
Catecholamines Modulate Metamorphosis in the Opisthobranch Gastropod Phestilla sibogae
13
Catecholamines Modulate Metamorphosis in the Opisthobranch Gastropod Phestilla sibogae ANTHONY PIRES, 1, * ROGER P. CROLL 2 , AND MICHAEL G. HADFIELD 3 1 Department of Biology, Dickinson College, Carlisle, Pennsylvania 17013; 2 Department of Physiology & Biophysics, Dalhousie University, Halifax, Nova Scotia, Canada B3H 4H7; and 3 University of Hawaii, Pacific Biomedical Research Center, Kewalo Marine Laboratory, 41 Ahui St., Honolulu, Hawaii 96813 Abstract. Larvae of the nudibranch Phestilla sibogae are induced to metamorphose by a factor from their adult prey, the coral Porites compressa. Levels of endogenous cat- echolamines increase 6 to 9 days after fertilization, when larvae become competent for metamorphosis. Six- to nine- day larvae, treated with the catecholamine precursor L-DOPA (0.01 mM for 0.5 h), were assayed for metamor- phosis in response to coral inducer and for catecholamine content by high-performance liquid chromatography. L-DOPA treatment caused 20- to 50-fold increases in dopa- mine, with proportionally greater increases in younger lar- vae, so that L-DOPA-treated larvae of all ages contained similar levels of dopamine. A much smaller (about twofold) increase in norepinephrine occurred in all larvae. The treat- ment significantly potentiated the frequency of metamor- phosis of 7- to 9-d larvae at low concentrations of inducer. In addition, L-DOPA treatment at 9 d increased aldehyde- induced fluorescence in cells that were also labeled in the controls, and revealed additional cells. However, all labeled cells were consistent with the locations of cells showing tyrosine-hydroxylase-like immunoreactivity. Catecholamines are likely to modulate metamorphosis in P. sibogae, but rising levels of catecholamines around the time of compe- tence are insufficient alone to account for sensitivity to inducer in competent larvae. Introduction The larvae of many marine invertebrates settle and meta- morphose in response to chemical signals in the environ- ment. In some taxa, larvae become competent to respond to such chemical inducers only after an obligate planktonic phase (Pechenik, 1990; Hadfield and Strathmann, 1996). In gastropod molluscs, the minimum time to competence ranges from several days to several weeks and may or may not require feeding and growth, depending on the species. For example, the onset of competence occurs in a predict- able, age-dependent fashion in the lecithotrophic larvae of the opisthobranch Phestilla sibogae (Miller and Hadfield, 1986), but is extremely variable in another opisthobranch, Haminaea callidegenita (Gibson, 1995). For planktotrophic larvae of the prosobranch genus Crepidula, the onset of competence is influenced by temperature, salinity, and nu- tritional history and is poorly predicted by size (Zimmerman and Pechenik, 1991; Pechenik et al., 1996). Even after the onset of competence, responsiveness to an inducer can be affected by a larva’s sensory history. Habituation to inducer has been studied in the gastropods Phestilla sibogae and Haliotis rufescens (Hadfield and Scheuer, 1985; Trapido- Rosenthal and Morse, 1986a, b). In the latter species, sen- sitization to an inducer by another external ligand has also been reported (Trapido-Rosenthal and Morse, 1985, 1986b), and both phenomenon have been related to regulation at the level of surface receptors for the inducer. The structural and functional transformations that under- lie the acquisition of competence and changes in respon- siveness to inducers are not well understood. Competence may depend on the completion of neural networks that mediate between the initial chemosensory transduction of inducer and the behavioral and developmental events of metamorphosis. The modulation of such networks is an- Received 6 August 1999; accepted 20 January 2000. * To whom correspondence should be addressed. Abbreviations: a-MMT, a-methyl-DL-m-tyrosine; CI, coral inducer; DA, dopamine; DHBA, dihydroxybenzylamine; DOPAC, dihydroxyphe- nylacetic acid; FaGlu, formaldehyde-glutaraldehyde; HPLC, high perfor- mance liquid chromatography; L-DOPA, L-3,4-dihydroxyphenylalanine; MBL-ASW, MBL artificial seawater; NE, norepinephrine; PBS, phos- phate-buffered saline; TH-LIR, tyrosine-hydroxylase-like immunoreactive. Reference: Biol. Bull. 198: 319 –331. (June 2000) 319
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Catecholamines Modulate Metamorphosis in the Opisthobranch Gastropod Phestilla sibogae

Catecholamines Modulate Metamorphosis in theOpisthobranch Gastropod Phestilla sibogae
ANTHONY PIRES,1,* ROGER P. CROLL2, AND MICHAEL G. HADFIELD3
1 Department of Biology, Dickinson College, Carlisle, Pennsylvania 17013;2 Department of Physiology& Biophysics, Dalhousie University, Halifax, Nova Scotia, Canada B3H 4H7; and3 University of Hawaii,Pacific Biomedical Research Center, Kewalo Marine Laboratory, 41 Ahui St., Honolulu, Hawaii 96813
Abstract. Larvae of the nudibranchPhestilla sibogaeareinduced to metamorphose by a factor from their adult prey,the coral Porites compressa.Levels of endogenous cat-echolamines increase 6 to 9 days after fertilization, whenlarvae become competent for metamorphosis. Six- to nine-day larvae, treated with the catecholamine precursorL-DOPA (0.01 mM for 0.5 h), were assayed for metamor-phosis in response to coral inducer and for catecholaminecontent by high-performance liquid chromatography.L-DOPA treatment caused 20- to 50-fold increases in dopa-mine, with proportionally greater increases in younger lar-vae, so thatL-DOPA-treated larvae of all ages containedsimilar levels of dopamine. A much smaller (about twofold)increase in norepinephrine occurred in all larvae. The treat-ment significantly potentiated the frequency of metamor-phosis of 7- to 9-d larvae at low concentrations of inducer.In addition,L-DOPA treatment at 9 d increased aldehyde-induced fluorescence in cells that were also labeled in thecontrols, and revealed additional cells. However, all labeledcells were consistent with the locations of cells showingtyrosine-hydroxylase-like immunoreactivity. Catecholaminesare likely to modulate metamorphosis inP. sibogae,butrising levels of catecholamines around the time of compe-tence are insufficient alone to account for sensitivity toinducer in competent larvae.
Introduction
The larvae of many marine invertebrates settle and meta-morphose in response to chemical signals in the environ-ment. In some taxa, larvae become competent to respond tosuch chemical inducers only after an obligate planktonicphase (Pechenik, 1990; Hadfield and Strathmann, 1996). Ingastropod molluscs, the minimum time to competenceranges from several days to several weeks and may or maynot require feeding and growth, depending on the species.For example, the onset of competence occurs in a predict-able, age-dependent fashion in the lecithotrophic larvae ofthe opisthobranchPhestilla sibogae(Miller and Hadfield,1986), but is extremely variable in another opisthobranch,Haminaea callidegenita(Gibson, 1995). For planktotrophiclarvae of the prosobranch genusCrepidula, the onset ofcompetence is influenced by temperature, salinity, and nu-tritional history and is poorly predicted by size (Zimmermanand Pechenik, 1991; Pecheniket al., 1996). Even after theonset of competence, responsiveness to an inducer can beaffected by a larva’s sensory history. Habituation to inducerhas been studied in the gastropodsPhestilla sibogaeandHaliotis rufescens(Hadfield and Scheuer, 1985; Trapido-Rosenthal and Morse, 1986a, b). In the latter species, sen-sitization to an inducer by another external ligand has alsobeen reported (Trapido-Rosenthal and Morse, 1985, 1986b),and both phenomenon have been related to regulation at thelevel of surface receptors for the inducer.
