
Histochemical localization of FMRFamide, serotonin and catecholamines in embryonic Crepidula...
14
&p.1:Abstract Recent reports indicate that neuronal elements develop in early larval stages of some Gastropoda from the Pulmonata and Opisthobranchia prior to the appear- ance of any ganglia of the future adult central nervous system (CNS). The present study describes similar early neuronal elements in Crepidula fornicata. A posterior FMRFamide-like immunoreactive (LIR) cell with anteri- orly projected fibers was observed in the trochophore stage. Additional FMRFamide-LIR and serotonin-LIR cells and fibers were found in the apical organ in the tro- chophore and early veliger stages. FMRFamide-LIR and serotonin-LIR projections to the velum and foot were also detected at this time. As the veliger developed, pe- ripheral FMRFamide-LIR and later catecholaminergic cells were located in the foot region. Also during this stage, catecholaminergic cells and processes were ob- served near the mouth. In addition, this study tentatively identified the first serotonin- and FMRFamide-LIR cells and fibers within the developing ganglia of the adult CNS, which appeared in close proximity to the earlier developing elements. These observations are consistent with the hypothesis that, in addition to its presumed role in the control of larval behaviors, the larval nervous system guides the development of the adult CNS. Larvae from the class Bivalvia and other invertebrate phyla also have neuronal elements marked by the presence of FMRFamide, serotonin, and catecholamines, and, there- fore, this study may provide additional insights into phy- logenetic relationships of the Gastropoda with other rep- resentatives of the Mollusca and different invertebrate phyla.&bdy: A. Introduction Several recent reports have described neuronal elements which appeared prior to the first cells of the cerebral or pedal ganglia in the Mollusca. For example, Croll and Voronezhskaya (1995, 1996a) identified neuronal ele- ments in the trochophore or early veliger stage of Lym- naea stagnalis (Linnaeus, 1758) (Pulmonata) using anti- bodies raised against the neuropeptide, Phe-Met-Arg-Phe- NH 2 (FMRFamide; Greenberg and Price 1992). FMRF- amide-like immunoreactive (LIR) cells were shown to de- velop in posterior regions of the embryo and to send pro- jections in an anterior direction. These projections termi- nated in the region of the future cerebral and pedal ganglia and near two large, flask-shaped FMRFamide-LIR apical cells. Additional neurons in the foot and mouth region were also revealed in L. stagnalis using techniques to lo- calize catecholamines (Voronezhskaya et al. 1999). Croll and Voronezhskaya (1996b) also identified FMRFamide- LIR elements in posterior regions in the trochophore stage from Aplysia californica (Cooper, 1863) (Opisthobrancia). In a slightly later stage of A. californica, Marois and Carew (1997a–c) revealed serotonin-LIR cells that com- prise part of an apical organ. Kempf et al. (1997) have shown that such apical cells appear to be general features of larvae from species of the Opisthobranchia. The FMRFamide-LIR and serotonin-LIR cells in these various species appeared in early stages prior to the first cells of the cerebral ganglia. Neuronal elements located outside the region of the future adult central nervous system (CNS) have also been identified in early larval stages from the Bivalvia using techniques to localize acetylcholinester- ase, catecholamines, and serotonin (Croll et al. 1997; Ku- lakovskiy and Flyachinskaya 1994; Raineri 1995; Raineri and Ospovat 1994). The presence of such early elements suggests the possibility that the Mollusca possess larval nervous systems which appear early in development and can become relatively extensive, but that have gone large- ly unrecognized in the past. Studies of early neuronal elements have been per- formed with species from the Pulmonata and Opistho- A.J.G. Dickinson ( ✉ ) · R.P. Croll Department of Biology, Dalhousie University, Halifax Nova Scotia, Canada e-mail: [email protected] A.J.G. Dickinson · J. Nason · R.P. Croll Department of Physiology and Biophysics, Dalhousie University Halifax, Nova Scotia, Canada&/fn-block: Zoomorphology (1999) 119:49–62 © Springer-Verlag 1999 ORIGINAL ARTICLE &roles:Amanda J.G. Dickinson · Janette Nason Roger P. Croll Histochemical localization of FMRFamide, serotonin and catecholamines in embryonic Crepidula fornicata (Gastropoda, Prosobranchia) &misc:Accepted: 10 February 1999
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Histochemical localization of FMRFamide, serotonin and catecholamines in embryonic Crepidula...

&p.1:Abstract Recent reports indicate that neuronal elementsdevelop in early larval stages of some Gastropoda fromthe Pulmonata and Opisthobranchia prior to the appear-ance of any ganglia of the future adult central nervoussystem (CNS). The present study describes similar earlyneuronal elements in Crepidula fornicata. A posteriorFMRFamide-like immunoreactive (LIR) cell with anteri-orly projected fibers was observed in the trochophorestage. Additional FMRFamide-LIR and serotonin-LIRcells and fibers were found in the apical organ in the tro-chophore and early veliger stages. FMRFamide-LIR andserotonin-LIR projections to the velum and foot werealso detected at this time. As the veliger developed, pe-ripheral FMRFamide-LIR and later catecholaminergiccells were located in the foot region. Also during thisstage, catecholaminergic cells and processes were ob-served near the mouth. In addition, this study tentativelyidentified the first serotonin- and FMRFamide-LIR cellsand fibers within the developing ganglia of the adultCNS, which appeared in close proximity to the earlierdeveloping elements. These observations are consistentwith the hypothesis that, in addition to its presumed rolein the control of larval behaviors, the larval nervoussystem guides the development of the adult CNS. Larvaefrom the class Bivalvia and other invertebrate phyla alsohave neuronal elements marked by the presence ofFMRFamide, serotonin, and catecholamines, and, there-fore, this study may provide additional insights into phy-logenetic relationships of the Gastropoda with other rep-resentatives of the Mollusca and different invertebratephyla.&bdy:
A. Introduction
Several recent reports have described neuronal elementswhich appeared prior to the first cells of the cerebral orpedal ganglia in the Mollusca. For example, Croll andVoronezhskaya (1995, 1996a) identified neuronal ele-ments in the trochophore or early veliger stage of Lym-naea stagnalis(Linnaeus, 1758) (Pulmonata) using anti-bodies raised against the neuropeptide, Phe-Met-Arg-Phe-NH2 (FMRFamide; Greenberg and Price 1992). FMRF-amide-like immunoreactive (LIR) cells were shown to de-velop in posterior regions of the embryo and to send pro-jections in an anterior direction. These projections termi-nated in the region of the future cerebral and pedal gangliaand near two large, flask-shaped FMRFamide-LIR apicalcells. Additional neurons in the foot and mouth regionwere also revealed in L. stagnalisusing techniques to lo-calize catecholamines (Voronezhskaya et al. 1999). Crolland Voronezhskaya (1996b) also identified FMRFamide-LIR elements in posterior regions in the trochophore stagefrom Aplysia californica(Cooper, 1863) (Opisthobrancia).In a slightly later stage of A. californica, Marois andCarew (1997a–c) revealed serotonin-LIR cells that com-prise part of an apical organ. Kempf et al. (1997) haveshown that such apical cells appear to be general featuresof larvae from species of the Opisthobranchia. TheFMRFamide-LIR and serotonin-LIR cells in these variousspecies appeared in early stages prior to the first cells ofthe cerebral ganglia. Neuronal elements located outsidethe region of the future adult central nervous system(CNS) have also been identified in early larval stages fromthe Bivalvia using techniques to localize acetylcholinester-ase, catecholamines, and serotonin (Croll et al. 1997; Ku-lakovskiy and Flyachinskaya 1994; Raineri 1995; Raineriand Ospovat 1994). The presence of such early elementssuggests the possibility that the Mollusca possess larvalnervous systems which appear early in development andcan become relatively extensive, but that have gone large-ly unrecognized in the past.
