
Light and electron microscopy study of the salivary glands of the carnivorous opisthobranch...
10
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
grupolusofona -
Category
Documents
-
view
4 -
download
0
Transcript of Light and electron microscopy study of the salivary glands of the carnivorous opisthobranch...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Tissue and Cell 41 (2009) 367–375
Contents lists available at ScienceDirect
Tissue and Cell
journa l homepage: www.e lsev ier .com/ locate / t i ce
Light and electron microscopy study of the salivary glands of the carnivorousopisthobranch Philinopsis depicta (Mollusca, Gastropoda)
Alexandre Lobo-da-Cunha a,b,∗, Íris Ferreira a, Rita Coelho c, Goncalo Calado c,d
a Laboratory of Cell Biology, Institute of Biomedical Sciences Abel Salazar (ICBAS), University of Porto, Largo Abel Salazar 2, 4099-003 Porto, Portugalb Centre of Marine and Environmental Research (CIIMAR), Porto, Portugalc Portuguese Institute of Malacology, Albufeira, Portugald Lusophone University of Humanities and Technologies, Lisbon and Institute for Marine Research, FCT/UNL, Caparica, Portugal
a r t i c l e i n f o
Article history:Received 23 January 2009Received in revised form 4 March 2009Accepted 5 March 2009Available online 11 April 2009
Keywords:CephalaspideaDigestive systemSecretory cellsHistochemistryCytochemistry
a b s t r a c t
Cephalaspideans are a group of opisthobranch gastropods that comprises carnivorous and herbivorousspecies, allowing an investigation of the relationship between these diets and the morphofunctional fea-tures of the salivary glands. In this study, the salivary glands of the carnivorous cephalaspidean Philinopsisdepicta were observed by light and electron microscopy. The secretory epithelium of these ribbon-shapedglands is formed by ciliated cells, granular cells and cells with apical vacuole. In ciliated cells the nucleusand most cytoplasmic organelles are located in the wider apical region and a very thin stalk reaches thebase of the epithelium. These cells possess significant amounts of glycogen. Granular cells are packed withelectron-dense secretory granules and also contain several cisternae of rough endoplasmic reticulum andGolgi stacks. The other type of secretory cell is mainly characterized by the presence of a large apical vac-uole containing secretion. These cells possess high amounts of rough endoplasmic reticulum cisternaeand several Golgi stacks. Vesicles with peripheral electron-dense material are also abundant, and seemto fuse to form the apical vacuole. The available data point out to a significant difference between thesalivary glands of carnivorous and herbivorous cephalaspidean opisthobranchs, with an intensificationof protein secretion in carnivorous species.
© 2009 Elsevier Ltd. All rights reserved.
1. Introduction
The opisthobranchs are a morphologically diversified group ofgastropods comprising between 5000 and 6000 species, almost allliving in marine and estuarine ecosystems (Rudman and Willan,1998; Wägele and Klussmann-Kolb, 2005). The cephalaspideansmake up the second largest taxon of opisthobranchs (Wägele, 2004;Malaquias et al., 2009a) including both carnivorous and herbivorousspecies (Kohn, 1983; Malaquias et al., 2009b).
The species under study here, Philinopsis depicta, was formerlyincluded in the genus Aglaja and belongs to the family Aglajidae(Gosliner, 1980; Malaquias et al., 2009a). Aglajids are active preda-tors that seek and eat other opisthobranchs or worms (Gosliner,1980; Martínez et al., 1993; Rudman and Willan, 1998). In Aglajids,the mouth is located at the anterior end of an oval or tubular buc-cal bulb with a strong muscular wall used for rapidly sucking inthe prey (Rudman, 1972c,d). In these carnivorous cephalaspideans
∗ Corresponding author at: Laboratory of Cell Biology, Institute of BiomedicalSciences Abel Salazar (ICBAS), University of Porto, Largo Abel Salazar 2, 4099-003Porto, Portugal.
E-mail address: [email protected] (A. Lobo-da-Cunha).
two short salivary glands are attached to the posterior region of thebuccal bulb near the connection between the buccal bulb and theoesophagus. The oesophagus widens shortly after leaving the buc-cal bulb to form a distensible crop where prey digestion is initiated.In the crop the soft tissues of the prey are dissolved and the nutritivefluid goes to the small stomach that is connected to the digestivegland. In molluscs, the digestive gland is formed by a vast numberof blind ending tubules that produces digestive enzymes and wherenutrient absorption largely occurs. The intestine is also connectedto the stomach and receives the indigested residues coming fromthe digestive gland (Rudman, 1972c).
Although relatively small, the salivary glands of Aglajids mayplay a role in the first stages of prey digestion. In gastropods, thesalivary glands secrete a mucous-containing fluid that at least insome species contains enzymes, suggesting that saliva contributesto digestion in the anterior part of the digestive tract (BolognaniFantin et al., 1982; Rittschof et al., 1990; Moura et al., 2004).
