
Allelic antigen and membrane-anchor epitopes of Paramecium primaurelia surface antigens
Upload
independentCategory
view
2download
0
[CANCER RESEARCH 56. 3808-3813, August 15, 1996]
Chromosome 7q Allelic Losses in Pancreatic Carcinoma1
Alberto Achille, Maria O. Biasi, Giuseppe Zamboni, Giuseppe Bogina, Alba R. Magalini, Paolo Pederzoli,Manuel Perucho, and Aldo Scarpa2
Istillilo di Anonimia Patologica ¡A.A., M. O. B.. C. Z.. G. B., A. R. M., A. S.¡and Dipanimeli!« di Scienze Chirurgiche ¡P.PJ. Università di Verona. Strada Le Grazie, 1-37134
Verona. Italy, and The Burham Institute ¡M.PJ. La Jolla, California 92037
ABSTRACT
During our DNA fingerprinting studies of paired normal and pancreatic cancer tissues using arbitrarily primed PCR, we noticed a bandshowing an apparent homozygous deletion in a pancreatic cancer cell lineand a decreased intensity in a number of primary cancers. That band wasassigned to chromosome 7. Such information led us to analyze chromosome 7 loss of heterozygosity (LOH) in a panel of 12 cryostat-enriched
primary pancreatic cancers and 2 pancreatic cancer cell lines, despite thereportedly low frequency of chromosome 7 LOH in xenograft-enrichedpancreatic cancers. Seventeen PCR-amplified CA-microsatellite polymorphic sites were analyzed. One of the two cell lines and eight common-type
cancers (including all five poorly differentiated and three of five moderately differentiated cancers) showed chromosome 7q LOH, whereas thetwo uncommon types of ductal cancer (one adenosquamous and onemucinous noncystic) scored negative. Our data suggest that chromosome7q LOH is a frequent event (80%) in cryostat-enrichable common pan
creatic ductal carcinomas, that is, those primarily of high cellularity. Thechromosome 7q smallest common deleted region described by our caseswas between 7q31.1 and 7q32.
INTRODUCTION
Alterations of recessive TSGs3 are critical events in the multistep
process leading to the development of cancers. Cytogenetic techniques have been used to determine the location of TSGs becauseconsistent deletions or inversions of part of a chromosome are indicative of the inactivation of a nearby TSG during neoplastic progression (1). However, cytogenetic techniques do not detect the entirespectrum of inactivating events during carcinogenesis, such as mi-
crodeletion, chromosomal loss with duplication, and homologousrecombination with defective chromatid (2). Therefore, more sensitivemolecular methods have been used to screen for genetic alterations intumors and to determine the SCDR involved in those alterations.These methods are based on the detection of LOH at chromosome-
specific polymorphic sites in DNA extracted from tumors, whencompared to DNA from matched normal tissues. Such allelotyping isfeasible by RFLP by Southern blot using VNTR DNA probes and/orPCR amplification of microsatellite repeats, provided there is heterozygosity for the studied loci and a neoplastic cellularity higher than50-60% in the cancer sample (3). Recently developed DNA fingerprinting techniques such as AP-PCR (4-6) are also being applied to
screen for chromosomal abnormalities in cancer.Pancreatic ductal cancer is a difficult neoplasm to study with such
techniques due to the characteristic host desmoplastic reaction and thehigh admixture of contaminating nonneoplastic inflammatory and
Received 4/12/96; accepted 6/18/96.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1Supported by grants from the Associazione Italiana Ricerca Cancro. Milan. Italy:
Banca Popolare di Verona. Italy; and Ministero Università e Ricerca Scientifica eTecnologica 60% and 40%, Rome. Italy; and NIH Grant CA38579 from ihe NationalCancer Institute.
2 To whom requests for reprints should be addressed. Phone: 39-45-8098-617; Fax:
39-45-8098-136.1The abbreviations used are: TSG, tumor suppressor gene: LOH. loss of helerozy-
gosity; AP-PCR. arbitrarily primed PCR; SCDR. smallest common deleted region; VNTR,variable number of tandem repeats.
stromal cells. To overcome such problems, neoplastic cell-enrichmenttechniques have been devised, such as cryostat-dissection (7) of
primary cancers and xenografting in nude mice (8, 9).During our studies of AP-PCR DNA fingerprinting of pancreatic
cancers, we noticed a band showing a decreased intensity in a numberof cancer samples with respect to matched normal tissues and anapparent homozygous deletion in a cell line. That band was assignedto chromosome 7. This led us to analyze chromosome 7 LOH in apanel of cryostat-enriched pancreatic cancer samples, despite thereportedly low frequency of chromosome 7 LOH in xenograft-en
riched pancreatic cancers (9). In this regard, it is worth noting thatcytogenetic studies of pancreatic cancer (10-12) showed that chro
mosome 7 is among those chromosomes most frequently affected bystructural abnormalities (32 of 66 cases, 48%), including partial lossesand breakpoints at which translocations or addition of material wereobserved. The LOH of chromosome 7q has been reported as a frequent abnormality in many different epithelial neoplasms (13, 14),including gastrointestinal malignancies (15-18), and the presence of
TSGs on 7q is suggested by several lines of evidence, including invitro studies (19, 20).
We explored chromosome 7 LOH in 12 pancreatic carcinomas witha cryostat-enriched neoplastic cellularity of at least 60% and in two
pancreatic cancer cell lines, using PCR amplification of matchednormal and cancer DNAs with a set of 17 CA-microsatellite repeat-
specific primers. Our results indicate that chromosome 7q LOH isfrequent (80%) in those common-type pancreatic carcinomas charac
terized primarily by a high neoplastic cellularity.
PATIENTS AND METHODS
Patients and Tumors. Twelve pancreatic adenocarcinomas with a neoplastic cellularity of at least 60% were obtained by cryostat enrichment of 87
frozen samples of cancers of ascertained pancreatic origin, collected between1984 and 1995 at the Pathology Department of the University of Verona.Ampullary. periampullary, and biliary cancers were excluded. Two pancreaticcancer cell lines, SK-PC-1 and IMIM-PC-2. whose cytogenetic and molecular
features have been reported (21, 22), and the matched normal tissue DNA fromthe same patients were also studied. The clinicopathological data of the 12patients are summarized in Table 1. All common-type ductal cancers were
graded using a combined histological and cytological grading system according to Kloeppel (23). They included five moderately differentiated and fivepoorly differentiated neoplasms. The remaining two cancers were a mucinousnoncystic cancer (PC 10) and an adenosquamous carcinoma (PCI 1).
