
Forelimb myology of carnivorous marsupials (Marsupialia ...
55
RESEARCH REPOSITORY This is the author’s final version of the work, as accepted for publication following peer review but without the publisher’s layout or pagination. The definitive version is available at: https://doi.org/10.1002/ar.23612 Warburton, N.M. and Marchal, C-R (2017) Forelimb myology of carnivorous marsupials (Marsupialia: Dasyuridae): Implications for the ancestral body plan of the Australidelphia. The Anatomical Record, 300 (9). pp. 1589-1608. http://researchrepository.murdoch.edu.au/id/eprint/38289/ Copyright: © 2017 Wiley Periodicals, Inc. It is posted here for your personal use. No further distribution is permitted.
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Forelimb myology of carnivorous marsupials (Marsupialia ...

RESEARCH REPOSITORY
This is the author’s final version of the work, as accepted for publication following peer review but without the publisher’s layout or pagination.
The definitive version is available at:
https://doi.org/10.1002/ar.23612
Warburton, N.M. and Marchal, C-R (2017) Forelimb myology of carnivorous marsupials (Marsupialia: Dasyuridae): Implications for the
ancestral body plan of the Australidelphia. The Anatomical Record, 300 (9). pp. 1589-1608.
http://researchrepository.murdoch.edu.au/id/eprint/38289/
Copyright: © 2017 Wiley Periodicals, Inc.
It is posted here for your personal use. No further distribution is permitted.

Forelimb myology of carnivorous marsupials (Marsupialia: Dasyuridae):
Implications for the ancestral body plan of the Australidelphia
Short title: Forelimb myology of carnivorous marsupials
Natalie M. Warburton1, 2 and Charlie-Rose Marchal 3
1School of Veterinary and Life Sciences, Murdoch University, 90 South Street, Murdoch,
Western Australia 6150
2Department of Earth and Planetary Sciences, Western Australian Museum, Kew Street,
Welshpool, Western Australia.
3L’Ecole Nationale Veterinaire de Toulouse, France
Corresponding author: Natalie Warburton email: [email protected]
This article has been accepted for publication and undergone full peer review but has not beenthrough the copyediting, typesetting, pagination and proofreading process which may lead todifferences between this version and the Version of Record. Please cite this article as an‘Accepted Article’, doi: 10.1002/ar.23612
This article is protected by copyright. All rights reserved.

2
ABSTRACT
Carnivorous marsupials of the family Dasyuridae represent a more generalized
anatomical condition of both craniodental and postcranial features in comparison to other groups
of Australidelphian marsupials. Plesiomorphic characters include polyprotodont dentition,
didactylous (rather than syndactylous) pedal morphology, the retention of clavicles and epipubic
bones, and an unossified patelloid. In light of the anatomy of the postcranial skeleton, we
hypothesized that the muscular anatomy of the Dasyuridae would likely display a range of
plesiomorphic traits. We performed gross anatomical dissection on the forelimbs of four species
of dasyurid marsupials to produce anatomical descriptions and muscle origin and insertion maps
for Dasyurus geoffroii, D. hallucatus and Phascogale tapoatafa, together with comparative notes
for Antechinus flavipes. These new descriptions were then compared with those of other
marsupials from the published literature in order to establish the principal patterns in forelimb
muscular anatomy. In nearly all aspects of anatomy, we found that the arrangement of the
muscular origins and insertions, and the relative degree of separation between muscle bellies
among dasyurids, provide a natural starting point from which the anatomies of other
Australidelphian marsupial groups can be derived.
Keywords: Appendicular muscles, marsupial, evolution, Dasyuromorphia, Muscular anatomy
Page 2 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

3
INTRODUCTION
The dasyurid marsupials (Marsupialia: Dasyuridae) comprise the principal group of
carnivorous marsupials endemic to Australia and Papua New Guinea, ranging in size from the
tiny insectivorous ningauis (Ningaui spp.) that weigh as little as 3 grams, to the Tasmanian devil
(Sarcophilus harrisii), which weighs around 9 kg (Morton et al., 1989). Many of the smaller
species resemble placental insectivores such as shrews in their body plan, while the medium to
larger sized species exhibit convergence with carnivorans such as mustelids or small felids
(Table 1). Within Dasyuridae, four tribes are recognized; tribes Dasyurini (Dasycercus,
Dasykaluta, Dasyuroides, Dasyurus, Myoictis, Neophascogale, Parantechinus, Phascolosorex,
Pseudantechinus, Sarcophilus) and Phascogalini (Antechinus, Murexia, Phascogale) are grouped
into subfamily Dasyurinae and tribes Sminthopsini (Antechinomys, Ningaui, Smithopsis) and
Planigalini (Planigale) within the subfamily Sminthopsinae (Baverstock et al., 1982; Archer,
1984; Krajewski et al., 1997; Krajewski et al., 2000; Westerman et al., 2015). Dasyuridae is
grouped together with the Myrmecobiidae (the numbat Myrmecobius fasciatus) and the now
extinct clade Thylacinidae to comprise the order Dasyuromorphia (Fig. 1).
Within the monophyletic marsupial clade Australidelphia (Szalay, 1982; Aplin and
Archer, 1987; Marshall et al., 1990; Amrine-Madsen et al., 2003; Horovitz and Sanchez-
Villagra, 2003; Meredith et al., 2008), the Dasyuridae represent a more generalized anatomical
condition of both craniodental and postcranial features in comparison to other groups. Four pairs
of upper incisors and three pairs of lower incisors in the dental formula of dasyurids places them
on the plesiomorphic side of the polyprotodont-diprotodont dichotomy (Archer, 1976; Morton et
al., 1989). Among Australian marsupials, the polyprotodont incisor arrangement is shared with
Peramelemorphia (bandicoots and bilbies) and Notoryctemorphia (marsupial moles), and is
Page 3 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

4
distinct from the Diprotodontia (wombats, koalas, possums and kangaroos) that possess only one
procumbent pair of lower incisors. In the postcranial skeleton, dasyurids are characterised the
possession of four or five digits on the pes. In contrast, Peramelemorphia and Diprotodontia
(convergently) have a derived syndactylous condition of the pes in which the often diminutive
second and third digits are anatomically and developmentally bound together (Jones, 1949;
Szalay, 1982; Morton et al., 1989; Archer and Hand, 2006). Other plesiomorphic postcranial
traits in dasyurids include the retention of a clavicle in the pectoral girdle, which is reduced or
absent in peramelemorphians (Gordon and Hulbert, 1989; Warburton et al., 2014) and
Notoryctes (Stirling, 1891; Warburton, 2006), the presence of epipubic bones (reduced or lost in
Notoryctes (Stirling, 1891; Warburton, 2006) and Thylacinus (Dixon, 1989)), and the possession
of a cartilaginous patelloid, which is distinct to the ossified patella found in peramelemorphians
(Jones, 1968; Reese et al., 2001; Warburton et al., 2015) and Notoryctes (Stirling, 1891;
Warburton, 2006). On the basis of these skeletal plesiomorphies, we hypothesize that the
muscular anatomy of dasyurids is likely to represent a primitive arrangement among Australian
marsupials.
Detailed accounts of the muscular anatomy of dasyurids are limited to a small number of
nineteenth and early twentieth century publications (MacAlister, 1870; Macalister, 1872;
Cunningham, 1878b, 1878a, 1881, 1882; MacCormick, 1886a, 1886b; Carlsson, 1926; Jones,
1949), and a few early comparative studies of marsupial myology (Young, 1879; Elftman, 1929;
Shrivastava, 1962). Here we revisit the forelimb myology of dasyurid marsupials with new
descriptions of muscle anatomy in four dasyurid species, which synonymize previous
descriptions with current anatomical nomenclature and provide a platform for a comparison of
forelimb structure among dasyurids and among Australidelphian marsupials more broadly. The
Page 4 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

5
four species dissected represent a diversity of body sizes, locomotor traits, and habitats, and
together with those published in the literature provide a solid framework for understanding the
comparative muscular anatomy of the dasyurid forelimb.
MATERIALS and METHODS
This study provides descriptions and observations of the forelimb muscle anatomy of four
dasyurid marsupials: the Chuditch, or western quoll (Dasyurus geoffroii Gould, 1841; n=3),
northern quoll (Dasyurus hallucatus Gould, 1842; n=3), brush-tailed phascogale (Phascogale
tapoatafa Meyer, 1793; n=2) and mardo (Antechinus flavipes Waterhouse, 1838; n=1). Cadaver
specimens for dissection of the two Dasyurus species and one Phascogale were obtained from
Western Australian Department of Parks and Wildlife (DPaW), the second Phascogale and the
Antechinus specimens were collected as opportunistic finds of specimens – the Phascogale had
been found dead post breeding (male die-off) and the Antechinus found drowned in a water
vessel on a domestic property (DPaW licence SF 6788 with approval from the Murdoch
University Animals Ethics Committee 2012-2015). While Dasyurus spp. are known to have
some degree of sexual dimorphism in body size (Table 1), and sexual dimorphism has been
demonstrated to alter the relative development of muscles in some marsupials (Warburton et al.,
2013), we believe it is unlikely to have a significant effect of muscle origin and insertion sites.
Further, given our relatively small samples sizes dictated by the opportunistic collection of
specimens, we did not consider sexual dimorphism to be testable within this study.
Page 5 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

6
All specimens were frozen upon initial collection and were defrosted prior to preparation.
Specimens were skinned, eviscerated and embalmed in a 10% formalin 4% glycerol solution for
one week, before being stored in 70% ethanol. Dissections were made of the right and left
forelimbs. Muscles were dissected in major functional groups: extrinsic muscles of the pectoral
limb, intrinsic muscles of the shoulder and brachium, muscles of the antebrachium, and intrinsic
muscles of the manus, using standard dissection techniques and equipment. Following dissection,
the bones were cleaned with a degreasing agent and used as the basis for illustrations. The bones
have been retained for future work within the School of Veterinary and Life Sciences, Anatomy
Department at Murdoch University, Western Australia.
As highlighted in Diogo et al (2016), marsupials (typified by Didelphis in that study)
have a more plesiomorphic musculature in comparison to many eutherian mammals. This
plesiomorphic musculature in marsupials is more complex, having a greater number of muscles.
In her descriptive studies of musculature of marsupials, Warburton (Warburton, 2006; Harvey
and Warburton, 2010; Warburton et al., 2011; Warburton et al., 2012; Warburton et al., 2014;
Warburton et al., 2015) has attempted, where possible, to standardize the marsupial muscle
nomenclature with that of the Nomina Anatomica Veterinaria (Schaller, 2007). The greater
complexity of muscles in marsupials and the highly derived condition of cursorial domestic
eutherians, however, has made this a challenging task, and one that was often fraught with
confusion. Extensive work by Diogo and colleagues on the comparative anatomy, homologies
and evolution of the muscles of the head and neck, culminating in Diogo et al. (2012), provide a
direct comparison of the nomenclature used in comparative anatomy and zoology with that of the
Nomina Anatomica Veterinaria. In this current study, we follow the nomenclature presented in
Diogo et al. (2016), which is based on extensive review of the literature on muscle development
Page 6 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

