
Toward Decoding the Principles of Cancer Metastasis Circuits
Upload
independentCategory
view
5download
0
Histochemical Evidence of Osteoclastic Degradation ofExtracellular Matrix in Osteolytic Metastasis OriginatingFrom Human Lung Small Carcinoma (SBC-5) CellsMINQI LI,1,2 NORIO AMIZUKA,1,2* KIICHI TAKEUCHI,1 PAULO H.L. FREITAS,1 YOSHIRO KAWANO,1
MASAAKI HOSHINO,1 KIMIMITSU ODA,3 KAYOKO NOZAWA-INOUE,1 AND TAKEYASU MAEDA1,2
1Division of Oral Anatomy, Niigata University Graduate School of Medical and Dental Sciences, Niigata, 951-8514, Japan2Center for Transdisciplinary Research, Niigata University, Niigata, 951-8514, Japan3Division of Biochemistry, Niigata University Graduate School of Medical and Dental Sciences, Niigata, 951-8514, Japan
KEY WORDS bone metastasis; osteoclast; matrix metalloproteinase-9; osteopontin; CD44
ABSTRACT The aim of this study was to assess the dynamics of osteoclast migration and thedegradation of unmineralized extracellular matrix in an osteolytic metastasis by examining a well-standardized lung cancer metastasis model of nude mice. SBC-5 human lung small carcinoma cellswere injected into the left cardiac ventricle of 6-week-old BALB/c nu/nu mice under anesthesia. At25–30 days after injection, the animals were sacrificed and their femora and/or tibiae wereremoved for histochemical analyses. Metastatic lesions were shown to occupy a considerable areaextending from the metaphyses to the bone marrow region. Tartrate resistant acid phosphatase(TRAPase)-positive osteoclasts were found in association with an alkaline phosphatase (ALPase)-positive osteoblastic layer lining the bone surface, but could also be localized in the ALPase-nega-tive stromal tissues that border the tumor nodules. These stromal tissues were markedly positivefor osteopontin, and contained a significant number of TRAPase-positive osteoclasts expressingimmunoreactivity for CD44. We thus speculated that, mediating its affinity for CD44, osteopontinmay serve to facilitate osteoclastic migration after their formation associated with ALPase-positiveosteoblasts. We next examined the localization of cathepsin K and matrix metallo-proteinase-9(MMP-9) in osteoclasts. Osteoclasts adjacent to the bone surfaces were positive for both proteins,whereas those in the stromal tissues in the tumor nests showed only MMP-9 immunoreactivity.Immunoelectron microscopy disclosed the presence of MMP-9 in the Golgi apparatus and in vesicu-lar structures at the baso-lateral cytoplasmic region of the osteoclasts found in the stromal tissue.MMP-9-positive vesicular structures also contained fragmented extracellular materials. Thus,osteoclasts appear to either select an optimized function, namely secreting proteolytic enzymesfrom ruffled borders during bone resorption, or recognize the surrounding extracellular matrix bymediating osteopontin/CD44 interaction, and internalize the extracellular matrices. Microsc. Res.Tech. 69:73–83, 2006. VVC 2006 Wiley-Liss, Inc.
INTRODUCTION
Bone is a frequent site of tumor metastasis originat-ing from breast, kidney, lung, and so forth. Colemanand Rubens (1987) have reported that nearly 70% ofpatients with breast cancer featured bone metastases.Histologically, three types of bone metastasis can beidentified: the intertrabecular, the osteolytic, and theosteoblastic metastasis (Moriwaki, 1987). Interestingly,the origin of the tumor seems to determine which willbe the resulting type of bone metastasis. As an illustra-tion, the osteolytic type is typically represented by bonemetastasis from breast carcinomas, having osteoclasticbone resorption markedly enhanced. On the otherhand, prostate carcinomas have been reported to causeosteoblastic lesions (Jacobs et al., 1979). Investigatorshave tried to explain why bone would be a site for carci-noma metastases, and both the ‘‘seed and soil theory’’(Paget, 1889) and the ‘‘hemodynamic theory’’ (Ewing,1928) are two viable justifications. Nonetheless, nomatter its origin or type, bone metastasis often makes
tumor patients suffering from severe complicationssuch as bone pain, pathological fracture, hypercalce-mia, and nerve compression syndromes (Firusianet al., 1976; Hall et al., 1983; Okada et al., 1984; Uro-vitz et al., 1977), adding more weight over an alreadycompromised life quality.
Using animal models and cell lines derived fromhuman carcinomas, bone metastasis has been thor-oughly studied to elucidate its intrinsic pathologicalevents. SBC-5 human small lung cancer cells havebeen used in animal models because of its selective col-onizination of long bones of nude mice and induction of
*Correspondence to: Prof. Norio Amizuka, DDS, Ph.D.; Center for Transdisci-plinary Research, Niigata University, Niigata 951-8514, Japan.E-mail: [email protected]
Received 1 September 2005; accepted in revised form 29 November 2005
Contract grant sponsor: Niigata University Research Project; Contract grantsponsor: Japan Society.
DOI 10.1002/jemt.20272
Published online inWiley InterScience (www.interscience.wiley.com).
VVC 2006 WILEY-LISS, INC.
MICROSCOPY RESEARCH AND TECHNIQUE 69:73–83 (2006)

osteolytic metastasis (Kakiuchi et al., 2003; Miki et al.,2000, 2004; Zhang et al., 2003). SBC-5 cells secreteparathyroid hormone-related peptide (PTHrP), a factorpreviously identified as causative for humoral hyper-calcemia of malignancy (Burtis et al., 1987; Horiuchiet al., 1987; Kemp et al., 1987; Strewler et al., 1987;Suva et al., 1987; Thiede and Rodan, 1988), and there-fore, stimulate osteoclastic activity in bone.
Bone resorption is believed to be responsible forosteolytic metastasis, as bone resorption and subse-quent tumor expansion were suppressed by uses ofantibodies for PTHrP or bisphosphonates that specifi-cally inhibit osteoclast bone resorption (Guise et al.,1996; Yoneda et al., 2000). Osteoclastic activity is regu-lated by alkaline phosphatase (ALPase)-positive osteo-blasts, which simultaneously express receptor activa-tor of NF-jB ligand (RANKL) in a physiological state.However, under nonphysiological or pathological cir-cumstances, e.g., bone grafting, osteoclastic migrationand subsequent bone resorption sometimes take placewithout associated ALPase-positive osteoblasts, andpreferentially occur prior to osteoblastic bone forma-tion (Hao et al., 2004). Also, devitalized bone oftenreveals osteoclastic resorption, followed by migrationof ALPase-positive cells (Kamijou et al., 1994; Piattelliet al., 1996; Piattelli et al., 1997). Therefore, onceformed in association with ALPase-positive osteoblasts,osteoclasts are likely to migrate and operate withoutbeing controlled by those cells to some extent. Sinceosteoclasts possess CD44 (Nakamura et al., 1995), theinterplay of CD44 with extracellular matrix proteinssuch as osteopontin may regulate osteoclastic motility(Chellaiah and Hruska, 2003).
