
Integumental amino acid uptake in a carnivorous predator mollusc ( Sepia officinalis, Cephalopoda
10
Integumental amino acid uptake in a carnivorous predator mollusc (Sepia officinalis, Cephalopoda) M. de Eguileor 1 , M.G. Leonardi 2 , A. Grimaldi 1 , G. Tettamanti 1 , L. Fiandra 2 , B. Giordana 2 , R. Valvassori 1 , G. Lanzavecchia 1 Abstract. The epithelial cells of the integument of body, arms and tentacles of Sepia officinalis present on their apical membrane a well-organised brush border and show the morphological and histochemical characteristics of a typical absorptive epithelium. The ability of the integument to absorb amino acids was investigated both in the arms incubated in vitro and in a purified preparation of brush border membrane vesicles (BBMV). Autoradiographic pictures of the integument after incubation of the arms in sea-water with or without sodium, showed that proline intake was Na + -dependent, whereas leucine intake appeared to be a largely cation- independent process. Time course experiments of labelled leucine, proline and lysine uptakes in BBMV evidenced that these amino acids are accumulated within the vesicles in the presence of an inwardly directed sodium gradient. The sodium-driven accumulation proves that cationic and neutral amino acids are taken up by the apical membrane of the epithelium of Sepia integument through a secondary active mechanism. For leucine, a 90% inhibition of the uptake was recorded in the presence of a large excess of the substrate. In agreement with the autoradiography results, an analysis of the cation specificity transport in BBMV showed that leucine uptake had a low cation specificity, whereas lysine and proline uptakes were Na + -dependent. An excess of lysine and proline, which share with alanine two different transport systems in the gill epithelium of marine bivalves, reduced eucine uptake. The possible role of the absorptive ability of the integument in a carnivorous mollusc is discussed. © 2000 Harcourt Publishers Ltd Keywords: amino acid uptake, brush border membrane vesicles, epidermal cells, arms, cuttlefish. Introduction The epithelial tissues of marine invertebrates are adapted to absorb dissolved organic material (DOM) across the body surface as shown by the ubiquitous presence of microvilli in direct contact with sea water (Preston, 1990). Amino acid absorption by the integument has been demonstrated in many phyla and different classes of soft-bodied marine invertebrates (Southward & Southward, 1968, 1980; Wright & Pajor, 1989; Preston, 1993). The transport of amino acids and other small organic compounds by epidermal cells may represent an essential contribution in nutrition (Wright et al., 1987; Stephens, 1988; Rice & Stephens, 1988; Wright & Manahan, 1989; Wright & Pajor, 1989; Preston, 1990;) and play a key role in cell volume regulation. It is well known that most marine invertebrates, which are osmocon- formers, maintain the elevate osmolality within cells and body fluids by means of a high level of small organic 1 DBSF University of Insubria Via J.H. Dunant 3,21100 Varese, Italy 2 Department of Biology, University of Milan Via Celoria 26,20133 Milano, Italy Received 22 November 1999 Accepted 12 July 2000 Correspondence to: Magda de Eguileor, DBSF-University of Insubria Via J.H. Dunant 3, 21100 Varese, Italy. Tel.: +39 0332 421310; Fax +39 0332 421300; E-mail: [email protected] 389 Tissue & Cell Tissue & Cell, 2000 32 (5) 389–398 © 2000 Harcourt Publishers Ltd doi: 10.1054/tice.2000.0127, available online at http://www.idealibrary.com
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Integumental amino acid uptake in a carnivorous predator mollusc ( Sepia officinalis, Cephalopoda

Integumental amino acid uptake in acarnivorous predator mollusc (Sepiaofficinalis, Cephalopoda)
M. de Eguileor1, M.G. Leonardi2, A. Grimaldi1, G. Tettamanti1,L. Fiandra2, B. Giordana2, R. Valvassori1, G. Lanzavecchia1
Tissue&Cell
Tissue & Cell, 2000 32 (5) 389–398© 2000 Harcourt Publishers Ltddoi: 10.1054/tice.2000.0127, available online at http://www.idealibrary.com
Abstract. The epithelial cells of the integument of body, arms and tentacles of Sepia officinalis present on theirapical membrane a well-organised brush border and show the morphological and histochemical characteristics ofa typical absorptive epithelium. The ability of the integument to absorb amino acids was investigated both in thearms incubated in vitro and in a purified preparation of brush border membrane vesicles (BBMV).Autoradiographic pictures of the integument after incubation of the arms in sea-water with or without sodium,showed that proline intake was Na+-dependent, whereas leucine intake appeared to be a largely cation-independent process. Time course experiments of labelled leucine, proline and lysine uptakes in BBMVevidenced that these amino acids are accumulated within the vesicles in the presence of an inwardly directedsodium gradient. The sodium-driven accumulation proves that cationic and neutral amino acids are taken up bythe apical membrane of the epithelium of Sepia integument through a secondary active mechanism. For leucine,a 90% inhibition of the uptake was recorded in the presence of a large excess of the substrate. In agreement withthe autoradiography results, an analysis of the cation specificity transport in BBMV showed that leucine uptakehad a low cation specificity, whereas lysine and proline uptakes were Na+-dependent. An excess of lysine andproline, which share with alanine two different transport systems in the gill epithelium of marine bivalves, reduced eucine uptake. The possible role of the absorptive ability of the integument in a carnivorous mollusc is discussed.© 2000 Harcourt Publishers Ltd
Keywords: amino acid uptake, brush border membrane vesicles, epidermal cells, arms, cuttlefish.