7
and anatomy, as well as dissection, to facilitate a unified nomenclature. A small number of
differences observed between our specimens and Didelphis required the use of additional muscle
nomenclature, for which we followed Warburton et al. (2014; m. pectoralis abdominis ) and
Fisher et al. (2009; mm. opponens, m. palmaris brevis).
Comparisons were made with other marsupial groups by dissection of Brush-Tailed
Possums (Trichosurus vulpecula; n=3) collected as road-killed cadavers under the same licence
as above, and Western Grey Kangaroo (Macropus fuliginosus; n=3) purchased from licenced
abattoirs. These specimens were fixed and treated following the protocols set out above. These,
together with extensive review of the anatomical literature (Table 2) and by investigation of
skeletal material held in the Western Australian Museum, enabled comparisons between
marsupial groups.
All muscle descriptions relate to D. geoffroii, unless otherwise stated. For other species,
remarks are made only where differences were noted.
RESULTS
Extrinsic muscles of the pectoral limb
M. trapezius
Three portions of the m. trapezius were identified: pars capitis, pars cervicis, pars
thoracica. In D. geoffroii (Fig. 2) pars capitis and cervicalis arose as a continuous sheet of fleshy
fibers from the entire length of the nuchal crest, and from along the supraspinous ligament from
Page 7 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

8
the occiput to the spinous process of T5. The cranial portion (frequently mentioned as a division
of the m. brachiocephalicus in eutherian groups) was thick and passed over the lateral aspect of
neck to insert along the cranial aspect of the lateral half of the clavicle, adjacent to the origin of
the fleshy cleidomastoid muscle. The cervical portion inserted with fleshy fibers along the length
of the scapular spine (Fig. 3b). The most proximal part passed over the acromion to insert by a
small aponeurotic connection onto the distal extremity of the deltoid crest, superficial to the
acromial and clavicular portions of the deltoid. The pars thoracica arose by fleshy fibers along
the spinous processes of T5-T12 and inserted by an aponeurotic sheet along the proximal one
third of the scapular spine (Fig. 3c). In D. hallucatus the thoracic part originated from T7-T13. In
P. tapoatafa the insertion of the cervical portion onto the scapular spine was by fleshy fibers
proximally and aponeurotic distally (Fig. 3e,f). In A. flavipes, the origin of the pars thoracica
extended only to T10.
M. levator claviculae (m. atlantoscapularis anticus ; m. omotransversarius; m.
atlantoacromialis; m. omotrachelien ventralis)
In all species, m. levator claviculae arose as thin muscular sheet from the transverse
process of the atlas (C1) and inserted onto the distal two-fifths of scapula spine and cranial
aspect of the acromion (D. geoffroii Fig. 2 inset; Fig. 3c; P. tapoatafa Fig. 3f; 4).
M. atlantoscapularis posticus (m. atlantoscapularis; m. omotrachelien dorsalis; m. levator
scapulae posticus; m. rhomboideus profundus)
In D. hallucatus and P. tapoatafa a distinct sheet of fleshy fibers representing m.
atlantoscapularis posticus was found, originating from the wing of the atlas and inserting onto
the proximal part of the scapular spine (P. tapoatafa Fig. 3f, 4). This muscle was similarly
present in A. flavipes. In D. geoffroii the m. atlantoscapularis posticus was represented by an
Page 8 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

9
extremely thin layer of aponeurotic tissue overlying the m. supraspinatus near the vertebral
border of the scapula.
M. latissimus dorsi
In Dasyurus spp., m. latissimus dorsi arose as a broad but thin muscular sheet from the
dorsal spines and lumbar fascia between the T4 and the last rib (13) (D. geoffroii Fig. 2). The
muscle inserted via a strong tendon onto the rugose teres tubercle on the medial humeral shaft,
immediately distal to, and fused with, the insertion of the teres major (D. geoffroii Figs. 5, 6a). In
P. tapoatafa the origin was from the spinous processes of T4-T11, and the lumbar fascia to the
level of L4 (Fig. 4) and the insertion was slightly cranial to that of the teres major (Fig. 6e). In A.
flavipes the origin extended further caudally, from spinous processes T5-T11 and the lumbar
fascia to S1. In A. flavipes, the sheet became divisible into two portions near its humeral
insertion, as observed in Dasycercus (Jones, 1949).
M. rhomboideus
M. rhomboideus pars capitis and pars cervicis formed a thin sheet of long fleshy fibers
from the occiput and medial nuchal crest, and from along spinous process C2-C7, which inserted
onto the cranial angle of the scapula (D. geoffroii Fig. 2; Fig. 3a,c). Pars thoracis was a much
thicker muscle portion, comprised of short fibers passing from the proximal thoracic spinous
processes to the vertebral border of scapula. In D. geoffroii the origin was between T1 and T3.
In P. tapoatafa the origin was between T1 and T5 and the insertion more localized to the cranial
angle of the scapula (Fig. 3d,f). In A. flavipes the origin extended to T6.
M. serratus anterior (m. serratus ventralis)
Page 9 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

10
In Dasyurus spp., m. serratus anterior pars cervicis (‘levator scapulae’) arose from the
transverse processes of the cervical vertebrae C2-C7 (D. geoffroii Fig. 2) and had a narrow
fleshy insertion onto the deep surface of the cranial angle and vertebral border of scapula (facies
serrata) (D. geoffroii Fig. 3a). Pars thoracis arose by fleshy fibers from the lateral aspect of the
first eight ribs and inserted onto the superficial aspect of the caudal angle of scapula, opposite the
seventh rib. In P. tapoatafa the cervical portion was relatively feebly developed (Fig. 3d).
M. pectoralis major (m. pectoralis superficialis)
M. pectoralis major comprised a short m. pectoralis transversus, originating from the
manubrium and medial half of the clavicle, and longer m. pectoralis descendens arising from the
ventral midline of the second to the terminal sternebrae. The insertions of the transverse and
descending portions were contiguous along the length of the pectoral ridge of humerus (P.
tapoatafa Figs. 4, 6g,h; D. geoffroii Fig. 6d). This was the same for all species dissected.
M. pectoralis minor (m. pectoralis profundus)
The m. pectoralis minor arose by fleshy fibers from the ventral midline of the second to
sixth sternebrae, deep to the pectoralis major. It inserted onto the crest of the greater trochanter
(D. geofroii Fig. 6d). The pectoralis minor, being considerably smaller in both extent and
development, was completely hidden by the pectoralis major. This was similar for the species
dissected.
M. pectoralis abdominis (m. pectoralis quartus)
In Dasyurus spp. the “m. pectoralis abdominis” was a small strap muscle that arose from
the distal sternum, on the deep, caudal surface of the m. pectoralis major pars descendens, and
Page 10 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

11
inserted onto the distal half of the pectoral ridge deep to the pectoralis major. In P. tapoatafa, the
muscle was only a thin ribbon of fibers from caudal rib, which became inseparable from the
superficial pectoralis major at its insertion.
M. subclavius
M. subclavius arose by fleshy fibers from lateral edge of the manubrium, adjacent to the
articulation of the first rib, and passed deep to the clavicle to insert onto the cranial aspect of the
capsule of the acromioclavicular joint and fascia of the m. supraspinatus.
Shoulder
Mm. deltoideus
In D. geoffroii, mm. deltoideus consisted of three distinct muscles: deltoideus clavicularis
(m. cleidobrachialis), deltoideus acromialis and deltoideus scapularis (Fig. 2 inset; Fig. 3d,c) all
inserting to the deltoid crest of the humerus (D. geoffroii Fig. 6d; P. tapoatafa Fig. 6g,h). M.
deltoideus clavicularis arose from the lateral half of the clavicle and inserted onto the distal end
of the deltopectoral crest, immediately proximal to the insertion of the transverse portion of the
superficial pectoral and adjacent to the insertion of the cervical trapezius. M. deltoideus
acromialis arose from the acromion (D. geoffroii Fig. 3d) and inserted via aponeurotic fibers
along the length of the deltoid crest of humerus. M. deltoideus scapularis arose by fleshy fibers
from the broad hamate process of the acromion and along the distal two-thirds of the posterior
border of the scapular spine. The insertion of the scapular portion was immediately distal to that
of the acromial part. The arrangement of three distinct muscle bellies was the same throughout
all specimens dissected (P. tapoatafa Fig. 3f, 4). The scapular and clavicular portions appeared
to be much more strongly developed in D. hallucatus than in D. geoffroii.
Page 11 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

12
M. supraspinatus
M. supraspinatus arose by fleshy fibers from the supraspinous fossa and the cranial edge
of the scapular spine (D. geoffroii Fig. 2 inset; Fig. 3c; P. tapoatafa Fig. 3f, 4). It inserted cranio-
medially onto the greater tuberosity of the humerus by both fleshy and aponeurotic fibers in
Dasyurus spp., and the tendon was tightly adherent to the capsule of the shoulder joint (Fig. 6d).
In P. tapoatafa (Fig. 6g,h) the insertion was via a strong tendon; no fleshy fibers were as
observed.
M. infraspinatus
M. infraspinatus arose from the infraspinous fossa and the caudal aspect of the scapular
spine by fleshy and aponeurotic fibers (D. geoffroii Fig. 2 inset; Fig. 3c; P. tapoatafa Fig. 3f, 4).
A strong tendon inserted onto the lateral aspect of the greater tuberosity of humerus (D. geoffroii
Fig. 6b,c; P. tapoatafa Fig. 6g,h). This arrangement was consistent in all species dissected.
M. teres minor
M. teres minor was a small, distinct muscle that arose from the infraglenoid rugosity and
ridge along the distal one-fifth of the caudal border of the scapula (D. geoffroii Fig. 3b; P.
tapoatafa Fig. 3e) and inserted immediately below the base of the greater trochanter (D. geoffroii
Fig. 6d P. tapoatafa Fig. 6g). M. teres minor appeared to be relatively smaller in D. hallucatus
when compared to D. geoffroii.
M. teres major
M. teres major arose from the thickened caudal angle of the scapula (D. geoffroii Fig. 2;
Fig. 3a-c, 5; P. tapoatafa Fig. 3d-f, 4) and inserted onto the proximo-lateral aspect of humeral
Page 12 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