Amongst gastropods the salivary glands have been studiedmainly in pulmonates (Serrano et al., 1996; Luchtel et al., 1997;Moura et al., 2004) and also in some species previously includedin the prosobranch assemblage (Minniti, 1986; Benmeradi andBenmeradi, 1992; Voltzow, 1994; Andrews et al., 1999). From thesestudies it becomes evident that the relative size and complexity of
0040-8166/$ – see front matter © 2009 Elsevier Ltd. All rights reserved.doi:10.1016/j.tice.2009.03.001

Author's personal copy
368 A. Lobo-da-Cunha et al. / Tissue and Cell 41 (2009) 367–375
such glands is very variable among species, ranging from glandsin which just a single type of secretory cell and small ciliated cellswere reported (Morton, 1955) to others comprising four or evenmore types of cells (Walker, 1970; Serrano et al., 1996).
Salivary glands were also histologically surveyed in opistho-branch gastropods, but in a limited number of species (Fretter,1939; Rudman, 1971, 1972a,b,c; Wägele et al., 1999), and theirultrastructure was previously investigated only in two herbiv-orous species: the anaspidean Aplysia depilans (Lobo-da-Cunha,2001, 2002) and the cephalaspidean Bulla striata (Lobo-da-Cunhaand Calado, 2008). Some nudibranch opisthobranchs do not con-tain salivary glands, but instead have ptyaline glands (also knownas oral glands) attached to the anterior part of the oesophagus,however this type of gland has not yet been studied in detail(Gosliner, 1994; Brodie, 2001, 2004). Therefore, in opisthobranchsthe current knowledge about salivary glands and other kinds ofglands from the anterior region of the digestive system is stillvery limited. Thus, in order to obtain further data for a compar-ative analysis of the digestive system regarding herbivorous andcarnivorous cephalaspidean opisthobranchs, the salivary glandsof P. depicta were studied with light and electron microscopymethods.
2. Materials and methods
2.1. Morphology
Six specimens of P. depicta (Renier, 1807) between 7 and 8 cm inlength were collected in Ria de Alvor, an estuary in the south coastof Portugal. The salivary glands were removed and plunged for 2 hat 4 ◦C in fixative containing 2.5% glutaraldehyde and 4% formalde-hyde (obtained from hydrolysis of paraformaldehyde) diluted with0.4 M cacodylate buffer pH 7.4 (final buffer concentration 0.28 M).After washing in buffer, the glands were post-fixed for 2 h with2% OsO4 in the same buffer at room temperature, dehydratedin ethanol and embedded in Epon. Semithin sections for lightmicroscopy were stained with methylene blue and azure II. Ultra-thin sections were stained with uranyl acetate and lead citrate,before being observed in a JEOL 100CXII transmission electronmicroscope operated at 60 kV.
For scanning electron microscopy, salivary glands fixed asreported above were washed in buffer, frozen in liquid nitrogen andtransversely fractured. After dehydration in ethanol, salivary glandpieces were critical point dried, coated with gold and observed ina JEOL scanning electron microscope.
Fig. 1. Anatomy of the salivary glands of P. depicta. (a) Portion of the digestive system showing the buccal bulb (B), the two salivary glands (arrows), crop (C) and digestivegland (DG). (b) Isolated gland in witch the central longitudinal duct (arrow) is visible. (c) Transverse semithin section of a salivary gland showing the central duct (arrow). (d)Longitudinal semithin section showing the lateral tubules (T) which are perpendicularly attached to the central duct. (e) Scanning electron microphotograph of a transverselyfractured gland showing many cilia (arrows) and granules in the cytoplasm of secretory cells (asterisk).

Author's personal copy
A. Lobo-da-Cunha et al. / Tissue and Cell 41 (2009) 367–375 369
Fig. 2. Semithin sections of P. depicta salivary glands. (a) The epithelium contains granular cells (GC), cells with apical vacuole (AV) containing secretory material (arrow) andciliated cells (arrowheads). Cilia (c) can be seen protruding from the epithelium and a thin layer of connective tissue (asterisk) underlies the epithelium. Methylene blue andazure II staining. (b) Tetrazonium reaction for protein detection strongly stains the secretory granules of granular cell and material within vesicles (arrowheads) and vacuole(arrows) of cell with apical vacuole. (c) PAS reaction also stains the secretory material located in apical vacuoles (arrow) and granular cells (GC), but in some of these cellsstaining is less intense (asterisk). (d) Alcian blue staining at pH 1 detects acid polysaccharides in the secretion of granular cells (GC), but the secretory material of cells withapical vacuole is stained with less intensity (arrows).
2.2. Histochemistry
Light microscopy techniques for the detection of proteins andpolysaccharides, based on procedures described by Ganter andJollès (1970), were applied to semithin sections (2 �m) of salivaryglands fixed and embedded as reported above. For all histochemi-cal techniques, the embedding medium was not removed from thesections before staining. After staining, sections were washed, airdried and mounted with DPX.
PAS—Semithin sections were treated with 1% periodic acidfor 10 min, washed and stained with Schiff reagent for 30 min.In control sections, the oxidation with periodic acid was omit-ted.
Alcian blue—Semithin sections were stained at 50 ◦C for approx-imately 48 h with a 0.5% solution of Alcian blue in 3% acetic acid at
pH 2.5 for detection of carboxylic acid groups of polysaccharides,and in HCl solution at pH 1.0 for detection of sulphated polysaccha-rides.
