in these experiments. A second protein band with a mo-hemagglutination or neutralize the virus in cell cultures.
AID VY 8224 / 6a21$$$343 10-14-96 13:36:12 vira AP: Virology

102 GIAMMARIOLI ET AL.
FIG. 1. SDS–PAGE and autoradiography of radiolabeled RRV-infected MA104 cell lysates immunoprecipitated with IgA monoclonal antibodies.Reference IgG MAbs are included: MAb 1A9 (VP8*), MAb 255/60 (VP6). For experimental details, see Materials and Methods.
lecular weight of about 65,000 Da was precipitated by all petitive binding assays are summarized in Tables 3 and4. No competition was found between antibodies di-VP4-specific MAbs, including control MAb 1A9 previously
mapped in the VP8 subfragment of RRV VP4. This protein rected at different RRV proteins. IgA MAbs 4G12 and 5B9showed a two-way competition with IgG antibodies 4F8was found in different preparations of RRV-infected ly-
sates, and probably represents a cytoplasmic proteolysis and 159 which map to the A antigenic region of RRV VP7.To a lesser extent, competition was also found with MAbproduct of native VP4. Five nonneutralizing IgA MAbs
(1D4, 2C5, 2F7, 2F8, and 3H5) selectively precipitated 5H3 that recognizes a distinct epitope of the same pro-tein. Among the VP8-specific IgA MAbs, at least fourVP6 from RRV lysates, whereas the two remaining neu-
tralizing MAbs (4G12 and 5B9) did not react with any distinct patterns of competition were observed with con-trol VP8 and VP5 IgG or IgM MAbs. Four IgA MAbs (2A10,RRV peptide. These latter antibodies were established
as being directed toward the VP7 protein by a competitive 3F12, 4B7, 4C2) showed no reactivity with any of thecontrol antibodies. IgA MAb 3G6 showed a one-way com-ELISA (see below) and by immunoperoxidase staining of
insect Sf-9 cells infected with recombinant baculovirus petition with IgM MAb M14, when the latter was usedas competing antibody. A further IgA MAb (5E2) showedexpressing RRV VP7 (Table 2) (the procedure was devel-
oped by Greenberg and co-workers and will be de- a two-way competition with IgG MAb 7A12 and was non-reciprocally competed by MAb M14. A completely distinctscribed in detail elsewhere; H. B. Greenberg et al., manu-
script in preparation). In this assay, a large number ofcontrol VP7 neutralizing MAbs were reactive (data not
TABLE 2shown).Immunoperoxidase Staining of Recombinant Baculovirus-Infected
Epitope competition ELISA Sf-9 Cells Expressing RRV Proteins by IgA mAbs
Competitive binding assays represent a common tech- Expressed proteinnique used to investigate the spatial relationship be- Monoclonal
antibody VP4 VP6 VP7 NSP2tween the epitopes recognized by different neutralizingVP4- and VP7-specific MAbs. The ability of IgA MAbs to
mAb 4G12 õ1000a õ1000 1,562,500 õ1000compete for binding to RRV with control IgG or IgM MAbsmAb 5B9 õ1000 õ1000 1,562,500 õ1000
of known epitope specificity was evaluated by using sec-ondary antibodies specific to the g-, m-, or a-chains of a Reciprocal of antibody dilution giving detectable staining of intracel-
lular antigen.mouse IgG, IgM, or IgA, respectively. The results of com-
AID VY 8224 / 6a21$$$343 10-14-96 13:36:12 vira AP: Virology
103IgA MONOCLONAL ANTIBODIES TO ROTAVIRUS SURFACE ANTIGENS
TABLE 3
ELISA Blocking Titers of Reference MAbs Tested in Competition with VP7-Specific IgA MAbs and RRV Antigens
Test IgA MAbCompeting Neutralization
MAb Ig class Region MAb 4G12 MAb 5B9 titer to RRV
MAb 60 IgG — õ200 õ200 õ200MAb 159 IgG A 6,400a 6,400 51,200MAb 4F8 IgG A 12,800 12,800 51,200MAb 5H3 IgG C 800b 400 12,800
a Competing antibody dilution giving more than 70% reduction in OD by test MAb.b Numbers in italic indicate nonreciprocal competition between antibodies.
pattern of reactivity was shown by three other VP8-IgA to resist neutralization by the IgA antibodies used fortheir selection.MAbs (1C4, 1E4, and 4B6) that presented a two-way com-
petition with three IgG antibodies (23, 1A9, 7A12) map-ping into a discontinuous major neutralization epitope of
Hemagglutination-inhibition analysis of neutralization-VP8. MAb 2B12 behaved slightly differently from the latter
escape mutants with MAbsIgA MAbs, in that it did not show competition with IgGMAb 7A12, whereas it competed with IgM MAb M14. Nine RRV variants, plaque isolated under the selective