Studies of early neuronal elements have been per-formed with species from the Pulmonata and Opistho-
A.J.G. Dickinson (✉) · R.P. CrollDepartment of Biology, Dalhousie University, HalifaxNova Scotia, Canadae-mail: [email protected]
A.J.G. Dickinson · J. Nason · R.P. CrollDepartment of Physiology and Biophysics, Dalhousie UniversityHalifax, Nova Scotia, Canada&/fn-block:
Zoomorphology (1999) 119:49–62 © Springer-Verlag 1999
O R I G I N A L A RT I C L E
&roles:Amanda J.G. Dickinson · Janette NasonRoger P. Croll
Histochemical localization of FMRFamide,serotonin and catecholamines in embryonic Crepidula fornicata(Gastropoda, Prosobranchia)
&misc:Accepted: 10 February 1999

branchia as well as the Bivalvia. However, species fromother taxa must be examined to make generalizations andcomparisons of neuronal development across the mollus-can phylum. One important molluscan group, which hasnot yet been examined extensively, is the Prosobranchia.The Prosobranchia differ from most of the Opisthobran-chia and the Pulmonata in terms of the gross anatomy ofthe adult nervous system by having chiastoneury, thecrossing of pleural–visceral connectives (Bullock andHorridge 1965). In addition, the neuronal cell bodies ofthe Prosobranchia are relatively small as compared to thegiant cell bodies found in the nervous system of the Pul-monata and Opisthobranchia (Bullock and Horridge1965). Such differences in the adult nervous systemsmay reflect changes in the morphology of the larval ner-vous systems. It is also believed that the Prosobranchiaare more representative of ancestral forms than the otherGastropoda (see Graham 1983). Thus, the morphologyof their larval nervous system may provide important in-sights into the phyletic position of the major molluscangroups. To date, the few neurodevelopmental studies ofmembers of the Prosobranchia have focused mostly onneurotransmitter phenotypes in later veliger stages (Bar-low and Truman 1992; Lin and Leise 1996b) with littleor no attention having been paid to younger specimens.
The purpose of this present study is to examine thedevelopment of early neuronal elements in Crepidulafornicata (Linné, 1758) (Prosobranchia). General embry-onic development of C. fornicatawas first reported byConklin (1897). Here, we focus on neuronal elements,which appear in embryonic stages, from first cleavage tothe time of hatching, using histological techniques to lo-calize FMRFamide, serotonin, and catecholamines. Allthree of these markers have previously revealed neuronsand fibers that possibly comprise the early larval nervoussystem in the Pulmonata and the Opisthobranchia and,therefore, would allow comparisons across the Gastropo-da. FMRFamide, serotonin, and catecholamines havealso been localized in early larval stages from the Bival-via (Croll et al. 1997; Kulakovskiy and Flyachinskaya1994) as well as other invertebrate phyla such as thePhoronida and the Polychaeta (for example, Hay-Schmidt 1990a–c; 1995) and, therefore, extend the possi-bility of such comparisons into the Mollusca and inverte-brate phyla.
B. Materials and methods
I. Animals
Embryos used in this study were gathered from brooding femaleslocated on the upper valve of locally gathered sea scallops andkept in the Aquatron facilities at Dalhousie University. Embryosgrow inside delicate egg capsules, which are united by a commonstem and covered by the parent’s shell (Conklin 1897). Each eggcapsule contains an albuminous fluid surrounding 200–300 em-bryos (Conklin 1897). Once removed from the adult, the eggclutches were kept at room temperature (22–25°C) in small con-tainers of artificial sea water (Instant Ocean) changed every 48 h.Sample specimens were taken every 12–24 h from each eggclutch. Under these conditions embryos generally took 11–12 days
to develop from first cleavage stage to the time of hatching. Thehatched veliger generally required another few weeks of life be-fore becoming competent to metamorphose into the juvenile stage(Conklin 1897; Pechenik 1984).
Although, embryonic development of C. fornicatawas previ-ously examined in detail (Conklin 1897), the timing of develop-mental stages was not provided. In this study, developmental stag-ing is based on the percentage of embryonic development elapsedbetween the first cleavage stage (0%=E0) and hatching(100%=E100). Morphological and behavioral features were exam-ined at regular intervals throughout embryogenesis. Embryonicdevelopment is divided into four stages: trochophore stage, pretor-sion veliger stage, early posttorsion veliger stage, and late posttor-sion veliger stage. Each stage is marked by general morphologicalchanges of the embryo and significant changes in the morphologyof neuronal elements.
II. Immunocytochemistry
Immunohistological procedures were performed according toCroll and Chiasson (1989) and Marois and Croll (1992). Embryoswere removed from their egg capsules and then fixed for 2–3 h in4% paraformaldehyde in phosphate-buffered saline (PBS; 50 mMNa2HPO4
.7H20 and 140 mM NaCl in distilled water adjusted topH 7.2) at room temperature for serotonin and FMRFamide immu-nocytochemistry or in cold (–20°C) methanol for tyrosine hydrox-ylase (TH) immunocytochemistry. They were then washed for5 min in PBS 2 or 3 times. The shells of older embryos (>E55)were decalcified with 10% EDTA in PBS for 30–45 min (Kempfet al. 1997; Marois and Carew 1997a–c). Embryos were nextwashed for 2–3 h in 4% Triton X-100 in PBS. The embryos that
50
Fig. 1A–C Schematic representation of Crepidula fornicataat thelate trochophore stage (E30–32). Figures on the leftrepresent rightside views and figures on the rightrepresent a frontal/dorsal orien-tation. A General morphology. B The FMRFamide-like immuno-reactive (LIR) posterior cell (FPC) with projections to the anteriorregion, and fibers forming an apical commissure (acm) in the ante-rior region. C Right and left serotonin-LIR apical cells (rSAC,lSAC), acm, and posteriorly projecting fibers&/fig.c:

had been fixed in 4% paraformaldehyde were next incubated inpolyclonal antibodies raised against FMRFamide or serotonin(both obtained from Incstar, Stillwater, Wis.). These antibodieswere diluted 1:500 in PBS with the addition of 1.0 normal goat se-rum (NGS) and 1% Triton X-100. Incubation periods lasting 48 hat 4°C or 12 h at room temperature gave comparable results. Theembryos were next rinsed 3 times with PBS (5 min each) and giv-en a final wash for 1 h before incubating in goat anti-rabbit anti-bodies conjugated to fluorescein isothiocyanate (FITC) or rhoda-mine. These secondary antibodies were obtained from Bio/CanScientific (Mississauga, Ontario) and diluted 1:50 in PBS with theaddition of 1% NGS and 1% Triton X-100. Double-labeling exper-iments were also performed with monoclonal antibodies raisedagainst serotonin obtained from Dimension Laboratories (Mississ-auga, Ontario). These latter antibodies were diluted 1:50 in PBSwith the addition of 1% normal sheep serum (NSS) and 1% TritonX-100. Incubation periods were the same as described above. Theembryos were next rinsed 3 times with PBS (5 min each) and giv-
en a final wash for 1 h before incubating in sheep anti-mouse seraconjugated to FITC or rhodamine. These secondary antibodieswere diluted 1:50 in PBS with the addition of 1% NGS and 1%Triton X-100.
The embryos that had been fixed with methanol were subse-quently washed and processed as described above, but were incu-bated in monoclonal antibodies raised against rat TH. TH is in-volved in the first step of catecholamine biosynthesis and, there-fore, its antibodies are used to indicate the presence of catechola-mines in tissues (Baldessarini 1996; Cooper et al. 1996). TH anti-bodies, obtained from Incstar, were diluted 1:50 in PBS with theaddition of 1% NSS and 1% Triton X-100.
All embryos processed for immunocytochemistry were mount-ed on glass slides in 3:1 glycerol to PBS for viewing on a LeitzAristoplan microscope equipped for epifluorescence. FITC fluo-rescence was viewed using a 450–490 nm excitation filter and a525/20-nm barrier filter, while rhodamine fluorescence wasviewed using a 530 to 560-nm excitation filter and a 580-nm long-pass barrier filter. Specimens were photographed with KodakTMAX 100 film.
As negative controls, embryos were processed without incuba-tion in primary antibody; such specimens exhibited no detectablefluorescence in somata or neurites except in the final stages beforehatching when large cells located along the rim of the velum wereobserved to be autofluorescent. Positive controls involved parallelprocessing of embryonic L. stagnalis which exhibited typicalstaining as described elsewhere (Croll and Chiasson 1989; Crolland Voronezhskaya 1996a, b; Marois and Croll 1992; Voronezhs-kaya et al. 1999).