Tetrazonium reaction—Semithin sections were treated for 10 minwith a freshly prepared 0.2% solution of fast blue salt B in veronal-acetate buffer (pH 9.2), washed and treated for 15 min with asaturated solution of �-naphthol in veronal-acetate buffer (pH9.2).
2.3. Ultrastructural cytochemistry
Glycogen detection—Ultrathin sections were collected on goldgrids, treated with 1% periodic acid for 30 min, washed in waterand treated for 4 h with a solution of 0.2% thiosemicarbazide in20% acetic acid. After being washed in 10% acetic acid and in water,

Author's personal copy
370 A. Lobo-da-Cunha et al. / Tissue and Cell 41 (2009) 367–375
Fig. 3. Ultrastructure of ciliated and granular cells in P. depicta salivary glands. (a) Apical region of ciliated cells showing cilia (arrows), mitochondria (M) and nucleus (N).(b) Mid portion of a ciliated cell, consisting of a thin strand of cytoplasm (asterisk) containing mitochondria (M) and a bundle of filaments (arrows). (c) Detection of glycogen(arrows) in the apical region of a ciliated cell. (d) Glycogen granules (arrows) are also very abundant in the thin mid and basal portions of ciliated cells. (e) Basal region of agranular cell showing the nucleus (N) several electron-dense secretory granules (SG) and rough endoplasmic reticulum cisternae (arrows).

Author's personal copy
A. Lobo-da-Cunha et al. / Tissue and Cell 41 (2009) 367–375 371
Fig. 4. Ultrastructure of secretory cells in P. depicta salivary glands. (a) In the Golgi stacks (G) of granular cells the trans face cisternae are usually filled with electron-densematerial (arrows). (b) In some granular cells the secretory granules (SG) fuse to form a large mass of secretion (asterisks). (c) Basal region of cells with apical vacuole showingnuclei (N) and vesicles (V) containing electron-dense secretory material attached to the membrane (arrows). (d) Detail of a cell with apical vacuole showing a large vesicle(V) with peripheral electron-dense material (arrows) and many cisternae of rough endoplasmic reticulum (RER).
Author's personal copy
372 A. Lobo-da-Cunha et al. / Tissue and Cell 41 (2009) 367–375
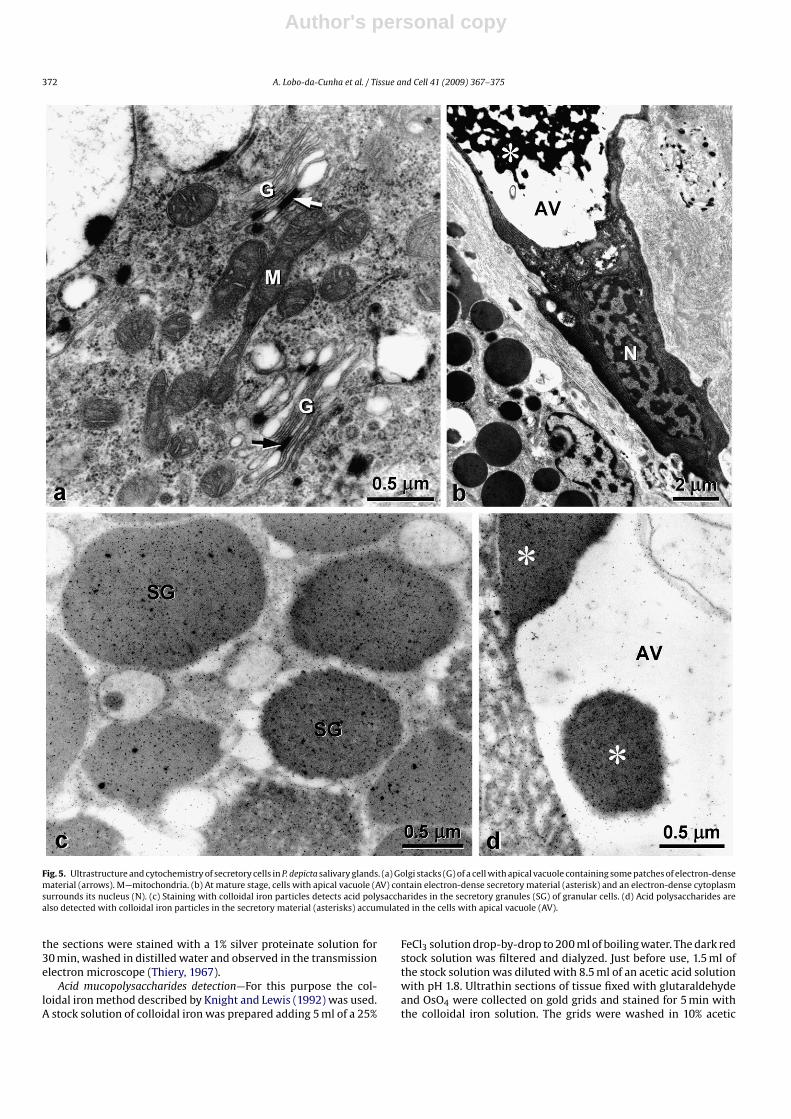
Fig. 5. Ultrastructure and cytochemistry of secretory cells in P. depicta salivary glands. (a) Golgi stacks (G) of a cell with apical vacuole containing some patches of electron-densematerial (arrows). M—mitochondria. (b) At mature stage, cells with apical vacuole (AV) contain electron-dense secretory material (asterisk) and an electron-dense cytoplasmsurrounds its nucleus (N). (c) Staining with colloidal iron particles detects acid polysaccharides in the secretory granules (SG) of granular cells. (d) Acid polysaccharides arealso detected with colloidal iron particles in the secretory material (asterisks) accumulated in the cells with apical vacuole (AV).
