
Vasopressin and social odor processing in the olfactory bulb and anterior olfactory nucleus

Cell Tissue Res (1994) 275:587-598 Cell&Tissue
Research �9 Springer-Verlag 1994
Histochemical and immunocytochemical study of the neurons from the rat olfactory placode V~ronique Pellier, Liliane Astic Laboratoire de Physiologie Neurosensorielle, UCB/Lyon I, F-69622 Villeurbanne Cedex, France
Received: 8 February 1993 / Accepted: 15 July 1993
migration of
Abstract. Immunocytochemical and histochemical meth- ods have been used to describe the neuronal population migrating from the rat olfactory placode and to analyze the spatio-temporal evolution of this neuronal migration during development. Several neuronal markers, such as binding to the lectin Ulex europaeus (UEA I) and the presence of neuron-specific enolase (NSE), olfactory marker protein (OMP), and luteinizing hormone-releas- ing hormone (LHRH), have been tested in order to deter- mine whether migrating neurons originate from both the medial and the lateral parts of the placode and whether they all express LHRH. Our data show that a large pop- ulation of differentiated migrating neurons can be identi- fied with an antibody against NSE from the 14th day of gestation and with UEA I one day later. Migrating neu- rons are closely associated with both the vomeronasal axon fascicles emerging from the medial pit and the olfac- tory axons originating from the lateral pit. However, the neuron migration from the lateral pit appears to be more discrete than that from the medial pit. No LHRH im- munoreactivity has been detected among neurons mi- grating from the lateral pit. Some neurons accompanying the olfactory axon fascicles exhibit a high level of matura- tion as shown by their OMP-positivity. Numerous neu- rons positive for both NSE and UEA I have also been observed within the presumptive olfactory nerve layer in early embryonic stages.
Key words: Olfactory placode - Cell migration - Neuron- specific enolase - Lectin - LHRH - Olfactory marker protein Development - Rat (Wistar, SPF)
Introduction
The earliest sign of olfactory organ development in verte- brate embryos is the appearance of olfactory placodes resembling epithelial thickenings in the anterolateral re-
Correspondence to: V. Pellier
gion of the head. In rat embryos of 12 days, the placodes invaginate to constitute simple olfactory pits that are lined by the pseudostratified olfactory epithelium; on embryonic day 13.5 (E13.5), a secondary recess is formed in the medial wall of either pit giving rise to the anlage of the vomeronasal organ (Lejour 1967; Farbman 1991). The olfactory nerves arise from the dorsal and lateral parts of the pit whereas vomeronasal and terminal nerves arise from the medially located vomeronasal organ.
An important cell migration process originating from the olfactory placode has been described in a wide variety of vertebrates during embryogenesis. At least two cell types have been proposed to migrate from the developing olfactory epithelium. Some of the migrating cells accom- panying the olfactory axon fascicles present epitheloid characteristics and are considered to be the precursors of the ensheathing cells often referred to in the literature as the olfaCtory Schwann cells (Van Campenhout 1956; Cuschieri and Bannister 1975 a,b; Bossy 1980; Mendoza et al. 1982; Farbman and Squinto 1985; Chuah and Au 1991 ; Valverde et al. 1992). It has been suggested that the precursors of these ensheathing cells are migrating basal cells (Marin-Padilla and Amieva 1989). Other migrating cells are probably neuroblasts, which are generally re- garded as precursors of the ganglion cells of the terminal nerve (Lejour 1967; Filogamo and Robecchi 1969; Men- doza et al. 1982; Farbman and Squinto 1985). Moreover, by combining luteinizing hormone-releasing hormone (LHRH) immunohistochemistry and thymidine autora- diography, it has been demonstrated that cerebral LHRH neurons originate in the medial olfactory placode and then migrate into the brain following the course of the terminal nerve (Schwanzel-Fukuda and Pfaff 1989; Wray et al. 1989 a,b). This migration of LHRH neurons from the medial pit has been described in various verte- brate species and is well-known, probably because neu- rons can be precociously identified by immunocytochem- istry. This is not the case for the ensheathing glial cells, which are immature in the early developmental stages of the olfactory placode (Farbman and Squinto 1985; Chuah and Au 1991). The data available on the process

588
of migra t ing neu rons from the nasal pit do no t make clear whether these neu rons or iginate exclusively from the medial par t of the pit or whether they also migrate from the lateral part. Likewise, it is unclear whether all migra t ing neu rons from the olfactory placode express the decapept ide L H R H .
We have used immunocy tochemica l and his tochemi- cal approaches to describe the neu rona l popu la t i on mi- gra t ing from the medial and lateral parts of the rat olfac- tory placode and to analyze the spa t io- tempora l evolu- t ion of this neu rona l migra t ion dur ing development . The n e u r o n a l markers tested were as follows: b ind ing to the lectin Ulex europaeus ( U E A I), which recognizes a fucose- con ta in ing glycoconjugate selectively expressed in olfac- tory p r imary neu rons (Barber 1989; L u n d h et al. 1989), and the presence of neuron-specif ic enolase (NSE), which is a cytoplasmic pro te in found specifically in neu rons and neu roendoc r ine cells (Marangos and Schmechel 1987) and the decapept ide L H R H . To de termine whether mi- grat ing n e u r o n a l elements exhibit a high level of differen- t iat ion, we have used an an t i body directed against olfac- tory marke r pro te in (OMP), which is expressed by ma- ture olfactory receptor cells ( F a r b m a n a n d Margol is 1980; Mont i -Graz iade i et al. 1980).
Materials and methods
Animals and tissue preparation
Rats (Wistar SPF) from 1-10 days old and embryos fi'om the 13rd (El3) to the 21st (E21) day of gestation were used in this study. Day 1 of pregnancy (El) was designated when we observed a vaginal plug. At least 3 specimens were used for each age group studied. Pregnant dams were anesthetized with an overdose of Nembutal and embryos were taken by Caesarean section. Embryos were quickly decapitated and the heads were immersed in cold Bouin's fixative for 3-4 h for younger embryos and overnight for the others. Most of the specimens were dehydrated through graded alcohols, cleared in xylene, and embedded in Paraplast. Serial sections (6- 9 gm thick) were cut in the sagittal and coronal planes, and were mounted on gelatinized slides. Some specimens were soaked in 15 % phosphate-buffered sucrose overnight at 4~ frozen in isopentane, and serially cut in 10-1~m-thick sagittal sections. After deep anesthe- sia with Nembutal, neonatal rats of 1, 3, 6, 9 and 10 days of age (day of birth = P1) were perfused transcardially with Ringer's solution and then with Bouin's fixative. Nasal cavities and olfactory bulbs were dissected and prepared for histology according to the same procedure.