51
Fig. 2A–D Neuronal elements present in the late trochophorestage (E30–32) of C. fornicata. A Right side view of an embryoshowing the FPC and fibers in the anterior region (acm). B Highermagnification showing the FPC and its fiber (arrows) in the vicin-ity of the kidney. C Right side view of an embryo showing therSAC, adjacent to it is the lSAC out of the plane of focus and theacm. Fibers extending in a posterior direction (arrows) are also in-dicated. D Dorsal orientation of an embryo showing both the lSACand right rSACand connecting fibers constituting an acm&/fig.c:

III. Catecholamine histofluorescence
The FaGlu technique of Furness et al. (1977) was also used to lo-calize catecholamines. Embryos were placed in a solution consist-ing of 4% paraformaldehyde, 0.5% glutaraldehyde, and 35% su-crose in PBS. Embryos were incubated in this FaGlu solution forat least 12 h, but were also found to remain stable for up to severalweeks. Embryos were then placed on glass slides, air dried forseveral hours, and then mounted in 3:1 glycerol:PBS. These em-bryos were viewed and photographed through the Leitz micro-scope equipped with a 355 to 425-nm excitation filter and a 460-nm long-pass barrier filter.
C. Results
I. Trochophore stage (E23–35)
The early trochophore stage embryo became mobile fol-lowing the development of prototrochal, pedal, and sto-modeal cilia. In addition, the shell gland and larval kid-neys formed during this stage (Fig. 1A).
1. FMRFamide-like immunoreactivity
As the embryo entered the trochophore stage (E22–24), asingle, FMRFamide-LIR cell appeared in its posterior re-gion. This FMRFamide-LIR posterior cell (FPC) was po-sitioned just ventral and to the right of the shell gland(Figs. 1B, 2A,B) and single processes projected anterior-ly from each side of the cell body. The fibers followedthe curvature of the body wall on each side and reachedthe midlength of the embryo near the larval kidney,which was also labeled with the anti-FMRFamide serum(Fig. 2A,B). Later in the trochophore stage (E30–32) theanteriorly projecting fibers from the FPC reached the an-terior region of the larva (Figs. 1B, 2A), where theycrossed the midline and ramified to form a prominentapical commissure (acm; Figs. 1B, 2A).
2. Serotonin-like immunoreactivity
The earliest detection of serotonin-like immunoreactivityoccurred in a late trochophore stage (E30–35; Figs. 1C,2C). A bilaterally symmetric pair of serotonin-LIR cellsappeared in the anterior region. These serotonin-LIR api-cal cells (SACs) were interconnected medially by a fewserotonin-LIR fibers, which appeared to intermingle withthe FMRFamide-LIR fibers within the acm (Figs. 1C,2D). Each SAC soma also extended a posterior projec-tion to the midlength of the embryo at a point where theyconverged upon the anteriorly directed FMRFamide-LIRfibers originating from the FPC (Figs. 1C, 2C).
II. Pretorsion veliger stage (E35–55)
As the embryo entered the veliger stage, the velar lobesand the foot began to grow. In addition, eyespots ap-peared at this stage (Fig. 3A).
1. FMRFamide-like immunoreactivity
By the early veliger stage (E35–37) the FPC became lessintensely labeled and its fibers became difficult to followinto anterior regions of the embryo. In contrast, fivemore brightly labeled FMRFamide-LIR cells appearedover the acm and are collectively referred to as FMRF-amide-LIR apical cells (FACs; Figs. 3B, 4A). A single,flask-shaped medial FAC (mFAC) appeared on the acmand a short process extended from this cell to the epithe-lium where it terminated. Two smaller, flask-shapedFACs (FAC2 and FAC3) appeared on each side of themFAC. These paired cells also had short processes ex-tending from their apices but were more intensely la-beled than the mFAC.
A loop of what appeared to be one or two fibers ranposteriorly and ventrally from each side of the acm. Thisloop extended to about the midlength of the embryo andby E40 it was displaced slightly to the right side(Fig. 3B). Also at this later time, two faintly labeled
52
Fig. 3A–C Schematic representation of C. fornicataat a pretor-sion veliger stage (E55). Figures on the leftrepresent right sideviews and figures on the rightrepresent a frontal/dorsal orienta-tion. The anterior region is to the right in each figure. A Generalmorphology. B Five FMRFamide-LIR apical cells (FACs). Themedial FAC (mFAC) and the two right FACs (rFAC2, 3) are la-belled. Two FMRFamide-LIR abdominal loop cells (FALC1, 2) arelocated in the middle region of the embryo and are connected byimmunoreactive fibers. Flask-shaped FMRFamide-LIR foot cells(FFC) are indicated. The FPC is present but its anteriorly directedprocesses (dashed line) are faint and often undetectable. C Rightand left serotonin-LIR cerebral cells (rSCC, lSCC) are located lat-eral to the SACs. The mFACalso appears to be very faintly seroto-nin-LIR. Immunoreactive fibers project to the foot and velar re-gions&/fig.c:

FMRFamide-LIR abdominal loop cells (FALC1 andFALC2) appeared on the right side of the posterior por-tion of the loop (Fig. 3B). During the early veliger stage(E35–37) fibers appeared in the developing foot. By E55there was increased branching in the foot region andFMRFamide-LIR foot cells (FFCs) appeared along theanterior margin of the appendage (Figs. 3B, 4B).
2. Serotonin-like immunoreactivity
During the early veliger stage (E35–37), the mFAC alsoappeared to be faintly serotonin-LIR (Fig. 3C). In addi-tion, serotonin-LIR fibers also began to appear in the ex-panding velar lobes (Fig. 3C). By E55, a new pair of
small, faintly labeled serotonin-LIR cells appeared later-al and slightly ventral to the acm within the region of thedeveloping cerebral ganglia. These cells are, therefore,tentatively identified as Serotonin-LIR cerebral cells(SCCs; Figs. 3C, 4D). Projections extended from eachSCC to the commissure near the SACs (Fig. 3C).
III. Early post torsion veliger stage (E55–65)
At approximately E55, the embryo underwent torsionwhich resulted in the 180° clockwise rotation of the viscer-al mass and mantle relative to the head–foot axis. The ve-lar lobes increased in size and ciliation (Fig. 5A). Duringthis time, velar cilia beat regularly as the embryo swamwithin the egg capsule. Cells and processes observed dur-ing this stage persisted until the time of hatching and,therefore, can be seen in the figures of later stages.
1. FMRFamide-like immunoreactivity
Torsion caused the twisting of the FMRFamide-LIR fi-bers in the middle region of the embryo resulting in theformation of a figure-eight pattern (Figs. 5B, 7A).FALC1 and FALC2, originally on the right, assumed
53
Fig. 4A–D Neuronal elements present in a pretorsion veligerstage C. fornicata. A Dorsal orientation of an embryo showing thefive (rFAC2, 3, lFAC 2, 3, mFAC) FACs near the acm. B Ventralorientation of the foot with the anterior region at the top of the fig-ure. The flask-shaped FFCs (FFC) are indicated. C Dorsal orienta-tion of an embryo revealing right and left SACs (rSAC, lSAC) nearthe acm. Arrows indicate fibers which extend into the posterior re-gion. D Higher magnification of a dorsal orientation of the anteri-or region of an embryo showing the SCCs (rSCC, lSCC) near theeyes. In this figure, the SACs are not in focus. Arrows indicate fi-bers which extend into the posterior region&/fig.c:

54
Fig. 5A–C Schematic representation of C. fornicata at an earlyposttorsion veliger stage (E65). Figures on the leftdepict rightside views and figures on the rightdepict a frontal/dorsal orienta-tion. A General morphology. B The figure-eight pattern formed bythe FMRFamide-LIR fibers at the midlength of the embryo is indi-cated. The FALCs (FALC1, 2) are located on the left side and anadditional FALC (FALC3) is indicated. Projections into the velarregion and additional FFCsare shown in the foot region. C A fig-ure-eight pattern of serotonin-LIR fibers at the midlength of theembryo is shown. Also a pair of SSCs (lSCC, rSCC) have ap-peared lateral to the SACs and each has a projection to the acm.Projections into the velar region are also indicated&/fig.c:
Fig. 6A–D Schematic representation of C. fornicata at a stagejust prior to hatching (E 100). Figures on the leftdepict right sideviews and figures on the rightdepict a frontal/dorsal orientation. AGeneral morphology. B An additional FALC (FALC4) is indicated.An additional pair of cells, the FMRFamide-LIR pedal cells(FPeC) are shown just posterior and ventral to the FACs. Also,FMRFamide-LIR innervations to the velum and foot are moreelaborate. C Two serotonin-LIR abdominal loop cells (SALCs) areshown, SALC1on the right and SALC2on the left side of the em-bryo. Serotonin-LIR innervation to the velum and foot are alsomore elaborate and form a similar pattern to the FMRFamide-LIRfibers. D The catacholaminergic oral cells (COC) are indicated.One large pair of flask-shaped cells and a few smaller pairs areshown to be interconnected with fibers. In addition, flask-shapedcatacholaminergic foot cells (CFC) are also shown, one medialpair sends fibers to the statocysts&/fig.c:
figure eight (Figs. 5B, 7A). There was increased innerva-tion to the foot region, but the FFCs became fainter byE55–60 and then undetectable after E65. From E55 untilthe time of hatching, there was increased innervation ofthe velar lobes (Fig. 5B). FMRFamide-LIR fibers origi-nating at the base of the acm projected into each velarlobe and branched repeatedly.