Introduction
The epithelial tissues of marine invertebrates are adapted toabsorb dissolved organic material (DOM) across the bodysurface as shown by the ubiquitous presence of microvilli in
1DBSF University of Insubria Via J.H. Dunant 3,21100 Varese, Italy2Department of Biology, University of Milan Via Celoria 26,20133 Milano, Italy
Received 22 November 1999Accepted 12 July 2000
Correspondence to: Magda de Eguileor, DBSF-University of Insubria Via J.H.Dunant 3, 21100 Varese, Italy. Tel.: +39 0332 421310; Fax +39 0332421300; E-mail: [email protected]
direct contact with sea water (Preston, 1990). Amino acidabsorption by the integument has been demonstrated inmany phyla and different classes of soft-bodied marineinvertebrates (Southward & Southward, 1968, 1980; Wright& Pajor, 1989; Preston, 1993). The transport of amino acidsand other small organic compounds by epidermal cells mayrepresent an essential contribution in nutrition (Wright etal., 1987; Stephens, 1988; Rice & Stephens, 1988; Wright& Manahan, 1989; Wright & Pajor, 1989; Preston, 1990;)and play a key role in cell volume regulation. It is wellknown that most marine invertebrates, which are osmocon-formers, maintain the elevate osmolality within cells andbody fluids by means of a high level of small organic
389

390 DE EGUILEOR ET AL.
compounds instead of inorganic osmolytes, since the formerare far less perturbing to the functional activity of proteins(Somero & Bowlus, 1983; Kinne, 1993).
Sepia officinalis is an active carnivorous predator as areall other Cephalopods and it lives, buried in the sand whennot seeking prey (Boucaud-Camou & Boucher-Rodoni,1983). The integument of cuttlefish presents a well organ-ised brush border on the body surface (Brocco, 1976;Singley, 1982; Packard, 1988) and it seems feasible, assuggested by Preston (1990) for epithelial tissues withmicrovilli, that these animals absorb organic compoundsdirectly from the environment, although the physiologicalrole of such ability in animals with an extensive digestivesystem, a closed circulatory system and a well-developedcentral nervous system is not immediately evident.
In the present work we provide evidence that a welldeveloped brush border is present also in the integumentalepithelial cells of arms and tentacles, and, on the basis of ahistochemical and immunohistochemical analysis, that boththe epithelium and the brush border have the conventionalfeatures typical of absorptive epithelia. Autoradiographicpictures of the integument, after incubation of the arms invitro with labelled leucine or proline in normal or sodium-free sea water, strongly suggested the ability of the epithe-lium to absorb the two amino acids with a differentsensitivity to the presence of Na+. To substantiate thisfinding, we characterised leucine and proline uptakes in apurified preparation of vesiculated brush border membranes(BBMV) obtained from the integument. BBMV represent auseful tool to study transport mechanisms, since it makes itpossible to separate the transport phenomenon from themetabolic event, and, due to the complete control of thecomposition of the internal and external compartments,allow a better understanding of transport processes. Theresults obtained with BBMV demonstrated that the integu-ment of Sepia officinalis is able to accumulate neutral andcationic amino acids via at least two transport systems withdifferent cation-sensitivity.
Materials and methods
Fifty specimens of Sepia officinalis, 3 months old, werekept in artificial sea water at 18°C and fed daily with liveshrimps.
Light microscopy and transmission electronmicroscopyFor routine transmission electron microscopy, anaesthetisedcuttlefishes were sectioned and fixed for 2 h in 2%glutaraldehyde in sea water. Specimens were washed in seawater and post fixed for 2 h with 1% osmic acid in cacody-late buffer (pH 7.2). After standard dehydration in anethanol series, specimens were embedded in an Epon-Araldite 812 mixture. Sections were cut with ReichertUltracut E ultrotome (Leica, Nussloch, Germany). Semithin
sections were stained by conventional methods (crystalviolet and basic fuchsin; Moore et al., 1960) and observedwith an optical microscope (Olympus, Tokyo, Japan). Thinsections were stained by uranyl acetate and lead citrate andwere observed with a Jeol 1010 electron microscope (Jeol,Tokyo, Japan).
HistochemistryArms and tentacles from anaesthetised cuttlefishes weredissected in 5 mm blocks. The specimens were rapidlyfrozen in liquid nitrogen. Cryosections (10 µm) wereobtained with a Reichert-Jung Frigocut 2800. The slideswere immediately used or stored at –20°C. The sectionswere stained according to Dubowitz & Brooke (1973) witha histoenzymatic kit (Bio-Optica, Milan, Italy). Variouscytoplasmic characteristics were assayed using severalhistochemical procedures: succinic acid dehydrogenasereaction (SDH); NADH-tetrazolium reductase (NADH-TR)and ATPase attributable to mitochondrial activity (i.e. to thesource of energy in cell metabolism); alkaline phosphatase(ALP), whose distribution is commonly close related withabsorptive function (Richards & Arme, 1982). For allreactions, controls involving the omission of the substratewere carried out. In all cases, negative results wereobtained, thus proving the specific nature of the reactions.Each experiment was repeated three times.
