
Summer surface waters in the Gulf of California: Prime habitat for biological N2 fixation

Author's personal copy
Changing climate in the Gulf of California
Salvador E. Lluch-Cota a,!, Alejandro Parés-Sierra b, Víctor O. Magaña-Rueda c, Francisco Arreguín-Sánchez d,Gastón Bazzino a, Hugo Herrera-Cervantes e, Daniel Lluch-Belda d
aCentro de Investigaciones Biológicas del Noroeste (CIBNOR), P.O. Box 128, La Paz, Baja California Sur 23000, MexicobCentro de Investigación Científica y Educación Superior de Ensenada (CICESE), P.O. Box 2732, Ensenada, Baja California 22800, MexicocCentro de Ciencias de la Atmósfera, Universidad Nacional Autónoma de México Circuito Exterior s/n, Ciudad Universitaria, Del. Coyoacán, 04510 DF, MexicodCentro Interdisciplinario de Ciencias Marinas (CICIMAR-IPN), P.O. Box 592, La Paz, Baja California Sur 23000, MexicoeCentro de Investigación Científica y Educación Superior de Ensenada, Unidad Baja California Sur (CICESE-La Paz), Miraflores 334, Frac. Bella Vista, La Paz, BCS 23050, Mexico
a r t i c l e i n f o
Article history:Available online 29 September 2010
a b s t r a c t
We conducted a four year interdisciplinary collaborative project focused in the Gulf of California, themost important fishing region for Mexico. We reviewed published reports, collected and analyzed phys-ical, chemical and ecological data sets, and developed models for the physical (atmosphere and ocean)and ecological components of this large marine ecosystem, to examine prevalent scientific questionsregarding climate variability and change in the region, covering three time scales (ENSO, decadal-to-interdecadal, and long-term trend). We were able to describe how the Gulf of California influences thenorthward propagation of coastal trapped Kelvin waves associated with El Niño (ENSO) events, andhow this signal, together with changes in the atmospheric forcing, results in a ENSO signature insidethe Gulf. For the decadal-to-multidecadal scales, we found coherent trends among series, and with thePacific Decadal Oscillation (PDO). The long-term temperature signal for the Gulf of California shows awarming that occurred in the mid 20th century, approximately a decade before that in the CaliforniaCurrent. This signal is coherent with fluctuations in the industrial fisheries catch records (sardine andshrimps). For the recent decades we found no significant sustained long-term trend in any of the timeseries of physical and ecological variables that we considered. Instead, variability seems to be fully dom-inated by the interaction of PDO and ENSO. We stress the urgent need for more modeling efforts and theestablishment of interdisciplinary (physical and biological) observation platforms for the marine environ-ment in the Gulf of California.
! 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Mexico is strongly committed to global climate change issues.The Federal Government has recently announced a national agendaon climate change, called ‘‘Programa Especial de Cambio Climáti-co”. This initiative demands rapid progress in national and regionalscientific research on the detection and attribution of anthropo-genic climate change impacts on ecosystems, and geographicaland cross sectors analyses of vulnerability and development ofadaptation strategies. Progress has been achieved in some aspectsof energy, tourism, agriculture, water management, and urbandevelopment (Martínez and Fernández, 2006); however, theadvances in other sectors remain limited, particularly for fisheries,even though it is recognized as a key sector for future food securityand sustainable development.
Fisheries systems are extremely complex, involving many fac-tors that contribute to the observed variability (e.g., natural forcingand interacting variability) and are influenced by many confound-ing factors (fishing dynamics and their direct and indirect impacts,internal ecosystem dynamics, land use and coastal development,etc.). The challenges associated with studying a system for whichobservations are often indirect, costly, and generally scarce makeit difficult to disentangle the impacts of anthropogenic activitiesfrom those of climate change.
A first step towards detection of climate change impacts inmarine ecosystems is to understand the natural variability. Toaccomplish this for the Gulf of California, we developed a multidis-ciplinary research project that was strongly inspired by theGLOBEC science plan (Lluch-Cota et al., 2006). The project involvedfive research groups, from different institutions and expertise:atmospheric scientists from the Center for Atmospheric Sciencesof the National University (CCA-Universidad Nacional Autónomade México), oceanographers from the Physical OceanographyDivision of CICESE (Centro de Investigación Científica y EducaciónSuperior de Ensenada), two groups from CICIMAR (Centro
0079-6611/$ - see front matter ! 2010 Elsevier Ltd. All rights reserved.doi:10.1016/j.pocean.2010.09.007
! Corresponding author. Address: Centro de Investigaciones Biológicas delNoroeste (CIBNOR), Mar Bermejo 195, Colonia Playa Palo de Santa Rita, La Paz,BCS 23090, Mexico. Tel.: +52 612 123 84 32; fax: +52 612 125 36 25.
E-mail address: [email protected] (S.E. Lluch-Cota).
Progress in Oceanography 87 (2010) 114–126
Contents lists available at ScienceDirect
Progress in Oceanography
journal homepage: www.elsevier .com/locate /pocean

Author's personal copy
Interdisciplinario de Ciencias Marinas del Instituto PolitécnicoNacional) dealing with ecosystem modeling and retrospectiveanalyses, and a group on Fisheries Oceanography from CIBNOR(Centro de Investigaciones Biológicas del Noroeste). This projectused retrospective analyses and model simulations to examinesome key questions for all interannual scales of variability: highfrequency (ENSO scale), decadal-to-interdecadal, and long-termtrends.
The Gulf of California is a strategic region for the marine fisher-ies in Mexico, with fully developed industries targeting the smallpelagic species (contributing up to 40% of total national marinecatch), shrimp (Mexico‘s most important fishery in terms of eco-nomic income and employment), jumbo squid (which has becomeimportant during the last 10–15 years), and yellowfin tuna. Localsmall-scale coastal fisheries are also active in the Gulf, represent-ing the most important fishery activities in terms of subsistenceand social values. Other relevant human activities include shrimpaquaculture, tourism, and agriculture (Lluch-Cota et al., 2007).
At the same time, this is a region of major conservation interest.The Gulf has one of the most diverse marine biological communi-ties in the world, with 4852 species of invertebrates (excludingcopepods and ostracods), 891 species of fish, and 222 tetrapod spe-cies. The degree of endemism is high for some taxa; for example,almost 16% of the macro-invertebrate species reported for the Gulfare endemic, most of them Mollusca and Arthropoda, and 10% ofthe fishes are endemic (Lluch-Cota et al., 2007). Several speciesof turtles, birds, and marine mammals are also of special interestfor conservation.
Physical and ecological dynamics are largely seasonal. Atmo-spheric wind forcing is dominated by weak winds blowing from
the southeast during the summer and strong winds blowing fromthe northwest during the winter. Ocean circulation is stronglyinfluenced by this forcing and, together with tidal cycles and air-sea heat exchange, defines the circulation pattern. These processespromote enrichment of nutrients by winter upwelling along theeastern coast and the topographically-induced year round tidalmixing that occurs around the Midriff islands (Angel de la Guardaand Tiburón, Fig. 1). Both of these mechanisms are easily detectedby satellite imagery.
1.1. ENSO scale
In the Gulf of California, El Niño-Southern Oscillation (ENSO) isthe most important interannual variability signal (Herrera-Cervan-tes et al., 2007). The magnitude of El Niño impacts and its regionalmanifestation depend, among other things, on the strength of theevents. During the last 30 years, for which some high quality envi-ronmental observational records exist for the Gulf of California,two major El Niño events occurred, in 1982–1983 and 1997–1998.
Two physical mechanisms have been proposed to link the Gulfand ENSO dynamics (Strub and James, 2002). These are the in-creased poleward (or decreased equatorward) winds caused bychanges in the mid-latitude atmospheric circulation related toENSO (Emery and Hamilton, 1985) and northward propagation ofcoastal trapped waves (Parés-Sierra and O‘Brien, 1989). Bakun(1996) noted that these coastal trapped waves bypass the entranceto the Gulf or travel into and around one or more of the interiortopographic basins and then re-emerge at the tip of the Baja Cali-fornia peninsula to continue poleward. Previous work has analyzedthis northward propagation at the continental scale (Lluch-Cota
01005001000150020002500300035004000
La Paz
Empalme
PacificOcean
116°W 113 110
25
Mexico mainland
28
Los Cabos
Midriff Islands
Depth (m)
Baja Californiapeninsula
Fig. 1. Geographic location of the Gulf of California and main bathymetric features.
S.E. Lluch-Cota et al. / Progress in Oceanography 87 (2010) 114–126 115
Author's personal copy
et al., 2001, 2003; Strub and James, 2002) and inside the Gulf (Soto-Mardones et al., 1999; Lavín et al., 2003), but because of the tem-poral resolution used in those analyses, the mechanisms by whichthese waves bypass or enter the Gulf remained unknown. Ripa(1990) and Beier (1997) used numerical models to propose thatsea level elevation changes and the circulation along the east andwest coasts of the Gulf of California could be attributed to baroclin-ic forcing at the mouth, as a Kelvin wave propagating into the Gulftowards the head, and going around to the Baja California coast. Tocontrast these modeling results and increase mesoscale dynamics,we posed
Question 1: How does northward propagation of coastal trappedwaves associated with ENSO interact with the Gulf of California(i.e. signal ‘‘jumping” the Gulf’s mouth or traveling around it)?
Many observations exist regarding ENSO impacts inside theGulf of California. However, comprehensive analyses of the mech-anisms by which ENSO changes the regional oceanographicdynamics are difficult because of a lack of sufficient spatial and
temporal coverage by scientific surveys. This has been approachedin this study by analyzing high-resolution satellite-derived SSTdata. Soto-Mardones et al. (1999) and Lavín et al. (2003) examinedSST for different periods (1984–1995 and 1984–2000, respec-tively), finding that both cold and warm ENSO episodes were evi-dent in the central region, south of the Midriff Islands but not inthe northern Gulf of California. Lavín et al. (2003) also compareda spatially averaged anomaly series of sea-surface temperaturefor the entire Gulf of California to the Southern Oscillation Index(SOI) and found a significant positive relationship during the stron-gest ENSO events. While these analyses have provided some clues,we decided to take advantage of numerical ocean modeling to fur-ther approach
Question 2: How are circulation patterns in the Gulf of Californiamodified by ENSO?