Tumor cells can change their surrounding microen-vironment into one that is appropriate for their growth,with degradation and subsequent remodeling ofunmineralized extracellular matrix apparently beingimportant for tumor cells to invade neighboring softtissues. Given that osteoclasts derive from the mono-cyte/macrophage lineage, they might accomplish endo-cytosis or phagocytosis (Everts et al., 1985; Heersche,1978). Albeit, there is no doubt that osteoclasts are re-sponsible for osteolytic metastasis. We speculated thatosteoclasts cells might have additional tasks in boneresorption such as engulfing of unmineralized extracel-lular matrix in the metastastic lesion.
To elucidate this possibility, we examined osteoclas-tic migration and the digestion of extracellular matrixas well as osteoclastic bone resorption in a well-stand-ardized lung cancer metastasis model of nude mice.
MATERIALS AND METHODSAnimals and Tissue Preparation
Six-weeks-old male BALB/c-nu/nu mice (CLEA Ja-pan Tokyo, Japan) were used in the experiments, andwere kept in a pathogen-free environment. The guide-lines for Animal Experimentation of Niigata Universi-ty were strictly observed in all animal experimentsconducted. According to Arguello et al. (1988), SBC-5human small lung cancer cells (105 cells per animal)were injected into the left cardiac ventricle under anes-thesia, with an intraperitoneal injection of 8% chloralhydrate (400 mg/100 g body weight). Animals were
housed having water and powder diet ad libitum. Uponconfirmation of an obvious bone metastasis in femoraand/or tibiae through soft X-ray analysis, at about 25–30 days after the injection, mice were subjected totranscardiac perfusion with a fixative of 4% parafor-maldehyde in a 0.1 M phosphate buffer (pH 7.4). Fem-ora and/or tibiae were removed, immersed in the samefixative for an additional 12 h, and those designated tolight microscopy observation were decalcified with 10%EDTA-2Na solution for 3 weeks at 48C. After decalcifi-cation, the specimens were dehydrated with ethanolsolutions in ascending concentrations prior to paraffin-embedding. The specimens for transmission electronmicroscopy (TEM) were postfixed with 1% OsO4 in0.1 M cacodylate buffer (pH 7.4) for 4 h at 48C. Theywere then dehydrated with ascending concentrationsof acetone, and embedded in epoxy resin (Epok: Poly-sciences, inc., Warrington, PA). The resulting ultrathinsections were observed under a TEM (H-7000; HitachiCo. Ltd, Tokyo, Japan) at 80 kV.
Immunohistochemistry for ALPase,Proliferating Cell Nuclear Antigen (PCNA),Osteopontin, Osteocalcin, CD44, Cathepsin K
and Type I Collagen
Dewaxed paraffin sections were pretreated with 0.3%hydrogen peroxide for 20 min, and then with 1% bovineserum albumin (BSA; Seologicals Proteins Inc. Kanka-kee, IL, USA) in PBS (1% BSA-PBS) for 30 min. Thesesections were incubated with primary antibodies of rab-bit serum to tissue nonspecific ALPase (Oda et al.,1999), mouse antiproliferating cell nuclear antigen(PCNA), a 36 kD auxiliary protein of DNA polymerase d(CN Biosciences, Inc. San Diego, CA), goat antiosteocal-cin (Biomedical Technologies, Inc., Stoughten, MA), ormouse anticathepsin K (Fuji Chemical Co., Takaoka, Ja-pan) at a dilution of 1:100 with 1% BSA-PBS for 2 h atroom temperature. Either antisera to rabbit osteopontinor rat anti-CD44 (BD Biosciences, San Jose, CA) wereapplied to the paraffin sections diluted at 1:2000 for 2 hat room temperature. For type I collagen immunohisto-chemistry, dewaxed-paraffin sections were subjected toenzyme digestion with 0.5% hyaluronidase (Sigma, St.Louis, MO, USA) at 378C for 1 h before being treatedwith hydrogen peroxide and 1% BSA-PBS. They werethen incubated with rabbit antiserum against type I col-lagen (Cosmo BIO Co. Ltd., Tokyo, Japan) diluted at1:3000 for 2 h at room temperature. Paraffin sectionstreated with primary antibodies were immersed inhorseradish peroxidase (HRP)-conjugated secondaryantibodies (Chemicon International Inc., Temecula, CA,USA) at a dilution of 1:100 for 1 h at room temperature.Visualization was achieved with diaminobenzidine(DAB) substrate, and the sections were then observedunder light microscopy.
Multiple-Staining for MMP-9, F4/80and TRAPase
Paraffin sections pretreated with hydrogen peroxi-dase and 1% BSA-PBS were incubated with goat anti-mouse matrix metalloproteinase (MMP)-9 (TECHNECorporation, Minneapolis, MN) at a dilution of 1:100for 2 h at room temperature. After immersion in HRP-
74 M. LI ET AL.

conjugated rabbit antigoat IgG (Chemicon), the immu-noreaction was visualized through DAB-staining. ForF4/80 immunohistochemistry, the same sections wereincubated with supernatant of rat hybridomas for 2 hat room temperature followed by ALPase-conjugatedantirat antibody (Kirkegaard & Perry laboratories Inc.Gaithersburg, MD, USA). For ALPase enzyme-reac-tion, sections were dipped in a mixture of 5 mg of naph-tol AS-BI phosphate (Sigma) as substrate and 18 mg offast blue RR salt (Sigma), diluted in 30 mL of 0.1 MTris-HCl buffer (pH 8.5) at 378C for 30 min. For TRA-Pase-detection, slides were rinsed with PBS and sub-merged in a mixture of 2.5 mg of naphthol AS-BI phos-phate, 18 mg of red violet LB salt, and 100 mM L(þ)tartaric acid (0.76 g) diluted in 30 mL of 0.1 M sodiumacetate buffer (pH 5.0) during 15 min at 378C. We alsoperformed double staining of F4/80 and TRAPase,ALPase and TRAPase, or osteopontin and TRAPase insome sections, thus aiming to confirm the spatial rela-tionship between these molecules. All the sections werefaintly counterstained with methyl green.