Immunocytochemistry and lectin binding
Brains and nasal regions were examined by a histochemical method for binding with UEA I and by immunocytochemical procedures for the localization of both NSE (rabbit polyclonal anti-NSE, [m- munotech, Marseille, France) and LHRH (rabbit anti-LHRH, LR1 kindly provided by Dr, Robert Benoit). Some sections were also examined for the localization of OMP (goat anti-OMP kindly pro- vided by Dr. Frank Margolis).
Biotinylated (Vector Laboratories, Burlingame, USA) and per- oxidase-conjugated (Sigma, St Louis, USA) UEA I lectins were used at dilutions of 5 gg/ml and 200 gg/ml, respectively in 0.1 M TRIS buffer, pH 7.2, containing 1 mM CaC12, 1 mM MgC12, 1 mM Mn- C12, 5% normal rat serum. Endogenous peroxidase activity was
blocked with 1% H202 in methanol for 30 min at room tempera- ture. To diminish the non-specific binding of proteins, sections were treated with 2% bovine serum albumin (BSA) in TRIS buffer. Sec- tions were then incubated with the conjugated lectins overnight at 4 ~ C. Sections with the biotinylated lectin were incubated in the avidin-biotin horseradish peroxidase complex (Vectastain ABC kit, Vector Laboratories) for 2 h at room temperature. For all sections, peroxidase activity was visualized by immersion in a solution con- taining 0.05% 3,3' diaminobenzidine tetrahydrochloride (DAB), 0.5% nickel chloride, 0.003% H202 in 0.05 M TRIS buffer, pH 7.6. Some slides were lightly counterstained with cresyl violet. To verify the specificity of the lectin, adjacent sections were incubated with TRIS buffer only.
Different immunocytochemical procedures were used for each antibody tested since they had different origins. For the antibody directed against NSE, all dilutions and rinses were made with 0.1 M TRIS buffer, pH 7.2. These steps were carried out in 0.1 M PBS, pH 7.4 for the antiserum against OMP and in 0.01 M PBS, pH 7.6 for the antiserum against LHRH. In all cases, endogenous peroxi- dase activity was blocked with 0.3% H20 z in methanol for 15 min and sections were treated with 0.1% Triton X-100 in the relevant buffer for 1 h. Sections were then incubated for 2 h in buffer contain- ing 5% BSA and 5%-10% normal goat or rabbit serum.
The immunocytochemical staining with antibodies against NSE and OMP was performed using the peroxidase anti-peroxidase (PAP) method. Sections were incubated in the primary antibodies: anti-NSE (prediluted) for 1-2 h at room temperature and anti-OMP (diluted 1:800) overnight at 4~ Sections were then treated with the appropriate secondary antibodies (diluted 1 : 100) for 1-2 h, and with the appropriate PAP complex (diluted 1:100) for 1-2 h at room temperature. Peroxidase was visualized as described previously for the lectin. Some sections were lightly counterstained with cresyl violet. The specificity of the immunostaining for each primary anti- body was verified by applying appropriate non-immune serum to adjacent sections.
The localization of LHRH was performed using the ABC tech- nique. Sections were incubated in the primary antibody (diluted 1 : 10 000) for 48-72 h at 4~ They were then incubated in biotiny- lated anti-rabbit IgG (diluted 1:300) for 1 h and then in the ABC complex for 2 h. Peroxidase was revealed according to the proce- dure described above. Some sections were lightly counterstained with cresyl violet. To demonstrate the specificity of the antiserum to LHRH, some sections were incubated with normal rabbit serum.
In order to visualize those cells expressing both NSE and UEA I, a double-labeling procedure was used. The sections were incubat- ed in the antibody against NSE and the reaction product was visu- alized with 4-C1-1 naphthol dissolved in 0.05 M TRIS buffer, pH 7.6 (Tramu et al. 1978). The sections were then washed in distilled water and mounted in glycerine. Micrographs were taken and then the blue reaction products formed with 4-C1-1 Naphthol were removed with acetone. Tissues were rehydrated in water. Antibodies were eluted by immersion for 1 min in an oxidizing solution of KMnO4 and H2SO 4 (1 vol 2.5% KMnO4, 1 vol 5% H2SO 4 in 100 vol dis- tilled water). Sections were then decolorized in 0.5% Na2S2SO 5 in distilled water, washed in running tap water, and reimmersed in buffer before carrying out the histochemical technique for the lectin as previously described. To verify the efficiency of the antibody elution, sections were treated with elution buffer after the first im- munocytochemical reaction and then reprocessed in a solution of DAB.
Results
Staining of cellular elements in the main olfactory epithelium
NSE-posi t ive react ions were first observed on fetal day 14. At this embryon ic stage, only few receptor cells were

589
Fig. la-f . Sagittal sections of the main olfactory epithelium of rat embryo stained with the antibody against NSE (a, b, e) and with UEA I lectin (d, e, t). a At El4, few receptor cells show NSE im- munoreactivity. Arrow indicates a stained dendrite. A thick strongly stained bundle of axons with cords of labeled migrating cells (arrow- heads) can be seen in the lamina propria, by Blood vessel, b At El5, numerous NSE-reactive receptor cells are observed throughout the
olfactory epithelium, ax Olfactory axon fascicles; by blood vessel, e At El8, the adult pattern of NSE-staining is observed. Only the deepest epithelial layers (arrowheads) are immunonegative, fl At El5, the UEA-I-binding receptor cells are rare. The cell body (c) and the dendrite (d) appear well-stained, e At El8, numerous UEA- I-stained primary neurons are observed, f At E21, the adult pattern of UEA-I-labeling is observed. All figures: x 500

590
NSE-immunoreactive above the background labeling. The staining was mainly located in the perikaryal region, but a few labeled dendrites could be also detected (Fig. la). Immunopositive neurons were distributed through- out the epithelial sheet. Thin immunoreactive axon fasci- cles were observed in the lamina propria. On El5, the number of primary NSE-positive neurons had markedly increased and their staining appeared to be more intense than at E14. The axon fascicles were densely labeled in the lamina propria, whereas most dendritic processes were immunonegative (Fig. lb). From the 16th day of gestation onward, most of the receptor cells exhibited moderate or strong NSE immunoreactivity that was mainly located in the perikaryon and the axon (Fig. lc). Cellular elements confined to the deepest epithelial layers were always negative.