Because ENSO is considered the strongest factor contributing toecological variability beyond the annual cycle, we found the con-trasting observations and reports on the ecological impacts of
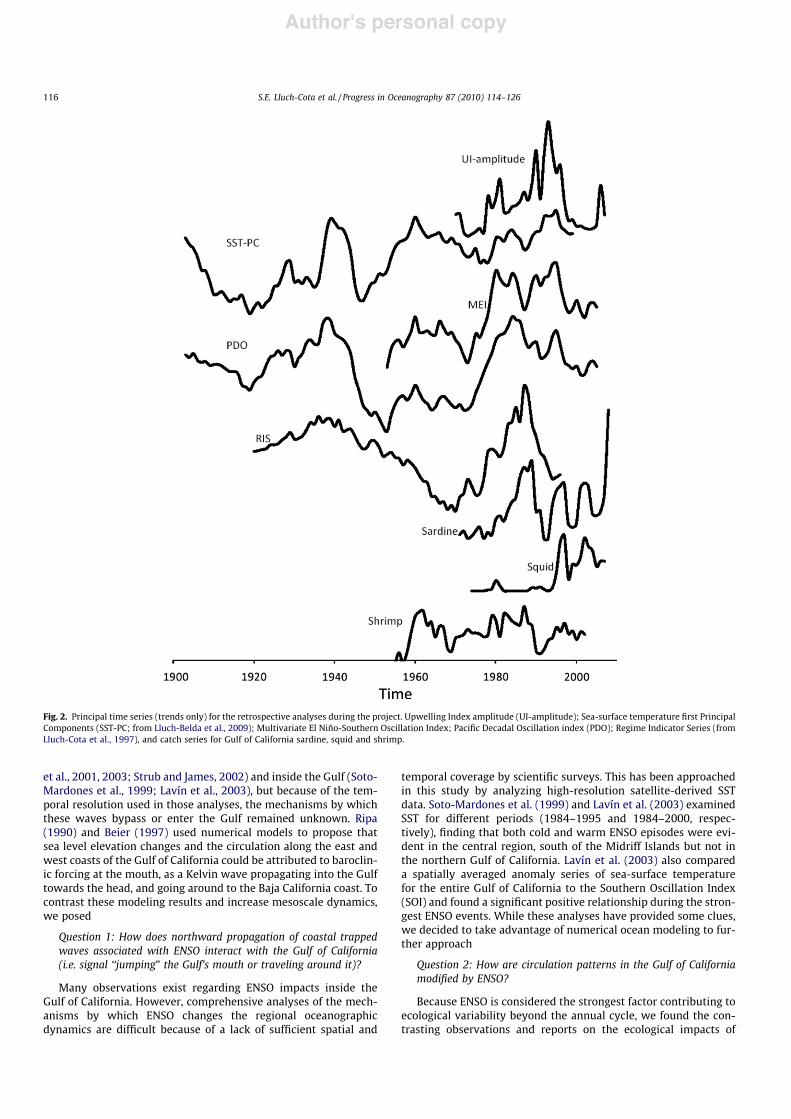
Fig. 2. Principal time series (trends only) for the retrospective analyses during the project. Upwelling Index amplitude (UI-amplitude); Sea-surface temperature first PrincipalComponents (SST-PC; from Lluch-Belda et al., 2009); Multivariate El Niño-Southern Oscillation Index; Pacific Decadal Oscillation index (PDO); Regime Indicator Series (fromLluch-Cota et al., 1997), and catch series for Gulf of California sardine, squid and shrimp.
116 S.E. Lluch-Cota et al. / Progress in Oceanography 87 (2010) 114–126

Author's personal copy
ENSO inside the Gulf particularly interesting; some of these aresummarized in Table 1 for the two strongest El Niño events inthe last 30 years. For example, for the same El Niño event differentauthors found phytoplankton biomass to decrease (Santamaría-delAngel et al., 1994) or remain unchanged (Valdez-Holguín and Lara-Lara, 1987), and for different ENSO events reproduction of brownpelicans succeeded (Jaques et al., 1996) or failed (Sydeman et al.,2001) in the midriff islands. Within the important limitationsposed by the quantity, quality and length of ecological time series,we examined the Gulf of California ecosystem response to ENSOthrough
Question 3: Why are there several contrasting observations on theecological consequences of ENSO inside the Gulf?
1.2. Decadal-to-multidecadal scale
Low frequency (decadal-to-multidecadal) climate variation hasbeen widely studied during the last 30 years, especially for theNorth Pacific. Alternation between periods of low frequency vari-ability and regime shifts have been documented for physical vari-ables such as temperature (both sea and air, instrumental andreconstructed; Wooster and Hollowed, 1995; Ware, 1995; Schwinget al., 2002), atmospheric pressure and associated wind fields(Miller and Schneider, 2000; Parrish et al., 2000), ocean currentsand advection (Wooster and Hollowed, 1995), upwelling intensity(Schwing and Mendelssohn, 1997; Bograd et al., 2001), and mixedlayer depth (Bernal and Chelton, 1984). Ecological changes in theNorth Pacific, e.g. changes in primary and secondary productivity(Polovina et al., 1995; Roemmich and McGowan, 1995;Lavaniegos-Espejo and Ohman, 2007), faunal distributions (Brintonand Townsend, 2003) and recruitment in several fish stocks (Lluch-Belda et al., 1989; Bakun, 1996; Mantua et al., 1997; Klyashtorin,2001; Norton, 1999; Hare and Mantua, 2000; Botsford, 2001;Botsford and Lawrence, 2002; Chavez et al., 2003) have been ob-served as well.
Although some underlying mechanisms of variability at thesefrequencies in the physical system (Wooster and Hollowed,1995; Schwing et al., 2010) and their coupling with ecosystemshave been hypothesized (Chavez et al., 2003; Lluch-Belda et al.,2003; Tourre et al., 2007), understanding is quite rudimentarycompared to that of ENSO processes. Furthermore, because of thenecessity for longer and more complete time series, and the manyuncertainties inherent in climate models on decadal and longertime scales, the regional manifestations of such variations are hardto investigate (IPCC, 2007; Lean and Rind, 2009). For these reasons,
we focused on the detection of decadal-to-multidecadal variability.For the retrospective analyses we asked
Question 4: Can we detect decadal-to-multidecadal variability inthe Gulf of California based on existing temperature and ecologicaldata?
One interesting aspect of decadal-to-multidecadal variability isthe potential regional scale modulation of the seasonal cycle.Bograd et al. (2002) and Palacios et al. (2004) have reportedchanges in the seasonality of the physical environment in the Cal-ifornia Current, and discussed potential implications for the pelagicecosystem. For the Gulf of California, Lluch-Cota (2000) also sug-gested a strong tendency towards increased seasonality (as thedifferences between summer and winter), particularly for theintensity of upwelling in the eastern central Gulf. Investigating thismodulation is critical to building future scenarios since phenolog-ical variations are one of the most common impacts attributed toclimate change (Edwards and Richardson, 2004). We explored thistopic through
Question 5: Is seasonality changing at the decadal-to-multidecadaltime scale?
1.3. Long-term trends
Determining how long-term trends in climate variability will bereflected in marine ecosystems is a critical question worldwide.Several studies and recent reviews already cover aspects of regio-nal signals of global change in the physical environment (Bakun,1990; IPCC, 2007; Lean and Rind, 2009), and ecosystem compo-nents (Hays et al., 2005; Höegh-Guldberg et al., 2007; Pörtnerand Knust, 2007; Stramma et al., 2008), and structure and func-tioning (Roessig et al., 2004; Harley et al., 2006). Currently thereare no formal detection reports for the Gulf of California, at leastbased on modeling and observational records. Thus it was relevantto evaluate
Question 6: Are there detectable trends, different from those asso-ciated with interannual variability, in the data and observationalrecords available for the Gulf of California? and Question 7: Towhat extent could interannual variability affect ecosystem func-tioning (trophic flows)?
We realize that, besides limited observational records, the onlyoptions to analyze long-term oceanographic and ecological trendsin the Gulf rely on paleo-reconstructions (e.g., Herguera-Garcíaet al., 2003; Pride et al., 1999) and modeling. We did not examine
Table 1Contrasting reports on the ecological impacts of El Niño-Southern Oscillation events in the Gulf of California.
Reference Observation 1982–1983
1997–1998
Santamaría-del Angel et al. (1994) Decreased satellite-derived pigment concentration in some areas XValdez-Holguín and Lara-Lara (1987) Phytoplankton biomasses in the southern Gulf similar to previous years XAlvarez-Borrego and Lara-Lara (1991) Primary production remain nearly constant in the central and northern areas XJiménez-Pérez and Lara-Lara (1988), Lavaniegos-Espejo and
Lara-Lara (1990)Zooplankton biomass remained unchanged in the central Gulf X
Cortés-Altamirano and Núñez-Pasten (1992) No HAB events in an area of frequent blooms (Mazatlán) during ENSO XHuato and Lluch-Belda (1987) Sardine concentrated around the Midriff Islands XThunell (1998) Diatoms diminished in distribution and abundance in the central Gulf XNevárez-Martínez et al. (2001) Collapse of sardine fishery XJaques et al. (1996) Brown pelican colonies breeding successfully in the northern Gulf region but the
opposite in the southX
Kahru et al. (2004) Decreased pigment concentration from satellite XCortés-Altamirano and Sierra-Beltrán (2001) Harmful Algal Blooms events decreased during the event XSánchez-Velasco et al. (2000) Sardine reproduction failure XLluch-Cota et al. (2007) Collapse of squid fishery XSydeman et al. (2001) Reduced reproductive success in Midriff Islands pelican colonies X
S.E. Lluch-Cota et al. / Progress in Oceanography 87 (2010) 114–126 117

Author's personal copy
paleo – data in our project. In regards to modeling, we built projec-tions based on regional models nested within Global Climate Mod-els to answer
Question 8: how do the IPCC-based climate projections downscalefor the Gulf of California region?
2. Methods
In this section, we describe the data and models developed dur-ing the project, with references to the technical and scientific re-ports in which these methods are described in greater detail.
2.1. Time series
There are few long-term observational time series of environ-mental variables in the Gulf of California, making the analysis oflow frequency variability difficult. However, Lluch-Belda et al.(2009) explored publicly available global SST datasets spanningover a century, and created four time series for the Gulf by recon-structing the International Comprehensive Ocean–AtmosphereData Set Release 2.0 (ICOADS) with the extended Reynolds sea-surface temperature (ERSST) data and compared the resultingseries with the few series that have been already reported(Herguera-García et al., 2003; Leal et al., 2001; Lavín et al., 2003;Dean et al., 2004). These series were analyzed here to describe dec-adal and long-term trends.
For higher spatial resolution, we examined satellite-deriveddata for sea-surface temperature and ocean color (Herrera-Cervantes et al., 2010). Since these databases span only a few(recent) decades, they were used to explore the impacts of high fre-quency variability (El Niño) and recent trends, as well as to quan-tify the interannual variations of the annual amplitude signal.