13
shaft. In Dasyurus spp. the insertion was fused with the inserting tendon of the m. latissimus
dorsi and the site of insertion was marked by a rugose teres tubercle on the humeral shaft (D.
geoffroii Fig. 6a). M. teres major appeared to be relatively larger in P. tapoatafa and A. flavipes.
Notably, in P. tapoatafa, m. teres major inserted over a larger area, and more distally than m.
latissimus dorsi, such that the muscle bellies overlapped somewhat at their humeral insertion
(Fig. 6e).
M. subscapularis
M. subscapularis was a multipennate muscle from the subscapular fossa, which covered
almost the entire medial surface of the scapula, as well as the supraspinatus fascia in the glenoid
notch (D. geoffroii Fig. 3a,b, 5; P. tapoatafa Fig. 3d,e). The insertion was by a thick tendon onto
the lesser humeral tuberosity (D. geoffroii Fig. 6a,d; P. tapoatafa Fig. 6e,f,h). This arrangement
was consistent in all species dissected.
Brachium
M. coracobrachialis
M. coracobrachialis was a short, flat muscle belly that arose via a thin tendon from the
coracoid process of scapula, medial to m. biceps brachii (D. geoffroii Fig. 3a,c, 5; P. tapoatafa
Fig. 3d,f), and travelled superficial to the tendon of the m. subscapularis to insert onto the
proximal medial humeral neck, immediately proximal to the insertion of the m. teres major (D.
geoffroii Fig. 6a; P. tapoatafa Fig. 6e). In D. hallucatus it was not possible to separate the tendon
of origin of the coracobrachialis from the coracoid tendon of the biceps brachii.
M. biceps brachii
Page 13 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

14
M. biceps brachii comprised two incompletely separable fusiform portions. The gleno-
ulnar head arose from the supraglenoid tubercle of the scapula (D. geoffroii Fig. 2, 5; P.
tapoatafa Fig. 3f, 4), passed through the shallow bicipital groove, and inserted onto the proximal
cranial aspect of the ulna, immediately distal to the insertion of the m. brachialis. The coraco-
radial head arose from the coracoid process of the scapula (D. geoffroii Fig. 3a,c; P. tapoatafa
Fig. 3d,f). This tendon did not enter the bicipital groove, but rather descended along the cranial
facet of the lesser tuberosity. The fusiform belly gave rise to a short tendon that inserted onto the
bicipital tubercle of the radius (Fig. 7). In D. geoffroii it was relatively easy to separate the two
bellies, while in D. hallucatus the distal ends of the muscle bellies were incompletely separable
(D. hallucatus Fig. 8).
M. brachialis
M. brachialis originated from the proximolateral humeral shaft, descended obliquely
along the lateral aspect of the humerus, between the deltoid ridge and the lateral supracondylar
ridge (D. geoffroii Figs. 2, 6a-d; P. tapoatafa Figs. 4, 6e-h). A short bundle of fibers arose from
the cranio-distal humerus to become blended with the much longer preceding portion. The
insertion was mostly by fleshy fibers onto cranial aspect of the ulna, immediately distal to the
coronoid process, and proximal to the insertion of the ulnar insertion of the m. biceps brachii
(Fig. 7). In D. hallucatus, the bony origin was much more extensive, from along much of the
length of the lateral aspect of the humeral shaft (effectively making the two separate origins in D.
geoffroii continuous). In P. tapoatafa and A. flavipes, the distal origin of the m. brachialis
surrounded the patent supracondylar fossa.
M. triceps brachii
Page 14 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

15
M. triceps brachii caput longum arose along one-fourth of caudal border of the scapula
adjacent to the glenoid, deep to, and slightly longer than, the origin of the m. teres minor (D.
geoffroii Figs. 2, 3b, 5; P. tapoatafa Figs. 3d,e, 4). The m. triceps brachii caput laterale arose
from the lateral aspect of the humeral neck (D. geoffroii Fig. 6b,c; P. tapoatafa Fig. 6f). The m.
triceps brachii caput mediale took its origin from the medial aspect of the humeral neck and
proximal two-thirds of humeral shaft (D. geoffroii Fig. 6a,b; P. tapoatafa Fig. 6e,f,h). The medial
and lateral heads were closely situated at their proximal origin and incompletely separable
throughout. The three muscular bellies converge to insert onto the olecranon process of the ulna
(D. geoffroii Fig. 7; P. tapoatafa Fig. 7).
M. dorsoepitrochlearis (m. tensor fascia antebrachii)
M. dorsoepitrochlearis arose by fleshy fibers from caudal margin of the m. latissimus
dorsi, close to its insertion on the humerus and passed over the superficial, medial aspect of the
m. triceps brachii. The insertion was by aponeurotic fibers into the medial fascia of the elbow
and olecranon (D. geoffroii Fig. 5). This muscle was relatively larger and thicker in D. geoffroii
than in D. hallucatus.
M. epitrochleoanconeus (m. anconeus internus; m. triceps brachii caput mediale
accessorium)
M. epitrochleoanconeus was a small fleshy muscle mass, passing from the distolateral
humerus, above the medial epicondyle (D. geoffroii Fig. 6b; P. tapoatafa Fig. 6f), to insert onto
the olecranon.
M. anconeus lateralis (m. anconeus externus; m. anconeus)
Page 15 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

16
M. anconeus lateralis arose from fascia over the lateral epicondyle and posterior aspect of
the humerus, surrounding the olecranon fossa (D. geoffroii Fig. 6b; P. tapoatafa Fig. 6f), and
inserted onto the lateral aspect of the olecranon. The muscle was much larger than the m.
epitrochleoanconeus. This arrangement was consistent in all of the species dissected.
Antebrachium – ventral division
M. pronator teres
M. pronator teres arose by fleshy and aponeurotic fibers from the cranial aspect of the
medial epicondyle of the humerus, proximal to the flexor carpi radialis and deep to the humeral
head of the flexor digitorum profundus (D. geoffroii Fig. 6a; P. tapoatafa Fig. 6e; D. hallucatus
Fig. 8). It inserted by fleshy fibers onto the proximal half of the craniomedial aspect of the radius
(D. geoffroii Fig. 7b). This arrangement was consistent in all of the species dissected.
M. flexor carpi radialis
M. flexor carpi radialis originated from the medial epicondyle, completely under cover of
humeral portion of the m. flexor digitorum profundus (D. geoffroii Fig. 6a; P. tapoatafa Fig. 6e;
D. hallucatus Fig. 8). The thin muscle belly was incompletely separable into two portions. Two
tendons arose in the distal half of the antebrachium. The dorsal portion gave a thin tendon to the
carpal fascia surrounding the proximoradial (scapholunate) bone of the carpus, and the ventral
belly gave a strong tendon that inserted onto the base of the second and third metacarpals (D.
geoffroii Fig. 9). In P. tapoatafa the insertion was only traced to metacarpal II.
M. flexor carpi ulnaris
Page 16 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

17
M. flexor carpi ulnaris had a broad, flat muscle belly that arose from the caudal aspect of
medial epicondyle and medial aspect of olecranon (D. geoffroii Fig. 6a,b P. tapoatafa Figs. 5, 6e;
D. hallucatus Fig. 8). The belly passed along the medial aspect of ulna to insert via a thick
tendon along the distal third of the antebrachium onto the pisiform bone (accessory carpal) and
via an extension of the tendon to the base of metacarpal V (D. geoffroii Fig. 9). In D. hallucatus
and P. tapoatafa, this insertion could be traced to the pisiform bone only.
M. palmaris longus
M. palmaris longus arose from the medial epicondyle of the humerus, between m. flexor
digitorum profundus and m. flexor carpi ulnaris (D. geoffroii Fig. 6a; P. tapoatafa Fig. 6e; D.
hallucatus Fig. 8). The long, thin belly passed along the ventral edge of the deep flexor to insert
into the deep fascia of ventral carpus over the flexor tendons to digits II-V (P. tapoatafa Fig. 5).
This arrangement was consistent in all the species dissected.
M. flexor digitorum superficialis (m. flexor sublimis, m. flexor perforatus)
M. flexor digitorum superficialis was a tiny slip of muscle from the distal superficial
aponeurosis of the m. flexor digitorum profundus (P. tapoatafa Fig. 5). The small tendons passed
through a superficial groove on deep flexor tendon and across the palm to insert onto the base of
the proximal phalanx of digits II-V (D. geoffroii Fig. 9). M. flexor digitorum superficialis
appeared relatively larger in A. flavipes than in the other species dissected.
M. flexor digitorum profundus (historically considered to include m. flexor longus pollicis, though see Diogo et al. 2016)
M. flexor digitorum profundus was a massive muscle and divisible into three thick
bellies: caput humerale, pars radiale and caput ulnare. The large, superficial belly arose from the
Page 17 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

18
medial flexor crest of the humerus proximal to the epicondyle (D. geoffroii Fig. 6a; P. tapoatafa
Figa. 5, 6e). The origin was so broad as to cover all of the muscles that arose from the medial
epicondyle. The ulnar belly, from the olecranon, was a more elongate muscle mass (D.
hallucatus Fig. 8). A short, deep belly arose in the distal half of the antebrachium from the
medial aspect of the radial shaft. The large tendons from each portion merged in the distal
antebrachium to pass through the carpal canal under the flexor retinaculum. In the palm, five
thick tendons were traced to the distal phalanx of each digit (D. geoffroii Fig. 9).
M. pronator quadratus
The fleshy fibers of the m. pronator quadratus passed between the medial aspects of distal
half of the radius and ulna, in the interosseous space (D. geoffroii Fig. 7b; P. tapoatafa Fig. 7d).
This arrangement was consistent in all of the species dissected.
Antebrachium – dorsal division
M. brachioradialis (M. supinator longus)
M. brachioradialis was relatively small muscle that arose from the most proximal tip of
the lateral supracondylar ridge, in close connection with the m. extensor carpi radialis (D.
geoffroii Figs. 2, 6c-d; P. tapoatafa Figs. 4, 6g,h). The thin muscle gave rise to a tiny tendon,
which passed on the medial aspect of the m. extensor carpi radialis, and deep to the tendon of the
m. abductor pollicis longus, to insert onto the radial aspect of the proximoradial carpal bone (D.
geoffroii Fig. 9). This arrangement was also present in the other species.
M. extensor carpi radialis
Page 18 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

19
M. extensor carpi radialis arose as a large single muscle belly from the lateral
supracondylar ridge (D. geoffroii Figs. 2, 6d; P. tapoatafa Fig. 4, 5, 6g,h) and inserted via an
obvious tendon onto the base of metacarpals II and III (D. geoffroii Fig. 9).
M. extensor digitorum communis
M. extensor digitorum communis consisted of many small bellies which arose from the
lateral humeral epicondyle via tendinous fibers, and also from the proximal radius via fleshy
fibers (D. geoffroii Figs. 2, 6c,d, 7; P. tapoatafa Figs. 4, 6g, 7). The tendons passed together deep
to the dorsal annular ligament of wrist before giving paired (lateral and medial) insertions to the
proximal phalanx of digits II-V (D. geoffroii Fig. 9).
M. extensor digiti minimi (m. extensor digitorum lateralis)
M. extensor digiti minimi arose from the lateral epicondyle of humerus adjacent to the
common digital extensor (D. geoffroii Figs. 2, 6c,d; P. tapoatafa Fig. 4, g). The muscle belly
gave rise to short tendon, which divided to insert onto the dorsal aspect proximal phalanx of
digits four and five (D. geoffroii Fig. 9; D. hallucatus Fig. 10).
M. extensor digitorum profundus (m. extensor secundi internodii pollicis + m. ext. indicis, medii digiti et quarti of McCormick)
M. extensor digitorum profundus arose as a single, thin belly from the olecranon (D.
geoffroii Fig. 7a). The muscle passed on the deep aspect of the lateral extensor and gave rise to
two tendons. The first tendon inserted onto fascia of the proximal phalanx of digit I; the second
tendon split into three, which ran to digits II, III and IV (D. geoffroii Fig. 9; D. hallucatus Fig.
10). In P. tapoatafa the origin was much longer, from along the proximal third of the ulna (Fig.
7c).
Page 19 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