Adequacy of Tissue Samples and DNA Extraction. Samples were selected based on the following criteria: (a) availability of frozen samples of bothtumor and normal tissue from the same patient. In fact, only the use of highquality genomic DNA obtainable from fresh or frozen tissues allows extensiveanalysis on different DNA sequences with a reasonable certainty of avoidingfalse results; and (b) a neoplastic cellularity of more than 60% in tumorspecimens, which is crucial for the loss of DNA sequences to be detectablewhen compared to normal tissue. For this purpose, neoplastic cellularity wasassessed on stained slides prepared during the cryostat dissection and was aconservative estimate of the number of neoplastic cells as a percentage of totalcells in the final sample. In the majority of samples, in which the neoplasticcells accounted for less than 60%. the neoplastic population was enriched byeither eliminating the portions of normal tissue from the frozen blocks orscraping neoplastic areas from slides. An adequate neoplastic cellularity of at
3808
Research. on February 9, 2016. © 1996 American Association for Cancercancerres.aacrjournals.org Downloaded from
7q LOH IN PANCREATIC CANCER
Table I Clinicopathe>lof>ical data of priman,- pancreatic carcinomas and ceti lines
DiameterMutations LOH at chromosome
CaseSK-PC-IIMIMPCIPC:PC3PC4PC5PC6PC7PC'SPC9PC10PC1IPC12Age(yr)/sex63/M52/M75/F60/F62/F51/M64/F68/F67/M48/M50/M78/M74/M36/FDitTerentiationPoorPoorModeratePoorModerateModerateModeratePoorModeratePoorPoor,
solidMucinousnoncysticAdenosquumousPoorCellularityCell
lineCellline70%70%60%60%70%80%60%70%70%60%60%70%(cm)3.53.52.50.674.5441.54126metastasisNoYesYesYesYesNoYesYesNoNoNoYesmoDOD/13DOD/8DOD/12DOD/16DOD/8DOD/10Lost
1mDOD/4AW/35AW/3DOD/5DOD/9Ki-ra/AspAspValValAspAspWild
typeWildtypeAspAspWild
typeValSerValpSfYesYesNot
doneYesYesNoYesYesNoNoNoYesNoYes17pNo
lossNolossLossLossLossNo
lossLossNILossLossNo
lossLossLossLossI8qNo
lossLossLossLossNILossNo
lossLossLossNINo
lossLossLossNo
loss9pNot
doneNotdoneNotdoneNotdoneNILossNo
lossNolossLossLossNo
lossNolossLossNot
done7qLossNo
lossLossLossLossNo
lossLossLossNo
lossLossLossNo
lossNolossLoss
" DOD. died of disease; AW, alive and well.'' Ki-raj mutations were determined by single-strand conformation polymorphism and direct DNA-sequencing of PCR-amplified exon 1. All
aspartic acid: Val. valin; Ser, serin.p53 mutations were determined by single-strand conformation polymorphism and direct DNA-sequencing of PCR-amplifiedexons 5-9. and the
analyzed by cloning and sequencing as described in Redston et al. (31).' NI. not informative.
mutations were at codon 12. Asp,
negative-scoring cases were further
least 60% was obtained in the 12 cases included in the present report out of the87 pancreatic cancer frozen samples dissected. DNA was prepared by phenol-chloroform extraction and ethanol precipitation from 30-100 six-jnm cryo-
static sections, checking the cell composition of the sample every 20 sections.To both confirm the success of cryostat enrichment and establish the
reliability of microsatellite markers for determining chromosomal loss in oursamples, we performed chromosome 9p. I7p, and I8q LOH analyses. In fact,alleile losses at these chromosomal arms are the most frequent chromosomalanomalies described in pancreatic cancer (7-9, 12). In addition, based on thecytogenetic information available for cell lines IMIM-PC-2 and SK-PC-1 (21),
we could use them as positive and negative controls, respectively, for chromosome 18 LOH. Such tests were particularly useful in the cases with a cancercellularity of around 60%, and from the 18 cases initially chosen after cryostatenrichment, 6 cases with borderline cellularity were eliminated because theydid not show, or showed questionable. LOH at chromosomes 9p, 17p, or 18q.
AP-PCR. The arbitrary primer used was AR3 (5'-GCGAATTCATG-TACGTCAGG-3'). DNA (70 ng) was incubated with 0.6 units of Taq polym-
erase (Perkin Elmer Cetus). 125 /xM of each deoxynucleotide triphosphate.0.13 PL]|a-"P]dCTP (Amersham. 3000 Ci/mmol), 10 mM Tris-HCI (pH 8.2),
50 mM KCI. 5 mM MgCU, 0.1% gelatin, and arbitrary primer (0.1 mM) in afinal volume of 15 fii. The reactions were carried out in a PT-100 thermalcycler (M. J. Research, Inc.) for 5 cycles at low stringency (94°Cfor 30 s, 50°Cfor 1 min, and 72°Cfor 1.5 min) and 25 cycles at high stringency (94°Cfor15 s, 60°Cfor 15 s, and 72°Cfor 1 min). The AP-PCR product (5 /xl) was
diluted in 13 id of dilution loading buffer (DLB) (0.01% each of bromphenolblue and xylene cyanol, 0.01 N NaOH. 0. l MEDTA. and 93% formamide), and3 /xl were electrophoresed in a 5% polyacrylamide gel containing 8 M urea setup by wedge spacers (0.4- 1.2 cm). After electrophoresis, the gel was trans
ferred to filter paper, was dryed under vacuum, and was exposed for multipletimes (ranging from 12-24 h) to X-ray films (Kodak X-OMAT AR). Each
experiment was performed in triplicate.Chromosomal Assignment of Band L of AR-3 AP-PCR Fingerprint. To
determine the chromosomal localization of band L (see "Results"), we used the
same AR-3 primer to amplify, with the same AP-PCR protocol described
above, the DNA (70 ng) from each of the 24 monochromosomal hybridsincluded in human/rodent somatic cell hybrid panel 2 (24). Hamster and mouseDNAs were used as controls. The chromosomal assignment was accomplishedby comparing the human fingerprints with those obtained from each mono-
chromosomal hybrid, as described (25). To confirm the assignment, band Lwas eluted from the gel and was cloned and sequenced, as previously described(4). Then, on the basis of the sequence obtained, two primers were synthesized(CAR31-up, CACATATGGCTACAGATTTC; CAR31-down, CTTCTAGG-GAAGATATAGCC) and were used to PCR-amplify human and human/rodent
monochromosomal hybrid DNAs.LOH Analysis. Chromosome 18q LOH was assessed using four microsat
ellite markers, which were D18S69, DI8S64, D18S61. and D18S58 (26). Allcases were tested for D18S69 and D18S64. Noninformative and negative-
scoring cases were sequentially tested for D18S61 and D18S58. Two micro-
satellite markers were used for each of chromosomes 9p and 17p, namely,IFN-a and D9S171 (8), and D17S513 and D17S559, respectively. The 17
microsatellite repeats used to analyze chromosome 7 LOH are reported inTable 2. All primers were obtained from the MapPairs collection (ResearchGenetics, Huntsville, AL).