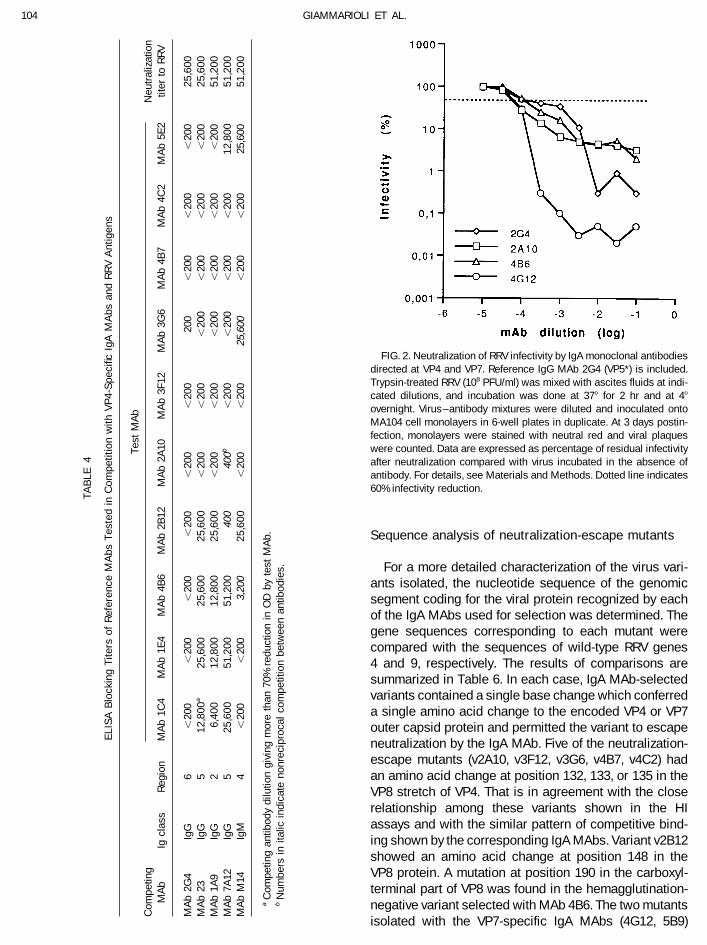
pressure of distinct IgA neutralizing MAbs, were exam-Neutralization curves of IgA MAbs ined by a HI test using a wide panel of monoclonal anti-
bodies (Table 5). Based on the results of assays, fourNeutralization curves were obtained for the 12 neu- variants (v2A10, v3G6, v4B7, and v4C2) could be as-
tralizing IgA MAbs by testing a wide range of ascites signed to a same major group of reactivity, all of themfluid dilutions against high titer RRV preparations in a being resistant to any of the corresponding MAbs.plaque inhibition assay. Figure 2 shows typical examples Among them, v4C2 showed only a partial resistance toof the curves obtained. All antibodies directed at VP8 MAb 2A10. All variants included in this group were alsocaused a reduction of virus infectivity in the order of 100- resistant to MAb 3F12. The viral mutant selected withfold or less in a range of ascites dilutions between 1:3200 this latter antibody (v3F12) proved only resistant to theand 1:32,000; further decrease of antibody dilution did homologous MAb, thus showing a one-way correlationnot result in higher rates of neutralization. A stronger with the former group of viruses. Hemagglutination byneutralization activity was shown by the two IgA MAbs mutant v5E2 was also resistant only to the 5E2 MAb. Into the VP7 protein, both of which were able to reduce addition to the MAb used for its selection, the VP8-spe-virus titer by about 4 log10 . Anti-VP8 and -VP7 IgA MAbs cific virus mutant v2B12 also showed lack of reactivityshowed neutralization curves similar to those observed with the IgM MAb M14 directed at the VP8 portion ofwith IgG MAbs of similar protein specificities (Ruggeri VP4. With this unique exception, all of the above mutantsand Greenberg, 1991). None of the VP6- or VP5-specific were sensitive to all of the VP4 and VP7 reference MAbsIgA MAbs generated in this study had any effect on virus tested. Variants v4G12 and v5B9 showed cross-resis-infectivity in this assay at 1:10 dilutions of ascites. tance to the corresponding MAbs and to reference MAbs
159 and 4F8, whereas they were sensitive to MAb 5H3.Isolation of neutralization-escape RRV mutants
Ten virus variants were selected by overlaying RRV- Neutralization of viral mutant v4B6 by MAbsinfected MA104 cell monolayers with neutralizing IgA
Although grown to titers exceeding 5 1 107 PFU/ml,MAbs and isolating RRV mutants which escaped IgAthe virus variant selected with MAb 4B6 showed no hem-MAb neutralization. Despite any efforts, RRV viral variantsagglutination property. Therefore, its reactivity with otherwere not selected when VP8-specific MAbs 1C4 and 1E4MAbs was examined by a microplate neutralizationwere included into the infection medium. All of the iso-assay (Table 5). In addition to the homologous antibody,lated variants could be grown to similar titers as wild-v4B6 also fully resisted neutralization by IgA MAbs 1C4,type RRV, following replication on cell cultures, in the1E4, and 5E2, and by reference MAbs 23, 1A9, and 7A12.presence or absence of the IgA MAbs that selected them.It also showed a low degree of resistance to all re-All variants except one, v4B6, were also able to aggluti-maining MAbs to VP8, but not to antibodies directed atnate human group O erythrocytes. After several pas-
sages in MA104 cells, all mutants maintained the ability VP5 or VP7.
AID VY 8224 / 6a21$$$343 10-14-96 13:36:12 vira AP: Virology
104 GIAMMARIOLI ET AL.
FIG. 2. Neutralization of RRV infectivity by IgA monoclonal antibodiesdirected at VP4 and VP7. Reference IgG MAb 2G4 (VP5*) is included.Trypsin-treated RRV (108 PFU/ml) was mixed with ascites fluids at indi-cated dilutions, and incubation was done at 377 for 2 hr and at 47
overnight. Virus–antibody mixtures were diluted and inoculated ontoMA104 cell monolayers in 6-well plates in duplicate. At 3 days postin-fection, monolayers were stained with neutral red and viral plaqueswere counted. Data are expressed as percentage of residual infectivityafter neutralization compared with virus incubated in the absence ofantibody. For details, see Materials and Methods. Dotted line indicates60% infectivity reduction.