ImmunocytochemistryCryosections (10 µm), incubated for 15 min with a solutionof 0.05 M NH4Cl to reduce auto-fluorescence, were washedwith phosphate-buffered saline solution (PBS) and thenincubated with the primary antisera (diluted 1:200) for30 min. The primary antibodies used were: (mouse) anti-human Alkaline Phosphatase (ALP) (clone 8B6) (SigmaImmunochemicals, St. Louis, MO) and (rabbit) anti-Na+/K+
ATPase α-1 (Upstate biotechnology, Lake Placid, NY).After incubation with the primary antibodies, washed speci-mens were incubated with the appropriate IgG antisera,tetramethylrhodamine isothiocyanate (TRITC) conjugated(diluted 1:100) (from Jackson, Immuno ResearchLaboratories, West Grove, PA), as a secondary antibody, for1 h in a dark moist chamber. The PBS buffer used forwashing and dilution of antibodies contained 2% bovineserum albumin (BSA).
Coverslips were mounted in Vectashield MountingMedium for fluorescence (Vector Laboratories,Burlingame, CA) and examined with a confocal lasermicroscope (laser λ, 568 nm for rhodamine) (MRC 1024,Bio-Rad Laboratories, CA) using X40 and X63 objectives(NA 1.30, 1.25). Confocal images were superimposed usingPhotoshop 5.0.
Incubation of arms and autoradiographyArms were ligated, severed from the animal and incubatedat 15°C in an artificial sea water with the following compo-sition: 460 mM NaCl, 10 mM KCl, 10 mM CaCl2, 27 mMMgSO4, 0.5 µM [3H] leucine 1 µCi/ml or 0.5 µM [3H]

AMINO ACID UPTAKE IN CUTTLEFISH INTEGUMENT 391
proline 1 µCi/ml. In the experiments without Na+, NaCl wasomitted and replaced by 920 mM sucrose. The incubationmedium was aerated and stirred by bubbling 100% O2. Afteran incubation of 30 min, the arms were washed twice inunlabelled artificial sea water with or without Na+, lightlyblotted and immediately frozen in liquid nitrogen.
Frozen sections of all specimens were cut at 12 µm thick-ness and collected on slides. These slides were coated withKodak fine-grain Autoradiography Stripping Plate AR 10emulsion, air-dried overnight in the dark and then stored at4°C for 3–5 weeks in an airtight lightproof box with silicagel dessicator. Slides were developed in a Kodak D-19developer, stained with hematoxylin or eosin and mountedin Permount.
Control slides containing specimens that had not beenexposed to radioactivity were treated in the same manner.
Isolation of integument, preservation and preparationof BBMVThe arm integument of anaesthetised cuttlefishes wasscraped, lightly blotted on filter paper, weighed, placed incryotubes, frozen in liquid nitrogen and then stored. At themoment of use, the cryotubes were kept in a 37°C waterbath until the tissue began to melt. BBMV were prepared byCa++-precipitation and differential centrifugation accordingto Giordana et al., 1982. The tissue was homogenised in 100mM mannitol, 10 mM Hepes-Tris at pH 7.6 and the finalpellet was resuspended by 10 passes through a 22 gaugeneedle in 600 mM mannitol, 10 mM Hepes-Tris at pH 7.6.The membrane protein concentration was assessed with theCoomassie Brilliant Blue G-250 protein assay (Pierce), withbovine serum albumin as standard, and adjusted to a finalconcentration of 2 mg/ml. The protein yield in homogenateand BBMV from 1 g of fresh tissue was respectively 21.00± 1.35 mg and 0.34 ± 0.04 mg (3 determinations). Thepurity of the vesicle preparation was assessed by measuringthe activity of the basolateral and brush border membranemarker enzymes in the crude homogenate and in the finalpellet: Na+/K+-ATPase (EC 3.6.1.3) was assayed accordingto Quigley and Gotterer (1969) and alkaline phosphatase(EC 3.1.3.1) was determined by measuring the release of p-nitrophenol from p-nitrophenylphosphate. Incubations werecarried out in a spectrophotometer (Ultrospec 3000Pharmacia Biotech, Cambridge, UK) with a thermostatised(25°C) cuvette holder, under conditions in which theactivity was proportional to protein concentrations and time.
Transport experimentsTransport experiments were performed in triplicate at 20 °Cby mixing 10 µl of the vesicle suspension with 10 µl of theradiolabeled incubation mixture, whose final composition isreported in the legends to the figures. At selected times, 20µl samples were withdrawn from the incubation mixture,diluted with 2 ml of ice-cold stop solution (80 mMmannitol, 400 mM NaCl, 10 mM Hepes-Tris at pH 7.6),filtered through a 0.45 µm pore size cellulose-nitrate filter
(MFS Micro Filtration System) and rapidly washed with 3ml of ice-cold stop solution. The filters were then dissolvedin a scintillation mixture (Ultima Gold, Packard) and theradioactivity counted in a liquid scintillation spectrometer(Packard, Tri-Carb model 300C). To measure the initialrates of amino acid transport, incubations lasting 10 s wereperformed in quadruplicate with an automated device.
All the figures report a typical experiment, but eachexperiment was repeated at least three times with differentBBMV preparations.