To analyze the variability of atmospheric drivers relevant to theecosystem dynamics in the Gulf, we updated the upwelling indextime series originally published in Lluch-Cota (2000) to completethe period 1970–2009. This index, based on locally registeredwinds at the Empalme meteorological station (see Fig. 1), success-fully represents the dynamics of the central Gulf (Lluch-Cota,2000).
Catch records were also updated for the main fisheries re-sources (Fig. 2). For the Pacific sardine fishery in the Gulf of Califor-nia, the time series originally published by Schwartzlose et al.(1999) was extended to the entire 1970–2009 period using thedata published by Lluch-Cota et al. (2007) and the most recent an-nual reports presented at the Small Pelagic Technical Committeeworkshops (Comité Técnico de Pelágicos menores). Lluch-Beldaet al. (2009) reconstructed a time series for shrimp catch basedon several sources, representing the entire Mexican Pacific sinceno subset for the Gulf was available; however, landings in the Gulfof California constitute about 80% of total shrimp catch (Lluch-Beldaet al., 2009).
2.2. Models
2.2.1. Atmospheric modelThe construction of future climate scenarios is based on down-
scaling the results of several General Circulation Models (GCMs) tothe Gulf of California region. The use of dynamic regional climatemodels, such as the Meteorological Research Institute (MRI) model(http://www.mri-jma.go.jp/) with 22 km ! 22 km spatial scale,and the PRECIS (40 km ! 40 km) model provide valuable climatechange information for Mexico and Central America (http://www.metoffice.com/research/hadleycentre/models/PRECIS.html).
We considered the A1B scenario from the IPCC Data Distribu-tion Centre for the simulations. Projections cover the present and
future (2070–2090), and a subset of differences between presentand future conditions. For present conditions, coherence betweenmodel outputs and observed wind fields were successfully tested.
2.2.2. Oceanographic modelOcean circulation in the Gulf of California was modeled using a
nested Regional Ocean Modeling System (ROMS) within a largerdomain covering the entire Mexican Pacific. ROMS_TOOLS(http://roms.mpl.ird.fr) was used for nesting the models. Thenested domain had a spatial resolution of nearly 3.8 km, with 20vertical layers and realistic bathymetry. The model reproducedseveral of the observed features of the Gulf, including the forma-tion of the seasonally alternating gyres in the central and southernGulf (Figueroa et al., 2003) resulting from barotropic instabilities(Allende-Arandía, 2005), the distribution of temperatures relatedto these gyres, the seasonal alternation of an extended cyclonicgyre during the summer, and substitution by an smaller anticy-clonic gyre during winter in the northern part of the Gulf (Lavínet al., 1997). Both models (i.e. the large scale Pacific model andthe nested Gulf of California model) were forced at the surfaceby winds and heat fluxes from the North American RegionalReanalysis (NARR; http://www.emc.ncep.noaa.gov/mmb/rreanl/).The Pacific model was forced at the lateral boundaries using salin-ity, temperature, sea level and velocity fields from the globalreanalysis model Simple Ocean Data Assimilation (SODA; Cartonet al., 2005). The Gulf of California model was forced at its lateralboundaries by the Pacific model and by tides obtained from theglobal tidal model of Oregon State University (Egbert and Erofeeva,2002).
2.2.3. Ecological modelsThe main approach to modeling the Gulf of California ecosys-
tems in this project employed ECOPATH with ECOSIM platform(Christensen and Pauly, 1992; Walters et al., 1997; Christensenet al., 2000). This suite of models has been widely used to assessthe direct and indirect impacts of fishing effort on the structureand function of the ecosystems characterized within the Gulf(Table 2).
We identified existing ecosystemmodels constructed with ECO-PATH, most of them for regions where shrimp trawling takes place.Shrimp fishing in diverse ecosystems within the Gulf revealed dif-ferent effects from fishing pressure. In the northern (Morales-Zárate et al., 2004; Lozano, 2006; Lercari et al., 2007; Lercari andArreguín-Sánchez, 2009) and central Gulf (Arreguín-Sánchezet al., 2002) there were well developed shrimp trawl fisheries withstable yields, although in the north there were conflicts betweenexploitation and conservation. On the other hand, the shrimp fish-ery in the central Gulf of California was well developed and yieldswere stable (Arreguín-Sánchez et al., 2002), and simulation exper-iments have been conducted to support ecosystem based fisheriesmanagement (Arreguín-Sánchez and Martínez-Aguilar, 2003). Sev-eral models were constructed for the southern Gulf; those by Salc-ido-Guevara and Arreguín-Sánchez (2007) and Arreguín-Sánchezet al. (2004) detail the impact of the shrimp trawl fishery on theecosystem, while others such as Díaz-Uribe et al. (2007) and Arre-guín-Sánchez et al. (2007) consider the impacts of different fleets.The simulation experiments were performed using the ecosystemtrophic model representing the shrimp trawling fishery in the cen-tral Gulf of California. Besides harvesting strategies, the forcing fac-tors included a climate function affecting only primary production(bottom up control), and we followed the propagation of this signalthrough the food web. We realize climate forcing also affects othergroups, complicating the simple assumption of bottom up control;however, to simulate those processes we need much better under-standing of the mechanisms or, at least, significant empiricalrelationships.
118 S.E. Lluch-Cota et al. / Progress in Oceanography 87 (2010) 114–126

Author's personal copy
3. Results
Environmental variability in the Gulf of California is more com-plex than many areas of the Eastern Pacific, due to interactionsfrom the low frequency, high-latitude Pacific Decadal Oscillation(PDO) and the high-frequency effects of tropical ENSO events(Lluch-Cota et al., 2001). Regional scale long-term climate trendsare particularly hard to investigate because no synoptic andlong-term climate proxies exist, and the coarse resolution of inter-polated global scale databases and model results cannot be appliedto study this long, thin sea. This section reports on the progress to-wards answering the scientific questions posed in the Introduction.
3.1. ENSO
Question 1: How does northward propagation of coastal trappedwaves associated to the ENSO interact with the Gulf of California(i.e. signal ‘‘jumping” the Gulf’s mouth or traveling around it)?
In Herrera-Cervantes et al. (2007), we analyzed the North Amer-ican coastal ocean propagation of the 1997 El Niño signal (from 10"to 40"N; including the entire Gulf of California) based on weeklysatellite-derived sea-surface temperature time series. Resultsshowed that the Gulf plays a significant role in the poleward prop-agation of ENSO-related coastal waves. The propagation time alongthe coasts of the Gulf, from the continental to the peninsular sidesof the mouth, was estimated at 3 weeks (bypassing the Midriff Is-lands), for a propagation speed of about 150 km day"1, whichagrees with the modeled phase speed for this signal (Beier,1997). Because the internal Rossby deformation radius for thesecoastal waves is narrower than the distance between the continen-tal and peninsular sides of the Gulf‘s mouth, it is likely that the sig-nal enters the Gulf and undergoes significant weakening throughfriction and dispersion as it travels into and around it. However,when the amplitude and duration of strong El Niño events resultin more than one peak of persistent warm SST and thermoclinedepth anomalies, coastal trapped waves could generate more in-tense poleward advection, where a first episode could enter theGulf along both coasts and weaken due to friction and dispersion,leaving the Gulf at the southern tip of the peninsula some3–4 weeks later, and continue its poleward propagation along thewest coast. Later episodes (2–3 months later) would be more con-sistent north of 24"N and reach higher latitudes. Previous observa-tions by Ryan and Noble (2002) are coherent with this proposal;they reported that after the detection of the 1997–1998 El Niñoin summer, there was a reinforcement of the signal starting inNovember, with no direct correlation to local winds. Furthermore,the described process explains the observation by many authorssuggesting that the poleward propagation of coastal SST anomaliesduring the 1997 El Niño seemingly bypassed the Gulf without anynoticeable delay (Lluch-Cota et al., 2001; Strub and James, 2002),
and also explains why we frequently fail to detect moderate toweak El Niño signals at the west coast of the Baja Californiapeninsula.
Question 2: How are circulation patterns in the Gulf of Californiamodified by ENSO?
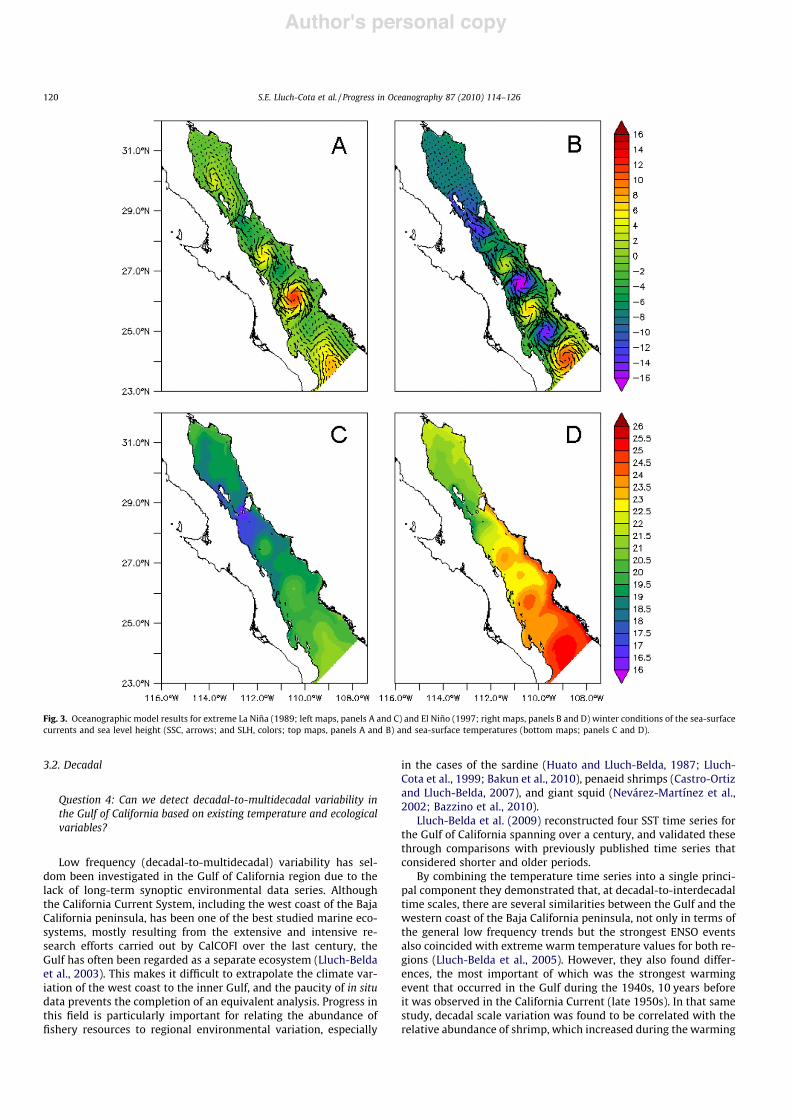
Using the oceanographic model, we explored the impacts on theGulf of California circulation of tropical ocean signals associatedwith El Niño/Southern Oscillation on the Gulf of California circula-tion. For this analysis, we compared the seasonal signal of temper-ature and circulation to the simulations of two extreme conditions,the 1997 El Niño and the 1989 La Niña. In addition to the strongdifferences in absolute winter temperatures, we found importantdifferences in the general patterns of the surface circulation. Thegeneral circulation pattern south of the large islands region wassimilar to the average climatology, dominated by alternating gyres.One important difference, however, was the intensity of the anticy-clonic gyres (Fig. 3). During the 1997 El Niño, the cyclonic and anti-cyclonic gyres were well-defined and of similar intensity, whileduring the 1989 La Niña the anticyclonic gyres virtually disap-peared. Another important difference occurred in the northernGulf, where the anticyclonic gyre was well-formed during LaNiña but disappeared during the El Niño event. These changesmay have caused differences in the vertical distributions of physi-cal properties and mixing dynamics, which may have influencedthe productivity and other biological processes.