Sequence analysis of neutralization-escape mutants
For a more detailed characterization of the virus vari-ants isolated, the nucleotide sequence of the genomicsegment coding for the viral protein recognized by eachof the IgA MAbs used for selection was determined. Thegene sequences corresponding to each mutant werecompared with the sequences of wild-type RRV genes4 and 9, respectively. The results of comparisons aresummarized in Table 6. In each case, IgA MAb-selectedvariants contained a single base change which conferreda single amino acid change to the encoded VP4 or VP7outer capsid protein and permitted the variant to escapeneutralization by the IgA MAb. Five of the neutralization-escape mutants (v2A10, v3F12, v3G6, v4B7, v4C2) hadan amino acid change at position 132, 133, or 135 in theVP8 stretch of VP4. That is in agreement with the closerelationship among these variants shown in the HIassays and with the similar pattern of competitive bind-ing shown by the corresponding IgA MAbs. Variant v2B12showed an amino acid change at position 148 in theVP8 protein. A mutation at position 190 in the carboxyl-terminal part of VP8 was found in the hemagglutination-negative variant selected with MAb 4B6. The two mutants
TA
BLE
4
ELI
SA
Blo
ckin
gT
iters
ofR
efer
ence
MA
bsT
este
din
Com
petit
ion
with
VP
4-S
peci
ficIg
AM
Abs
and
RR
VA
ntig
ens
Test
MA
bC
ompe
ting
Neu
tral
izat
ion
MA
bIg
clas
sR
egio
nM
Ab
1C4
MA
b1E
4M
Ab
4B6
MA
b2B
12M
Ab
2A10
MA
b3F
12M
Ab
3G6
MA
b4B
7M
Ab
4C2
MA
b5E
2tit
erto
RR
V
MA
b2G
4Ig
G6
õ20
0õ
200
õ20
0õ
200
õ20
0õ
200
200
õ20
0õ
200
õ20
025
,600
MA
b23
IgG
512
,800
a25
,600
25,6
0025
,600
õ20
0õ
200
õ20
0õ
200
õ20
0õ
200
25,6
00M
Ab
1A9
IgG
26,
400
12,8
0012
,800
25,6
00õ
200
õ20
0õ
200
õ20
0õ
200
õ20
051
,200
MA
b7A
12Ig
G5
25,6
0051
,200
51,2
0040
040
0bõ
200
õ20
0õ
200
õ20
012
,800
51,2
00M
Ab
M14
IgM
4õ
200
õ20
03,
200
25,6
00õ
200
õ20
025
,600
õ20
0õ
200
25,6
0051
,200
aC
ompe
ting
antib
ody
dilu
tion
givi
ngm
ore
than
70%
redu
ctio
nin
OD
byte
stM
Ab.
bN
umbe
rsin
italic
indi
cate
nonr
ecip
roca
lco
mpe
titio
nbe
twee
nan
tibod
ies.
isolated with the VP7-specific IgA MAbs (4G12, 5B9)
AID VY 8224 / 6a21$$$343 10-14-96 13:36:12 vira AP: Virology
105IgA MONOCLONAL ANTIBODIES TO ROTAVIRUS SURFACE ANTIGENS
TA
BLE
5
Com
pari
son
ofH
IT
iters
ofV
P4-
and
VP
7-S
peci
ficM
onoc
lona
lA
ntib
odie
sA
gain
stV
iral
Ant
igen
icV
aria
nts
and
Par
enta
lR
RV
Vira
lva
riant
ste
sted
MA
bIg
clas
sP
rote
inv2
A10
v3F1
2v3
G6
v4B
7v4
C2
v2B
12v5
E2
v4G
12v5
B9
v4B
6a
MA
b2A
10Ig
AVP
80
90
10
90
90
31
01
10
3M
Ab
2B12
IgA
VP8
00
00
00
80
00
03
MA
b3F
12Ig
AVP
80
110
110
110
110
111
01
10
3M
Ab
3G6
IgA
VP8
011
00
110
110
110
01
00
04
MA
b4B
6Ig
AVP
80
00
01
01
01
00
11M
Ab
4B7
IgA
VP8
011
01
011
011
011
00
11
10
5M
Ab
4C2
IgA
VP8
010
00
100
100
101
01
10
5M
Ab
5E2
IgA
VP8
01
00
10
10
10
010
00
010
MA
b1C
4Ig
AVP
80
00
00
00
00
011
MA
b1E
4Ig
AVP
80
00
01
01
01
10
11M
Ab
4G12
IgA
VP7
00
00
01
00
80
80
1M
Ab
5B9
IgA
VP7
00
01
01
01
00
10
90
100
1M
Ab
2G4
IgG
VP5
01
00
10
00
00
01
MA
b23
IgG
VP8
00
00
00
00
00
9M
Ab
1A9
IgG
VP8
00
00
01
01
00
10M
Ab
7A12
IgG
VP8
01
00
01
01
10
8M
Ab
M14
IgM
VP8
01
11
10
81
11
03
MA
b15
9Ig
GVP
70
10
00
00
04
06
01
MA
b4F
8Ig
GVP
70
10
00
10
05
05
0M
Ab
5H3
IgG
VP7
01
00
00
00
10
10
Not
e.H
Itit
ers
wer
ede
term
ined
for
each
ofth
ein
dica
ted
mA
bsag
ains
tR
RV
neut
raliz
atio
n-es
cape
varia
nts
and
com
pare
dw
ithth
etit
ers
ofth
esa
me
mA
bsvs
pare
ntal
RR
V.Th
eva
lues
inth
eta
ble
repr
esen
tlo
g 2tit
ervs
varia
nt0
log 2
titer
svs
RR
V.D
iffer
ence
sin
titer
betw
een
varia
ntan
dpa
rent
alR
RVú
log 2
3.2
(10-
fold
)ar
ehi
ghlig
hted
inbo
ld.
aD
ata
for
v4B
6ar
efr
omFR
Nas
says
.
