
Diet of Octopus bimaculatus Verril, 1883 (Cephalopoda: Octopodidae) in Bahía De Los Ángeles, Gulf...
11
BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. Diet of Octopus bimaculatus Verril, 1883 (Cephalopoda: Octopodidae) in Bahía De Los Ángeles, Gulf of California Author(s): Elisa Jeanneht Armendáriz Villegas , Bertha Patricia Ceballos-Vázquez , Unai Markaida , Andrés Abitia-Cárdenas , Marco Antonio Medina-López and Marcial Arellano-Martínez Source: Journal of Shellfish Research, 33(1):305-314. 2014. Published By: National Shellfisheries Association DOI: http://dx.doi.org/10.2983/035.033.0129 URL: http://www.bioone.org/doi/full/10.2983/035.033.0129 BioOne (www.bioone.org ) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use . Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.
Transcript of Diet of Octopus bimaculatus Verril, 1883 (Cephalopoda: Octopodidae) in Bahía De Los Ángeles, Gulf...

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, researchlibraries, and research funders in the common goal of maximizing access to critical research.
Diet of Octopus bimaculatus Verril, 1883 (Cephalopoda: Octopodidae) in Bahía DeLos Ángeles, Gulf of CaliforniaAuthor(s): Elisa Jeanneht Armendáriz Villegas , Bertha Patricia Ceballos-Vázquez , Unai Markaida ,Andrés Abitia-Cárdenas , Marco Antonio Medina-López and Marcial Arellano-MartínezSource: Journal of Shellfish Research, 33(1):305-314. 2014.Published By: National Shellfisheries AssociationDOI: http://dx.doi.org/10.2983/035.033.0129URL: http://www.bioone.org/doi/full/10.2983/035.033.0129
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, andenvironmental sciences. BioOne provides a sustainable online platform for over 170 journals and books publishedby nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiriesor rights and permissions requests should be directed to the individual publisher as copyright holder.

DIET OF OCTOPUS BIMACULATUS VERRIL, 1883 (CEPHALOPODA: OCTOPODIDAE) IN
BAHIA DE LOS ANGELES, GULF OF CALIFORNIA
ELISA JEANNEHT ARMENDARIZ VILLEGAS,1 BERTHA PATRICIA CEBALLOS-VAZQUEZ,1
UNAI MARKAIDA,2 ANDRES ABITIA-CARDENAS,1 MARCO ANTONIO MEDINA-LOPEZ3
AND MARCIAL ARELLANO-MARTINEZ1*1Instituto Politecnico Nacional-CICIMAR, Av. Instituto Politecnico Nacional s/n. Col. Playa Palo deSanta Rita, La Paz, Baja California Sur, 23096 Mexico; 2Colegio de la Frontera Sur, Unidad Campeche,Calle 10361 #264, Col. Centro, Campeche, Campeche, Mexico; 3Universidad Autonoma de BajaCalifornia Sur, Carretera al Sur km. 5, La Paz, Baja California Sur, Mexico
ABSTRACT Two hundred sixty-one octopuses were obtained from August 2006 to June 2007 in Bahıa de Los Angeles, BC,
Mexico. Sizes ranged from 58–190 mm in mantle length. Diet was determined from 3 sources: the digestive tract analysis (hard
rests), accumulations of hard prey remaining in refuges, and live prey present during capture. Ripe females had the greatest
fullness weight index (FWI) whereas spawning/spent females had the lowest. During the spring, female and male octopuses
showed the greatest FWI, whereas in summer they showed the lowest, coinciding with the spawning/spent stage. A total of 76 prey
items from 8 phyla were found, withMollusca being the most important phylum and xanthid crabs the most important prey year-
round.During autumn andwinter, more bivalves were consumed, whereasmore crabs were consumed in spring.Males fedmainly
on crabs during all gonad development stages, but spentmales fedmostly onmolluscs. In contrast, females fedmostly onmolluscs,
except ripe females, which includedmore crabs in their diet. The octopusOctopus bimaculatus appears to be a specialist consumer, and
this selectivity could be a consequence of different energetic demands of each sex during the gonad ripening process.
KEY WORDS: food preference, diet, midden, digestive tract analysis, California two-spotted octopus, Octopus bimaculatus
INTRODUCTION
The California 2-spotted octopus Octopus bimaculatus Ver-ril, 1883, is 1 of the most abundant species in the commercialcatches of the Gulf of California coast. It is distributed from
Santa Barbara, CA, to the southern Baja California Peninsula,and within the Gulf of California from Puerto Penasco, Sonoraand San Felipe, BC, to Bahıa de La Paz, BCS. (Hochberg &
Fields 1980). It inhabits intertidal and subtidal rocky coasts,where it finds appropriate refuges to hide from predators andspawn (Ambrose 1982).
Octopuses play an important role in the marine environment
as both predators and prey (Nixon 1987, Guerra 1992). Anunderstanding of their trophic relationships would help indetermining the structure and function of marine ecosystems
(Cherel & Hobson 2005). In addition, knowledge of theirfeeding habits is a key aspect that has a bearing on the survivaland growth of these organisms in captivity (Nixon 1966).
Octopuses consume a wide variety of prey, including crusta-ceans and molluscs (Nixon 1987); however, their diet can beinfluenced by preference and by prey availability (Ambrose1984), which has consequences for the community. Also,
modifications to their feeding behavior and food ingestion areassociated with the gonad ripening process (Cortez et al. 1995),although this aspect has not been well studied despite its
biological relevance. In general, a decrease in food ingestionhas been observed in ripe and broody females of differentoctopus species in captivity (Joll 1976, Wodinsky 1978, Mather
1980, Smale & Buchan 1981), as well as in the wild (Lopez-Uriarte et al. 2010). This has been confirmed indirectly by a highincidence of ripe females with empty stomachs (Quetglas et al.
2005) as well as by a decrease of energetic reserves in the
digestive gland (O�Dor & Wells 1978, Cortez et al. 1995,Zamora & Olivares 2004) and in muscle (Zamora & Olivares
2004).In general, the study of wild octopus diets is based on
stomach content analysis; however, the fragmented state of thestomach contents, caused by the mechanical action of the beak,
complicates the analysis. In this sense, only prey with hardpieces can be identified, such as molluscs (shells smaller than theaperture of themandibular apparatus), crustaceans (exoskeletal
fragments), and fish (bones) (Villanueva 1993, Quetglas et al.1998, Hernandez-Lopez 2000, Cardoso et al. 2004, Quetglaset al. 2005). Another technique used in diet studies is based on
the fact that octopuses discard large, hard prey parts, such ascrustacean exoskeletons, mollusc shells, or sea urchin tests,which accumulated around their refuge and can be identified
and quantified (Ambrose 1982, Ambrose 1983, Ambrose &Nelson 1983, Mather & O�Dor 1991, Dodge & Scheel 1999,Smith 2003). In the literature both techniques are used sepa-rately and combined (Smale & Buchan 1981, Cortez et al. 1995),
and a third technique, based on direct observation of in situfeeding, has been also used (Smith 2003), which gives a betterapproximation to the feeding habits of these organisms.
The objective of the current study was to determine the dietofOctopus bimaculatus in Bahıa de Los Angeles, combining thethree sampling techniques, and analyzing the diet variation in
relation to season, sex, and gonad development stage.
MATERIALS AND METHODS
This research was carried out on a dense octopus populationin Bahıa de Los Angeles, Mexico, on the northwest coast of theBaja California Peninsula (28�55# N, 113�30# W) within theGulf of California. This bay is surrounded by numerous small
islands (Barnard & Grady 1968); it has a sandy bottom, and*Corresponding author. E-mail: [email protected]
DOI: 10.2983/035.033.0129
Journal of Shellfish Research, Vol. 33, No. 1, 305–314, 2014.