UEA I binding first appeared on fetal day 15 and its developmental pattern of expression was similar to that of NSE. At E 15 and E 16, only few scattered receptor cells showed moderate labeling spreading in all cell regions
(Fig. ld). In the lamina propria, axon fascicles that were negative at El5, became weakly positive at E16. The number of UEA-I-positive neurons subsequently in- creased rapidly (Fig. le,f), and an adult pattern of stain- ing was observed from the 21st day of gestation. Most of the positive receptor cells exhibited moderate or strong labeling in all cell regions, and axon fascicles appeared well-labeled in the lamina propria. Cellular elements that were confined to the deepest epithelial layers remained negative.
Staining of cells migrating from the lateral olfactory placode
On El3, thin olfactory axon fascicles accompanied by numerous migrating cells emerged from the lateral part of the olfactory placode, and grouped together in the lamina propria where they formed a large fibrillo-cellular aggregate having no contact with the developing fore-
Fig. 2a-d. Sagittal sections through the lateral part of the olfactory pit from embryonic rats on days 14 (a) and 15 (b, e, d). Reaction with the antibody against NSE. a At E14, NSE-immunoreactive axons and their associated migrating cells form a large fibrillo-cellular aggregate or neural blastema (nb) in the mesenchyme. A thick fiber bundle with cords of migrating cells (arrow) extends obliquely from the caudo-lateral part of the olfactory pit (op) to the blastema, t Telencephalic vesicle; vno anlage of the vomeronasal organ, x 100. b At E15, immunoreactive axon fascicles with their associated mi-
grating cells (arrows) leave the dorsal and lateral parts of the olfac- to ry pit (op) and group together into the neural blastema (nb). t Telencephalic vesicle, x 70. e At higher magnification, the reactive migrating cells (arrow) accompanying an olfactory axon fascicle (ax) have a round or fusiform shape with a nucleus that occupies a large part of the cell perikaryon, oe Olfactory epithelium, x 550. d Nu- merous strongly NSE-immunoreactive cells can be seen in the neu- ral blastema (nb). oe Olfactory epithelium, x 550

591
Fig. 3a--d. Sagittal sections through the lateral part of the olfactory pit from 15-day-old embryonic rats. Reaction with UEA I (a, b, d) and the antibody against NSE (c). a Only few migrating cells (ar- rows) exhibit strong UEA I binding along the olfactory axon fasci- cles (ax). A noticeable number of positive cells can be observed in the neural blastema (nb). oe Olfactory epithelium, x 250. b At higher magnification, a UEA-I-binding migrating cell exhibiting a short positive process (arrow) can be seen along an olfactory axon fascicle (ax). oe Olfactory epithelium, x 625. c, d The double-label procedure shows that only a subset of the NSE-immunoreactive cells (c) is UEA-I-positive (d) along the olfactory axon fascicles (ax)
leaving the olfactory epithelium (oe) and in the neural blastema (nb). t Telencephalic vesicle, x 250
Figs. 4, 5. Sagittal sections through the lateral part of the olfactory pit from embryonic rats on days 16 and 18.
Fig. 4. Strongly NSE-immunoreactive migrating cells (arrowheads) are observed, singly or in small clusters, along the olfactory axon fascicles (ax) emerging from the olfactory epithelium (oe). x 250
Fig. ft. The arrow indicates a UEA-I-binding cell leaving the olfac- tory epithelium (oe). x 625

592
brain. This aggregate that Lejour (1967) has termed the neural blastema corresponds to the presumptive olfacto- ry nerve layer. At this embryonic stage, neither NSE im- munoreactivity nor UEA I binding was detected along olfactory fascicles and in the neural blastema.
On E14, the neural blastema lay nearer to the telen- cephalic vesicle and its rostral region was in direct con- tact with the developing brain (Fig. 2a). At this stage, olfactory axons with their associated cells appeared strongly NSE-immunoreactive (Figs. la,2a). A thick fiber bundle with cords of migrating cells was seen extending obliquely from the caudo-lateral part of the nasal pit up to the neural blastema. As shown in Fig. la, NSE-im- munoreactive migrating cells were closely clumped to- gether. Unlike the antibody against NSE, the UEA I lect- in bound only to few cellular elements in the neural blastema and the labeling was weak (data not shown).
At 15 days of gestation, the neural blastema, of which the rostral portion capped the ventral surface of the ros- tral forebrain, became more prominent by the addition of olfactory fascicles emerging from the dorsal parts of the olfactory pit (Fig. 2b). The migrating cells accompanying these fascicles were strongly NSE-immunoreactive. They were characterized by a round or fusiform shape and their nucleus occupied a large part of the cell perikaryon (Fig. 2c). The labeling was confined to the cytoplasm. In the neural blastema, a large number of cells also exhibit- ed strong NSE-staining (Fig. 2d). With the UEA I lectin, only few migrating cells exhibited strong positive staining along the negative olfactory axons, whereas a noticeable number of positive cells were observed in the neural blastema (Fig. 3a). Some of the stained cellular elements had short labeled processes (Fig. 3b). Experiments using the double-label procedure with the antibody against NSE and UEA I showed that the lectin recognized only a subset of the NSE-immunoreactive cells in the blastema and a few migrating cells along the olfactory axons (Fig. 3c,d).
On El6, the neural blastema whose the caudal part was now in direct contact with the telencephalic vesicle was no longer separated from the brain, and thus became the definitive olfactory nerve layer. Single or clusters of 2-5 migrating cells that exhibited strong NSE immunore- activity and UEA I binding were observed along thin axon fascicles emerging from the olfactory epithelium lin- ing developing ethmoid turbinates (Fig. 4). Some of these cells were seen migrating from the olfactory epithelium (Fig. 5). The large fibrillo-cellular bundle, which formed a bridge between the caudal part of the olfactory pit and the blastema in earlier embryonic stages, was no longer observed. Strongly lectin-positive cells were also seen scattered throughout the olfactory nerve layer (Fig. 6), but their number was reduced compared with El5 em- bryos. No positive cells were seen penetrating the pre- sumptive bulb beyond the olfactory nerve layer. With the NSE antibody, no immunoreactive cells could be detect- ed within the olfactory nerve layer because of the strong labeling of the olfactory fibers.
A decrease in the number of NSE- and UEA I-posi- tive cells was observed during the next few embryonic days in the lamina propria. From the 18th day of gesta-
tion, the few lectin-binding cells found in the olfactory nerve layer appeared to be mainly confined to the periph- ery of this layer. From postnatal day 9, no positive cellu- lar elements were detected in these two structures.
Some nasal cavity sections of 16- and 18-day fetuses and of newborn rats (P3) were reacted with the antiserum against OMP. At El6, neither the olfactory epithelium nor the migrating cells showed OMP immunoreactivity. For the two other developmental stages, a few strongly OMP-positive cells were observed along axon fascicles or at the periphery of the olfactory nerve layer (Fig. 7).