Results
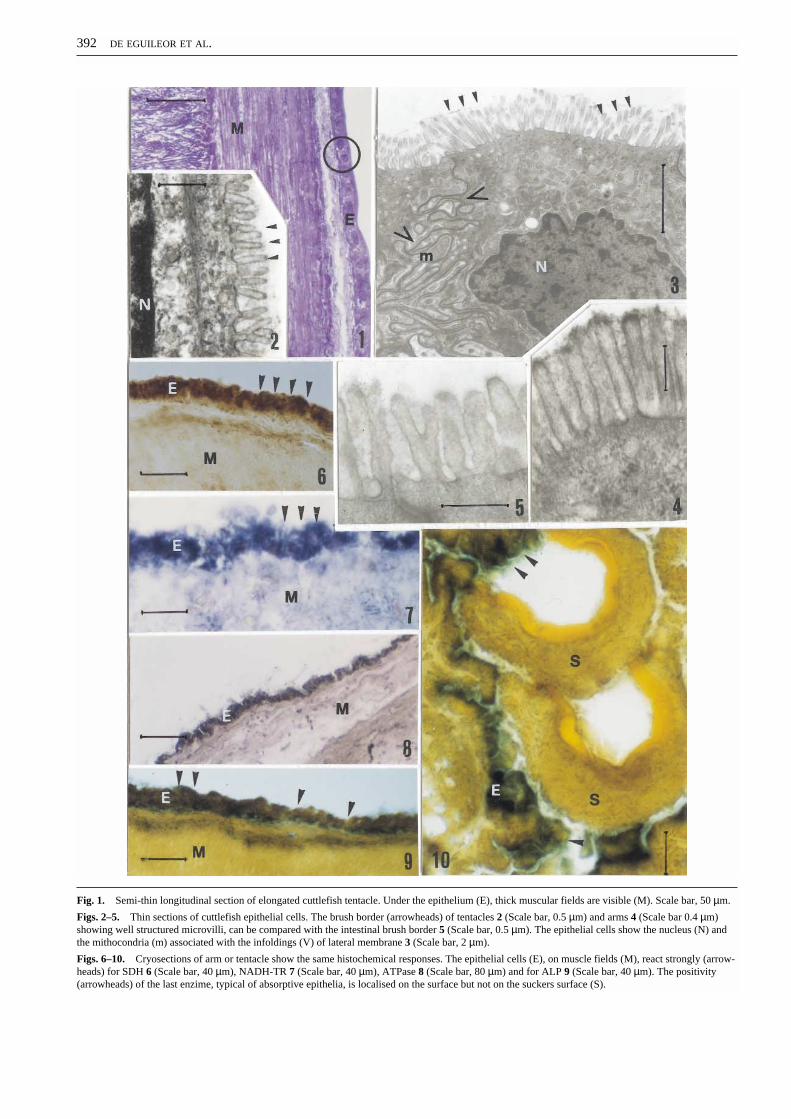
The epithelium of the integument in cuttlefish body, as justdescribed by Brocco (1976), Singley (1982) and Packard(1988), is characterised on the free margin by a brush borderof closely pressed microvilli. We found this specialisationalso on arms and tentacles (Figs. 1–4). It is important toremember that both arms and tentacles, used in preycapture, are structured in analogous manner with the differ-ence concerning the presence or not of a circular musclelayer (Kier, 1982, 1991). In these two structures, themicrovillar distribution and organisation were ratherdifferent: in the former case microvilli were closely resem-bling those present on the body and the intestinal wall (Figs.4–5) and in the latter case, microvilli were short and sparse(Fig. 2). Moreover the epidermal cells showed, in additionto the apical microvillar surface, the lateral side infoldingsclosely associated with mitochondria (Fig. 3).
In all cases (arm and tentacle surfaces) the epithelial cellsshowed the same histochemical picture with a strong posi-tivity for SDH (Fig. 6), NADH-TR (Fig. 7) and ATPase(Fig. 8) suggesting a high mitochondrial activity. In additiona strong positivity was evident for alkaline phosphatasemarking the apical portion of the epithelia (as typicallyobserved in many absorptive epithelia) but not thearm/tentacle sucker epithelia (Figs. 9–10).
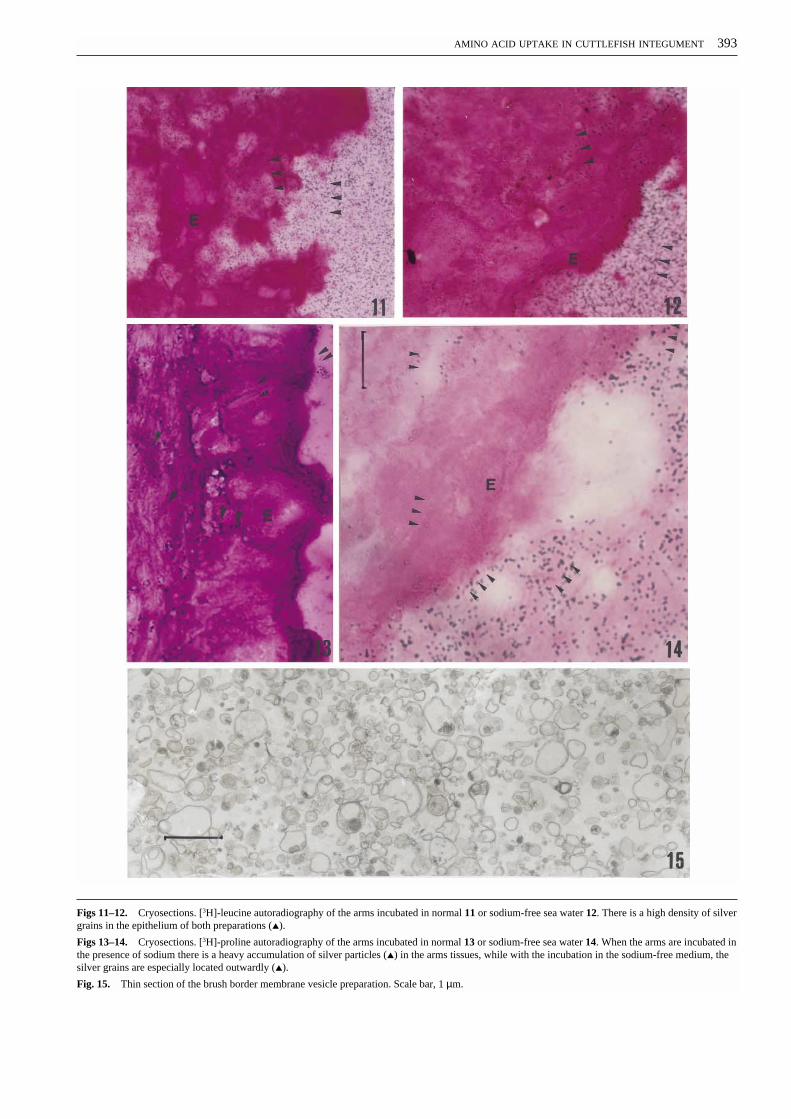
To establish if the integument of Sepia officinalis canabsorb amino acids from the environment, we performedexperiments with arms ligated, severed and isolated in vitro.The arms were incubated with 0.5 µM labelled leucine orproline in normal or Na+ free sea water. The autoradiog-raphy shows that leucine is taken up by the integument andcrosses the epithelial layer both in the presence (Fig. 11) andin the absence of sodium (Fig. 12), whereas for proline aheavy accumulation of silver particles in the tissue occurredonly in specimens incubated in the presence of sodium(Figs. 13, 14).