The structural and functional transformations that under-lie the acquisition of competence and changes in respon-siveness to inducers are not well understood. Competencemay depend on the completion of neural networks thatmediate between the initial chemosensory transduction ofinducer and the behavioral and developmental events ofmetamorphosis. The modulation of such networks is an-
Received 6 August 1999; accepted 20 January 2000.* To whom correspondence should be addressed.Abbreviations: a-MMT, a-methyl-DL-m-tyrosine; CI, coral inducer;
DA, dopamine; DHBA, dihydroxybenzylamine; DOPAC, dihydroxyphe-nylacetic acid; FaGlu, formaldehyde-glutaraldehyde; HPLC, high perfor-mance liquid chromatography;L-DOPA, L-3,4-dihydroxyphenylalanine;MBL-ASW, MBL artificial seawater; NE, norepinephrine; PBS, phos-phate-buffered saline; TH-LIR, tyrosine-hydroxylase-like immunoreactive.
Reference:Biol. Bull. 198: 319–331. (June 2000)
319

other possible cause of both age- and context-dependentshifts in responsiveness to inducers. Such networks arepoorly characterized due to the difficulty of neurophysio-logical work in larvae (Arkettet al.,1989; Barlow, 1990). Incontrast, a wealth of information is available on the neuralbases of many behaviors in adult gastropods, whose nervoussystems are much more accessible. A common principle isthat monoamine neurotransmitters and neurohormones of-ten play critical roles in initiating and controlling behavior(Katz, 1995; Kabotyanskiet al.,1998). For example, dopa-mine is important in the control of several neural processesthat are normally associated with a chemosensory stimulus:it has been shown to elicit feeding motor programs inseveral pulmonates (Quinlanet al., 1997; Trimble andBarker, 1984; Wieland and Gelperin, 1983), modulate theactivity of olfactory interneurons inLimax maximus(Gel-perin et al., 1993), and activate hunting behavior in theopisthobranchClione limacina (Norekyan and Satterlie,1993).
Given that gastropod veligers contain monoamines andother neuromodulatory transmitters (Kempfet al., 1992,1997; Pireset al.,1997), it is reasonable to ask whether suchcompounds participate in the control of metamorphosis(Pires and Hadfield, 1991; Pireset al., 1997; Couper andLeise, 1996; Froggett and Leise, 1999). We have usedveligers ofP. sibogae,which settle and metamorphose inresponse to an inducer derived from their adult prey, thecoralPorites compressa(Hadfield, 1978; Hadfield and Pen-nington, 1990). The levels of catecholamines in these larvaeincrease several-fold within a period of 4–5 days afterhatching, during which the larvae become competent formetamorphosis.a-methyl-DL-m-tyrosine (a-MMT), an in-hibitor of the catecholamine-generative enzyme tyrosinehydroxylase (Uretskyet al., 1975; Bernabeet al., 1996),depletes catecholamines and inhibits induction of metamor-phosis by coral inducer (Pireset al.,1997). We now reportthat coral-induced metamorphosis is potentiated by ex-posure of competent larvae toL-DOPA, the syntheticprecursor of catecholamines. This treatment substantiallyenhanced endogenous norepinephrine (NE) and espe-cially dopamine (DA).L-DOPA treatment also restoredcatecholamine levels and ability to metamorphose inlarvae that had been treated witha-MMT. Histochemicalstudy of catecholamine-containing tissues suggested thatenhancement of catecholamines was localized in popula-tions of cells in locations consistent with those of cellsexhibiting tyrosine-hydroxylase-like immunoreactivity.Although these results support a role for catecholaminesin the control of metamorphosis, experiments withyounger larvae indicated that rising levels of cat-echolamines were not by themselves sufficient to accountfor the onset of competence.
Materials and Methods
Larval culture
Larvae were cultured as described by Miller and Hadfield(1986) with modifications given in Pires and Hadfield(1991). Larval ages are given as days postfertilization. Alllarvae were hatched at 5 d and held unfed in culture untilneeded. Veligers ofPhestilla sibogaeare facultative plank-totrophs; under our culture conditions larvae begin to attainmetamorphic competence at 7 d and by 10 d nearly allindividuals are competent without having fed (Kempf andHadfield, 1985; Miller and Hadfield, 1986).
Manipulation of catecholamine biosynthesis
Two sets of experiments were performed to test howmanipulating catecholamine biosynthesis affected metamor-phosis. In the first, content of endogenous catecholaminesand frequency of metamorphosis were determined in 9-dlarvae after treatment witha-MMT, L-DOPA (both fromSigma Chemical Co., St. Louis, MO), or both. (Endogenouscatecholamines are depleted bya-MMT (Pireset al.,1997),and preliminary experiments done for this study indicatedthat they are enhanced byL-DOPA at the concentrationsused here.) Five replicates were performed on differentculture batches on different dates. In each replicate, 1500–2000 larvae were taken from culture on day 8 and evenlydivided among four treatments, all carried out in MBLartificial seawater (MBL-ASW; Cavanaugh, 1956) bufferedto pH 8.2 with 10 mM Trizma: (1) 24 h in MBL-ASWalone; (2) 24 h in 1 mM a-MMT; (3) 21 h in MBL-ASWfollowed by 0.5 h in 1025 M L-DOPA followed by 2 h inMBL-ASW; (4) 21 h in 1 mM a-MMT followed by 0.5 h in1025 M L-DOPA plus 1 mM a-MMT followed by 2 h in 1mM a-MMT. L-DOPA was applied as a 0.5-h pulse fol-lowed by a MBL-ASW rinse becauseL-DOPA and othercatechols oxidize rapidly in seawater and the oxidativeproducts themselves affect metamorphosis (Pires and Had-field, 1991). All treatments were carried out in 20-ml vol-ume in 62-mm glass Stender dishes. At the end of the drugtreatments, a sample of each group was homogenized foranalysis of catecholamines by high-performance liquidchromatography (HPLC); the remainder was used to assaymetamorphosis.
The second set of experiments tracked catecholaminecontent, frequency of metamorphosis, and onset of compe-tence in larvae of increasing ages after modest or extremeenhancement of catecholamines by different doses ofL-DOPA. Four replicate series were performed, beginningon different dates with different culture batches. In eachreplicate series about 1000 six-day-old larvae were pipettedfrom their culture into MBL-ASW, and then divided intothree treatments, all in MBL-ASW: (1) 2–3 h in MBL-ASWalone; (2) 0.5 h in 53 1027 M L-DOPA followed by 2 h in
320 A. PIRES ET AL.

MBL-ASW; (3) 0.5 h in 1025 M L-DOPA followed by 2 hin MBL-ASW. A sample of each group was homogenizedfor analysis of catecholamines by HPLC; the remainder wasused to assay metamorphosis. This experiment was repeatedon days 7, 8, and 9 with larvae freshly drawn from the sameculture in each replicate series. Nine-day-old larvae did notreceiveL-DOPA treatments; frequency of metamorphosisand catecholamine levels in these oldest larvae served forcomparison with younger animals from the same batch thathad been treated withL-DOPA on previous days.
Preparation of metamorphic inducer and assay ofmetamorphosis
Frequency of metamorphosis was assayed in response topreparations of the natural inducer. This “coral inducer”(CI) was made by incubating a few “fingers” ofPoritescompressa(about 50–75 g wet weight) in an 11.5-cm glassbowl filled with MBL-ASW. After 24 h the CI was filteredthrough coffee-filter paper, adjusted to 32 ppt salinity, andfrozen in aliquots at220°C. Aliquots to be used on a givenday were thawed to room temperature and diluted with freshMBL-ASW to make a graded dilution series of CI rangingup 0.5 times (0.53) full strength (50% v/v CI in MBL-ASW). All experiments on a given culture batch of larvaeused freshly thawed aliquots of the same CI. In each assay,20–40 larvae were transferred to 37-mm glass Stenderdishes containing 3 ml 0.053, 0.13, 0.23, or 0.53 CI orMBL-ASW control. (For larvae that had been treated witha-MMT, the dilution series of CI used to assay metamor-phosis also contained 1 mM a-MMT to maintain inhibitionof catecholamine synthesis for the duration of the assay.)Assays were scored after 24 h. Individuals that had evacu-ated the larval shell were counted as having metamorphosed(Hadfield, 1978). Effects of CI concentration and of drugtreatments on arcsine-transformed frequencies of metamor-phosis were analyzed by ANOVA with Bonferroni adjust-ment for multiple comparisons (Zolman, 1993), using Data-Desk 6.1 statistical analysis software (Data Description,Ithaca, NY).