305

large rocks with fissures and cavities that are used by octopusesas a refuge. The mouth of the bay adjoins Channel de Ballenas,
and the water exchange between the channel and the bay allowsthe mixing of cold water (rich in nutrients) with warm baywaters, which provide primary and secondary productivity inthe area year-round and results in high biodiversity (Bustos-
Serrano et al. 1996, Delgadillo-Hinojosa et al. 1997).Samples were obtained from August 2006 to June 2007
between 0800 and 1100 HR by diving with an air compressor
‘‘hookah’’ and catching the octopuses with hooks. Each month,20–30 different-sizeOctopus bimaculatuswere caught at a depthof 5–12 m. Along with each octopus, recent accumulations of
hard prey remains found in the refuge were collected. Some preythatwere being ingestedby the octopuses at the timeof capturewerealso collected and were recorded as ‘‘live prey’’ observations.
In the laboratory, sex (by direct observation of gonads),
total weight (TW) in grams, and the mantle length (ML) andtotal length (TL) in millimeters were recorded for each organ-ism. The digestive tract was extracted and stored at –20�C for
additional analysis. The gonad development stage was deter-mined histologically as immature, developing, ripe, spawning,or spent (Olivares-Paz et al. 2001, Rodrıguez-Rua et al. 2005).
After thawing, the digestive tract (crop and stomach)contents were weighed and preserved in 70% alcohol foradditional analysis. The hard rests were used to identify the
prey. The fullness weight index (FWI) was calculated as thepercent ratio of the weight of the digestive tract contents tothe total weight of the organism less the empty digestive tractweight (Hyslop 1980). To assess significant differences in FWI
for females, males, and combined among seasons, and gonaddevelopment stages, Kruskal–Wallis tests were used.
Taxonomic Identification of Prey
Prey were identified to the minimum possible taxonomic
level (with each considered as 1 item) using specialized keysaccording to the taxonomic group: crustaceans (Brusca 1980,Hendrickx 1995, Kerstitch & Bertsch 2007), bivalves andgastropods (Keen 1971,Brusca 1980, Poutiers 1995), fish (Heemstra
1995, Sommer 1995), polychaetes (Brusca & Brusca 2005), seaurchins and ophiurans (Kerstitch & Bertsch 2007). Echiuranworms and sipunculids were identified by the presence of chetae
in the digestive tract (Fisher 1946, Brusca 1980, Brusca & Brusca2005). Food in an advanced state of digestion was labeled asunidentified organic matter and was quantified as 1 U for each
digestive tract in the analysis of frequency of occurrence (FO) only.
Diet Analysis
The diet was analyzed by taking into account informationobtained with the three sampling methods (digestive tract,accumulation of prey remains in the refuge, and observation
of live prey). The importance of each item (prey category) wasdetermined with two quantitative methods: FO (calculated asthe percent ratio of the total number of stomachs containing
a given item with respect to the total number of stomachscontaining food) (Cailliet et al. 1996) and number (calculated asthe percent ratio of the total number of individuals of a prey
item with respect to the total number of prey items).Data were grouped by season—summer (August and Sep-
tember 2006), autumn (October to December 2006), winter
(January to March 2007), and spring (April to June 2007)—bysex, and by gonad development stage (immature, developing,
ripe, spawning, and spent) (assigned histologically to the samespecimens by Castellanos-Martınez 2008). The two immaturemales were grouped with the developing males and the fourspawning/spent females were also grouped because they were of
similar size and they fed on the same prey. To compare diets,items were grouped as follows: crabs, bivalves, gastropods,echiurans, fish, and other prey.
To determine the general diet width by sex and by gonaddevelopment stage, Levin�s index (B) was used with the frequencyof occurrence. B values more than 3.0 denote generalist predators;
B values less than 3.0 denote specialist predators (Hurlbert 1978).
RESULTS
Two hundred sixty-one Octopus bimaculatus were collected,120 (46%) were female and 141 (54%) were male. Malesmeasured from 58–190 mm in ML (average ML, 114 ± 29 mm
(SD)) and weighed between 115 g and 2,200 g TW (average TL,635 ± 434 g). Females measured from 54–240 mm in ML(average ML, 118 ± 37 mm) and weighed between 130 g and
3,415 g TW (average TW, 720 ± 585 g). After reviewing the 261octopuses sampled, only 203 contained food; 83% had food inthe digestive tract, 39% had accumulations of prey remains in
their refuge, and 4% had live prey during capture (Fig. 1).From the total sample, it was possible to assign a gonadal
development stage to 87 females and 97 males only, and 5females contained no food. For the analysis of diet composition
by gonad development stage, 82 females with food wereanalyzed: 41 immature, 14 developing, 23 ripe, and 4 spawn-ing/spent (1 spawning, 3 spent). In addition, 97 males with food
were analyzed: 18 immature/developing (2 immature, 16 de-veloping), 47 ripe, and 17 spawning.
General Diet Description
A total of 1,138 prey corresponding to 76 items or preycategories were identified, belonging to 8 phyla: Arthropoda,
Mollusca, Echiura, Sipuncula, Echinodermata, Ectoprocta,Annelida, and Chordata. All these phyla were recorded in thedigestive tract, 4 were recorded in prey accumulations, and only
3 were noted in live prey (Table1).
Figure 1. Frequency of octopuses with food according to the different
sampling techniques. Digestive tract, digestive tract + midden,
midden, midden + live prey, live prey, without food.
ARMENDARIZ VILLEGAS ET AL.306
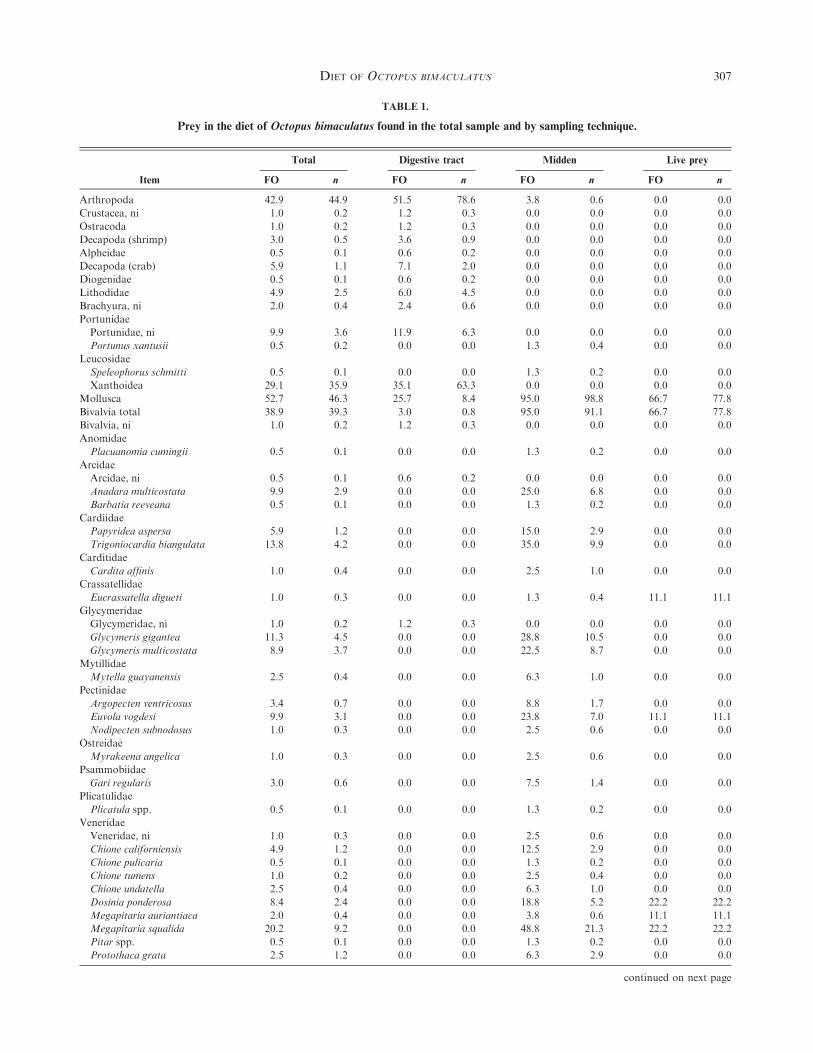
TABLE 1.
Prey in the diet of Octopus bimaculatus found in the total sample and by sampling technique.