AID VY 8224 / 6a21$$822410-14-96 13:36:12 vira AP: Virology
106 GIAMMARIOLI ET AL.
TABLE 6
Gene 4 and 9 Nucleotide and Amino Acid Changes in RRV Variants Selected by Neutralizing IgA MAbs
RRV Gene Nucleotide Amino acidvariant (protein) change (bp No.) change (aa No.) Region
v2A10 4 (VP4) C-A (407) Ala-Asn (133) Newv3F12 4 (VP4) C-T (407) Ala-Val (133) Newv3G6 4 (VP4) T-G (403) Tyr-Asp (132) Newv4B7 4 (VP4) C-A (412) Gln-Lys (135) Newv4C2 4 (VP4) A-G (413) Gln-Arg (135) Newv2B12 4 (VP4) A-G (452) Gln-Arg (148) 4v4B6 4 (VP4) C-T (578) Ser-Leu (190) 5v4G12 9 (VP7) A-G (328) Asn-Asp (94) Av5B9 9 (VP7) A-G (329) Asn-Ser (94) A
showed a mutation of base 328 and 329, respectively, antigens including viruses (Lycke and Holmgren, 1986;corresponding to an amino acid change at position 94. Chen and Strober, 1990), was used in subsequent oral
administration of rotavirus. Hence, pups were primedwith an initial oral RRV dose which induced diarrhea, andDISCUSSIONwere boosted orally with a combination of RRV virions
A great deal of interest has been centered on defining and cholera toxin in order to induce secondary immunethe basis of immunity to rotaviruses due to the worldwide responses.clinical importance of these enteric pathogens. The pres-
The suitability of this immunization protocol appearsence of neutralizing antibody in the intestine has been
to be supported by the identification of a high level ofcorrelated with protection against rotavirus disease in
neutralizing antibodies to RRV in most murine sera takenseveral animal model systems (Snodgrass and Well,after a single booster dose of rotavirus. In addition, a1976; Saif et al., 1983; Offit et al., 1986b; Matsui et al.,proportion close to 50% of cell supernatants from primary1989). Detailed analyses of the antigenic domains in bothfusion cultures showed reactivity with RRV antigen inrotavirus neutralization proteins, VP4 and VP7, have beenan IgA-specific ELISA, suggesting a specific targeting toconducted with the use of monoclonal antibodies andimmune cells involved in secretory antibody response.viral neutralization-escape mutants (Taniguchi et al.,In fact, all MAbs tested were mostly produced in the form1987, 1988a; Burns et al., 1988; Shaw et al., 1988; Zhouof dimers, and therefore may be suitable for recognitionet al., 1994), and the presence of at least three neutraliza-by the poly-Ig receptor that mediates transport of secre-tion epitopes on VP4 and VP7 has been reported. Neu-tory antibodies through mucosal epithelia (Kraehenbuhltralizing antibodies of IgG or IgM immunoglobulinand Neutra, 1992).classes resulting from parenteral immunization of mice
Although the MAbs described in this paper representwith rotavirus have been used in these studies. However,only a sample of the cultures obtained, it is of interest thatit has been emphasized by several investigators (Dhara-10 of 12 neutralizing antibodies which were randomlykul et al., 1988; Losonsky et al., 1988; Conner et al., 1991;selected for the study were shown to be directed at VP4.Merchant et al., 1991; Coulson et al., 1992) that theseSince VP4 is stoichiometrically underrepresented relativehumoral responses are not likely to fully represent theto the VP7 on rotaviruses, this suggests that the VP4local immune mechanisms which mediate protection inprotein may be particularly efficient at eliciting neutraliz-the intestinal mucosa.ing immune responses at the intestinal mucosa (ShawIn this study, we have analyzed the neutralizing secre-et al., 1988; Ward et al., 1988, 1993) and, as opposed totory IgA response to rotavirus antigens and defined IgA-IgG immune response studies, suggests that the VP7specific neutralization sites in the VP4 and VP7 proteinsprotein may play a secondary role in mucosally derivedof RRV. As in natural infections, we have developed anneutralizing immunity. On the other hand, a possible con-oral immunization strategy that made it possible to iso-tribution of cholera toxin in enhancing the neutralizinglate a library of murine IgA monoclonal antibodies toimmune response to VP4 more than to VP7 cannot beRRV. First, we have taken advantage of the fact that,ruled out, even though no data are available to sustainalthough adult Balb/c mice are not susceptible to rotavi-the hypothesis of a differential stimulation of antigenicrus disease, diarrhea can be induced in pups up to 2responses by cholera toxin. Virus-specific IgA antibodiesweeks of age by oral administration of live virions (Rie-have also been shown to be produced by lamina propriapenhoff et al., 1982). Second, cholera toxin, which haslymphocytes as a result of parenteral immunization withbeen reported to strongly enhance mucosal immune re-
sponses to a great variety of soluble and particulated rotavirus (Coffin et al., 1995). Comparison of the epitope
AID VY 8224 / 6a21$$$343 10-14-96 13:36:12 vira AP: Virology

107IgA MONOCLONAL ANTIBODIES TO ROTAVIRUS SURFACE ANTIGENS
specificity of IgA responses following oral or parenteral Isolation of MAbs to this epitope has been reported forseveral animal and human strains of rotavirus (Mackowimmunization with rotavirus could provide definite infor-
mation on the importance of the inoculation route in de- et al., 1988b; Taniguchi et al., 1988b), and Shaw and co-workers (1987) have presented evidence of the directtermining the epitope pattern of the immune response.
Characterization of the two VP7-specific IgA mono- involvement of this epitope in protection among childrenvaccinated with RRV. Our results may indicate a relativelyclonal antibodies which select for variants with mutations
at amino acid 94 confirms previous findings of an anti- low immunogenicity of the VP5 heterotypic epitope in gutimmune responses and its underrepresentation follow-genically highly reactive epitope at this position (Dyall et
al., 1986; Mackow et al., 1988a; Taniguchi et al., 1988a). ing oral immunization.Immune responses to oral immunization identify VP8Neutralizing antibodies to this domain have been iso-
lated for both animal and human strains of rotavirus, and as the major immunogenic element of both VP4 andRRV. The VP8 peptide has been both shown to carrythe role of this epitope in in vivo protection has been
clearly demonstrated (Matsui et al., 1989). Both IgG and the viral hemagglutinin (Fiore et al., 1991) and sug-gested to play an important role in virus binding to hostIgM antibodies directed at this epitope have been shown
to exert a strong neutralizing effect on RRV and to act by cells (Ruggeri and Greenberg, 1991), and therefore itis likely to be highly accessible on the virion surface.single-hit kinetics (Ruggeri and Greenberg, 1991). Prelim-
inary experiments indicate that the neutralizing IgA MAbs From competitive binding studies and gene sequenceanalysis of viral neutralization-escape mutants, our IgAdescribed here are at least as efficient at neutralizing
RRV in vitro as previously described IgG and IgM MAbs MAbs to VP8 are directed toward at least three distinctneutralization epitopes. One of these, recognized by(Ruggeri and Greenberg, 1991). Some of the MAbs to the