The reaction mixture (10 /xl) contained 20 ng of genomic DNA. the properpair of primers (0.5 /xM of each). 125 /MMof each deoxynucleotide triphosphate, 1 x PCR buffer [10 mM Tris-HCI (pH 8.2). 1.5 mM MgCU, 50 mM KCI,and 0.1% gelatin], 0.1 /xl (0.5 units) Taq polymerase, and 0.1 /xl [a-32P]dCTP(3000 Ci/mmol). After heating at 94°Cfor 4 min, PCR was performed for 30cycles of 30 s at 94°C.30 s at 55°C,and 30 s at 72°C.The reaction products
(5 /xl) were diluted 1:10 with DLB and were denatured, and 3 /xl wereelectrophoresed for 2-3 h according to the length of amplified fragments at 60W in 6% polyacrylamide (5% crosslinker, 0.4-mm thick) gel containing 8 Murea. Before loading, the gel was pre-run for 30 min at 60 W. After electro
phoresis, the gel was fixed on paper, was dried under vacuum, and wasexposed to X-ray film for multiple times (ranging from 12-24 h) at room
temperature.Determination of AlÃeleLoss. Only patients heterozygous for a given
DNA sequence were considered as informative, whereas the presence of eitherhomozygosity or an unclear distinction between paternal and maternal alÃeleswas considered as uninformative. Allelic imbalances consistent with LOHwere scored when there was a loss of intensity of one alÃelein the tumorsample with respect to the matched alÃelefrom normal tissue, and the relativeintensity of the two alÃeles in the tumor DNA differed from the relativeintensity in the nonneoplastic tissue DNA by a factor of at least 1.5 (26, 27).
Table 2 Cumulative results of chromosome 7 LOH in 12 priman' pancreatic cancersand cell lines IMIM-PC-2 ami SK-PC-I
MarkerD7S641D7S547D7S435D7S555EGFRD7S80IELNCOLIA2D7S796D7S471D7S522D7S649D7S495D7SI805TCRBD7S636D
75550Location7p22-p217p21-pl37pl5.17pl27pl2-pll.27qll.21-qll.237qll-q21.17q21.3-q227q227q3
1.l-q31.27q31.17q31.27q31.3-q327q347q357q35-q367q36Informative
cases(%)9
(64%)9(64%)7(50%)9
(64%)10(71%)7
(50%)6(42%)1
1(79%)11(79%)11(79%)8
(57%)11(79%)8
(57%)12(86%)11
(79%)8(57%)7
(50%)Cancers
withLOH00001104664g57642FrequencyofLOH0%0%0%0%10%14%0%36%55%55%50%73%62%58%54%50%29%
3809
Research. on February 9, 2016. © 1996 American Association for Cancercancerres.aacrjournals.org Downloaded from
7q LOH IN PANCREATIC CANCER
The evaluation of the intensity of the signal between the different alÃeleswasdetermined (using multiple exposure times) by visual examination by fourindependent reviewers and was quantitated by densitometry (GS-670 scanningdensitometer equipped with Molecular Analysis software; Bio-Rad). A comparison of these allelic imbalances with the results of the AP-PCR fingerprints
regarding chromosome 7 band L permitted us to decide whether these imbalances were due to allelic losses or gains. The results of allelic losses wereconsidered reliable only if reproducible in replicate experiments performed bytwo independent researchers (A. A. and M O. B.). The percentage of LOH wasexpressed as the number of cases where LOH was exhibited over the numberof hétérozygotesfor that particular sequence.
Statistical Analysis. The normality of the LOH percentage distributionswas tested using the Kolmogorov-Smirnov continuous cumulative distribution
test (28).
RESULTS
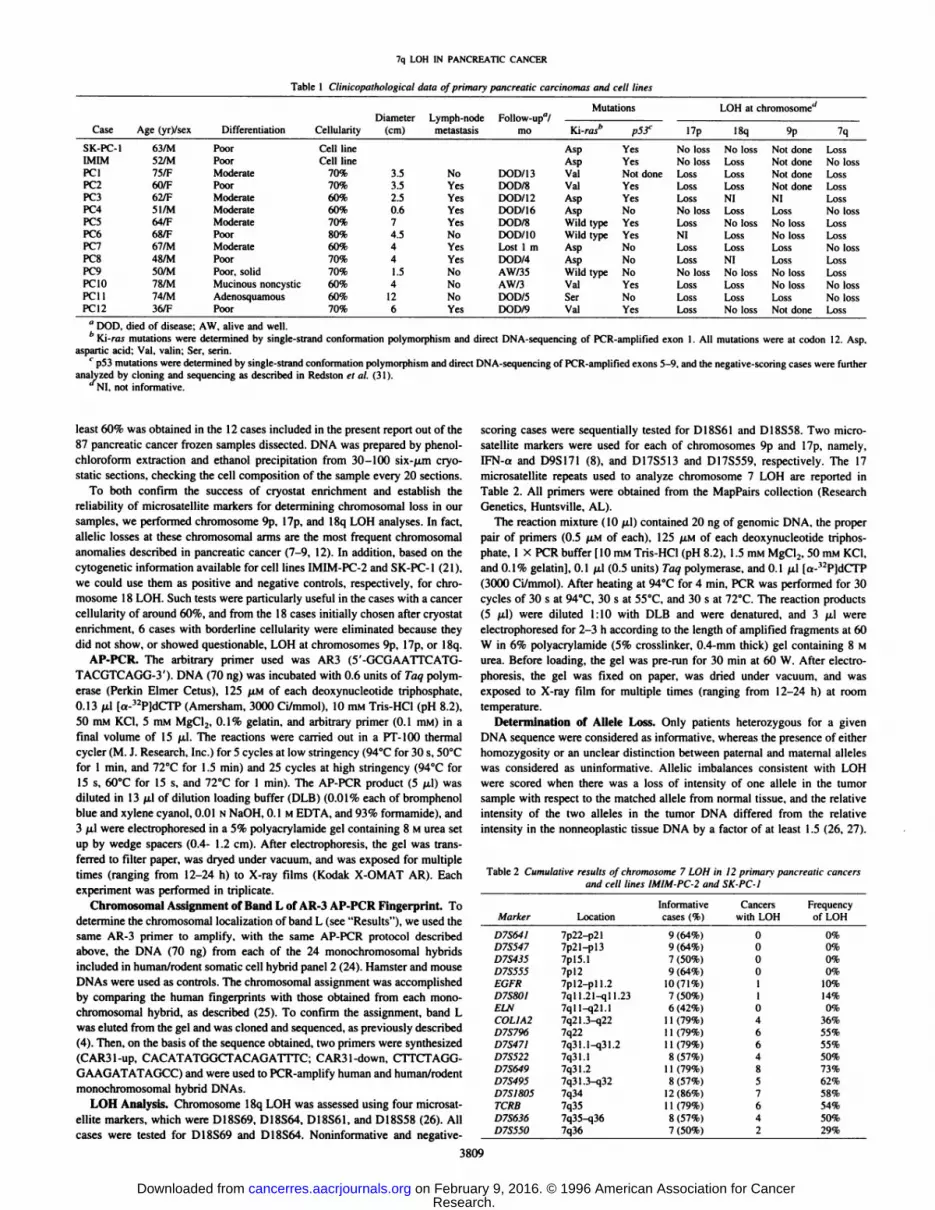
AP-PCR. Among the abnormalities noticed in AP-PCR finger
prints of pancreatic cancer cell lines, a nonpolymorphic band, named"L", showed an apparent homozygous deletion in the COLO357 cell
line (Fig. 1/4). When AP-PCR fingerprints from a panel of pancreatic
cancer cell lines and primary cancers (including 10 cases of thepresent study) were compared with matched DNAs from normaltissues, band L showed a decrease in intensity in about 50% of thecases, including cell line SK-PC-1 and cancers PCI, PC2, and PCS
(Fig. IS). No change in band L intensity was seen in cell lineIMIM-PC-2 and cancers PCS, PC9, and PC 10, whereas cancers PC4
- band L
B IMIM PCI PC2 SKPC1 PCS
NTNTNTNTNT
- bandL
Fig. 1. AP-PCR analysis with AR-3 arbitrary primer of pancreatic cancer cell lines (A)and of paired normal and cancer cells (fl|. A portion of the gel including band L is shown.jVand T indicate normal and cancer DNA. respectively. Band L is absent in the COLO357cell line and shows a reduced intensity (arrowheads) in cancers PCI. PC2, and PCS andin cell line SK-PC-1, when compared to corresponding normal DNA. Cell line IMIM-PC-2 shows no abnormality in band L.