For any given embryonic stage, no LHRH immunore- activity was detected along the olfactory fascicles emerg- ing from the lateral part of the nasal pit or in the olfacto- ry nerve layer formed by these fascicles.
Staining of cells migrating from the medial olfactory placode
NSE immunoreactivity first occurred in vomeronasal ax- on fascicles and their numerous associated cells by El4. The staining of axons and migrating cells, which was moderate at E14, became intense on the 15th day of ges- tation (Fig. 8a). Migrating cells appeared to be closely packed together, and formed thick cords as they left the epithelium of the vomeronasal organ anlage and inter- mingled with vomeronasal fascicles (Fig. 8b). These cords of cells were observed throughout the nasal septum. On E15, moderately staining NSE-immunoreactive cells were also seen within the vomeronasal epithelium along the intraepithelial course of axon fascicles (Fig. 8b). At this developmental stage, NSE-immunoreactive fibers ac- companied by few stained cells were observed entering the forebrain medially, between the developing olfactory bulbs. On the next 3 embryonic days, thick fibrillo-cellu- lar bundles could still be observed coursing across the nasal septum to the forebrain. In 18-day embryos, a clus- ter of strongly NSE-positive cells was observed medially at the level of the presumptive cribiform plate of the eth- moid bone. This group of cells was different from the developing ganglion terminal, which was located on the medial-caudal surface of the olfactory bulb and in which NSE-positive cells could also be seen (data not shown). From the 21st day of gestation, the migrating NSE-la- beled cells associated with the vomeronasal bundles ap- peared to be preferentially located at the level, or beyond, the cribiform plate pores and were only occasionally ob- served singly or in small clusters of few cells throughout the nasal septum.
The first binding activity with the lectin was observed on El5. Compared with NSE, UEA I bound to only a few migrating cells and the labeling had a supranuclear location that probably corresponded to the Golgi ap- paratus. From El6 onward, an increased number of UEA-I-positive migrating cells was observed but these cells always represented a subset of the NSE-positive cell population found along the course of the vomeronasal fascicles and/or the terminal nerve (Fig. 9).
LHRH reactivity was first detected on E15 in the brain and in the medial part of the neural blastema where

Figs. 6, 7. Sagittal sections through the lateral part of the nasal cavity from embryonic rats on days 16 (6) and 18 (7)
Fig. 6. UEA-I-binding cells are scattered throughout the developing olfactory nerve layer (onI). ob Olfactory bulb. x 250
Fig. 7. A few strongly OMP-immunoreactive migrating cells (ar- rows) lie along the olfactory axon fascicles and at the periphery of the olfactory nerve layer (onl). oe Olfactory epithelium, x 300
Fig. 8a, b. Sagittal section through the medial part of the olfactory pit from a 15-day-old embryonic rat. Reaction with the antibody against NSE. a Numerous NSE-immunoreactive cells (arrowheads) migrate along the vomeronasal axon fascicles toward the telen- cephalic vesicle (t). nb Neural blastema; vno anlage of the vomeronasal organ, x 60. h Higher magnification of the
vomeronasal axon fascicles and their associated migrating cells, vno Anlage of the vomeronasal organ, x 375
Fig. 9. Sagittal section through the anlage of the vomeronasal organ (vno) from a 16-day-old embryonic rat showing numerous UEA-I- positive migrating cells along the vomeronasal axon fascicles (ax). x 300
Figs. 10, 11. Sagittal sections through the medial part of the olfacto- ry pit reacted with antiserum against LHRH
Fig. 10. At embryonic day 15, the first immunoreactive-LHRH neu- rons (arrows) are detected in the medial part of the neural blastema (nb) where the vomeronasal and/or terminal nerves converge. x 320
594
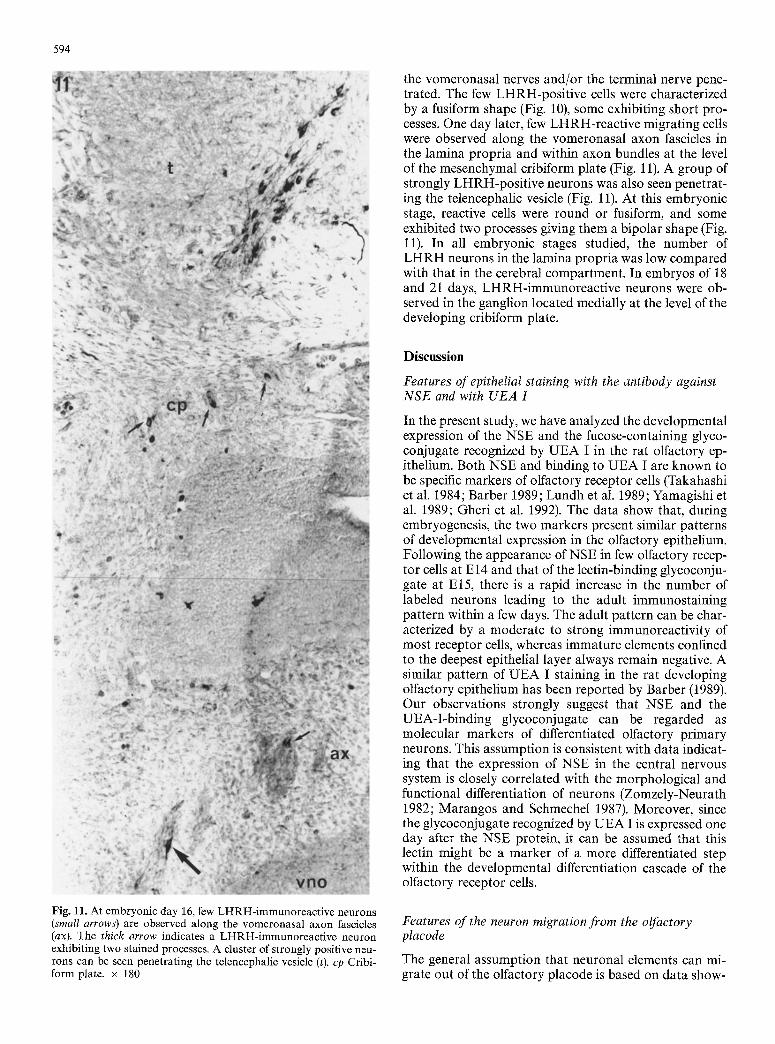
the vomeronasal nerves and/or the terminal nerve pene- trated. The few LHRH-positive cells were characterized by a fusiform shape (Fig. 10), some exhibiting short pro- cesses. One day later, few LHRH-reactive migrating cells were observed along the vomeronasal axon fascicles in the lamina propria and within axon bundles at the level of the mesenchymal cribiform plate (Fig. 11). A group of strongly LHRH-positive neurons was also seen penetrat- ing the telencephalic vesicle (Fig. 11). At this embryonic stage, reactive cells were round or fusiform, and some exhibited two processes giving them a bipolar shape (Fig. 11). In all embryonic stages studied, the number of LHRH neurons in the lamina propria was low compared with that in the cerebral compartment. In embryos of 18 and 21 days, LHRH-immunoreactive neurons were ob- served in the ganglion located medially at the level of the developing cribiform plate.