Question 3: Why are there several contrasting observations onthe ecological consequences of ENSO inside the Gulf?
In Herrera-Cervantes et al. (2007, 2010), we explored the vari-ability of satellite-derived SST and chlorophyll fields using Empir-ical Orthogonal Functions (EOF) and their relationships to ENSOspatial signatures based on the distribution of correlation coeffi-cients between the Southern Oscillation Index (SOI) and tempera-ture and chlorophyll anomalies for the time series of each pixel.We showed, based on the coherence between the spatial distribu-tion of the EOF amplitudes and the SOI-temperature correlationcoefficients, that interannual variability of the chlorophyll pigmentconcentration in the Gulf of California was dominated by the ENSOinfluence. Spatially, the pattern showed the highest variability andstronger correlation with ENSO along the east coast (the upwellingcorridor) and southern Midriff island region (where tidal mixingoccurs), and lower variability and weaker correlation with ENSOalong the western margin. This pattern was also observed forSST. The proposed physical processes behind this pattern werethe disruption of the wind-driven upwelling corridor along the eastcoast of the Gulf, together with the propagation of ENSO-relatedcoastal waves being smoothed by bottom friction. This spatial sig-nature explained the apparent contradictory field observationsregarding the ecological impacts.
Table 2Existing Ecopath-based ecological models for the Gulf of California.
Reference Region Focus/ecosystem
Morales-Zárate et al. (2004) Northern Benthic: shrimpLercari et al. (2007), Lercari and Arreguín-Sánchez (2009) Northern Benthic: multispecies fisheriesLozano (2006) Northern BenthicHernández-Herrera and Cisneros-Mata (unpublished) Central Epipelagic and coastal ecosystemArreguín-Sánchez et al. (2002) Central Benthic: shrimpGorostieta Monjaraz (unpublished) Central western CoastalArreguín-Sánchez and Martínez-Aguilar (2003) Central Pelagic: sardineArreguín-Sánchez and Calderón (2002) Central Multispecies fisheriesRosas-Luis et al. (2008) Central Pelagic: squidArreguín-Sánchez et al. (2004, 2007), Díaz-Uribe (unpublished), Díaz-Uribe et al. (2007) Southern western (La Paz Bay) Coastal: artisanal fisheriesZetina-Rejón et al. (2001, 2003, 2004) Southern eastern (Huizache-Caimanero) Coastal: multispeciesSalcido-Guevara and Arreguín-Sánchez (2007) Southern Eastern Coastal
S.E. Lluch-Cota et al. / Progress in Oceanography 87 (2010) 114–126 119
Author's personal copy
3.2. Decadal
Question 4: Can we detect decadal-to-multidecadal variability inthe Gulf of California based on existing temperature and ecologicalvariables?
Low frequency (decadal-to-multidecadal) variability has sel-dom been investigated in the Gulf of California region due to thelack of long-term synoptic environmental data series. Althoughthe California Current System, including the west coast of the BajaCalifornia peninsula, has been one of the best studied marine eco-systems, mostly resulting from the extensive and intensive re-search efforts carried out by CalCOFI over the last century, theGulf has often been regarded as a separate ecosystem (Lluch-Beldaet al., 2003). This makes it difficult to extrapolate the climate var-iation of the west coast to the inner Gulf, and the paucity of in situdata prevents the completion of an equivalent analysis. Progress inthis field is particularly important for relating the abundance offishery resources to regional environmental variation, especially
in the cases of the sardine (Huato and Lluch-Belda, 1987; Lluch-Cota et al., 1999; Bakun et al., 2010), penaeid shrimps (Castro-Ortizand Lluch-Belda, 2007), and giant squid (Nevárez-Martínez et al.,2002; Bazzino et al., 2010).
Lluch-Belda et al. (2009) reconstructed four SST time series forthe Gulf of California spanning over a century, and validated thesethrough comparisons with previously published time series thatconsidered shorter and older periods.
By combining the temperature time series into a single princi-pal component they demonstrated that, at decadal-to-interdecadaltime scales, there are several similarities between the Gulf and thewestern coast of the Baja California peninsula, not only in terms ofthe general low frequency trends but the strongest ENSO eventsalso coincided with extreme warm temperature values for both re-gions (Lluch-Belda et al., 2005). However, they also found differ-ences, the most important of which was the strongest warmingevent that occurred in the Gulf during the 1940s, 10 years beforeit was observed in the California Current (late 1950s). In that samestudy, decadal scale variation was found to be correlated with therelative abundance of shrimp, which increased during the warming
Fig. 3. Oceanographic model results for extreme La Niña (1989; left maps, panels A and C) and El Niño (1997; right maps, panels B and D) winter conditions of the sea-surfacecurrents and sea level height (SSC, arrows; and SLH, colors; top maps, panels A and B) and sea-surface temperatures (bottom maps; panels C and D).
120 S.E. Lluch-Cota et al. / Progress in Oceanography 87 (2010) 114–126

Author's personal copy
with an unexplained 3-year lag, as well as with sardine, which in-creased during the cooling periods.
Question 5: Is seasonality changing at the decadal-to-multideca-dal time scale?
Strong seasonal amplitude is evident in virtually every physicaland ecological variable in the Gulf area (Lluch-Cota et al., 1999).For example, when compared to nearby sites on the west coastof Baja California and in the Eastern Tropical Pacific off Nayarit,Mexico, the seasonal amplitude of sea-surface temperature andpigment concentration in the central Gulf is more than double. In-side the Gulf, the amplitude of physical variables tends to increasefrom south to north. In the northern area, long-term studies havedocumented dramatic changes in sea-surface water temperaturesfrom a warm-temperate environment during the winter, as shoretemperature drops to lows of 8–12 "C, to a tropical-like summer,where nearshore temperatures rise to more than 30 "C. Havingsuch an extreme seasonality, and realizing that phenology changeshave been widely recognized as critical impacts of climate changein several different ecosystems (Edwards and Richardson, 2004),we analyzed the seasonal variability in temperature and pigmentconcentration, and the potential modulation from other scales ofnatural variation. We found that the annual temperature ampli-tude is highly variable between years, with changes occurringeverywhere in the Gulf except for the region around the Midriff Is-land (Herrera-Cervantes et al., 2007). Statistical relationships be-tween amplitude variations and the PDO were insignificant,while links to ENSO were more evident, with the largest ampli-tudes occurring during La Niña periods.
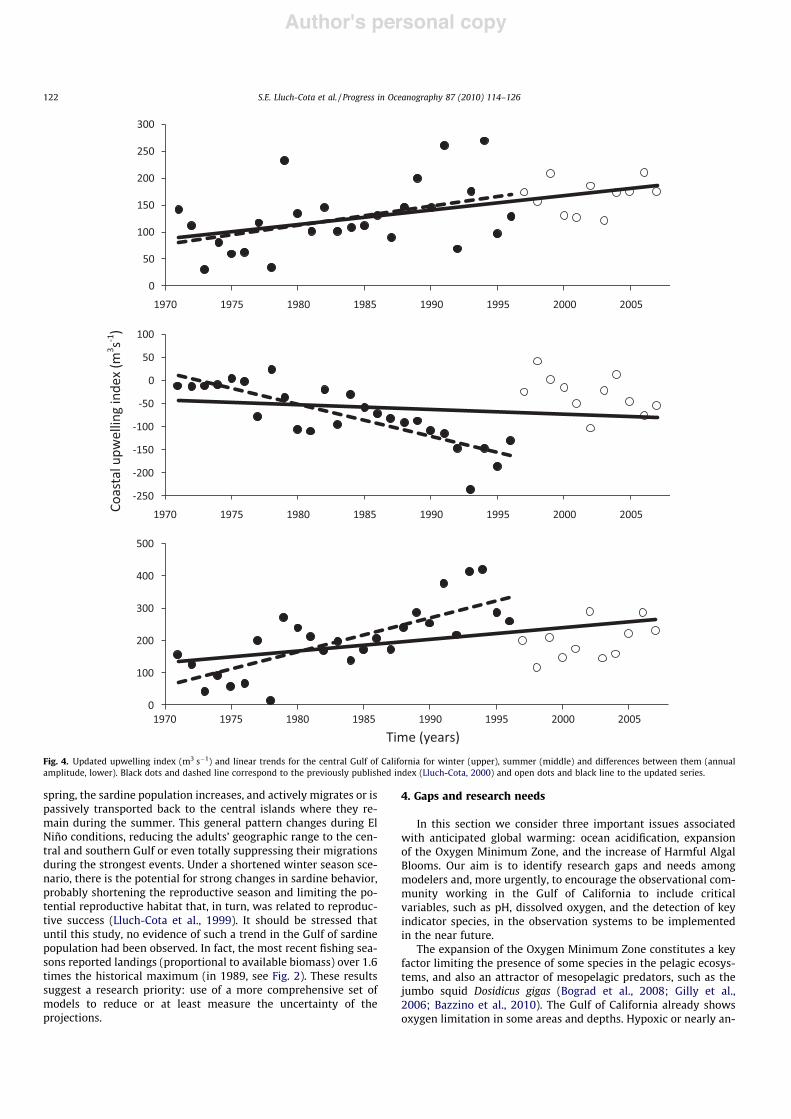
The computation and analysis of an upwelling index for the cen-tral Gulf of California completed a decade ago (Lluch-Cota, 2000)suggested a sustained trend of increasing annual amplitude, whichled the author to speculate about linkages with climate change. Theupdated version of this index portrays a different picture (Fig. 4). Itsuggests that the annual amplitude time series was highly variable,mostly composed of a period of relatively small differences betweenwinter and summer, from 1970 to the early 1980s (except for the1976–1981 period) an increasing trend and maximum differencesin the late 1990s, then a sharp decline afterwards. The general shapeof the curve strongly resembles the PDO (see Fig. 2).