the sections were stained with a 1% silver proteinate solution for30 min, washed in distilled water and observed in the transmissionelectron microscope (Thiery, 1967).
Acid mucopolysaccharides detection—For this purpose the col-loidal iron method described by Knight and Lewis (1992) was used.A stock solution of colloidal iron was prepared adding 5 ml of a 25%
FeCl3 solution drop-by-drop to 200 ml of boiling water. The dark redstock solution was filtered and dialyzed. Just before use, 1.5 ml ofthe stock solution was diluted with 8.5 ml of an acetic acid solutionwith pH 1.8. Ultrathin sections of tissue fixed with glutaraldehydeand OsO4 were collected on gold grids and stained for 5 min withthe colloidal iron solution. The grids were washed in 10% acetic

Author's personal copy
A. Lobo-da-Cunha et al. / Tissue and Cell 41 (2009) 367–375 373
acid and finally in distilled water before being observed with theelectron microscope.
3. Results
3.1. Anatomy and light microscopy observations
In P. depicta, two ribbon-shaped salivary glands are linked to theposterior region of the buccal bulb close to the connection betweenthe buccal bulb and the oesophagus, but their posterior tip is notattached to the wall of the digestive tract (Fig. 1a). In average theseglands are about 5 mm long, about 600 �m wide and 120–150 �mthick. A central duct runs along its length (Fig. 1b and c) and oneach side of this duct the gland is formed by a single row of tubulesperpendicularly attached to the central duct (Fig. 1d). The apicalsurface of the epithelium of both central duct and lateral tubulesbears a dense coat of cilia, which are clearly visible in scanningelectron microscopy images of fractured glands (Fig. 1e).
In the central duct and lateral tubules the simple epithelium hasan average thickness of 50 �m and contains two different types ofsecretory cells as well as very thin ciliated cells. The base of theepithelium is covered by a very thin outer layer of connective tis-sue (Fig. 2a). In one kind of secretory cell the cytoplasm is filledwith large granules and therefore these cells were named “gran-ular cells” (Fig. 2a). These granules become strongly stained afterboth tetrazonium reaction for protein detection (Fig. 2b) and PASreaction for polysaccharide detection (Fig. 2c). In some granularcells staining intensity was weaker after the histochemical reac-tions, but this seems to reflect differences in maturation stage ratherthan different cell types (Fig. 2c). The secretory granules were alsostained with Alcian blue at pH 1 for sulphated polysaccharide detec-tion (Fig. 2d), but they do not stain at pH 2.5. The main feature ofthe other kind of secretory cell is a large apical vacuole contain-ing secretory material, and the cells of this type were called “cellswith apical vacuole” (Fig. 2a). The secretory material within theapical vacuole was strongly stained after histochemical detectionof proteins, and several vesicles in the cytoplasm also contain asmall drop of positively stained material (Fig. 2b). The secretionin the apical vacuole is also PAS positive (Fig. 2c) and althoughwith much less intensity, Alcian blue staining only at pH 1 couldbe detected in the secretory material accumulated in the vacuole(Fig. 2d).
3.2. Transmission electron microscopy observations
In the epithelium the thin ciliated cells are numerous, form-ing small clusters between the secretory cells. Their nucleus andmost cytoplasmic organelles including some mitochondria andlysosomes are located in the apical region which is the wider regionof the cell (Fig. 3a). The ciliated cells possess a thin and long cyto-plasmic stalk that crosses the epithelium to reach the basal lamina,
containing a bundle of filaments in addition to some mitochondria(Fig. 3b). Their cytoplasm also contains large number of glycogenparticles both in the apical region and in the thin stalk (Fig. 3c andd), but glycogen deposits were not detected in the secretory cells ofthese glands.
The granular cells contain numerous spherical electron-densesecretory granules, but the immature granules have a lower elec-tron density. In the cytoplasm around the granules cisterns of roughendoplasmic reticulum are abundant (Fig. 3e) and several Golgistacks with flattened cisterns are also present in these cells. At thecis face of the Golgi apparatus the cisterns have an electron-lucentcontent, whereas at the trans face they are filled with electron-dense secretory material (Fig. 4a). In some cells the granules havefused to form a large mass of electron-dense secretory materialfilling most of the cytoplasm, just with a few non-fused granulesaround it (Fig. 4b).