Extraction of catecholamines and analysis by HPLC
For each determination of catecholamine content, 150–250 larvae were counted and pipetted into a glass homog-enization tube on ice. The tube was briefly centrifuged(,1503 g), and excess water was drawn off to concentratelarvae in a volume of about 25ml. Homogenization oflarvae and extraction, separation, identification, and quan-tification of catecholamines by HPLC were modified fromprocedures described previously (Pireset al., 1997). Insummary, larvae were homogenized in a perchlorate buffer;supernatants of centrifuged homogenates were extractedover acid-washed alumina (Bioanalytical Systems) in 1.5MTris buffer (pH 8.6). Alumina was washed in deionized
water and transferred to microcentrifuge filters, where ex-cess water was spun off and catecholamines were elutedwith 0.1 M perchloric acid. Efficiency of extraction ofcatecholamines, typically 60%–80%, was corrected with adihydroxybenzylamine (DHBA) internal standard added toall larval homogenates. Catecholamines were separated on aBioanalytical Systems MF-6213 analytical column (10033.2 mm, 3mm C-18 reversed phase packing). The aqueousportion of the mobile phase contained 100 mM monochlo-roacetic acid, 1.3 mM Na2EDTA, and 1.3 mM sodium octylsulfate. Mobile phase pH was adjusted between 3.0 and 3.3with NaOH. The organic portion of the mobile phase was2%–4% (v/v) acetonitrile. Flow rate was set at 1.0 ml/min.Catecholamines were detected with a Bioanalytical SystemsLC-4C amperometric electrochemical detector with glassycarbon working electrode set at an oxidizing potential of650 mV against a Ag/AgCl reference electrode. Detectoroutput was low-pass filtered (1.0 Hz cutoff) and sent to aHewlett-Packard 3395 integrator that printed chromato-grams and calculated peak areas and retention times. Cate-cholamine contents were expressed as picomoles of freebase per larva.
Histochemistry
At least 50 larvae were obtained from each group afterscoring for frequency of metamorphosis in two replicates ofthe first set of experiments described above. In addition,several hundred 9-d larvae were taken directly from culturewithout drug treatment. All larvae were immobilized in a1:1 mixture of MBL-ASW and 7.5% (w/v) MgCl2 in water.These larvae were gathered into 1.5-ml microcentrifugetubes and spun gently (,1503 g rpm) for 5–10 s and werethen processed using the formaldehyde-glutaraldehyde(FaGlu) procedure modified from Furnesset al. (1977) asfollows: The MBL-ASW/MgCl2 was replaced with FaGlusolution of 4% paraformaldehyde and 0.55% glutaraldehydein phosphate-buffered saline (PBS: 50 mM Na2HPO4 27H2O and 140 mM NaCl, pH 7.4). After 18 h, the fixedlarvae were decalcified in 10% EDTA for 10 min, washedbriefly in PBS, and then dehydrated through an ascendingethanol series (70%, 80%, 90%, 95%, 100%, 100%; 2min/step). The larvae were then cleared and mounted onglass slides with methyl salicylate. The slides were viewedthrough a Zeiss Axiophot microscope equipped for ultravi-olet (UV) epifluorescence with a filter block containing365-nm excitation and 420-nm longpass barrier filters andphotographed using Kodak T-MAX 400 film. Negative con-trols were also prepared with no glutaraldehyde added to thefixative. Such control preparations exhibited none of theblue-green fluorescence reported in this study.
321CATECHOLAMINES MODULATE METAMORPHOSIS

Immunocytochemistry
Several hundred 9-d larvae taken directly from culturewithout any drug treatment were also processed for immu-nocytochemistry using monoclonal antibodies raised againsttyrosine hydroxylase (TH; IncStar #22941, Stillwater, MN)and procedures similar to those employed previously ingastropods (Herna´di et al., 1993; Voronezhskayaet al.,1999; Crollet al., 1999).
Larvae were first immobilized with MgCl2 as describedabove and then fixed in 100% methanol at220°C. (Prelim-inary experiments had indicated that such methanol fixationyielded more intense labeling than using paraformaldehydein another gastropod,Lymnaea stagnalis,in which cat-echolaminergic neurons have been well characterized [Crolland Chiasson, 1990; Elekeset al., 1991; Voronezhskayaetal., 1999; Crollet al.,1999].) After 2–4 h, the fixed larvaewere decalcified in 10% EDTA and then placed in a block-ing solution of PBS with the addition of 2% bovine serumalbumin and 1% Triton X-100. This and subsequent stepswere performed at 4°C. After 12 h, larvae were transferredto the primary anti-TH antibody diluted 1:50 in the blockingsolution for 48 h. After three 30-min washes, larvae wereincubated overnight in a 1:50 dilution of fluorescein iso-thiocyanate-labeled polyclonal goat anti-mouse antibodies(Sigma #F5262). Following several more washes in PBS,larvae were placed on glass slides and mounted in 3:1glycerol in 0.1M Tris buffer (pH 8.5) with the addition of2% n-propyl gallate (Giloh and Sedat, 1982).
Preparations were examined either on the Zeiss Axiophotmicroscope equipped with a filter block containing 450–490 nm excitation and 515–565 nm bandpass barrier filtersor on a Zeiss Axiovert microscope equipped for confocallaser scanning (Zeiss model LSM 510; excitation 488 nmwith 505–550 nm barrier filter). In the latter case, photo-graphs were produced by superpositioning stacks of 20–40images obtained through stepped sequences of focal planesat about 1.0-mm intervals.
Negative control experiments used identical proceduresexcept that the primary antibody was omitted. No labelingwas observed in these control preparations.
Results
Depletion and enhancement of catecholamines incompetent larvae
NE and DA were present in quantifiable amounts inlarvae of all ages and all drug treatment groups used in thisstudy. Figure 1 shows chromatograms of catecholaminestandards and of alumina extracts of 9-d larvae that hadbeen treated with MBL-ASW only, the tyrosine hydroxy-lase inhibitora-MMT (1 mM), or the catecholamine pre-cursorL-DOPA (1025 M). Numbers of larvae and extractionefficiencies were very similar between treatments, so in-
spection of peaks roughly indicates relative amounts ofcatecholamines per larva. These chromatograms show thequalitative effects of catecholamine depletion bya-MMTand enhancement byL-DOPA, compared to larvae held inMBL-ASW. Enhancement of DA is especially evident inthe chromatogram ofL-DOPA-treated larvae (Fig. 1D). Inaddition, this chromatogram shows a prominent peak iden-
Figure 1. Chromatograms of catecholamine standards, and of aluminaextracts of 9-d larvae ofPhestilla sibogaefrom one replicate of anexperiment in which catecholamine biosynthesis was manipulated. Stan-dards (A) contained 0.32 pmol each ofL-DOPA, norepinephrine, anddihydroxybenzylamine (DHBA), and 1.6 pmol of dopamine. (B) Extract of210 larvae treated with MBL artificial seawater only; (C) extract of 210larvae treated with 1 mM a-methyl-DL-m-tyrosine; (D) extract of 222larvae treated with 1025 M L-DOPA. Current scale of detector changes atpoints marked by vertical arrows.
322 A. PIRES ET AL.

tified as the dopamine metabolite dihydroxyphenylaceticacid (DOPAC) on the basis of its co-elution with authenticDOPAC over a range of mobile phase pH and acetonitrileconcentrations. Test extractions revealed that recovery ofDOPAC was not accurately estimated by our DHBA inter-nal standard, so we do not report quantitative values forDOPAC.
Treatment of 9-d larvae with a 0.5-h pulse of 1025 ML-DOPA followed by a 2-h incubation in MBL-ASW re-sulted in an average 26-fold enhancement of endogenousDA (Fig. 2A), compared to larvae exposed to MBL-ASWalone. In contrast, DA was depleted to 49% of MBL-ASWcontrol values in larvae that had been exposed toa-MMT (1mM for 24 h). DA was elevated to 11 times the value ofMBL-ASW controls in larvae that received a combinedtreatment ofa-MMT and L-DOPA (1 mM a-MMT for 21 h,followed by anL-DOPA pulse as described above but in thecontinued presence ofa-MMT). A similar pattern of deple-tion and enhancement of NE was observed following the
above treatments (Fig. 2B), although the magnitude of theseeffects was much smaller than was observed for DA.