aa 94 epitope have been shown to induce heterotypic IgA MAb 2B12, has been previously reported for RRVand mapped at amino acid 148 by a cross-reactiveneutralization and protection, whereas most others ap-
pear to be G-serotype specific (Dyall et al., 1986; Mackow neutralizing MAb (Mackow et al., 1988b). A secondgroup of three IgA MAbs (1C4, 1E4, 4B6) showed recip-et al., 1988a; Taniguchi et al., 1988a). Further studies are
necessary to verify whether our IgA MAbs are cross- rocal binding competition with a series of MAbs pre-viously described to select mutations at amino acidreactive and neutralize G3 serotype rotaviruses other
than RRV. residues 100, 188, and 194 (Mackow et al., 1988b; Zhouet al., 1994). Of them, only MAb 4B6 was successfullyBecause of the wide generation of neutralizing anti-
bodies to the aa 94 domain of VP7 obtained by parenteral used to isolate a neutralization-escape mutant (v4B6)which presents a mutation at amino acid 190. It is ofimmunization, the isolation of orally stimulated IgA MAbs
with this antigenic specificity suggests that the aa 94 interest that this virus mutant, although grown to highinfectious titers similarly to parental RRV, did not showneutralization site is of particular importance to viral neu-
tralization by all immunization routes. The presentation any ability to agglutinate erythrocytes, resembling theSA-11 neutralization-escape mutant to MAb 23 re-of the VP7 antigen in a properly folded conformation has
been acknowledged as being critical for inducing VP7- ported by Zhou and colleagues (1994). The MAb 23variant was also shown to carry a substitution of aminospecific neutralizing immune responses (Andrew et al.,
1992; Dormitzer et al., 1994). In fact, several studies using acid 194 of VP4, suggesting that residues 190 and 194are critical for HA function. In contrast, mutation identi-recombinant VP7 to induce immunity to rotavirus in ani-
mals have had limited success (Andrew et al., 1987, 1990; fied at amino acid 188, selected by MAb 7A12 (Mackowet al., 1988b), does not affect viral hemagglutination.Arias et al., 1986; McCrae and McCorquodale, 1987;
McGonigal et al., 1992). Our data indicate that an immu- These data also suggest that the HA domain and thecell-binding domain on rotavirus may be localized innologically reactive conformation of the VP7 protein is
presented to the mucosal immune system by oral immu- different sites on VP4, as previously hypothesized(Mendez et al., 1993).nization.
One major difference noted in the oral immune re- A strict functional correlation between distant portionsof the VP8 molecule is suggested by finding that variantsponse to RRV administration is the overwhelming neu-
tralizing IgA response to the VP4 protein, specifically to v4B6 is fully resistant to neutralization by IgG MAbs 23and 7A12 (selecting amino acid changes at 194 and 188,the 28-kDa amino-terminal trypsin cleavage fragment,
VP8. None of the neutralizing antibodies was directed at respectively) and by MAb 1A9, selecting amino acidchange at residue 100. Whether this indicates a closethe VP5 portion of VP4, which was previously shown to
contain an epitope involved in heterotypic neutralization spatial relation between the two epitopes on folded VP4in the virion or a conformational alteration induced by aand protection (Offit et al., 1986b; Shaw et al., 1986). The
VP5 cross-reactive neutralization epitope maps to the aa change in amino acid at a distant site is not known atpresent. However, it should be noted that v4B6 also393 region of VP4 by neutralization-escape mutants and
has been illustrated in a cryoelectron microscopy study showed some degree of resistance to neutralization byvirtually all the MAbs to VP8 tested suggesting a minorof the VP4 spikes of SA-11 rotavirus (Prasad et al., 1990).
AID VY 8224 / 6a21$$$343 10-14-96 13:36:12 vira AP: Virology

108 GIAMMARIOLI ET AL.
but more general alteration of the antigenic structure of ponent of humoral IgG neutralization responses to paren-teral immunization. As a result, unique VP4-specific intes-the protein. A close functional correlation with the 4B6
epitope seems to be shown by IgA MAbs 1C4 and 1E4 tinal secretory IgA responses to oral RRV vaccinationneed to be considered in the rational development ofwhich not only share a similar competitive binding pat-
tern with MAb 4B6 but were also unable to neutralize rotavirus vaccines.the v4B6 mutant. Interestingly, we were not able to isolatea neutralization-escape mutant to either MAb 1C4 or 1E4. ACKNOWLEDGMENTSIt is possible that the epitopes recognized by these MAbs
This research project received financial support from the WHO/are located in a site that is essential to the virion. A UNDP Programme for Vaccine Development. Dr. Harry B. Greenbergthird epitope not previously recognized on RRV VP8 is is VA Medical Investigator.identified by a group of five IgA MAbs selecting for mu-tants with amino acid changes at positions 132, 133, or REFERENCES135. This epitope appears to be totally unrelated to the
Andrew, M. E., Boyle, D. B., Coupar, B. E., Reddy, D., Bellamy, A. R.,antigenic domains previously described on RRV by Shaw and Both, G. W. (1992). Vaccinia–rotavirus VP7 recombinants protectand co-workers (1986) and, to our knowledge, only two mice against rotavirus-induced diarrhoea. Vaccine 10, 185–191.
Andrew, M. E., Boyle, D. B., Coupar, B. E., Whitfeld, P. L., Both, G. W.,IgG antibodies to this site have been described. Theseand Bellamy, A. R. (1987). Vaccinia virus recombinants expressingMAbs were assigned to the NP1a epitope of the simianthe SA11 rotavirus VP7 glycoprotein gene induce serotype-specificSA-11 strain of rotavirus, and one of them was shown toneutralizing antibodies. J. Virol. 61, 1054–1060.
select for change at amino acid 136 (Burns et al., 1988; Andrew, M. E., Boyle, D. B., Whitfeld, P. L., Lockett, L. J., Anthony, I. D.,Zhou et al., 1994). The SA-11 MAbs were found to com- Bellamy, A. R., and Both, G. W. (1990). The immunogenicity of VP7,
a rotavirus antigen resident in the endoplasmic reticulum, is en-pete in epitope-binding ELISA with antibodies directedhanced by cell surface expression. J. Virol. 64, 4776–4783.at different epitopes in both subpeptides of VP4, whereas
Arias, C. F., Ballado, T., and Plebanski, M. (1986). Synthesis of the outer-only one of our five IgA MAbs showed a low degree ofcapsid glycoprotein of the simian rotavirus SA11 in Escherichia coli.
competition with RRV MAb M14 (selecting for amino acid Gene 47, 211–219.148 change; Mackow et al., 1988b). However, competi- Bastardo, J. W., McKimm, B. J., Sonza, S., Mercer, L. D., and Holmes,
I. H. (1981). Preparation and characterization of antisera to electro-tion seen with M14 was not reciprocal, and since M14phoretically purified SA11 virus polypeptides. Infect. Immun. 34, 641–is an IgM antibody, the effect observed may simply be647.due to steric hindrance.