To extend the study on the ability of the epithelium toabsorb amino acids by means of Na+-dependent and Na+-independent processes, as observed in the intact integument,we prepared brush border membrane vesicles from thistissue by Ca++-precipitation. The final preparation is shownin Fig. 15. The purity of this preparation was evaluated fromthe enrichment factor (ratio of the enzyme specific activity
392 DE EGUILEOR ET AL.
Fig. 1. Semi-thin longitudinal section of elongated cuttlefish tentacle. Under the epithelium (E), thick muscular fields are visible (M). Scale bar, 50 µm.
Figs. 2–5. Thin sections of cuttlefish epithelial cells. The brush border (arrowheads) of tentacles 2 (Scale bar, 0.5 µm) and arms 4 (Scale bar 0.4 µm)showing well structured microvilli, can be compared with the intestinal brush border 5 (Scale bar, 0.5 µm). The epithelial cells show the nucleus (N) andthe mithocondria (m) associated with the infoldings (V) of lateral membrane 3 (Scale bar, 2 µm).
Figs. 6–10. Cryosections of arm or tentacle show the same histochemical responses. The epithelial cells (E), on muscle fields (M), react strongly (arrow-heads) for SDH 6 (Scale bar, 40 µm), NADH-TR 7 (Scale bar, 40 µm), ATPase 8 (Scale bar, 80 µm) and for ALP 9 (Scale bar, 40 µm). The positivity(arrowheads) of the last enzime, typical of absorptive epithelia, is localised on the surface but not on the suckers surface (S).
AMINO ACID UPTAKE IN CUTTLEFISH INTEGUMENT 393
Figs 11–12. Cryosections. [3H]-leucine autoradiography of the arms incubated in normal 11 or sodium-free sea water 12. There is a high density of silvergrains in the epithelium of both preparations (▲).
Figs 13–14. Cryosections. [3H]-proline autoradiography of the arms incubated in normal 13 or sodium-free sea water 14. When the arms are incubated inthe presence of sodium there is a heavy accumulation of silver particles (▲) in the arms tissues, while with the incubation in the sodium-free medium, thesilver grains are especially located outwardly (▲).
Fig. 15. Thin section of the brush border membrane vesicle preparation. Scale bar, 1 µm.

394 DE EGUILEOR ET AL.
in the final vesicle suspension and in the homogenate) of thetwo enzymes alkaline phosphatase and Na+/K+-ATPasespecific markers for the apical plasma membrane and forbasolateral membrane, respectively, in absorptive epithelia.Their presence in these specific areas of the epithelialmembrane in the integument of Sepia officinalis is demon-strated by their immunochemical localisation (Figs. 16–18).Table 1 shows that the activity of alkaline phosphatase, butnot of Na+/K+-ATPase, is increased in BBMV in compar-ison with that in homogenate, and confirms that the finalfraction is enriched in brush border membranes.
A typical time course of the uptake of 0.5 µM leucineinto BBMV in the presence of an inwardly directed Na+-gradient is reported in Fig. 19. Leucine was translocated andtransiently accumulated within the vesicles. The amino acidaccumulation value (ratio between the maximal and thefinal uptake values) was only fourfold: possibly this value isunderestimated, if the equilibrium value was not yet reachedafter 120 min. Longer incubation times could not beperformed because the transport activity, measured as initialrates, was appreciably decreased in BBMV maintained atroom temperature for longer times (data not shown).
Lysine and proline uptakes with time were also measuredin the presence of a sodium gradient. Fig. 20 shows thatboth lysine and proline were accumulated within the vesi-cles, even if in both cases the occurrence of the maximalpeak was delayed compared to that of leucine and the equi-librium value was clearly not yet reached after 120 min.Even if the height and the shape of the time course curvesare strictly linked to the kinetic characteristics of the trans-port system (Leonardi et al., 1993), other membrane proper-ties such as a low passive permeability for the cation, soluteand/or water, as observed in bivalves (Wright et al., 1987;Pajor & Wright, 1989), may cause a delay in the attainmentof the maximum value.
