
Gulf Coast Phragmites
14
538 American Journal of Botany 99(3): 538–551. 2012. American Journal of Botany 99(3): 538–551, 2012; http://www.amjbot.org/ © 2012 Botanical Society of America Even though most biologists would agree that a species rep- resents a fundamental biological unit, many different species concepts have been suggested (De Queiroz, 2007). Frequently mentioned in textbooks is the biological species concept (BSP) which implies that individuals capable of interbreeding and producing fertile offspring belong to the same species (Mayr, 1942). While this definition generally works well for verte- brates, it may be difficult to apply to plants. One reason is poly- ploidization, a phenomenon common in plants but rare in animals. Polyploidization following hybridization, resulting in allopolyploidy, may re-establish disomic segregation, allowing hybrids to escape infertility. The genus Phragmites is one of many plant genera where ap- plication of the species concept is challenging partly due to poly- ploidy and associated interfertility barriers. Phragmites is distributed from the tropics to the cold temperate areas of both hemispheres, and includes five species, which are hardly distin- guishable morphologically and each showing cytological variation. Four of the species, P. frutescens H. Scholz, P. japonicus Steud., P. karka (Retz.) Trin. ex Steud., and P. mauritianus Kunth, have restricted geographical distributions (Clevering and Lissner, 1999), whereas P. australis (Cav.) Trin. ex Steud. is widely distributed in temperate and subtropical areas. Phylogeographically, most P. aus- tralis genotypes belong to a poorly structured group, with largely reticulate interrelationships, and a distribution as wide as the genus as a whole (Lambertini et al., 2006). The small number of species within the genus, the lack of diploids, and the occurrence of several ploidy levels are characteristics typical of old “taxonomic com- plexes” (Stebbins, 1971). The lack of genetic structure within the species might indicate that there is sufficient reproductive contact among populations within the complex to prevent differentiation and, ultimately, speciation. The reproductive contact can occur as cross-pollination between geographically distant parents, but also as a consequence of seed dispersal by wind and migratory birds, rafting of eroded plant fragments and marsh sods, as well as human-aided spread of seeds and rhizome fragments. Dispersal of Phragmites by humans is known to have oc- curred repeatedly at various geographical scales. The spreading of a European P. australis lineage in North America is particu- larly well documented, and has been addressed primarily because of its aggressive behavior and the associated effects on 1 Manuscript received 11 August 2011 revision accepted 19 December 2011. The authors thank D. Hauber for his valuable comments on the manuscript, L. Achenbach for assistance with sampling in the Mississippi River delta, K. Saltonstall for coding the new haplotypes, and the curators of herbaria at Aarhus University, Aarhus, Denmark (AAU) and Louisiana State University, Baton Rouge, Louisiana, USA (LSU) for allowing sampling from museum specimens. Special thanks are due to colleagues and friends who sent Phragmites leaves, seeds, or rhizomes from various parts of the world. The project was funded by the Danish Council for Independent Research, Natural Sciences, project 10-083195, and in part by the Louisiana Sea Grant Program and its LaBorde Chair, awarded to Professor Hans Brix. The Louisiana Sea Grant Program is a component of the National Sea Grant Program maintained by the National Oceanic and Atmospheric Administration of the U.S. Department of Commerce. 4 Author for correspondence (email: [email protected]) doi:10.3732/ajb.1100396 TRACING THE ORIGIN OF GULF COAST PHRAGMITES (POACEAE): A STORY OF LONG-DISTANCE DISPERSAL AND HYBRIDIZATION 1 CARLA LAMBERTINI 2,4 , IRVING A. MENDELSSOHN 3 , MATS H. G. GUSTAFSSON 2 , BIRGIT OLESEN 2 , TENNA RIIS 2 , BRIAN K. SORRELL 2 , AND HANS BRIX 2 2 Department of Bioscience, Plant Biology, Aarhus University, Ole Worms Allé 1, 8000 Aarhus C, Denmark; and 3 Department of Oceanography and Coastal Sciences, School of the Coast and Environment, Louisiana State University, 3263 Energy, Coast and Environment Building, Baton Rouge, Louisiana 70803 USA • Premise of the study: Long-distance dispersal can affect speciation processes in two opposing ways. Dispersal can promote geographic isolation or it can bring together geographically distant and distantly related genotypes, thus counteracting local differentiation. We used the Gulf Coast of North America (GC), a “hot spot” of reed diversity and evolutionary dynamics, as a model system to study the diversification processes within the invasive, cosmopolitan, polyploid grass Phragmites. • Methods: Genetic diversity was studied using collections representing all species of the genus and from all continents (except Antarctica). A range of molecular markers, including chloroplast and nuclear sequences, microsatellites, and AFLPs, was ana- lyzed to detect DNA variation from the population to the species level and to infer phylogenetic relationships across continents. • Key results: An interspecific hybrid, Phragmites mauritianus × P. australis, and four P. australis cp-DNA haplotypes from Africa, Europe, and North America have been dispersed to the GC and interbreed with each other. • Conclusions: Long-distance dispersal and weak breeding barriers appear to be recurring phenomena, not only in the GC, but worldwide. We present data strongly suggesting that interspecific hybridization and introgression among different Phragmites species take place and appear to have contributed significantly to the diversification processes within the genus. Hence, the application of traditional species concepts within Phragmites might be inappropriate. Key words: Gene flow; Gulf Coast; hybridization; introgression; invasion; long-distance dispersal; Phragmites australis; Phragmites mauritianus
Transcript of Gulf Coast Phragmites

538
American Journal of Botany 99(3): 538–551. 2012.
American Journal of Botany 99(3): 538–551, 2012; http://www.amjbot.org/ © 2012 Botanical Society of America
Even though most biologists would agree that a species rep-resents a fundamental biological unit, many different species concepts have been suggested ( De Queiroz, 2007 ). Frequently mentioned in textbooks is the biological species concept (BSP) which implies that individuals capable of interbreeding and producing fertile offspring belong to the same species ( Mayr, 1942 ). While this defi nition generally works well for verte-brates, it may be diffi cult to apply to plants. One reason is poly-ploidization, a phenomenon common in plants but rare in animals. Polyploidization following hybridization, resulting in allopolyploidy, may re-establish disomic segregation, allowing hybrids to escape infertility.
The genus Phragmites is one of many plant genera where ap-plication of the species concept is challenging partly due to poly-ploidy and associated interfertility barriers. Phragmites is distributed from the tropics to the cold temperate areas of both hemispheres, and includes fi ve species, which are hardly distin-guishable morphologically and each showing cytological variation. Four of the species, P. frutescens H. Scholz, P. japonicus Steud., P. karka (Retz.) Trin. ex Steud., and P. mauritianus Kunth, have restricted geographical distributions ( Clevering and Lissner, 1999 ), whereas P. australis (Cav.) Trin. ex Steud. is widely distributed in temperate and subtropical areas. Phylogeographically, most P. aus-tralis genotypes belong to a poorly structured group, with largely reticulate interrelationships, and a distribution as wide as the genus as a whole ( Lambertini et al., 2006 ). The small number of species within the genus, the lack of diploids, and the occurrence of several ploidy levels are characteristics typical of old “ taxonomic com-plexes ” ( Stebbins, 1971 ). The lack of genetic structure within the species might indicate that there is suffi cient reproductive contact among populations within the complex to prevent differentiation and, ultimately, speciation. The reproductive contact can occur as cross-pollination between geographically distant parents, but also as a consequence of seed dispersal by wind and migratory birds, rafting of eroded plant fragments and marsh sods, as well as human-aided spread of seeds and rhizome fragments.
Dispersal of Phragmites by humans is known to have oc-curred repeatedly at various geographical scales. The spreading of a European P. australis lineage in North America is particu-larly well documented, and has been addressed primarily because of its aggressive behavior and the associated effects on
1 Manuscript received 11 August 2011 revision accepted 19 December 2011. The authors thank D. Hauber for his valuable comments on the
manuscript, L. Achenbach for assistance with sampling in the Mississippi River delta, K. Saltonstall for coding the new haplotypes, and the curators of herbaria at Aarhus University, Aarhus, Denmark (AAU) and Louisiana State University, Baton Rouge, Louisiana, USA (LSU) for allowing sampling from museum specimens. Special thanks are due to colleagues and friends who sent Phragmites leaves, seeds, or rhizomes from various parts of the world. The project was funded by the Danish Council for Independent Research, Natural Sciences, project 10-083195, and in part by the Louisiana Sea Grant Program and its LaBorde Chair, awarded to Professor Hans Brix. The Louisiana Sea Grant Program is a component of the National Sea Grant Program maintained by the National Oceanic and Atmospheric Administration of the U.S. Department of Commerce.
4 Author for correspondence (email: [email protected])
doi:10.3732/ajb.1100396
TRACING THE ORIGIN OF GULF COAST PHRAGMITES (POACEAE): A STORY OF LONG-DISTANCE DISPERSAL AND HYBRIDIZATION 1
CARLA LAMBERTINI 2,4 , IRVING A. MENDELSSOHN 3 , MATS H. G. GUSTAFSSON 2 , BIRGIT OLESEN 2 , TENNA RIIS 2 , BRIAN K. SORRELL 2 , AND HANS BRIX 2
2 Department of Bioscience, Plant Biology, Aarhus University, Ole Worms All é 1, 8000 Aarhus C, Denmark; and 3 Department of Oceanography and Coastal Sciences, School of the Coast and Environment, Louisiana State University, 3263 Energy,
Coast and Environment Building, Baton Rouge, Louisiana 70803 USA
• Premise of the study: Long-distance dispersal can affect speciation processes in two opposing ways. Dispersal can promote geographic isolation or it can bring together geographically distant and distantly related genotypes, thus counteracting local differentiation. We used the Gulf Coast of North America (GC), a “ hot spot ” of reed diversity and evolutionary dynamics, as a model system to study the diversifi cation processes within the invasive, cosmopolitan, polyploid grass Phragmites .
• Methods: Genetic diversity was studied using collections representing all species of the genus and from all continents (except Antarctica). A range of molecular markers, including chloroplast and nuclear sequences, microsatellites, and AFLPs, was ana-lyzed to detect DNA variation from the population to the species level and to infer phylogenetic relationships across continents.
• Key results: An interspecifi c hybrid, Phragmites mauritianus × P. australis, and four P. australis cp-DNA haplotypes from Africa, Europe, and North America have been dispersed to the GC and interbreed with each other.
• Conclusions: Long-distance dispersal and weak breeding barriers appear to be recurring phenomena, not only in the GC, but worldwide. We present data strongly suggesting that interspecifi c hybridization and introgression among different Phragmites species take place and appear to have contributed signifi cantly to the diversifi cation processes within the genus. Hence, the application of traditional species concepts within Phragmites might be inappropriate.
Key words: Gene fl ow; Gulf Coast; hybridization; introgression; invasion; long-distance dispersal; Phragmites australis ; Phragmites mauritianus

539March 2012] LAMBERTINI ET AL. — GULF COAST PHRAGMITES
molecular markers, to detect variation patterns from the popula-tion to the species level. Inclusion of both the maternally inherited chloroplast (cp-) sequences and biparentally inherited markers, like nuclear SSR (Simple Sequence Repeats or microsatellites) can potentially identify cases of hybridization between different lineages. In addition to the cp-sequences and nuclear microsatel-lites developed by Taberlet et al. (1991) and Saltonstall (2001 , 2002, 2003 ), we explored genetic variation in other regions of the chloroplast ( rpl 16 intron, Posno et al., 1986 ; Zhang, 2000 ) and of the nucleus (sequences and AFLPs) not associated with repetitive sequences. Repetitive sequences might not be optimal to address intercontinental relationships as their high mutation rates might increase the likelihood of homoplasy.