Immunoelectron Microscopic Observation
After decalcification with 5% EDTA, bone specimenswere sectioned 100 lm in thickness by a Microslicer(Dosaka EM, Kyoto, Japan). The sections were pre-treated with 0.3% hydrogen peroxide for 30 min, with1% BSA-PBS for 5 h, and incubated with goat anti-mouse MMP-9 (TECHNE) for 48 h at 48C. After wash-ing in PBS for 12 h, they were submerged in HRP-con-jugated rabbit antigoat IgG (Chemicon) at a dilution of1:100 during 24 h at 48C. DAB staining was accom-plished, and the sections were postfixed with 1% OsO4
in 0.1 M cacodylate buffer (pH 7.4) for 3 h at 48C, dehy-drated with ascending concentrations of acetone, andembedded in epoxy resin (Polysciences) prior to TEMobservations.
RESULTSMetastatic Tumor Nest and Bone Resorption
Soft X-ray analysis revealed translucencies in theaffected femora, a representative image of the onset ofa metastatic lesion (data not shown). Histologicalobservations demonstrated a huge mass of metasta-sized tumor cells occupying previous metaphyseal andbone marrow spaces, with obvious trabecular resorp-tion (Fig. 1A). Metastatic lesions consisted of many tu-mor nodules and intervening layers of stromal tissue,and PCNA-immunoreactivity was identified mainly inthe metastasized tumor cells (Figs. 1D and 1E). A thicklayer of the stromal tissue between the tumor nodulesand bone surfaces was recognized (Figs. 1B and 1C),and the portion closer to the bone surface revealedALPase-reactivity (Figs. 2A and 2B). In contrast,ALPase was absent in those cell layers facing the tu-mor nests, as well as in the stromal tissues in betweentumor nodules (Fig. 2C). At the bone surfaces, manyTRAPase-positive osteoclasts were found accompaniedby ALPase-reactive osteoblastic cells. Remarkably,TRAPase-positive osteoclasts and F4/80-immunoposi-tive monocyte/macrophages were also observed in thestromal tissues amid the tumor nodules (Fig. 2D),though none of these cell types could be found insidethe tumor nodules.
Immunolocalization of Osteopontin and CD44 inthe Metastatic Lesions
An immunoreactivity for type I collagen was foundin the stromal layer in metastatic lesion though not soabundant as seen in the bone matrix (Fig. 3A). Whenexamined under TEM, unmineralized extracellularmatrices such as collagen fibrils and amorphous mate-rials were present in the vicinity of osteoclasts(Fig. 3B). The osteoclasts, characterized by irregularlyshaped nuclei, many mitochondria, and absence ofruffled borders, often formed coated pits on their cellmembranes, facing these extracellular matrices. Unliketype I collagen, osteopontin immunoreactivity wasbroadly and intensely found in the stromal tissuesencompassing tumor nodules in the metastasizedlesion (Fig. 3C). Both the stromal tissues between tu-mor nests and bone surfaces (Fig. 3D) and thoseencompassing tumor nodules (Fig. 3E) showed a strongimmunoreaction for osteopontin. Given the affinity ofCD44 to osteopontin, we next examined CD44 immuno-histochemistry in metastatic lesion. Serial sections dis-closed the corresponding distribution of F4/80 (mono-cyte/macrophage lineage) and CD44-immunoreactiv-ities, even though CD44-immunoreactive cells had arelative higher population when compared with that ofF4/80-positive cells (Compare Figs. 4A and 4B). Mutu-ally, F4/80 reactive monocyte/macrophages and CD44-positive cells were present in the stromal tissues of themetastatic lesion (Compare Figs. 4C and 4D). As well,TRAPase-positive osteoclasts and some CD44-reactivecells colocalized, and TRAPase-positive osteoclastspopulated osteopontin-immunoreactive stromal tissues(Figs. 4E and 4F).