Analysis of variance revealed significant effects of cul-ture batch (P , 0.01), CI concentration, drug treatment and(CI 3 drug) interaction (allP , 0.0001) on frequency ofmetamorphosis in 9-d larvae. No larva in any treatmentmetamorphosed in the absence of CI. Treatments witha-MMT and L-DOPA that modulated endogenous catechol-amine levels had dramatic and opposite effects on the fre-quency of coral-induced metamorphosis (Fig. 3). Bonfer-roni post-hoc tests compared effects of individual drugtreatments. Larvae that had been held 24 h in 1 mM a-MMTmetamorphosed at much lower frequencies than controllarvae held in MBL-ASW (P , 0.0001). Larvae that hadreceived a 0.5-h pulse of 1025 M L-DOPA, ending 2 hbefore exposure to CI, metamorphosed at significantlyhigher frequencies than did MBL-ASW controls when chal-lenged with dilute preparations of CI (P , 0.0001). How-ever, mean frequencies of metamorphosis were not signifi-cantly different in these two groups at the highestconcentration of CI. Larvae that received a combined treat-ment ofa-MMT and L-DOPA metamorphosed at frequen-cies that also were not significantly different than those ofMBL-ASW controls.
Histochemistry
Nine-day larvae taken from two replicates of the exper-iment documented in Figures 2 and 3 were subjected to
Figure 3. Frequency of metamorphosis of larvae ofPhestilla sibogaeafter 24-h exposure to varying dilutions of coral inducer on day 9 postfer-tilization. Prior to inducer, larvae were subjected to the following treat-ments: 1 mM a-methyl-DL-m-tyrosine (a-MMT, filled circles), 1 mMa-MMT plus 1025 M L-DOPA (filled triangles), 1025 M L-DOPA (opentriangles), or MBL artificial seawater control (open circles). Each pointrepresents the mean (61 SEM) of 5 replicate trials. Double asterisksindicate significant difference (P , 0.01) from MBL artificial seawatercontrol at a given dilution of coral inducer.
Figure 2. Content of dopamine (A) and norepinephrine (B) at 9 dpostfertilization in larvae ofPhestilla sibogaeafter treatment with 1 mMa-methyl-DL-m-tyrosine (a-MMT), 1 mM a-MMT plus 1025 M L-DOPA,1025 M L-DOPA, or MBL artificial seawater control. Each column repre-sents the mean (61 SEM) of 5 replicate trials.
323CATECHOLAMINES MODULATE METAMORPHOSIS

FaGlu histochemistry to visualize catecholamine-containingcells. The staining patterns in normal larvae without anyprior drug treatment and those incubated in MBL-ASW orMBL-ASW plus a-MMT were all similar. Patches of 3 to 6faint, blue-green fluorescent cells were reliably located inthe propodial region of the foot of larvae from all of thesegroups (Fig. 4A). In addition, 2 to 4 fluorescent cells were
located near the mouth, 1 to 2 cells were located morelateral to the mouth and near the base of the velum, and afinal 2 to 4 fluorescent cells were located in regions anteriorto the eyes (Fig. 4B).
The intensity of fluorescence and the numbers of fluores-cent cells were markedly enhanced following treatment withL-DOPA alone or withL-DOPA plus a-MMT. In these
Figure 4. Aldehyde-induced histofluorescence. Frontal (A) and lateral (B) views of control larvae showinga few dimly fluorescent cells (arrowheads) in the region of the propodium, lateral to the mouth and near theapical sensory organ anterior to the eye. One or more cells were also sometimes observed near the base of thevelum. No cells were generally found toward the distal tip of the foot to which the operculum is attached. Frontal(C) and lateral (D) views of larvae that were previously incubated inL-DOPA. Many more cells fluorescedbrightly in such larvae. Most of these fluorescent cells were located in two rows running along the length of thefoot (arrowheads in C) and in a patch on each side of the propodium. Several of these cells (arrowheads in D)appeared to be vase-shaped with slender processes projecting into the overlying epithelium. Calibration barsequal approximately 50mm.
324 A. PIRES ET AL.

larvae, the largest concentrations of fluorescent cells wereagain found in the propodial regions of the foot. Each of thebilateral patches contained 10 to 20 brightly fluorescentcells (Fig. 4C). In addition, several fluorescent cells werelocated in lateral bands along the length of the foot. At leastsome of these cells appeared to possess apical processesextending through the overlying epithelium (Fig. 4D).
Larvae treated with eitherL-DOPA or L-DOPA plusa-MMT also contained many brightly fluorescent cells inthe head region. Several cells were routinely observedaround the mouth, near the base of the velum (Fig. 4C), andanterior to the eyes. Thus the distribution of the cells in headregions was similar to that observed in normal larvae and inthose incubated in MBL-ASW alone, or in MBL-ASW plusa-MMT. However, the cells observed in the head regions oflarvae treated withL-DOPA or L-DOPA plusa-MMT ap-
peared to be more intensely labeled and slightly more nu-merous.
Immunocytochemistry
Tyrosine-hydroxylase-like immunoreactive (TH-LIR)cells were labeled in 9-d larvae. The highest concentrationof TH-LIR cells was located in bilateral patches in thepropodium (Fig. 5A). Each patch contained about 40 to 50cells (Fig. 5B). These cells appeared to be bipolar with oneprocess penetrating the overlying epithelium and a longaxon that projected to the region of the pedal ganglia.Similar cells were located at lesser densities on lateral edgesalong the length of the foot (Fig. 5C). Between 2 and 4TH-LIR cells were located at each side near the mouth, and2 to 4 additional cells were located more laterally near the
Figure 5. Tyrosine-hydroxylase-like immunoreactivity shown in projection patterns of normal larvae,reconstructed from multiple optic sections from a confocal microscope. (A) Lateral view of larva with velar lobesfolded medially by mounting. A concentration of immunoreactive, subepithelial cells was labeled in thepropodium. These cells appeared to send axons into the developing pedal ganglia. Large numbers of other cellswere located along the length of the foot. Other cells were scattered near the mouth and velum, and in regionsof the head near and dorsal to the apical sensory organ and eyes. (B) Magnified and more frontal view of somata(larger arrowheads) in propodium. Many of these cells possessed processes (slender arrows) that penetrated theoverlying epithelium. (C) A smaller number of somata (arrowheads) with processes (slender arrows) are locatednear the distal tip of the foot. Calibration bar in A equals approximately 57mm. Calibration bars in B and C equalapproximate 12mm.
325CATECHOLAMINES MODULATE METAMORPHOSIS
base of the velum on each side. At least one of these lattercells appeared to project an axon into the velum. An addi-tional 4 to 6 somata were located anterior to the eyes, inregions near and also more dorsal to the apical sensoryorgan. Although TH-LIR fibers were contained in pathwaysrunning through the pedal and cerebral ganglia, no TH-LIRsomata were located in these ganglia.