Item
Total Digestive tract Midden Live prey
FO n FO n FO n FO n
Arthropoda 42.9 44.9 51.5 78.6 3.8 0.6 0.0 0.0
Crustacea, ni 1.0 0.2 1.2 0.3 0.0 0.0 0.0 0.0
Ostracoda 1.0 0.2 1.2 0.3 0.0 0.0 0.0 0.0
Decapoda (shrimp) 3.0 0.5 3.6 0.9 0.0 0.0 0.0 0.0
Alpheidae 0.5 0.1 0.6 0.2 0.0 0.0 0.0 0.0
Decapoda (crab) 5.9 1.1 7.1 2.0 0.0 0.0 0.0 0.0
Diogenidae 0.5 0.1 0.6 0.2 0.0 0.0 0.0 0.0
Lithodidae 4.9 2.5 6.0 4.5 0.0 0.0 0.0 0.0
Brachyura, ni 2.0 0.4 2.4 0.6 0.0 0.0 0.0 0.0
Portunidae
Portunidae, ni 9.9 3.6 11.9 6.3 0.0 0.0 0.0 0.0
Portunus xantusii 0.5 0.2 0.0 0.0 1.3 0.4 0.0 0.0
Leucosidae
Speleophorus schmitti 0.5 0.1 0.0 0.0 1.3 0.2 0.0 0.0
Xanthoidea 29.1 35.9 35.1 63.3 0.0 0.0 0.0 0.0
Mollusca 52.7 46.3 25.7 8.4 95.0 98.8 66.7 77.8
Bivalvia total 38.9 39.3 3.0 0.8 95.0 91.1 66.7 77.8
Bivalvia, ni 1.0 0.2 1.2 0.3 0.0 0.0 0.0 0.0
Anomidae
Placuanomia cumingii 0.5 0.1 0.0 0.0 1.3 0.2 0.0 0.0
Arcidae
Arcidae, ni 0.5 0.1 0.6 0.2 0.0 0.0 0.0 0.0
Anadara multicostata 9.9 2.9 0.0 0.0 25.0 6.8 0.0 0.0
Barbatia reeveana 0.5 0.1 0.0 0.0 1.3 0.2 0.0 0.0
Cardiidae
Papyridea aspersa 5.9 1.2 0.0 0.0 15.0 2.9 0.0 0.0
Trigoniocardia biangulata 13.8 4.2 0.0 0.0 35.0 9.9 0.0 0.0
Carditidae
Cardita affinis 1.0 0.4 0.0 0.0 2.5 1.0 0.0 0.0
Crassatellidae
Eucrassatella digueti 1.0 0.3 0.0 0.0 1.3 0.4 11.1 11.1
Glycymeridae
Glycymeridae, ni 1.0 0.2 1.2 0.3 0.0 0.0 0.0 0.0
Glycymeris gigantea 11.3 4.5 0.0 0.0 28.8 10.5 0.0 0.0
Glycymeris multicostata 8.9 3.7 0.0 0.0 22.5 8.7 0.0 0.0
Mytillidae
Mytella guayanensis 2.5 0.4 0.0 0.0 6.3 1.0 0.0 0.0
Pectinidae
Argopecten ventricosus 3.4 0.7 0.0 0.0 8.8 1.7 0.0 0.0
Euvola vogdesi 9.9 3.1 0.0 0.0 23.8 7.0 11.1 11.1
Nodipecten subnodosus 1.0 0.3 0.0 0.0 2.5 0.6 0.0 0.0
Ostreidae
Myrakeena angelica 1.0 0.3 0.0 0.0 2.5 0.6 0.0 0.0
Psammobiidae
Gari regularis 3.0 0.6 0.0 0.0 7.5 1.4 0.0 0.0
Plicatulidae
Plicatula spp. 0.5 0.1 0.0 0.0 1.3 0.2 0.0 0.0
Veneridae
Veneridae, ni 1.0 0.3 0.0 0.0 2.5 0.6 0.0 0.0
Chione californiensis 4.9 1.2 0.0 0.0 12.5 2.9 0.0 0.0
Chione pulicaria 0.5 0.1 0.0 0.0 1.3 0.2 0.0 0.0
Chione tumens 1.0 0.2 0.0 0.0 2.5 0.4 0.0 0.0
Chione undatella 2.5 0.4 0.0 0.0 6.3 1.0 0.0 0.0
Dosinia ponderosa 8.4 2.4 0.0 0.0 18.8 5.2 22.2 22.2
Megapitaria auriantiaca 2.0 0.4 0.0 0.0 3.8 0.6 11.1 11.1
Megapitaria squalida 20.2 9.2 0.0 0.0 48.8 21.3 22.2 22.2
Pitar spp. 0.5 0.1 0.0 0.0 1.3 0.2 0.0 0.0
Protothaca grata 2.5 1.2 0.0 0.0 6.3 2.9 0.0 0.0
continued on next page
DIET OF OCTOPUS BIMACULATUS 307
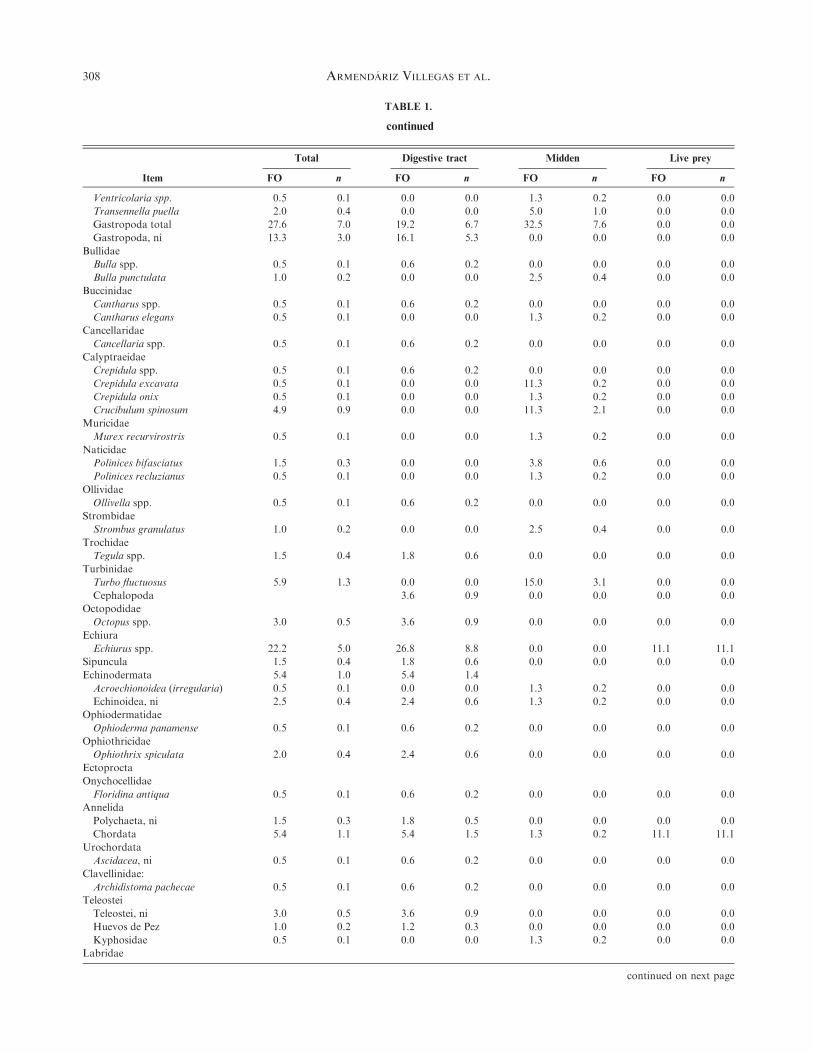
TABLE 1.