MATERIALS AND METHODS
Sampling — We randomly collected 157 samples of Phragmites australis along the GC of United States from Texas to Florida. The outer marshes of the Mississippi River delta were densely sampled by helicopter. Additional samples (46) were collected from museum specimens at Louisiana State University Her-barium and covered the period 1927-2006 (Appendix S1 – see Supplemental Data with the online version of this article).
The genetic diversity of the GC populations was compared with a global col-lection, which included Phragmites australis specimens from all continents (ex-cept Antarctica) and all Phragmites species. Genotypes to include in the study were selected with phylogeographic criteria from the living collection in the experimental fi elds at Aarhus University ( Lambertini et al., 2006 ). The number of samples was progressively increased in areas phylogenetically related to the GC specimens, and decreased from areas that consistently appeared distantly related. The fi nal sample sets and their use in the various analyses are shown in Appendix S1 (see Supplemental Data with the online version of this article).
DNA extraction — DNA was isolated with the E.Z.N.A. SP Plant DNA kit (Omega Bio-Tek, Inc., Norcross, Georgia, USA) from young leaves stored in silica gel, after grinding the plant material in a mortar with liquid nitrogen. The DNA concentration was measured with a Nanodrop spectrophotometer ND-1000 (Saveen & Werner, Limhamn, Sweden) and diluted to 10 ng/ µ l. Her-barium collections were treated as described in Lambertini et al. (2008) .
Microsatellites — Ten microsatellite primers were developed by Saltonstall ( 2003 ) to study the North American populations of Phragmites australis . Of these, we chose 6 primer sets (Pagt4, Pagt8, Pagt9, Pagt13, Pagt14, and Pagt22) based on their variation within the global sample set and their scoring properties.
Twenty ng DNA were added to 18 µ l mastermix consisting of 10 µ l 2xMaser-mix (VWR International, Arlington Heights, Illinois, USA), 10 pmol forward and reverse primers and sterile water to reach the fi nal volume of 20 µ l. Ampli-fi cation was run in a Peltier Thermal Cycler PTC-200-MJ Research under the following conditions: 94 ° C for 3 min, 40 cycles of 94 ° C for 30 s, annealing temperature for 40 s, 72 ° C for 40 s, followed by 72 ° C for 7 min. Annealing temperatures were: 50 ° C for primers Pagt8, Pagt9, Pagt13, and Pagt22; 54 ° C for Pagt4; and 62 ° C for Pagt14. Forward primers were fl uorochrome (cy) labeled.
The PCR product was diluted 20 × with sterile water prior to electrophoresis on a 7% acrylamide gel (Reprogel-Long Read, GE Healthcare, Piscataway, New Jersey, USA) in an ALF Express II DNA Analysis System (Amersham Pharmacia Biotech, Inc., Piscataway, New Jersey, USA). Five µ l of diluted PCR product were added to 3 µ l loading dye (GE Healthcare) and 1 µ l each of 100 and 300 bp internal sizers (GE Healthcare). DNA was denatured in the PTC-200 PCR at 94 ° C for 5 min prior to loading onto the gel. Electrophoresis conditions were 1500 V, 55 ° C, 120 min. The fi rst and the last lanes of the gel were loaded with 50-500 bp external sizers (GE Healthcare). The chromatograms were scored manually.
Ploidy level — The number of alleles ranged from 1 to 6 in some indi-viduals. Pagt22, Pagt9, and Pagt4 amplifi ed three alleles in a considerable number of GC samples. Pagt14 amplifi ed up to six alleles in some of the samples with three alleles at loci Pagt22, Pagt9, and Pagt4. Considering that we counted up to 70 chromosomes in one GC individual with three alleles (by root tip squash) and that the highest number of alleles that we found
native marsh ecosystems ( Chambers et al., 1999 ; Saltonstall, 2002, 2003 ; McCormick et al., 2010 ; Meyerson et al., 2010b ; Paul et al., 2010 ; Saltonstall, 2011 ). In addition to the intro-duced European lineage, two distantly related P. australis lineages occur in North America ( Saltonstall et al., 2004 ): 1) P. australis subsp. americanus Saltonstall, P. M. Peterson & Soreng, endemic to North America and found in much of Can-ada and in the United States; and 2) P. australis var. berlandieri (E. Fourn.) C. F. Reed, found in the southern United States along the Gulf Coast ( Saltonstall, 2002 ; Saltonstall et al., 2004 ) and in the Southwest ( Meyerson et al., 2010a ), as well as in Central America and Asia ( Saltonstall, 2002 ). The native and the European P. australis lineages are sympatric in the northern part of the United States and in Canada, but the European lin-eage has displaced most of the native stands in large parts of North America. It is believed that the native and the European P. australis lineages do not interbreed due to nonoverlapping fl owering times, the lack of intermediate morphological forms, as well as genetic evidence ( Saltonstall, 2003, 2011 ). However, recent experimental crossings have demonstrated that the two lineages can interbreed (Meyerson et al., 2010b ) and intermedi-ate genetic patterns have been found in Canadian populations ( Paul et al., 2010 ). However, direct evidence of recombination is still missing from the wild. While interbreeding between dis-tantly related lineages of P. australis has been reported and re-ferred to as “ hybridization, ” interspecifi c hybridization in wild populations has received much less attention. This is probably due to the fact that, apart from the widespread P. australis , all other species of the genus have restricted distributions far from Europe and North America, and are generally poorly known and taxonomically controversial.
The Gulf Coast of North America (GC) is one area where Phragmites australis is particularly diverse, with distantly re-lated genotypes coexisting, especially in the interior marshes of the Mississippi River delta ( Hauber et al., 1991 ; Pellegrin and Hauber, 1999 ; Saltonstall, 2002, 2003 ; White et al., 2004 ; Lambertini et al., 2006 ; Howard et al., 2008 ; Hauber et al., 2011 ). Hauber et al. (2011) reported that at least four chloro-plast DNA haplotypes are found in the delta. In addition, there is both morphological and cytological variation, with at least 2n = 4 x and 6 x represented ( Fournier et al., 1995 ; Chambers et al., 1999 ; Clevering and Lissner, 1999 ; Hauber et al., 2011 ). In a previous study, GC P. australis var. berlandieri was found to be closely related to the tropical African species P. mauritianus ( Lambertini et al., 2006 ). As phylogenetic relationships in Phragmites are still not fully understood, and the similarities with P. mauritianus imply relationships across continents with another species, we analyzed genetic diversity in the GC in the context of a large collection of reference samples representing all species of the genus and its global distribution. This ap-proach allows conclusions regarding relationships across taxa and long geographic distances. Another advantage of a global phylogeographic study is that the pattern of relationships pro-vides a tool to distinguish ancestral from newly evolved alleles, which might have been acquired by genetic recombination in a determined area or by mutations. It is in this perspective that we reconstruct the history of GC Phragmites and analyze gene fl ow in the GC area, with the ultimate goal to understand 1) the relationship between P. australis and P. mauritianus , 2) the im-plications of long-distance dispersal on gene fl ow at the popula-tion level, and 3) to what extent Phragmites taxa meet the criteria of the biological species concept. Given the broad taxonomic and geographic approach, we analyzed a range of

540 AMERICAN JOURNAL OF BOTANY [Vol. 99
Two µ l of template DNA were added to 10 µ l 2xMasermix (VWR International), 10 pmol forward and reverse primers, and sterile water to reach the fi nal volume of 20 µ l. PCR conditions were 94 ° C for 3 min, 40 cycles of 94 ° C for 30 s, annealing temperature for 40 s, 72 ° C for 1 min, followed by 72 ° C for 7 min. Annealing tem-peratures were: 50 ° C for trn T- trn L and rbc L- psa I and 62 ° C for rpl 16 and waxy .
The amplifi ed product was sent to Macrogen Inc. (Seoul, Korea) for Sanger sequencing. Most of the variation detected by trn T- trn L and rbc L- psa I sequences was due to varying numbers of repeats at microsatellite loci. To make sure that this variation was not due to PCR or sequencing artifacts, we reamplifi ed samples yielding sequences differing from those obtained from GenBank. Such samples were sequenced with both forward and re-verse primers.
Waxy bands — Besides the waxy gene itself amplifi ed by primers F-for and M-bac ( Mason-Gamer et al., 1998 ), additional products appeared in several PCR reactions. As the presence/absence of two particular bands (of about 100 and 200 bp) appeared as a distinctive trait among the GC samples, we isolated such bands with the E.Z.N.A. Gel Extraction kit (Q-spin column) and se-quenced them. Sequences were subjected to a BLAST search in GenBank, but no signifi cant match was found. The 100 and 200 bp sequences were neither alignable with each other nor with any part of the waxy gene sequence of about 1000 bp. Presence/absence of these two bands was assessed in the global sample set.
Sequences data analysis — Sequences were aligned using Bioedit ver. 7.0 Sequence Alignment Editor ( Hall, 1999 ). After the fi rst round of automatic alignment with the option “ clustal ” , we completed the alignment manually. We downloaded all Phragmites trn T- trn L and rbc L- psa I sequences from NBCI (Saltonstall, 2003 ; Vachon and Freeland, 2011 ; Hauber et al., 2011 ) and in-cluded them in our alignment. The variable parts of the sequences were care-fully checked on the chromatograms to make sure that the changes were real and not due to poor sequence quality.
The aligned matrices were analyzed in a median-joining network ( Bandelt et al., 1999 ) with the program Network ver. 4.5.1.6. Phylogenetic Network Constructions (Fluxus Technologies Ltd., Clare, Suffolk, UK). A separate net-work was run for each chloroplast region.
AFLPs — A subset of the GC genotypes (Appendix S1; see Supplemental Data with the online version of this article) was AFLP fi ngerprinted following the protocol described in Lambertini et al. (2006) . Several samples from our previous study were amplifi ed and used as a reference to align the new samples from the GC in the global AFLP binary matrix. The AFLPs were perfectly re-producible and it was possible to identify the peaks scored in the previous study. However, of the 128 fragments previously used, only 105 could be reliably scored in the new GC specimens. Lower transmittance in the detection equip-ment made some of the peaks appear under the detection limit (set with a peak shape of 10 and a minimum height of 1%), while still recognizable in the chro-matograms among many other small peaks. For this reason we preferred to ex-clude such fragments from the analysis. The results presented in Lambertini et al. (2006) did not change with the 105-fragment matrix, so we used this re-duced matrix for the statistical analyses.
AFLP data analysis — The AFLP data were used to analyze the position of the GC genotypes in the global phylogeographic pattern of Phragmites pre-sented in Lambertini et al. (2006) . To this end, a median-joining network ( Bandelt et al., 1999 ) was constructed using the program Network. The network combines minimum spanning trees ( Kruskal, 1956 ) with maximum parsimony ( Farris, 1970 ) and introduces hypothetical missing ancestral genotypes, repre-sented as median vectors (nodes).
The AFLP data set was also divided into the populations identifi ed by se-quences and microsatellites and analyzed with the population genetic program GenAlEx. We ran a Principal Coordinate Analysis (PCA) based on the Euclid-ean metrics generated by the genetic distance algorithm implemented for binary data ( Peakall and Smouse, 2006 ). The fi rst two axes explained 57% of the varia-tion and the third axis an additional 16%.
Population pairwise genetic identities were calculated with the program PopGen according to Nei ( Nei, 1978 ). Nei ́ s unbiased genetic identities are minimally affected by differences in sample size ( Gorman and Renzi, 1979 ) and unbiased by small sample sizes.
Nei ́ s Gene Diversity ( Nei, 1973 ) and Shannon Information Index ( Lewontin, 1972 ) were calculated with the program PopGen and were compared with those obtained with microsatellite binary data.
within GC individuals was six, we considered samples with three to six alleles as hexaploids ( x = 12 is the basic chromosome number in Phragmites , Gorenfl ot et al., 1972 ; Raicu et al., 1972 ). Hexaploids were found in Phrag-mites accessions of the GC also by Clevering and Lissner (1999) and Hauber et al. (2011) .