Enhancement of catecholamines in precompetent larvae
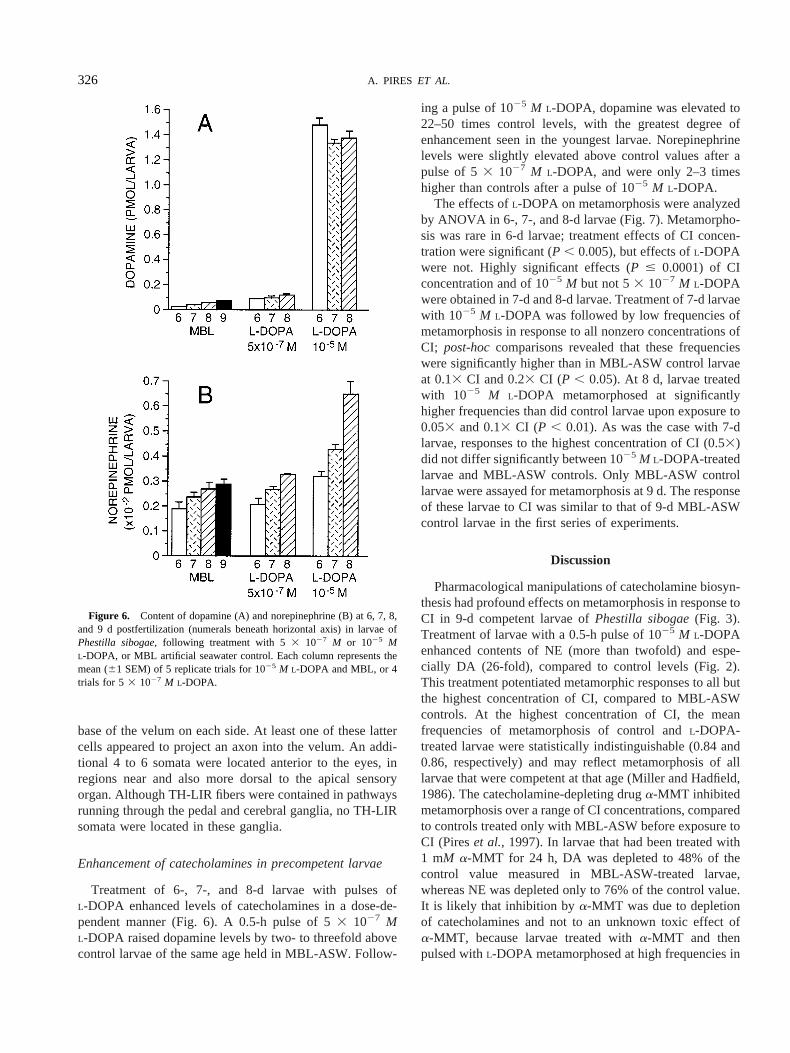
Treatment of 6-, 7-, and 8-d larvae with pulses ofL-DOPA enhanced levels of catecholamines in a dose-de-pendent manner (Fig. 6). A 0.5-h pulse of 53 1027 ML-DOPA raised dopamine levels by two- to threefold abovecontrol larvae of the same age held in MBL-ASW. Follow-
ing a pulse of 1025 M L-DOPA, dopamine was elevated to22–50 times control levels, with the greatest degree ofenhancement seen in the youngest larvae. Norepinephrinelevels were slightly elevated above control values after apulse of 53 1027 M L-DOPA, and were only 2–3 timeshigher than controls after a pulse of 1025 M L-DOPA.
The effects ofL-DOPA on metamorphosis were analyzedby ANOVA in 6-, 7-, and 8-d larvae (Fig. 7). Metamorpho-sis was rare in 6-d larvae; treatment effects of CI concen-tration were significant (P , 0.005), but effects ofL-DOPAwere not. Highly significant effects (P # 0.0001) of CIconcentration and of 1025 M but not 53 1027 M L-DOPAwere obtained in 7-d and 8-d larvae. Treatment of 7-d larvaewith 1025 M L-DOPA was followed by low frequencies ofmetamorphosis in response to all nonzero concentrations ofCI; post-hoccomparisons revealed that these frequencieswere significantly higher than in MBL-ASW control larvaeat 0.13 CI and 0.23 CI (P , 0.05). At 8 d, larvae treatedwith 1025 M L-DOPA metamorphosed at significantlyhigher frequencies than did control larvae upon exposure to0.053 and 0.13 CI (P , 0.01). As was the case with 7-dlarvae, responses to the highest concentration of CI (0.53)did not differ significantly between 1025 M L-DOPA-treatedlarvae and MBL-ASW controls. Only MBL-ASW controllarvae were assayed for metamorphosis at 9 d. The responseof these larvae to CI was similar to that of 9-d MBL-ASWcontrol larvae in the first series of experiments.
Discussion
Pharmacological manipulations of catecholamine biosyn-thesis had profound effects on metamorphosis in response toCI in 9-d competent larvae ofPhestilla sibogae(Fig. 3).Treatment of larvae with a 0.5-h pulse of 1025 M L-DOPAenhanced contents of NE (more than twofold) and espe-cially DA (26-fold), compared to control levels (Fig. 2).This treatment potentiated metamorphic responses to all butthe highest concentration of CI, compared to MBL-ASWcontrols. At the highest concentration of CI, the meanfrequencies of metamorphosis of control andL-DOPA-treated larvae were statistically indistinguishable (0.84 and0.86, respectively) and may reflect metamorphosis of alllarvae that were competent at that age (Miller and Hadfield,1986). The catecholamine-depleting druga-MMT inhibitedmetamorphosis over a range of CI concentrations, comparedto controls treated only with MBL-ASW before exposure toCI (Pireset al., 1997). In larvae that had been treated with1 mM a-MMT for 24 h, DA was depleted to 48% of thecontrol value measured in MBL-ASW-treated larvae,whereas NE was depleted only to 76% of the control value.It is likely that inhibition bya-MMT was due to depletionof catecholamines and not to an unknown toxic effect ofa-MMT, because larvae treated witha-MMT and thenpulsed withL-DOPA metamorphosed at high frequencies in
Figure 6. Content of dopamine (A) and norepinephrine (B) at 6, 7, 8,and 9 d postfertilization (numerals beneath horizontal axis) in larvae ofPhestilla sibogae,following treatment with 53 1027 M or 1025 ML-DOPA, or MBL artificial seawater control. Each column represents themean (61 SEM) of 5 replicate trials for 1025 M L-DOPA and MBL, or 4trials for 5 3 1027 M L-DOPA.
326 A. PIRES ET AL.

response to CI even in the continued presence ofa-MMT.Larvae that received this combined treatment ofL-DOPAanda-MMT had levels of DA and NE that were higher thanthose of MBL-ASW controls but lower than those measuredin larvae treated withL-DOPA only. The mechanism by
which a-MMT depletes endogenous DA, at least in mam-malian nervous systems, involves both inhibition of tyrosinehydroxylase activity bya-MMT and displacement of storedDA by a decarboxylated metabolite ofa-MMT (Uretsky etal., 1975; Bernabeet al.,1996). The latter mechanism mayhelp explain why larvae treated with botha-MMT andL-DOPA contained less DA and NE than those treated withL-DOPA alone. Taken together, our manipulations of cate-cholamine biosynthesis withL-DOPA anda-MMT in com-petent larvae show that depletion of endogenous cat-echolamines is accompanied by inhibition of CI-inducedmetamorphosis, whereas enhancement of catecholamines isaccompanied by potentiation of metamorphic responses toCI. Thus catecholamines appear to serve as endogenousmodulators of metamorphosis in competent larvae ofP.sibogae.
Catecholamine-containing cells in competent larvae werelabeled by FaGlu histochemistry (Fig. 4) in order to maplocations of these putative modulators of metamorphosis,and to test the anatomical specificity of catecholamine en-hancement byL-DOPA. Intensity of labeling and numbersof labeled cells were dramatically enhanced by treatment oflarvae with L-DOPA, without a concomitant increase inbackground fluorescence. Thus, the quantitative enhance-ment of catecholamine levels measured by HPLC in suchlarvae was localized in specific populations of cells. Thelocations of these cells were consistent with the locations ofcells labeled with antibodies to tyrosine hydroxylase (Fig.5). Immunocytochemistry combined with confocal micros-copy, however, revealed larger numbers of cells than didFaGlu histochemistry. The former method presumably la-beled a catecholamine-generative enzyme rather than cat-echolamines themselves, and yielded preparations withlower background fluorescence. Unfortunately, differencesin fixative solutions for FaGlu and immunocytochemistryprevented double-labeling of cells in this study. Nonethe-less, it remains likely that the cells labeled with FaGluwhose catecholamine contents were increased byL-DOPAtreatment were cells that are capable of synthesizing cat-echolamines, but normally contain concentrations below thelevel of detection by histochemistry.