Fig. 3. The assignment of AP-PCR band L Io chromosome 7 (see Fig. 2) was confirmed
by PCR amplification of DNA from the monochromosomal human/rodent somatic cellhybrid panel with primers designed from the sequence of the cloned L band. Numbers, thehuman chromosome contained in each hybrid. Hamster and mouse DNAs were used ascontrols. The expected PCR product of 163 bp is visible in the lanes of human andchromosome 7 hybrid DNAs.
and PC7 showed an increased intensity with respect to matchednormal tissue DNAs.
Chromosomal Localization of Band L by AR-3 AP-PCR Fingerprint. The PCR amplification with AR-3 primer of DNA from
monochromosomal human/rodent cell hybrid panel 2 allowed theassignment of band L to chromosome 7 (Fig. 2). This assignment wasconfirmed by PCR amplification of the same hybrid panel withprimers designed from the sequence of the cloned band L (availablefrom the authors; Fig. 3).
Adequacy of Samples: Chromosome 9p, 17p, and 18q LOH.Microsatellite allelotyping of chromosome 18 correctly recognizedthe loss of one chromosome in cell line IMIM-PC-2 and the retentionof both chromosomes in SK-PC-1. Eight of 11 informative cases
(72%) showed 18q LOH, whereas 9 of 11 (82%) and 4 of 8 (50%)informative cases showed 17p and 9p LOH, respectively (Table 1). Inparticular, the suitability of the five samples with the lowest cellularity(cases PC3, PC4, PC7, PC10, and PC11) was confirmed by thedetection of LOH in at least two of the tested loci. The results of thefrequency of LOH at these loci are consistent with those reported byother groups (7-9, 12, 29).
Chromosome 7 LOH. We then used 17 PCR-amplifted microsat
ellite repeats to screen the 12 pancreatic cancers and the two cell linesfor LOH on chromosome 7. The results are summarized in Table 2.Allelic losses were detected on 7q in 8 of the 12 cancers (67%) and incell line SK-PC-1. Only one case also showed 7p LOH. Representa
tive results of 7q LOH are shown in Figs. 4 and 5, where examples ofthe interpretation of results are also detailed. Insights concerning thelosses in positive-scoring cases are given in Figs. 6 and 7. The cases
showing 7q LOH included all five poorly differentiated cancers andthree of the five moderately differentiated carcinomas (8 of 10, 80%),whereas the two uncommon types of ductal pancreatic cancer, themucinous noncystic (PC 10) and the adenosquamous (PC 11), scorednegative. In particular, three cases showed LOH encompassing theentire q arm (PC 1, PC2, and PCS), together with the allelic loss of aportion of the p arm in one case (PCS). Case PC6 and cell lineSK-PC-1 showed a 7q partial LOH including the telomeres, whereas
the remaining four cases showed interstitial allelic losses. Overlapping
Fig. 2. Chromosomal localization of AP-PCR band L, usingPCR amplification with AR-3 primer of DNA from 24 mono-chromosomal human/rodent somatic cell hybrids. Numbers, thehuman chromosome contained in each hybrid. Hamster andmouse DNAs were used as controls. A band corresponding tohuman AP-PCR band L is visible only in the lane of the hybridcontaining chromosome 7 (arrowheads).
3810
Research. on February 9, 2016. © 1996 American Association for Cancercancerres.aacrjournals.org Downloaded from