We found other specimens in the global sample with more than two alleles at some of the loci (three or four). Normally Phragmites tetraploids have two alleles because genome diploidization occurred after polyploidization ( Gorenfl ot et al., 1979 ; Gaudreault et al., 1989 ). We therefore considered sam-ples with three and four alleles as possible euploids higher than 2n = 4 x and entered them as hexaploids. 2n = 6 x is a compromise for cytologically unknown genomes, to cover all allelic variation and not overestimate allelic frequencies with octoploids or higher ploidy levels in the case of hexaploids or recent tetra-ploids that are still under genomic rearrangement ( Soltis and Soltis, 1993 ; Otto and Whitton, 2000 ).
Microsatellite data analysis — Considering the polysomic variation in the dataset, microsatellites data were initially translated into a binary matrix (1= presence, 0= absence of homologous alleles) and analyzed with Neighbor Joining (NJ) (PAUP* ver. 4.0.b10; Swofford, 1998 ). The GC genotypes were fi rst analyzed independently and subsequently together with the global sample set. One NJ tree was run for each SSR locus. The global NJ analysis identifi ed groups with identical microsatellites at several loci, which represented not only GC genotype groups, but included also individuals from other continents. As the intercontinental grouping of identical samples was consistent through-out the SSR data set, we tested the genetic relationships among the geo-graphical areas represented in each group with a population assignment test. The presence of polysomic populations is incompatible with microsatellite analysis programs, which are designed for codominant diploid organisms. Rather than excluding polysomies, represented by several samples and one entire GC group, we addressed the problem in two different ways. We entered hexaploids with three codominant markers in a diploid matrix where each hexaploid indi-vidual had three diploid entries, whereas disomic tetraploids had only one dip-loid entry. We used GenAlEx ver. 6.4 ( Peakall and Smouse, 2006 ) to run the frequency-based assignment test of Paetkau et al., (1995) . This test calculates the probability that an allele match among populations occurs by chance and assigns each genotype to the population with the lowest likelihood of a random match. DNA profi le probability is calculated as the log transformed product (multiplied − 1 times) of the genotype ́ s allelic frequencies across all loci in each of the investigated population ( Peakall and Smouse, 2006 ). We also calcu-lated genetic identities with the program PopDist ver.1.2.1 ( Guldbrandtsen et al., 2000 ) to correct errors in the estimation of allelic frequencies due to polyploid genotypes. Tomiuk and Loeschcke ́ s Identity Estimating Measure ( Tomiuk and Loeschcke, 1991 ) implemented in PopDist is the only algo-rithm that can be used to compare allelic frequencies among populations of different ploidy levels. For this analysis disomic and polysomic allelic patterns were considered as independent populations, even within the same geographi-cal area. Standard error was calculated with jack-knife resampling (mean of individual jack-knife estimates) ( Guldbrandtsen et al., 2009 ). Similar results obtained from population assignment test and Tomiuk and Loeschcke ́ s Identity Estimating Measure, as well as from jack-knife analysis, indirectly supported the results of the population assignment test.
The “ 3-entry ” matrix covers all allelic variation but is not representative of the allelic combinations that might be segregated into gametes by hexaploid individuals and thus might contribute to gene fl ow. To evaluate gene fl ow pos-sibilities, we calculated allelic frequencies with the “ 3-entry ” matrix (with GenAlEx) and obtained an overview of all alleles present (and absent) in the GC and in the global sample set. We used the presence/absence dataset to dis-tinguish possibly plesio morphic alleles present in the GC gene pool and its phylogenetically related populations from novel alleles, which have likely evolved in the GC (Appendix S2).
Gene Diversity ( Nei, 1973 ) and Shannon Information Index ( Lewontin, 1972 ) were calculated with the program PopGene ver. 1.32 Population Genetic Analysis ( Yeh and Boyle, 1997 ) to compare genetic diversity levels between phylogenetically related populations. For this analysis we used the binary ma-trix of SSR data because results were directly compared to those obtained from the AFLP dataset.
Sequences — The rpl 16 chloroplast intron ( Posno et al., 1986 ; Zhang, 2000 ), trn T- trn L chloroplast intergenic spacer ( Taberlet et al., 1991 ; Saltonstall, 2001 ), rbc L- psa I chloroplast intergenic spacer ( Saltonstall, 2001 and 2002 ), and the Grass- waxy nuclear gene ( Mason-Gamer et al., 1998 ) were amplifi ed.
541March 2012] LAMBERTINI ET AL. — GULF COAST PHRAGMITES
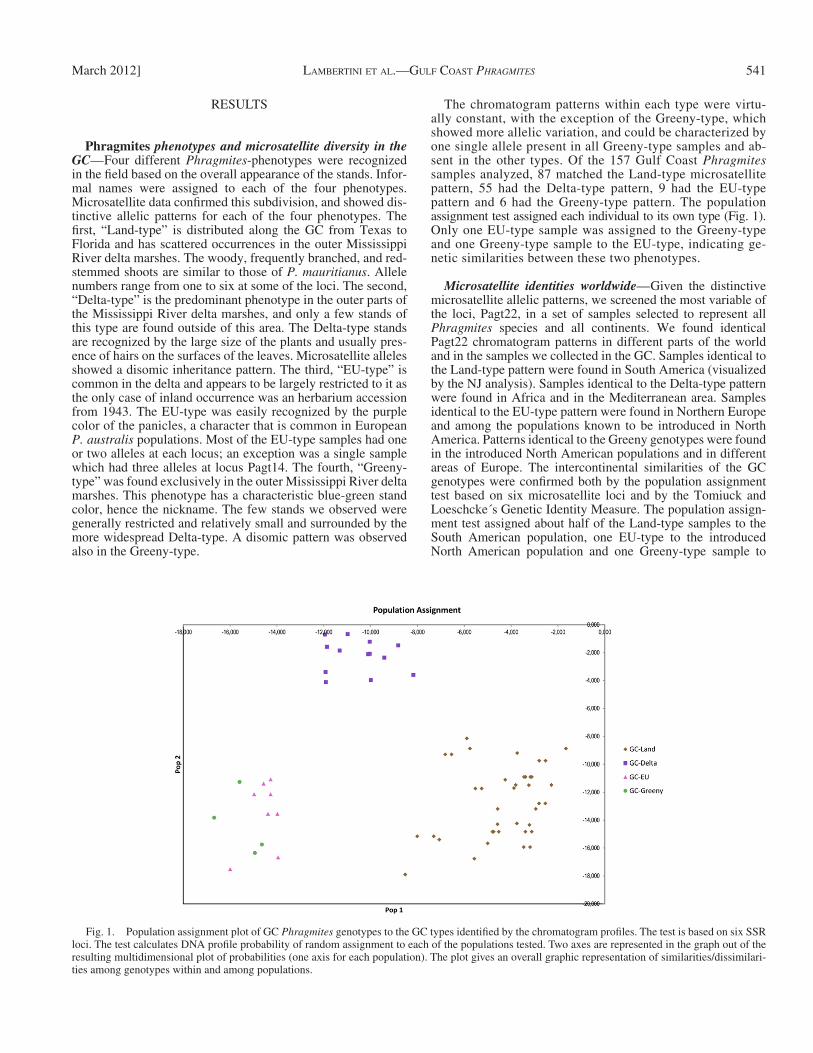
The chromatogram patterns within each type were virtu-ally constant, with the exception of the Greeny-type, which showed more allelic variation, and could be characterized by one single allele present in all Greeny-type samples and ab-sent in the other types. Of the 157 Gulf Coast Phragmites samples analyzed, 87 matched the Land-type microsatellite pattern, 55 had the Delta-type pattern, 9 had the EU-type pattern and 6 had the Greeny-type pattern. The population assignment test assigned each individual to its own type ( Fig. 1 ). Only one EU-type sample was assigned to the Greeny-type and one Greeny-type sample to the EU-type, indicating ge-netic similarities between these two phenotypes.
Microsatellite identities worldwide — Given the distinctive microsatellite allelic patterns, we screened the most variable of the loci, Pagt22, in a set of samples selected to represent all Phragmites species and all continents. We found identical Pagt22 chromatogram patterns in different parts of the world and in the samples we collected in the GC. Samples identical to the Land-type pattern were found in South America (visualized by the NJ analysis). Samples identical to the Delta-type pattern were found in Africa and in the Mediterranean area. Samples identical to the EU-type pattern were found in Northern Europe and among the populations known to be introduced in North America. Patterns identical to the Greeny genotypes were found in the introduced North American populations and in different areas of Europe. The intercontinental similarities of the GC genotypes were confi rmed both by the population assignment test based on six microsatellite loci and by the Tomiuck and Loeschcke ́ s Genetic Identity Measure. The population assign-ment test assigned about half of the Land-type samples to the South American population, one EU-type to the introduced North American population and one Greeny-type sample to
RESULTS
Phragmites phenotypes and microsatellite diversity in the GC — Four different Phragmites -phenotypes were recognized in the fi eld based on the overall appearance of the stands. Infor-mal names were assigned to each of the four phenotypes. Microsatellite data confi rmed this subdivision, and showed dis-tinctive allelic patterns for each of the four phenotypes. The fi rst, “ Land-type ” is distributed along the GC from Texas to Florida and has scattered occurrences in the outer Mississippi River delta marshes. The woody, frequently branched, and red-stemmed shoots are similar to those of P. mauritianus . Allele numbers range from one to six at some of the loci. The second, “ Delta-type ” is the predominant phenotype in the outer parts of the Mississippi River delta marshes, and only a few stands of this type are found outside of this area. The Delta-type stands are recognized by the large size of the plants and usually pres-ence of hairs on the surfaces of the leaves. Microsatellite alleles showed a disomic inheritance pattern. The third, “ EU-type ” is common in the delta and appears to be largely restricted to it as the only case of inland occurrence was an herbarium accession from 1943. The EU-type was easily recognized by the purple color of the panicles, a character that is common in European P. australis populations. Most of the EU-type samples had one or two alleles at each locus; an exception was a single sample which had three alleles at locus Pagt14. The fourth, “ Greeny-type ” was found exclusively in the outer Mississippi River delta marshes. This phenotype has a characteristic blue-green stand color, hence the nickname. The few stands we observed were generally restricted and relatively small and surrounded by the more widespread Delta-type. A disomic pattern was observed also in the Greeny-type.
Fig. 1. Population assignment plot of GC Phragmites genotypes to the GC types identifi ed by the chromatogram profi les. The test is based on six SSR loci. The test calculates DNA profi le probability of random assignment to each of the populations tested. Two axes are represented in the graph out of the resulting multidimensional plot of probabilities (one axis for each population). The plot gives an overall graphic representation of similarities/dissimilari-ties among genotypes within and among populations.
542 AMERICAN JOURNAL OF BOTANY [Vol. 99
P. mauritianus samples from Uganda and Burkina Faso and in P. frutescens ( Table 2 ).
The EU-type sequences were identical to Haplotype 4 (Saltonstall, NCBI accession no. AY016327), to the intro-duced North American and the European Phragmites austra-lis populations, as well as to part of the African/Mediterranean samples.