2. Serotonin-like immunoreactivity
During early posttorsion stages the serotonin-LIR fibersformed similar patterns to the FMRFamide-LIR fi-bers described above; torsion also caused the twisting ofthe serotonin-LIR fibers in the middle region of the em-bryo resulting in the formation of a figure-eight pattern(Figs. 5C, 7B). Serotonin-LIR fibers originating at thebase of the acm projected into each velar lobe and thefoot where the fibers branched repeatedly (Fig. 5C).
IV. Late posttorsion veliger stage to hatching (E65–100)
During this time the velar lobes, foot, and shell contin-ued to increase in size (Fig. 6A). In addition, the opercu-
new positions on the left side of the body (Figs. 7A,10C). In addition, torsion resulted in more dorsal posi-tions of the fibers comprising the figure eight, FALC1,FALC2, and the FPC (Figs. 5B). At E55, FALC3 ap-peared on the right side along the posterior loop of the

lum and statocysts also appeared as the veliger ap-proached the time of hatching (Fig. 6A).
1. FMRFamide-like immunoreactivity
At approximately E65 a pair of FMRFamide-LIR pedalcells (FPeCs) appeared near the base of the acm(Figs. 6B, 10A). Each of these FPeCs extended two pro-cesses which met a fiber projecting from the base of theacm. Velar innervation continued to increase and, byE70–75, an FMRFamide-LIR fiber grew along the rim ofeach of the two velar lobes. By E100, these fibers metdorsally where the lobes connect. During this time atleast one additional FALC (FALC4) was faintly labeled(Figs. 6B, 7A).
2. Serotonin-like immunoreactivity
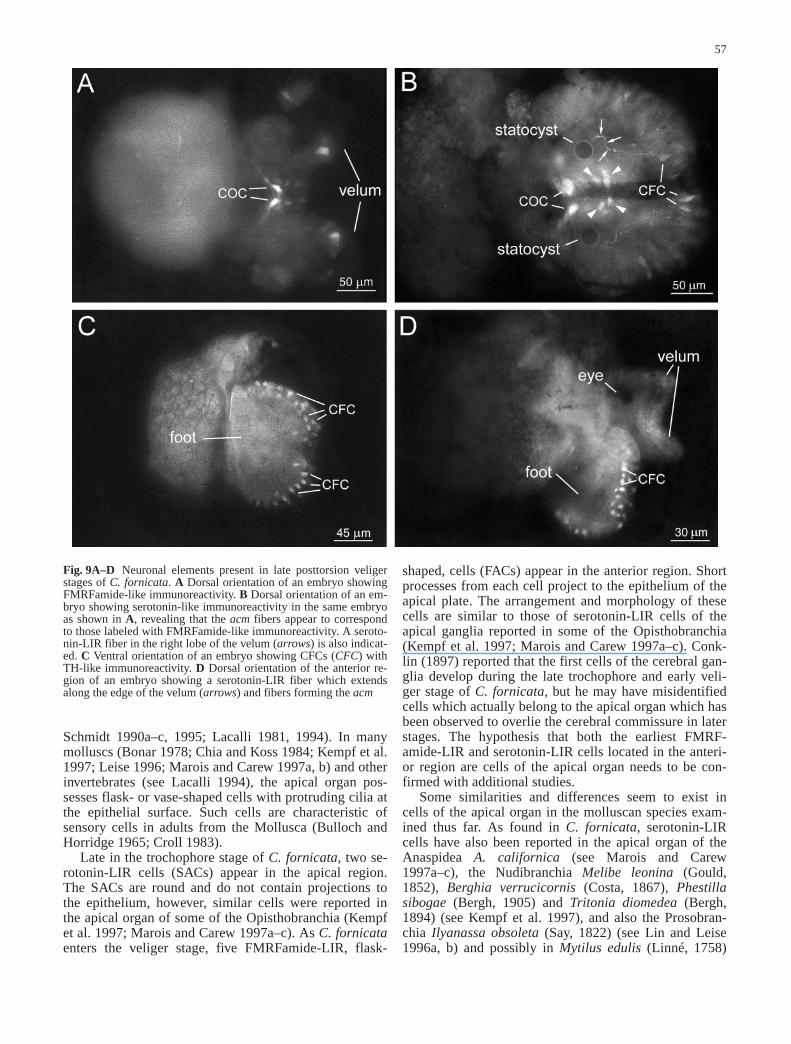
By E70–75 two serotonin-LIR abdominal loop cells(SALCs) appeared. SALC1 was located on the right,near FALC3, while SALC2 was located on the left, nearFALC1 and FALC2 (Figs. 6C, 7B, 10D). Unfortunately,double labeling for serotonin and FMRFamide was un-successful or appeared quite faint. It was, therefore, notpossible to show precisely where the FMRFamide-LIRcells and fiber tracts were in relation to those identifiedwith serotonin-like immunoreactivity. However, duringlate embryonic stages the acm appeared to be double la-beled by both anti-FMRFamide and anti-serotonin(Fig. 9A,B). Velar innervation continued to increase and,by E70–75, a serotonin-LIR fiber grew along the rim ofeach of the two velar lobes. By E100, these fibers alsomet dorsally where the lobes connect (Fig. 9D).
3. Catecholamines (TH-LIR, FaGlu)
It was not until E65–70 that cells became detectable us-ing FaGlu histochemistry or immunocytochemistry forthe detection of TH (Fig. 6D). At this stage both FaGlu-induced fluorescence and TH-like immunoreactivitywere located in numerous catecholaminergic foot cells(CFCs; Figs. 6D, 8B–D, 9C). The CFCs appeared to beflask-shaped with a slender portion that extends to theepithelial surface of the foot. Also at this stage, the Fa-Glu method revealed a pair of large, flask-shaped cate-cholaminergic oral cells (COCs) just dorsal to the mouth(Figs. 6D, 8A,B). By E70–75, additional COCs appearedand were interconnected with a network of fibers(Figs. 6D, 8B). The somata and fibers of the COCs werenot TH-LIR. Also by E70–75 the CFCs became morenumerous. Two slightly brighter medial CFCs at the dis-tal extreme of the foot sent projections toward the regionwhere the statocysts were located (Figs. 6D, 8B).
D. Discussion
None of the major recent reviews of neurodevelopmentin the Mollusca mentioned the presence of neuronal ele-ments in very early stages (Bullock 1985; Kandel 1979;Kandel et al. 1981; Raven 1966). The appearance of suchelements, therefore, might not have been expected in tro-chophore stages of C. fornicata. The present study notonly shows the early appearance of neuronal elementsbut also presents evidence for a relatively extensive ner-vous system formed during embryonic development.This early nervous system includes a posterior FMRF-amide-LIR cell, an apical organ with innervation to thevelum and foot, and numerous peripheral neurons.