7q LOH IN PANCREATIC CANCER
MarterO Oo. D,
uCX
M S~T Ñ~T Ñ~T
O7S 649
D7S 495
O7S 1805
TCRB
cell population. These results are at variance with those obtained inxenograft-enriched cancers, which showed a 17% frequency of 7q
LOH (9). Such a discrepancy might depend on the different enrichingprocedures. These include cryostat dissection of cancer samples, xe-nografting in nude mice, and the use of short-term cultures. Each of
these expedients introduces some bias in the selection of cases.Cryostat enrichment is successful in only about 20% of cases, whichrepresent those with a neoplastic cellularity higher than the averagecommon pancreatic cancer. Xenografting, which allows the neoplasticenrichment of at least 2-5 times, may also introduce some bias in the
M&rker
D7S801
tO <«•IA vu
AAAANT NT NT NT
D7S 636
07S 550
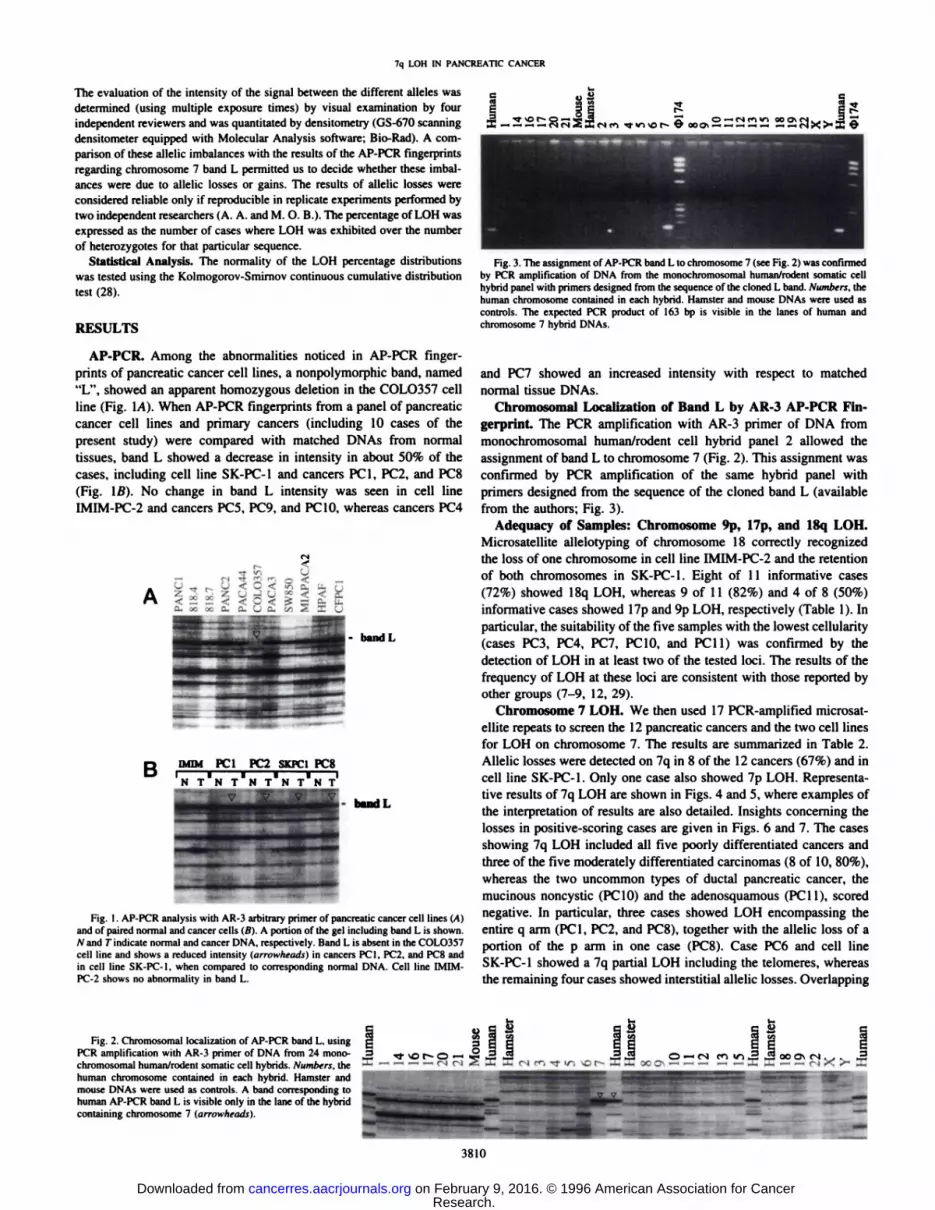
Fig. 4. Analysis of six PCR-amplified microsatellite loci spanning chromosome7q31.2-qler in two primary pancreatic cancers (PCI and PC2) and two pancreatic cancercell lines (SK-PC-I and IMIM-PC-2). Arrowheads, the presence of allelic losses in tumor(TÃcompared to matched normal tissue (AO DNA. In PCI. the losses of the upper D7S649.D7SÃŒ805.and TCRB alÃeles,and of the lower D7S495 alÃeleare visible. PC2 shows lossesin D7S649. D7S1805. and D7S636. In cell line SK-PC-I. the losses of the upper alÃeleinD7S649 and D7S550 loci, and of the lower alÃelein TCRB and D7S636 loci are evident,whereas the D7S1805 locus is not informative. Marker D7S495 in SK-PC-1 was considered as noninformative because the pattern obtained in cancer may be assigned to eitherthe loss of the lower alÃeleor the presence of an unrepaired mismatch error during DNAreplication (allelic shift). The latter phenomenon may be found occasionally, with afrequency of 0.3% per microsatellite locus (see Ref. 9). For the same reason. D7S495 wasconsidered uninformative in PC2. Cell line IMIM-PC-2 shows no losses.
deletions of the cases showing interstitial allelic losses indicate thatthe SCDR is enclosed between D7S522 and D7S1805 loci, spanningchromosomal region 7q31.1-7q32 (Fig. 7). No significant correlationwas found between allelic loss of chromosome 7 and clinicopatho-
logical data of the patients analyzed.No evidence of microsatellite instability (suggestive of the RER +
replication error phenotype) was found in the present cases. Rareallelic shifts extending or shortening the normal alÃelesize were foundin three cases, involving one or two single alÃeles/case (Fig. 4).
Statistical Analysis. The analysis of the histogram of the dataindicated that they had a normal distribution (Ref. 28; P < 0.001), aswould be expected by a stochastic process such as the inactivation ofaTSG.
DISCUSSION
The panel of pancreatic cancers in the present study is not representative of the more common low-cellularity pancreatic ductal cancers, but only of those 15-20% of cases containing a relatively highproportion of cancer cells and therefore cryostat-enrichable to a neo