The Delta-type sequences were identical to Haplotype 4a (Hauber et al., NCBI accession no. JF271678) and to those of Phragmites australis samples from Senegal, North Africa, and the Mediterranean area. The trn T- trn L sequences revealed three cpDNA haplotypes within the Greeny-type. The Greeny1-type sequences were identical to the EU-type (Haplotype 4). The Greeny2-type sequences were identical to Haplotype 19 (Hauber et al., NCBI accession no. accession no. JF271679) and ap-peared as intermediate between the European Haplotype 4 and the native North American Haplotype 9 (Saltonstall, NCBI accession no. AF457396), with which it shared an 8 bp indel. The indel was otherwise present in other North American haplo-types (Haplotype 17, Saltonstall NCBI accession no. AY714215; Haplotype 14, Saltonstall NCBI accession no. AF457400; Hap-lotype 2, Saltonstall NCBI accession no. AY016325; Haplo-type E1, Vachon and Freeland NCBI accession no. GQ468791; Haplotype E3, Vachon and Freeland NCBI accession no. GQ468792; Haplotype E4, Vachon and Freeland NCBI acces-sion no. GQ468793; Haplotype M2, Vachon and Freeland NCBI accession no. GQ468794), in all samples of the native North American population and in one P. mauritianus sample from Zambia. Considering the indel as a single mutation event, the Greeny2-type is more similar to the European Haplotype 4 (one change) than to the North American native Haplotype 9 (three changes). The Greeny3-type sequence matched Hap-lotype 3 (Saltonstall, NCBI accession no. AY016326). Like
EU-type. About half of the samples of the introduced North American population were assigned to the GC EU-type and one to the Greeny-type. Four South American samples were as-signed to the GC Land-type and one European sample to the GC Greeny-type ( Fig.2 ). Tomiuk and Loeschcke ́ s population pairwise genetic identities confi rmed that the Land – type micro-satellite pool was more similar to the South American population than to any other population. The Delta-type appeared most similar to the African/Mediterranean population, the EU-type more similar to the introduced North American population than to the European population, whereas the Greeny-type was more similar to the European population ( Table 1 ).
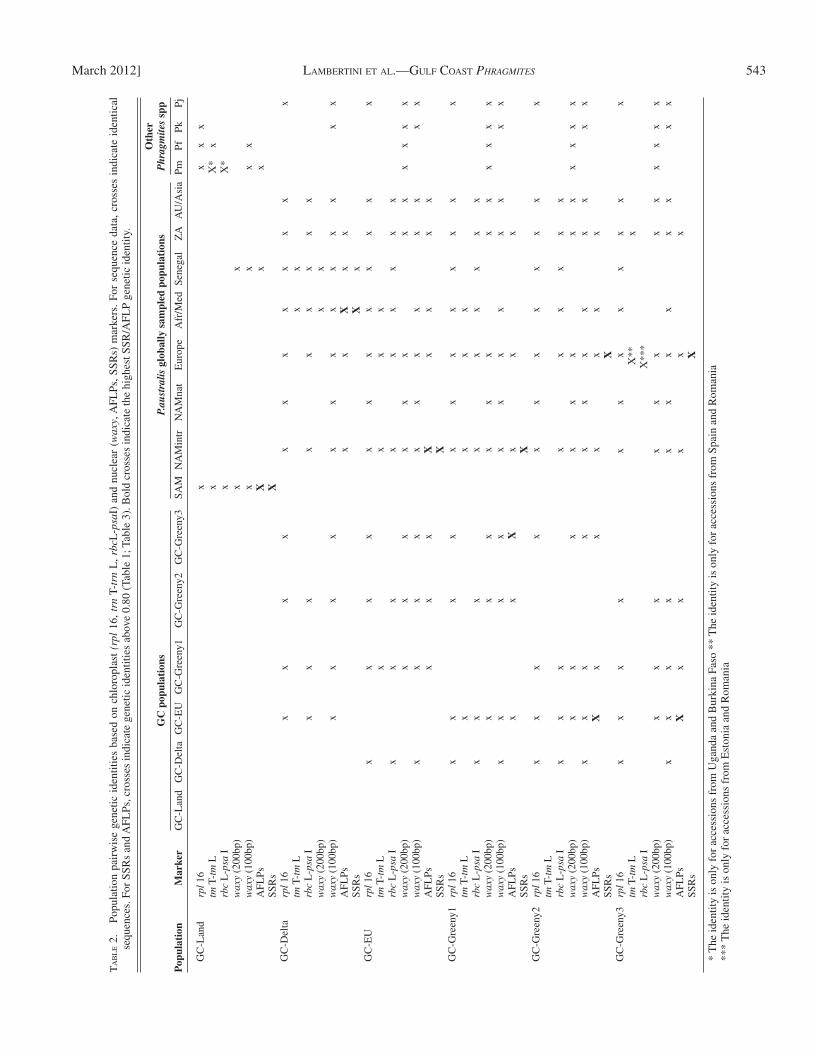
Intercontinental relationships revealed by cpDNA se-quences — rpl16 — An indel divided the Phragmites samples into two groups. Absence of the mutation was shared by Phragmites australis Land-type (Lambertini et al., NCBI accession no. JN565034), P. australis from South America and the species P. mauritianus (samples from tropical Africa), P. frutescens (samples from Greece) and P. karka (samples from tropical Asia and North Australia). Presence of the mutation (Lambertini et al., accession no. JN565035) was seen in Delta-, EU-, Greeny-type, North American intro-duced and native populations, European, African/Mediterra-nean, Australian and Asiatic populations of P. australis, and in P. japonicus ( Table 2 ).
trnT-trnL — The Land-type sequences (Lambertini et al., NCBI accession no. HQ664450) were almost identical to hap-lotype 5 (Saltonstall, NCBI accession no. AY016328). They differed only in the number of repeats at one microsatellite lo-cus. Sequences identical to the Land-type were found in the South American Phragmites australis samples, as well as in
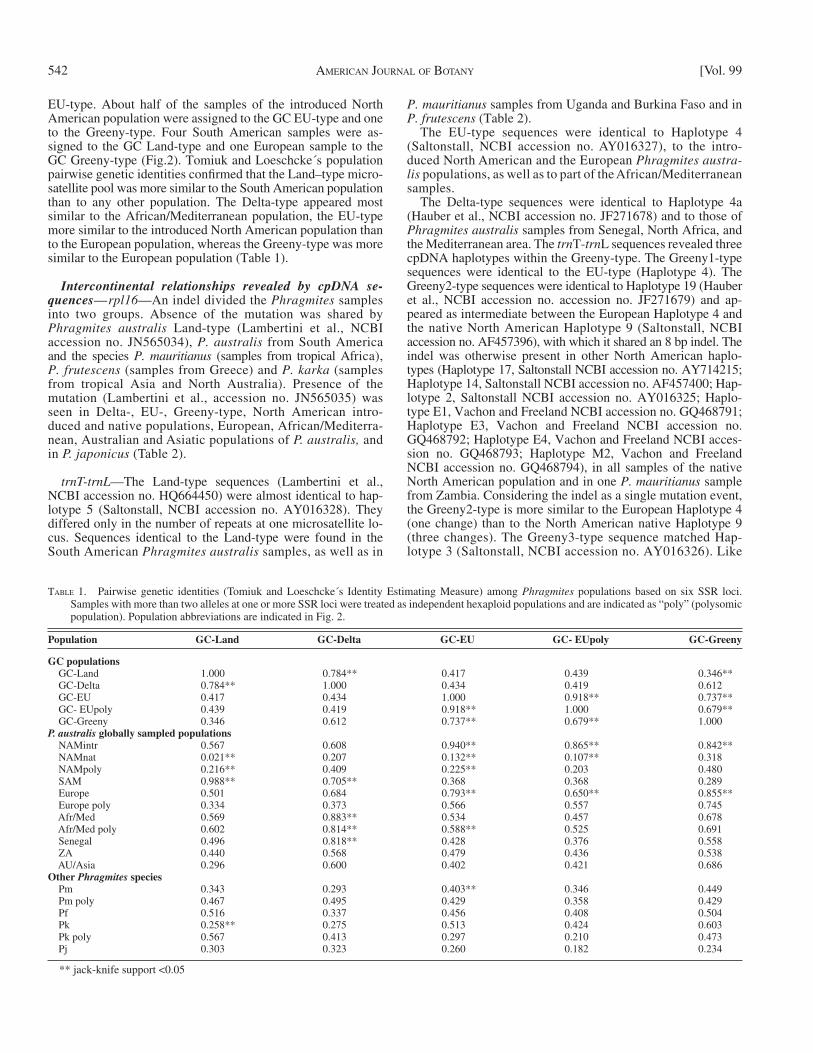
TABLE 1. Pairwise genetic identities (Tomiuk and Loeschcke ́ s Identity Estimating Measure) among Phragmites populations based on six SSR loci. Samples with more than two alleles at one or more SSR loci were treated as independent hexaploid populations and are indicated as “ poly ” (polysomic population). Population abbreviations are indicated in Fig. 2.
Population GC-Land GC-Delta GC-EU GC- EUpoly GC-Greeny
GC populations GC-Land 1.000 0.784** 0.417 0.439 0.346** GC-Delta 0.784** 1.000 0.434 0.419 0.612 GC-EU 0.417 0.434 1.000 0.918** 0.737** GC- EUpoly 0.439 0.419 0.918** 1.000 0.679** GC-Greeny 0.346 0.612 0.737** 0.679** 1.000 P. australis globally sampled populations NAMintr 0.567 0.608 0.940** 0.865** 0.842** NAMnat 0.021** 0.207 0.132** 0.107** 0.318 NAMpoly 0.216** 0.409 0.225** 0.203 0.480 SAM 0.988** 0.705** 0.368 0.368 0.289 Europe 0.501 0.684 0.793** 0.650** 0.855** Europe poly 0.334 0.373 0.566 0.557 0.745 Afr/Med 0.569 0.883** 0.534 0.457 0.678 Afr/Med poly 0.602 0.814** 0.588** 0.525 0.691 Senegal 0.496 0.818** 0.428 0.376 0.558 ZA 0.440 0.568 0.479 0.436 0.538 AU/Asia 0.296 0.600 0.402 0.421 0.686 Other Phragmites species Pm 0.343 0.293 0.403** 0.346 0.449 Pm poly 0.467 0.495 0.429 0.358 0.429 Pf 0.516 0.337 0.456 0.408 0.504 Pk 0.258** 0.275 0.513 0.424 0.603 Pk poly 0.567 0.413 0.297 0.210 0.473 Pj 0.303 0.323 0.260 0.182 0.234
** jack-knife support < 0.05
543March 2012] LAMBERTINI ET AL. — GULF COAST PHRAGMITES
TA
BL
E 2
. Po
pula
tion
pair
wis
e ge
netic
ide
ntiti
es b
ased
on
chlo
ropl
ast
(rpl
16,
trn
T- t
rn L
, rb
c L- p
sa I)
and
nuc
lear
( w
axy ,
AFL
Ps,
SSR
s) m
arke
rs.
For
sequ
ence
dat
a, c
ross
es i
ndic
ate
iden
tical
se
quen
ces.
For
SSR
s an
d A
FLPs
, cro
sses
indi
cate
gen
etic
iden
titie
s ab
ove
0.80
( Ta
ble
1 ; T
able
3 ).
Bol
d cr
osse
s in
dica
te th
e hi
ghes
t SSR
/AFL
P ge
netic
iden
tity.