The Internalization of UnmineralizedExtracellular Matrices by Osteoclasts
in the Stromal Tissue
The osteoclasts attached to bone surfaces, but notthose in the tumor stromal tissues, were intenselyimmunopositive for cathepsin K (Fig. 5A, 5B). Immu-nolocalization of MMP-9 spatially matched that ofTRAPase-positive osteoclasts (Figs. 5C and 5F). At ahigher magnification, MMP-9 could be found sharing acommon region with TRAPase-positive osteoclasts lin-ing bone surfaces (Figs. 5D and 5G), as well as in TRA-Pase-positive cells in the stromal tissues (Figs. 5E and5H). Thus, osteoclasts resorbing bone were positiveboth for cathepsin K and MMP-9, but those in the stro-mal tissues expressed MMP-9 only. Under TEM, osteo-clasts in the stromal tissues have demonstrated inter-nalization of extracellular matrix, as evidenced by theamorphous organic materials in the pits of cell mem-branes and the cytoplasmic vesicles (Figs. 5I and 5J),which fused with the MMP-9-reactive vesicles (Fig. 5K).
Macrophages are well known to show MMP-9 inpathological states (Hibbs, 1992; Welgus et al., 1990;Xie et al., 1994), but in our experiments, F4/80-positivemonocyte/macrophages were not positive for this enzymewhen assessed through a triple staining for MMP-9,F4/80, and TRAPase (Fig. 6A). On the other hand, TRA-Pase-positive osteoclasts attaching bone surfaces in-tensely expressed MMP-9. Under TEM observations,osteoclasts on bone surfaces displayed well-formed ruffledborders, numerous mitochondria, and irregularly-shaped
75HISTOLOGICAL EXAMINATIONS ON BONE METASTASIS
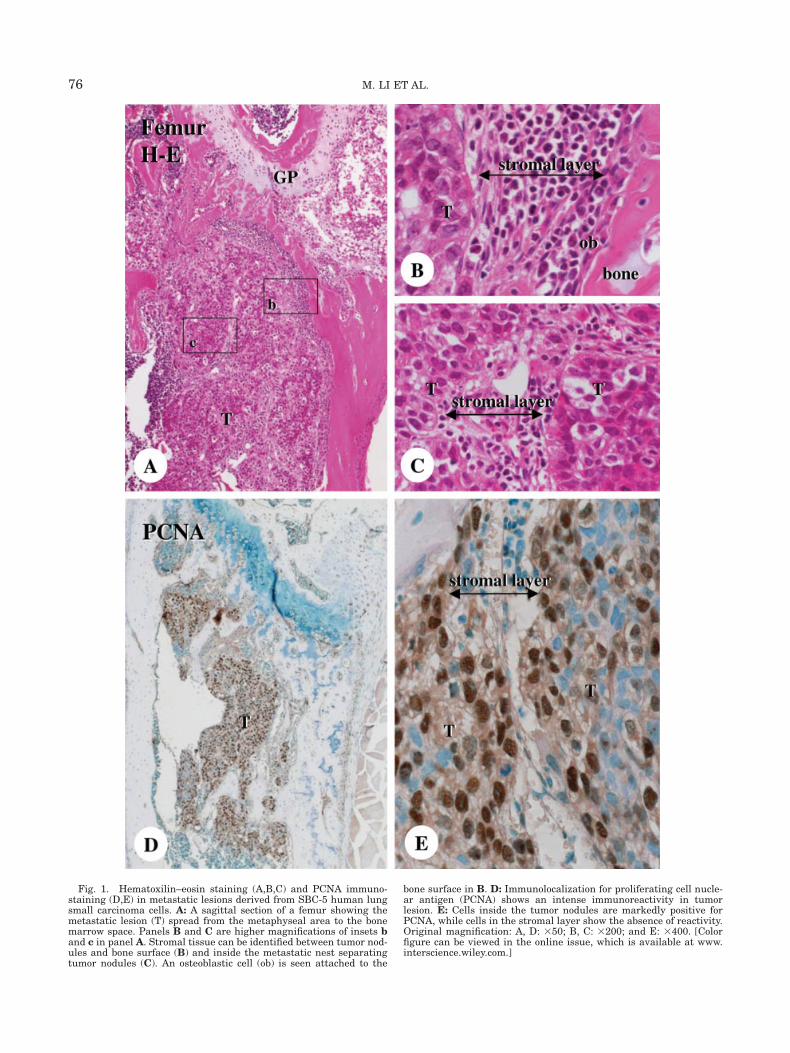
Fig. 1. Hematoxilin–eosin staining (A,B,C) and PCNA immuno-staining (D,E) in metastatic lesions derived from SBC-5 human lungsmall carcinoma cells. A: A sagittal section of a femur showing themetastatic lesion (T) spread from the metaphyseal area to the bonemarrow space. Panels B and C are higher magnifications of insets band c in panel A. Stromal tissue can be identified between tumor nod-ules and bone surface (B) and inside the metastatic nest separatingtumor nodules (C). An osteoblastic cell (ob) is seen attached to the
bone surface in B. D: Immunolocalization for proliferating cell nucle-ar antigen (PCNA) shows an intense immunoreactivity in tumorlesion. E: Cells inside the tumor nodules are markedly positive forPCNA, while cells in the stromal layer show the absence of reactivity.Original magnification: A, D: 350; B, C: 3200; and E: 3400. [Colorfigure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
76 M. LI ET AL.
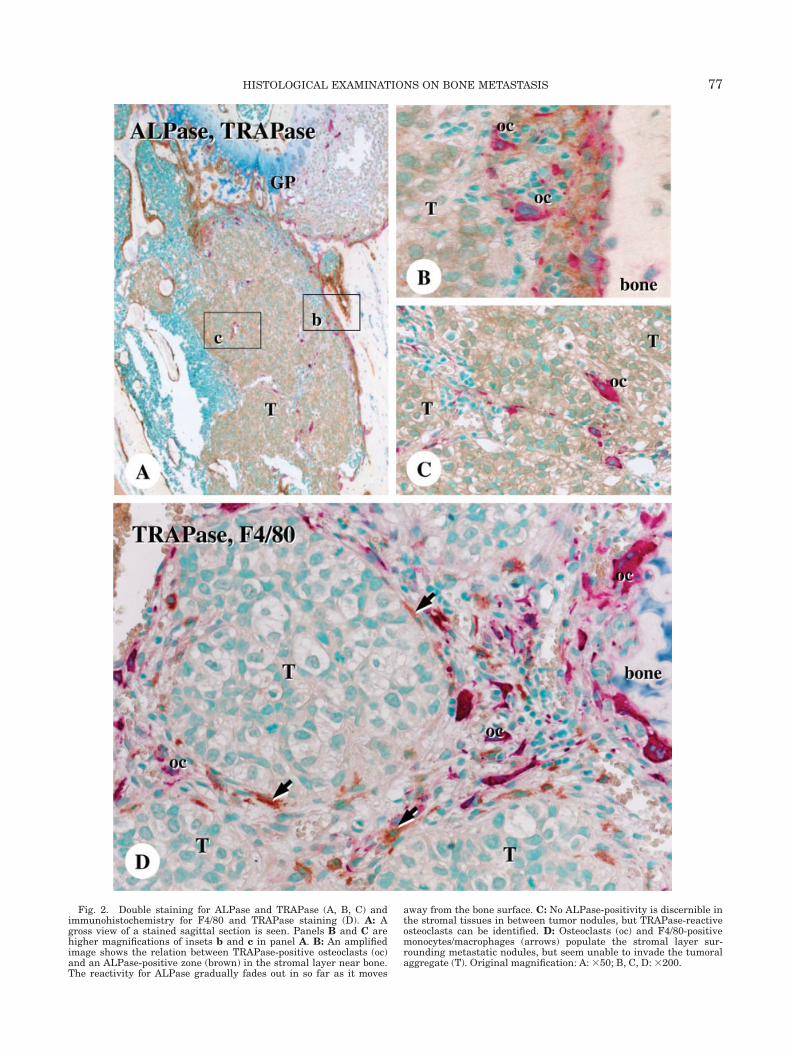
Fig. 2. Double staining for ALPase and TRAPase (A, B, C) andimmunohistochemistry for F4/80 and TRAPase staining (D). A: Agross view of a stained sagittal section is seen. Panels B and C arehigher magnifications of insets b and c in panel A. B: An amplifiedimage shows the relation between TRAPase-positive osteoclasts (oc)and an ALPase-positive zone (brown) in the stromal layer near bone.The reactivity for ALPase gradually fades out in so far as it moves
away from the bone surface. C: No ALPase-positivity is discernible inthe stromal tissues in between tumor nodules, but TRAPase-reactiveosteoclasts can be identified. D: Osteoclasts (oc) and F4/80-positivemonocytes/macrophages (arrows) populate the stromal layer sur-rounding metastatic nodules, but seem unable to invade the tumoralaggregate (T). Original magnification: A: 350; B, C, D: 3200.