The location and morphology of TH-LIR cells hint atmechanisms by which catecholamines might modulatemetamorphosis. Several of these cells are located near, andperhaps within, the apical sensory organ, which has beenimplicated as the chemosensory organ responsible for trans-duction of the CI stimulus (Kempfet al.,1997; Hadfieldetal., 2000). Future studies must further examine the exactlocations and morphologies of these cells, since they eithermay be chemosensory cells themselves or may modulatesynaptic interactions between primary chemosensory neu-rons and postsynaptic cells in a signaling pathway. Dopa-mine is known to modulate the activity of procerebralolfactory interneurons in the pulmonate molluscLimax
Figure 7. Frequency of metamorphosis of larvae ofPhestilla sibogaeafter 24-h exposure to varying dilutions of coral inducer, following 0.5-htreatment with 53 1027 M (squares) or 1025 M (triangles)L-DOPA, orMBL-artificial seawater control (circles) on day 6 (A), 7 (B), 8 (C), or 9 (D,control only) postfertilization. Each point represents the mean (61 SEM)of 5 replicate trials of 1025 M L-DOPA and MBL, or 4 trials for 53 1027
M L-DOPA. Asterisks indicate significant difference (*,P , 0.05; **, P ,0.01) from MBL artificial seawater control at a given dilution of coralinducer.
327CATECHOLAMINES MODULATE METAMORPHOSIS

maximus(Gelperinet al.,1993) as well as several aspects ofcentral olfactory processing in amphibians and mammals(Duchamp-Viretet al.,1997; Dotyet al.,1998). One or twopairs of TH-LIR cells were located at the base of the velum,and at least one pair appeared to send an axon distally intothe ipsilateral velar lobe. Dopamine inhibits ciliary beatingin gill epithelium of the bivalveMytilus edulis(Catapaneetal., 1978; Aielloet al.,1986) and also inhibits velar ciliarybeating in the larvae of this species (Beiras and Widdows,1995a). Dopaminergic innervation of the velum ofP. sibo-gaemight possibly serve to inhibit ciliary beating as part ofthe settlement response to CI that precedes the initiation ofmetamorphosis (Hadfield, 1978). Many cells exhibiting TH-LIR and aldehyde-induced histofluorescence were presentin the propodium of the foot. These cells possessed apicalprocesses extending to the surface, a characteristic of sen-sory neurons; in immunocytochemical preparations some ofthese same cells were also seen to have axonal projectionsto the pedal ganglia. Similar cells have been labeled byTH-LIR in anterior regions of the foot in embryonic andpostembryonic stages of the pulmonateLymnaea stagnalis,and by aldehyde-induced histoflourescene in ventral ante-rior regions of the foot in pediveligers of the bivalvesMytilus edulisandPlacopecten magellanicus(Croll et al.,1997; Voronezhskayaet al.,1999; Crollet al.,1999). InP.sibogaethe propodium becomes enlarged around the timeof metamorphic competence, mostly due to the develop-ment of mucus-secreting glands (Bonar and Hadfield,1974). Settlement in response to CI is accompanied bysecretion of mucus from the foot, and normally there isextensive contact between the propodium and the substra-tum. Therefore, potentiation of a sensory or motor functionof the foot associated with a settlement response to CI is ameans by which catecholaminergic cells might promote theinitiation of metamorphosis. Sensory neurons in the antero-lateral propodial ganglia of veligers of the nudibranchOn-chidoris bilamellataare depolarized by a soluble settlementcue that is derived from barnacle prey of adultO. bilamel-lata (Chia and Koss, 1989; Arkettet al.,1989). Neither suchcells nor such ganglia have been described in veligers ofP.sibogae,but we cannot rule out the possibility that homol-ogous cells in this species might have a role in chemosen-sory responses to CI.
Our final set of experiments investigated the relationshipbetween levels of endogenous catecholamines and the onsetof metamorphic competence inP. sibogae.Under our cul-ture conditions, larvae normally begin to attain competenceon day 7; most are competent on day 8, and nearly all arecompetent on day 10 (Miller and Hadfield, 1986). Previouswork had indicated that larval contents of DA and NEincrease severalfold during the period of 4 or 5 days be-tween hatching and attainment of maximal competence(Pires et al., 1997). In larvae of the bivalveCrassostreagigas,there is a sharp increase in NE content that coincides
with the onset of competence (Coon and Bonar, 1986); inthat species, dopamine and norepinephrine have been fur-ther implicated in the control, respectively, of settlementand metamorphosis (Coon and Bonar, 1987; Bonaret al.,1990; Beiras and Widdows, 1995b). These observations,together with the correlation of catecholamine enhancementand potentiation of metamorphosis in 9-d larvae in thepresent study, led us to ask whether the timing of compe-tence in larvae ofP. sibogaecould be influenced by phar-macological enhancement of catecholamines. Larvae aged6, 7, and 8 d were treated with either a low (53 1027 M)or high (1025 M) dose ofL-DOPA, or with MBL-ASW,then assayed for catecholamine contents (Fig. 6) and formetamorphic response to CI (Fig. 7) as in the first set ofexperiments. The higher dose was the same as that used inthe first set of experiments on 9-d competent larvae and alsoyielded dramatic increases in contents of catecholamines,especially DA, in younger larvae. Dopamine increased to asimilar high level, 1.3–1.5 pmol/larva, in 6-, 7-, and 8-dlarvae. These levels greatly exceed the range of valuesnormally found in larvae and juveniles ofP. sibogae(Pireset al.,1997), and indicate that the ability to convertL-DOPAto DA and store it is already well developed by 6 d. Thelower dose ofL-DOPA brought about much more modestenhancements of DA and NE within their normal physio-logical ranges, in most cases close to levels typically foundin older (9-d) competent larvae. Frequency of metamorpho-sis was always very low in 6-d larvae, never exceeding 0.1even at the highest concentration of CI, and was not signif-icantly affected by either dose ofL-DOPA. In 7-d and 8-dlarvae the higher dose but not the lower dose ofL-DOPAsignificantly potentiated metamorphosis in response to CI,compared to controls treated with MBL-ASW.
The results of these final experiments clearly indicate thatthe increases in catecholamine levels that naturally occuraround the time of competence, mimicked by exposure tothe lower dose ofL-DOPA, are not sufficient to confercompetence.Post-hocanalysis of metamorphosis at eachconcentration of CI in 7-d and 8-d larvae revealed that thehigher dose ofL-DOPA significantly increased frequenciesof metamorphosis at low concentrations of CI, but not at thehighest concentration. After thisL-DOPA treatment, theresponses of larvae to CI saturated at frequencies of meta-morphosis that were not significantly different from theresponses of MBL-ASW control larvae to the highest con-centration of CI. This suggests that potentiation of meta-morphosis byL-DOPA operated mostly in larvae that hadalready become competent. However, we cannot excludethe possibility that the apparent differences between 1025 ML-DOPA and MBL-ASW control treatments at the highestconcentration of CI in Figure 7B and C reflect a smallpositive effect of catecholamine enhancement on the onsetof competence.
How might catecholamines modulate metamorphosis in
328 A. PIRES ET AL.

competent larvae ofP. sibogaein nature? Both DA and NEare very weak inducers of metamorphosis when bath-ap-plied in the absence of CI (Pires and Hadfield, 1991).Therefore it seems unlikely that either of these serves as themajor neurotransmitter in a signaling pathway that directlymediates the induction of metamorphosis. The inhibition ofcoral-induced metamorphosis in larvae depleted of cat-echolamines bya-MMT (Fig. 3) suggests that catecholamin-ergic cells may modulate metamorphosis in the sense ofgating or enabling (Harris-Warrick, 1988) a primary signal-ing pathway. This sort of modulatory function has beendemonstrated in the serotonergic cerebral giant cells of thepulmonateLymnaea stagnalis,which enable activation of afeeding motor program by other interneurons (Yeomanetal., 1994). Pharmacological depletion of serotonin in thisspecies lowers the proportion of animals that respond to afood stimulus, and increases the latency and diminishes theintensity of responses that do occur (Kemeneset al.,1990).In similar fashion, depletion of catecholamines in modula-tory neurons in larvae ofP. sibogaemay lower the propor-tion of animals that respond to CI.