Fig. 11. At embryonic day 16, few LHRH-immunoreactive neurons (small arrows) are observed along the vomeronasal axon fascicles (ax). The thick arrow indicates a LHRH-immunoreactive neuron exhibiting two stained processes. A cluster of strongly positive neu- rons can be seen penetrating the telencephalic vesicle (t). cp Cribi- form plate. • 180
Discussion
Features of epithelial staining with the antibody against NSE and with UEA I
In the present study, we have analyzed the developmental expression of the NSE and the fucose-containing glyco- conjugate recognized by UEA I in the rat olfactory ep- ithelium. Both NSE and binding to UEA I are known to be specific markers of olfactory receptor cells (Takahashi et al. 1984; Barber 1989; Lundh et al. 1989; Yamagishi et al. 1989; Gheri et al. 1992). The data show that, during embryogenesis, the two markers present similar patterns of developmental expression in the olfactory epithelium. Following the appearance of NSE in few olfactory recep- tor cells at El4 and that of the lectin-binding glycoconju- gate at El5, there is a rapid increase in the number of labeled neurons leading to the adult immunostaining pattern within a few days. The adult pattern can be char- acterized by a moderate to strong immunoreactivity of most receptor cells, whereas immature elements confined to the deepest epithelial layer always remain negative. A similar pattern of UEA I staining in the rat developing olfactory epithelium has been reported by Barber (1989). Our observations strongly suggest that NSE and the UEA-I-binding glycoconjugate can be regarded as molecular markers of differentiated olfactory primary neurons. This assumption is consistent with data indicat- ing that the expression of NSE in the central nervous system is closely correlated with the morphological and functional differentiation of neurons (Zomzely-Neurath 1982; Marangos and Schmechel 1987). Moreover, since the glycoconjugate recognized by UEA I is expressed one day after the NSE protein, it can be assumed that this lectin might be a marker of a more differentiated step within the developmental differentiation cascade of the olfactory receptor cells.
Features of the neuron migration from the olfactory placode
The general assumption that neuronal elements can mi- grate out of the olfactory placode is based on data show-

595
ing migrating cells with a Koelle-positive reaction for acetylcholinesterase (Filogamo and Robecchi 1969) or having ultrastructural features similar to those of some cells observed within the developing olfactory epithelium (Mendoza et al. 1982). Using immunocytochemical stain- ing of NSE, we have demonstrated a large population of migrating neurons originating from the epithelium of the olfactory pit. These neurons are closely associated not only with the vomeronasal axon fascicles emerging from the medial region of the pit, but also with olfactory axons originating from the lateral region. One should empha- size that the migration of neurons from the lateral pit appears to be more discrete than that from the medial pit. As also noted by Schwanzel-Fukuda et al. (1985), a thick fibrillo-cellular bundle emerging from the olfactory ep- ithelium is observed exclusively at E14 and E15, whereas the thick cords of NSE-stained neurons accompanying the vomeronasal nerves and/or the terminal fibers are seen until at least the 18th day of gestation. As soon as the turbinates begin to develop, multiple thin olfactory fascicles emerge that are associated with only single or clusters of few neurons. The migrating neurons can be immunocytochemically identified from the 14th day of gestation and their initial expression of the NSE protein is concomitant with the appearance of NSE reactivity in some receptor cells in the olfactory epithelium. Because NSE is expressed only by differentiated olfactory recep- tor cells, it can be assumed that NSE-immunoreactive migrating cells correspond to differentiated postmitotic neurons.
The migration of differentiated neurons along the vomeronasal and olfactory axon fascicles is confirmed by the presence of cells expressing the glycoconjugate recog- nized by UEA I lectin. The expression of other cell-sur- face glycoconjugates, such as the CC2 antigen, has also been reported for some neurons migrating from the olfac- tory placode (Schwarting et al. 1991, 1992). In the early stages of migration, however, the spatio-temporal pat- tern of UEA I labeling appears to be different from that seen for NSE. A remarkable feature of this pattern is that UEA-I-binding neurons are first observed at E14 in the fibrillo-cellular aggregate located in the mesenchyme, whereas no lectin labeling is seen in the olfactory epithe- lium from which these neurons presumably originate. One day later, an increased number of cells in the fibrillo- cellular aggregate express the lectin-bound glycoconju- gate, whereas few UEA-I-positive cells are now seen as- sociated with axon fascicles. Such observations strongly suggest that some cell differentiation occurs along the migratory route, at least in earlier developmental stages of the pit. From E18, when most receptor cells express the lectin-bound glycoconjugate, it is more probable that the neurons migrating from the olfactory epithelium are also lectin-positive.
Cytoplasmic OMP is a typical protein of mature olfac- tory receptor neurons, whose the expression during onto- genesis has been analyzed in detail in rats and mice (Farbman and Margolis 1980; Monti-Graziadei et al. 1980; Margolis 1988). As also reported by Schwanzel- Fukuda et al. (1992), no OMP labeling can be detected among migrating cells associated with the vomeronasal
nerves and/or the terminal fibers. Nervertheless, we have seen few OMP-immunoreactive migrating cells accompa- nying the olfactory axon fascicles in El8 and P3 animals. Recent studies have also shown the presence of OMP-im- munoreactive cells along the olfactory nerves in rats from E16 (Baker and Farbman 1993; Valverde et al. 1993). Mature migrating neurons have previously been ob- served in studies using the B determinant of ABH blood group antigens, which can be regarded, like OMP, as a marker of mature olfactory receptor cells in rats (Astic et al. 1989). The B antigen is expressed by migrating cells associated with olfactory axons in rat embryos from the 16th day of gestation (Mollicone et al. 1985; Pellier et al. 1992), and the presence of the B antigen in these migrat- ing cells coincides with the appearance of B reactivity in few receptor cells in the olfactory epithelium (Astic et al. 1989). These data seem to indicate that mature neurons migrate from the olfactory epithelium when the latter has reached a specific degree of maturation.