Burns, J. W., Greenberg, H. B., Shaw, R. D., and Estes, M. K. (1988).In agreement with the epitope competition data, the Functional and topographical analyses of epitopes on the hemagglu-
five virus neutralization-escape mutants with selected tinin (VP4) of the simian rotavirus SA11. J. Virol. 62, 2164–2172.Burns, J. W., Siadat-Pajouh, M., Krishnaney, A. A., and Greenberg, H. B.amino acid changes at residues 132–135 were normally
(1996). Protective effect of rotavirus VP6-specific IgA monoclonalneutralized by the whole panel of reference MAbs to VP4antibodies that lack neutralizing activity. Science 272, 104–107.and VP7 used in this study. The aa 132–135 mutants
Chen, K.-S., and Strober, W. (1990). Cholera holotoxin and its B subunitwere also fully neutralized by all other IgA MAbs belong- enhance Peyer’s patches B cell responses induced by orally adminis-ing to the groups described above. Altogether, these data tered influenza virus: Disproportionate cholera toxin enhancement
of the IgA B cell response. Eur. J. Immunol. 20, 433–436.indicate that the aa 132–135 epitope is completely sepa-Coffin, S. E., Klinek, M., and Offit, P. A. (1995). Induction of virus-specificrated from other known antigenic domains of RRV VP4.
antibody production by lamina propria lymphocytes following intra-The pattern of resistance to MAb neutralization was sub-muscular inoculation with rotavirus. J. Infect. Dis. 172, 874–878.
stantially similar for four of the five mutants which Conner, M. E., Gilger, M. A., Estes, M. K., and Graham, D. Y. (1991).showed loss of reactivity with only the MAbs belonging Serologic and mucosal immune response to rotavirus infection in
the rabbit model. J. Virol. 65, 2562–2571.to the group. Unexpectedly, the fifth virus variant v3F12Coulson, B. S., Grimwood, K., Hudson, I. L., Barnes, G. L., and Bishop,escaped neutralization by only the homologous IgA MAb.
R. F. (1992). Role of coproantibody in clinical protection of childrenIt is possible that the alanine to valine change at positionduring reinfection with rotavirus. J. Clin. Microbiol. 30, 1678–1684.
135 selected in variant v3F12 does not significantly alter Dharakul, T., Riepenhoff, T. M., Albini, B., and Ogra, P. L. (1988). Distri-the reactivity of other MAbs since both residues share bution of rotavirus antigen in intestinal lymphoid tissues: Potential
role in development of the mucosal immune response to rotavirus.similar aliphatic side chains, but this change is sufficientClin. Exp. Immunol. 74, 14–19.to prevent binding of the selecting MAb. In all other mu-
Dormitzer, P. R., Both, G. W., and Greenberg, H. B. (1994). Presentationtants of this group, the mutations detected involved aof neutralizing epitopes by engineered rotavirus VP7s expressed by
charge modification of the amino acid selected. Further recombinant vaccinia viruses. Virology 204, 391–402.studies in animal models will indicate the importance of Dyall, S. M., Lazdins, I., Tregear, G. W., and Holmes, I. H. (1986). Loca-
tion of the major antigenic sites involved in rotavirus serotype-spe-this epitope in protective immunity.cific neutralization. Proc. Natl. Acad. Sci. USA 83, 3465–3468.The high frequency of isolation of IgA MAbs which
Estes, M. K., Graham, D. Y., and Mason, B. B. (1981). Proteolytic en-select for amino acid changes at residues 132–135 ofhancement of rotavirus infectivity: Molecular mechanisms. J. Virol.
VP4 suggest that orally induced neutralizing IgA re- 39, 879–888.sponses also involve an immunodominant antigenic site Fiore, L., Greenberg, H. B., and Mackow, E. R. (1991). The VP8 fragment
of VP4 is the rhesus rotavirus hemagglutinin. Virology 181, 553–563.on the virion VP8 protein that may not be a normal com-
AID VY 8224 / 6a21$$$343 10-14-96 13:36:12 vira AP: Virology

109IgA MONOCLONAL ANTIBODIES TO ROTAVIRUS SURFACE ANTIGENS
Flores, J., Myslinski, J., Kalica, A. R., Greenberg, H. B., Wyatt, R. G., Kapi- Mackow, E. R., Shaw, R. D., Matsui, S. M., Vo, P. T., Dang, M. N., andGreenberg, H. B. (1988b). The rhesus rotavirus gene encoding pro-kian, A. Z., and Chanock, R. M. (1982). In vitro transcription of two
human rotaviruses. J. Virol. 43, 1032–1037. tein VP3: Location of amino acids involved in homologous and heter-ologous rotavirus neutralization and identification of a putative fusionGerna, G., Battaglia, M., Milanesi, G., Passarani, N., Percivalle, E., and
Cattaneo, E. (1984). Serotyping of cell culture-adapted subgroup 2 region. Proc. Natl. Acad. Sci. USA 85, 645–649.Matsui, S. M., Offit, P. A., Vo, P. T., Mackow, E. R., Benfield, D. A., Shaw,human rotavirus strains by neutralization. Infect. Immun. 43, 722–
729. R. D., Padilla, N. L., and Greenberg, H. B. (1989). Passive protectionagainst rotavirus-induced diarrhea by monoclonal antibodies to theGorziglia, M., Larralde, G., Kapikian, A. Z., and Chanock, R. M. (1990).
Antigenic relationships among human rotaviruses as determined by heterotypic neutralization domain of VP7 and the VP8 fragment ofVP4. J. Clin. Microbiol. 27, 780–782.outer capsid protein VP4. Proc. Natl. Acad. Sci. USA 87, 7155–7159.