Sepia officinalis, as several marine species (Preston,1990; Kinne, 1993), presents a high cellular amino acidconcentration (Robertson, 1965), therefore amino aciduptake in vivo has to be performed against a steep chemicalgradient (Stephens, 1988; Wright & Pajor, 1989; Preston,1990, 1993) so that amino acid accumulation has to be ener-gised by the downhill gradient of a driver cation. The speci-ficity for the driver cation was tested by measuring leucine,lysine and proline initial uptake rates in the presence of aLi+-, Na+- or K+-gradient. The cation specificity of the trans-port agency involved in leucine uptake is poor (Fig. 21),since both lithium and potassium can efficiently substitutefor sodium in catalysing leucine uptake. On the contrary,proline initial rate was significantly higher in the presenceof Na+ compared to that in the presence of Li+ or K+ (Fig.21) and lysine uptake appears to require Na+, even if lithiumand potassium can partially support the amino acid uptake(Fig. 21).
In the gills of Mytilus edulis the uptake of neutral aminoacids occurs via two distinct transport systems: an alanine-lysine (AK) pathway with a poor cation selectivity and analanine-proline (AP) pathway with a high cation selectivity
(Wright & Pajor, 1989). Therefore, the inhibition inducedby a large excess (1 mM) of lysine and proline on 0.5 µMleucine uptake at 10 s was tested (Fig. 22). Lysine caused a42% and proline a 22% inhibition of leucine uptake (percentinhibitions were referred to the control uptake, 8.32 ± 0.10pmol/10 s/mg protein, after subtraction of the residualuptake in the presence of 1 mM leucine, 0.64 ± 0.01pmol/10 s/mg protein). Therefore these two amino acidsseem to share with leucine the same transport system, evenif the inhibition of leucine transport by the two amino acidsin Sepia officinalis is far less remarkable than that exertedon alanine in M. edulis (Pajor & Wright, 1989; Wright &Pajor, 1989).
DISCUSSION
The morphological and immunohistochemical characterisa-tion of Sepia officinalis integument carried out in thepresent study has evidenced that the epithelial cells possessthe classical features of absorptive epithelia (Figs. 1–10). Asstressed by Preston (1990), the epithelial tissues specificallyadapted for absorption invariably have microvilli, and thepositive histochemical responses obtained in cuttlefish armsand tentacles are comparable to those observed in thoseepithelia in which passage of nutritive substances regularlyoccurs, i.e. the gut epithelium of invertebrate and vertebrate(Barnet & Munday, 1972; Adibi & Mecer, 1973), theexternal integument of parasitic helminths (Mettrick &Podesta, 1974), adult/young adhering zones of broodingleeches (de Eguileor et al., 1994), the larvae ofStrongylocentrotus (Davis et al., 1985), the gutless organ-isms such as the pogonophora (Southward & Southward,1968, 1980) (Figs. 6–10).
The presence of an extensive array of microvilli on thebody surface is a rather surprising finding in an activestrictly carnivorous animal. In addition, it seems peculiarthat arms and tentacles, capturing and attacking structuresexposed to physical stress, have a surface with microvilli(Figs. 2–4, 16).
We investigated the ability of Sepia officinalis integu-ment to absorb amino acids by uptake experiments in theisolated arms. The autoradiographic pictures clearly showthat the labelled amino acids are taken up from the externalmilieu and suggest that proline transport might be depen-dent on the sodium gradient, whereas leucine uptake wouldnot (Figs. 11–14).
To determine if concentrative transport systems areinvolved in this amino acid transport, brush bordermembrane vesicles were purified from Sepia officinalis armintegument. Electron microscopy (Fig. 15) and the enrich-ment of the marker enzyme alkaline phosphatase (Table 1)provide evidence that the final preparation containedmembrane vesicles enriched in brush border.
The transient intravesicular accumulation of leucinewhich occurred in the presence of an inwardly directedsodium gradient (Fig. 19) indicates that the uptake is

AMINO ACID UPTAKE IN CUTTLEFISH INTEGUMENT 395
Fig. 16. Thin section of arm epithelial cells (E) at low magnification to easily identify the next immunofluorescence staining. Scale bar, 3 µm.
Fig. 17. Cryosection. Superimposed confocal image. The immunofluorescence staining using anti-ALP antibody showed a localization of the enzyme atthe level of the brush border membrane.
Fig. 18. Cryosection. Superimposed confocal image. The immunofluorescence staining using anti-Na+/K+ ATPase, an integral membrane proteincomplex responsible of electrochemical gradients of Na+ and K+ across the plasma membrane, was localized in the baso-lateral portion of epithelial cellmembranes.
performed by a carrier-mediated mechanism, further provedby the 90% inhibition of leucine uptake by an excess of thesubstrate (Fig. 22). As suggested by the experiments withthe arms incubated in vitro (Figs. 11–12), the transportagency involved in leucine uptake appears to have a lowspecificity for Na+ (Fig. 21) and recognises also cationicamino acids, since an excess of lysine caused a 42% inhibi-tion of leucine uptake (Fig. 22). Lysine, but also proline, isindeed absorbed in Sepia integument by a cotransport mech-anism, as shown by the presence of the peak (Fig. 20) and ofan increased uptake (Fig. 21) in the presence of the sodiumgradient.