77HISTOLOGICAL EXAMINATIONS ON BONE METASTASIS
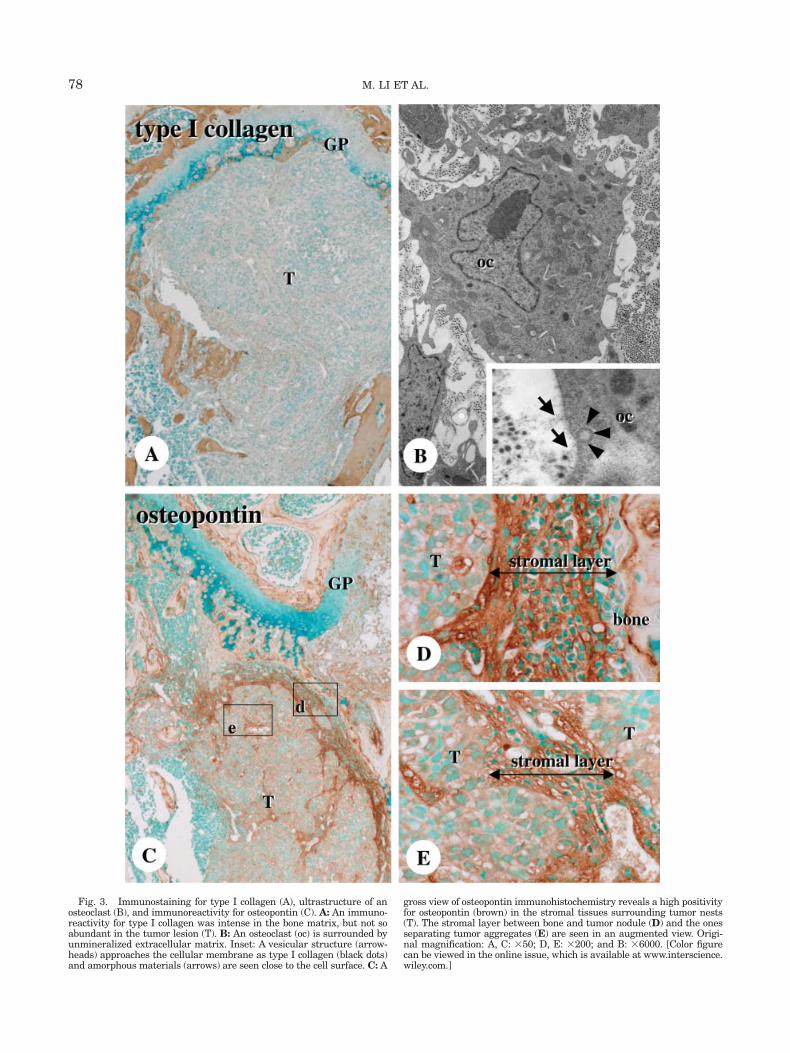
Fig. 3. Immunostaining for type I collagen (A), ultrastructure of anosteoclast (B), and immunoreactivity for osteopontin (C). A: An immuno-reactivity for type I collagen was intense in the bone matrix, but not soabundant in the tumor lesion (T). B: An osteoclast (oc) is surrounded byunmineralized extracellular matrix. Inset: A vesicular structure (arrow-heads) approaches the cellular membrane as type I collagen (black dots)and amorphous materials (arrows) are seen close to the cell surface. C: A
gross view of osteopontin immunohistochemistry reveals a high positivityfor osteopontin (brown) in the stromal tissues surrounding tumor nests(T). The stromal layer between bone and tumor nodule (D) and the onesseparating tumor aggregates (E) are seen in an augmented view. Origi-nal magnification: A, C: 350; D, E: 3200; and B: 36000. [Color figurecan be viewed in the online issue, which is available at www.interscience.wiley.com.]
78 M. LI ET AL.

Fig. 4. Immunostaining for F4/80 (A, C), CD44 (B, D, F), and dou-ble staining for osteopontin and TRAPase (E) in serial sections.A, B: In low-powered images, F4/80 reactive monocyte/macrophages(brown in A) and CD44-positive cells (brown in B) colocalize in serialsections, though the population of CD44-immunoreactive cells seemsto be relatively higher. C, D: F4/80-positive monocyte/macrophages
(arrows) are found identical to CD44-positive cells (an arrowhead) inthe stromal tissues (Compare panels C and D). E, F: TRAPase-posi-tive osteoclasts (oc) in the osteopontin-reactive stromal tissue (brownin E) are found to coincide with CD44-positive cells (brown in F) inserial sections. Original magnification: A, B: 350; C, D: 3200; and E,F: 3400.
79HISTOLOGICAL EXAMINATIONS ON BONE METASTASIS
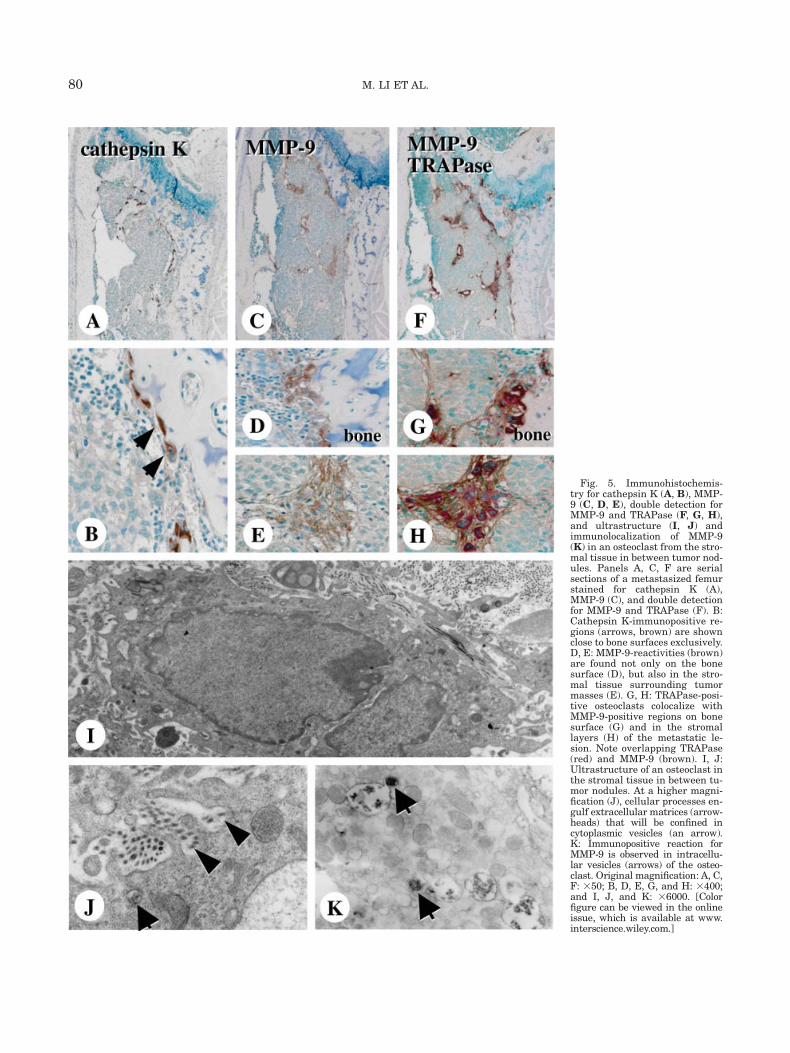
Fig. 5. Immunohistochemis-try for cathepsin K (A, B), MMP-9 (C, D, E), double detection forMMP-9 and TRAPase (F, G, H),and ultrastructure (I, J) andimmunolocalization of MMP-9(K) in an osteoclast from the stro-mal tissue in between tumor nod-ules. Panels A, C, F are serialsections of a metastasized femurstained for cathepsin K (A),MMP-9 (C), and double detectionfor MMP-9 and TRAPase (F). B:Cathepsin K-immunopositive re-gions (arrows, brown) are shownclose to bone surfaces exclusively.D, E: MMP-9-reactivities (brown)are found not only on the bonesurface (D), but also in the stro-mal tissue surrounding tumormasses (E). G, H: TRAPase-posi-tive osteoclasts colocalize withMMP-9-positive regions on bonesurface (G) and in the stromallayers (H) of the metastatic le-sion. Note overlapping TRAPase(red) and MMP-9 (brown). I, J:Ultrastructure of an osteoclast inthe stromal tissue in between tu-mor nodules. At a higher magni-fication (J), cellular processes en-gulf extracellular matrices (arrow-heads) that will be confined incytoplasmic vesicles (an arrow).K: Immunopositive reaction forMMP-9 is observed in intracellu-lar vesicles (arrows) of the osteo-clast. Original magnification: A, C,F: 350; B, D, E, G, and H: 3400;and I, J, and K: 36000. [Colorfigure can be viewed in the onlineissue, which is available at www.interscience.wiley.com.]