The absence of NSE immunoreactivity and UEA I binding in the nasal mesenchyme at El3, when axon fas- cicles and migrating cells form a fibrillo-cellular aggre- gate, does not exclude the possibility that some undiffer- entiated postmitotic neurons might be present among the migrating cells found in the lamina propria. It should be noted that the only marker to be expressed in cells leav- ing the olfactory epithelium at these early embryonic stages is the neural cell adhesion molecule (NCAM). In 13-day-old rat embryos, staining of migrating cells has been observed in the nasal mesenchyme with the mono- clonal antibody Neu 5, which recognizes NCAM epi- topes (Cart et al. 1989). In embryonic mice, the presence of NCAM-immunoreactive migrating cells from the ol- factory pit is concomitant with the appearance of the migratory process on the 10th day of gestation (Schwanzel-Fukuda et al. 1992). However, although NCAM is expressed by most of the postmitotic olfactory primary neurons including those that are undifferentiat- ed (Calof and Chikaraishi 1989; Terkelsen et al. 1989), and by olfactory ensheathing glial cells (Miragall and Dermietzel 1992), the nature of these NCAM-positive migrating cells remains unclear. Indeed, during the early developmental stages of the olfactory pit, it is difficult to distinguish, immunocytochemically, migrating neurons from the ensheathing cells that are also thought to origi- nate from the developing olfactory epithelium (Cuschieri and Bannister 1975b; Farbman and Squinto 1985; Mar- in-Padilla and Amieva 1989; Chuah and Au 1991; Valverde et al. 1992). Thus, the initial staining of migrat- ing neurons with NSE is seen from El4, whereas Chuah and Au (1991) have observed that ensheathing cells ex- press the glial fibrillary acidic protein (GFAP), a specific marker of this cell type (Barber and Lindsay 1982), only from the 16th day of gestation.
There is a convergence of data indicating that the LHRH neuronal system in mammalian, avian, and am- phibian species originates from the medial olfactory pla- code, and that LHRH neurons migrate into the brain along the terminal nerve and/or the olfactory nerve dur- ing development (Schwanzel-Fukuda and Pfaff 1989, 1990; Wray et al. 1989a,b; Ronnekleiv and Resko 1990;

596
Daikoku-Ishido et al. 1990; Murakami et al. 1991, 1992; Norgren and Lehman 1991). As we had demonstrated that migrating neurons also originate from the lateral olfactory pit, it became pertinent to verify whether some of these migrating cells might be LHRH neurons. No LHRH immunoreactivity has been detected in neurons migrating from the olfactory epithelium; LHRH-positive neurons are exclusively found throughout the course of the vomeronasal nerves and/or the terminal nerve. The absence of LHRH-immunoreactive neurons in the axon fascicles originating from the lateral part of the olfactory pit has also recently been reported by Schwanzel-Fukuda et al. (1992) and by Baker and Farbman (1993). These observations confirm that the LHRH neuronal system originates exclusively from the anlage of the vomeronasal organ. The spatio-temporal pattern of the migration of the LHRH neurons is comparable to that previously re- ported in rats (Schwanzel-Fukuda et al. 1985; Jennes t989; S~t~il6 et al. 1992). Nevertheless, we have seen no evidence of either stained neurons in the vomeronasal epithelium or continuous cords of LHRH cells across the nasal mesenchyme, as described in mice with the same LR1 antiserum (Schwanzel-Fukuda and Pfaff 1989; Wray et al. 1989a,b; Zheng et al. 1992). Even when using an antiserum against rat gonadotropic hormone-releasing hormone associated peptide (28-56, rGAP), only occa- sional immunoreactive neurons have been detected in the rat vomeronasal epithelium from E13.5 (Daikoku-Ishido et al. 1990). Thus, the differences observed between rats and mice do not appear to be related to the antibodies tested, but seem to reflect species specificities. The ab- sence of cells expressing the mature decapeptide in the pit epithelium of the rat has led Schwanzel-Fukuda and Pfaff (1991) to propose that LHRH neurons in this species do not begin to synthesize the decapeptide until they mi- grate out onto the nasal septum. A confirmation of this hypothesis has recently been demonstrated by $6t~16 et al. (1992) who have shown that the number of LHRH- positive cells detected in the nasal placodal area during fetal life is always lower than that found in the brain.
The developmental pattern of the olfactory nerve lay- er as shown with NSE and UEA I is similar to that described by Lejour (1967) who used an alkaline phos- phatase reaction, and by Schwanzel-Fukuda et al. (1992) using NCAM as marker. In the early developmental stages, this pattern is mainly characterized by the forma- tion of a fibrillo-cellular aggregate in the nasal mes- enchyme. This mass, which corresponds to the presump- tive olfactory nerve layer, migrates and comes into direct contact with the developing forebrain. Our study has provided evidence for the presence of a large number of strongly NSE-immunoreactive and UEA-I-binding neu- rons in this presumptive olfactory nerve, which is mainly formed by the olfactory axon fascicles originating from the dorsal and lateral parts of the olfactory pit. In addi- tion to these neural elements, the developing olfactory nerve layer contains immature ensheathing cells that are known to migrate from the olfactory epithelium (Lejour 1967; Doucette 1989; Marin-Padilla and Amieva 1989; Valverde et al. 1992). Although a large and increasing number of glial ensheathing cells resulting from local mi-
totic activity has been reported during embryonic devel- opment (Valverde et al. 1992), the number of NSE-im- munoreactive and UEA-I-binding cells show a dramatic decrease from the 16th day of gestation; no stained cells are detected in the olfactory nerve layer beyond postnatal day 9. We do not know what is the significance of this disappearance of the NSE-immunoreactive and UEA-I- binding neurons from the developing olfactory nerve lay- er. There is no direct evidence that these neurons are incorporated into the bulbar neural network as proposed by Valverde et al. (1992), since no lectin-binding cells can be seen penetrating the olfactory bulb. What is more probable is that the glycoconjugate is not expressed by the olfactory bulb except for transient weak binding in the mitral cell perikarya between the 18th and 21th days of gestation. Such a transient expression of UEA-I- bound glycoconjugate may be related to some recogni- tion process between the olfactory axons and their targets. Specific carbohydrates expressed on the cell sur- face are thought to play a prominent role in cell-cell in- teractions (Pfenninger and Mayli&Pfenninger 1979; Schwarting and Yamamoto 1988). In addition, one can- not exclude the possibility that neurons migrating from the olfactory pit might undergo dedifferentiation or alter their phenotype after migration and thus become unde- tectable. The neuron decrease might also be the result of programmed cell death. Furthermore, during embryoge- nesis, neurons migrating from the olfactory epithelium might play a prominent role in the guidance of outgrow- ing axons in the nasal mesenchyme without having specific targets in the central nervous system. Pro- grammed cell death following neurogenesis can occur and is usually dependent on target influences (Cunning- ham 1982; Oppenheim 1991).