3.3. Long-term trends (climate change detection)
Ecosystems are permanently under the influence of diverseinteracting forcing modes, including anthropogenic changes andnatural climate variability. For this reason, the detection and attri-bution of anthropogenic climate change is a statistical ‘‘signal-in-noise” problem (IPCC, 2007). Detection implies an observed statis-tically significant change in climate, and attribution would ideallybe the dissection of that change signal into different sources of var-iation; however, the actual possibility for discerning attribution isthrough formal statistical tests to demonstrate that certainchanges are unlikely to be triggered by the internal variability(IPCC, 2007). The first step is the identification of long-term trendsor shifts in the observed signals, typically temperature.
Question 6: Are there detectable trends, different from those asso-ciated with interannual variability, in the data and observationalrecords available for the Gulf of California?
The analysis reported in Lluch-Belda et al. (2009) showed that,when considering sea-surface temperature for the Gulf of Califor-nia during the entire 20th century, a positive long-term warmingtrend can be observed, similar to that reported for the CaliforniaCurrent, but comparatively slower. However, when one considersonly the second part of the century (since the early 1960s), whenthe global surface air temperature series shows the most dramatic
warming, no significant trends can be detected and, in fact, theslope is negative. The Gulf of California time series shares severalfeatures with the PDO and the Multivariate ENSO Index (MEI).Dean et al. (2004) analyzed a paleo-reconstruction from the Gulf,and recognized periodicities of 10–20 years, particularly in sedi-ments deposited during the 19th century, related to warm phasesof the PDO. Herguera-García et al. (2003), also based on paleo-reconstructions, reported that the average wintertime tempera-tures have been similar during the last 170 years and were coolerand warmer between 1700 and 1830, suggesting that thesechanges were related to intensification and relaxation of El Niño/La Niña events on decadal periods.
Based on much shorter time series of satellite-derived chloro-phyll in the Gulf, Herrera-Cervantes et al. (2010) identified a neg-ative relationship with the MEI during a period considered assustained warming (1999–2006), similar to what has been ob-served for most of the inter-tropical world ocean by Behrenfeldet al. (2006), and related to ocean stratification, one of the expectedeffects of climate change. It must be noted that this short period isused as an analogue for a warming trend, even when it is mostlydriven by the return to normal conditions after the strong 1997–1998 El Niño (Behrenfeld et al., 2006).
Question 7: To what extent could natural variability affect ecosys-tem functioning (trophic flows)?
We simulated three scales of variability (artificial series) repre-senting high frequency (ENSO-like), low frequency (bi-decadal cy-cles) and long-term trends (monotonic), and used them to forceprimary production in the ecological model. This is an oversimpli-fication of the climate impacts on ecosystems, since such variabil-ity will also affect other trophic levels directly, but it allowed us toisolate part of the bottom up control dynamics. Other mechanismsforcing upper trophic levels are not understood well enough to de-scribe patterns.
The high frequency variability produced a rapid ecosystem re-sponse, but resilience (the ecosystem capacity to return to stabilityafter a perturbation) overcame the impacts relatively quickly.However, when the low frequency forcing was introduced, shiftingconditions occurred and the ecosystem reorganized into differentstates. Our modeling exercise suggests it is important for manage-ment to consider changes in the vulnerability of ecosystems overtime. During different decadal periods (i.e. temperature regimes),the system showed different resilience; thus, intense fishing sce-narios associated with periods of high vulnerability could signifi-cantly impact the structure and functioning of the ecosystem,while similar fishing pressure could result in sustainable practicesduring periods of high system resilience.
Question 8: how do the IPCC-based climate projections down-scale for the Gulf of California region?
The project generated an extensive and publically availabledatabase containing the model runs for present and future climate(2070–2090) derived from the IPCC scenarios. We highlight thateven using the A1B1 emission scenario, differences between futureand present conditions tend to be relatively small for the Gulf ofCalifornia region, particularly during the winter season, whichwas our focus due to increased upwelling, sardine reproductionand stronger El Niño impacts.
We also found that while differences in summer and winter sur-face winds were rather small, the transitional seasons, spring andautumn, showed strong differences due to reduced northwesterlywinds. Fig. 5 shows the monthly distribution of the future surfacewind fields and the differences between future and present. Atpresent, southward movements of sardine for reproduction, fromthe Midriff Islands to the central Gulf, start in mid-to-late autumn,when the coastal wind-driven upwelling begins. During the early
S.E. Lluch-Cota et al. / Progress in Oceanography 87 (2010) 114–126 121
Author's personal copy
spring, the sardine population increases, and actively migrates or ispassively transported back to the central islands where they re-main during the summer. This general pattern changes during ElNiño conditions, reducing the adults’ geographic range to the cen-tral and southern Gulf or even totally suppressing their migrationsduring the strongest events. Under a shortened winter season sce-nario, there is the potential for strong changes in sardine behavior,probably shortening the reproductive season and limiting the po-tential reproductive habitat that, in turn, was related to reproduc-tive success (Lluch-Cota et al., 1999). It should be stressed thatuntil this study, no evidence of such a trend in the Gulf of sardinepopulation had been observed. In fact, the most recent fishing sea-sons reported landings (proportional to available biomass) over 1.6times the historical maximum (in 1989, see Fig. 2). These resultssuggest a research priority: use of a more comprehensive set ofmodels to reduce or at least measure the uncertainty of theprojections.
4. Gaps and research needs
In this section we consider three important issues associatedwith anticipated global warming: ocean acidification, expansionof the Oxygen Minimum Zone, and the increase of Harmful AlgalBlooms. Our aim is to identify research gaps and needs amongmodelers and, more urgently, to encourage the observational com-munity working in the Gulf of California to include criticalvariables, such as pH, dissolved oxygen, and the detection of keyindicator species, in the observation systems to be implementedin the near future.
The expansion of the Oxygen Minimum Zone constitutes a keyfactor limiting the presence of some species in the pelagic ecosys-tems, and also an attractor of mesopelagic predators, such as thejumbo squid Dosidicus gigas (Bograd et al., 2008; Gilly et al.,2006; Bazzino et al., 2010). The Gulf of California already showsoxygen limitation in some areas and depths. Hypoxic or nearly an-
Fig. 4. Updated upwelling index (m3 s"1) and linear trends for the central Gulf of California for winter (upper), summer (middle) and differences between them (annualamplitude, lower). Black dots and dashed line correspond to the previously published index (Lluch-Cota, 2000) and open dots and black line to the updated series.
122 S.E. Lluch-Cota et al. / Progress in Oceanography 87 (2010) 114–126

Author's personal copy
oxic environments have been detected in very shallow water in thesouthern Gulf (less than 100 m depth; Hendrickx, 2001). Off theSinaloa coast (southeastern part of the Gulf), the OMZ extendsroughly between depths of 150 and 750 m, and is completely de-void of benthic macrofauna (i.e., invertebrates and fishes). A dis-tinct, moderately diverse fauna is found from around 800 m to2300 m where oxygen concentrations increase to 2.0–2.5 ml l"1
(Hendrickx, 2001). The northern Gulf is not affected by severe hy-poxia along the sea floor.
We are not aware of any report on acidification of the Gulf ofCalifornia region; however, we identify this as an important issuesince no monitoring exists and the region is located in the bound-ary of aragonite saturation levels, limiting coral reef survival (Kley-pas et al., 2001). Most of the coral reefs within the Gulf have beenclassified as existing within marginal environmental conditions(Kleypas et al., 1999), close to the limits of minimum temperatures(Guaymas, Espiritu Santo, La Paz), high temperature (ConcepcionBay), high phosphate (Cabo Pulmo, Espiritu Santo, La Paz), lightpenetration (Guaymas, Concepcion Bay, Espiritu Santo, La Paz),and low aragonite saturation (Espiritu Santo, La Paz, Guaymas).For all the ocean acidification projections based on IPCC scenarios(Höegh-Guldberg et al., 2007), the Gulf of California ends outside ofthe suitable habitat for coral reefs. Further, as noted by theseauthors, synergies with other pressures are likely to occur, increas-ing the sensitivity of coral reef communities.
Harmful algal blooms (HAB) have been associated with massivemortality of marine organisms in the Gulf of California (sea birds,turtles, fish, and marine mammals; Sierra-Beltrán et al., 1997).The only relatively long and consistent HAB time series in the Gulfof California region, (22 years at Mazatlan Bay) shows that thenumber of toxic species and the frequency and duration of eventsare increasing (Cortés-Altamirano et al., 1999). Furthermore, non-native species have been observed, driving strong changes in phy-toplankton community structure changes (Sierra-Beltrán et al.,1998). Recently, Flores-Trujillo et al. (2009) analyzed the paleo-re-cords of the dinoflagellate Gymnodinium catenatum during the last#500 years, in the southern Gulf of California. This species is acommon component of the toxic algal blooms off the MexicanPacific coasts and in the Gulf where public health problems, humanmortality, and significant losses to aquaculture have been docu-mented (Alonso-Rodríguez and Páez-Osuna, 2003; Hernández-Becerril et al., 2007). The results obtained by Flores-Trujillo et al.(2009) indicate that G. catenatum abundance seems to bepositively/negatively modulated by La Niña/El Niño conditions,reflecting an inverse relationship with sea-surface temperature.
5. Conclusions
Through the interdisciplinary GLOBEC-like approach adoptedby this research project, we examined prevalent scientific ques-
Fig. 5. Atmospheric model-based surface wind projections for the regional domain including the Gulf of California, considering the A1B1 IPCC scenario. The vector maps showthe differences between future (2070–2090) and present (1990–2010) for each month.
S.E. Lluch-Cota et al. / Progress in Oceanography 87 (2010) 114–126 123

Author's personal copy
tions regarding climate variability and change in the Gulf of Cali-fornia, covering three time scales (ENSO, decadal-to-interdecadal,and long-term trend). The major challenge was, and remains, thelack of sufficient information in terms of time series length, geo-graphical coverage, and observational variables. However, we wereable to describe how the Gulf of California influences the north-ward propagation of coastal trapped Kelvin waves associated withEl Niño events, and how this signal, together with changes in theatmospheric forcing, results in a ENSO signature inside the Gulf.We also made an important observation that annual temperatureamplitude was larger during La Niña periods. In addition, we wereable to reconstruct environmental time series that allowed us todescribe decadal-to-multidecadal scales of variability, where wefound coherent trends with the PDO. The long-term temperaturesignal for the Gulf of California shows a warming that occurredin the mid 20th century, approximately a decade before that inthe California Current. This signal is coherent with fluctuations inthe catch records of industrial fisheries for sardine and shrimps.We found no significant sustained long-term trends in any of thetime series of physical and ecological variables that we considered.Instead, variability seems to be fully dominated by the interactionof PDO and ENSO.