continued
Item
Total Digestive tract Midden Live prey
FO n FO n FO n FO n
Ventricolaria spp. 0.5 0.1 0.0 0.0 1.3 0.2 0.0 0.0
Transennella puella 2.0 0.4 0.0 0.0 5.0 1.0 0.0 0.0
Gastropoda total 27.6 7.0 19.2 6.7 32.5 7.6 0.0 0.0
Gastropoda, ni 13.3 3.0 16.1 5.3 0.0 0.0 0.0 0.0
Bullidae
Bulla spp. 0.5 0.1 0.6 0.2 0.0 0.0 0.0 0.0
Bulla punctulata 1.0 0.2 0.0 0.0 2.5 0.4 0.0 0.0
Buccinidae
Cantharus spp. 0.5 0.1 0.6 0.2 0.0 0.0 0.0 0.0
Cantharus elegans 0.5 0.1 0.0 0.0 1.3 0.2 0.0 0.0
Cancellaridae
Cancellaria spp. 0.5 0.1 0.6 0.2 0.0 0.0 0.0 0.0
Calyptraeidae
Crepidula spp. 0.5 0.1 0.6 0.2 0.0 0.0 0.0 0.0
Crepidula excavata 0.5 0.1 0.0 0.0 11.3 0.2 0.0 0.0
Crepidula onix 0.5 0.1 0.0 0.0 1.3 0.2 0.0 0.0
Crucibulum spinosum 4.9 0.9 0.0 0.0 11.3 2.1 0.0 0.0
Muricidae
Murex recurvirostris 0.5 0.1 0.0 0.0 1.3 0.2 0.0 0.0
Naticidae
Polinices bifasciatus 1.5 0.3 0.0 0.0 3.8 0.6 0.0 0.0
Polinices recluzianus 0.5 0.1 0.0 0.0 1.3 0.2 0.0 0.0
Ollividae
Ollivella spp. 0.5 0.1 0.6 0.2 0.0 0.0 0.0 0.0
Strombidae
Strombus granulatus 1.0 0.2 0.0 0.0 2.5 0.4 0.0 0.0
Trochidae
Tegula spp. 1.5 0.4 1.8 0.6 0.0 0.0 0.0 0.0
Turbinidae
Turbo fluctuosus 5.9 1.3 0.0 0.0 15.0 3.1 0.0 0.0
Cephalopoda 3.6 0.9 0.0 0.0 0.0 0.0
Octopodidae
Octopus spp. 3.0 0.5 3.6 0.9 0.0 0.0 0.0 0.0
Echiura
Echiurus spp. 22.2 5.0 26.8 8.8 0.0 0.0 11.1 11.1
Sipuncula 1.5 0.4 1.8 0.6 0.0 0.0 0.0 0.0
Echinodermata 5.4 1.0 5.4 1.4
Acroechionoidea (irregularia) 0.5 0.1 0.0 0.0 1.3 0.2 0.0 0.0
Echinoidea, ni 2.5 0.4 2.4 0.6 1.3 0.2 0.0 0.0
Ophiodermatidae
Ophioderma panamense 0.5 0.1 0.6 0.2 0.0 0.0 0.0 0.0
Ophiothricidae
Ophiothrix spiculata 2.0 0.4 2.4 0.6 0.0 0.0 0.0 0.0
Ectoprocta
Onychocellidae
Floridina antiqua 0.5 0.1 0.6 0.2 0.0 0.0 0.0 0.0
Annelida
Polychaeta, ni 1.5 0.3 1.8 0.5 0.0 0.0 0.0 0.0
Chordata 5.4 1.1 5.4 1.5 1.3 0.2 11.1 11.1
Urochordata
Ascidacea, ni 0.5 0.1 0.6 0.2 0.0 0.0 0.0 0.0
Clavellinidae:
Archidistoma pachecae 0.5 0.1 0.6 0.2 0.0 0.0 0.0 0.0
Teleostei
Teleostei, ni 3.0 0.5 3.6 0.9 0.0 0.0 0.0 0.0
Huevos de Pez 1.0 0.2 1.2 0.3 0.0 0.0 0.0 0.0
Kyphosidae 0.5 0.1 0.0 0.0 1.3 0.2 0.0 0.0
Labridae
continued on next page
ARMENDARIZ VILLEGAS ET AL.308

Molluscs were most abundant (52.7% FO), mainly in di-gestive tracts and accumulations, followed by arthropods
(crustaceans) (42.9%) and echiurans (22.2%). Similarly, mol-luscs were the most abundant phylum in the diet (46.3%N) andincluded the greatest number of prey categories, followed by
arthropods (crustaceans; 44.9%), and echiurans (5%; Table 1).Unidentified organic matter was the most common item in
the digestive tract (45.8% FO), which highlights the high
digestion state of prey. When considering all sampling tech-niques, the most frequent item was xanthid crabs (29%),followed by Echiurus spp. (22.2%), Megapitaria squalida
(20.2%), and, in minor amounts, Trigoniocardia biangulata,unidentified gastropods, Glycymeris gigantea, portunid crabs,Euvola vogdesi, and Glycymeris multicostata (Table 1).
Considering all sampling techniques, the xanthid crabs
represent a third of the diet (36% N), followed by the bivalveMegapitaria squalida, the worms Echiurus spp., Glycymerisgigantea, Trigoniocardia biangulata, Glycymeris multicostata,
and portunid crabs (Table 1).
Seasonal Changes in Diet Composition
During winter and autumn, a high frequency of octopuseshad food (considering all sampling techniques; 84% and 83%,respectively), whereas during summer only 55% had food. The
greatest abundance of food items from digestive tracts wasrecorded during autumn, from accumulations during winter,and from live prey during spring (Fig. 1). Significant differencesin FWI combined values were found between spring and
summer (Kruskal–Wallis, H ¼ 11.42, P < 0.05; Table 2).Considering all sampling techniques, seasonal changes in
diet composition were observed (Fig. 2). Xanthid crabs were
a main item consumed during all seasons, although withdifferent importance. During summer, in addition to xanthidcrabs (27% FO, 22% N) and echiuran worms (32% FO, 12%
N), octopuses also consumed portunid crabs, the concavescallop Euvola vogdesi, and fish in similar proportions. Duringautumn, xanthid crabs (22% FO, 28% N) and echiuran worms(32%FO, 10%N) were also important, but the consumption of
bivalves such as Megapitaria squalida (14% FO, 8% N) andTrigoniocardia biangulata (15% FO, 6% N) increased. During
winter, more prey was noted, but Echiurus spp. were notconsumed; xanthid crabs were important (26% FO, 26% N),but molluscs such as M. squalida, Glycymeris gigantea, T.
biangulata, and Dosinia ponderosa (all more than 24% FO)increased in the diet. In spring, the crabs dominated theOctopusbimaculatus diet completely—in particular, xanthid crabs (61%
N), which were present in almost half of all digestive tracts (47%FO). With regard to the unidentified organic matter, autumnhad the highest FO value (52%), followed by summer (46%),
winter (40%), and spring (30%).