It should be noted that the capacity of the olfactory epithelium to provide migrating neural elements does not appear to be a typical feature of embryogenesis. The oc- currence of neuron migration has also been reported in newborn and adult rats after bulbectomy (Graziadei and Monti-Graziadei 1986; Monti-Graziadei 1992) and in cases of brain transplants of olfactory mucosa (Morrison and Graziadei 1983; Barber and Jensen 1988; Monti- Graziadei and Morrison 1988; Monti-Graziadei and Graziadei 1989). Under these experimental conditions, however, most if not all migrating cells have been identi- fied as immature olfactory neurons resembling globose basal cells. Moreover, no differentiation process seems to occur along the migratory route to the olfactory bulb or into the host tissue. Thus, although the functions of these migrating cells remain to be determined during embryo- genesis and in bulbectomized postnatal animals, they are probably different.
Acknowledgements. This research was supported by the CNRS (URA 180). We express our thanks to Dr. D. Saucier for the thor- ough reading and helpful comments on this manuscript, and to Dr. R.M. Babitch for reviewing the English.
References Astic L, Le Pendu J, Mollicone R, Saucier D, Oriol R (1989) Cellu-
lar expression of H and B antigens in the rat olfactory system during development. J Comp Neurol 289:386-394

597
Baker H, Farbman AI (1993) Olfactory afferent regulation of the dopamine phenotype in the fetal rat olfactory system. Neuro- science 52:115-134
Barber PC (1989) Ulex europaeus agglutinin I binds exclusively to primary olfactory neurons in the rat nervous system. Neuro- science 30:1-9
Barber PC, Jensen S (1988) Olfactory tissue interactions studied by intraocular transplantation. In: Margolis FL and Getchell TV (eds) Molecular neurobiology of the olfactory system. Plenum Press, New York, pp 333-350
Barber PC, Lindsay RM (1982) Schwann cells of the olfactory nerves contain glial fibrillary acidic protein and resemble astro- cytes. Neuroscience 7: 3077-3090
Bossy J (1980) Development of olfactory and related structures in staged human embryos. Anat Embryol (Berl)161:225-236
Calof AL, Chikaralshi DM (1989) Analysis of neurogenesis in a mammalian neuroepithelium: proliferation and differentiation of an olfactory neuron precursor in vitro. Neuron 3:115-127
Carr V, Farbman AI, Lidow MS, Colletti LM, Hempstead JL, Mor- gan JI (1989) Developmental expression of reactivity to mono- clonal antibodies generated against olfactory epithelia. J Neu- rosci 9:1179-1198
Chuah MI, Au C (1991) Olfactory Schwann cells are derived from precursor cells in the olfactory epithelium. J Neurosci Res 29:172-180
Cunningham TJ (1982) Naturally occurring neuron death and its regulation by developing neural pathways. Int Rev Cytol 79:163-186
Cuschieri A, Bannister LH (1975a) The development of the olfacto- ry mucosa in the mouse: light microscopy. J Anat 119:277-286
Cuschieri A, Bannister LH (1975b) The development of the olfacto- ry mucosa in the mouse: electron microscopy. J Anat 119:471- 49
Daikoku-Ishido H, Okamura Y, Yanalhara N, Daikoku S (1990) Development of the hypothalamic luteinizing hormone-releas- ing hormone-containing neuron system in the rat: in vivo and in transplantation studies. Dev Biol 140:374-38
Doucette R (1989) Development of the nerve fiber layer in the olfactory bulb of mouse embryos. J Comp Neurol 285:514-52
Farbman AI (1991) Developmental neurobiology of the olfactory system. In: Getchell TV, Bartoshuck LM, Doty RL, Snow JB (eds) Smell and taste in health and disease. Raven Press, New York, pp 19-33
Farbman AI, Margolis FL (1980) Olfactory marker protein during ontogeny: immunohistochemical localization. Dev Biol 74:205- 215
Farbman AI, Squinto LM (1985) Early development of olfactory receptor cell axons. Dev Brain Res 19:205~13
Filogamo G, Robecchi MG (1969) Neuroblasts in the olfactory pits in mammals. Acta Anat 73 [Suppl 56]: 182-187
Gheri G, Gheri Bryk S, Balboni GC (1992) Identification of sugar residues in human fetal olfactory epithelium using lectin histo- chemistry. Acta Anat 145:167-174
Graziadei PPC, Monti-Graziadei GA (1986) Neurogenesis and plasticity of the olfactory sensory neurons. Ann NY Acad Sci 457:127-142
Jennes L (1989) Prenatal development of the gonadotropin-releas- ing hormone-containing systems in rat brain. Brain Res 482:97- 108
Lejour M (1967) Activit6 de quatre enzymes d6phosphorylants au cours de la morphogenbse du palais primaire chez le rat (phos- phatase alcaline, phosphatase acide, ATP- et AMPases). Arch Biol (Li6ge) 78 : 389-450
Lundh B, Brockstedt U, Kristensson K (1989) Lectin-binding pat- tern of neuroepithelial and respiratory epithelial cells in the mouse nasal cavity. Histochem J 21:33-43
Marangos PJ, Schmechel DE (1987) Neuron specific enolase, a clin- ically useful marker for neurons and neuroendocrine cells. Annu Rev Neurosci 10:269-295
Margolis, FL (1988) Molecular cloning of olfactory specific gene products. In: Margolis FL, Getchell TV (eds) Molecular neuro-
biology of the olfactory system. Plenum Press, New York, pp 237-265
Marin-Padilla M, Amieva MR (1989) Early neurogenesis of the mouse olfactory nerve: Golgi and electron microscopic studies. J Comp Neurol 288:339-352
Mendoza AS, Breipohl W, Miragall F (1982) Cell migration from the chick olfactory placode: a light and electron microscopic study. J Embryol Exp Morphol 69:47-59
Miragall F, Dermietzel R (1992) Immunocytochemical localization of cell adhesion molecules in the developing and mature olfacto- ry system. Microsc Res Technique 23:157-172
Mollicone R, Trojan J, Oriol R (1985) Appearance of H and B antigens in primary sensory cells of the rat olfactory apparatus and inner ear. Dev Brain Res 17:275-279
Monti-Graziadei AG (1992) Cell migration from the olfactory neu- roepithelium of neonatal and adult rodents. Dev Brain Res 70:65 74
Monti-Graziadei AG, Graziadei PPC (1989) Experimental studies on the olfactory marker protein. V. Olfactory marker protein in the olfactory neurons transplanted within the olfactory bulb. Brain Res 484:157-167