80 M. LI ET AL.

Fig. 6. Triple staining for F4/80, MMP-9, and TRAPase (A), andultrastructure (B, C) and immunolocalization of MMP-9 (D) in osteo-clasts adjacent to bone surface. A: TRAPase-positive osteoclasts (oc)are shown to possess the MMP-9-reactivity (brown), but F4/80-posi-tive macrophages/monocytes (m/) do not. B, C: An osteoclast adjacent
to bone surface shows its exuberant ruffled border (rb). D: Positivereaction for MMP-9 is seen in the Golgi complex, small vesicles (anarrow), and the ruffled border (an arrowhead). Original magnifica-tion: A: 3400; B, C, and D: 36000.
81HISTOLOGICAL EXAMINATIONS ON BONE METASTASIS

nuclei (Fig. 6B). MMP-9 was localized in the Golgi appa-ratus, cytoplasmic vesicles close to the ruffled borders,and narrow extracellular regions flanked by ruffled bor-ders (Fig. 6C).
DISCUSSION
After injection with SBC-5 human lung small carci-noma cells, mice femora featured large metastaticlesions with osteoclasts playing several roles, namely,bone resorption, migration in association with osteo-pontin/CD44 interaction, and participation in matrixdegradation. Our approach unveiled that osteoclasticactivities in metastatic lesions are not restricted tobone resorption, although it is apparent that osteoclas-tic bone resorption is the major contributory event forosteolytic metastasis, in agreement with other reportswith SBC-5 cell line (Kakiuchi et al., 2003; Miki et al.,2000, 2004; Zhang et al., 2003). Osteoblasts seem to beessential for osteoclast formation and activation, andin addition, tumor-derived factors may enhance theosteoblastic functions in osteoclastogenesis. Miki et al.(2000) have reported a positive correlation betweenPTHrP production by human lung cancer cells and thedevelopment of bone metastasis in mice. The molecularmechanisms of osteoclastic bone resorption involveinterleukin (IL)-6, IL-1, tumor necrosis factor-a, andmacrophage inflammatory protein-1a, as well asPTHrP (David Roodman, 2001; Kwan et al., 2004; Oka-matsu et al., 2004). Besides, bone matrix would repre-sent a bank of growth factors such as transforminggrowth factors and insulin-like growth factor, whichwould be released in case of bone resorption. Severalinvestigators have proposed a model in which tumorcells would secrete factors that stimulate osteoblasticstromal cells, this stimulation enhancing osteoclasticresorption; bone degradation would release embeddedgrowth factors, which in turn activate tumor cells(Choki et al., 1998; Ciftci et al., 2003). It seems likelythat these complex cell-to-cell and cell signaling mech-anisms would fit as an explanation for the events inbone metastatic lesions.
It is likely that osteopontin, once released from bonematrix by accelerated osteoclastic bone resorption,would accumulate in the stromal tissues of the meta-static lesion. Osteoclasts and macrophages were shownto possess CD44, and both cell types could be found inthe stromal tissues. Chellaiah and Hruska (2003) havesuggested that osteopontin acts as an autocrine factorfor osteoclastic motility, mediated by avb3 and CD44.Under the pathological circumstance of bone metasta-sis, osteopontin may enhance osteoclastic migration to-ward the stromal tissues of the metastatic lesion bymediating interaction with these molecules. It is note-worthy that, despite the absence of ALPase-reactiveosteoblasts, osteoclasts could successfully come into tu-mor stromal tissues, which appears just seemingly con-tradictable to previous reports (Owens and Chambers,1993; Suda et al., 1999; Takahashi et al., 1988). How-ever, in vivo study has shown that even without associ-ating ALPase-positive osteoblasts, osteoclasts couldaccumulate artificial bone defects containing bone sub-stitutes (Hao et al., 2004). In addition, devitalized boneoften undergoes an initial osteoclastic resorption, fol-lowed by migration of ALPase-positive cells (Kamijou
et al., 1994; Piattelli et al., 1996, 1997). Therefore, wepostulate that osteoclasts, after being generated inassociation with ALPase-positive osteoblastic cells, couldsurvive and migrate to some extent without having osteo-blasts in a supporting role. Tumor nodules failed to dis-play osteopontin, and osteoclasts did not invade tumornodules as they did to the stromal tissues. Supposingthat tumor cells are to secrete chemotactic factors induc-ing osteoclastic migration, the osteoclasts should be rec-ognized inside the tumor nodules, but it did not happenin our study. Therefore, attachment to an extracellularmatrix, including osteopontin appears to be a reasonableexplanation for the site-specific migration of osteoclastsin our experiments.
It is of paramount importance to elucidate the biolog-ical functions of osteoclasts in the stromal tissues ofthe metastatic lesion. Osteoclast generally secretes ca-thepsin K (Drake et al., 1996; Inaoka et al., 1995; Saha-ra and Sasaki, 2001; Yamaza et al., 1998), and expressMMP-9, also referred to as gelatinase B (Tezuka et al.,1994, Okada et al., 1995), a molecule that acts in coop-eration with collagenase to degrade bone matrix(Sakiyama et al., 1994). As shown in Figure 5K, MMP-9-immunoreactive vesicles were found in the cytoplasmicregion of the osteoclasts, which often fused to vacuolescontaining organic materials, indicative of internalizedextracellular substances. Therefore, degradation of inter-nalized materials in osteoclasts follows the general path-way of primary lysosomes with MMP-9 and endosomes asseen in other cell types, without secreting this enzymeextracellularly. In contrast to MMP-9, we failed to detectcathepsin K in the osteoclasts from stromal tissues (Figs.5A and 5B), indicating that cathepsin K is a proteolyticenzyme acting on collagen digestion specifically at theresorption lacunae coincident with cathepsin K. MMP-9was seen in resorption lacunae adjacent to bone-resorbingosteoclasts. In addition, the presence of cytoplasmicvacuoles and their close proximity to ruffled borders indi-cates that osteoclasts secrete MMP-9 through theirruffled borders. Thus, osteoclasts appear to recognize thesurrounding extracellular matrix, and select an optimizeddegradation pathway, namely MMP-9 secretion fromruffled borders, or digestion of internalized materials.