20
M. extensor carpi ulnaris (m. ulnaris lateralis)
M. extensor carpi ulnaris arose by fleshy fibers from the lateral humeral epicondyle (D.
geoffroii Fig. 6c,d; P. tapoatafa Fig. 6g). The insertion was to the lateral carpal bone (hamate),
the adjoining intercarpal ligaments, the capsule of the lateral carpometacarpal joint and the base
of metacarpal V (D. geoffroii Figs. 2,9). In P. tapoatafa (Fig. 4) the insertion was restricted to
the carpal bones.
M. supinator (supinator brevis)
M. supinator was a very small muscle from the lateral humeral epicondyle, originating
deep to m. extensor carpi radialis (D. geoffroii Fig. 6c,d; P. tapoatafa Fig. 6g). The fleshy fibers
passed obliquely to insert onto the proximal one-quarter of the cranial aspect of the radius (D.
geoffroii Fig. 7a,b; P. tapoatafa Fig. 7c,d).
M. abductor pollicis longus (M. abductor digiti I longus; M. extensor ossis metacarpi
pollicis, M. extensor carpi obliquus)
M. abductor pollicis longus arose along the entire length of the cranial border of radius,
interosseus membrane and ulnar shaft, deep to m. extensor carpi radialis and the common digital
extensor (D. geoffroii Figs. 2, 7a; P. tapoatafa Fig. 4, 7c; D. hallucatus Fig. 10). The thick,
flattened tendon of m. abductor pollicis longus emerged from between the two muscles, and
followed an oblique course to insert onto the base of metacarpal I (D. geoffroii Fig. 9). The
oblique tendon of m. abductor pollicis longus passed superficial to the tendons of m. extensor
carpi radialis (D. hallucatus Fig. 10).
Manus
Page 20 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

21
The following descriptions of the intrinsic muscles of the hand pertain to D. geoffroii and
D. hallucatus; the origins and insertions could not be traced with confidence in the smaller P.
tapoatafa and A. flavipes.
Mm. lumbricales
Four lumbricales arose from between the clefts of the thick mesial tendon of the deep
digital flexor and inserted onto the extensor expansions of the deep aponeurosis of the digits (II-
V).
Mm. contrahentes (mm. adductores digitorum breves) (D. hallucatus Fig. 11)
The short adductor of the thumb, m. adductor digiti I (pollicis) arose from the flexor
retinaculum and deep palmar fascia. The muscle belly passed obliquely to insert onto the fascia
over the metacarpophalangeal joint and base of the proximal phalanx of the first digit. The
contrahentes of digits II (adductor indicis) and IV (adductor annularis) arose from the palmar
carpal ligament (superficial to the flexor breves) and inserted to the lateral base of proximal
phalanx II and medial aspect of the base of proximal phalanx IV respectively. The contrahens of
digit V (m. adductor digiti minimi) arose from the ulnar side of the flexor retinaculum. The
muscle lay in an oblique orientation and we traced the insertion only as far as the fascia over the
head of the fifth metacarpal and the fifth metacarpophalangeal joint.
Mm. flexores breves profundi
The short digital flexors were represented by ten small muscular bellies, arranged in
pairs, which arose from the fascia over the carpo-metacarpal joints, and passed to each of the five
digits (D. hallucatus Fig. 11). The paired lateral and medial muscles for each of the five digits
Page 21 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

22
inserted by fleshy fibers onto their respective side of base of the proximal phalanx and
surrounding fascia. The most radial of the flexores breves, often referred to as m. flexor pollicis
brevis, arose from the tough fascia over the trapezoid, at the base of the second metacarpal and
inserted onto the radial aspect of the base of proximal phalanx of digit I. The most ulnar of the
flexores breves, the short flexor of the digit V, arose from the tough fascia at base of metacarpal
III and inserted onto the base of the proximal phalanx of digit V, immediately distal to the
insertion of m. flexor carpi ulnaris.
M. abductor digiti I (pollicis) brevis
The short abductor of the pollex was a small muscle, which arose from the radial distal
carpal bone (trapezium) and inserted onto the fascia on the radial side of the
metacarpophalangeal joint of digit I (D. geoffroii Fig. 9; D. hallucatus Fig. 11).
M. opponens pollicis
The m. opponens pollicis was a tiny fusiform muscle belly that originated from the radial
aspect of the thenar eminence, passing from the trapezoid to insert onto the fascia surrounding
the radial aspect of the proximal interphalangeal joint of the first digit (D. hallucatus Fig. 11).
M. abductor digiti minimi
The abductor of the fifth digit arose as fleshy fibers from the fascia between the pisiform
bone and hamate and inserted directly onto the fascia of the metacarpophalangeal joint of digit V
(D. hallucatus Fig. 11).
M. palmaris brevis
Page 22 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

23
The m. palmaris brevis was a fleshy muscle originating from the hypothenar eminence
over the pisiform bone. It inserted by a short tendon onto the ulnar side proximal phalanx of the
fifth digit (D. hallucatus Fig. 11).
M. intermetacarpales
Four intermetacarpales were observed, lying between adjacent metacarpals (D. hallucatus
Fig. 10). They had a bipennate arrangement, arising from the adjacent sides of the proximal ends
of the metacarpal bones. The m. intermetacarpales of the first and second intermetacarpal space
inserted to radial side of the proximal phalanges of the second and third digit respectively, in the
region of the insertion of the flexores breves and adjacent dorsal extensor fascia. The third and
fourth intermetacarpales similarly arise from the adjacent bases of the metacarpals and inserted
onto the ulnar aspect of the respective digits.
Discussion
Variation within Dasyuridae
The dissections and review of the literature performed in this study revealed a highly
conservative pattern of muscular anatomy within the Dasyuridae. The muscular anatomy of the
two species of Dasyurus dissected here corresponded very closely with that of D. viverrinus
described by MacCormick (1886a). The anatomical variation between species, with respect to
the origins and insertions of muscles within the forelimb was very slight, particularly in light of
intraspecific variation observed here and also discussed in the literature (MacAlister, 1870;
Page 23 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

24
Macalister, 1872; Fisher et al., 2009). The principle source of variation between species was the
relative surface area of the fleshy muscle origins; the general pattern being that muscle origins
covered a relatively larger area in species of smaller body size. For instance, the origin of the m.
brachialis in P. tapoatafa covered a relatively larger area on the lateral aspect of the humerus
than the same muscle in D. hallucatus, which in turn had relatively greater areas of origin than in
D. geoffroii. This pattern was even more pronounced when A. flavipes was considered, and
especially so for the origin and strengthened insertion of m. latissimus dorsi (also in Dasycercus;
Jones, 1949) and the extended thoracic origin of m. rhomboideus. Conversely, S. harrisii
reportedly has relatively small area of origin of m. trapezius pars thoracica and insertion of m.
trapezius pars capitis to the lateral one-quarter to one-third (rather than half) of the clavicle
(MacAlister, 1870; Macalister, 1872). The larger muscle attachments in P. tapoatafa and A.
flavipes in comparison to Dasyurus spp. likely reflect differences in limb posture with
corresponding differences in size and behaviour. Biewener (1989) demonstrated that while small
mammals run with more flexed limbs and crouched postures that favour manoeuvrability and
accelerative capability, larger mammals utilize more upright postures in which the limbs are
more closed aligned with ground forces in order to reduce bone stresses and muscle force
requirements. Antechinus flavipes, as the smallest animal within our sample, indeed displays a
more crouched posture with highly flexed limbs, which would require relatively greater muscle
force to move, and thus larger muscles relative to its body size.
The relatively larger attachments of muscles including the medial head of the m. triceps
brachii, m. brachialis, m. flexor digitorum profundus and m. abductor longus pollicis in P.
tapoatafa (in comparison to Dasyurus spp.) may also be correlated with adaptation for arboreal
behaviours in this species. As well as having a relatively more flexed limb posture, which in
Page 24 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

25
arboreal animals brings the centre of gravity closer to the supporting surface (tree trunk or
branch) and improves stability in the complex three-dimensional arboreal habitus (Cartmill,
1985), P. tapoatafa also has a more sprawling, abducted limb posture during climbing. The
relatively large size of the m. teres major, an adductor as well as extensor of the shoulder, also
represents an adaption for improved grasping during vertical climbing as has been reported in
other climbing mammals (Taylor, 1974; Warburton et al., 2011).
The myological descriptions of Sarcophilus harrisii (MacAlister, 1870; Macalister, 1872)
suggest a number of subtle differences relating to the extent of muscle attachments in this species
including the m. latissimus dorsi, the insertion of the m. teres major (though this was variable
between species in our observations), extension of the m. subclavius to dorsal aspect of scapula,
indistinct m. teres minor, incomplete separation of the acromial and clavicular heads of the
deltoid, and larger lateral head of the m. triceps brachii. Without quantitative data on muscle size
and architecture, functional comparisons are difficult and will await future studies.
Primitive retentions in Dasyuridae and the patterns of myology in Australidelphian marsupials
The anatomy of the m. trapezius has been identified as one of the key differences
between marsupials and placental mammals, and in particular the relationship between the
trapezius and the deltoid muscle group (Shrivastava, 1962). The trapezius of dasyurids forms a
broad, continuous sheet of muscle from along the nuchal crest and the mid-dorsal line of the
cervical and thoracic regions that wraps around the lateral aspect of the neck to have a substantial
insertion along the clavicle, the length of the scapular spine and the deltoid crest of the humerus.
This arrangement appears likely to represent the plesiomorphic condition among
Australidelphian marsupials. The broad continuous origin from the nuchal crest/occipital region,
Page 25 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

26
cervical and thoracic spine is consistent with that of other marsupial groups including didelphids
(Jenkins and Weijs, 1979), peramelids (Parsons, 1903; Filan, 1990; Warburton et al., 2014), and
diprotodonts (Young, 1882; Sonntag, 1922; Boardman, 1941; Barbour, 1963). The marsupial
moles (Notoryctes spp.) are unusual in the distinct separation of the spinotrapezius, but retains
the continuous origin of the cranial and cervical portions (Warburton, 2006). Among marsupials
generally, the cranial portion of the trapezius passing around the neck is thick and fleshy in
comparison to the much thinner thoracic portion, and has a robust insertion to the clavicle. There
is some degree of fusion of these anterior fibers of the trapezius with the clavicular deltoid in
Phascolarctos, Pseudocheirus, Phalanger (Sonntag, 1922), and more substantially in Notoryctes
(Warburton, 2006) and Phascolomys (Sonntag, 1922). In dasyuromorphians, however, while the
insertion of the trapezius was very closely arranged with the insertion of the deltoids, blending of
fibers was not apparent. In the aclaviculate peramelids, the very thick anterior trapezius
continues to the distal deltoid crest forming a strong cephalohumeral muscle (Warburton et al.,
2014). This highly derived condition presents a similar arrangement to that observed in placental
mammals where the clavicle is reduced or absent, particularly among the Carnivora (Wood,
1870; Davison, 1944; Miller, 2012).
The m. omotransversarius of veterinary nomenclature, derived from the levator scapulae
(Arlamowska-Palider and Zablocki, 1972; Diogo et al., 2016), presents a nomenclatorial
difficulty. The plesiomorphic condition for both marsupials and eutherian mammals is the
presence of two distinct muscles, atlantoscapularis anticus (considered more appropriately
named levator claviculae by Diogo et al. 2016) and atlantoscapularis posticus. Thus the
veterinary m. omotransversarius of domestic species represents only the atlantoscapularis
anticus, in the derived condition in which the atlantoscapularis posticus is absent. The presence
Page 26 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