The results here presented suggest that neutral andcationic amino acids are absorbed by Sepia integumentthrough a secondary active mechanism, in agreement withwhat found in other molluscs (Wright et al., 1987; Rice &Stephens, 1988; Stephens, 1988; Wright & Manahan, 1989;
Wright & Pajor, 1989; Preston, 1990). However, the neutralamino acid leucine crosses the apical membrane throughpathways with different features as compared to thoseobserved in Mytilus, since only 40% or less of leucineuptake was inhibited by either lysine or proline, while aninhibition of 84% was observed in marine bivalves.
The ability of cuttlefish to absorb amino acids by theintegument via a secondary active mechanism, in order toaccumulate them in cells and body fluids, is shared by manyother molluscs (Wright et al., 1987; Rice & Stephens; 1988,Stephens, 1988; Wright & Manahan, 1989; Wright & Pajor,1989; Preston, 1990) and may represent a phylogeneticalremnant.
The integument could, alternatively, be the largestabsorptive surface in S. officinalis, since in a specimen10 cm long it has an extension of tenfold the intestinal one.In this rough estimate we have a value by defect of the entire

396 DE EGUILEOR ET AL.
Fig. 19. Time course of leucine uptake in BBMV in the presence of aNa+-gradient. BBMV were incubated in a medium of the following finalcomposition: 300 mM mannitol, 10 mM Hepes-Tris at pH 7.6, 260 mMNaCl, 0.5 µM [3H] leucine 30 µCi/ml. Each point is the mean ± S.E. of anexperiment in triplicate. When not given, S.E. bars were smaller thansymbols.
Table 1. Enzyme activities in homogenate and BBMV.
Enzyme Homogenate BBMV R.E.mU/mg protein mU/mg protein
Na+-K+ ATPase 76.5 ± 8.9 93.3 ± 9.8 1.2 ± 0.1Alkaline phosphatase 4.8 ± 0.6 27.6 ± 2.4 5.9 ± 0.2
R.E.: relative enrichment factor, is the ratio between BBMV andhomogenate specific activities (mU/mg of protein). Means ± S.E. of threedifferent preparations.
body surface. In fact we assumed that the body and armscould be compared to a frustum of cones and tentacles andgut to cilinders. The effective absorptive surface per unitarea could be comparable, if we consider that the length andnumber of microvilli appear to be similar in body/armintegumental and intestinal epithelial cells (Figs. 2–5).Although no information is available on the amino acidtransport capability of the intestinal brush border, it seems
Fig. 20. Time course of lysine and proline uptake in BBMV in the presence composition: 300 mM mannitol, 10 mM Hepes-Tris at pH 7.6, 260 mM NaCl,of an experiment in triplicate. When not given, S.E. bars were smaller than sym
feasible that the remarkable absorptive properties of armsand body surface may play a significant role in nutrition andin other aspects of S. officinalis physiology. For example,even if cuttlefishes rely primarily on prey capture as afeeding source, they could exploit the absorptive ability oftheir integument to gain amino acids from DOM asosmolytes for the regulation of their internal environment.In fact, S. officinalis, like other marine molluscs whichutilise nitrogenous solutes as major intracellular osmoticeffectors (Somero & Bowlus, 1983), has a high intracellularamino acid concentration (Preston, 1993). Concurrently,amino acids could represent a supplement to traditional foodand an additional energy source when the animals remainburied under the sand, where the dissolved organic matter isvery high. The superficial absorption could be also of directadvantage to arm and tentacle muscle metabolism, particu-larly active because of the non-stop use of these appendagesto catch the prey, to dig in the sand or to perceive, like asearch periscope, any presence in the environment.
The intriguing problems of the functional properties andof the physiological role of integumental amino acid trans-port in cuttlefish is under current investigation.
of a Na+-gradient. BBMV were incubated in a medium of the following final 0.5 µM [3H] lysine or [3H] proline 30 µCi/ml. Each point is the mean ± S.E.bols.

AMINO ACID UPTAKE IN CUTTLEFISH INTEGUMENT 397
Fig. 21. Effect of different monovalent cations on the initial uptake rates of leucine, lysine and proline. BBMV were incubated in a medium of thefollowing final composition: 300 mM mannitol, 10 mM Hepes-Tris at pH 7.6, 260 mM LiCl or NaCl or KCl, 0.5 µM [3H] leucine or [3H] lysine or [3H]proline 30 µCi/ml. Means ± S.E. of an experiment in triplicate. Difference with the uptake in the presence of sodium was tested by Student’s t test.*p < 0.05; **p < 0.01; ***p< = 0.001.
Fig. 22. Inhibition of leucine uptake by different amino acids. BBMV were incubated for 10 s in a buffer of the following final composition: 300 mMmannitol, 10 mM Hepes-Tris at pH 7.6, 0.5 µM [3H] leucine 30 µCi/ml, 1 mM leucine, lysine or proline. Means ± S.E. of an experiment in triplicate.Difference with the uptake in the absence of inhibitor was tested by Student’s t test: *p<0.05; **p<0.01; ***p<0.001
ACKNOWLEDGEMENTS
We are grateful to Maria Luisa Guidali for technicalassistance.
REFERENCES
Adibi, S.A., Mercer, D.W. 1973. Protein digestion in human intestine asreflected in luminal, mucosal and plasma amino acid concentrationafter meals. J. Clin. Invest., 52, 1586–1594.

398 DE EGUILEOR ET AL.
Barnet, J.E.G., Munday, K.A. 1972. Structural requirements for activeintestinal sugar transport in the small intestine in the hamster. In:Transport across the intestine. W.L. Barland, and P.D. Samuel, eds.Churchill Livingstone, London.