At younger stages of development, the cells with apical vacuolehave a basal nucleus (Fig. 4c) and a clear cytoplasm containing alarge amount of flattened rough endoplasmic reticulum cisternae,frequently in parallel arrays (Fig. 4d). Several mitochondria andvesicles of variable sizes with highly dense material attached tothe vesicle membrane are also common in these cells (Fig. 4c andd). Their numerous Golgi stacks are formed by cisternae that canshow dilated rims and a few patches of electron-dense material thatresembles the electron-dense material found in the vesicles andapical vacuole (Fig. 5a). As the cell matures, those vesicles fuse witheach other forming the large apical vacuole containing a variableamount of electron-dense secretory material in a clear back-ground.At the final stages of maturation, the cytoplasm becomes graduallydenser and the cell shrinks to attain a goblet shape with a dilatedapical zone containing a very large vacuole (Fig. 5b).
The colloidal iron staining for acid polysaccharides confirms thepresence of these compounds in the electron-dense secretory mate-rial of both types of secretory cells (Fig. 5c and d), whereas in thecells with apical vacuole the electron-lucent region of vesicles andvacuoles were almost free of colloidal iron particles (Fig. 5d).
4. Discussion
Light microscopy was used to describe salivary gland cells insome species of cephalaspideans (Fretter, 1939; Rudman, 1971,1972a,b,c), but without ultrastructural data it is difficult to comparecells accurately, and until now the ultrastructure of cephalaspideansalivary glands was reported only in P. depicta (current article) andB. striata (Lobo-da-Cunha and Calado, 2008). Ultrastructural dataare also available from A. depilans an anaspidean opisthobranch(Lobo-da-Cunha, 2001, 2002).
The general architecture of the salivary glands is simple and sim-ilar in P. depicta (Fig. 6) and B. striata (Lobo-da-Cunha and Calado,2008). In these two cephalaspideans, a central canal runs alongthe length of the thin ribbon-shaped glands dividing them in two
Fig. 6. Schematic representation of a segment of the salivary glands of P. depicta, showing the lateral tubes (T) and the central duct (arrow).

Author's personal copy
374 A. Lobo-da-Cunha et al. / Tissue and Cell 41 (2009) 367–375
halves, each formed by a single row of tubules perpendicularlyattached to the central duct. However, some differences are evidentbetween the salivary glands of P. depicta and the salivary glands ofherbivorous cephalaspideans of the genera Bulla and Haminoea. Inthese herbivores the salivary glands are longer and the posteriortip is attached to the exterior of the oesophageal wall (Rudman,1971), whereas in P. depicta and other Aglajids the posterior tipsof the salivary glands lie free in the body cavity (Rudman, 1972c).But, the most important differences between the salivary glandsof P. depicta and B. striata are the characteristics of the secretorycells. In both species two types of secretory cells were found inthese glands. In B. striata, the most common secretory cell typewas considered a granular mucocyte (Lobo-da-Cunha and Calado,2008) following a classification proposed for the salivary gland cellsof pulmonate gastropods (Serrano et al., 1996). In these cells, thesecretory vesicles contain small electron-dense peripheral spots ofsecretory proteins whereas the remaining space within each vesi-cle contains electron-lucent acid polysaccharides (Lobo-da-Cunhaand Calado, 2008). The other type of secretory cell found in B. stri-ata possesses large electron-lucent vesicles or vacuoles unstainedby light microscopy techniques, which were named vacuolated cellsfollowing the above mentioned classification. However, this last celltype contains acid polysaccharides in the secretory vacuoles (Lobo-da-Cunha and Calado, 2008) and could be better called “vacuolatedmucocyte”. In the salivary glands of A. depilans these two cells typesare also present, but a third type of secretory cell with cilia andsecretory granules containing proteins and neutral polysaccharideswas also reported (Lobo-da-Cunha, 2001, 2002). Thus, consideringthe ultrastructural and histochemical data, the secretory cells of thesalivary glands from the carnivorous P. depicta seem significantlydifferent from the cells previously described in the salivary glandsof herbivorous opisthobranchs.
The granular cells of P. depicta salivary glands are clearly pro-tein secreting cells producing dense secretory granules, resemblingcell types found in the salivary glands of some terrestrial pul-monate gastropods (Beltz and Gelperin, 1979; Moreno et al., 1982;Moya et al., 1992; Serrano et al., 1996). Typically, serous cells haveelectron-dense secretory granules containing proteins that can beassociated with limited amounts of polysaccharides (Tandler andPhillips, 1993), but in the granular cells of P. depicta the electron-dense secretory granules are strongly PAS positive and include acidpolysaccharides or oligosaccharides in addition to a high concen-tration of proteins, and could be considered as seromucous cells(Tandler and Phillips, 1993). Alcian blue staining of these granules atpH 1 demonstrates the presence of strongly acidic sulphate groupsin polysaccharides or glycoproteins, since carboxylic acid groupsare not ionized at this pH and thus unable to link with the cationicAlcian blue dye (Ganter and Jollès, 1970). The lack of staining at pH2.5 suggests the absence of carboxylic acid groups that would beionized at this pH. Moreover, it was demonstrated that some typesof sulphated glycoproteins stain at both pH 1 and 2.5, while oth-ers are stained only at the most acidic pH levels, that is, betweenpH 1.5 and 0.5 (Jones and Reid, 1973a,b), like those found in thesecretory cells of P. depicta salivary glands. In the granular muco-cytes of B. striata the secretory vesicles also include proteins andacid polysaccharides but these components are segregated withineach vesicle (Lobo-da-Cunha and Calado, 2008), while in P. depictathey are mixed.