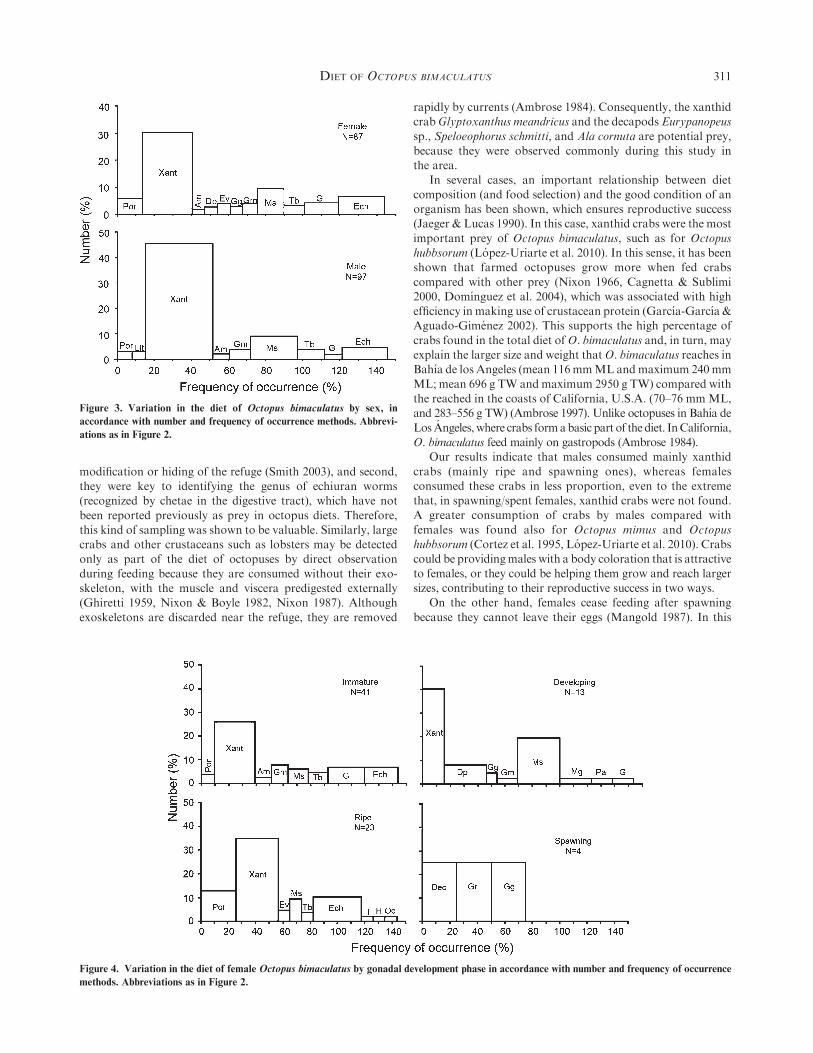
Diet Composition by Sex
The FWIs by sex are presented in Tables 2 and 3. Both,
males and females consumed xanthid crabs preferably; how-ever, the importance of secondary prey was different betweenthem. Females consumed, in order of importance, more
echiuran worms (24% FO, 7% N) followed by Megapitariasqualida (14% FO, 10% N), whereas males inverted them:M. squalida (25% FO, 9% N) followed by echiurans (24% FO,
5%N; Fig. 3). Crabs were consumedmore bymales than females,whereas bivalves were consumed less by males than females.
Diet Composition by Gonad Development Stage
Significant differences were found in FWIs by gonad de-velopment stage in females (Kruskal-Wallis, P¼ 0.14; Table 3).The greatest value was recorded for ripe females (0.53 ± 0.45),
and the lowest for spawning/spent females (0.02 ± 0.01). Nosignificant differences were found in FWIs for the gonaddevelopment stages of males.
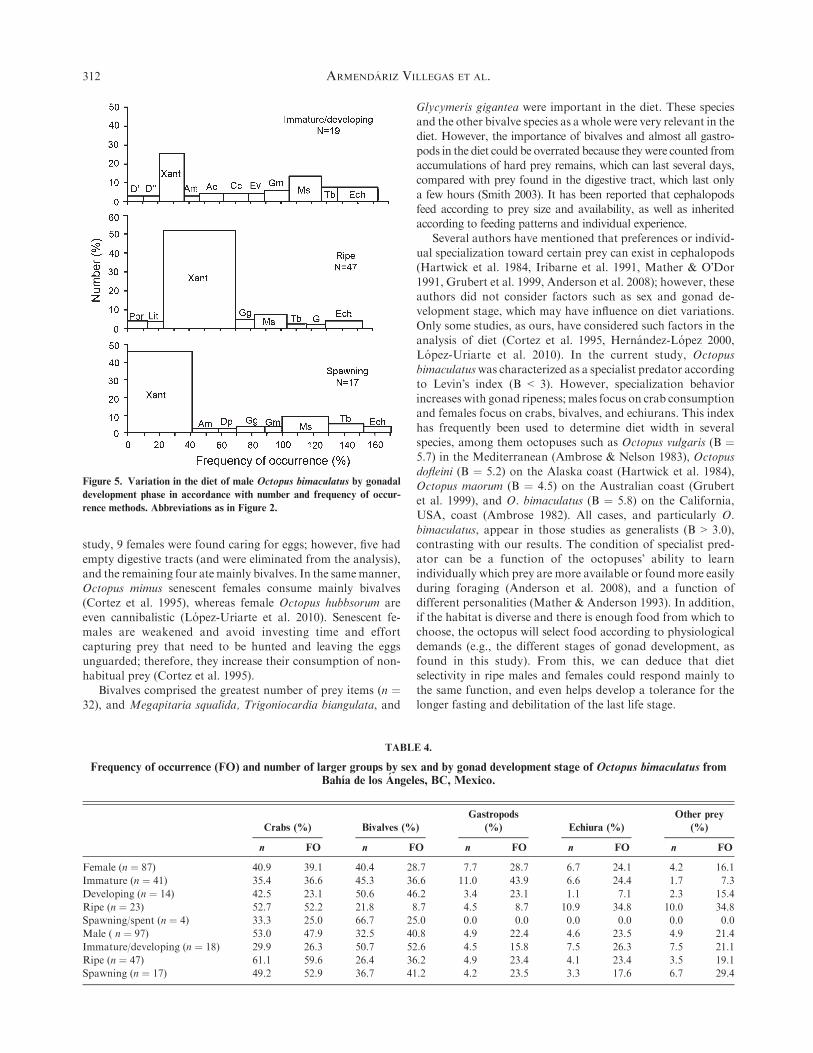
In general, a tendency toward increasing crab consumptionwith gonad development was observed, with the greatest crabconsumption by ripe octopuses (Figs. 4 and 5, and Table 4).
Bivalves seemed to have a greater importance to immature anddeveloping stages of both males and females, but their impor-tance diminished during the ripe stage. Consumption ofechiuran and ‘‘other prey’’ (such as fish, echinoderms, and
octopuses) was the maximum in ripe females. Immature femaleshad a diet composed mainly of xanthid crabs, gastropods,echiuran worms, and, in a lesser proportion, Megapitaria
squalida and Glycymeris multicostata bivalves (Fig. 4). De-veloping females consumed in number more xanthid crabs,but in frequency more M. squalida. Ripe females consumed
mainly xanthid and portunid crabs, and increased the con-sumption of echiurans. The spawning/spent females consumedless prey. Two females contained only unidentified organic
TABLE 1.
continued
Item
Total Digestive tract Midden Live prey
FO n FO n FO n FO n
Halichoeres nicholsi 0.5 0.1 0.0 0.0 0.0 0.0 11.1 11.1
UOM 38.9 0.0 45.8 0.0 0.0 0.0 0.0 0.0
Total general 296.6 100 187.4 100 350 100 100 100
Total values for larger groups are not indented in the left column. FO, frequency of occurrence; ni, not identified; UOM, unidentified organic matter.
TABLE 2.
Female, male, and combined fullness weight index by season.
Summer Autumn Winter Spring
Female 0.30 ± 0.47 0.42 ± 0.53 0.28 ± 0.41 0.36 ± 0.40
Male 0.08 ± 0.07 0.25 ± 0.36 0.31 ± 0.35 0.41 ± 0.37
Combined 0.20 ± 0.36 0.32 ± 0.44 0.29 ± 0.37 0.39 ± 0.38
DIET OF OCTOPUS BIMACULATUS 309

matter; one contained a unidentified decapod and one con-tained 2 bivalves (Glycymeris gigantea and Gari regularis).
Immature/developing males had the most diverse diet. Theirmost important prey were xanthids, Megapitaria squalida,echiurans, and bivalves such as Glycymeris multicostata and
Trigoniocardia biangulata (Fig. 5). The diet of ripe and spawn-ing males was based almost exclusively on xanthid crabs, andbivalves were secondary prey (mainly M. squalida). However,
bivalves were more important in spent males.
Diet Width
According to Levin�s index obtained in this study for thewhole sample (2.6), Octopus bimaculatus is a specialist con-sumer. The Levin�s index calculated for females was greater(2.8) than for males (2.2), but both are categorized as specialist
consumers. In the same manner, the Levin�s index obtained foreach gonad development stage (immature females, 2.3; devel-oping females, 1.4; ripe females, 1.7; immature/developing
males, 2.7; ripe males, 2.0; spawning males, 1.2), confirm thatO. bimaculatus is a specialist consumer.