Greenberg, H. B., Flores, J., Kalica, A. R., Wyatt, R. G., and Jones, R. Mazanek, M. B., Kaetzel, C. S., Lamm, M. E., Fletcher, D., and Nedrud,J. G. (1992). Intracellular neutralization of virus by immunoglobulin A(1983a). Gene coding assignments for growth restriction, neutraliza-
tion and subgroup specificities of the W and DS-1 strains of human antibodies. Proc. Natl. Acad. Sci. USA 89, 6901–6905.McCrae, M. A., and McCorquodale, J. G. (1987). Expression of a majorrotavirus. J. Gen. Virol. 64, 313–320.
Greenberg, H. B., Valdesuso, J., Van Wyke, K., Midthun, K., Walsh, M., bovine rotavirus neutralization antigen (VP7c) in Escherichia coli.Gene 55, 9–18.McAulliffe, V., Wyatt, R. G., Kalica, A. R., Flores, J., and Hoshino, Y.
(1983b). Production and preliminary characterization of monoclonal McGonigal, T. P., Turon, M. C., Komar, H. K., Kister, S. E., and Smith,R. E. (1992). Expression of the gene coding for the major outer capsidantibodies directed at two surface proteins of rhesus rotavirus. J.
Virol. 47, 267–275. protein of SA-11 rotavirus in a baculovirus system. Virus Res. 23,135–150.Hoshino, Y., Sereno, M. M., Midthun, K., Flore, J., Kapikian, A. Z., and
Chanock, R. M. (1985). Independent segregation of two antigenic Mendez, E., Arias, C. F., and Lopez, S. (1993). Binding to sialic acidsis not an essential step for the entry of animal rotaviruses into epithe-specificities (VP3 and VP7) involved in neutralization of rotavirus
infectivity. Proc. Natl. Acad. Sci. USA 82, 8701–8704. lial cells in culture. J. Virol. 67, 5253–5259.Merchant, A. A., Groene, W. S., Cheng, E. H., and Shaw, R. D. (1991).Kaetzel, C. S., Robinson, J. K., Chintalacharuvu, K. R., Vaerman, J., and
Lamm, M. E. (1991). The polymeric immunoglobulin receptor (secre- Murine intestinal antibody response to heterologous rotavirus infec-tion. J. Clin. Microbiol. 29, 1693–1701.tory component) mediates transport of immune complexes across
epithelial cells: A local defense function for IgA. Proc. Natl. Acad. Mills, J. V., Van Kirk, J. E., Wright, P. F., and Chanock, R. M. (1971). Exper-imental respiratory syncitial virus infection of adults. Possible mecha-Sci. USA 88, 8796–8800.
Kalica, A. R., Flores, J., and Greenberg, H. B. (1983). Identification of nisms of resistance to infection and illness. J. Immunol. 107, 123–130.the rotaviral gene that codes for hemagglutination and protease-
enhanced plaque formation. Virology 125, 194–205. Nagesha, H. S., and Holmes, I. H. (1991). VP4 relationships betweenporcine and other rotavirus serotypes. Arch. Virol. 116, 107–118.Kalica, A. R., Greenberg, H. B., Wyatt, R. G., Flores, J., Sereno, M. M.,
Kapikian, A. Z., and Chanock, R. M. (1981). Genes of human (strain Offit, P. A., and Blavat, G. (1986a). Identification of the two rotavirusgenes determining neutralization specificities. J. Virol. 57, 376–378.Wa) and bovine (strain UK) rotaviruses that code for neutralization
and subgroup antigens. Virology 112, 385–390. Offit, P. A., Shaw, R. D., and Greenberg, H. B. (1986b). Passive protectionagainst rotavirus-induced diarrhea by monoclonal antibodies to sur-Kapikian, A. Z., and Chanock, R. M. (1990). Rotaviruses. In ‘‘Virology’’
(B. N. Fields, D. M. Knipe, R. M. Chanock, M. S. Hirsch, L. M. Joseph, face proteins vp3 and vp7. J. Virol. 58, 700–703.Prasad, B. V., Burns, J. W., Marietta, E., Estes, M. K., and Chiu, W. (1990).T. P. Monath, and B. Roizman, Eds.), 2nd ed., pp. 1353–1404. Raven
Press, New York. Localization of VP4 neutralization sites in rotavirus by three-dimen-sional cryo-electron microscopy. Nature (London) 343, 476–479.Kraehenbuhl, J.-P., and Neutra, M. R. (1992). Molecular and cellular
basis of immune protection of mucosal surfaces. Physiol. Rev. 72, Prasad, B. V., Wang, G. J., Clerx, J. P., and Chiu, W. (1988). Three-dimen-sional structure of rotavirus. J. Mol. Biol. 199, 269–275.853–880.
Laemmli, U. K. (1970). Cleavage of structural proteins during the assem- Renegar, K. B., and Small, P. A., Jr. (1991). Immunoglobulin A mediationof murine nasal anti-influenza virus immunity. J. Virol. 65, 2146–2148.bly of the head of bacteriophage T4. Nature (London) 227, 680–685.
Lazdins, I., Sonza, S., Dyall-Smith, M., Coulson, B., and Holmes, I. (1985). Riepenhoff, T. M., Lee, P. C., Carmody, P. J., Barrett, H. J., and Ogra,P. L. (1982). Age-dependent rotavirus-enterocyte interactions. Proc.Demonstration of an immunodominant neutralization site by analysis
of antigenic variants of SA11 rotavirus. J. Virol. 56, 317–319. Soc. Exp. Biol. Med. 170, 146–154.Ruggeri, F. M., and Greenberg, H. B. (1991). Antibodies to the trypsinLiew, F. Y., Russell, S. M., Appleyard, G., Brand, C. M., and Beale, J.
(1984). Cross-protection in mice infected with influenza A virus by cleavage peptide VP8 neutralize rotavirus by inhibiting binding ofvirions to target cells in culture. J. Virol. 65, 2211–2219.the respiratory route is correlated with local IgA antibody rather than
serum antibody or cytotoxic T cell reactivity. Eur. J. Immunol. 14, Sabara, M., Barrington, A., and Babiuk, L. A. (1985). Immunogenicity ofa bovine rotavirus glycoprotein fragment. J. Virol. 56, 1037–1040.350–356.