27
of both these muscles in all dasyurids considered here, consistent with previous findings in
Dasyurus (MacCormick, 1886a; "acromio-trachelien") and Phascogale (Cunningham, 1882),
reflects the plesiomorphic condition for dasyuromorphians; note that for S. harrisii, while only
the ventral portion was observed in MacAlister (1870; "omo-atlantic"), both portions were
identified in (Macalister, 1872; "trachelo-acromiales muscles"). Both muscles are present in
Didelphis (Coues, 1871; Jenkins and Weijs, 1979; atlanto-acromialis and atlanto-scapularis) and
Caluromys (Argot, 2001; omotransversarius and levator scapulae dorsalis), as well as many
eutherian mammals and thus the presence of two muscles has been interpreted as the
plesiomorphic arrangement in all mammals (Arlamowska-Palider and Zablocki, 1972; Diogo and
Abdala, 2010; Diogo et al., 2016). In Diprotodontian marsupials (Sonntag, 1922; Boardman,
1941; "omo-cleido-transversarius"; Harvey and Warburton, 2010; for dorsal portion see "serratus
ventrals cervicis: atlantoscapularis"; Warburton et al., 2011), the two muscles often have a
broadly continuous insertion along the length of the scapular spine. In Phascolomys and
Phascolarctos there appears to be a greater separation between the two muscles, with the levator
claviculae apparently reduced and incorporated into the m. serratus ventralis as “levator anguli
scapulae” (Sonntag, 1922) or not reported at all (Macalister, 1865, 1870; Young, 1882).
Marsupial taxa in which the atlantoscapularis posticus has become reduced or absent tend to be
those with a more strongly adducted forelimb posture and emphasis on limb movements in the
parasagittal plane for terrestrial locomotion and/or digging (Chaeropus (Parsons, 1903), Isoodon
(Filan, 1990; Warburton et al., 2014), Notoryctes (Warburton, 2006), and thus the absence of this
muscle should be considered the derived condition amongst marsupials. A similar conclusion has
been made for eutherian mammals (Arlamowska-Palider and Zablocki, 1972). The presence of
the two muscles (atlantoscapularis anterior and posterior, or omotransversarius and rhomboideus
Page 27 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

28
profundus) is a common condition for Mephitidae and Mustelidae, lineages with an extremely
diverse range of locomotory habits (Ercoli et al., 2015), contrary to some other carnivorans
(ursids; Davis, 1964; canids; Miller, 2012) . Together, the m. omotransversarius and m.
rhomboideus profundus are efficient to advance and rotate the scapula in the parasagittal plane,
and to laterally flex the neck. Canis, however, a more specialised cursor, possess only the m.
levator claviculae (m. omotransversarius) (Miller, 2012) and thus the absence or reduction of the
atlantoscapularis posticus (m. rhomboideus profundus) muscle may be linked to a reduction of
these muscles functions in animals which tend to have a more restricted suite of limb
movements. Support for this hypothesis is the apparent absence of the atlantoscapularis posticus
in Thylacinus (Cunningham, 1882 reported in MacCormick 1886a) and in cursorial placental
mammals more generally (Chauveau, 1891; Miller, 2012).
The nomenclature of the pectoral muscles in marsupials has historically been very
variable (Boardman, 1941; Horiguchi, 1981; Warburton, 2006; Diogo et al., 2009; Harvey and
Warburton, 2010; Warburton et al., 2011; Warburton et al., 2014). As far as can be ascertained
from this study, it would appear that the following generalisations might be made regarding the
plesiomorphic condition of this group. Firstly, the m. pectoralis major (superficialis) is by far the
largest component of the pectoral group arising from the length of the sternum, and having an
elongate insertion along the length of pectoral ridge of the humerus. The division between a
cranial pars transversus and caudal pars descendens can usually be made, and the pars
transversus from the manubrium often has some fibers of clavicular origin. Secondly, the m.
pectoralis minor (profundus) is generally relatively small in comparison to the major, almost
always being completely hidden by the more superficial muscle and thus is very different to the
condition in Canis (Miller, 2012) and eutherian mammals more generally. Finally, the abdominal
Page 28 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

29
portion of the pectorales “m. pectoralis abdominis (quartus)” of Australidelphian marsupials
appears to be a derivative of the superficial pectoralis major, rather than the pectoralis minor. We
found a very close association between the origin superficial and abdominal portions; only in P.
tapoatafa was the origin of the m. pectoralis abdominis positioned caudal to the margin of the m.
pectoralis descendens. The insertion of m. pectoralis abdominis is variable amongst marsupials,
though we found it underlay the proximal part of the insertion of the superficial pectoral in most
cases and suggest this may be the plesiomorphic condition for Australidelphian taxa. Where the
insertion is more closely opposed that of the m. pectoralis profundus, as in some
peramelemorphians (Warburton et al., 2014), we hypothesize that this reflects a derived
condition which corresponds to a greater emphasis on movements of limb retraction in a
parasagittal plane. Diogo et al. (2016) found the abdominal portion to be part of the pectoralis
minor in Didelphis, but notes that the derivation of the abdominal pectoral is likely to be
variable.
The intrinsic muscles of the forelimb in dasyurids provides a useful reference for
interpreting the anatomies of other Australiadelphian marsupials. The coracobrachialis was
found to be generally single (contra Cunningham, 1882; double in Phascogale) as is typical of
marsupials (Diogo et al., 2016), with the reported exceptions of two bellies in Phascolarctos
(Young, 1882) and Phalangista (Cunningham, 1882), and possible absence in in Chaeropus
(Parsons, 1903) and Notorytes (Warburton, 2006). As summarised in (Warburton et al., 2014),
two heads of the biceps are generally found, and correspond to the long and short heads of
eutherian mammals (Diogo et al., 2016). There is, however, often some degree of fusion either
proximally, as in Phascolarctos (Young, 1882), Vombatus and Phalanger (Sonntag, 1922), and
Trichosurus (Barbour, 1963), or distally, as in D. hallucatus (this study) among marsupials. The
Page 29 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

30
retention of only a single coraco-radial head of m. biceps brachii in Chaeropus (Parsons, 1903)
and Notorytes (Warburton, 2006) clearly represents a derived condition. The m. flexor carpi
radialis is generally small with insertions to metacarpals II and III, while m. flexor carpi ulnaris
is relatively much larger with the typical insertion to the pisiform occasionally extending to
metacarpal V (MacAlister, 1870; Macalister, 1872; Young, 1882; Hopwood, 1974). We found
both m. flexor digitorum superficialis and distinct m. palmaris longus to be present in dasyurids,
as it is in Didelphis (Coues, 1871) and Australian marsupials generally (Barbour, 1963;
Warburton et al., 2014); both of these muscles are typically very small and have a strong
connection to the relatively massive and complex m. flexor digitorum profundus. Diogo and
Abdala (2010) and Diogo et al., (2016), following the detailed descriptions of the flexor tendons
of the didelphid manus in Abdala et al. (2006) provide detailed commentary on the variable
nature of the palmaris longus, which in Didelphis and some eutherian mammals is represented by
two muscle bellies, and may in fact be derived variably from either the flexor digitorum
superficialis, flexor carpi ulnaris and/or occasionally from the flexor carpi radialis. The “gain” of
a second head (palmaris longus internus) was considered a derived trait in Didelphis (Diogo et
al., 2016; p.1250), with which we agree given that only one palmaris longus appears typical of
dasyurids and most Austalidelphian marsupials. The intrinsic muscles of the manus, derived
from the ventral division of the forelimb, appear in dasyurids to reflect the general arrangement
described in detail in Lewis (1989).
Nine muscles were found to be present in the extensor division of the forearm of the
dasyurids studied: m. brachioradialis, m. supinator, m. extensor carpi radialis, m. extensor carpi
ulnaris, m. extensor digitorum communis, m. extensor digitorum profundus (“indicis”), m.
extensor digiti minimi, m. abductor pollicis longus, together with the m. anconeus lateralis which
Page 30 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

31
is a derivative of this group (Lewis, 1989). This appears to reflect the typical arrangement of this
group within Australidelphian marsupials. Both m. brachioradialis (supinator longus) and m.
supinator (brevis) are present in dasyurids, as they are in Didelphis (Diogo et al., 2016),
reflecting the plesiomorphic condition among mammals generally. M. brachioradialis has been
lost in Peramelemorphia (Parsons, 1903; Warburton et al., 2014) and Notoryctes (Warburton,
2006). M extensor digitorum communis in dasyurids was found to have insertions to the four
ulnar digits, as is typical of most marsupials (Coues, 1871; Barbour, 1963; note this was
mistakenly reported in Warburton et al., 2014 to go to all five digits), and considered the
plesiomorphic condition by Lewis (1989). Derived conditions have been noted in Phascolarctos
(Young, 1882; Sonntag, 1922), Chaeropus (Parsons, 1903) and Notoryctes (Wilson, 1894;
Warburton, 2006) in which the number of insertions has been reduced. Barbour (1964)
considered the m. extensor digitorum profundus to represent the combined m. extensor pollicis
longus and m. extensor indicis within marsupials, but notes significant variation between species;
many authors have described these as separate muscles. Lewis (1989) describes a “proprius”
division of the deep extensor mass with insertions to the first, second and third digits to be
general pattern. Given that we traced insertions to at least four digits in our specimens, as did
Macalister (1872) and Jones (1949) for Sarcophilus and Dasycercus respectively, we suggest
that the general condition of this muscle group likely involved insertion to at least the four radial
digits (pollicis, indicis, medius and annularis) in dasyurids. Barbour (1963) noted that in many
marsupials including Dasycercus (Jones, 1949), the insertion of m. extensor digiti minimi
extends to both the fourth and fifth digits, as did MacCormick (1886a) for D. viverrinus and
Cunningham (1882) for Phascogale, Cuscus and Thylacinus. Lewis (1989) and Diogo et al.
(2016) highlight the insertion to both digits four and five to be the plesiomorphic arrangement
Page 31 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