The cells with apical vacuole also produce a mixture of pro-teins and polysaccharides, but their secretion stains less intenselywith Alcian blue than the secretion of granular cells. These twotypes of secretory cells are not very clearly distinct consideringthe results of the histochemical analysis, but morphologically theyare very different. The small vesicles in the basal region of thecells with apical vacuole morphologically resemble the vesicles ofgranular mucocytes of B. striata (Lobo-da-Cunha and Calado, 2008)
and A. depilans (Lobo-da-Cunha, 2001), because all of them con-tain peripheral electron-dense material. However, in P. depicta thecentral electron-lucent region of those vesicles was not stained bycolloidal iron particles, which were observed over the electron-dense protein containing material, while in granular mucocytes theiron particles were found just in the electron-lucent region of thevesicles (Lobo-da-Cunha, 2002; Lobo-da-Cunha and Calado, 2008).Thus, in the salivary glands of P. depicta both secretory cells typesproduce a mixture of proteins and acid polysaccharides. Differenttypes of proteins and polysaccharides must be secreted by each celltype, but additional studies are needed to clarify the roles of thesecells in the production of specific saliva components. In the herbivo-rous B. stiata and A. depilans mucus production clearly predominatein the salivary glands (Lobo-da-Cunha, 2001, 2002; Lobo-da-Cunhaand Calado, 2008) whereas in the carnivorous P. depicta both secre-tory cell types are more engaged in protein production. Thus, P.depicta saliva should be richer in proteins, probably including diges-tive enzymes, while in those herbivorous species mucus seems tobe a more important component of the saliva to facilitate the agglu-tination and ingestion of algae fragments or diatoms.
In the salivary glands of P. depicta and B. striata there are alsoslender ciliated cells intermingled in the secretory epithelium, butin P. depicta the cytoplasm of these cells is filled with glycogen gran-ules while in the accordant cells of B. striata just small amountsof glycogen were found (Lobo-da-Cunha and Calado, 2008). Mostprobably, the ciliated cells generate currents responsible for theflow of saliva towards the buccal cavity, and the glycogen stored inthese cells may be useful as an energy reserve to maintain ciliarymotion. However, in B. striata the granular mucocytes contain largeamounts of glycogen (Lobo-da-Cunha and Calado, 2008) that werenot reported in the secretory cells of P. depicta or A. depilans salivaryglands (Lobo-da-Cunha, 2001).
According to several phylogenetic studies the anaspideans andcephalaspideans s.str. can be regarded as sister groups (Mikkelsen,1996, 2002; Vonnemann et al., 2005), and since macroalgae anddiatoms are the food sources for most species within these groupsit can be assumed that the ancestor of both groups was a herbivore(Wägele, 2004; Malaquias et al., 2009b). Herbivory was also foundto be the plesiomorphic condition by superimposing the diet ontoa molecular cephalaspidean family-level phylogeny (Malaquias etal., 2009b). Thus, carnivorous cephalaspideans like P. depicta shouldhave had and a herbivorous ancestor. In this context, the study of thesalivary glands is especially interesting, since the features of theseglands must be correlated with both ancestry and adaptations tospecific food sources. So far, the salivary glands of opisthobranchswere studied in detail in very few species, but the presence ofgranular mucocytes and vacuolated mucocytes with great ultra-structural similarities in the herbivorous anaspidean A. depilans andcephalaspidean B. striata (Lobo-da-Cunha, 2001; Lobo-da-Cunhaand Calado, 2008) can represent an ancestral feature of their sali-vary glands. Similar cell types were also reported in the salivaryglands of herbivorous or omnivorous terrestrial pulmonates (Beltzand Gelperin, 1979; Serrano et al., 1996) and therefore seem to bewidespread among gastropods. On the other hand, the replacementof mucous cells by seromucous cells suggests and increment in pro-tein secretion and a reduction of mucus in the saliva in P. depicta,and this could represent an adaptation to a carnivorous diet in thelatter species. Since Aglajids ingest opisthobranchs or worms bysuction and except in Odontoglaja they do not have radula (Gosliner,1980; Rudman and Willan, 1998) probably they do not need to pro-duce mucus for agglutination of food particles, while herbivorousspecies that use the radula to scrap surfaces require more mucus forthat purpose. Moreover, it was suggested that the mucus coveringprey body may aid its own ingestion by Aglajids (Rudman, 1972c).However, this hypothesis needs to be confirmed by future studiesin other species.