I. Posterior FMRFamide-LIR cell
During early trochophore stages of C. fornicata, a single,large, FMRFamide-LIR cell (the FPC) appeared in the
55
Fig. 7A, B Neuronal elements present in a late posttorsion veligerstage of C. fornicata. The anterior region is to the right in eachfigure. A Dorsal orientation of an embryo revealing the FACs(rFAC 2,3, lFAC2, mFAC) and the four FALCs (FALC 1–4).B Dorsal orientation of an embryo showing the SACs (rSAC,lSAC) and the two SALCs (SALC1, 2) &/fig.c:

posterior region of the embryo and extended anterior ax-ons which terminated in an acm. A similarly positionedFMRFamide-LIR soma and its projections have also beenreported in early developmental stages of L. stagnalis(see Croll and Voronezhskaya 1996a). This central, early,FMRFamide-LIR, anteriorly projecting (c-EFAP) cell inL. stagnalisis positioned in the posterior region near thedeveloping shell gland outside the visceral loop similar tothe FPC of C. fornicata. The fibers from the c-EFAP alsoproject anteriorly and terminate in the region of the futurecerebral and pedal ganglia and near two cells beneath theapical plate. In addition, Croll and Voronezhskaya(1996a) reported that projections from the c-EFAP cellalso appear to form a scaffolding upon which the adultCNS develops. The similar morphology, position, timing,
and transmitter phenotype of the posterior FMRFamide-LIR cells in the two species suggests that they might behomologous. Although the most posterior FMRFamide-LIR cells in L. stagnalisand C. fornicataare quite simi-lar, some significant differences exist. For example, earlyveliger stages of L. stagnalisalso possesses additionalFMRFamide-LIR posterior cells which were not detectedin C. fornicata. Preliminary reports suggest the additionalpresence of FMRFamide-LIR cells in posterior regions ofthe trochophore stage of A. californica (see Croll andVoronezhskaya 1996a, b) but clearly more species need tobe examined to ascertain the generality of the posteriorFMRFamide-LIR cells and to examine differences insuch cells across the Mollusca.
II. Apical organ
An apical organ has been reported in some of the Gastro-poda (Bonar 1977, 1978; Chia and Koss 1984; Kempf etal. 1991, 1997; Lin and Leise 1996a, b; Marois andCarew 1997c; Moor 1983; Page 1993; Raineri 1995;Raineri and Ospovat 1994) and the Bivalvia (Raineri1995; Raineri and Ospovat 1994) as well as many otherswimming invertebrate larvae (see for example, Hay-
56
Fig. 8A–D Neuronal elements present in late posttorsion veligerstages of C. fornicata. The anterior region is to the right in eachfigure. A Ventral orientation of an embryo using FaGlu methodsshowing the two larger flask-shaped COCs (COC). B Ventral ori-entation of an embryo using FaGlu methods showing the largerCOCs and smaller pairs of flask-shaped cells (arrowheads) in theoral region. A fiber (arrows) extends from each statocyst to a pairof CFCs (CFC). C Ventral orientation of an embryo showingCFCs using FaGlu methods. D Right side view of an embryo us-ing FaGlu methods also showing the CFCs&/fig.c:
Schmidt 1990a–c, 1995; Lacalli 1981, 1994). In manymolluscs (Bonar 1978; Chia and Koss 1984; Kempf et al.1997; Leise 1996; Marois and Carew 1997a, b) and otherinvertebrates (see Lacalli 1994), the apical organ pos-sesses flask- or vase-shaped cells with protruding cilia atthe epithelial surface. Such cells are characteristic ofsensory cells in adults from the Mollusca (Bulloch andHorridge 1965; Croll 1983).
Late in the trochophore stage of C. fornicata, two se-rotonin-LIR cells (SACs) appear in the apical region.The SACs are round and do not contain projections tothe epithelium, however, similar cells were reported inthe apical organ of some of the Opisthobranchia (Kempfet al. 1997; Marois and Carew 1997a–c). As C. fornicataenters the veliger stage, five FMRFamide-LIR, flask-
shaped, cells (FACs) appear in the anterior region. Shortprocesses from each cell project to the epithelium of theapical plate. The arrangement and morphology of thesecells are similar to those of serotonin-LIR cells of theapical ganglia reported in some of the Opisthobranchia(Kempf et al. 1997; Marois and Carew 1997a–c). Conk-lin (1897) reported that the first cells of the cerebral gan-glia develop during the late trochophore and early veli-ger stage of C. fornicata, but he may have misidentifiedcells which actually belong to the apical organ which hasbeen observed to overlie the cerebral commissure in laterstages. The hypothesis that both the earliest FMRF-amide-LIR and serotonin-LIR cells located in the anteri-or region are cells of the apical organ needs to be con-firmed with additional studies.
Some similarities and differences seem to exist incells of the apical organ in the molluscan species exam-ined thus far. As found in C. fornicata, serotonin-LIRcells have also been reported in the apical organ of theAnaspidea A. californica (see Marois and Carew1997a–c), the Nudibranchia Melibe leonina (Gould,1852), Berghia verrucicornis(Costa, 1867), Phestillasibogae (Bergh, 1905) and Tritonia diomedea(Bergh,1894) (see Kempf et al. 1997), and also the Prosobran-chia Ilyanassa obsoleta(Say, 1822) (see Lin and Leise1996a, b) and possibly in Mytilus edulis(Linné, 1758)
57
Fig. 9A–D Neuronal elements present in late posttorsion veligerstages of C. fornicata. A Dorsal orientation of an embryo showingFMRFamide-like immunoreactivity. B Dorsal orientation of an em-bryo showing serotonin-like immunoreactivity in the same embryoas shown in A, revealing that the acm fibers appear to correspondto those labeled with FMRFamide-like immunoreactivity. A seroto-nin-LIR fiber in the right lobe of the velum (arrows) is also indicat-ed. C Ventral orientation of an embryo showing CFCs (CFC) withTH-like immunoreactivity. D Dorsal orientation of the anterior re-gion of an embryo showing a serotonin-LIR fiber which extendsalong the edge of the velum (arrows) and fibers forming the acm&/fig.c:

(Bivalvia; see Croll et al. 1997). Also, as observed in C.fornicata, FMRFamide-LIR cells, which may be withinthe apical organ, have been identified in later veligerstages of the opisthobranch P. sibogae(see Kempf et al.1991). However, studies of A. californica do not reportthe existence of FMRFamide-LIR cells under the apicalplate (Croll and Voronezhskaya 1996b). Two FMRF-amide-LIR flask-shaped cells were reported in L. stagn-alis (Pulmonata; see Croll and Voronezhskaya 1995,1996a). These cells were also observed to be TH-LIR,indicating the presence of catecholamines (Voronezhs-kaya et al. 1999). Based on the timing, position, andmorphology, these two cells are also possibly remnants
of an apical organ. The velar structures of L. stagnalisare reduced, presumably due to its intercapsular directdevelopment, and, therefore, it is possible that an apicalorgan has also been reduced to a few cells during evolu-tion.