32
(“extensor digiti quarti et quinti”). The insertion of the lateral extensor in Chaeropus to digit IV
only (Parsons, 1903), and the complete absence of this muscle in Notoryctes (Wilson, 1894;
Warburton, 2006), reflect the highly modified manus in these species.
Derived features of Dasyuridae
In only one instance do we consider the general muscular anatomy of dasyurids to present
a derived state among Australidelphian marsupials. The m. extensor carpi radialis, is described as
being “double” Didelphis; Diogo et al. 2016) and Trichosurus (Barbour, 1963) with separate
long and short heads to the second and third metacarpals respectively. Barbour (1963), later
confirmed by Lewis (1989), regarded the presence of two fairly distinct extensor carpi radialis
muscles as being the plesiomorphic condition among marsupials. Thus the arrangement here for
dasyurids in which a single (combined) belly with insertions to both the second and third
metacarpals is clearly derived from the former pattern. Further modification of this has been
found in D. viverrinus (MacCormick, 1886a), Chaeropus (Parsons, 1903) and Notoryctes
(Wilson, 1894; Warburton, 2006), in which only a single insertion is retained.
Shrivistava (1962) suggested that a tripartite arrangement of the m. deltoideus, in which
cleido-, acromion- and spino-deltoids are clearly separable from one another is plesiomorphic to
an arrangement in which there is some degree of fusion between the various parts, as in
Phascolomys and Phascolarctos (Sonntag, 1922) and large Macropus spp. (Boardman, 1941;
Hopwood, 1974). In contrast, Diogo et al. (2016), on the basis of dissections and literature
review of Didelphis and many eutherian mammals, consider a single deltoideus acromialis and
clavicularis (rather than these being two separate muscles) to be plesiomorphic for all mammals.
Page 32 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

33
If the latter is true, then the differentiated arrangement in dasyuromorphians must be considered
derived, though we reserve judgement on this point.
CONCLUSIONS
It is interesting that there are very few points of difference between Didelphis (Diogo et
al. 2016) and dasyurid marsupials: the abdominal pectoral muscle appears to be part of the
pectoralis major in dasyurids, as opposed to the pectoralis minor in Didelphis; the absence of a
cleidoacromialis in dasyurids; separate cleido- and acromial deltoids in dasyurids; single
palmaris longus and single extensor carpi radialis in dasyurids. Diogo et al. (2016) demonstrated
that Didelphis has undergone relatively fewer changes in muscular anatomy from the last
common ancestor of marsupial and eutherian mammals, than many eutherian groups including
rodents and primates. The relatively few differences between dasyurids and Didelphis, suggest
that the dasyurids have similarly undergone relatively few changes in forelimb muscular
anatomy from the last common ancestor of the two therian mammal clades. Indeed, in some
aspects, such as the single palmaris longus, the tripartite arrangement of the deltoid and the
presence of m. opponens pollicis and m. palmaris brevis, dasyurids appear to retain more
plesiomorphic traits than Didelphis. As hypothesized in the introduction, the musculature of
dasyurid marsupials does indeed appear to correlate with their relatively generalized body forms
and skeletal morphology among Australidelphian marsupials, and thus the anatomy described
herein provides a useful framework for understanding the evolutionary changes that have
occurred in other Australidelphian marsupials.
Page 33 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

34
Marsupials are characterized by giving birth to young that are in an altricial state relative
to their eutherian counterparts. There are different grades of development among marsupials
neonates, and the dasyurid quolls, or “native cats”, (Dasyurus spp.) are among the most highly
altricial at birth (Hill and Hill, 1955). While embryonic in appearance and form, marsupial
neonates also have a range of heterochronic changes, particularly of the craniofacial region and
forelimb, that enable them to make their way to their mothers’ pouch and to suckle (Smith, 2006;
Keyte and Smith, 2010). It is interesting to note, then, that despite their highly embryonic state at
birth, the anatomy of the forelimb muscles in dasyurids appears to develop in line with that of
other marsupials, and that there appears to be no ongoing effects on the arrangement of the
muscles resulting from any heterochronic patterning during development.
ACKNOWLEDGEMENTS
The authors would like to sincerely thank D. Nottle, Z. Wong and J. Hong of the
Murdoch University Veterinary Anatomy Department for their assistance in the laboratory. We
thank the editor and reviewers, especially R. Diogo and M. Ercoli, for the time they gave in
preparing detailed comments and suggestions for the improvement of the manuscript.
Page 34 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

35
LITERATURE CITED
Abdala V, Moro S, Flores DA. 2006. The flexor tendons in the didelphid manus. Mastozool Neotrop 13:193-204.
Amrine-Madsen HA, Scally M, Westerman M, Stanhope MJ, Krajewski C, Springer MS. 2003. Nuclear gene sequences provide evidence for the monophyly of australidelphian marsupials. Mol Phylogenet Evol 28:186-196.
Aplin K, Archer M. 1987. Recent advances in marsupial systematics with a new syncretic classification. In: Archer M, editor. Possums and Opossums: Studies in Evolution. Sydney: Surrey Beatty & Sons & Roy Zool Soc NSW. p xv–lxxii.
Archer M. 1976. The dasyurid dentition and its relationships to that of didelphis, thylacinids, borhyaenids (Marsupicarnivora) and peramelids (Peramelina: Marsupialia). Aust J Zool Supp 39:1-34.
Archer M. 1984. The Australian marsupial radiation. In: Clayton MAG, editor. Vertebrate Zoogeography and Evolution in Australasia. Perth: Hesperian Press. p 633-808.
Archer M, Hand SJ. 2006. The Australian marsupial radiation. In: Merrick JR, Archer M, Hickey GM, Lee MSY, editors. Evolution and biogeography of Australiasian vertebrates. Oatlands, NSW: Australia Auscipub. p 575-646.
Argot C. 2001. Functional-adaptive anatomy of the forelimb in the Didelphidae, and the paleobiology of the Paleocene marsupials Mayulestes ferox and Pucadelphys andinus. J Morphol 247:51-79.
Arlamowska-Palider A, Zablocki J. 1972. Musculus omotransversarius in the light of comparative anatomy. Acta Theriol (Warsz) 17:381-398.
Barbour RA. 1963. The musculature and limb plexus of Trichosurus vulpecula. Aust J Zool 11:488-610.
Baverstock PR, Archer M, Adams M, Richardson BJ. 1982. Genetic relationships among 32 species of Australian dasyurid marsupials. In: Archer M, editor. Carnivorous marsupials. Chipping Norton, NSW: Surrey Beatty & Sons. p 641-650.
Biewener AA. 1989. Scaling body support in mammals: limb posture and muscle mechanics. Science 245:45-48.
Boardman W. 1941. On the anatomy and functional adaptation of the thorax and pectoral girdle in the wallaroo (Macropus robustus). J Linn Soc NSW 66:349-387.
Carlsson A. 1914. Uber Dendrolagus dorianus. Zoologische Jahrbuecher Abteilung fuer Systematic Oekol Geograph der Tiere 36:547-617.
Carlsson A. 1926. Über den Bau des Dasyuroides byrnei und seine Beziehungen zu den übrigen Dasyuridae. Acta Zool 7:249-275.
Cartmill M. 1985. Climbing. In: Hildebrand M, Bramble DM, Liem KF, Wake DB, editors. Functional Vertebrate Morphology. Cambridge: Belknap Press. p 73–88.
Chauveau A. 1891. The comparative anatomy of the domesticated animals. 2nd English ed. New York: D. Appleton.
Cheng C. 1955. The development of the shoulder region of the opossum, Didelphis virginiana, with special reference to musculature. J Morphol 97:415-471.
Coues E. 1871. The osteology and myology of Didelphis virginiana. Mem Bost Soc Nat Hist 2:41-149.
Crowther MS. 2008. Yellow-footed antechinus, Antechinus flavipes (Waterhouse, 1838). In: Van Dyck S, Strahan R, editors. The Mammals of Australia. Sydney: Reed Holland. p 86-88.
Page 35 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

36
Cunningham DJ. 1878a. The Intrinsic Muscles of the Hand of the Thylacine (Thylacinus cynocephalus), Cuscus (Phalangista maculata), and Phascogale (Phascogale calura). J Anat Physiol 12:434-444.
Cunningham DJ. 1878b. The Nerves of the Fore-Limb of the Thylacine (Thylacinus cynocephalus or harrisii) and Cuscus (Phalangista maculata). J Anat Physiol 12:427–433.
Cunningham DJ. 1881. The nerves of the hind-limb of the thylacine (Thylacinus harrisii or cynocephalus) and cuscus (Phalangista maculata). J Anat 15:265–277.
Cunningham DJ. 1882. Report on Some Points in the Anatomy of the thylacine (Thylacinus cynocephalus), cuscus (Phalangista maculata), and phascogale (Phascogale calura), collected during the Voyage of H. M. S. Challenger in the years 1873-1876; with an account of the Comparative Anatomy of the Intrinsic Muscles and Nerves of the Mammalian Pes. Voy H M S Challenger, Zool V, pt. XVI.
Davis DD. 1964. The giant panda: a morphological study of evolutionary mechanisms. Field Zool Mem 3:1-339.
Davison A. 1944. Mammalian anatomy with special reference to the cat. Revised by Stromstein FA, editor. Sixth Edition. Philadelphia: The Blakiston Company.
Diogo R, Abdala V. 2010. Muscles of vertebrates - comparative anatomy, evolution, homologies and development. Oxford: Taylor and Francis.
Diogo R, Abdala V, Aziz MA, Lonergan N, Wood BA. 2009. From fish to modern humans; comparative anatomy, homologies and evolution of the pectoral and forelimb musculature. J Anat 214:694-716.
Diogo R, Bello-Hellegouarch G, Kohlsdorf T, Esteve-Altava B, Molnar JL. 2016. Comparative Myology and Evolution of Marsupials and Other Vertebrates, With Notes on Complexity, Bauplan, and “Scala Naturae”. Anat Rec 299:1224-1255.
Diogo R, Pastor F, De Paz F, Potau JM, BelloSHellegouarch G, Ferrero EM, Fisher RE. 2012. The head and neck muscles of the serval and tiger: homologies, evolution, and proposal of a mammalian and a veterinary muscle ontology. Anat Rec 295:2157-2178.
Dixon JM. 1989. Thylacinidae. In: Walton WD, Richardson BJ, editors. Fauna of Australia. 1B Mammalia. Canberra: Australian Government Publishing Service. p 549-559.
Elftman HO. 1929. Functional adaptations of the pelvis in marsupials. Bull Am Mus Nat Hist 58:189-232.
Ercoli MD, Alvarez A, Stafanini MI, Busker F, Morales MM. 2015. Muscular anatomy of the forelimbs of the lesser grison (Galictis cuja), and a functional and phylogenetic overview of mustelidae and other Caniformia. J Mamm Evol 22:57-91.
Filan SL. 1990. Myology of the head and neck of the bandicoot (Marsupialia; Peramelemorphia). Aust J Zool 38:617-634.
Fisher RE, Adrian B, Barton M, Holmgren J, Tang SY. 2009. The phylogeny of the red panda (Ailurus fulgens): evidence from the forelimb. J Anat 215:611-635.
Gordon G, Hulbert AJ. 1989. Peramelidae. In: Walton DW, Richardson BJ, editors. Fauna of Australia. 1B Mammalia. Canberra: Australian Government Publishing Service. p 603-624.
Harvey KJ, Warburton NM. 2010. Forelimb musculature of kangaroos with particular emphasis on the tammar wallaby Macropus eugenii (Desmarest, 1817). Aust Mammal 32:1-9.
Page 36 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