Pop
ulat
ion
Mar
ker
GC
pop
ulat
ions
P.
aust
ralis
glo
bally
sam
pled
pop
ulat
ions
O
ther
P
hrag
mite
s sp
p
GC
-Lan
dG
C-D
elta
GC
-EU
GC
-Gre
eny1
GC
-Gre
eny2
GC
-Gre
eny3
SAM
NA
Min
trN
AM
nat
Eur
ope
Afr
/Med
Sene
gal
ZA
AU
/Asi
aPm
PfPk
Pj
G
C-L
and
rpl 1
6
x
xx
x tm
T- t
m L
xX
*x
rbc
L- p
sa I
xX
* w
axy
(200
bp)
xx
wax
y (1
00bp
)x
xx
xA
FLPs
X
xx
SSR
s X
GC
-Del
ta rp
l 16
xx
xx
xx
xx
xx
xx
tm T
- tm
Lx
x rb
c L
- psa
Ix
xx
xx
xx
xx
wax
y (2
00bp
)x
x w
axy
(100
bp)
xx
xx
xx
xx
xx
xx
xA
FLPs
xx
X
xx
SSR
s X
x
G
C-E
U rp
l 16
xx
xx
xx
xx
xx
xx
tm T
- tm
Lx
xx
x rb
c L
- psa
Ix
xx
xx
xx
xx
wax
y (2
00bp
)x
xx
xx
xx
xx
xx
x w
axy
(100
bp)
xx
xx
xx
xx
xx
xx
AFL
Psx
xx
X
xx
xx
SSR
s X
GC
-Gre
eny1
rpl 1
6x
xx
xx
xx
xx
xx
x tm
T- t
m L
xx
xx
rbc
L- p
sa I
xx
xx
xx
xx
x w
axy
(200
bp)
xx
xx
xx
xx
xx
xx
wax
y (1
00bp
)x
xx
xx
xx
xx
xx
xA
FLPs
xx
X
xx
xSS
Rs
X
G
C-G
reen
y2 rp
l 16
xx
xx
xx
xx
xx
xx
tm T
- tm
L rb
c L
- psa
Ix
xx
xx
xx
xx
wax
y (2
00bp
)x
xx
xx
xx
xx
xx
x w
axy
(100
bp)
xx
xx
xx
xx
xx
xx
AFL
Ps X
x
xx
xx
xSS
Rs
X
G
C-G
reen
y3 rp
l 16
xx
xx
xx
xx
xx
xx
tm T
- tm
LX
**x
rbc
L- p
sa I
X**
* w
axy
(200
bp)
xx
xx
xx
xx
xx
xx
wax
y (1
00bp
)x
xx
xx
xx
xx
xx
xA
FLPs
X
xx
xx
xSS
Rs
X
* T
he id
entit
y is
onl
y fo
r ac
cess
ions
fro
m U
gand
a an
d B
urki
na F
aso
** T
he id
entit
y is
onl
y fo
r ac
cess
ions
fro
m S
pain
and
Rom
ania
***
The
iden
tity
is o
nly
for
acce
ssio
ns f
rom
Est
onia
and
Rom
ania

544 AMERICAN JOURNAL OF BOTANY [Vol. 99
Fig. 2. Population assignment plot of GC and globally sampled genotypes to Phragmites populations worldwide, identifi ed by chromatogram profi les and NJ analysis. The test is based on 6 SSR loci. Abbreviations: GC-Land : Land-type; GC-Delta : Delta-type; GC-EU : EU-type; GC-Greeny : Greeny types (Greeny1, Greeny2, Greeny3); NAMintr : P. australis introduced North American population; NAMnat : P. australis native North American popula-tion; SAM : P. australis South American population; Europe : P. australis European population; Afr/Med : P. australis North African and Mediterranean population; Senegal : P. australis Senegal population; ZA : P. australis South African population; AU/Asia : P. australis Australian and Asian population; Pm : P. mauritianus ; Pf : P. frutescens ; Pk : P. karka ; Pj : P. japonicus .
Saltonstall (2002) , we found this haplotype in African P. aus-tralis accessions from South Africa and Namibia, but we also found it in European samples from Romania and Spain.
rbcL-psaI — The Land-type sequences were identical to Hap-lotype 3 (Saltonstall, NCBI accession no. AY016334), to the South American Phragmites australis samples and to the P. mauritianus accessions from Uganda and Burkina Faso. The Delta-, EU-, Greeny1- and Greeny2-type sequences were identical to Haplotype 4 (Saltonstall, NCBI accession no. AY0116335) and to P. australis samples from the introduced North American population, Europe, Africa (including Senegal, South Africa, and Namibia), and the Mediterranean area. The Greeny 3-type sequences (Lambertini et al., NCBI accession nr. HQ664451) were identical to European P. australis samples from Romania and Estonia ( Table 2 ).
Relationships with Phragmites mauritianus — Chloroplast DNA sequences revealed that the Phragmites australis Land-type is closely related to P. mauritianus , if, in fact, it is not P. mauritianus itself. The Delta-, EU-, and Greeny-types appear instead to share P. australis cpDNA haplotypes. Microsatellite alleles suggested that the Delta-type might be a hybrid between the Land-type and the EU-type. The Delta-type shares alleles with the Land-type (e. g., locus Pagt22, allele 187) and with the EU-type (allele 202), and gene fl ow is not contradicted at any of the microsatellite loci (online Appendix S2). The Delta-type ap-pears therefore to be a P. australis × P. mauritinaus interspecifi c hybrid. Interestingly, cpDNA sequences ( Table 2 ) and micro-satellites ( Table 1 ) indicated that the closest relatives of the Delta-type are in Africa, where the two Phragmites species co-exist, and in the Mediterranean area, where P. australis is
sympatric with P. frutescens . In a previous study ( Lambertini et al., 2006 ) P. frutescens appeared to be nested within P. mauri-tianus . P. frutescens differed from the Land-type in just a few changes in the rbc L- psa I haplotype ( Table 2 ). We therefore searched for evidence of hybridization in more conserved markers than microsatellites.
Nuclear sequences — The waxy gene primers amplifi ed a fragment of about 200 bp ( Fig.3 ) in addition to the waxy region itself. This fragment was amplifi ed only in Delta- and Land-types and was 99.9% homologous in the two types. The frag-ment was present in all Delta- and Land-type samples and absent in all EU- and Greeny-type samples. Amplifi cation of the waxy gene in a subset of our globally sampled Phragmites collection showed the presence of yet another fragment of ca. 100 bp in some of the samples. This fragment invariably oc-curred together with the 200 bp fragment in the Land-type, in South American samples, and in samples from Senegal ( Fig. 3 , Table 2 ). Samples of P. mauritianus and P. frutescens only had the 100 bp fragment. Samples of P. australis from North Africa and the Mediterranean area and the Delta-type only had the 200 bp fragment. None of the fragments occurred in any other sam-ples ( Table 2 ). The homology of the 100 bp fragment was 100% in P. mauritianus , Senegal, South American, and Land-type samples (Lambertini et al., NCBI accession no. JF317301). The homology of the 200 bp fragment was 100% in Delta-type, North African, Mediterranean, and in some of the Senegal sam-ples (Lambertini et al., NCBI accession no. JF317300). The two substitutions in the 200 bp fragment that distinguished Land- from Delta-type, were common to the Land-type, South Ameri-can and part of the Senegal samples (Lambertini et al., NCBI accession no. JF317299). The two variants of the same 200 bp

545March 2012] LAMBERTINI ET AL. — GULF COAST PHRAGMITES
Africa and Europe ( Fig. 4 ). The Land-type, together with the South American and Senegal populations, had intermediate distances from the African/European P. australis group and P. mauritianus and P. frutescens populations. The Senegal pop-ulation, which shared the cpDNA haplotype with P. australis and had the waxy hybrid fi ngerprint, appeared closer to P. australis than to P. mauritianus / P. frutescens . The GC Land-type and the South American population, which also had the double waxy band, but P. mauritianus cpDNA haplotype, were closer to P. mauritianus / P. frutescens .
The unbiased genetic identities of Land-type AFLPs were highest with the Phragmites australis South American popula-tion (0.90) followed by P. mauritianus (0.86) and the P. austra-lis Senegal population (0.83). Genetic identities with the P. frutescens population (0.76) were lower than with the P. aus-tralis African/Mediterranean and the South African populations
sequence indicate that Delta-type is not directly related to Land-type in this DNA fragment. Rather, Delta- and Land-type have independent origins. Surprisingly, the interspecifi c hybrid ap-peared not to be Delta-type as suspected, but the Land-type, which like the South American and Senegal populations carried the double waxy band, with the 100 bp fragment inherited from P. mauritianus / P. frutescens and the 200 bp fragment from the North African/Mediterranean P. australis population ( Fig. 3 ).
AFLPs — A global AFLP data set generated as part of a pre-vious study ( Lambertini et al., 2006 ) provided the possibility to test the robustness of the results with an independent sample set and a different source of nuclear DNA. The Principal Coordi-nates Analysis (PCA) of population pairwise differences showed that the Delta, EU, and the three Greeny cpDNA haplo-types were closest to the Phragmites australis populations from
Fig. 3. Agarose gel of waxy sequence pattern in the GC. 1-3 Delta-type samples; 4-8 Greeny (1-2-3)-type samples; 9-12 EU-type samples; 13-16 Land-type samples. Waxy gene is the fragment present in all samples about 1000 bp long. The additional fragment of 200 bp ( waxy 200) is common to Delta- and Land-types. The 100 bp fragment ( waxy 100) is present only in Land-type.
Fig. 4. PCA plot of pairwise Euclidean distances among Phragmites populations worldwide based on AFLPs. Senegal, GC Land-type, and SAM popu-lations appear as intermediate between P. australis populations (NAMintr , Europe, GC-EU, GC-Greeny1-2-3, GC-Delta, Afr/Med and ZA), P. mauritianus (Pm), and P. frutescens (Pf). Red bands represent the 100 bp waxy fragment; blue bands represent the 200 bp waxy fragment. These DNA fragments are present only in the indicated populations.

546 AMERICAN JOURNAL OF BOTANY [Vol. 99
Origins of the GC-types — The AFLP dataset ( Lambertini et al., 2006 ) was used to place the GC genotypes in a wider phylogeographic context ( Fig. 5 ). Phragmites is a taxonomi-cally isolated genus, and even the closest relatives are very distant. Hence, there is a difference in magnitude between intra- and intergeneric branch lengths. Any marker that is in-formative within the genus is unalignable outside of the ge-nus. Correspondingly, markers providing information about intergeneric relationships are invariable, or nearly so, within Phragmites ( Lambertini et al., 2006 ). For this reason, the rooting point (basalmost node) in the Phragmites genus is still unknown. The direction of migration, and indirectly, evolu-tion can be, however, assessed by comparing levels of diver-sity within Phragmites populations. Populations from source areas are expected to have higher genetic diversity than intro-duced populations derived from them, which typically would have originated from a limited number of founders. There-fore, to trace the directions of migration of the GC reed types in the unrooted AFLP phylogeny ( Fig. 5 ) we compared ge-netic diversity levels within the GC-types and their closely related populations. The AFLP and microsatellite datasets yielded matching results ( Table 4 ). The lower levels of ge-netic diversity within each GC-type compared to its closest relatives suggest that the Delta-type was introduced to the GC from the African/Mediterranean population, that the EU-type was recruited from the North American introduced population and that the three Greeny-types originated either from the North American introduced population and/or from Europe directly. Inconsistent results were obtained for the Land-type and its South American and African relatives ( P. mauritianus , Senegal, and African/Mediterranean P. australis populations). Due to the limited number of samples from South America and tropical Africa, it was not possible to determine migration routes among these populations based on genetic diversity measurements.
The AFLP median joining network indicated that the Delta-, EU-, and the three Greeny-types of the GC have an origin within Phragmites australis . For the Land-type, the AFLP network suggested an origin in South America where AFLP diversity is
(0.79) and of the same order of magnitude as with the European and introduced North American populations ( Table 3 ) .
Of the 105 AFLP fragments entered in the AFLP binary ma-trix, the Land-type shared 59 fragments with both Phragmites mauritianus and the African/Mediterranean P. australis sam-ples. Eleven fragments were present only in Land-type and in the African/Mediterranean samples and absent from P. mauri-tianus, and eight fragments were shared by Land-type and P. mauritianus and absent from the African/Mediterranean acces-sions of P. australis . The remaining 27 fragments, present in the African/ Mediterranean and/or P. mauritianus samples, were absent in Land-type samples. The number of shared frag-ments increased to 65 when the Land-type was compared with the Senegal P. australis samples, while the number of alleles exclusive to P. australis or P. mauritianus shared by the Land-type with the Senegal samples decreased to 8 and 6 respectively, indicating different degrees of introgression between the two hybrids following hybridization.