III. Innervation of the velum
In C. fornicata, serotonin- and FMRFamide-LIR fibersextend from the apical or cerebral cells into the develop-ing velum. The origin of such innervation has been thesource of confusion in molluscan species. Velar innerva-tion had been thought to originate from the cerebral gan-glia (Arkett et al. 1987; Carter 1926, 1928; Mackie et al.1976), but more recently it has been shown that axonsmay actually emerge from cells of the apical organ insome of the Opisthobranchia (Kempf et al. 1997; Maroisand Carew 1997a–c) and I. obsoleta(Prosobranchia; seeLin and Leise 1996a, b). In addition, velar innervationhas also been reported from cells that may comprise anapical organ in M. edulis(Bivalvia; see Croll et al. 1997;Kulakovskiy and Flyachinskaya 1994; Raineri 1995;Raineri and Ospovat 1994). Although the origin of velarinnervation could not be determined in C. fornicatafrom
58
Fig. 10A–DNeuronal elements present in C. fornicatajust prior tohatching (E 100). A Dorsal orientation of an embryo showing in-creased FMRFamide-LIR innervation of the velum and to the foot(arrowheads). FPECs (FPeC), two FALCs (FALC1, 2), and non-specifically labeled cells (arrow) along the edge of the velum areindicated. B Dorsal orientation of an embryo showing serotonin-LIR innervation of the velum (arrowheads) and non-specificallylabeled cells (arrows). C Dorsal orientation of an embryo showingthe FPC (FPC), two FALCs (FALC1, 2), FACs (rFAC2, 3, lFAC2,mFAC), and fibers composing the acm. D Dorsal orientation of anembryo showing two SALCs (SALC1, 2), fibers forming the acm,and non-specifically labeled cells (arrows)

this present study, it is likely that such extensive innerva-tion arises from the apical cells since fibers were ob-served in velar regions prior to the first cells of the cere-bral or pedal ganglia. As the veliger develops, innerva-tion of velar regions becomes more elaborate. Since suchvelar innervation most likely controls locomotion andfeeding currents generated by the cilia, the amount andpattern of innervation probably correspond to changes inciliary movements at the time of hatching.
Different biochemical phenotypes have been shown toexist in the innervation of the velum. Serotonin-LIR inner-vation of the velar lobes exists in C. fornicataas in manymolluscan species (Kempf et al. 1997; Lin and Leise1996b; Marois and Carew 1997a–c). Evidence that seroto-nin stimulates cilia, has been reported in both adult forms(Audesirk and Willows 1979; Murakami 1987; Syed andWinlow 1989) and in larval stages (Beiras and Widdows1995; Koshtoyants et al. 1961). In bivalve molluscs, cate-cholaminergic innervation of the velum has also been re-ported in mid and late veliger stages (Croll et al. 1997).Catecholamines were not observed in velar regions of C.fornicataby the time of hatching, but may develop in laterveliger stages (Croll et al. 1997). In C. fornicata, FMRF-amide-like immunoreactivity is also observed in velar in-nervation, but this neuropeptide has not been reported invelar regions in any other molluscan species thus far.
IV. Innervation in the foot
It is probable that there is both efferent and afferent in-nervation of the foot in larval stages of C. fornicata. In-nervation in the foot consists of both FMRFamide- andserotonin-LIR processes. Such processes can be ob-served to project to the developing foot as the embryoenters the veliger stage. At this stage, identifiable cells ofthe cerebral and pedal ganglia were not observed and,therefore, innervation of the foot most likely originatesfrom cells of the apical organ. Similarly, axons were sug-gested to project from apical cells to the foot in someopisthobranchs (Kempf et al. 1991, 1997; Lin and Leise1996a; Marois and Carew 1997a–c) and the Prosobran-chia I. obsoleta(Lin and Leise 1996a, b). Such innerva-tion may control the pedal ciliary band reported to be in-volved with feeding (D’Asaro 1969) and locomotion(Diefenbach et al. 1991).
Flask-shaped FFCs with morphologies typical of pu-tative sensory cells were also observed in early stages.Such cells later disappeared but similar cells, CFCs wereobserved using techniques to localize catecholamines. Itis, therefore, possible that the cells of the foot switchedtransmitter phenotype during development. The presenceof both FMRFamide- and TH-like immunoreactivity inthe apical cells of L. stagnalissuggests that expression ofone transmitter does not preclude the expression of theother and the possibility of neurotransmitter switching,which has also been reported in the nervous system oflarval stages of the hornworm moth Manduca sexta(Linné, 1763) (see Tublitz and Sylwester 1990). On the
other hand, it is also possible that the FMRFamide-LIRcells are replaced by a new set of catecholaminergiccells. Similar cells positive for catecholamines were alsoreported in the foot of L. stagnalis (Gastropoda; seeVoronezhskaya et al. 1999) and in the Bivalvia M. edulisand Placopecten magellanicus(Gmelin, 1790) (see Crollet al. 1997). In larvae of Onchidoris bilamellata(Linné,1767) (Opisthobranchia), a ganglion exists containingsimilar flask-shaped cells in the propodial region of thefoot (Arkett et al. 1989). The precise function of theseputative sensory cells in the foot of C. fornicata is un-known, but they may be involved in metamorphosis, sim-ilar to the presumed function of the cells in O. bilamell-ata (see Arkett et al. 1989).
V. Neurons associated with the oral region
Catecholaminergic cells (COCs) are found near the mouthin prehatching stages of C. fornicata.These neurons alsohave a flask-shape typical of sensory cells in adult Mollus-ca (Bulloch and Horridge 1965; Croll 1983). Similar cate-cholaminergic cells were also described in the oral regionof a veliger stage of P. magellanicusand M. edulis(Bival-via; see Croll et al. 1997). Around metamorphosis, small,flask-shaped, catecholaminergic cells were also reportedaround the mouth of L. stagnalis (Gastropoda; seeVoronezhskaya et al. 1999). Sensory cells in the oral re-gion may control aspects of feeding, such as the detectionof differences in nutritional quality of food (Baldwin andNewell 1995). The appearance of the oral cells just priorto the time of hatching, when yolk supplies become de-pleted and the larva must start to feed on its own, is alsoconsistent with the possibility that such cells in oral re-gions might be involved in feeding in C. fornicata.
The catecholaminergic cells surrounding the mouth ofC. fornicatawere not TH-LIR. Similarly, catecholamin-ergic cells which were not TH-LIR have also been re-ported in L. stagnalis(see Croll et al. 1999; Voronezhs-kaya et al. 1999) and Homarus gammarus(Linné, 1758)(Crustacea; Cournil et al. 1994) and from the AmphibiaXenopus laevis(Duadin, 1803) and Rana ridibunda(Pallas, 1771) (see Gonzalez et al. 1994). The levels ofTH may be below detection by the antibody or a differentisoform of the TH enzyme may be present in such cellsand is undetected by the antibody. Alternatively, the cate-cholaminergic cells which were not TH-LIR may lack theability to synthesize catecholamines, but accumulatethem using an uptake mechanism (Gonzalez et al. 1994).
VI. The development of the adult CNS
During very early veliger stages (E35–40) of C. fornica-ta, serotonin- and additional FMRFamide-LIR fibers be-gin to develop along a pathway formed by the first ante-rior projections of the FPC. After torsion, such fiberstake on the figure-eight pattern typical of chiastoneury inthe adult CNS. Thus, by E55–60, a framework that be-
59

comes the ganglionic connectives of the adult CNS ap-pears to be laid down. In addition, the first serotonin-and/or FMRFamide-LIR cells of all the major ganglia ofthe adult CNS are observed by the time of hatching.
A pair of small serotonin-LIR cells (SCCs), whichmay be among the earliest residents of the cerebral gan-glia, appeared at E55. The cells appear near the develop-ing eyespots, slightly posterior and lateral to the acm.The location of these cells in the embryo is similar tothat of the first serotonergic cells reported in L. stagnalis(see Marois and Croll 1992). In addition to cells of thecerebral ganglia, the first cells of the other major gangliaof the adult CNS may possibly be present in embryonicstages of C. fornicata. For example, at approximatelyE65, two small FPeCs appear in regions which seem tocorrespond to the future site of the two pedal ganglia inC. fornicata. The FALCs and SALCs correspond to re-gions where the supra- and subintestinal ganglia and vis-ceral ganglia appear in prehatching stages. The presentstudy tentatively identifies the connectives and manycells of the adult CNS which appear in C. fornicatapriorto the time of hatching. The timing of such cells of theadult CNS is consistent with reports in other proso-branch species by D’Asaro (1969). Furthermore, gan-gliogenesis has been reported to take place just after thetime of hatching in I. obsoleta(see Lin and Leise 1996b)and, therefore, it is probable that the first cells developslightly earlier. The presence of cerebral and pedal cellsat hatching stages in some of the Opisthobranchia hasbeen previously reported (Kriegstein 1977; Page 1992a,b). Also by hatching stages in A. californica, FMRF-amide-LIR cells can be observed in regions correspond-ing to future sites of the ganglia which comprise theadult CNS (R.P. Croll, A.J.G. Dickinson, and E.E.L.Voronezhskaya unpublished observations).