Thus, osteoclasts play important roles in degrada-tion of unmineralized extracellular matrices in the lungcancer cell (SBC-5) bone metastasis. Osteopontin mayserve to facilitate osteoclastic migration in the metastaticlesion, by mediating its affinity to CD44 on osteoclasticcell membranes, which may lead to immediate osteolysis.Osteoclasts appear to utilize an optimized pathway ofmatrix degradation, MMP-9 secretion from ruffled bor-ders or digestion of internalized materials.
ACKNOWLEDGMENTS
The authors express their gratitude to Ono ChemicalCo., Tokyo for having kindly provided the SBC-5human small lung cancer cells used in this study.
REFERENCES
Arguello F, Baggs RB, Frantz CN. 1988. A murine model of experi-mental metastasis to bone and bone marrow. Cancer Res 48:6876–6881.
Burtis WJ, Wu T, Bunch C, Wysolmerski JJ, Isogna KL, Weir EC,Broadus AE, Stewart AF. 1987. Identification of a novel 17,000-dal-ton parathyroid hormone-like adenylate cyclase-stimulating pro-
82 M. LI ET AL.

tein from a tumor associated with humoral hypercalcemia of malig-nancy. J Biol Chem 161:7151–7156.
Chellaiah MA, Hruska KA. 2003. The integrin a(v)b(3) and CD44 reg-ulate the actions of osteopontin on osteoclast motility. Calcif TissueInt 72:197–205.
Choki I, Sourla A, Reyes-Moreno C, Koutsilieris M. 1998. Osteoblast-derived growth factors enhance adriamycin-cytostasis of MCF-7human breast cancer cells. Anticancer Res 18:4213–4224.
Ciftci K, Su J, Trovitch PB. 2003. Growth factors and chemotherapeu-tic modulation of breast cancer cells. J Pharm Pharmacol 55:1135–1141.
Coleman RE, Rubens RD. 1987. The clinical course of bone metasta-ses from breast cancer. Br J Cancer 55:61–66.
David Roodman G. 2001. Biology of osteoclast activation in cancer. JClin Oncol 19:3562–3571.
Drake FH, Dodds RA, James IE, Connor JR, Debouck C, RichardsonS, Lee-Rykaczewski E, Coleman L, Rieman D, Barthlow R, Hast-ings G, Gowen M. 1996. Cathepsin K, but not cathepsins B, L, or S,is abundantly expressed in human osteoclasts. J Biol Chem271:12511–12516.
Everts V, Aronson DC, Beertsen W. 1985. Phagocytosis of bone colla-gen by osteoclasts in two cases of pycnodysostosis. Calcif Tissue Int37:25–31.
Ewing JA. 1928. A treatise on tumor. 3rd ed. Philadelphia: W.B. Saun-ders.
Firusian N, Mellin P, Schmidt CG. 1976. Results of 89strontium ther-apy in patients with carcinoma of the prostate and incurable painfrom bone metastases: a preliminary report. J Urol 116:764–768.
Guise TA, Yin JJ, Taylor SD, Kumagai Y, Dallas M, Boyce BF, YonedaT, Mundy GR. 1996. Evidence for a causal role of parathyroid hor-mone-related protein in the pathogenesis of human breast cancer-mediated osteolysis. J Clin Invest 98:1544–1549.
Hall SM, Buzdar AU, Blumenschein GR. 1983. Cranial nerve palsiesin metastatic breast cancer due to osseous metastasis without in-tracranial involvement. Cancer 52:180–184.
Hao H, Amizuka N, Oda K, Fujii N, Ohnishi H, Okada A, Nomura S,Maeda T. 2004. A histological evaluation on self-setting a-tricalciumphosphate applied in the rat bone cavity. Biomaterials 25:431–442.
Heersche JN. 1978. Mechanism of osteoclastic bone resorption: a newhypothesis. Calcif Tissue Res 26:81–84.
Hibbs MS. 1992. Expression of 92 kDa phagocyte gelatinaseby inflammatory and connective tissue cells. Matrix Suppl 1:51–57.
Horiuchi N, Caulfield MP, Fisher JE, Goldman ME, Mckee GR, Rea-gan JE, Levy JJ, Nutt RF, Rodan SB, Schofield TL, Clemens TL,Rosenblatt M. 1987. Similarity of synthetic peptide from human tu-mor to parathyroid hormone in vivo and in vitro. Science 238:1566–1568.
Inaoka T, Bilbe G, Ishibashi O, Tezuka K, Kumegawa M, Kokubo T.1995. Molecular cloning of human cDNA for cathepsin K: novel cys-teine proteinase predominantly expressed in bone. Biochem Bio-phys Res Commun 206:89–96.
Jacobs SC, Pikna D, Lawson RK. 1979. Prostatic osteoblastic factor.Invest Urol 17:195–198.
Kakiuchi S, Daigo Y, Tsunoda T, Yano S, Sone S, Nakamura Y. 2003.Genome-wide analysis of organ-preferential metastasis of humansmall cell lung cancer in mice. Mol Cancer Res 1:485–499.
Kamijou T, Nakajima T, Ozawa H. 1994. Effects of osteocytes onosteoinduction in the autogenous rib graft in the rat mandible.Bone 15:629–637.
Kemp BE, Moseley JM, Rodda CP, Ebeling PR, Wettenhall REH, Sta-pleton D, Diefenbach-Jagger N, Ure F, Michelangeli VP, SimmonsHA, Raisz LG, Martin TJ. 1987. Parathyroid hormone-related pro-tein of malignancy: active synthetic fragments. Science 238:1568–1570.
Kwan Tat S, Padrines M, Theoleyre S, Heymann D, Fortun Y. 2004.IL-6, RANKL, TNF-a/IL-1: interrelations in bone resorption patho-physiology. Cytokine Growth Factor Rev 15:49–60.
Miki T, Yano S, Hanibuchi M, Sone S. 2000. Bone metastasis modelwith multiorgan dissemination of human small-cell lung cancer(SBC-5) cells in natural killer cell-depleted SCID mice. Oncol Res12:209–217.