DISCUSSION
Prey proportions can vary daily or seasonally, and thereforeoctopuses can exhibit feeding cycles that reflect this availability(Hanlon & Messenger 1998). In this sense, there are two
possible explanations for the significantly low FWIs found insummer. On one hand, they could be attributed to a fasterdigestion as a consequence of warmer temperatures (26–32�C)reported in Bahıa de Los Angeles during summer (Yee-Duarteet al. 2009); the complete digestion of stomach contents isestimated to last 16 hours at 14�C and 12 hours at 18–19�C(Boucher-Rodoni et al. 1987). On the other hand, a greaternumber was found during the summer in spawning/spentoctopuses (26% females and 31% males) compared with other
seasons (Castellanos-Martınez 2008), when it consumes lessfood. In this sense, it was found that ripe females had
significantly greater FWIs whereas spawning/spent femaleshad significantly lower FWIs than the other stages. It has beennoted that females decrease food ingestion considerably dur-
ing egg care (Mather 1980, Mangold 1987, Cortez et al. 1995,Lopez-Uriarte et al. 2010), although this decrease can be similarin males and females (Quetglas et al. 2005). Garcıa-Garcıa and
Aguado-Gimenez (2002) reported greater ingestion rates forOctopus vulgaris females before maturity. Mather (1980) alsomentions that physiological changes during female maturation
cause a notable increase in food ingestion 2–3 wk before egglaying, when females need more nutrients and energy for eggproduction.
In agreement with our results, it is better to use the combined
three sampling methods (digestive tract, accumulation of preyremains in the refuge, and observation of live prey) for dietanalysis in octopuses. The prey obtained with each technique
used was as expected—in accumulations, mainly molluscs; instomach contents, crabs. Samples of live prey (being eaten byoctopuses) were useful in two senses: first to confirm that the
bivalves found in accumulations were used not only for
Figure 2. Seasonal variation in the diet of Octopus bimaculatus in accordance with number and frequency of occurrence methods.
TABLE 3.
Female, male, and combined fullness weight index by gonad
development stage.
Immature Developing Ripe Spawning/Spent
Female 0.41 ± 0.52 0.25 ± 0.29 0.53 ± 0.45 0.02 ± 0.01
Immature/
Developing Ripe Spawning
Male 0.42 ± 1.01 0.41 ± 0.64 0.30 ± 0.34
ARMENDARIZ VILLEGAS ET AL.310
modification or hiding of the refuge (Smith 2003), and second,they were key to identifying the genus of echiuran worms(recognized by chetae in the digestive tract), which have not
been reported previously as prey in octopus diets. Therefore,this kind of sampling was shown to be valuable. Similarly, largecrabs and other crustaceans such as lobsters may be detectedonly as part of the diet of octopuses by direct observation
during feeding because they are consumed without their exo-skeleton, with the muscle and viscera predigested externally(Ghiretti 1959, Nixon & Boyle 1982, Nixon 1987). Although
exoskeletons are discarded near the refuge, they are removed
rapidly by currents (Ambrose 1984). Consequently, the xanthidcrabGlyptoxanthus meandricus and the decapods Eurypanopeus
sp., Speloeophorus schmitti, and Ala cornuta are potential prey,because they were observed commonly during this study inthe area.
In several cases, an important relationship between diet
composition (and food selection) and the good condition of anorganism has been shown, which ensures reproductive success(Jaeger & Lucas 1990). In this case, xanthid crabs were the most
important prey of Octopus bimaculatus, such as for Octopushubbsorum (Lopez-Uriarte et al. 2010). In this sense, it has beenshown that farmed octopuses grow more when fed crabs
compared with other prey (Nixon 1966, Cagnetta & Sublimi2000, Domınguez et al. 2004), which was associated with highefficiency in making use of crustacean protein (Garcıa-Garcıa &Aguado-Gimenez 2002). This supports the high percentage of
crabs found in the total diet ofO. bimaculatus and, in turn, mayexplain the larger size and weight thatO. bimaculatus reaches inBahıa de los Angeles (mean 116mmML andmaximum 240mm
ML; mean 696 g TW andmaximum 2950 g TW) compared withthe reached in the coasts of California, U.S.A. (70–76 mmML,and 283–556 g TW) (Ambrose 1997). Unlike octopuses in Bahıa de
Los Angeles,where crabs formabasic part of the diet. InCalifornia,O. bimaculatus feed mainly on gastropods (Ambrose 1984).
Our results indicate that males consumed mainly xanthid
crabs (mainly ripe and spawning ones), whereas femalesconsumed these crabs in less proportion, even to the extremethat, in spawning/spent females, xanthid crabs were not found.A greater consumption of crabs by males compared with
females was found also for Octopus mimus and Octopushubbsorum (Cortez et al. 1995, Lopez-Uriarte et al. 2010). Crabscould be providingmales with a body coloration that is attractive
to females, or they could be helping them grow and reach largersizes, contributing to their reproductive success in two ways.
On the other hand, females cease feeding after spawning
because they cannot leave their eggs (Mangold 1987). In this
Figure 3. Variation in the diet of Octopus bimaculatus by sex, in
accordance with number and frequency of occurrence methods. Abbrevi-
ations as in Figure 2.
Figure 4. Variation in the diet of female Octopus bimaculatus by gonadal development phase in accordance with number and frequency of occurrence
methods. Abbreviations as in Figure 2.
DIET OF OCTOPUS BIMACULATUS 311
study, 9 females were found caring for eggs; however, five had
empty digestive tracts (and were eliminated from the analysis),and the remaining four atemainly bivalves. In the samemanner,Octopus mimus senescent females consume mainly bivalves(Cortez et al. 1995), whereas female Octopus hubbsorum are
even cannibalistic (Lopez-Uriarte et al. 2010). Senescent fe-males are weakened and avoid investing time and effortcapturing prey that need to be hunted and leaving the eggs
unguarded; therefore, they increase their consumption of non-habitual prey (Cortez et al. 1995).
Bivalves comprised the greatest number of prey items (n ¼32), and Megapitaria squalida, Trigoniocardia biangulata, and
Glycymeris gigantea were important in the diet. These speciesand the other bivalve species as a whole were very relevant in the
diet. However, the importance of bivalves and almost all gastro-pods in the diet could be overrated because theywere counted fromaccumulations of hard prey remains, which can last several days,compared with prey found in the digestive tract, which last only
a few hours (Smith 2003). It has been reported that cephalopodsfeed according to prey size and availability, as well as inheritedaccording to feeding patterns and individual experience.
Several authors have mentioned that preferences or individ-ual specialization toward certain prey can exist in cephalopods(Hartwick et al. 1984, Iribarne et al. 1991, Mather & O�Dor
1991, Grubert et al. 1999, Anderson et al. 2008); however, theseauthors did not consider factors such as sex and gonad de-velopment stage, which may have influence on diet variations.Only some studies, as ours, have considered such factors in the
analysis of diet (Cortez et al. 1995, Hernandez-Lopez 2000,Lopez-Uriarte et al. 2010). In the current study, Octopusbimaculatuswas characterized as a specialist predator according
to Levin�s index (B < 3). However, specialization behaviorincreases with gonad ripeness; males focus on crab consumptionand females focus on crabs, bivalves, and echiurans. This index
has frequently been used to determine diet width in severalspecies, among them octopuses such as Octopus vulgaris (B ¼5.7) in the Mediterranean (Ambrose & Nelson 1983), Octopus
dofleini (B ¼ 5.2) on the Alaska coast (Hartwick et al. 1984),Octopus maorum (B ¼ 4.5) on the Australian coast (Grubertet al. 1999), and O. bimaculatus (B ¼ 5.8) on the California,USA, coast (Ambrose 1982). All cases, and particularly O.
bimaculatus, appear in those studies as generalists (B > 3.0),contrasting with our results. The condition of specialist pred-ator can be a function of the octopuses� ability to learn
individually which prey are more available or foundmore easilyduring foraging (Anderson et al. 2008), and a function ofdifferent personalities (Mather & Anderson 1993). In addition,
if the habitat is diverse and there is enough food from which tochoose, the octopus will select food according to physiologicaldemands (e.g., the different stages of gonad development, asfound in this study). From this, we can deduce that diet
selectivity in ripe males and females could respond mainly tothe same function, and even helps develop a tolerance for thelonger fasting and debilitation of the last life stage.