Author's personal copy
A. Lobo-da-Cunha et al. / Tissue and Cell 41 (2009) 367–375 375
Acknowledgements
The authors thank Mr. João Carvalheiro and Miss Joana Car-valheiro for the reproduction of the microphotographs, and Mr.Emanuel Monteiro for technical assistance during sample prepa-ration for SEM. Rita Coelho holds a grant from the “Fundacão paraa Ciência e a Tecnologia”, Portugal (BDE 15577/2005) and GoncaloCalado also holds a grant from the “Fundacão para a Ciência e a Tec-nologia”, Portugal (BPD 7133/2001). This work was supported byICBAS and CIMAR.
References
Andrews, E.B., Page, A.M., Taylor, J.D., 1999. The fine structure and function of theanterior foregut glands of Cymatium intermedius (Cassoidea: Ranellidae). J. Moll.Stud. 65, 1–19.
Beltz, B., Gelperin, A., 1979. An ultrastructural analysis of the salivary system of theterrestrial mollusc Limax maximus. Tissue Cell 11, 31–50.
Benmeradi, S.S., Benmeradi, N., 1992. Ultrastructure et activité de la gland salivairede Patella safiana Lmk (Mollusque, Gastropode). 8 Congress National de la SociétéFrancaise de Malacologie. Actes de Colloque 13, 107–112.
Bolognani Fantin, A.M., Bolognani, L., Ottaviani, E., Franchini, A., 1982. The digestiveapparatus of Murex brandaris (L) and Trunculariopsis trunculus (L.). Z. Mikrosk.Anat. Forsch. 96, 561–582.
Brodie, G.D., 2001. Some comparative histological aspects of the dendrodorid generaDoriopsilla and Dendrodoris (Opisthobranchia: Nudibranchia). Boll. Mallacol. 37,99–104.
Brodie, G.D., 2004. An unusual dendrodorid: redescription of the tropical nudi-branch Dendrodoris atromaculata (Alder and Hancock 1864) (Anthobranchia:Doridoidea: Dendrodorididae). Zootaxa 503, 1–13.
Fretter, V., 1939. The structure and function of the alimentary canal of sometectibranch molluscs, with a note on excretion. Trans. R. Soc. Edinb. 59, 599–646.
Ganter, P., Jollès, G., 1970. Histochimie Normal et Pathologique, vol. 2. Gauthier-Villars, Paris.
Gosliner, T.M., 1980. Systematics and phylogeny of the Aglajidae (Opisthobranchia:Mollusca). Zool. J. Linnean Soc. 68, 325–360.
Gosliner, T.M., 1994. Gastropoda: Opisthobranchia. In: Harrison, F.W., Kohn, A.J.(Eds.), Microscopic Anatomy of Invertebrates, vol. 5, Mollusca I. Wiley-Liss, NewYork, pp. 253–355.
Jones, R., Reid, L., 1973a. The effect of pH on Alcian Blue staining of epithelial acid gly-coproteins I. Sialomucins and sulphomucins (singly or in simple combinations).Histochem. J. 5, 9–18.
Jones, R., Reid, L., 1973b. The effect of pH on Alcian Blue staining of epithelial acidglycoproteins II. Human bronchial submucosal gland. Histochem. J. 5, 19–27.
Knight, D.P., Lewis, P.R., 1992. General cytochemical methods. In: Glauert, A.M.(Ed.), Practical Methods in Electron Microscopy, vol. 14. Elsevier, Amsterdam,pp. 79–138.
Kohn, A.J., 1983. Feeding biology of gastropods. In: Saleuddin, A.S.M., Wilbur, K.M.(Eds.), The Mollusca, vol. 5, part 2. Academic Press, New York, pp. 1–63.
Lobo-da-Cunha, A., 2001. Ultrastructural and histochemical study of the salivaryglands of Aplysia depilans (Mollusca Opisthobranchia). Acta Zool. 82, 201–212.
Lobo-da-Cunha, A., 2002. Cytochemical localisation of lysosomal enzymes andacidic mucopolysaccharides in the salivary glands of Aplysia depilans (Opistho-branchia). J. Submicrosc. Cytol. Pathol. 34, 217–225.
Lobo-da-Cunha, A., Calado, G., 2008. Histology and ultrastructure of the salivaryglands in Bulla striata (Mollusca Opisthobranchia). Inv. Biol. 127, 33–44.
Luchtel, D.L., Martin, A.W., Deyrup-Olsen, I., Boer, H.H., 1997. Gastropoda: Pulmonata.In: Harrison, F.W., Kohn, A.J. (Eds.), Microscopic Anatomy of Invertebrates, vol.6B, Mollusca II. Wiley-Liss, New York, pp. 459–718.
Malaquias, M.A.E., Dodds, J.M., Bouchet, P., Reid, D.G., 2009a. A molecular phylogenyof the Cephalaspidea sensu lato (Gastropoda: Euthyneura): architectibranchiaredefined and Runcinacea reinstated. Zool. Scr. 38, 23–41.
Malaquias, M.A., Bericibar, E., Reid, E.D., 2009b. Reassessment of the trophic posi-tion of Bullidae (Gastropoda: Cephalaspidea) and the importance of diet in theevolution of cephalaspidean gastropods. J. Zool. 227, 88–97.