Boucaud-Camou E., Boucher-Rodoni, R. 1983. Feeding and digestion inCephalopods. In: The Mollusca. P.W. Hochachka (ed) Vol. 2.Academic Press, London, 149–187.
Brocco, S.L. 1976. The ultrastructure of the epidermis, dermis,iridophores, leucophores and chromatophores of Octopus dofleini,Martini (Cephalopoda: Octopoda). Dissertation, University ofWashington.
Davis, J., Keenan, C., Stephens, G. 1985. Na+-dependent amino acid trans-port in bacteria-free urchin larvae. J. Comp. Physiol., B156, 121–127.
Dubowitz, V., Brooke, M.H. 1973. ‘Muscle biopsy: A modern approach’.Saunders, London.
de Eguileor, M., Daniel, S., Giordana, B., Lanzavecchia, G., Valvassori,R. 1994. Trophic exchanges between parent and young during devel-opment of Glossiphonia complanata (Annelida, Hirudinea). J. Exp.Zool., 269, 389–402.
Giordana, B., Sacchi, V.F., Hanozet, G.M. 1982. Intestinal amino acidabsorption in lepidopteran larvae. Biochim. Biophys. Acta, 692, 81–88.
Kinne, R.K.H. 1993. The role of organic osmolytes in osmoregulation:from Bacteria to Mammals, J. Exp. Zool., 265, 346–355.
Kier, W.M. 1982. The functional morphology of the musculature of squid(Loliginidae) arms and tentacles. J. Morphol., 172, 179–192.
Kier, W.M. 1991. Squid cross-striated muscle: the evolution of a special-ized muscle fiber type. Bull. Marin. Sci., 49, 389–403.
Leonardi M.G., Andrietti, F., Bonfanti, P., Sacchi, V.F. 1993. Theoreticalanalysis of cotransport: its use in alanine uptake in plasma membranevesicles, Pathobiology, 61, 222–229.
Mettrick, D.F., Podesta, R.B. 1974. Ecological and physiological aspectsof helminth-host interactions in the mammalian gastrointesinal canal.Adv. Parasitol., 12, 183–278.
Moore, M.D., Mumaw, V., Schonberg, M.D. 1960. Optical microscopy ofultrathin tissue sections. J. Ultrastruct. Res., 4, 113–116.
Packard, A. 1988. The skin of Cephalopods (Coleoids): general andspecial adaptations. In: The Mollusca. Form and function. E.R.Trueman and M.R. Clarke, (eds) Academic Press, London, 37–67.
Pajor, A.M., Wright, S.H. 1989. Uptake of lysine and proline via separate
α-neutral amino acid transport pathways in Mytilus gill brush bordermembranes. J. Membrane Biol., 107, 237–247.
Preston R.L. 1990. Sodium amino acid cotransport systems in marineinvertebrates. In: Comparative aspects of sodium amino acid cotrans-port systems. R.K.H. Kinne ed. Comp. Physiol., Vol. 7, Karger, Basel,pp 1–129.
Preston R.L. 1993. Transport of amino acids by marine invertebrates. J. Exp. Zool., 265, 410–421.
Quigley, J.P., Gotterer, G.S. 1969. Properties of a high specific activity(Na+/K+) stimulated ATPase from rat intestinal mucosa. Biochim.Biophys. Acta, 173, 456–469.
Rice, M.A., Stephens, G.C. 1988. Influx, net flux and transepithelial fluxof amino acid in the hardshell clam Mercenaria mercenaria (Linne):influence of salinity. Comp. Biochem. Physiol. A, 89, 631–636.
Richards, S., Arme, C. 1982. Integumentary uptake of dissolved organicmaterials by earthworms. Pedobiologia 23, 358–366.
Robertson, J. 1965. Studies on the chemical composition of muscle tissue.III The mantle tissue of cephalopod muscle. J. Exp. Biol. 42, 153–175.
Singley, C.T. 1982. Histochemistry and fine structure of the citodermalepithelium of the sepiolid squid Euprymna scolopes. Malacologia, 23,177–192.
Somero, G.N., Bowlus, R.D. 1983. Osmolytes and methabolic end prod-ucts of molluscs: the design of compatible solute systems. In: TheMollusca. A.S.M. Saleuddin and K.M. Wilbur (eds) Vol 5. AcademicPress, London, 77–100.
Southward, A., Southward, E. 1968. Uptake and incorporation of glycineby Pogonophores. Nature, 218, 875–876.
Southward, A., Southward, E. 1980. The significance of dissolved organiccompounds in the nutrition of Siboglium ekmani and other smallspecies of Pogonophora. J. Mar. Biol. Ass. UK, 60, 1005–1034.
Stephens, G. 1988. Epidermal amino acid transport in marine inverte-brates. Biochim. Biophys. Acta, 947, 113–138.
Wright S.H., Manahan, D.T. 1989. Integumental nutrient uptake byaquatic organisms. Annu. Rev. Physiol. 51, 585–600.
Wright S.H., Secomb, T.W., Bradley, T.J. 1987. Apical membrane perme-ability of Mytilus gill: influence of ultrastructure, salinity and competi-tive inhibitors on amino acid fluxes. J. Exp. Biol., 129, 205–230.
Wright, S.H., Pajor, A.M. 1989. Mechanism of integumental amino acidtransport in marine bivalves. Am. J. Physiol. 257, R473–R483.




