Figure 5. Variation in the diet of male Octopus bimaculatus by gonadal
development phase in accordance with number and frequency of occur-
rence methods. Abbreviations as in Figure 2.
TABLE 4.
Frequency of occurrence (FO) and number of larger groups by sex and by gonad development stage of Octopus bimaculatus fromBahıa de los Angeles, BC, Mexico.
Crabs (%) Bivalves (%)
Gastropods
(%) Echiura (%)
Other prey
(%)
n FO n FO n FO n FO n FO
Female (n ¼ 87) 40.9 39.1 40.4 28.7 7.7 28.7 6.7 24.1 4.2 16.1
Immature (n ¼ 41) 35.4 36.6 45.3 36.6 11.0 43.9 6.6 24.4 1.7 7.3
Developing (n ¼ 14) 42.5 23.1 50.6 46.2 3.4 23.1 1.1 7.1 2.3 15.4
Ripe (n ¼ 23) 52.7 52.2 21.8 8.7 4.5 8.7 10.9 34.8 10.0 34.8
Spawning/spent (n ¼ 4) 33.3 25.0 66.7 25.0 0.0 0.0 0.0 0.0 0.0 0.0
Male ( n ¼ 97) 53.0 47.9 32.5 40.8 4.9 22.4 4.6 23.5 4.9 21.4
Immature/developing (n ¼ 18) 29.9 26.3 50.7 52.6 4.5 15.8 7.5 26.3 7.5 21.1
Ripe (n ¼ 47) 61.1 59.6 26.4 36.2 4.9 23.4 4.1 23.4 3.5 19.1
Spawning (n ¼ 17) 49.2 52.9 36.7 41.2 4.2 23.5 3.3 17.6 6.7 29.4
ARMENDARIZ VILLEGAS ET AL.312

The high productivity near Bahıa de Los Angeles probablyprovidesOctopus bimaculatus with more food options. It shows
a selective behavior toward prey (crabs and bivalves), as theoptimum foraging theory predicts. When the food density ishigh, predators specialize on good-quality prey and ignore preywith lower food value (Schoener 1971).
ACKNOWLEDGMENTS
We thank Instituto Politecnico Nacional (SIP 20060858 and20070360) and Pronatura Noroeste for funding this work.
J. Armendariz-Villegas was a fellow student of PIFI (IPN) andCONACyT, and the results presented here are part of her MS
thesis. B. P. Ceballos-Vazquez, M. Arellano-Martınez, and A.Abitia-Cardenas received grants from SIBE (COFAA) andEDI (IPN). They and U. Markaida are members of SNI-CONACyT. We thank the following specialized personnel
who helped in prey identification: M. Dıaz and E. Gonzalez-Navarro (UABCS), and E. Felix-Pico, L. Burnes-Romo, andD.Herrero-Perezrul (CICIMAR-IPN). Thanks to Myla Young
and Emmanuel O. Vaughan for revising the English manuscript.
LITERATURE CITED
Ambrose, R. F. 1982. Shelter utilization by the molluscan cephalopod
Octopus bimaculatus. Mar. Ecol. Prog. Ser. 7:67–73.
Ambrose, R. F. 1983.Midden formation by octopuses: the role of biotic
and abiotic factors. Mar. Behav. Physiol. 10:137–144.
Ambrose, R. F. 1984. Food preferences, prey availability and the diet of
Octopus bimaculatus Verril. J. Exp. Mar. Biol. Ecol. 77:29–44.
Ambrose, R. F., M. A. Land & F. G. Hochberg. 1997. Octopus
bimaculatus. In:M. A. Land&F. G.Hochberg, editors. Proceedings
of the workshop on the fishery and market potential of octopus in
California. Washington, DC: Smithsonian Institution. pp. 11–22.
Ambrose, R. F. & B. Nelson. 1983. Predation byOctopus vulgaris in the
Mediterranean. Mar. Ecol. (Berl.) 4:251–261.
Anderson, R. C., J. Wood & J. A. Mather. 2008. Octopus vulgaris in
the Caribbean is a specializing generalist. Mar. Ecol. Prog. Ser.
371:199–202.
Barnard, J. L. & J. R. Grady. 1968. A biological survey on Bahıa de Los
Angeles. Gulf of California, Mexico: general account. San Diego
Soc. Nat. Hist. I15:51–66.
Boucher-Rodoni, R., E. Boucaud-Camou & K. Mangold. 1987. Feed-
ing and digestion. In: P. R. Boyle, editor. Cephalopod life cycle. Vol.
II: comparative reviews. London: Academic Press. pp. 85–108.
Brusca, R. C. 1980. Common intertidal invertebrates of the Gulf of
California, 2nd edition. Tucson, AZ: University of Arizona Press.
513 pp.
Brusca, R. C. & G. J. Brusca. 2005. Invertebrados. Madrid: McGraw
Hill-Interamericana. 1005 pp.
Bustos-Serrano, H., R. Millan-Nunez & R. Cajal-Medrano. 1996.
Efecto de la marea en la productividad organica primaria en una
laguna costera del Canal de Ballenas, Golfo de California. Cienc.
Mar. 22:215–233.
Cagnetta, P. & A. Sublimi. 2000. Productive performance of the
common octopus (Octopus vulgaris C.) when fed on a monodiet:
recent advances in Mediterranean aquaculture finfish species di-
versification. Zaragoza, Spain: CIHEAM. 47:331–336.
Cailliet, M. G., M. S. Love & A. W. Ebeling. 1986. Fishes: a field and
laboratory manual on their structure identification and natural
history. Belmont, CA: Wadsworth. 194 pp.
Cardoso, F., P. Villegas & C. Estrella. 2004. Observaciones sobre la
biologıa de Octopus mimus (Cephalopoda: Octopoda) en la costa
Peruana. Rev. Peru. Biol. 11:45–50.
Castellanos-Martınez, S. 2008. Biologıa reproductiva del pulpoOctopus
bimaculatus Verril, 1883 en Bahıa de los Angeles, Baja California,
Mexico. MS thesis. Instituto Politecnico Nacional. 98 pp.
Cherel, Y. & K. A. Hobson. 2005. Stable isotopes, beaks and predators:
a new tool to study ecology of cephalopods, including giant and
colossal squids. Proc. R. Soc. Lond. 272:1601–1607.
Cortez, T., B. G. Castro & A. Guerra. 1995. Feeding dynamics of
Octopus mimus (Mollusca: Cephalopoda) in northern Chile waters.
Mar. Biol. 123:497–503.
Delgadillo-Hinojosa, F., G. Gaxiola-Castro, J. A. Segovia-Zavala, A.
Munoz-Barbosa & M. V. Orozco-Borbon. 1997. The effect of
vertical mixing on primary production in a bay of the Gulf of
California. Estuar. Coast. Shelf Sci. 45:135–148.
Dodge, R. & D. Scheel. 1999. Remains of the prey, recognizing the
midden piles of Octopus dofleini (Wulker). Veliger 42:260–266.
Domınguez, P., G. Gaxiola-Cortes & C. Rosas-Vazquez. 2004. Ali-
mentacion y nutricion de moluscos cefalopodos: avances recientes y
perspectivas futuras. In: L. E. Cruz-Suarez, M. D. Ricque, M. G.
Nieto-Lopez, D. Villarreal, U. Scholz & M. Gonzalez, editors.
Avances en Nutricion Acuıcola. Memorias del VII Simposium
Internacional de Nutricion Acuıcola. Hermosillo, Mexico: Univer-
sidad de Sonora. pp. 16–19.
Fisher, W. K. 1946. Echiuroid worms of the North Pacific Ocean. Proc.
U.S. Nat. Mus. 96:215–292.
Garcıa-Garcıa, B. & F. Aguado-Gimenez. 2002. Influence of diet on
growing and nutrient utilization in the common octopus (Octopus
vulgaris). Aquaculture 211:173–184.
Ghiretti, F. 1959. Cephalotoxin: the crab-paralyzing posterior salivary
glands in the cephalopods. Nature 182:1192–1193.