The results from this project contribute to a better understand-ing of natural variability and other forcing factors that interact inthe Gulf of California. However, we are also aware of the long roadahead and the urgency of knowledge to better manage this largemarine ecosystem to be prepared for future environmentalchanges. In the short term, we envision as urgent the use of a morecomprehensive set of climate and oceanographic models, and theestablishment of interdisciplinary (physical and biological) obser-vation platforms for the marine environment in the Gulf.
Acknowledgments
This was an integration of the project SEMARNAT-CONACYT0002. DLB y FAS received support from the National PolytechnicInstitute through COFAA and EDI.
References
Allende-Arandía, M.E., unpublished. Giros de Mediana escala en el Golfo deCalifornia: balance energético, estudio numérico. M.S. Thesis, Departamentode Oceanografía Física, Centro de Investigación Científica y de EducaciónSuperior de Ensenada, Baja California, Mexico.
Alonso-Rodríguez, R., Páez-Osuna, F., 2003. Nutrients, phytoplankton and harmfulalgal blooms in shrimp ponds: a review with special reference to the situationin the Gulf of California. Aquaculture 219 (1–4), 317–336.
Alvarez-Borrego, S., Lara-Lara, J.R., 1991. The physical environment and primaryproductivity of the Gulf of California. In: Dauphin, J.P., Simoneit, B.R.T. (Eds.),The Gulf and Peninsular Province of the Californias, vol. 47. AmericanAssociation of Petroleum Geologists Memoir, pp. 555–567.
Arreguín-Sánchez, F., Arcos, E., Chavez, E.A., 2002. Flows of biomass and structure inan exploited benthic ecosystem in the Gulf of California, Mexico. EcologicalModelling 156, 167–183.
Arreguín-Sánchez, F., Calderón, L.E., 2002. Evaluating harvesting strategies forfisheries in the Central Gulf of California ecosystem. In: The Use of EcosystemModels to Investigate Multispecies Management Strategies for CaptureFisheries. Fisheries Centre Research Reports 10(2), 135–141.
Arreguín-Sánchez, F., Martínez-Aguilar, S., 2003. Manejo adaptativo de la pesqueríade sardina, Sardinops caeruleus, del Golfo de California. In: Quiñónez-Velázquez, C., Elorduy-Garay, J.F. (Eds.), Ambiente y pesquería de pelágicosmenores en el noroeste de México. Instituto Politécnico Nacional, México, pp.1–16.
Arreguín-Sánchez, F., Hernández-Herrera, A., Ramírez-Rodríguez, M., Pérez-España,H., 2004. Optimal management scenarios for the artisanal fisheries in theecosystem of La Paz Bay, Baja California Sur, México. Ecological Modelling 172,373–382.
Arreguín-Sánchez, F., del Monte-Luna, P., Díaz-Uribe, J.G., Gorostieta, M., Chávez,E.A., Ronzón-Rodríguez, R., 2007.Trophic model for the ecosystem of La Paz Bay,Southern Baja California peninsula, Mexico. Fisheries Centre Research Reports15(6), 134–160.
Bakun, A., 1990. Global climate change and intensification of coastal oceanupwelling. Science 247, 198–201.
Bakun, A., 1996. Patterns in the Ocean: Ocean Processes and Marine PopulationDynamics. University of California Sea Grant, San Diego, California, USA, incooperation with Centro de Investigaciones Biológicas de Noroeste, La Paz, BajaCalifornia Sur, Mexico. 323 p.
Bakun, A., Babcock, E.A., Lluch-Cota, S.E., Santora, C., Salvadeo, C.J., 2010. Issues ofecosystem-based management of forage fisheries in ‘‘open‘‘ non-stationaryecosystems: the example of the sardine fishery in the Gulf of California. Reviewsin Fish Biology and Fisheries 20, 9–29.
Bazzino, G., Gilly, W.F., Markaida, U., Salinas-Zavala, C.A., Ramos-Castillejos, J., 2010.Horizontal migrations, vertical habitat utilization and diet of the jumbo squid(Dosidicus gigas) in the Pacific Ocean off Baja California Sur, Mexico. Progress inOceanography 86 (1–2), 59–71.
Behrenfeld, M.J., O‘Malley, R.T., Siegel, D.A., McClain, C.R., Sarmiento, J.L.,Feldman, G.C., Milligan, A.J., Falkowski, P.G., Letelier, R.M., Boss, E.S., 2006.Climate driven trends in contemporary ocean productivity. Nature 444, 752–755.
Beier, E., 1997. A numerical investigation of the annual variability in the Gulf ofCalifornia. Journal of Physical Oceanography 27, 615–632.
Bernal, P.A., Chelton, D.B., 1984. Low frequency and large scale biological variabilityin the California Current, 1949–1978. Expert consultation to examine changesin abundance and species composition of neritic fish resources. FAO FisheriesReport (FAO) 291, 713–730.
Bograd, S.J., Chereskin, T.K., Roemmich, D., 2001. Transport of mass, heat, salt,and nutrients in the southern California Current System: annual cycle andinterannual variability. Journal of Geophysical Research 106 (C5), 9255–9275.
Bograd, S., Schwing, F., Mendelssohn, R., Green-Jessen, P., 2002. On the changingseasonality over the North Pacific. Geophysical Research Letters 29 (9).doi:10.1029/2001GL013790.
Bograd, S.G., Castro, C.G., Di Lorenzo, E., Palacios, D.M., Bailey, H., Gilly, W., Chavez,F.P., 2008. Oxygen declines and the shoaling of the hypoxic boundary in theCalifornia Current. Geophysical Research Letters 35, L12607. doi:10.1029/2008GL034185.
Botsford, L.W., 2001. Physical influences on recruitment to California Currentinvertebrate populations on multiple scales. ICES Journal of Marine Science 58(5), 1081–1091.
Botsford, L.W., Lawrence, C.A., 2002. Patterns of co-variability among CaliforniaCurrent chinook salmon, coho salmon, Dungeness crab, and physicaloceanographic conditions. Progress in Oceanography 53 (2–4), 283–305.
Brinton, E., Townsend, A., 2003. Decadal variability in abundances of the dominanteuphausiid species in southern sectors of the California Current. Deep-SeaResearch II 50, 2449–2472.
Carton, J.A., Giese, B.S., Grodsky, S.A., 2005. Sea level rise and the warming of theoceans in the SODA ocean reanalysis. Journal of Geophysical Research 110,C09006.
Castro-Ortiz, J.L., Lluch-Belda, D., 2007. Low frequency variability of fishingresources, climate, and ocean. Fisheries Research 85, 186–196.
Chavez, F.P., Ryan, J., Lluch-Cota, S.E., Niquen, C.M., 2003. From anchovies tosardines and back: multidecadal change in the Pacific Ocean. Science 299, 217–221.
Christensen, V., Pauly, D., 1992. Ecopath II—a software for balancing steady-stateecosystem models and calculating network characteristics. EcologicalModelling 61, 169–185.
Christensen, V., Walters, C.J., Pauly, D., 2000. Ecopath with Ecosim Version 4, Helpsystem!. University of British Columbia, Fisheries Centre, Vancouver, Canadaand ICLARM, Penang, Malaysia.
Cortés-Altamirano, R., Núñez-Pasten, A., 1992. Doce años (1979–1990) de registrosde mareas rojas en la Bahía de Mazatlán, Sin., México. Anales del Instituto deCiencias del Mar y Limnología 19, 113–121.
Cortés-Altamirano, R., Licea-Durán, S., Gómez-Aguirre, S., 1999. Evidencias deaumento de microalgas nocivas en la bahía de Mazatlán, Sin., México. In:Tresierra-Aguilar, A.E., Culquichicon-Malpica, Z.G. (Eds.), Proceedings del VIIICongreso Latinoamericano sobre Ciencias del Mar. Universidad de Trujillo,Lima, Perú.
Cortés-Altamirano, R., Sierra-Beltrán, A.P., 2001. Mitigación de microalgas nocivas,su impacto negativo en las Pesquerías, Acuacultura y Salud Pública. In: Alejo-Armenta, C.A., Gastelum, J.A. (Eds.), Memorias del Primer Foro Estatal de Cienciay Tecnología: Sinaloa, soluciones para el desarrollo, vol. 1. Gobierno del Estadode Sinaloa y Consejo Estatal de Ciencia y Tecnología, México.
Dean, W., Pride, C., Thunell, R., 2004. Geochemical cycles in sediments deposited onthe slopes of the Guaymas and Carmen Basins of the Gulf of California over thelast 180 years. Quaternary Science Reviews 23, 1817–1833.
Díaz-Uribe, J.G., unpublished. Estrategias de evaluación para el manejo de lapesquería artesanal del huachinango (Lutjanus peru) en el Sur del Golfo deCalifornia. Ph.D. Thesis. Centro de Investigaciones Biológicas del Noroeste. LaPaz, Baja California Sur, Mexico.
Díaz-Uribe, J.G., Arreguín-Sánchez, F., Cisneros-Mata, M.A., 2007. Multispeciesperspective for small-scale fisheries management: a trophic analysis of La PazBay in the Gulf of California, Mexico. Ecological Modelling 201, 205–222.
Edwards, M., Richardson, A.J., 2004. Impact of climate change on marine pelagicphenology and trophic mismatch. Nature 430, 881–884.
Egbert, G., Erofeeva, S., 2002. Efficient inverse modeling od baroscopic ocean tides.Journal of Atmospheric and Oceanic Technology 19, 183–204.
Emery, W., Hamilton, K., 1985. Atmospheric forcing of interannual variability in thenortheast Pacific Ocean: connections with El Niño. Journal of GeophysicalResearch 90, 857–868.
124 S.E. Lluch-Cota et al. / Progress in Oceanography 87 (2010) 114–126

Author's personal copy
Figueroa, J.M., Marinone, S.G., Lavin, M.F., 2003. A description of geostrophic gyresin the southern Gulf of California. In: Velasco-Fuentes, O.U., Sheinbaum, J.,Ochoa, J. (Eds.), Nonlinear Processes in Geophysical Fluid Dynamics. KluwerAcademic Publishers, The Netherlands, pp. 237–255.
Flores-Trujillo, J.G., Helenes, J., Herguera, J.C., Orellana-Cepeda, E., 2009.Palynological record (1483–1994) of Gymnodinium catenatum in PescaderoBasin, southern Gulf of California, Mexico. Marine Micropaleontology 73, 80–89.