Potentiation of coral-induced metamorphosis ofL-DOPAwas observed only after enhancement of catecholaminecontents, especially DA, far beyond normal physiologicalvalues. Although we have demonstrated that these sub-stances load into specific populations of cells, we remainignorant of the extent of physiological release of cat-echolamines in these experiments and of the conditions thatfavor catecholamine release generally. Indeed, there is ev-idence in other systems that stored catecholamines are seg-regated into releasable and non-releasable pools (Kelly,1993; Reimeret al., 1998). Enhancement of release inL-DOPA-treated larvae would depend both on the presenceof some of the elevated catecholamine contents in releasablecellular pools and on the level of activity in those cells. Theenhancement of catecholamine release in our experimentsmay have been much more modest than the enhancement oftotal catecholamine contents, especially if we did not pro-vide adequate stimuli to evoke activity in the appropriatecells. Under natural conditions, such catecholaminergicmodulatory cells, carrying a normal complement of trans-mitter, might potentiate responses to CI by increasing theiractivity in the context of sensory cues or behaviors thatprecede or accompany natural settlement. The activity ofserotonergic modulatory cerebral giant cells of adult mol-luscs has been shown byin vivo recording to be strikinglydependent on behavioral context—for example, whether theanimal is quiescent, locomoting, or feeding, and whether ornot a food stimulus is present (Weisset al.,1978; Yeomanet al., 1994). Most assays of metamorphic responses tonatural inducers inP. sibogaeand other marine inverte-brates have been performed with filtered extracts of induc-ers in laboratory glassware under static conditions. Intrigu-ingly, responses to CI in larvae ofP. sibogaeare potentiated
by mechanical agitation of the assay dishes (Pecheniket al.,1995). More assays that better incorporate the sensory rich-ness of a larva’s experience leading up to natural metamor-phosis might uncover behavioral contexts that allow mod-ulation of responses to inducers.
Acknowledgments
This work was supported by Dickinson College, NaturalSciences and Engineering Research Council of Canadagrant OPG 38863, and U.S. Office of Naval Research grantN00014-95-1-1015.
Literature Cited
Aiello, E., E. Hager, C. Akiwumi, and G. B. Stefano. 1986. An opioidmechanism modulates central and not peripheral dopaminergic controlof ciliary activity in the marine musselMytilus edulis. Cell. Mol.Neurobiol.6: 17–30.
Arkett, S. A., F.-S. Chia, J. I. Goldberg, and R. Koss. 1989. Identifiedsettlement receptor cells in a nudibranch veliger respond to a specificcue.Biol. Bull. 176: 155–160.
Barlow, L. A. 1990. Electrophysiological and behavioral responses oflarvae of the red abalone (Haliotis rufescens) to settlement-inducingsubstances.Bull. Mar. Sci.46: 537–554.
Beiras, R., and J. Widdows. 1995a. Effects of the neurotransmittersdopamine, serotonin and norepinephrine on the ciliary activity ofmussel (Mytilus edulis) larvae.Mar. Biol. 122: 597–603.
Beiras, R., and J. Widdows, 1995b. Induction of metamorphosis inlarvae of the oysterCrassostrea gigasusing neuroactive compounds.Mar. Biol. 123: 327–334.
Bernabe, J., E. Proshlyakova, A. Sapronova, A. Trembleau, A. Calas,and M. Ugrumov. 1996. Pharmacological model of catecholaminedepletion in the hypothalamus of fetal and neonatal rats and its appli-cation.Cell. Mol. Neurobiol.16: 617–624.
Bonar, D. B., and M. G. Hadfield. 1974. Metamorphosis of the marinegastropodPhestilla sibogaeBergh (Nudibranchia: Aeolidacea). I. Lightand electron microscopic analysis of larval and metamorphic stages.L.Exp. Mar. Biol. Ecol.16: 227–255.
Bonar, D. B., S. L. Coon, M. Walch, R. M. Weiner, and W. Fitt. 1990.Control of oyster settlement and metamorphosis by endogenous andexogenous chemical cues.Bull. Mar. Sci.46: 484–498.
Catapane, E. J., G. B. Stefano, and E. Aiello. 1978.Pharmacologicalstudy of the reciprocal dual innervation of the lateral ciliated gillepithelium by the CNS ofMytilus edulis(Bivalvia). J. Exp. Biol.74:101–113.
Cavanaugh, G. M. 1956. Formulae and Methods VI of the MarineBiological Laboratory Chemical Room.Marine Biological Laboratory,Woods Hole, Massachusetts.
Chia, F.-S., and R. Koss. 1989. The fine structure of the newly discov-ered propodial ganglia of the veliger larva of the nudibranchOnchido-ris bilamellata. Cell Tissue Res.256: 17–26.
Coon, S. L., and D. B. Bonar. 1986. Norepinephrine and dopaminecontent of larvae and spat of the Pacific oyster,Crassostrea gigas. Biol.Bull. 171: 632–639.
Coon, S. L., and D. B. Bonar. 1987. Pharmacological evidence thatalpha-1 adrenoceptors mediate metamorphosis of the Pacific oysterCrassostrea gigas. Neuroscience23: 1169–1174.
Couper, J. M., and E. M. Leise. 1996. Serotonin injections inducemetamorphosis in larvae of the gastropod molluscIlyanassa obsoleta.Biol. Bull. 191: 178–186.
Croll, R. P., and B. J. Chiasson. 1990. Distribution of catecholaminesand of immunoreactivity to substances like vertebrate enzymes for the
329CATECHOLAMINES MODULATE METAMORPHOSIS

synthesis of catecholamines within the central nervous system of thesnail,Lymnaea stagnalis. Brain Res.525: 101–114.
Croll, R. P., D. L. Jackson, and E. E. Voronezhskaya. 1997.Cate-cholamine-containing cells in larval and postlarval bivalve molluscs.Biol. Bull. 193: 116–124.
Croll, R. P., E. E. Voronezhskaya, L. Hiripi, and K. Elekes. 1999.Development of catecholaminergic neurons in the pond snail,Lymnaeastagnalis: II. Postembryonic development of central and peripheralneurons.J. Comp. Neurol.404: 297–309.
Doty, R. L., C. Li, R. Bagla, W. Huang, C. Pfeiffer, G. M. Brosvic, andJ. M. Risser. 1998. SKF 38393 enhances odor detection perfor-mance.Psychopharmacology136: 75–82.
Duchamp-Viret, P., V. Coronas, J. C. Delaleu, E. Moyse, and A.Duchamp. 1997. Dopaminergic modulation of mitral cell activity inthe frog olfactory bulb: a combined radioligand binding-electrophysi-ological study.Neuroscience79: 203–216.
Elekes, K., G. Kemenes, L. Hiripi, M. Geffard, and P. R. Benjamin.1991. Dopamine-immunoreactive neurons in the central nervous sys-tem of the pond snailLymnaea stagnalis. J. Comp. Neurol.37: 214–224.
Froggett, S. J., and E. M. Leise. 1991. Metamorphosis in the marinesnail Ilyanassa obsoleta,yes or NO?Biol. Bull. 196: 57–62.
Furness, J. B., M. Costa, and W. W. Blessing. 1977.Simultaneousfixation and production of catecholamine fluorescence in central ner-vous tissue by perfusion with aldehydes.Histochem. J.9: 745–750.
Gelperin, A., L. D. Rhines, J. Flores, and D. W. Tank. 1993. Coherentnetwork oscillations by olfactory interneurons: modulation by endog-enous amines.J. Neurophysiol.69: 1930–1939.
Gibson, G. 1995. Why be choosy? Temporal changes in larval sensitivityto several naturally-occurring metamorphic inducers in the opistho-branchHaminaea callidegenita. J. Exp. Mar. Biol. Ecol.194: 9–24.
Giloh, H., and J. W. Sedat. 1982. Fluorescence microscopy: reducedphotobleaching of rhodamine and fluorescein protein conjugates byn-propyl gallate.Science217: 1252–1255.