Intercontinental relationships revealed by nuclear mark-ers — The population pairwise genetic identities of the GC-types based on AFLP and microsatellite data generally showed the same relationships ( Table 1 , Table 3 ). Only in the case of the three Greeny-types, the AFLP data indicated additional rela-tionships, not evident in the microsatellite data. The three Greeny-types showed higher AFLP similarities with each other and with the EU-type than was revealed by microsatellite iden-tities ( Table 2 , Table 3 ). The highest microsatellite identities of the Greeny1-type were with the introduced North American population (0.830), whereas Greeny2- and Greeny3-types were most similar to the European population (0.811 and 0.790, re-spectively). AFLPs confi rmed microsatellite relationships of the three Greeny-types, but showed similarly high identities with the introduced North American and European populations, as well as with the South African population ( Table 3 ). The genetic identities with the native North American population were much lower, both in the microsatellite (0.248 for Greeny1-type; 0.102 for Greeny2-type; 0.208 for Greeny3-type) and AFLP data ( Table 3 ).
TABLE 3. Pairwise genetic identities (Nei ́ s Unbiased Genetic Identity) among Phragmites populations worldwide based on AFLPs.
Populations GC-Land GC-Delta GC-EU GC-Greeny1 GC-Greeny2 GC-Greeny3
GC populations GC-Land 1.0000 0.7387 0.7254 0.6891 0.7682 0.6468 GC-Delta 0.7387 1.0000 0.7788 0.7424 0.7550 0.7309 GC-EU 0.7254 0.7788 1.0000 0.8649 0.8819 0.9076 GC-Greeny1 0.6891 0.7424 0.8649 1.0000 0.8447 0.8911 GC-Greeny2 0.7682 0.7550 0.8819 0.8447 1.0000 0.8139 GC-Greeny3 0.6468 0.7309 0.9076 0.8911 0.8139 1.0000 P. australis globally sampled populations NAMintr 0.7508 0.8387 0.9414 0.8584 0.8488 0.8820 NAMnat 0.6290 0.7032 0.6756 0.6365 0.6244 0.6205 SAM 0.8950 0.6973 0.7254 0.7026 0.7385 0.6588 Europe 0.7646 0.8306 0.9299 0.8681 0.8708 0.8667 Afr/Med 0.7882 0.9127 0.8554 0.7891 0.8210 0.7774 Senegal 0.8306 0.9095 0.7839 0.7663 0.7911 0.7247 ZA 0.7892 0.8386 0.8780 0.8748 0.8505 0.8655 AU/Asia 0.7051 0.7604 0.8177 0.7570 0.7789 0.7462 Other Phragmites species Pm 0.8620 0.6808 0.7411 0.7160 0.6960 0.6530 Pf 0.7594 0.5857 0.6927 0.6667 0.6412 0.6246 Pk 0.7280 0.6649 0.6998 0.6826 0.6720 0.6233 Pj 0.7176 0.7353 0.7553 0.7304 0.6916 0.7305

547March 2012] LAMBERTINI ET AL. — GULF COAST PHRAGMITES
is inconsistent with the high genetic identities between these two populations shown both by microsatellites (0.89) and AFLPs (0.91) ( Tables 1 and 3 ).
Gene fl ow within the GC gene pool revealed by genetic interrelationships — To explain the confl icting result for the Delta-type revealed by the AFLP network vs. the AFLP/ microsatellites genetic identities, we focused on the small portion of alleles that the GC Delta genotypes do not share with their African/Mediterranean relatives. No AFLP alleles in this category were found, whereas a few microsatellite al-leles were present in the Delta-type while absent in the Afri-can/Mediterranean accessions. An example is allele no. 179 at locus Pagt14 (Appendix S2), which was absent from all Af-rican accessions, while present in the Delta-, Land-type, and in the South American population. Allele no. 179 occurs in the closely related Land-type and the South American popula-tion. Its presence in the Delta-type is most easily explained by recombination with the Land-type. Alternative cases of gene fl ow among the GC-types include alleles common to Land-, Greeny-, and EU-types. Allele no. 184 at locus Pagt 22 and allele no. 192 at locus Pag14 were absent in all South American and African samples, but present in the Land-, EU-, and Greeny-types, as well as in the introduced North Ameri-can populations and in the European samples (Appendix S2).
higher ( Fig. 5 ). Strikingly, this analysis introduced two ances-tral nodes for the South American group, one in common with a P. mauritianus sample from Uganda (in agreement with cpDNA sequences) and one with a P. australis sample from Senegal (in agreement with waxy sequences).
Evolutionary rates in the GC — Although the global median joining network based on AFLPs is not an optimal method to determine the absolute evolutionary rates in the GC popula-tions, the network gives an idea of the extent of evolutionary divergence that has occurred in the GC, and also an estimate of the time of establishment of the GC-types ( Fig. 5 ). The intro-duced North American genotypes (light blue nodes), which ap-parently established themselves in North America during the last 200 yr ( Chambers et al., 1999 ) can serve as a time refer-ence. The Land-type appears to be a well-defi ned group with moderate variation, indicating a long independent history. In contrast, the Delta-, EU-, and the three Greeny-types fall largely within the broader Phragmites australis pool compared with more differentiated groups, such as the Australian/Asiatic group (red branch) and the native North American group (black branch). This suggests a relative recent introduction of these Phragmites types to the GC. In spite of the short history in the GC, the network shows a remarkable isolation of the Delta-type from the African/Mediterranean genotypes of P. australis . This
Fig. 5. Global median joining network of all Phragmites genotypes (GC and global sampling) based on AFLPs. The global phylogeographic pattern of interrelationships was analyzed in Lambertini et al. (2006) .
548 AMERICAN JOURNAL OF BOTANY [Vol. 99
and taxonomic sampling is necessary for the correct interpre-tation of population interrelationships, even when analyzed in a local context. The study of population relationships using microsatellites alone may yield misleading results because distant genetic relationships are not evident due to the high mutation rates of microsatellites. Optimally, other techniques should be combined with microsatellites to cover a broad taxonomic spectrum. On the contrary, both nuclear microsat-ellite markers (in particular locus Pagt 22) and trn T- trn L and rbc L- psa I cp-sequences, which are based on repetitive se-quences, proved to have a strong phylogeographic resolution to infer relationships among closely related genotypes across continents.
It is only with a global sampling and the combination of several methods that we can recognize the genetic dynamics in the GC and conclude that gene fl ow and introgressive hy-bridization are likely to occur in the GC area. The common occurrence of the alleles in the GC types could alternatively be explained by independent mutations and/or multiple dis-persal events from each of the source populations. The fi rst explanation would entail a far higher number of mutations/evolutionary events and inconsistencies among markers than observed here. Hence, frequent independent mutations can be dismissed as a less parsimonious hypothesis. As to the second possible explanation, the low genetic variation within each GC-type, compared to the native populations, as well as the limited number of alleles and almost invariable chromatogram patterns, testify against multiple introductions. Ancestral polymorphisms could be implied, but then higher genetic sim-ilarities among the GC-types and among their native popula-tions, especially with Phragmites mauritianus populations, would be expected. The reason why Hauber et al. (2011) did not fi nd evidence of gene fl ow, using the same microsatellites as we did, is probably their exclusion of polysomic markers that were incompatible with codominant microsatellite analyses. Some of these actually showed recombining alleles in our analyses.
This suggests gene fl ow also between the European related GC genotypes and the Land-type. In general, gene fl ow among the GC-genotypes appears to be the most parsimonious inter-pretation of the pattern of allele distribution in the GC gene pool. Alternative explanations entail a large number of paral-lel gains and losses of alleles.
DISCUSSION
As suggested by the visually apparent differences in pheno-type, the Mississippi River delta is a “ hot spot ” of Phragmites diversity. The area is home to fi ve distinct cpDNA haplotypes differing in geographic origin, time of introduction, and ge-netic traits. Their phylogeographic relationships show that long-distance dispersal is a recurring phenomenon within Phragmites and that reproductive barriers among haplotypes, as well as among the extant Phragmites species, are weak. We provide evidence of at least two hybridization events between P. mauritianus and P. australis , and of back-crossing of one of these hybrids with the paternal species in the GC area. The implications reach beyond the GC area and concern the entire genus. An additional example of interspecifi c hybridization within the genus was provided by Chu et al. (2011) and in-volves P. australis and P. japonicus in South Korea. As in our study, no intermediate morphological form was observed and the authors suggested that the P. australis phenotype might be dominant in the hybrids. The cosmopolitan, and ecologically unrivalled, P. australis, can interbreed with two of its four con-geners (interbreeding with P. frutescens and P. karka has never been tested). One possible interpretation of these fi ndings is that P. australis might constitute a hybrid swarm. This however needs to be tested further, particularly in areas where several species coexist.
Another important implication of this study is that, given the frequency of long-distance dispersal, a broad geographic
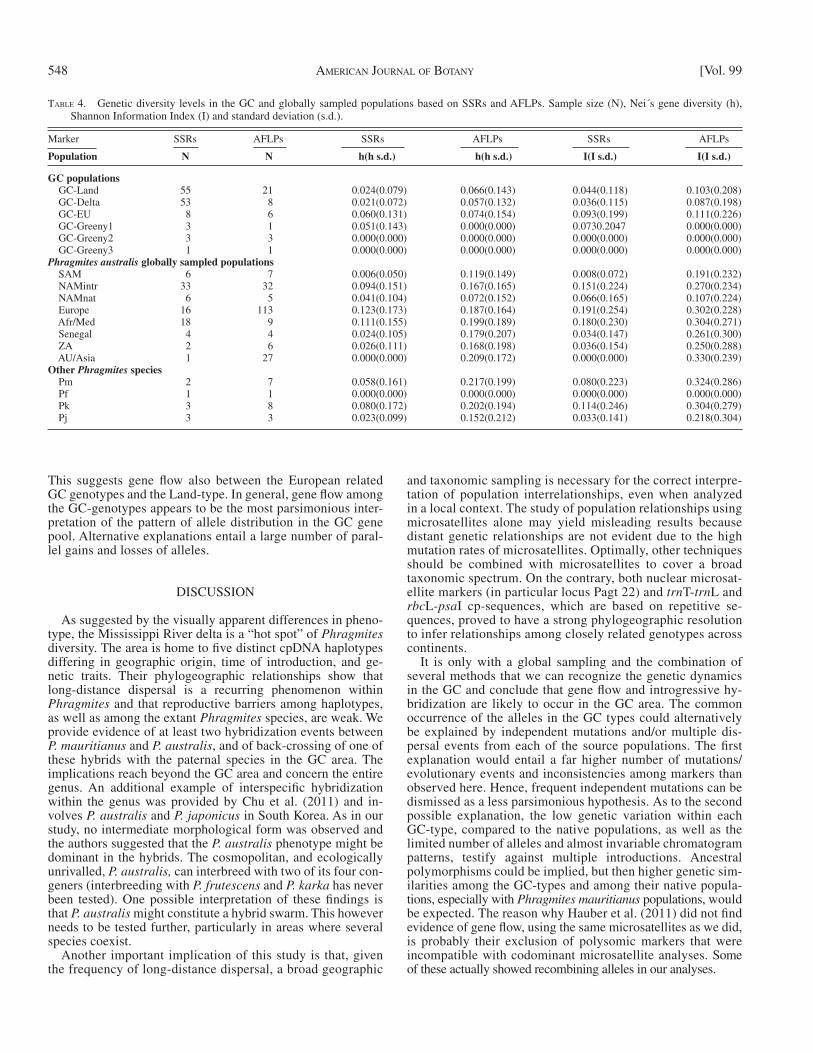
TABLE 4. Genetic diversity levels in the GC and globally sampled populations based on SSRs and AFLPs. Sample size (N), Nei ́ s gene diversity (h), Shannon Information Index (I) and standard deviation (s.d.).
Marker SSRs AFLPs SSRs AFLPs SSRs AFLPs
Population N N h (h s.d.) h (h s.d.) I (I s.d.) I (I s.d.)