Gilly, W.F., Markaida, U., Baxter, C.H., Block, B.A., Boustany, A., Zeidberg, L.,Reisenbichler, K., Robison, B., Bazzino, G., Salinas-Zavala, C.A., 2006. Vertical andhorizontal migrations by jumbo squid, Dosidicus gigas, revealed by electronictagging. Marine Ecology Progress Series 324, 1–17.
Gorostieta Monjaraz, M., unpublished. Dinámica de la estructura trófica delecosistema de Bahía Concepción, B.C.S., México. M.S. Thesis, CentroInterdisciplinario de Ciencias Marinas del Instituto Politécnico Nacional, LaPaz, Baja California Sur, Mexico.
Hare, S.R., Mantua, N.J., 2000. Empirical evidence for North Pacific regime shifts in1977 and 1989. Progress in Oceanography 47, 103–145.
Harley, C.D.G., Randall Hughes, A., Hultgren, K.M., Miner, B.G., Sorte, C.J.B., Thornber,C.S., Rodriguez, L.F., Tomanek, L., Williams, S.L., 2006. The impacts of climatechange in coastal marine systems. Ecology Letters 9, 228–241.
Hays, G.C., Richardson, A.J., Robinson, C., 2005. Climate change and marineplankton. Trends in Ecology and Evolution 20 (6), 337–344.
Hendrickx, M.E., 2001. Occurrence of a continental slope deep-water decapodcrustacean community along the edge of the minimum oxygen zone in thesoutheastern Gulf of California, Mexico. Belgian Journal of Zoology 131, 71–86.
Herguera-García, J.C., Bernal-Franco, G.R., Molina-Cruz, A., 2003. Decadal surfaceocean variability in the lower Gulf of California: records for the past 300 years.Geofisica Internacional 42, 397–406.
Hernández-Herrera, A., Cisneros-Mata, M.A., unpublished. Biomass flux model forthe epipelagic and coastal ecosystem in the Gulf of California. Reporte Interno.Centro Interdisciplinario de Ciencias Marinas del Instituto Politécnico Nacional,La Paz, Baja California Sur, México.
Hernández-Becerril, D.U., Alonso-Rodríguez, R., Álvarez-Góngora, C., Barón-Campis,S.A., Ceballos-Corona, G., Herrera-Silveira, J., Meave del Castillo, M.E., Juárez-Ruíz, N., Merino-Virgilio, F., Morales-Blake, A., Ochoa, J.L., Orellana-Cepeda, E.,Ramírez-Camarena, C., Rodríguez-Salvador, R., 2007. Toxic and harmful marinephytoplankton and microalgae (HABs) in Mexican coasts. Journal ofEnvironmental Science and Health Part A 42, 1349–1363.
Herrera-Cervantes, H., Lluch-Cota, D.B., Lluch-Cota, S.E., Gutiérrez de Velasco, G.,2007. The ENSO signature in sea-surface temperature in the Gulf of California.Journal of Marine Research 65, 589–605.
Herrera-Cervantes, H., Lluch-Cota, S.E., Lluch-Cota, D.B., Gutierrez de Velasco, G.,Lluch-Belda, D., 2010. ENSO influence on satellite-derived chlorophyll trends inthe Gulf of California. Atmósfera 23, 253–262.
Höegh-Guldberg, O., Mumby, P.J., Hooten, A.J., Steneck, R.S., Greenfield, P., Gomez,E., Harvell, C.D., Sale, P.F., Edwards, A.J., Caldeira, K., Knowlton, N., Eakin, C.M.,Iglesias-Prieto, R., Muthiga, N., Bradbury, R.H., Dubi, A., Hatziolos, M.E., 2007.Coral reefs under rapid climate change and ocean acidification. Science 318,1737–1742.
Huato, L., Lluch-Belda, D., 1987. Mesoscale cycles in the series of environmentalindices related to the sardine fishery in the Gulf of California. CaliforniaCooperative Oceanic Fisheries Investigations Reports 28, 128–134.
IPCC, 2007. Climate change 2007: the physical science basis. In: Solomon, S., Qin, D.,Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L. (Eds.),Contribution of Working Group I to the Fourth Assessment Report of theIntergovernmental Panel on Climate Change. Cambridge University Press,Cambridge United Kingdom and New York, NY, USA, p. 996.
Jaques, D.L., Strong, C.S., Keeney, T.W., 1996. Brown pelican roosting patterns andresponses to disturbance at Mugu lagoon and other nonbreeding sites in thesouthern California bight, Tech. Rep. 54, US Department of the Interior, NationalBiological Service, Cooperative Park Studies Unit, The University of Arizona,Tucson, Arizona. 63 p.
Jiménez-Pérez, L.C., Lara-Lara, J.R., 1988. Zooplancton biomass and copepodcommunity structure in the Gulf of California during the 1982–1983 El Niñoevent. California Cooperative Oceanic Fisheries Investigations Reports 12, 122–128.
Kahru, M., Marinone, S.G., Lluch-Cota, S.E., Parés-Sierra, A., Mitchell, B.G., 2004.Ocean-color variability in the Gulf of California: scales from days to ENSO.Deep-Sea Research II 51, 139–146.
Kleypas, J.A., McManus, J.W., Meñe, L.A.B., 1999. Environmental limits to coral reefdevelopment: where do we draw the line? American Zoology 39, 146–159.
Kleypas, J.A., Buddemeier, R.W., Gattuso, J.P., 2001. The future of coral reefs in an ageof global change. International Journal of Earth Science 90, 426–437.
Klyashtorin, L.B., 2001. Climate change and long-term fluctuations of commercialcatches: the possibility of forecasting. Rome, FAO Fisheries Tech. Paper410. 98 p.
Lavaniegos-Espejo, B.E., Lara-Lara, J.R., 1990. Zooplankton of the Gulf of Californiaafter the 1982 1983 El Niño event: biomass distribution and abundance. PacificScience 44 (3), 297–310.
Lavaniegos-Espejo, B.E., Ohman, M.D., 2007. Coherence of long-term variations ofzooplankton in two sectors of the California Current System. Progress inOceanography 75 (1), 42–69.
Lavín, M.F., Durazo, R., Palacios, E., Argote, M.L., Carrillo, L., 1997. Lagrangianobservations of the circulation in the northern Gulf of California. Journal ofPhysical Oceanography, 27, 2298–2305.
Lavín, M.F., Palacios-Hernandez, E., Cabrera, C., 2003. Sea surface temperatureanomalies and trend in the Gulf of California. Geofísica Internacional 42, 363–375.
Leal, A., López-Martínez, J., Chávez, E.A., Hernández-Vázquez, S., Méndez-Tenorio, F.,2001. Interannual variability of the reproductive period of the brown shrimp,Farfantepenaeus californiensis (Holmes, 1900) (Decapoda, Natantia).Crustaceana 74, 839–851.
Lean, J.L., Rind, D.H., 2009. How will earth‘s surface temperature change in thefuture decades? Geophysical Research Letters 36, L15708.
Lercari, D., Arreguín-Sánchez, F., Le Quesne, W., 2007. An ecosystem simulationmodel of the northern Gulf of California. In: Le Quesne, W.J.F., Arreguín-Sánchez, F., Heymans, S.J.J. (Eds.), INCOFISH Ecosystem Models: Transiting fromEcopath to Ecospace. Fisheries Centre Research Reports 15(6). Fisheries Centre,University of British Columbia, pp. 100–113.
Lercari, D., Arreguín-Sánchez, F., 2009. An ecosystem modelling approach toderiving viable harvest strategies for multispecies management of theNorthern Gulf of California. Aquatic Conservation: Marine and FreshwaterEcosystems 19 (4), 384–397.
Lluch-Belda, D., Crawford, R.J.M., Kawasaki, T., MacCall, A.D., Parrish, R.H.,Schwartzlose, R.A., Smith, P.E., 1989. World-wide fluctuations of sardine andanchovy stocks: the regime problem. South African Journal of Marine Science 8,195–205.
Lluch-Belda, D., Lluch-Cota, D.B., Lluch-Cota, S.E., 2003. The temperate-tropicaleastern Pacific ecotone: Baja California’s biological transition zones. Journal ofOceanography 59, 503–513.
Lluch-Belda, D., Lluch-Cota, D.B., Lluch-Cota, S.E., 2005. Changes in marine faunaldistributions and ENSO events in the California Current. FisheriesOceanography 14, 458–467.
Lluch-Belda, D., del Monte-Luna, P., Lluch-Cota, S.E., 2009. 20th Century variabilityof the Gulf of California SST. California Cooperative Oceanic FisheriesInvestigations Reports 50, 147–154.
Lluch-Cota, D.B., Hernández-Vázquez, S., Lluch- Cota, S.E., 1997. Empiricalinvestigation on the relationship between climate and small pelagic globalregimes and El Niño-southern Oscillation (ENSO). FAO Fisheries Circular No.934. FAO, Rome. 48 p.
Lluch-Cota, D.B., Wooster, W.S., Hare, S.R., 2001. Sea surface temperature variabilityin coastal areas of the Northeastern Pacific related to the El Niño-SouthernOscillation and the Pacific Decadal Oscillation. Geophysical Research Letters 28,2029–2032.
Lluch-Cota, D.B., Wooster, W.S., Hare, S.R., Lluch-Belda, D., Parés-Sierra, A., 2003.Principal modes and related frequencies of sea surface temperaturevariability in the Pacific coast of North America. Journal of Oceanography59, 477–488.
Lluch-Cota, S.E., 2000. Coastal upwelling in the eastern Gulf of California.Oceanologica Acta 23, 731–740.
Lluch-Cota, S.E., Lluch-Cota, D.B., Lluch-Belda, D., Nevárez-Martínez, M.O., Parés-Sierra, A., Hernández-Vázquez, S., 1999. Variability of sardine catch as related toenrichment, concentration, and retention processes in the central Gulf ofCalifornia. California Cooperative Oceanic Fisheries Investigations Reports 40,184–190.
Lluch-Cota, S.E., Pares-Sierra, A., Magaña-Rueda, V.O., Arreguín-Sánchez, F., 2006.Modeling climate change in the Gulf of California. GLOBEC InternationalNewsletter 12 (2), 70–71.
Lluch-Cota, S.E., Aragón-Noriega, E.A., Arreguín-Sánchez, F., Aurioles-Gamboa, D.,Bautista-Romero, J.J., Brusca, R.C., Cervantes-Duarte, R., Cortéz-Altamirano, R.,Del-Monte-Luna, P., Esquivel-Herrera, A., Fernández, G., Hendrickx, M.E.,Hernández-Vázquez, S., Herrera-Cervantes, H., Kahru, M., Lavín, M., Lluch-Belda, D., Lluch-Cota, D.B., López-Martínez, J., Marinone, S.G., Nevárez-Martínez,M.O., Ortega-García, S., Palacios-Castro, E., Parés-Sierra, A., Ponce-Díaz, G.,Ramírez-Rodríguez, M.E., Salinas-Zavala, C.A., Schwartzlose, R.A., Sierra-Beltrán,P.A., 2007. The Gulf of California: review of ecosystem status and sustainabilitychallenges. Progress in Oceanography 73, 1–26.