37
Hill JP, Hill WC. 1955. The growthSstages of the pouchSyoung of the native cat (Dasyurus viverrinus) together with observations on the anatomy of the newSborn young. Trans Zool Soc Lond 28:349-352.
Hopwood PR. 1974. The intrinsic musculature of the pectoral limb of the Eastern Grey Kangaroo (Macropus major (Shaw) Macropus giganteus (Zimm)). J Anat 118:445-468.
Horiguchi M. 1981. A comparative anatomical study of the pectoral muscle group in the brindled bandicoot (Isoodon macrourus Gould 1842) an Australian marsupial. Acta Anat Nippon 56:375-399.
Horovitz I, Sanchez-Villagra MR. 2003. A morphological analysis of marsupial mammal higher-level phylogenetic relationships. Cladistics 19:181-212.
Jenkins FA, Weijs WA. 1979. The functional anatomy of the shoulder in the Virginia opossum (Didelphis virginiana). J Zool 188:379-410.
Jones FW. 1949. The study of a generalised marsupial (Dasycercus cristicauda Krefft). Trans Zool Soc 26:409-501.
Jones FW. 1968. The Mammals of South Australia. Adelaide: Government Printer. Jones M. 1995. Tasmanian devil, Sarcophilus harrisii. In: Strahan R, editor. The Mammals of
Australia. NSW, Australia: Reed Books Australia. p 82-84. Keyte AL, Smith KK. 2010. Developmental origins of precocial forelimbs in marsupial neonates.
Development 137:4283-4294. Krajewski C, Blacket M, Buckley L, Westerman M. 1997. A Multigene Assessment of
Phylogenetic Relationships within the Dasyurid Marsupial Subfamily Sminthopsinae. Mol Phylogenet Evol 8:236-248.
Krajewski C, Wroe S, Westerman M. 2000. Molecular evidence for the pattern and timing of cladogenesis in dasyurid marsupials. Zool J Linn Soc 130:375-404.
Lewis OJ. 1989. Functional morphology of the evolving hand and foot. Oxford: Clarendon Press. Macalister A. 1872. XIX. The muscular anatomy of the koala (Phascolarctos cinereus). Ann
Mag Nat Hist 10:127-135. MacAlister A. 1870. XVI. On the myology of the Wombat (Phascolomys wombata) and the
Tasmanian devil (Sarcophilus ursinus). Ann Mag Nat Hist 5:153-173. Macalister A. 1872. Further observations on the myology of Sarcophilus ursinus. Ann Mag Nat
Hist 10:17-20. MacCormick A. 1886a. The myology of the limbs of Dasyurus viverrinus. A. Myology of the
forelimb. J Anat Physiol 21:103-137. MacCormick A. 1886b. The myology of the limbs of Dasyurus viverrinus. B. Myology of the
hindlimb. J Anat Physiol 21:199-226. Marshall LG, Case JA, Woodburne MO. 1990. Phylogenetic relationships of the families of
marsupials. Current Mammal 2:433-505. Masters P. 2008. Crest-tailed mulgara, Dasycercus cristicauda (Kreft, 1867). In: Van Dyck S,
Strahan R, editors. The Mammals of Australia. Sydney: Reed Holland. p 49-50. Meredith RW, Westerman M, Case JA, Springer MS. 2008. A Phylogeny and Timescale for
Marsupial Evolution Based on Sequences for Five Nuclear Genes. J Mamm Evol 15:1-36.
Miller ME. 2012. Miller's anatomy of the dog. Howard E. Evans, George C. Christensen, editors. 2nd ed. Philadelphia: Saunders.
Page 37 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

38
Morton SR, Dickman CR, Fletcher TP. 1989. Dasyuridae. In: Walton DW, Richardson BJ, editors. Fauna of Australia. 1B Mammalia. Canberra: Australian Government Publishing Service. p 560-582.
Oakwood M. 2008. Northern quoll, Dasyurus hallucatus (Gould, 1842). In: Van Dyck S, Strahan R, editors. The Mammals of Australia. Sydney: Reed Holland. p 57-59.
Parsons FG. 1896. On the anatomy of Petrogale xanthopus, compared with that of other kangaroos. Proc Zool Soc Lond 64:683-714.
Parsons FG. 1903. Anatomy of the pig-footed bandicoot (Chaeropus castanotis). J Linn Soc Lond 29:64-80.
Reese S, Pfuderer U, Bragulla H, Loeffler K, Budras K. 2001. Topography, structure and function of the patella and the patelloid in marsupials. Anat Histol Embryol 30:289-294.
Rounsevell DE, Mooney N. 1995. Thylacine, Thylacinus cynocephalus. In: Strahan R, editor. The Mammals of Australia. NSW, Australia: Reed Books. p 164-165.
Serena M, Soderquist T. 2008. Western quoll, Dasyurus geoffroii (Gould, 1841). In: Van Dyck S, Strahan R, editors. The Mammals of Australia. Sydney: Reed Holland. p 54-56.
Schaller O. 2007 Illustrated Veterinary Anatomical Nomenclature. Stuttgart: Enke Verlag. Shrivastava RK. 1962. The deltoid musculature of the Marsupialia. Am Midl Nat 67:305-320. Smith KK. 2006. Craniofacial development in marsupial mammals: developmental origins of
evolutionary change. Dev Dyn 235:1181-1193. Soderquist T, Rhind S. 2008. Brush-tailed Phascogale, Phascogale tapoatafa (Meyer, 1793). In:
Van Dyck S, Strahan R, editors. The Mammals of Australia. Sydney: Reed Holland. p 105-107.
Sonntag CF. 1922. On the myology and classification of wombat, koala and phalanger. Proc Zool Soc Lond 92:863-869.
Stirling EC. 1891. Description of a new genus and species of Marsupialia, Notoryctes typhlops. Trans Roy Soc S Aust 14:154-187.
Szalay FS. 1982. A new appraisal of marsupial phylogeny and classification. In: Archer M, editor. Carnivorous Marsupials. Sydney: Roy Zool Soc NSW. p 612-640.
Taylor ME. 1974. The functional anatomy of the forelimb of some African Viverridae (Carnivora). J Morphol 143:307-336.
Warburton NM. 2006. Functional morphology of marsupial moles (Marsupialia: Notoryctidae). Verhandlungen des Naturwissenschaftlichen Vereins Hamburg 42:39-149.
Warburton NM. 2014. Relicts, Reproduction and Reintroductions - A century of marsupial research in Western Australia. J Roy Soc W Aust 97:65-85.
Warburton NM, Bateman PW, Fleming PA. 2013. Sexual selection on forelimb muscles of western grey kangaroos (Skippy was clearly a female). Biol J Linn Soc 109:923-931.
Warburton NM, Gregoire L, Jacques S, Flandrin C. 2014. Adaptations for digging in the forelimb muscle anatomy of the southern brown bandicoot (Isoodon obesulus) and bilby (Macrotis lagotis). Aust J Zool 61:402-419.
Warburton NM, Harvey KJ, Prideaux GJ, O'Shea JE. 2011. Functional morphology of the forelimb of living and extinct tree-kangaroos (Marsupialia: Macropodidae). J Morphol 272:1230-1244.
Warburton NM, Malric A, Yakovleff M, Leonard V, Cailleau C. 2015. Hind limb myology of the southern brown bandicoot Isoodon obesulus and greater bilby Macrotis lagotis (Marsupialia: Peramelemorphia). Aust J Zool 63:147-162.
Page 38 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

39
Warburton NM, Yakovleff M, Malric A. 2012. Anatomical adaptations of the hind limb musculature of tree-kangaroos for arboreal locomotion (Marsupialia: Macropodinae). Aust J Zool 60:246-258.
Westerman M, Krajewski C, Kear BP, Meehan L, Meredith RW, Emerling CA, Springer MS. 2015. Phylogenetic relationships of dasyuromorphian marsupials revisited. Zool J Linn Soc 176:686-701
Wilson JT. 1894. On the myology of Notoryctes typhlops with comparative notes. Trans Roy Soc S Aust 18:3-74.
Wood JT. 1870. On a group of varieties of the muscles of the human neck, shoulder, and chest, with their transitional forms and homologies in the Mammalia. Phil Trans Roy Soc Lond 160:83-116.
Young AH. 1879. The intrinsic muscles of the marsupial hand. J Anat Physiol 14:149-167. Young AH. 1882. The muscular anatomy of the koala. J Anat 16:217-242.
FIGURE CAPTIONS
Figure 1. Simplified phylogenetic tree of the Dasyuromorphia (modified from Westerman et al.
2015; branch lengths are not indicative).
Figure 2. Superficial lateral view of the right forelimb muscles in Dasyurus geoffroii; inset:
shoulder region with trapezius removed.
Figure 3. Muscle attachments on the right scapula of Dasyurus geoffroii (A, B, C) and
Phascogale tapoatafa (D, E, F). (A,D) medial view, (B,E) caudal view and (C,F) lateral view
(D,G). Atl, m. atlantoscapularis posticus; BiB, m. biceps brachii; Cor, m. coracobrachialis; DeA,
m. deltoideus acromialis; DeS m. deltoideus scapularis; Inf, m. infraspinatus; LCl, m. levator
Page 39 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

40
claviculae; OmH, m. omohyoideus; Rho, m. rhomboideus; SAn, m. serratus anterior; Sub, m.
subscapularis; Sup, m. supraspinatus; TMj, m. teres major; TMn, m. teres minor; TrC, m.
trapezius pars cervicalis; TrT, m. trapezius pars thoracis; TLn, m. triceps caput longum.
Figure 4. Lateral view of the right forelimb muscles in Phascogale tapoatafa, with m. trapezius
and mm. rhomboideus removed.
Figure 5. Superficial medial view of the right forelimb muscles in Dasyurus geoffroii.
Figure 6. Muscle attachments on the right humerus of Dasyurus geoffroii (A-D) and Phascogale
tapoatafa (E-H). (A,E) medial view, (B,F) caudal view and (C,G) lateral view, (D, H) cranial
view. ALt, m. anconeus lateralis; Bra, m. brachialis; BRd, m. brachioradialis; Cor, m.
coracobracialis; Del, m. deltoideus; ECR, m. extensor carpi radialis; ECU, m. extensor carpi
ulnaris; EDC, m. extensor digitorum communis; EDM, m. extensor digiti minimi; Epi, m.
epitrochleoanconeus; FCR, m. flexor carpi radialis; FCU, m. flexor carpi ulnaris; FDP, m. flexor
digitorum profundus; Inf, m. infraspinatus; LtD, m. latissimus dorsi; PaL, m. palmaris longus;
PeM, m. pectoralis major; Pmn, m. pectoralis minor; PrT, m. pronator teres; Spr, m. supinator;
Sub, m. subscapularis; Sup, m. supraspinatus; TLt, m. triceps caput lateral; TMd, m. triceps
caput medial; TMj, m. teres major; TMn, m. teres minor.
Figure 7. Muscle attachments on the right radius, ulna and carpus of Dasyurus geoffroii (A,B)
and Phascogale tapoatafa (C, D). (A,C) lateral view and (B, D) medial view. ALt, m. anconeus
lateralis; APL, M. abductor pollicis longus; BiB, m. biceps brachii; Bra, m. brachialis; EDC, m.
extensor digitorum communis; EDP, m. extensor digitorum profundus; Epi, m.
epitrochleoanconeus; FDP, m. flexor digitorum profundus; PrQ, m. pronator quadratus; PrT, m.
Page 40 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