GC populations GC-Land 55 21 0.024(0.079) 0.066(0.143) 0.044(0.118) 0.103(0.208) GC-Delta 53 8 0.021(0.072) 0.057(0.132) 0.036(0.115) 0.087(0.198) GC-EU 8 6 0.060(0.131) 0.074(0.154) 0.093(0.199) 0.111(0.226) GC-Greeny1 3 1 0.051(0.143) 0.000(0.000) 0.0730.2047 0.000(0.000) GC-Greeny2 3 3 0.000(0.000) 0.000(0.000) 0.000(0.000) 0.000(0.000) GC-Greeny3 1 1 0.000(0.000) 0.000(0.000) 0.000(0.000) 0.000(0.000) Phragmites australis globally sampled populations SAM 6 7 0.006(0.050) 0.119(0.149) 0.008(0.072) 0.191(0.232) NAMintr 33 32 0.094(0.151) 0.167(0.165) 0.151(0.224) 0.270(0.234) NAMnat 6 5 0.041(0.104) 0.072(0.152) 0.066(0.165) 0.107(0.224) Europe 16 113 0.123(0.173) 0.187(0.164) 0.191(0.254) 0.302(0.228) Afr/Med 18 9 0.111(0.155) 0.199(0.189) 0.180(0.230) 0.304(0.271) Senegal 4 4 0.024(0.105) 0.179(0.207) 0.034(0.147) 0.261(0.300) ZA 2 6 0.026(0.111) 0.168(0.198) 0.036(0.154) 0.250(0.288) AU/Asia 1 27 0.000(0.000) 0.209(0.172) 0.000(0.000) 0.330(0.239) Other Phragmites species Pm 2 7 0.058(0.161) 0.217(0.199) 0.080(0.223) 0.324(0.286) Pf 1 1 0.000(0.000) 0.000(0.000) 0.000(0.000) 0.000(0.000) Pk 3 8 0.080(0.172) 0.202(0.194) 0.114(0.246) 0.304(0.279) Pj 3 3 0.023(0.099) 0.152(0.212) 0.033(0.141) 0.218(0.304)

549March 2012] LAMBERTINI ET AL. — GULF COAST PHRAGMITES
and Australia ( Saltonstall, 2002 ), as well as North America ( Saltonstall 2002, 2003 ; Howard et al., 2008 ; Meyerson et al., 2010a; Hauber et al., 2011 ). Our samples of P. karka collected in tropical Asia and North Australia also showed similarities to Haplotype I. However, accessions of P. mauritianus from Uganda and Burkina Faso and Land-type share the 100 bp waxy fragment, absent in P. karka , and identical chloroplast and nu-clear sequences. Ward (2010) recognized several morphologi-cal similarities between the GC P. australis var. berlandieri and the tropical species P. karka that led to a taxonomic de-termination of P. australis var. berlandieri as P. karka . Phragmites karka and P. mauritianus are highly similar mor-phologically and very closely related genetically. It is there-fore likely to confuse the two species based on morphology only. Ward ́ s identifi cation of a tropical Phragmites species in the GC is thus in agreement with our results. Our trn T- trn L sequences differ from those of Saltonstall (2002) in the num-ber of repeats at a microsatellite locus. Repeated amplifi ca-tions and sequencing sessions consistently gave the same result. Considering the identity of Land-type markers with the African samples, it appears that two variants of this sequence are present in the GC. However, as mononucleotide repetitive regions in the trnT-trnL region were not previously used to defi ne haplotypes ( Saltonstall, 2002 ), the Land-type can be referred as Haplotype I.
Delta-type — The Delta-type appears to be an introduction independent of the Land-type, as unequivocally demonstrated by the trn T- trn L sequences and the 200 bp band amplifi ed by the waxy primers. The source population is in the North Afri-can/Mediterranean area. The presence/absence pattern of the 200 bp fragment amplifi ed by the waxy primers indicates that the African/Mediterranean population of Phragmites australis might also be the paternal lineage of the Land-type hybrid. It is therefore not surprising that the Delta-type can interbreed with the Land-type in the GC. The African distribution of Land-type ́ s paternal lineage supports the hypothesis of an African origin also for Land-type and entails two possible interpreta-tions of the introductory history of these two haplotypes to the GC. The Land- and the Delta-type might have been dispersed together from a mixed population in which parents and hy-brids coexisted, or they might have been established by sepa-rate introductions at different times. Our dataset supports an ancient introduction for Land-type and a recent introduction for Delta-type. The genetic identities of Delta-type with the native African/Mediterranean population at the microsatellite and AFLP loci and the phylogeographic topology of the AFLP network are inconsistent with a long history of allopatric evo-lution. The Delta-type ́ s haplotype appears to be closely re-lated to the European Haplotype M and identical to Haplotype M1 ( Hauber et al., 2011 ). The same nickname “ Delta ” was used by Hauber et al. (2011) to describe this morphological phenotype in the GC.
EU-type — Genetic identities and genetic diversity measure-ments suggest that the EU-type has recently been introduced from the European invasive North American population of Phragmites australis . The southward dispersal of the EU-type implicates the Mississippi River as the logical means of disper-sal. However we did not fi nd North American native genotypes in the Mississippi River delta, suggesting either strong ecologi-cal differences between the two North American lineages or alternative dispersal paths for the EU-type. A larger number of
The US Southwest offers another opportunity to test intro-gressive hybridization. Here GC Phragmites is present together with the North American native and introduced populations ( Meyerson et al., 2010a ; Kulmatiski et al., 2011). However, stands consisting of more than one type are rare. Experimental crossings can confi rm or dismiss the preconditions for inter-breeding, although the case of North America shows that green-house populations do not always react like wild populations (Meyerson et al., 2010b; Paul et al., 2010 ; Saltonstall, 2011 ). Several microsatellite markers should ideally be developed spe-cifi cally for Land-, Delta-, and the Greeny-types to increase support for cases of parentage. In any case, several examples show that conclusions about gene fl ow might differ depending on the type of DNA variation analyzed ( Muir and Schloetterer, 2005 ; Lexer et al., 2006 ).
In the following, the history of the GC phenotypes and of their phylogeographic relationships is presented. We could not have addressed gene fl ow in the Gulf Coast area without recon-structing their relationships, and in particular the relationship with Phragmites mauritianus , previously described in Lambertini et al. (2006) , because species boundaries and differences in ploidy levels represent breeding barriers in many species. On the contrary, as often is the case in plants, the biological species concept appears not to be applicable to the variation seen in Phragmites .
Land-type — Among the Phragmites lineages coexisting in the marshes of the Mississippi River delta, the Land-type ap-pears to be a hybrid between P. mauritianus and P. australis . This is supported by identical cpDNA sequences as P. mauri-tianus , and several nuclear homologies from P. australis acces-sions from the African and Mediterranean area ( waxy bands and AFLP fragments), which are absent in P. mauritianus . The Land-type ́ s closest relatives are the South American geno-types, which seem to have the same hybrid origin and share several similarities both in the chloroplast and nuclear genomes. Even though it is not evident from this dataset if the Land-type evolved from the South American group or the reverse, they obviously share a common ancestor. We hypothesize that the presence of this hybrid in the Americas is the result of a com-mon founder from Africa. This is based on the present distribu-tion ranges of P. mauritianus and P. australis , which largely overlap in the African tropical region. The variation in the bands amplifi ed by the waxy primers indicates Senegal to be a hybrid zone. However, Senegal ’ s hybrids have different cpDNA sequences from Land-type and South American acces-sions, and therefore cannot be confi rmed as the source popula-tion. This introduces other possible scenarios for the origins of the American populations. A previously larger distribution of P. mauritianus in tropical America could alternatively explain an American origin of this hybrid as well as the similarities between Land-type genotypes and the South American popula-tion. More samples from South America, as well as from tropi-cal Africa, need to be analyzed to address conclusively the origin, dispersal, and evolution of this Phragmites haplotype in the Americas.
Using the nomenclature introduced by Saltonstall (2002) to identify native and introduced haplotypes of Phragmites aus-tralis in North America, which is based on the combination of the trn T- trn L and rbc L- psa I sequences, the Land haplotype is closely related with Haplotype I. Haplotype I has been formally described as P. australis var. berlandieri ( Saltonstall et al., 2004 ) and has previously been found in South America, Asia,

550 AMERICAN JOURNAL OF BOTANY [Vol. 99
Even though the three Greeny haplotypes are distinctive and appear to have independent biogeographical relationships, the Greeny genotypes share several nuclear alleles, all of which are present in the introduced North American population and/or in Europe. All three Greeny-types also have high AFLP genetic identities with the South African population, whereas only the Greeny3-type shares the trn T- trn L sequence with the South Af-rican population. It may be possible that the nuclear homolo-gies among the three types derive from intensive gene fl ow in the GC area. However, the extent of such similarities appears too high, given the time available, to have been acquired in the GC, as well as in other parts of North America. A common ancestral gene pool, most likely European, appears a more plausible explanation for these similarities. Like the three Greeny-types, the European population has high AFLP ge-netic identities with the South African population. Even though several long-distance biogeographical relationships remain to be fully explored, the range of possibilities has been consider-ably reduced. The results specifi cally point to Phragmites aus-tralis in North America, Danube River delta, Spain, South Africa, and to the African species P. mauritianus to trace the history of the Greeny-types.
LITERATURE CITED
BANDELT , H. J. , P. FORSTER , AND A. ROEHL . 1999 . Median-joining net-works for inferring intraspecifi c phylogenies. Molecular Biology and Evolution 16 : 37 – 48 .
CHAMBERS , R. M. , L. A. MEYERSON , AND K. SALTONSTALL . 1999 . Expansion of Phragmites australis into tidal wetlands of North America. Aquatic Botany 64 : 261 – 273 .
CHU , H. , W. K. CHO , Y. JO , W.-I. I. KIM , Y. RIM , AND R.-Y. KIM . 2011 . Identifi cation of natural hybrids in Korean Phragmites using haplo-type and genotype analyses. Plant Systematics and Evolution 293 : 247 – 253 .
CLEVERING , O. A. , AND J. LISSNER . 1999 . Taxonomy, chromosome numbers, clonal diversity and population dynamics of Phragmites australis. Aquatic Botany 64 : 175 – 208 .
DE QUEIROZ , K. 2007 . Species concepts and species delimitation. Systematic Biology 56 : 879 – 886 .
FARRIS , J. S. 1970 . Methods for computing Wagner trees. Systematic Zoology 19 : 83 – 92 .
FOURNIER , W. , D. P. HAUBER , AND D. A. WHITE . 1995 . Evidence of in-frequent sexual propagation of Phragmites australis throughout the Mississippi River delta. American Journal of Botany 82 (6, Supplement) : 71 .
GAUDREAULT , S. , D. A. WHITE , AND D. P. HAUBER . 1989 . Phragmites australis : An analysis of reproductive differences in two adjacent pop-ulations in the Mississippi River delta. American Journal of Botany 76 (6, Supplement) : 103 .
GORENFLOT , R. , P. RAICU , D. CARTIER , I. CIOBANU , V. STOIAN , AND S. STAICU . ( 1972 ) Le complexe polyploide du Phragmites communis Trin. Comptes rendus hebdomadaire des S é ances de l ’ Acad é mie des Sciences. S é rie D, Sciences naturalles, Paris 274: 1501-1504.
GORENFLOT, R. , M. SANEI-CHARIAT PANAHI , AND J. LIEBERT . ( 1979 ) Le com-plexe polyploide du Phragmites australis (Cav.) Trin. ex Steud. en Iran. Revue de Cytologie et de Biologie v é getales-le Botaniste. Paris 2: 67-81.
GORMAN , G. C. , AND J. RENZI JR . 1979 . Genetic distance and heterozy-gosity estimates in electrophoretic studies: Effects of sample size. Copeia 2 : 242 – 249 .
GULDBRANDTSEN , B. , J. TOMIUK , AND V. LOESCHCKE . 2000 . Popdist, version 1.1.1: A program to calculate population genetic distance and identity measures . Journal of Heredity 91 : 178 – 179 .