Martínez, E., Ballesteros, M., Ávila, C., Dantart, Cimino, G., 1993. La familia Aglajidae(Opisthobranchia: Cephalaspidea) en la Península Ibérica. Iberus 11, 15–29.
Mikkelsen, P.M., 1996. The evolutionary relationships of Cephalaspidea s.l (Gas-tropoda: Opisthobranchia): a phylogenetic analysis. Malacologia 37, 375–442.
Mikkelsen, P.M., 2002. Shelled opisthobranchs. Adv. Mar. Biol. 42, 67–136.Minniti, F., 1986. Morphological and histochemical study of pharynx of Leiblein,
salivary glands and gland of Leiblein in the carnivorous Gastropoda Amy-clina tinei Maravigna and Cyclope neritea Lamarck (Nassariidae: ProsobranchiaStenoglossa). Zool. Anz. 217, 14–22.
Moura, K.R.S., Terra, W.R., Ribeiro, A.F., 2004. The functional organization of thesalivary gland of Biomphalaria straminea (Gastropoda: Planorbidae): secretorymechanisms and enzymatic determinations. J. Moll. Stud. 70, 21–29.
Moreno, F.J., Pinero, J., Hidalgo, J., Navas, P., Aijón, J., Lópes-Campos, J.L., 1982. His-tochemical and ultrastructural studies on the salivary glands of Helix aspersa(Mollusca). J. Zool. 196, 343–354.
Morton, J.E., 1955. The functional morphology of Otina otis, a primitive marine pul-monate. J. Mar. Biol. Assoc. U.K. 34, 113–150.
Moya, J., Serrano, M.T., Ângulo, E., 1992. Ultrastructure of the salivary glands of Arionater (Gastropoda Pulmonata). Biol. Struct. Morph. 4, 81–87.
Rittschof, D., Kratt, C.M., Clare, A.S., 1990. Gastropod predation sites: the role ofpredator and prey in chemical attraction of the hermit crab Clibanarius vittatus.J. Mar. Biol. Assoc. U.K. 70, 583–596.
Rudman, W.B., 1971. Structure and functioning of the gut in the Bullomorpha(Opisthobranchia) Part 1. Herbivores. J. Nat. Hist. 5, 647–675.
Rudman, W.B., 1972a. Structure and functioning of the gut in the Bullomorpha(Opisthobranchia) Part 2. Acteonidae. J. Nat. Hist. 6, 311–324.
Rudman, W.B., 1972b. Structure and functioning of the gut in the Bullomorpha(Opisthobranchia) Part 3. Philinidae. J. Nat. Hist. 6, 459–474.
Rudman, W.B., 1972c. Structure and functioning of the gut in the Bullomorpha(Opisthobranchia) Part 4. Aglajidae. J. Nat. Hist. 6, 547–560.
Rudman, W.B., 1972d. A comparative study of the genus Philinopsis Pease, 1860(Aglajidae Opisthobranchia). Pac. Sci. 26, 381–399.
Rudman, W.B., Willan, R.C., 1998. Opistobranchia. In: Beesley, P.L., Ross, G.J.B., Wells,A. (Eds.), Mollusca: The Southern Synthesis. Fauna of Australia, vol. 5. CSIROPublishing, Melbourne, pp. 915–1035.
Serrano, T., Gómez, B.J., Ângulo, E., 1996. Light and electron microscopy study ofthe salivary gland secretory cells of Helicoidea (Gastropoda Stylommatophora).Tissue Cell 28, 237–251.
Tandler, B., Phillips, C.J., 1993. Structure of serous cells in salivary glands. Microsc.Res. Tech. 26, 32–48.
Thiery, J.P., 1967. Mise en évidence des polysaccharides sur coupes fines en micro-scopie électronique. J. Microsc. 6, 987–1018.
Voltzow, J., 1994. Gastropoda: Prosobranchia. In: Harrison, F.W., Kohn, A.J. (Eds.),Microscopic Anatomy of Invertebrates, vol. 5, Mollusca I. Wiley-Liss, New York,pp. 111–252.
Vonnemann, V., Schrödl, M., Klussmann-Kolb, A., Wägele, H., 2005. Reconstructionof the phylogeny of the Opisthobranchia (Mollusca: Gastropoda) by means of18S and 28S rRNA gene sequences. J. Moll. Stud. 71, 113–125.
Wägele, H., 2004. Potential key characters in Opisthobranchia (Gastropoda Mollusca)enhancing adaptive radiation. Org. Div. Evol. 4, 175–188.
Wägele, H., Klussmann-Kolb, A., 2005. Opisthobranchia (MolluscaGastropoda)—more than just slimy slugs. Shell reduction and its implicationson defence and foraging. Front. Zool. 2, 3.
Wägele, H., Brodie, G.D., Klussmann-Kolb, A., 1999. Histological investigations onDendrodoris nigra (Stimpson, 1855) (Gastropoda, Nudibranchia Dendrodoridi-dae). Moll. Res. 20, 79–94.
Walker, G., 1970. Light and electron microscope investigations on the salivary glandsof the slug Agriolimax reticulatus (Müller). Protoplasma 71, 111–126.




