Lozano, H., unpublished. Historical ecosystem modelling of the upper Gulf ofCalifornia (Mexico): following 50 years of change. Ph.D. Thesis. University ofBritish Columbia, Canada.
Mantua, N.J., Hare, S.R., Zhang, Y., Wallace, J.M., Francis, R.C., 1997. A Pacificinterdecadal climate oscillation with impacts on salmon production. Bulletin ofAmerican Meteorology Society 78, 1069–1079.
Martínez, J., Fernández, A., 2006. Cambio climático: una visión desde México.Instituto Nacional de Ecología – Secretaría de Medio Ambiente y RecursosNaturales, Mexico. 350 p
Miller, A.J., Schneider, N., 2000. Interdecadal climate regime dynamics in the NorthPacific Ocean: theories, observations and ecosystem impacts. Progress inOceanography 47 (2–4), 355–379.
Morales-Zárate, M.V., Arreguín-Sánchez, F., López-Martínez, J., Lluch-Cota, S.E.,2004. Ecosystem trophic structure and energy flux in the Northern Gulf ofCalifornia, México. Ecological Modelling 174, 331–345.
Nevárez-Martínez, M.O., Lluch-Belda, D., Cisneros-Mata, M.A., Santos-Molina, J.P.,Martínez-Zavala, M.D.L.A., Lluch-Cota, S.E., 2001. Distribution and abundance ofthe Pacific sardine (Sardinops sagax) in the Gulf of California and their relationwith the environment. Progress in Oceanography 49, 565–580.
Nevárez-Martínez, M.O., Rivera-Parra, G.I., Morales-Bojórquez, E., López-Martínez,J., Lluch-Cota, D.B., Miranda-Mier, E., Cervantes-Valle, C., 2002. The jumbo squid(Dosidicus gigas) fishery of the Gulf of California and its relation toenvironmental variability. Investigaciones Marinas 30 (1), 193–194.
S.E. Lluch-Cota et al. / Progress in Oceanography 87 (2010) 114–126 125

Author's personal copy
Norton, J.G., 1999. Apparent habitat extensions of dolphin fish (Coryphaenahippurus) in response to climate transients in the California Current. ScientiaMarina 6 (3–4), 239–260.
Palacios, D.M., Bograd, S.J., Mendelssohn, R., Schwing, F.B., 2004. Long-term andseasonal trends in stratification in the California Current, 1950–1993. Journal ofGeophysical Research 109, 1–12.
Parés-Sierra, A., O’Brien, J.J., 1989. The seasonal and interannual variability of theCalifornia Current System: a numerical model. Journal of Geophysical Research93 (C3), 3159–3180.
Parrish, R.H., Schwing, F.B., Mendelssohn, R., 2000. Mid-latitude wind stress: theenergy source for climatic shifts in the North Pacific Ocean. FisheriesOceanography 9, 224–238.
Polovina, J.J., Mitchum, G.T., Evans, G.T., 1995. Decadal and basin-scale variation onthe mixed layer depth and the impact on the biological production in thecentral and North Pacific, 1960–88. Deep-Sea Research 42, 1701–1716.
Pörtner, H.O., Knust, R., 2007. Climate change affects marine fishes through theoxygen limitation of thermal tolerance. Science 315, 95–97.
Pride, C., Thunell, R., Sigman, D., Keigwin, L., Altabet, M., Tappa, E., 1999. Nitrogenisotopic variations in the Gulf of California since the last deglaciation: responseto global climate change. Paleoceanography 14 (3), 397–409.
Ripa, P., 1990. Seasonal circulation in the Gulf of California. Annales Geophysicae 8,559–564.
Roemmich, D., McGowan, J., 1995. Climatic warming and the decline of zooplanktonin the California Current. Science 267, 1324–1326.
Roessig, J.M., Woodley, C.M., Cech Jr., J.J., Hansen, L.J., 2004. Effects of global climatechange on marine and estuarine fishes and fisheries. Reviews in Fish Biologyand Fisheries 14, 251–275.
Rosas-Luis, R., Salinas-Zavala, C.A., Koch, V., del Monte-Luna, P., Morales-Zárate, V.,2008. Importance of jumbo squid Dosidicus gigas (Orbigny, 1835) in the pelagicecosystem of the central Gulf of California. Ecological Modelling 218 (1–2),149–161.
Ryan, H.F., Noble, M., 2002. Sea level response to ENSO along the central Californiacoast: how the 1997–1998 event compares with the historic record. Progress inOceanography 54 (1–4), 149–169.
Salcido-Guevara, L.A., Arreguín-Sánchez, F. 2007. A benthic ecosystem model of theSinaloa Continental Shelf, Mexico. In: Le Quesne, W., Arreguín-Sánchez, F.Heymans, S.J. (Eds.), INCOFISH Ecosystem Models: Transiting from Ecopath toEcospace. Fisheries Center, University of British Columbia, Canada, 188p.
Sánchez-Velasco, L., Shirasago, B., Cisneros-Mata, M.A., Avalos-García, C., 2000.Spatial distribution of small pelagic fish larvae in the Gulf of California and itsrelation to El Niño 1997–1998. Journal of Plankton Research 22, 1611–1618.
Santamaría del Angel, E., Alvarez-Borrego, S., Muller-Karger, F.E., 1994. The 1982–1984 El Niño in the Gulf of California as seen in coastal zone color scannerimagery. Journal of Geophysical Research 99, 7423–7431.
Schwartzlose, R.A., Alheit, J., Bakun, A., Baumgartner, T., Cloete, R., Crawford, R.J.M.,Fletcher, W.J., Green-Ruiz, Y., Hagen, E., Kawasaki, T., Lluch-Belda, D., Lluch-Cota, S.E., MacCall, A.D., Matsuura, Y., Nevares-Martinez, M.O., Parrish, R.H., Roy,C., Serra, R., Shust, K.V., Ward, N.M., Zuzunaga, J.Z., 1999. Worldwide large-scalefluctuations of sardine and anchovy populations. South African Journal ofMarine Science 21, 289–347.
Schwing, F.B., Mendelssohn, R., 1997. Increased coastal upwelling in the CaliforniaCurrent System. Journal of Geophysical Research 102 (C2), 3421–3438.
Schwing, F.B., Murphree, T., deWitt, L., Green, P.M., 2002. The evolution of oceanicand atmospheric anomalies in the northeast Pacific during the El Nino and LaNina events of 1995–2001. Progress in Oceanography 54, 459–491.
Schwing, F.B., Mendelssohn, R., Bograd, S.J., Overland, J.E., Wang, M., Ito, S., 2010.Climate change, teleconnection patterns, and regional processes forcing marinepopulations in the Pacific. Journal of Marine Systems 79 (3–4), 245–257.
Sierra-Beltrán, A., Palafox-Uribe, M., Grajales-Montiel, J., Cruz-Villacorta, A., Ochoa,J.L., 1997. Sea bird mortality at Cabo San Lucas: evidence that domoic acid isspreading. Toxicon 35, 447–454.
Sierra-Beltrán, A.P., Cruz, A., Nuñez, E., Del Villar, L.M., Cerecero, J., Ochoa, J.L., 1998.An overview of the marine food poisoning in Mexico. Toxicon 36, 1493–1502.
Soto-Mardones, L., Marinone, S.G., Parés-Sierra, A., 1999. Time and spatialvariability of sea surface temperature in the Gulf of California. CienciasMarinas 25, 1–30.
Stramma, L., Johnson, G.C., Sprintall, J., Mohrholz, V., 2008. Expanding oxygen-minimum zones in the tropical oceans. Science 320, 655.
Strub, P.T., James, C., 2002. Altimeter-derived surface circulation in the large-scalePacific gyres. Part 2: 1997–1998 El Niño anomalies. Progress in Oceanography53, 185–214.
Sydeman, W.J., Hester, M.M., Thayer, J.A., Gress, F., Martin, P., Buffa, P., 2001. Climatechange, reproductive performance and diet composition of marine birds in thesouthern California Current system, 1969–1997. Progress in Oceanography 49,309–329.
Thunell, R., 1998. Seasonal and annual variability in particle fluxes in the Gulf ofCalifornia: a response to climate forcing. Deep-Sea Research I 45, 2059–2083.
Tourre, Y.M., Lluch-Cota, S.E., White, W.B., 2007. Global multi-decadal ocean climateand small-pelagic fish population. Environmental Research Letters 2, 1–9.
Valdez-Holguín, J.E., Lara-Lara, J.R., 1987. Productividad primaria en el Golfo deCalifornia: efectos de El Niño 1982–1983. Ciencias Marinas 13, 34–50.
Walters, C., Christensen, V., Pauly, D., 1997. Structuring dynamic models ofexploited ecosystems from trophic mass-balance assessments. Reviews ofFish Biology and Fisheries 7, 1–34.
Ware, D.M., 1995. A century and a half of change in the climate of the NE Pacific.Fisheries Oceanography 4, 267–277.
Wooster, W.S., Hollowed, A.B., 1995. Decadal scale variations in the easternsubarctic Pacific. I. Winter ocean conditions. In: Beamish, R.J. (Eds.), Climatechange and northern fish populations. Canadian Special Publication of Fisheriesand Aquatic Sciences, 121, 81–85.
Zetina-Rejón, M.J., Arreguín-Sánchez, F., Chavéz, E.A., 2001. Using an ecosystemmodeling approach to assess the management of a mexican coastal lagoonsystem. California Cooperative Oceanic Fisheries Investigations Reports 42, 88–96.
Zetina-Rejón, M.J., Arreguín-Sánchez, F., Chavéz, E.A., 2003. Trophic structure andflows of energy in the Huizache–Caimanero lagoon complex on the Pacific coastof Mexico. Estuarine, Coastal and Shelf Science 57, 803–815.
Zetina-Rejón, M.J., Arreguín-Sánchez, F., Chavéz, E.A., 2004. Exploration ofharvesting strategies for the management of a Mexican coastal lagoon fishery.Ecological Modelling 172, 361–372.
126 S.E. Lluch-Cota et al. / Progress in Oceanography 87 (2010) 114–126

This article appeared in a journal published by Elsevier. The attached
copy is furnished to the author for internal non-commercial research
and education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling or
licensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of the
article (e.g. in Word or Tex form) to their personal website or
institutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies are
encouraged to visit:
http://www.elsevier.com/copyright
Copyright © 2022 FDOKUMEN




