plastic cellularity of at least 60%. We could obtain only 12 casessuitable for LOH studies out of 87 samples screened by cryostatsectioning. Based on this consideration, our results indicate thatchromosome 7q LOH is frequent (80%) in those common-type pan
creatic carcinomas primarily characterized by a relatively high cancer
ELN
COL1A2
D7S 796
D7S471
D7S 522
D7S 649
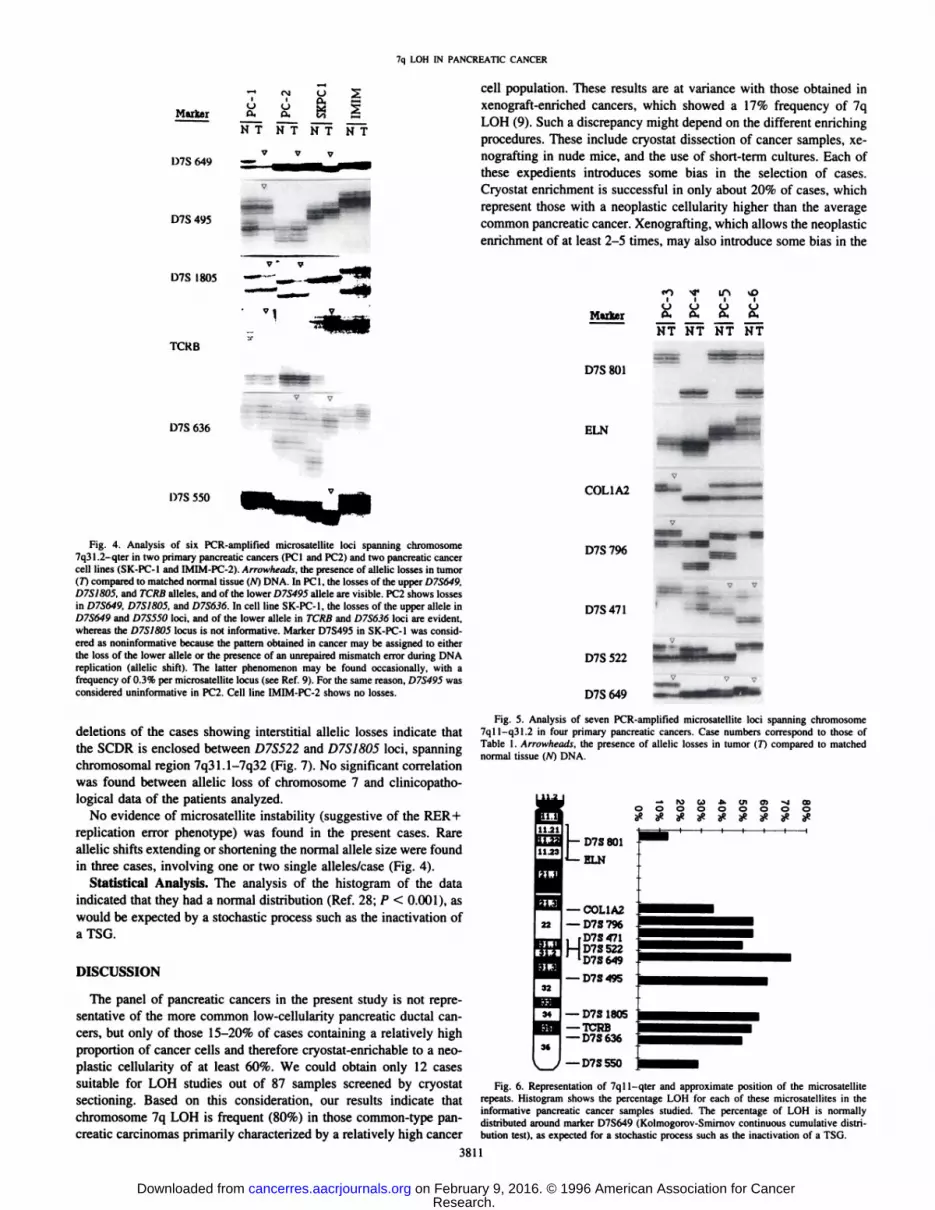
Fig. 5. Analysis of seven PCR-amplified microsatellite loci spanning chromosome7qll-q31.2 in four primary pancreatic cancers. Case numbers correspond to those ofTable 1. Arnmheads, the presence of allelic losses in tumor (7~)compared lo matched
normal tissue (/V) DNA.
ft» (B A M •¿�O O O O O
D7S801
ELN
—¿�COL1A2
22 | —¿�D7S 796
ID7S 471D7S 522D7S 649
I —¿�D7S 495
Hi
sii —¿�D7S 1805—¿�TCRB—¿�D7S 636
—¿�D7S550
Fig. 6. Representation of 7qll-qter and approximate position of the microsatelliterepeats. Histogram shows the percentage LOH for each of these microsatellites in theinformative pancreatic cancer samples studied. The percentage of LOH is normallydistributed around marker D7S649 (Kolmogorov-Smimov continuous cumulative distribution test), as expected for a stochastic process such as the inactivation of a TSG.
3811
Research. on February 9, 2016. © 1996 American Association for Cancercancerres.aacrjournals.org Downloaded from

7q LOH IN PANCREATIC CANCER
—¿�D7S Ml—¿�D7S 547
—¿�D7S 435
—¿�D7S555—¿�EOF
bD7S801
ELN
—¿�COL1A222 | —¿�D7S 796
rD7S471D7S 522
1D7S649
I—D7S495
Hi
~0-~00~_0"o"0-000000O00oooQ"0o000000gOC?^00OO—Q?0:?0000000D*•O0"0-"~-0c-~„70o-X0oo0-—0*«*Ä•*O*••fi0"piO'«0«**«4«fi000"*0m*»0.4*e000Q:-"***0a**.'-•-0--"„0**»•e*0o-rO0Ä*»**••k00-~-0„?Q**•hE00o-000-7QO«"g00C'o0?0C•**0C00sen«
—¿�D7S 1805—¿�TCRB—¿�D7S636
—¿�D7S550
Fig. 7. Deletions suggested by the alleile losses. Open columns, region is retained;shaded columns, loss or reduction to homozygosity of the region. The length of anycolumn is arbitrary because we possess no exact information on the precise limits of adeterminate deletion. •¿�.LOH: O. both alÃelesare retained: O. noninformative loci. Right.SCDR obtained by this analysis. Tap. case numbers.
panels obtained. For instance, all xenografts show p53 mutations (8),suggesting a selection for either that particular 60-70% of pancreatic
cancers containing p53 mutations (30, 31) or, sometimes, a selectiontor subclones within a cancer population (9, 32).
Cytogenetic studies of 66 pancreatic cancers with abnormal karyo-types (10-12) have shown that chromosome 7 is among the chromo
somes most frequently affected by structural abnormalities (32 of 66cases, 48%), including partial losses and breakpoints at which trans-
locations or addition of material were observed. Losses occurredmainly at 7q (9 of 66 cases, 14%) when compared to 7p (3 of 66 cases,4%), whereas breakpoints (20 of 66 cases, 29%) were equally distributed between the two chromosomal arms. The 7q losses were terminalin seven cases, whereas the remaining two were interstitial. All three7p and almost all 7q losses were represented by large deletionsincluding the telomeres. Numerical abnormalities of chromosome 7were frequently represented by gains ( 14 of 58 cases, 24%) and rarelyby losses of the entire chromosome (4 of 66 cases, 6%; Refs. 10-12).