Monti-Graziadei AG, Morrison EE (1988) Experimental studies on the olfactory marker protein. IV. Olfactory marker protein in the olfactory neurons transplanted within the brain. Brain Res 455:401-406
Monti-Graziadei GA, Stanley RS, Graziadei PPC (1980) The olfac- tory marker protein in the olfactory system of the mouse during development. Neuroscience 5:1239-1252
Morrison EE, Graziadei PPC (1983) Transplants of olfactory mu- cosa in the rat brain. I. A light microscopic study of transplant organization. Brain Res 279:241-245
Murakami S, Seki T, Wakabayashi K, Aral Y (1991) The ontogeny of luteinizing hormone-releasing hormone (LHRH) producing neurons in the chick embryo: possible evidence for migrating LHRH neurons from the olfactory epithelium expressing a high- ly polysialylated neural cell adhesion molecule. Neurosci Res 12:421-431
Murakami S, Kikuyama S, Arai Y (1992) The origin of the luteiniz- ing hormone-releasing hormone (LHRH) neurons in newts (Cynops pyrrhogaster): the effect of olfactory placode ablation. Cell Tissue Res 269:21-27
Norgren RB, Lehman MN (1991) Neurons that migrate from the olfactory epithelium in the chick express luteinizing hormone- releasing hormone. Endocrinology 128:1676-1678
Oppenheim RW (1991) Cell death during development of the ner- vous system. Annu Rev Neurosci 14:453-501
Pellier V, Astic L, Mollicone R, Oriol R (1992) Etude de la migra- tion cellulalre au niveau du systbme olfactif p&iph6rique au cours du d6veloppement. Abstr Soc Neurosci Fr 1:251
Pfenninger KH, Mayli6-Pfenninger MF (1979) Surface glycoconju- gates in the differentiating neuron. In: Margolis RU, Margolis RK (eds) Complex carbohydrates of nervous tissue. Plenum Press, New York, pp 185-191
Ronnekleiv OK, Resko JA (1990) Ontogeny of gonadotropin-re- leasing hormone-containing neurons in early fetal development of rhesus macaques. Endocrinology 126:498-511
Schwanzel-Fukuda M, Pfaff DW (1989) Origin of luteinizing hor- mone-releasing hormone neurons. Nature 338:161-164
Schwanzel-Fukuda M, Pfaff DW (1990) The migration of luteinizing hormone-releasing hormone (LHRH) neurons from the medial olfactory placode into the medial basal forebrain. Experientia 46: 956-96
Schwanzel-Fukuda M, Pfaff DW (1991) Migration of LHRH-im- munoreactive neurons from the olfactory placode rationalizes olfacto-hormonal relationships. J Steroid Biochem Mol Biol 39:565-572
Schwanzel-Fukuda M, Morrell JI, Pfaff DW (1985) Ontogenesis of neurons producing luteinizing hormone-releasing hormone (LHRH) in the nervus terminalis of the rat. J Comp Neurol 238: 348-364

598
Schwanzel,Fukuda M, Abraham S, Crossin KL, Edelman GM, Pfaff DW (1992) Immunocytochemical demonstration of neural cell adhesion molecule (NCAM) along the migration route of luteinizing hormone-releasing hormone (LHRH) neurons in mice. J Comp Neurol 321:1-18
Schwarting GA, Yamamoto M (1988) Expression of glycoconju- gates during development of the vertebrate nervous system. Bioessays 9:19-23
Schwarting GA, Drinkwater DL, Crandall JE, Tobet SA (1991) Luteinizing hormone-releasing hormone (LHRH) neurons mi- grate along vomeronasal and olfactory axons which express CC2 glycoconjugates in rats. Soc Neurosci Abstr 17:427
Schwarting GA, Deutsch G, Gattey DM, Crandall JE (1992) Glyco- conjugates are stage- and position-specific cell surface molecules in the developing olfactory system. 2. Unique carbohydrate anti- gens are topographic markers for selective projection patterns of olfactory axons. J Neurobiol 23:130-142
S6t/tl6 G, Hagino N, Dittrich E (1992) Ontogenesis of the GnRH neuron system in the rat. A quantitative immunohistochemical study with special reference to the extra cerebral GnRH-positive cells and the occupation of intracerebral termination fields. Neu- ropeptides 21:93-103
Takahashi S, Iwanaga T, Takahashi Y, Nakano Y, Fujita T (1984) Neuron-specific enolase, neurofilament protein and S-100 protein in the olfactory mucosa of human fetuses. An immuno- histochemical study. Cell Tissue Res 238:231-234
Terkelsen OBF, Bock E, Mollg~rd K (1989) NCAM and Thy-1 in special sense organs of the developing mouse. Anat Embryol (Berlin) 179: 311-3 t 8
Tramu G, Pillez A, Leonardelli J (1978) An efficient method of antibody elution for the successive or simultaneous localization of two antigens by immunocytochemistry. J Histochem Cy- tochem 26:322-324
Valverde F, Santacana M, Heredia M (1992) Formation of an olfac- tory glomerulus: morphological aspects of development and or- ganization. Neuroscience 49:255-275
Valverde F, Heredia M, Santacana M (1993) Characterization of neuronal cell varieties migrating from the olfactory epithelium during prenatal development in the rat. Immunocytochemical study using antibodies against olfactory marker protein (OMP) and luteinizing hormone-releasing hormone (LH-RH). Dev Brain Res 71:209-220
Van Campenhout E (1956) Le d~veloppement embryonnalre corn- par6 des nerfs olfactif et auditif. Acta Otorhinolaryngol Belg 11:279-287
Wray S, Grant P, Gainer H (1989a) Evidence that cells expressing luteinizing hormone-releasing hormone mRNA in the mouse are derived from progenitor cells in the olfactory placode. Proc Natl Acad Sci USA 86:813~8136
Wray S, Nieburgs A, Elkabes S (1989b) Spatiotemporal cell expres- sion of luteinizing hormone-releasing hormone in the prenatal mouse: evidence for an embryonic origin in the olfactory pla- code. Dev Brain Res 46:309-318
Yamagishi M, Hasegawa S, Nakano Y, Takahashi S, Iwanaga T (1989) Immunohistochemical analysis of the olfactory mucosa by use of antibodies to brain proteins and cytokeratin. Ann Otol Rhinol Laryngol 98:384-388
Zheng LM, Pfaff DW, Schwanzel-Fukuda M (1992) Electron micro- scopic identification of luteinizing hormone-releasing hormone- immunoreactive neurons in the medial olfactory placode and basal forebrain of embryonic mice. Neuroscience 46:407M18
Zomzely-Neurath CE (1982) Nervous-system-specific proteins: 14- 3-2 protein, antigen alpha and neuron-specific enolase. Scand J Immunol 15 [Suppl 9]: 1-40
Copyright © 2022 FDOKUMEN




