GULDBRANDTSEN , B. , J. TOMIUK , AND V. LOESCHCKE . 2009 . POPDIST, version 1.2.0: User ’ s guide. http://genetics.agrsci.dk/~bernt/pop-gen/userguide.pdf [accessed 15 December 2011].
EU-type samples should therefore be analyzed before defi ni-tively ruling out the hypothesis of a direct introduction from Europe. Our data suggest that there is gene fl ow between the EU- and the Land-type. Considering the closer phylogenetic relationships of the EU-type with the Delta-type, it appears likely that the EU-type can also interbreed with the conspecifi c Delta-type. This hypothesis is supported by the presence/absence pattern of microsatellites. Due to the strong genetic similarities between their native populations, it was however not possible to distin-guish between alleles originally present in the ancestral popula-tions and alleles shared due to recombination in the GC area.
The EU-type has haplotype M ( Saltonstall, 2002 ). Identical microsatellite alleles were found in the North American intro-duced populations and along the coasts of the North Sea in Hol-land and Belgium and along the Baltic Sea in Denmark and Finland. The EU-type appears to be the same morphological phenotype that Hauber et al. (2011) defi ned as “ Short ” .
Greeny-types — The similarities in morphology and nuclear DNA among the three Greeny-types suggest a common origin or a long history of sympatry of these genotypes prior to estab-lishment in the GC. On the contrary, the cpDNA sequences identify three distinctive lineages. The Greeny1-type has Hap-lotype M ( Saltonstall, 2002 ) in common with the EU-type, but differs from the EU-type in the microsatellite alleles. As both sets of microsatellites are present both in the introduced North American and European populations, they are not helpful in as-sessing from which population it is derived.
Hauber et al. (2011) suggested a local mutation in Haplotype M to explain the origin of Haplotype AD, which corresponds to our Greeny2-type. The hypothesis of Hauber et al. considers the 8 bp indel in the trnt - trn L region as independently derived in the native North American population on the one hand and in the Haplotype AD, alias Greeny2-type, on the other. Further evidence, from our data, of a European origin from Haplotype M is the shared European nuclear alleles. Another, novel hy-pothesis is that the Greeny2-type is a mutant of one native North American haplotype. The 8 bp indel seems an exclusive trait of the native North American haplotypes and of Phragmites mau-ritianus (the only uncertain cases are the trn T- trn L Haplotypes E1, E3, E4, and M2 ( Vachon and Freeman, 2011 ) of which we could not fi nd information concerning the rbc L- psa I sequences. In other words, it is hard to say if the Greeny2-type originated from the European, or from the North American introduced or native populations. It shares the same nuclear alleles as Euro-pean and North American introduced populations, but its cp-sequences have been found only among North American na-tive samples and in P. mauritianus . To resolve the issue of the single vs. multiple origin of the 8 bp indel, more samples from North America and of P. mauritianus need to be included in the analysis.
The Greeny3-type, or new Haplotype AI (K. Saltonstall, per-sonal communication), shares the chloroplast with European samples from the Danube River delta in Romania. We cannot assess if the Danube River delta is the source area of the Greeny3- type or if the origin is in yet another unsampled area. The apparently restricted distribution of this haplotype in the Danube River delta makes it less likely as source area. The cp-DNA sequences indicate relationships also with one single sample in Spain and a geographically widespread population in South Africa and Namibia. Strikingly, both our Romanian and South African samples, with trn T -trn L sequences identical to the Greeny3-type, are octoploids.

551March 2012] LAMBERTINI ET AL. — GULF COAST PHRAGMITES
HALL , T. A. ( 1999 ) Bioedit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series 41: 95-98.
HAUBER , D. P. , K. SALTONSTALL , D. A. WHITE , AND C. S. HOOD . 2011 . Genetic variation in the common reed, Phragmites australis , in the Mississippi River delta marshes: Evidence for multiple introductions. Estuaries and Coasts 34 : 851 – 862 .
HAUBER D. P. , D. A. WHITE , S. P. POWERS , AND F. R. DEFRANCESCH. ( 1991 ) Isozyme variation and correspondence with unusual infrared refl ec-tance patterns in Phragmites australis (Poaceae). Plant Systematics and Evolution 178: 1-8.
HOWARD , R. J. , S. E. TRAVIS , AND B. A. SIKES . 2008 . Rapid growth of a Eurasian haplotype of Phragmites australis in a restored brackish marsh in Louisiana, USA. Biological Invasions 10 : 369 – 379 .
KRUSKAL , J. B. 1956 . On the shortest spanning subtree of the graph and the travelling salesman problem. Proceedings of the American Mathematical Society 7 : 48 – 57 .
KULMATISKI , A. , K. H. BEARD , L. A. MEYERSON , J. R. GIBSON , AND K. E. MOCK . 2011 . Nonnative Phragmites australis invasion into Utah wetlands. Western North American Naturalist 70 : 541 – 552 .
LAMBERTINI , C. , J. FRYDENBERG , M. H. G. GUSTAFSSON , AND H. BRIX . 2008 . Herbarium specimens as a source of DNA for AFLP fi ngerprinting of Phragmites (Poaceae): Possibilities and limitations. Plant Systematics and Evolution 272 : 223 – 231 .
LAMBERTINI , C. , M. H. G. GUSTAFSSON , J. FRYDENBERG , J. LISSNER , M. SPERANZA , AND H. BRIX . 2006 . A phylogeographic study of the cosmopolitan genus Phragmites (Poaceae) based on AFLPs. Plant Systematics and Evolution 258 : 161 – 182 .
LEWONTIN , R. C. 1972 . Apportionment of human diversity. Evolutionary Biology 6 : 381 – 398 .
LEXER , C. , A. KREMER , AND R. J. PETIT . 2006 . Shared alleles in sympat-ric oaks: Recurrent gene fl ow is a more parsimonious explanation than ancestral polymorphism. Molecular Ecology 15 : 2007 – 2012 .
MASON-GAMER , R. J. , C. F. WEIL , AND E. A. KELLOGG . 1998 . Granule-bound starch synthase: Structure, function and phylogenetic utility. Molecular Biology and Evolution 15 : 1658 – 1673 .
MAYR , E. ( 1942 ) Systematics and the origin of species. Columbia University Press, New York, New York, USA.
MCCORMICK , M. K. , K. M. KETTENRING , H. M. BARON , AND D. F. WHIGHAM . 2010 . Spread of invasive Phragmites australis in estuaries with dif-fering degrees of development: Genetic patterns, Allee effects and in-terpretation. Journal of Ecology 98 : 1369 – 1378 .
MEYERSON , L. A. , A. M. LAMBERT , AND K. SALTONSTALL . 2010a . A tale of three lineages: Expansion of common reed ( Phragmites australis ) in the U.S. Southwest and Gulf Coast. Invasive Plant Science and Management 3 : 515 – 520 .
MEYERSON , L. A. , D. V. VIOLA , AND R. N. BROWN . 2010b . Hybridization of invasive Phragmites australis with a native subspecies in North America. Biological Invasions 12 : 103 – 111 .
MUIR , G. , AND C. SCHLOETTERER . 2005 . Evidence of shared ancestral poly-morphism rather than recurrent gene fl ow at microsatellite loci dif-ferentiating two hybridizing oaks ( Quercus spp.). Molecular Ecology 14 : 549 – 561 .
NEI , M. 1973 . Analysis of gene diversity in subdivided populations. Proceedings of the National Academy of Sciences, USA 70 : 3321 – 3323 .
NEI , M. 1978 . Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 89 : 583 – 590 .
OTTO , S. P. , AND J. WHITTON . 2000 . Polyploid evidence and evolution. Annual Review of Genetics 34 : 401 – 437 .
PAETKAU , D. , W. CALVERT , I. STIRLING , AND C. STROBECK . 1995 . Microsatellite analysis of population structure in Canadian polar bears. Molecular Ecology 4 : 347 – 354 .
PAUL , J. , N. VACHON , C. J. GARROWAY , AND J. R. FREELAND . 2010 . Molecular data provide strong evidence of natural hybridization between native and introduced lineages of Phragmites australis in North America. Biological Invasions 12 : 2967 – 2973 .
PEAKALL , R. , AND P. E. SMOUSE . 2006 . GenAlEx 6: Genetic analysis in Excel. Population genetic software for teaching and research. Molecular Ecology Notes 6 : 288 – 295 .
PELLEGRIN , D. , AND D. P. HAUBER . 1999 . Isozyme variation among popula-tions of the clonal species, Phramites australis (Cav.) Trin. ex Steudel. Aquatic Botany 63 : 241 – 259 .
POSNO , M. , A. VLIET , AND G. S. P. GROOD . 1986 . The gene for Spirodela oligorhiza chloroplast ribosomal protein homologous to E. coli ribosomal protein L16 is split by a large intron near its 5 ′ end: Structure and expression. Nucleic Acids Research 14 : 3181 – 3195 .
RAICU , P. , S. STAICUS , V. STOIAN , AND T. ROMAN . 1972 . The Phragmites australis Trin. chromosome complement in the Danube delta. Hydrobiologia 39 : 83 – 89 .
SALTONSTALL , K. 2001 . A set of primers for the amplifi cation of noncod-ing regions of chloroplast DNA in the grasses. Molecular Ecology Notes 1 : 76 – 78 .
SALTONSTALL , K. 2002 . Cryptic invasion by a non-native genotype of the common reed, Phragmites australis , into North America. Proceedings of the National Academy of Sciences, USA 99 : 2445 – 2449 .
SALTONSTALL , K. 2003 . Microsatellite variation within and among North American lineages of Phragmites australis. Molecular Ecology 12 : 1689 – 1702 .
SALTONSTALL , K. 2011 . Remnant native Phragmites australis maintains high genetic diversity despite multiple threats. Conservation Genetics 12 : 1027 – 1033 .
SALTONSTALL , K. , P. M. PETERSON , AND R. J. SORENG . 2004 . Recognition of Phragmites australis subsp. americanus (Poaceae: Arundinoideae) in North America: Evidence from morphological and genetic analyses. Sida 21 : 683 – 692 .
SOLTIS , D. E. , AND P. S. SOLTIS . 1993 . Molecular data and the dynamic na-ture of polyploidy. Critical Reviews in Plant Sciences 12 : 243 – 273 .
STEBBINS , G. L. ( 1971 ) Chromosomal evolution in higher plants. Edward Arnold (Publishers) Ltd., London, UK.
SWOFFORD , D. L. ( 1998 ) PAUP, version 4.. Phylogenetic analysis using parsimony (and other methods). Sinauer, Sunderland, Massachusetts, USA.
TABERLET , P. , L. GIELLY , G. PAUTOU , AND J. BOUVET . 1991 . Universal primers for amplifi cation of three non-coding regions of chloroplast DNA. Plant Molecular Biology 17 : 1105 – 1109 .
TOMIUK , J. , AND V. LOESCHCKE . 1991 . A new measure of genetic iden-tity between populations of sexual and asexual species. Evolution; International Journal of Organic Evolution 45 : 1685 – 1694 .
VACHON , N. , AND J. R. FREEMAN . 2011 . Phylogeographic inference from chloroplast DNA: Quantifying the effects of mutations in repetitive and non-repetitive sequences. Molecular Ecology Resources 11 : 279 – 285 .
WARD , D. B. 2010 . North America has two species of Phragmites (Gramineae). Castanea 75 : 394 – 401 .
WHITE , D. A. , D. P. HAUBER , AND C. S. HOOD . 2004 . Clonal differences in Phragmites australis from the Mississippi River delta. Southeastern Naturalist (Steuben, ME) 3 : 531 – 544 .
YEH , F. C. , AND T. J. B. BOYLE . 1997 . Population genetic analysis of co-dominant and dominant markers and quantitative traits. Belgian Journal of Botany 129 : 157 .
ZHANG , W. 2000 . Phylogeny of the grass family (Poaceae) from rpl16 intron sequence data. Molecular Phylogenetics and Evolution 15 : 135 – 146 .




