VII. Conclusion
The present study has provided morphological evidenceof neuronal elements that may constitute a larval nervoussystem in trochophore and early veliger stages of C.fornicata. While relatively extensive, this nervoussystem was nonetheless largely undetected in earlierstudies of molluscan neurogenesis. Such elements as theposterior FMRFamide-LIR cell, the apical organ, andcells in the foot and the oral region are probably respon-sible for control of various behaviors and physiologicalfunctions in larval stages. In addition, this report alsosuggests the possibility that the larval nervous systemforms a scaffolding along which the adult nervoussystem develops. Such a hypothesis is consistent with thesuggested role of FMRFamide-LIR cells and processesin trochophore and early veliger stages of the pulmonate,L. stagnalis(see Croll and Voronezhskaya 1995, 1996a).The larval nervous system has also been shown to be im-portant in guiding the development of the adult CNS inother invertebrate groups such as in the Annelida (La-calli 1981, 1982) and in the Insecta (Bate 1976; Caudy
and Bentley 1986; Goodman and Shatz 1993). In addi-tion, neurotransmitters themselves may function to mod-ulate neurodevelopment through controlling morphoge-netic events in both insects and vertebrates (Buznikov etal. 1996; Lauder 1988) and shaping neuronal circuitry(Budnik et al. 1986; Haydon et al. 1987). They are alsoknown to control channel properties, concentrations ofsecond messengers and other transmitters in developingneuronal elements (reviewed by Baker and Croll 1996;Kater and Mills 1991; Meirer et al. 1991; Spitzer 1991).Neuronal elements in larval forms from the Nemertini,Phoronida, and Polychaeta also contain FMRFamide, se-rotonin, and catecholamines (Hay-Schmidt 1990a–c;1995). For example, the nervous system in the larvae ofPolygordius lacteus(Schieder 1868) (Polychaeta; seeHay-Schmidt 1995) shows neuronal elements containingsuch substances in the apical organ, in nerves associatedwith ciliary bands, and in the oral region. Thus similari-ties and differences in structure and development of ner-vous systems may also prove to be useful in phylogenet-ic studies as well as provide important insights into thebasic mechanisms of neurogenesis.
&p.2:Acknowledgements Research described in the present report wasconducted in partial fulfillment of requirements for an M.Sc. in bi-ology at Dalhousie University. We thank members of the thesis ex-amining committee (I.A. Meinertzhagen, B.K. Hall, E. Leise) andD.L. Jackson for useful suggestions which were incorporated intothis report and also M. Handa for technical assistance in prelimi-nary stages of this study. Funding was provided by a grant fromthe Natural Sciences and Engineering Research Council of Canadato R.P.C.
References
Arkett SA, Chia F, Goldberg JI, Koss R (1989) Identified settle-ment receptor cells in a nudibranch veliger respond to specificcue. Biol Bull 176:155–160
Arkett SA, Mackie GO, Singla CL (1987) Neuronal control of cili-ary locomotion in a gastropod veliger (Calliostoma). Biol Bull173:513–526
Audesirk GM, Willows E, Willows AOD (1979) The role of sero-tonin in the control of pedal ciliary activity by identified neu-rons in Tritonia diomedea. Comp Biochem Physiol 62:87–91
Baker MW, Croll RP (1996) Modulation of in vivo neuronalsprouting by serotonin in the adult CNS of the snail. Cell MolNeurobiol 16:561–576
Baldessarini RJ (1996) Drugs and the treatment of psychiatric dis-orders. In: Hardman JG, Limbird LE (eds) Goodman and Gil-man’s pharmacological basis of therapeutics. 9th edn. Mc-Graw-Hill, New York
Baldwin NS, Newell RIE (1995) Feeding rate responses of oysterlarvae (Crassostrea virginica) to seston quantity and composi-tion. J Exp Mar Biol Ecol 189:77–91
Barlow LA, Truman JW (1992) Patterns of serotonin and SCP im-munoreactivity during metamorphosis of the nervous systemof the red abalone, Haliotis rufescens. J Neurobiol 23:829–844
Bate CM (1976) Pioneer neurons in an insect embryo. Nature 260:54–55
Beiras R, Widdows J (1995) Effects of the neurotransmitters dopa-mine, serotonin and norepinephrine on the ciliary activity ofmussel (Mytilis edulis) larvae. Mar Biol 122:597–603
Bonar DB (1977) Morphogenesis at metamorphosis in opistho-branch molluscs. In: Chia F, Rice ME (eds) Settlement andmetamorphosis of marine invertebrate larvae. Elsevier, NewYork
60

Bonar DB (1978) Ultrastructure of a cephalic sensory organ in lar-va of the gastropod Phestilla sibogae(Aeolidacea, Nudibran-chia). Tissue Cell 10:153–165
Budnik V, Martin-Morris L, White K (1986) Perturbed pattern of cat-echolamine-containing neurons in mutant Drosophiladeficient inthe enzyme dopa decarboxylase. J Neurosci 6:6682–6691
Bullock TH, Horridge GA (1965) Structure and function in thenervous systems of invertebrates. Freeman, San Francisco
Bullock AGM (1985) Development and plasticity of the molluscannervous system. In: Neurobiology and behavior, part 1. Wil-lows (ed) AOD 8 The Mollusca. Academic Press, Toronto
Buznikov GA, Shumkler YB, Lauder JM (1996). From oocyte toneuron: do neurotransmitters function in the sane waythroughout development? Cell Mol Neurol 16:537–559
Carter GS (1926) On the nervous control of the velar cilia of thenudibranch veliger. J Exp Biol 4:1–26
Carter GS (1928) On the structure of the cells bearing the velarcilia of the nudibranch veliger. J Exp Biol 6:97–109
Caudy M, Bentley D (1986) Pioneer growth cone steering along aseries of neuronal and non-neuronal cues of different affinities.J Neurosci 6:1781–1795
Chia F, Koss R (1984) Fine structure of the cephalic sensory organin the larva of the nudibranch Rostanga pulchra(Mollusca,Opisthobranchia, Nudibranchia). Zoomorphology 104:131–139
Conklin EG (1897) The embryology of Crepidula. J Morphol 13:1–230
Cooper JR, Bloom FE, Roth RH (1996) The biochemical basis ofneuropharmacology, 7th edn. Oxford University Press, New York
Cournil I, Helluy SM, Beltz BS (1994) Dopamine in the lobsterHomarus gammarus. I. Comparative analysis of dopamine andtyrosine hyroxylase immunoreactivities in the nervous systemof the juvenile. J Comp Neurol 344:455–469
Croll RP (1983) Gastropod chemoreception. Biol Rev 58:293–319Croll RP, Chiasson BJ (1989) Post-embryonic development of se-
rotonin-like immunoreactivity in the central nervous system ofthe snail, Lymnaea stagnalis. J Comp Neurol 280:122–142
Croll RP, Voronezhskaya EE (1995) Early FMRFamide-like im-munoreactive cells in gastropod neurogenesis. Acta Biol Hung46:295–303
Croll RP, Voronezhskaya EE (1996a) Early elements in gastropodneurogenesis. Dev Biol 173:344–347
Croll RP, Voronezhskaya EE (1996b) Early neurodevelopment inAplysia, Lymnaeaand Helisoma. Soc Neurosci Abstr 22:1948
Croll RP, Jackson DL, Voronezhskaya EE (1997) Catecholamine-containing cells in larval and post-larval bivalve molluscs. BiolBull 193:116–124
Croll RP, Voronezhskaya EE, Hiripi L, Elekes K (1999) Develop-ment of catecholaminergic neurons in the pond snail, Lymnaeastagnalis II: Postembryonic development of central and pe-ripheral cells. J Comp Neurol 404:297–309
D’Asaro CN (1969) The comparative embryogenesis and early or-ganogenesis of Bursa corrugataPerry and Distorsio clathrata(Gastropoda: Prosobranchia). Malacologia 9:349–389