Molecular techniques are more sensitive than cytogenetic analysisin detecting the loss of genetic material wherever it is located, eitherin its natural position or in the context of complex translocations.However, both RFLP using VNTR probes (7) and PCR-based micro-
satellite allelotyping (9) cannot formally determine whether thechange in the ratio between alÃelesis due to the losses of one or thegains of the other. This is particularly important for chromosome 7,which has been found to be triploid in 24% of pancreatic cancers (12).However, the interpretation that the allelic imbalances detected in ourcases by means of microsatellite allelotyping reflect allelic losses issupported by the detection of a decreased intensity of the AP-PCR
band assigned to chromosome 7. It has been demonstrated that decreased intensity of AP-PCR bands in tumor tissue reflects allelic
losses, whereas increased band intensity in tumor DNA reflects the
presence of extra copies of these sequences, which could be due togene amplification or tumor cell aneuploidy (4). In addition, ourfindings in cell line SK-PC-1. which contains three copies of chro
mosome 7 (21), well illustrate the fact that chromosomal gains mayactually hide a loss of genetic material. In this cell line: (a) micro-
satellite allelotyping results, that is, the retention of heterozygosity at7pter-q21 and the reduction of homozygosity at 7q21-qter, suggest
that a somatic recombination with a breakpoint at 7q21 occurred; and(b) AP-PCR evidence of reduced intensity of chromosome 7 band L
suggests that a homologous recombination with a defective chromatidwas the specific event, and that the duplicated chromosome was therecombinant-defective one (33). In fact, no change in the intensity of
nonpolymorphic band L would be observed if either the duplicatedchromosome was the normal one or no loss of material had occurredin the area where the sequence amplified by the AP-PCR primer is
located.The presence of TSG(s) on 7q is suggested by several lines of
evidence, including in vitro experiments and the detection of frequent7q deletions by either cytogenetic or molecular techniques in differenttypes of epithelial neoplasms. In vitro experiments showed that theintroduction of a normal chromosome 7 in a carcinoma cell lineresulted in inhibition of tumorigenicity of those cells when injectedinto nude mice, and that the insertion of intact human chromosome 7to immortalized human fibroblast cell lines having 7q31-32 LOH
restored the senescence properties in the cells (19, 20). Deletions in7q, with breakpoints varying from q 11 to q34. have been described bycytogenetic analysis in many different malignancies, including esoph-
ageal and gastric cancers (34, 35). The occurrence of interstitialdeletions and the variability of breakpoints in the different cancersfavor the view that gene loss, rather than hybrid gene formation, is thesignificant event resulting from chromosomal deletion. This is furthersuggested by cases of gastric and uterine tumors with simple chromosomal changes, in which the sole abnormality was interstitial 7qlosses (34. 36, 37). Also, molecular studies using VNTR probesshowed frequent 7q LOH in gastric (32%; Ref. 13) and breast (40%;Ret. 14) cancers . Recent studies using microsatellite markers reportedhigher frequencies of 7q LOH in colon, head-neck, prostate, ovary,
and breast cancers. In these five types of cancer, the SCDR waslocalized at 7q31.1-32, pointing to this region as a possible TSG-harboring site (15-18). The SCDR of chromosome 7q in pancreatic
cancer, described by our cases, overlaps that described above.Chromosome 7q losses including the region 7q31-35 have been
associated with worse prognosis when compared to matched stage IIIand IV cases in gastric (13) and breast cancers (14). Due to theparticular aggressiveness of pancreatic cancer and the small numberof cases, it is not possible to draw conclusions about the clinicalsignificance of 7q LOH in this tumor from this study.
ACKNOWLEDGMENTS
We thank Dr. N. L. Lemoine of the Imperial Cancer Research Fund.Hammersmith Hospital. London. United Kingdom, tor furnishing the pancreatic cancer cell lines shown in Fig. \A. We are also indebted to Dr. FranciscoX. Real, of the IstituìMunicipal de Investigado' Médica.Barcelona, Spain, for
supplying the DNA of pancreatic cancer cell-lines SK-PC-1 and IMIM-PC-2
together with their matched normal DNA. We also thank Dr. S. Orlandini forher skillful technical contribution.
REFERENCES
1. Knudson. A. G. Antioncogenes and human cancer. Proe. Nati. Acad. Sci. USA. 90:10914-10921. 1993.
2. James. C. D.. Carlbom. E.. Nordenskjold. M.. Collins. V. P.. and Cavenee. W. K.Mitotic recombination of chromosome 17 in astrocytomas. Proc. Nati. Acad. Sci.USA. «6.-2858-2862. 1989.
3812
Research. on February 9, 2016. © 1996 American Association for Cancercancerres.aacrjournals.org Downloaded from

7q LOH IN PANCREATIC CANCER
3. Louis. D. N., von Deimling. A., and Seizinger. B. R. A (CA)n dinucleotide repeatassay for evaluating loss of allelic heterozygosity in small and archival human braintumor specimens. Am. J. Pathol., 141: 777-782, 1992.
4. Peinado. M. A.. Malkhosyan. S.. Velazquez. A., and Perucho. M. Isolation andcharacterization of allelic losses and gains in colorectal tumors by arbitrarily primedPCR. Proc. Nati. Acad. Sci. USA. 89: 10065-10069. 1992.
5. lonov. A.. Peinado. M. A., Malkhosyan. S.. Shibata. D.. and Perucho. M. Ubiquitoussomatic mutations in simple repeated sequences reveal a new mechanism for coloniecarcinogenesis. Nature (Lond.). 363: 558-561. 1993.
6. Kohno. T.. Morishita. K.. Takano. H.. Shapiro. D. N.. and Yokota. J. Homozygousdeletion at chromosome 2q33 in human small-cell lung carcinoma identified byarbitrarily primed PCR genomic fingerprinting. Oncogene. 9: 103-108, 1994.
7. Seymour. A. B., Hruban. R. H.. Redston. M.. Caldas. C.. Powell. S. M.. Kinzler.K. W., Yeo. C. J., and Kern, S. E. Allelotype of pancreatic adenocarcinoma. CancerRes.. 54: 2761-2764. 1994.
8. Caldas. C.. Hahn. S. A., da Costa. L. T.. Redston. M. S.. Schulte. M.. Seymour. A. B.,Weinstein. C. L.. Hruban. R. H., Yeo, C. J.. and Kern. S. E. Frequent somaticmutations and homozygous deletions of the pió (A/75/) gene in pancreatic adenocarcinoma. Nat. Genet.. 8: 27-32. 1994.
9. Hahn, S. A., Seymour. A. B.. Shamsul Hoque, A. T. M.. Schutte, M., da Costa. L. T.,Redslon. M. S.. Caldas. C.. Weinslein. C. L.. Fischer. A., Yeo. C. J.. Hruban. R. H..and Kern. S. E. Allelotype of pancreatic adenocarcinoma using xenografi enrichment.Cancer Res.. 55: 4670-4675. 1995.
10. Johansson, B.. Bardi, G.. Heim. S.. Mandahl, N.. Martens. F.. Bak-Jensen, E.,Andrén-Sandberg, A., and Mitelman, F. Nonrandom chromosomal rearrangements inpancreatic carcinomas. Cancer (Phila.). 69: 1674-1681. 1992.
11. Bardi. G., Johansson. B., Pandis. N.. Mandahl. N.. Bak-Jensen. E.. Andrén-Sandberg.A.. Mitelman. F., and Heim, S. Karyotypic abnormalities in tumours of the pancreas.Br. J. Cancer, 67: 1106-1112, 1993.
12. Griffin. C. A.. Hruban. R. H.. Morsberger. L. A.. Elligham. T.. Long. P. P.. Jaffee.E. M.. Haunda. K. M.. Bohlander, S. K.. and Yeo. C. J. Consistent chromosomeabnormalities in adenocarcinoma of the pancreas. Cancer Res.. 55: 2394-2399. 1995.
13. Kuniyasu. H.. Yasui, W., Yokozaki, H.. Akagi, M.. Akama, Y., Kitahara, K.. Fujii.K.. and Tahara, E. Frequent loss of heterozygosity of the long arm of chromosome 7is closely associated with the progression of human gastric carcinomas. Int. J. Cancer.59: 597-600. 1994.
14. Bieche. !.. Chámpeme. M. H.. Matifas. F.. Hacene. K.. Callahan. R., and Lidereau, R.Loss of heterozygosity on chromosome 7q and aggressive primary breast cancer.Lancet. 339: 139-143, 1992.