Losonsky, G. A., Rennels, M. B., Lim, Y., Krall, G., Kapikian, A. Z., and Saif, L. J., Redman, D., Smith, K., and Theil, K. (1983). Passive immunityto bovine rotavirus in newborn calves fed colostrum supplementsLevine, M. M. (1988). Systemic and mucosal immune responses to
rhesus rotavirus vaccine MMU 18006. Pediatr. Infect. Dis. J. 7, 388– from immunized or non-immunized cows. Infect. Immun. 41, 1118–1131.393.
Lycke, N., and Holmgren, J. (1986). Strong adjuvant properties of cholera Saif, L. J., Rosen, B. I., and Parwani, A. V. (1994). Animal rotavirus. In‘‘Viral Infections of the Gastrointestinal Tract’’ (A. Z. Kapikian, Ed.),toxin on gut mucosal immune responses to orally presented anti-
gens. Immunology 59, 301– 308. pp. 279–368. Dekker, New York.Sanger, F., Nicklen, S., and Coulson, A. R. (1977). DNA sequencing withMackow, E. R., Barnett, J. W., Chan, H., and Greenberg, H. B. (1989).
The rhesus rotavirus outer capsid protein VP4 functions as a hemag- chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74, 5463–5467.glutinin and is antigenically conserved when expressed by a baculo-
virus recombinant. J. Virol. 63, 1661–1668. Shaw, R. D., Fong, K. J., Losonsky, G. A., Levine, M. M., Maldonado, Y.,Yolken, R., Flores, J., Kapikian, A. Z., Vo, P. T., and Greenberg, H. B.Mackow, E. R., Shaw, R. D., Matsui, S. M., Vo, P. T., Benfield, D. A., and
Greenberg, H. B. (1988a). Characterization of homotypic and hetero- (1987). Epitope-specific immune responses to rotavirus vaccination.Gastroenterology 93, 941–950.typic VP7 neutralization sites of rhesus rotavirus. Virology 165, 511–
517. Shaw, R. D., Mackow, E. R., Dyall, S. M., Lazdins, I., Holmes, I. H., and
AID VY 8224 / 6a21$$$344 10-14-96 13:36:12 vira AP: Virology

110 GIAMMARIOLI ET AL.
Greenberg, H. B. (1988). Serotypic analysis of VP3 and VP7 neutral- Taniguchi, K., Morita, Y., Urasawa, T., and Urasawa, S. (1987). Cross-ization escape mutants of rhesus rotavirus. J. Virol. 62, 3509–3512. reactive neutralization epitopes on VP3 of human rotavirus: Analysis
Shaw, R. D., Stoner-Ma, D., Estes, M., and Greenberg, H. (1985). Spe- with monoclonal antibodies and antigenic variants. J. Virol. 61, 1726–cific enzyme-linked immunoassay for rotavirus serotypes 1 and 3. J. 1730.Clin. Microbiol. 22, 286–291. Taniguchi, K., Nishikawa, K., Urasawa, T., Urasawa, S., Midthun, K.,
Shaw, R. D., Vo, P. T., Offit, P. A., Coulson, B. S., and Greenberg, H. B. Kapikian, A. Z., and Gorziglia, M. (1989). Complete nucleotide se-(1986). Antigenic mapping of the surface proteins of rhesus rotavirus. quence of the gene encoding VP4 of a human rotavirus (strain K8)Virology 155, 434–451. which has unique VP4 neutralization epitopes. J. Virol. 63, 4101–
Snodgrass, D., and Well, P. (1976). Rotavirus infection in lambs: Studies 4106.on passive protection. Arch. Virol. 52, 201–205. Taniguchi, K., Urasawa, S., and Urasawa, T. (1985). Preparation and
Solari, R., and Kraehenbhul, J.-P. (1985). The biosynthesis of secretory characterization of neutralizing monoclonal antibodies with differentcomponent and its role in the transepithelial transport of IgA dimer. reactivity patterns to human rotaviruses. J. Gen. Virol. 66, 1045–1053.Immunol. Today 6, 17–20. Ward, R. L., Knowlton, D. R., Schiff, G. M., Hoshino, Y., and Greenberg,
Svensson, L., Sheshberadaran, H., Vene, S., Norrby, E., Grandien, M., H. B. (1988). Relative concentrations of serum neutralizing antibodyand Wadell, G. (1987). Serum antibody responses to individual viral
to VP3 and VP7 proteins in adults infected with a human rotavirus.polypeptides in human rotavirus infections. J. Gen. Virol. 68, 643–
J. Virol. 62, 1543–1549.651.
Ward, R. L., McNeal, M. M., Sander, D. S., Greenberg, H. B., and Bern-Taniguchi, K., Hoshino, Y., Nishikawa, K., Green, K. Y., Maloy, W. L.,stein, D. I. (1993). Immunodominance of the VP4 neutralization pro-Morita, Y., Urasawa, S., Kapikian, A. Z., Chanock, R. M., and Gorziglia,tein of rotavirus in protective natural infections of young children. J.M. (1988a). Cross-reactive and serotype-specific neutralization epi-Virol. 67, 464–468.topes on VP7 of human rotavirus: Nucleotide sequence analysis of
Zagursky, R., Baumeister, K., Lomax, N., and Berman, M. (1985). Dideoxyantigenic mutants selected with monoclonal antibodies. J. Virol. 62,DNA sequencing directly from plasmid DNA using reverse tran-1870–1874.scriptase. Gene Anal. Tech. 2, 5–11.Taniguchi, K., Maloy, W. L., Nishikawa, K., Green, K. Y., Hoshino, Y.,
Zhou, Y. J., Burns, J. W., Morita, Y., Tanaka, T., and Estes, M. K. (1994).Urasawa, S., Kapikian, A. Z., Chanock, R. M., and Gorziglia, M.Localization of rotavirus VP4 neutralization epitopes involved in anti-(1988b). Identification of cross-reactive and serotype 2-specific neu-body-induced conformational changes of virus structure. J. Virol. 68,tralization epitopes on VP3 of human rotavirus. J. Virol. 62, 2421–
2426. 3955–3964.
AID VY 8224 / 6a21$$$344 10-14-96 13:36:12 vira AP: Virology




