Hadfield, M. G. 1978. Metamorphosis in marine molluscan larvae: ananalysis of stimulus and response. Pp. 165–175 inSettlement andMetamorphosis of Marine Invertebrate Larvae,F.-S. Chia and M. Rice,eds. Elsevier/North-Holland, New York.
Hadfield, M. G., and J. T. Pennington. 1990. Nature of the metamor-phic signal and its internal transduction in larvae of the nudibranchPhestilla sibogae. Bull. Mar. Sci.46: 455–464.
Hadfield, M. G., and D. Scheuer. 1985. Evidence for a soluble meta-morphic inducer inPhestilla sibogae:ecological, chemical and biolog-ical data.Bull. Mar. Sci.37: 556–566.
Hadfield, M. G., and M. F. Strathmann. 1996. Variability, flexibilityand plasticity in life histories of marine invertebrates.Oceanol. Acta19: 323–334.
Hadfield, M. G., E. A. Meleshkevitch, and D. Y. Boudko. 2000. Theapical sensory organ of a gastropod veliger is a receptor for settlementcues.Biol. Bull. 198: 67–76.
Harris-Warrick, R. M. 1988. Chemical modulation of central patterngenerators. Pp. 285–331 inNeural Control of Rhythmic Movements inVertebrates,A. H. Cohen, S. Rossignol, and S. Grillner, eds. Wiley,New York.
Hernadi, L., S. Juhos, and K. Elekes. 1993. Distribution of tyrosine-hydroxylase-immunoreactive and dopamine-immunoreactive neuronsin the central nervous system of the snail,Helix pomatia. Cell TissueRes.274: 503–513.
Kabotyanski, E. A., D. A. Baxter, and J. H. Byrne. 1998. Identificationand characterization of catecholaminergic neuron B65, which initiatesand modifies patterned activity in the buccal ganglia ofAplysia. J. Neu-rophysiol.79: 605–621.
Katz, P. S. 1995. Intrinsic and extrinsic neuromodulation of motorcircuits.Curr. Opin. Neurobiol.5: 799–808.
Kelly, R. B. 1993. Storage and release of neurotransmitters.Cell 72/Neuron10 suppl.: 43–53.
Kemenes, G., L. Hiripi, and P. R. Benjamin. 1990. Behavioural andbiochemical changes in the feeding system ofLymnaeainduced by thedopamine and serotonin neurotoxins 6-hydroxydopamine and 5,6-di-hydroxytryptamine.Philos. Trans. R. Soc. Lond. B Biol. Sci.329:243–255.
Kempf, S. C., and M. G. Hadfield. 1985. Planktotrophy by the lecitho-trophic larvae of a nudibranch,Phestilla sibogae(Gastropoda).Biol.Bull. 169: 119–130.
Kempf, S. C., G. V. Chun, and M. G. Hadfield, 1992. An immunocy-tochemical search for potential neurotransmitters in larvae ofPhestillasibogae (Gastropoda, Opisthobranchia).Comp. Biochem. Physiol.101C: 299–305.
Kempf, S. C., L. R. Page, and A. Pires. 1997. Development of seroto-nin-like immunoreactivity in the embryos and larvae of nudibranchmollusks with emphasis on the structure and possible function of theapical sensory organ.J. Comp. Neurol.386: 507–528.
Miller, S. E., and M. G. Hadfield. 1986. Ontogeny of phototaxis andmetamorphic competence in larvae of the nudibranchPhestilla sibogaeBergh (Gastropoda: Opisthobranchia).J. Exp. Mar. Biol. Ecol.97:95–112.
Norekyan, T. P., and R. Satterlie. 1993. Neuronal analysis of huntingbehavior of the pteropod molluscClione limacina. Neurosci. Behav.Physiol.23: 11–23.
Pechenik, J. A. 1990. Delayed metamorphosis by larvae of benthicmarine invertebrates: Does it occur? Is there a price to pay?Ophelia32: 63–94.
Pechenik, J. A., M. G. Hadfield, and L. S. Eyster. 1995. Assessingwhether larvae of the opisthobranch gastropodPhestilla sibogaeBerghbecome responsive to three chemical cues at the same age.J. Exp. Mar.Biol. Ecol. 191: 1–17.
Pechenik, J. A., K. Hammer, and C. Weise. 1996. The effect ofstarvation on acquisition of competence and postmetamorphic perfor-mance in the marine prosobranch gastropodCrepidula fornicata. J.Exp. Mar. Biol. Ecol.199: 137–152.
Pires, A., and M. G. Hadfield. 1991. Oxidative breakdown products ofcatecholamines and hydrogen peroxide induce partial metamorphosisin the nudibranchPhestilla sibogaeBergh (Gastropoda: Opistho-branchia).Biol. Bull. 180: 310–317.
Pires, A., S. L. Coon, and M. G. Hadfield. 1997. Catecholamines anddihydroxyphenylalanine in metamorphosing larvae of the nudibranchPhestilla sibogaeBergh.J. Comp. Physiol. A181: 187–194.
Quinlan, E. M., B. C. Arnett, and A. D. Murphy. 1997. Feedingstimulants activate an identified dopaminergic interneuron that inducesthe feeding motor program inHelisoma. J. Neurophysiol.78: 812–824.
Reimer, R. J., E. A. Fon, and R. H. Edwards. 1998. Vesicular neuro-transmitter transport and the presynaptic regulation of quantal size.Curr. Opin. Neurobiol.8: 405–412.
Trapido-Rosenthal, H., and D. E. Morse. 1985. L-a, v-diamino acidsfacilitate GABA induction of larval metamorphosis in a gastropodmollusc (Haliotis rufescens). J. Comp. Physiol. B155: 403–414.
Trapido-Rosenthal, H., and D. E. Morse. 1986a. Availability of che-mosensory receptors is down-regulated by habituation of larvae to amorphogenetic signal.Proc. Natl. Acad. Sci. USA83: 7658–7662.
Trapido-Rosenthal, H., and D. E. Morse. 1986b. Regulation of recep-tor-mediated settlement and metamorphosis in larvae of a gastropodmollusc (Haliotis rufescens). Bull. Mar. Sci.39: 383–392.
Trimble, D. L., and D. L. Barker. 1984. Activation by dopamine ofpatterned motor output from the buccal ganglia ofHelisoma trivolvis.J. Neurobiol.15: 37–48.
Uretsky, N. J., S. R. Snodgrass, and A. V. Lorenzo. 1975.Studies onthe mechanism of depletion of striatal dopamine bya-methyl-m-ty-rosine.J. Pharmacol. Exp. Ther.195: 465–479.
330 A. PIRES ET AL.

Voronezhskaya, E. E., L. Hiripi, K. Elekes, and R. P. Croll. 1999.Development of catecholaminergic neurons in the pond snail,Lymnaeastagnalis:I. Embryonic development of dopamine-containing neuronsand dopamine-dependent behaviors.J. Comp. Neurol.404: 285–296.
Weiss, K. R., J. L. Cohen, and I. Kupfermann. 1978. Modulatorycontrol of buccal musculature by a serotonergic neuron (metacerebralcell) in Aplysia. J. Neurophysiol.41: 181–203.
Wieland, S. J., and A. Gelperin. 1983. Dopamine elicits feeding motorprogram inLimax maximus. J. Neurosci.3: 1735–1745.
Yeoman, M. S., A. W. Pieneman, G. P. Ferguson, A. Ter Maat, and
P. R. Benjamin. 1994. Modulatory role for the serotonergic cerebralgiant cells in the feeding system of the snail,Lymnaea.I. Fine wirerecording in the intact animal and pharmacology.J. Neurophysiol.72:1357–1371.
Zimmerman, K. M., and J. A. Pechenik. 1991. How do temperatureand salinity affect relative rates of growth, morphological differentia-tion, and time to metamorphic competence in larvae of the marinegastropodCrepidula plana? Biol. Bull. 180: 372–386.
Zolman, J. F. 1993. Biostatistics: Experimental Design and StatisticalInference.Oxford University Press, New York.
331CATECHOLAMINES MODULATE METAMORPHOSIS




