Grubert, M. A., V. A. Wadley & R. W. White. 1999. Diet and feeding
strategy of Octopus maorum in southeast Tasmania. Bull. Mar. Sci.
65:441–451.
Guerra, A. 1992. Fauna Iberica, Vol. 1:Mollusca, Cephalopoda.Museo
Nacional de Ciencias Naturales. Madrid: Consejo Superior de
Investigaciones Cientıficas. 327 pp.
Hanlon, R. & J. B.Messenger. 1998. Cephalopod behavior. Cambridge:
University Press. 232 pp.
Hartwick, E. B., R. F. Ambrose & S. M. C. Robinson. 1984. Den
utilization and the movements of tagged Octopus dofleini. Mar.
Behav. Physiol. 11:95–110.
Heemstra, P. C. 1995. Serranidae. In: W. F. Fischer, W. Krup, C.
Schneider, K. Sommer, E. Carpenter & V. H. Niem, editors. Guıa
FAO para la identificacion de especies para los fines de la pesca. Vol.
III: Pacıfico Centro-Oriental. Rome: FAO. pp. 1565–1613.
Hendrickx, M. E. 1995. Cangrejos. In: W. F. Fischer, W. Krup, C.
Schneider, K. Sommer, E. Carpenter & V. H. Niem, editors. Guıa
FAO para la identificacion de especies para los fines de la pesca. Vol.
I: Pacıfico Centro-Oriental. Rome: FAO. pp. 565–636.
Hernandez-Lopez, J. L. 2000. Biologıa, ecologıa y pesca del pulpo
comun (Octopus vulgaris, Cuvier 1797) en aguas de Gran Canaria.
PhD diss., University Las Palmas Gran Canaria. 210 pp.
Hochberg, F. G. & W. G. Fields. 1980. Cephalopods: the squids and
octopuses. In: R.Morris, D. Abbot &E. Haderlie, editors. Intertidal
invertebrates of California. Stanford, CA: Stanford University
Press. pp. 429–444.
Hurlbert, S. H. 1978. The measurement of the niche overlap and some
relatives. Ecology 59:67–77.
Hyslop, E. J. 1980. Stomach content analysis: a review of methods and
their applications. J. Fish Biol. 17:411–429.
Iribarne, O. O., M. E. Fernandez &H. Zucchini. 1991. Prey selection by
the small Patagonian octopusOctopus tehuelchusD�Orbigny. J. Exp.
Mar. Biol. Ecol. 148:271–281.
DIET OF OCTOPUS BIMACULATUS 313

Jaeger, R. G. & J. Lucas. 1990. On evaluation of foraging strategies
through estimates of reproductive success. In: R. Hughes, editor.
Behavioral mechanisms of food selection. Alemania: Springer-
Verlag. pp. 83–94.
Joll, L. M. 1976. Mating, egg-laying and hatching of Octopus tetricus
(Mollusca: Cephalopoda) in the laboratory. Mar. Biol. 36:327–333.
Keen, M. 1971. Sea shells of tropical west America: marine mollusks
from California to Peru. Stanford, CA: Stanford University Press.
1064 pp.
Kerstitch, A. N. &H. Bertsch. 2007. Sea of Cortez marine invertebrates:
a guide for the Pacific coast, Mexico to Peru. Monterey, CA: Sea
Challengers. 124 pp.
Lopez-Uriarte, E., E. Rios-Jara & M. E. Gonzalez-Rodrıguez. 2010.
Diet and feeding habits of Octopus hubbsorum Berry, 1953 in the
Central Mexican Pacific. Veliger 51:26–42.
Mangold, K. 1987. Reproduction. In: P. Boyle, editor. Cephalopod life
cycles. London: Academic Press. pp. 157–200.
Mather, J. 1980. Some aspects of food intake inOctopus joubiniRobson.
Veliger 22:286–290.
Mather, J. A. & R. C. Anderson. 1993. Personalities of octopuses
(Octopus rubescens). J. Comp. Psychol. 107:336–340.
Mather, J. A. & R. K. O�Dor. 1991. Foraging strategies and predation
risk shape the natural history of juvenileOctopus vulgaris.Bull.Mar.
Sci. 49:256–269.
Nixon,M. 1966. Changes on bodyweight and intake of food byOctopus
vulgaris. J. Zool. 150:1–9.
Nixon, M. 1987. Cephalopod diets. In: P. R. Boyle, editor. Cephalopod
life cycle. Vol. II: comparative reviews. London: Academic Press.
pp. 201–217.
Nixon, M. & P. R. Boyle. 1982. Hole-drilling in crustaceans by Eledone
cirrhosa (Mollusca: Cephalopoda). J. Zool. 196:439–444.
O�Dor, R.K. &M. J.Wells. 1978. Reproduction versus somatic growth:
hormonal control in Octopus vulgaris. J. Exp. Biol. 77:15–31.
Olivares-Paz, A., M. Zamora-Covarrubias, P. Portilla-Reyes & O.
Zuniga-Romero. 2001. Estudio histologico de la ovogenesis y
maduracion ovarica en Octopus mimus (Cephalopoda: Octopodi-
dae) de la II region de Chile. Estud. Oceanol. 20:13–22.
Poutiers, J. M. 1995. Gasteropodos. In: W. F. Fischer, W. Krup, C.
Schneider, K. Sommer, E. Carpenter & V. H. Niem, editors. Guıa
FAO para la identificacion de especies para los fines de la pesca. Vol.
I: Pacıfico Centro-Oriental. Rome: FAO. pp. 223–297.
Quetglas, A., F. Alemany, A. Carbonell, P. Merella & P. Sanchez. 1998.
Biology and fishery of Octopus vulgaris Cuvier, 1797, caught by
trawlers in Mallorca (Balearic Sea, western Mediterranean). Fish.
Res. 36:237–249.
Quetglas, A., M. Gonzalez & I. Franco. 2005. Biology of the upper-
slope cephalopod Octopus salutii from the western Mediterranean
Sea. Mar. Biol. 146:1131–1138.
Rodrıguez-Rua, A., I. Pozuelo, M. A. Prado, M. J. Gomez & M. A.
Bruzon. 2005. The gametogenic cycle ofOctopus vulgaris (Mollusca:
Cephalopoda) as observed on the Atlantic coast of Andalusia (south
of Spain). Mar. Biol. 147:927–933.
Schoener, T. W. 1971. Theory of feeding strategies. Annu. Rev. Ecol.
Syst. 11:396–404.
Smale, M. J. & P. R. Buchan. 1981. Biology of Octopus vulgaris off the
east coast of South Africa. Mar. Biol. 65:1–12.
Smith, C. D. 2003. Diet of Octopus vulgaris in False Bay, South Africa.
Mar. Biol. 143:1127–1133.
Sommer, C. 1995. Kyphosidae. In: W. F. Fischer, W. Krup, C.
Schneider, K. Sommer, E. Carpenter & V. H. Niem, editors. Guıa
FAO para la identificacion de especies para los fines de la pesca. Vol.
II: Pacıfico Centro-Oriental. Rome: FAO. pp. 1195–1200.
Villanueva, R. 1993. Diet andmandibular growth ofOctopus magnificus
(Cephalopoda). South Afr. J. Mar. Sci. 13:121–126.
Wodinsky, J. 1978. Feeding behaviour of broody female Octopus
vulgaris. Anim. Behav. 26:803–813.
Yee-Duarte, J. A., B. P. Ceballos-Vazquez & M. Arellano-Martınez.
2009. Variacion de los ındices morfofisiologicos de la almeja mano
de leon Nodipecten subnodosus (Sowerby, 1835), en Bahıa de los
Angeles, B.C., Golfo de California. Oceanides 24:91–99.
Zamora, C. M. & P. A. Olivares. 2004. Variaciones bioquımicas e
histologicas asociadas al evento reproductivo de la hembra de
Octopus mimus (Mollusca: Cephalopoda). Int. J. Morphol. 22:207–
216.
ARMENDARIZ VILLEGAS ET AL.314




