Miki T, Yano S, Hanibuchi M, Kanematsu T, Muguruma H, Sone S.2004. Parathyroid hormone-related protein (PTHrP) is responsiblefor production of bone metastasis, but not visceral metastasis, byhuman small cell lung cancer SBC-5 cells in natural killer cell-depleted SCID mice. Int J Cancer 108:511–515.
Moriwaki S. 1987. Histopathology of metastatic bone tumors. Jpn JCancer Chemother 14:1680–1687.
Nakamura H, Kenmotsu S, Sakai H, Ozawa H. 1995. Localization ofCD44, the hyaluronate receptor, on the plasma membrane of osteo-cytes and osteoclasts in rat tibiae. Cell Tissue Res 280:225–233.
Oda K, Amaya Y, Fukushi-Irie M, Kinameri Y, Ohsuye K, Kubota I,Fujimura S, Kobayashi J. 1999. A general method for rapid purifi-cation of soluble versions of glycosylphosphatidylinositol-anchoredproteins expressed in insect cells: an application for human tissue-nonspecific alkaline phosphatase. J Biochem (Tokyo) 126:694–699.
Okada Y, Ogawa M, Horikoshi N, Inagaki J, Inoue K, Ikeda K, UsuiN, Nakada H, Adachi K, Mukaiyama T. 1984. Hypercalcemia inbreast cancer. Gan To Kagaku Ryoho 11:1468–1474.
Okada Y, Naka K, Kawamura K, Matsumoto T, Nakanishi I, FujimotoN, Sato H, Seiki M. 1995. Localization of matrix metalloproteinase 9(92-kilodalton gelatinase/type IV collagenase ¼ gelatinase B) in osteo-clasts: implications for bone resorption. Lab Invest 72:311–322.
Okamatsu Y, Kim D, Battaglino R, Sasaki H, Spate U, Stashenko P.2004. MIP-1 g promotes receptor-activator-of-NF-j-B-ligand-inducedosteoclast formation and survival. J Immunol 173:2084–2090.
Owens J, Chambers TJ. 1993. Macrophage colony-stimulating factor(M-CSF) induces migration in osteoclasts in vitro. Biochem BiophysRes Commun 195:1401–1407.
Paget S. 1889. The distribution of secondary growths in cancer of thebreast. Lancet 1:571–573.
Piattelli A, Scarano A, Corigliano M, Piattelli M. 1996. Presence ofmultinucleated giant cells around machined, sandblasted andplasma-sprayed titanium implants: a histological and histochemicaltime-course study in rabbit. Biomaterials 17:2053–2058.
Piattelli A, Scarano A, Alberti LD, Piattelli M. 1997. Histological andhistochemical analyses of acid and alkaline phosphatases aroundhydroxyapatitecoated implants: a time course study in rabbit. Bio-materials 18:1191–1194.
Sahara T, Sasaki T. 2001. Effects of brefeldin-A: potent inhibitor of in-tracellular protein transport on ultrastructure and resorptive func-tion of cultured osteoclasts. Anat Rec 263:127–138.
Sakiyama H, Inaba N, Toyoguchi T, Okada Y, Matsumoto M, MoriyaH, Ohtsu H. 1994. Immunolocalization of complement C1s and ma-trix metalloproteinase 9 (92kDa gelatinase/type IV collagenase) inthe primary ossification center of the human femur. Cell Tissue Res277:239–245.
Strewler GJ, Stern PH, Jacob JW, Eveloff J, Klein RF, Leung SC,Rosenblatt M, Nissenson RA. 1987. Parathyroid hormonelikeprotein from human renal carcinoma cells: structural and func-tional homology with parathyroid hormone. J Clin Invest 80:1803–1807.
Suda T, Takahashi N, Udagawa N, Jimi E, Gillespie MT, Martin TJ.1999. Modulation of osteoclast differentiation and function by thenew members of the tumor necrosis factor receptor and ligand fami-lies. Endocr Rev 20:345–357.
Suva LJ, Winslow GA, Wettenhall RE, Hammonds RG, Moseley JM,Diefenbach-Jagger H, Rodda CP, Kemp BE, Rodriguez H, Chen EY,Hudson PJ, Martin TJ, Wiod WI. 1987. A parathyroid hormone-related protein implicated in malignant hypercalcemia: cloning andexpression. Science 237:893–896.
Takahashi N, Akatsu T, Udagawa N, Sasaki T, Yamaguchi A, MoseleyJM, Martin TJ, Suda T. 1988. Osteoblastic cells are involved inosteoclast formation. Endocrinology 123:2600–2602.
Tezuka K, Nemoto K, Tezuka Y, Sato T, Ikeda Y, Kobori M, Kawa-shima H, Eguchi H, Hakeda Y, Kumegawa M. 1994. Identificationof matrix metalloproteinase 9 in rabbit osteoclasts. J Biol Chem269:15006–15009.
Thiede MA, Rodan GA. 1988. Expression of a calcium-mobilizingparathyroid hormone-related peptide in lactating mammary tissue.Science 242:278–280.
Urovitz EP, Fornasier VL, Czitrom AA. 1977. Sternal metastases andassociated pathological fractures. Thorax 32:444–448.
Welgus HG, Campbell EJ, Cury JD, Eisen AZ, Senior RM, Wilhelm SM,Goldberg GI. 1990. Neutral metalloproteinases produced by humanmononuclear phagocytes. Enzyme profile, regulation, and expressionduring cellular development. J Clin Invest 86:1496–1502.
Xie B, Dong Z, Fidler IJ. 1994. Regulatory mechanisms for the expres-sion of type IV collagenases/gelatinases in murine macrophages.J Immunol 152:3637–3644.
Yamaza T, Goto T, Kamiya T, Kobayashi Y, Sakai H, Tanaka T. 1998.Study of immunoelectron microscopic localization of cathepsin K inosteoclasts and other bone cells in the mouse femur. Bone 23:499–509.
Yoneda T, Michigami T, Yi B, Williams PJ, Niewolna M, Hiraga T.2000. Actions of bisphosphonate on bone metastasis in animal mod-els of breast carcinoma. Cancer 88:2979–2988.
Zhang H, Yano S, Miki T, Goto H, Kanematsu T, Muguruma H,Uehara H, Sone S. 2003. A novel bisphosphonate minodronate(YM529) specifically inhibits osteolytic bone metastasis producedby human small-cell lung cancer cells in NK-cell depleted SCIDmice. Clin Exp Metastasis 20:153–159.
83HISTOLOGICAL EXAMINATIONS ON BONE METASTASIS
Copyright © 2022 FDOKUMEN




