Diefenbach TJ, Koehncke NK, Goldberg JI (1991) Characteriza-tion and development of rotational behavior in Helisomaem-bryos: role of endogenous serotonin. J Neurobiol 22:922–934
Furness JB, Costa M, Wilson AJ (1977) Water-stable fluoropho-res, produced by reaction with aldehyde solutions, for the his-tochemical localization of catechol- and indolethylamines.Histochemistry 52:159–170
Gonzalez A, Marin O, Tuinhof R, Smeets WJ (1994) Ontogeny ofcatecholamine systems in the central nervous system of anuranamphibians; an immunohistochemical study with antibodiesagainst tyrosine hydroxylase and dopamine. J Comp Neurol346:63–79
Goodman CS, Shatz CJ (1993) Developmental mechanisms thatgenerate precise patterns of neuronal connectivity. Cell 72:77–98
Graham A (1983) Evolution within the Gastropoda: Prosobran-chia. In: Wilber KM (ed) Evolution. 10 The Mollusca. Aca-demic Press, Toronto
Greenberg MJ, Price DA (1992) Relationships among the FMRF-amide-like peptides. Prog Brain Res 92: 25–37
Haydon PG, McCobb DP, Kater SB (1987) The regulation of neu-rite outgrowth, growth cone motility, and electrical synapto-genesis by serotonin. J Neurobiol 18:197–215
Hay-Schmidt A (1990a) Distribution of catecholamine-containing,serotonin-like and neuropeptide FMRFamide-like immunore-active cells and processes in the nervous system of the pili-dium larva (Nemertini). Zoomorphology 109:231–244
Hay-Schmidt A (1990b) Distribution of catecholamine-containing,serotonin-like and neuropeptide FMRFamide-like immunore-active neurons and processes in the nervous system of the ac-tinotroch larva of Phoronis muelleri(Phoronida). Cell TissueRes 259:105–118
Hay-Schmidt A (1990c) Distribution of catecholamine-containing,serotonin-like and neuropeptide FMRFamide-like immunore-active neurons and processes in the nervous system of the ear-ly actinotroch larva of Phoronis vancouverensis(Phoronida):distribution and development. Can J Zool 68:1525–1536
Hay-Schmidt A (1995) The larval nervous system of PolygordiuslacteusSchieder, 1868 (Polygordiidae, Polychaeta): immuno-cytochemical data. Acta Zool (Stockh) 76:121–140
Kandel ER (1979) Behavioral biology of Aplysia. Freeman, SanFrancisco
Kandel ER, Kriegstein A, Schacher S (1981) Development of thecentral nervous system of Aplysia in terms of the differentia-tion of its specific identifiable cells. Neurosci 5:2033–2063
Kater SB, Mills LR (1991) Regulation of growth cone behavior bycalcium. J Neurosci 11:891–899
Kempf SC, Chun GV, Hadfield MG (1991) An immunocytochemi-cal search for potential neurotransmitters in larvae of Phestillasibogae (Gastropoda, Opisthobranchia). Comp BiochemPhysiol 101C:299–305
Kempf SC, Page LR, Pires A (1997) Development of serotonin-like immunoreactivity in the embryos and larvae of nudibranchmollusks with emphasis on the structure and possible functionof the apical sensory organ. J Comp Neurol 386:507–528
Koshtoyants KS, Buznikov GA, Manukhin BN (1961) The possi-ble role of 5-hydroxy-tryptamine in the motor activity of em-bryos of some marine gastropods. Comp Biochem Physiol 3:20–26
Kriegstein AR (1977) Development of the nervous system ofAplysia californica. Proc Nat Acad Sci USA 74:375–378
Kulakovskiy EY, Flyachinskaya LP (1994) Formation of elementsof the regulatory systems in larval development of Mytilusedulis. Hydrobiol J 30:85–94
Lacalli TC (1981) Structure and development of the apical organin trochophores of Spirobranchus polycerus, Phyllodoce ma-culataand Phyllodoce mucosa(Polychaeta). Proc R Soc LondB 212:381–402
Lacalli TC (1982) The nervous system and ciliary band ofMuller’s larva. Proc R Soc Lond B 217:37–58
Lacalli TC (1994) Apical organs, epithelial domains and origin ofthe chordate central nervous system. Am Zool 34:533–541
Lauder JM (1988) Neurotransmitters as morphogens. Prog BrainRes 73:365–387
Leise EM (1996) Selective retention of the fluorescent dye DA-SPEI in a larval gastropod mollusc after paraformaldehyde fix-ation. Microsc Res Tech 33:496–500
Lin M, Leise EM (1996a) NADPH-Diaphorase activity changesduring gangliogenesis and metamorphosis in the gastropodmollusc Ilyanassa obsoleta. J Comp Neurol 374:194–203
Lin M-F, Leise EM (1996b) Gangliogenesis in the prosobranchgastropod Ilyanassa obsoleta. J Comp Neurol 374:180–193
Mackie GO, Singla CL, Thiriot-Quievreux C (1976) Nervous con-trol of ciliary activity in gastropod larvae. Biol Bull 151:182–199
Marois R, Carew TJ (1997a) Fine structure of the apical ganglionand its serotonergic cells in the larva of Aplysia californica.Biol Bull 192:388–398
Marois R, Carew TJ (1997b) Ontogeny of serotonergic neurons inAplysia californica. J Comp Neurol 386:477–490
61

Marois R, Carew TJ (1997c) Projection patterns and target tissuesof serotonergic cells in larval Aplysia californica. J CompNeurol 386:491–506
Marois R, Croll RP (1992) Development of serotonergic cellswithin the embryonic central nervous system of the pond snail,Lymnaea stagnalis. J Comp Neurol 322:255–265
Meirer E, Hertz L, Schousboe A (1991) Neurotransmitters as de-velopmental signals. Neurochem Int 19:1–15
Moor B (1983). Organogenesis. In: Wilbur KM (ed) Development.The Mollusca. Academic Press, Toronto
Murakami A (1987) Control of ciliary beat frequency in the gill ofMytilus. Comp Biochem Physiol 86C:273–279
Page LR (1992a) New interpretation of a nudibranch central ner-vous system based on ultrastructural analysis of neurodevelop-ment in Melibe leonina. I. Cerebral and visceral loop ganglia.Biol Bull 182:348–365
Page LR (1992b) New interpretation of a nudibranch central ner-vous system based on ultrastructural analysis of neurodevelop-ment in Melibe leonina. II. Pedal, pleural and labial ganglia.Biol Bull 182:366–381
Page LR (1993) Developmental analysis reveals labial and sura-dular ganglia and the primary framework of the nervoussystem in nudibranch gastropods. J Neurobiol 24:1443–1459
Pechenik JA (1984) The relationship between temperature,growth rate, and duration of planktonic life for larvae of the
gastropod Crepidula fornicata(L.). J Exp Mar Biol Ecol 74:241–257
Raineri M (1995) Is a mollusc an evolved bent metatrochophore?A histochemical investigation of neurogenesis in Mytilus(Mollusca: Bivalvia). J Mar Biol Assoc UK 75:571–592
Raineri M, Ospovat M (1994) The initial development of gangliarrudiments in a posterior position in Mytilus galloprovincialis(Mollusca: Bivalvia). J Mar Biol Assoc UK 74:73–77
Raven CP (1966) Morphogenesis: the analysis of molluscan devel-opment. Pergamon Press, Oxford
Spitzer NC (1991) A developmental handshake: neuronal controlof ionic currents and their control of neuronal differentiation. JNeurobiol 22:659–673
Syed NI, Winlow W (1989) Morphology and electrophysiology ofneurons innervating the ciliated locomotor epithelium in Lym-naea stagnalis(L.). Comp Biochem Physiol 93A:633–644
Tublitz NJ, Sylwester AW (1990) Postembryonic alteration oftransmitter phenotype in individually identified peptidergicneurons. J Neurosci 10:161–168
Voronezhskaya EE, Hiripi L, Elekes K, Croll RP (1999) Develop-ment of catecholaminergic neurons in the pond snail, Lymnaeastagnalis. I. Embryonic development of dopaminergic neuronsand dopamine-dependent behaviors. J Comp Neurol 404:285–296
&/fig.c:
62




