15. Zenklusen. J. C., Bieche, I., Lindereau. R.. and Conti. C. J. (C-A) microsatellite repeat
D7S522 is the most commonly deleted region in human primary breast cancer. Proc.Nati. Acad. Sci. USA, 91: 12155-12158. 1994.
16. Zenklusen. J. C., Thompson. J. C.. Troncoso, P., Kagan. J., and Conti, C. J. Loss ofheterozygosity in human primary prostate carcinomas: a possible tumor suppressorgene at 7q3l.l. Cancer Res.. 54: 6370-6373, 1994.
17. Zenklusen. J. C.. Thompson. J. C.. Klein-Szanto. A. J.. and Conti. C. J. Frequent lossof heterozygosity in human primary squamous cell and colon carcinomas at 7q31.1:evidence for a broad range tumor suppressor gene. Cancer Res.. 55: 1347-1350.
1995.18. Zenklusen. J. C.. Weitzel. J. N.. Ball. H. G.. and Conti, C. J. Allelic loss al 7q31.1 in
human priman ovarian carcinomas suggests the existence of a tumor suppressorgene. Oncogene. //: 359-363. 1995.
19. Ogata. T., Ayusawa. D., Namba, M.. Takahashi. E.. Oshimura, M.. and Oishi. M.Chromosome 7 suppresses indefinite division of nontumorigenic immortalized human
fibroblast cell lines KMST-6 and SUSM-1. Mol. Cell. Biol.. 13: 6036-6043. 1993.20. Zenklusen, J. C.. Oshimura. M.. Barret. J. C.. and Conti. C. J. Inhibition of tumori-
genicity of a murine squamous cell carcinoma (SCC) cell line by a putative tumoursuppressor gene on human chromosome 7. Oncogene. 9: 2817-2825. 1994.
21. Vila1, M. R., Llórela.J., Schüssler.M. H.. Berrozpe. G., Welt. S., and Real, F. X. New
pancreas cancer cell lines that represent distinct slages of ductal differentiation. Lab.Invest.. 72: 395-404, 1995.
22. Berrozpe. G.. Schaeffer. J.. Peinado. M. A.. Real. F. X., and Perucho. M. Comparativeanalysis of mutations in the p53 and Ki-ra.v genes in pancreatic cancer. Int. J. Cancer.58: 185-191. 1994.
23. Kloeppel. G. Pancreatic Pathology. New York: Churchill Livingstone, 1984.24. Drwinga, H. L.. Toji. L. H., Kim, C. H., Greene, A. E., and Mulivor. R. A. NIGMS
human/rodent somatic cell hybrid mapping panel I and 2. Genomics. 16: 311-314,1993.
25. Yasuda, J.. Navarro, M.. Malkhosyan. S.. Velazquez. A.. Arribas, R.. Sekiya. T.. andPerucho. M. Chromosomal assignment of human DNA fingerprint sequences bysimultaneous hybridization to arbitrarily primed PCR products from human/rodentmonochromosome cell hybrids. Genomics, 34: 1-5, 1996.
26. Jen, J.. Kim. H.. Pianladosi. S.. Liu. Z.. Levitt. R.. Sistonen. P.. Kinzler, K.,Vogelstein. B., and Hamilton. S. Allelic loss of chromosome I8q and prognosis incolorectal cancer. N. Engl. J. Med.. 331: 213-221. 1994.
27. Gruis. N.. Abeln. E.. Bardoel, A.. Delvilee. P.. Frants, R.. and Comelisse, C.PCR-based microsalellitc polymorphisms in the detection of loss of heterozygosity infresh and archival tumour tissue. Br. J. Cancer, 68: 308-313, 1993.
28. Zar, J. H. Biostatislical Analysis. Englewood Cliffs. NJ: McGraw-Hill. Inc., 1991.
29. Johansson. B.. Bardi. G.. Pandis. N.. Gorunova, L., Backman, P.. Mandahl. N.,Dawiskiba. S.. Andren Sandberg, A., Heim. S.. and Mitelman, F. Karyotypic patternof pancreatic adenocarcinomas correlates with survival and tumor grade. Int. J.Cancer, 5«:8-13, 1994.
30. Scarpa. A.. Capelli. P., Mukai. K.. Zamboni. G.. Oda. T.. lacono. C., and Hirohashi.S. Pancreatic adenocarcinomas frequently show p53 gene mutations. Am. J. Pathol..142: 1534-1543. 1993.
31. Redston. M. S.. Caldas, C.. Seymour. A. B.. Hruban. R. H.. da Costa. L.. Yeo, C. J..and Kern, S. E. p53 mutations in pancreatic carcinoma and evidence of commoninvolvement of homocopolymer tracts in DNA microdeletions. Cancer Res., 54:3025-3033. 1994.
32. Gorunova. L.. Johansson. B.. Dawiskiba. S.. Andrén-Sandberg, A.. Jin. Y., Mandahl.
N.. Heim. S., and Mitelman. F. Massive cytogenetic heterogeneity in a pancreaticcarcinoma: 54 karyotypically unrelated clones. Genes Chromosomes & Cancer. 14:259-266, 1995.
33. Hansen. M., and Cavenee. W. Genetics of cancer predisposition. Cancer Res., 47:5518-5527, 1987.
34. Xiao. S., Geng. J. S.. Feng, X. L., Liu, X. Q., Liu. Q. Z.. and Li. P. Cytogenetic studiesof eight primary gastric cancers. Cancer Genet. Cytogenel.. 58: 79-84, 1992.
35. Xiao. S.. Feng. X. L.. Geng. J. S.. Yan. F. C.. Liu. Q. Z., and Li. P. Cytogeneticstudies of five primary esophageal cancers. Cancer Genet. Cytogenet.. 55: 197-205.
1991.36. Pandis. N.. Karaiskos. C.. Bardi. G.. Sfikas. K.. Tserkezoglou. A.. Fotiou. S., and
Heim. S. Chromosome analysis of uterine adenomyosis. Cancer Genet. Cytogenet..80: 118-120. 1995.
37. Osella, P., Carlson. A.. Wyandt. H.. and Milunsky. A. Cylogenelic studies of eightsquamous cell carcinomas of the head and neck: deletion of 7q. a possible primarychromosomal event. Cancer Genet. Cytogenet., 59: 73-78, 1992.
3813
Research. on February 9, 2016. © 1996 American Association for Cancercancerres.aacrjournals.org Downloaded from

1996;56:3808-3813. Cancer Res Alberto Achille, Maria O. Biasi, Giuseppe Zamboni, et al. Chromosome 7q Allelic Losses in Pancreatic Carcinoma
Updated version
http://cancerres.aacrjournals.org/content/56/16/3808
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on February 9, 2016. © 1996 American Association for Cancercancerres.aacrjournals.org Downloaded from
Copyright © 2022 FDOKUMEN




