41
pronator teres; Spr, m. supinator; TMd, m. triceps caput medial, TLn, m. triceps caput longum;
TLt, m. triceps caput lateral.
Figure 8. Medial view of the intrinsic forelimb muscles in Dasyurus hallucatus (right), humeral
head m. flexor digitorum profundus reflected.
Figure 9. Muscle attachments of the right manus of Dasyurus geoffroii, A dorsal view, B palmar
view.
Figure 10. Deep lateral view of the extensor muscles in the right forearm of Dasyurus hallucatus.
Figure 11. Deep intrinsic muscles of the right manus of Dasyurus hallucatus.
Page 41 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.
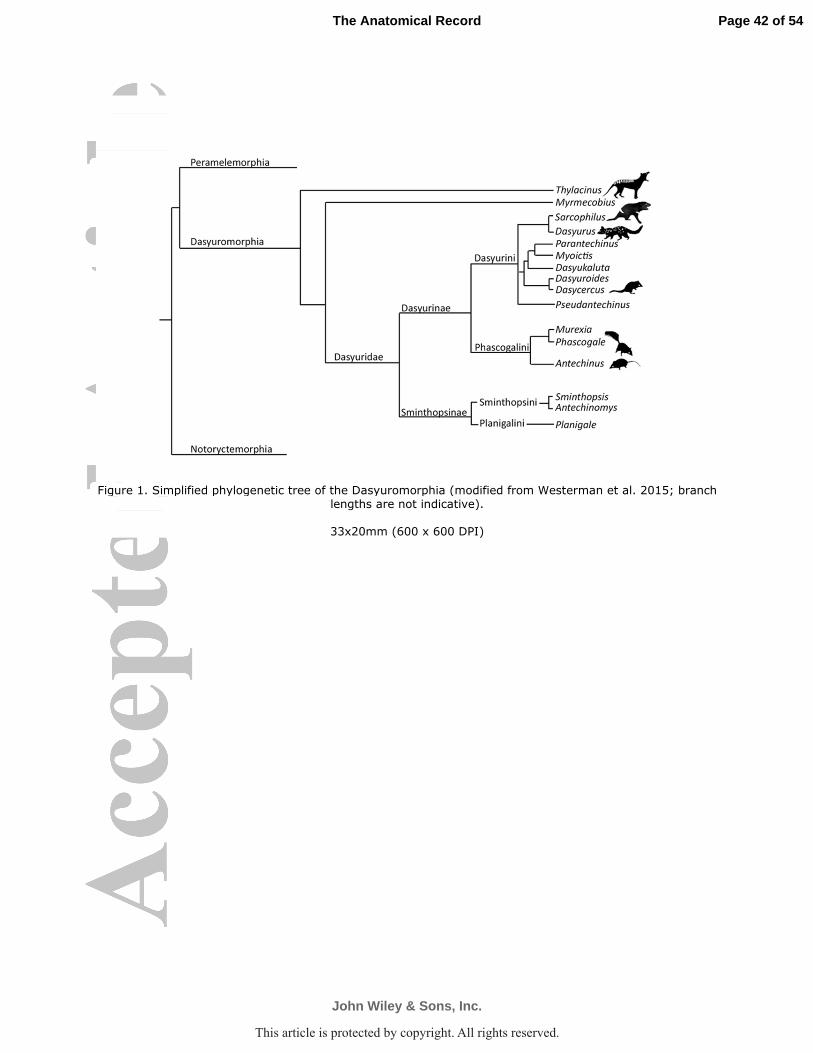
Figure 1. Simplified phylogenetic tree of the Dasyuromorphia (modified from Westerman et al. 2015; branch lengths are not indicative).
33x20mm (600 x 600 DPI)
Page 42 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.
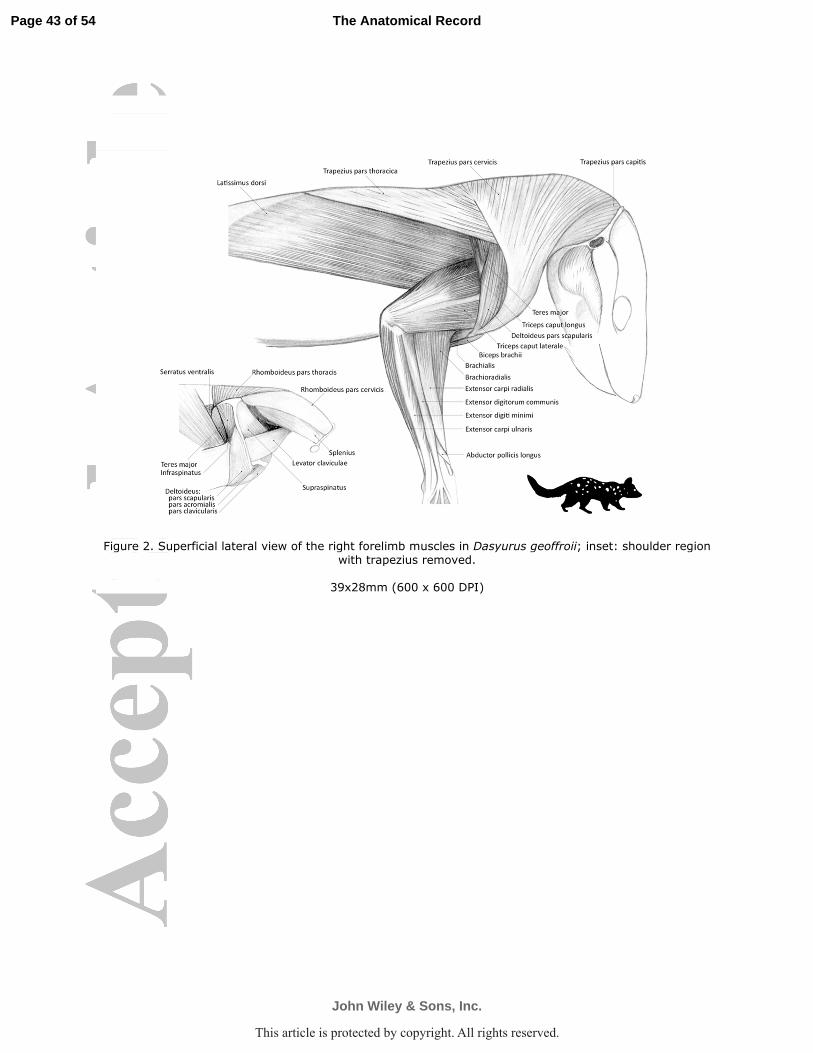
Figure 2. Superficial lateral view of the right forelimb muscles in Dasyurus geoffroii; inset: shoulder region with trapezius removed.
39x28mm (600 x 600 DPI)
Page 43 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.
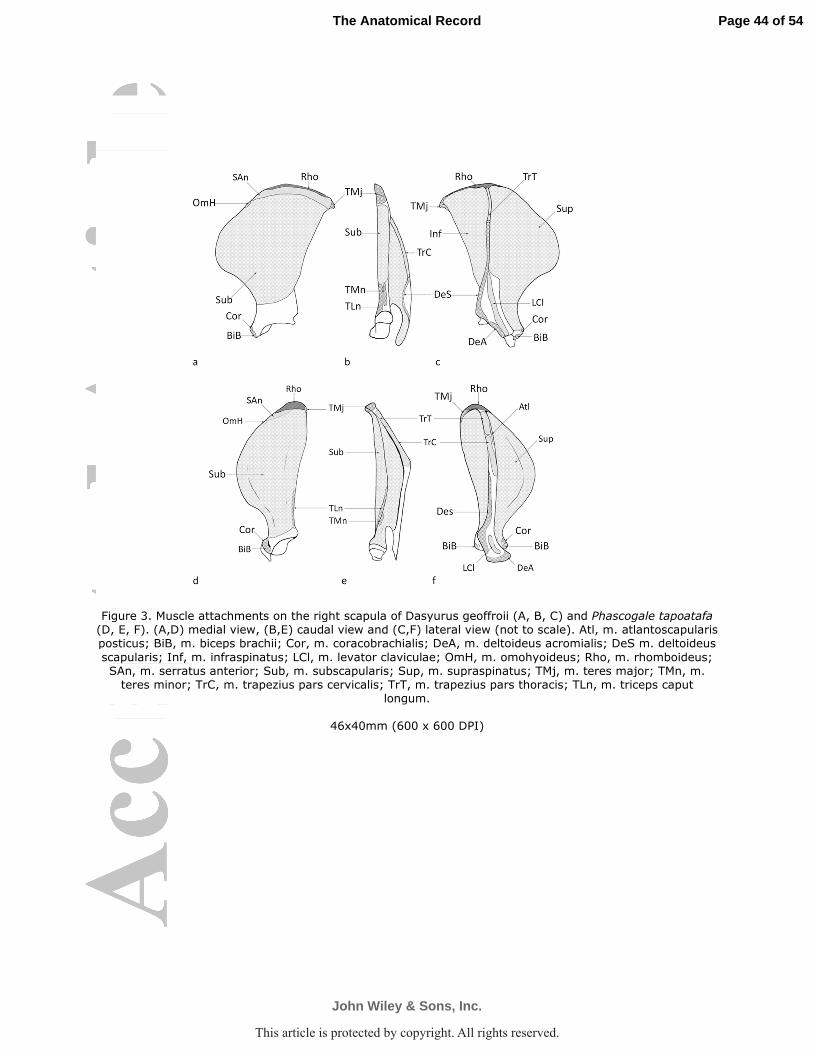
Figure 3. Muscle attachments on the right scapula of Dasyurus geoffroii (A, B, C) and Phascogale tapoatafa (D, E, F). (A,D) medial view, (B,E) caudal view and (C,F) lateral view (not to scale). Atl, m. atlantoscapularis posticus; BiB, m. biceps brachii; Cor, m. coracobrachialis; DeA, m. deltoideus acromialis; DeS m. deltoideus
scapularis; Inf, m. infraspinatus; LCl, m. levator claviculae; OmH, m. omohyoideus; Rho, m. rhomboideus; SAn, m. serratus anterior; Sub, m. subscapularis; Sup, m. supraspinatus; TMj, m. teres major; TMn, m. teres minor; TrC, m. trapezius pars cervicalis; TrT, m. trapezius pars thoracis; TLn, m. triceps caput
longum.
46x40mm (600 x 600 DPI)
Page 44 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.
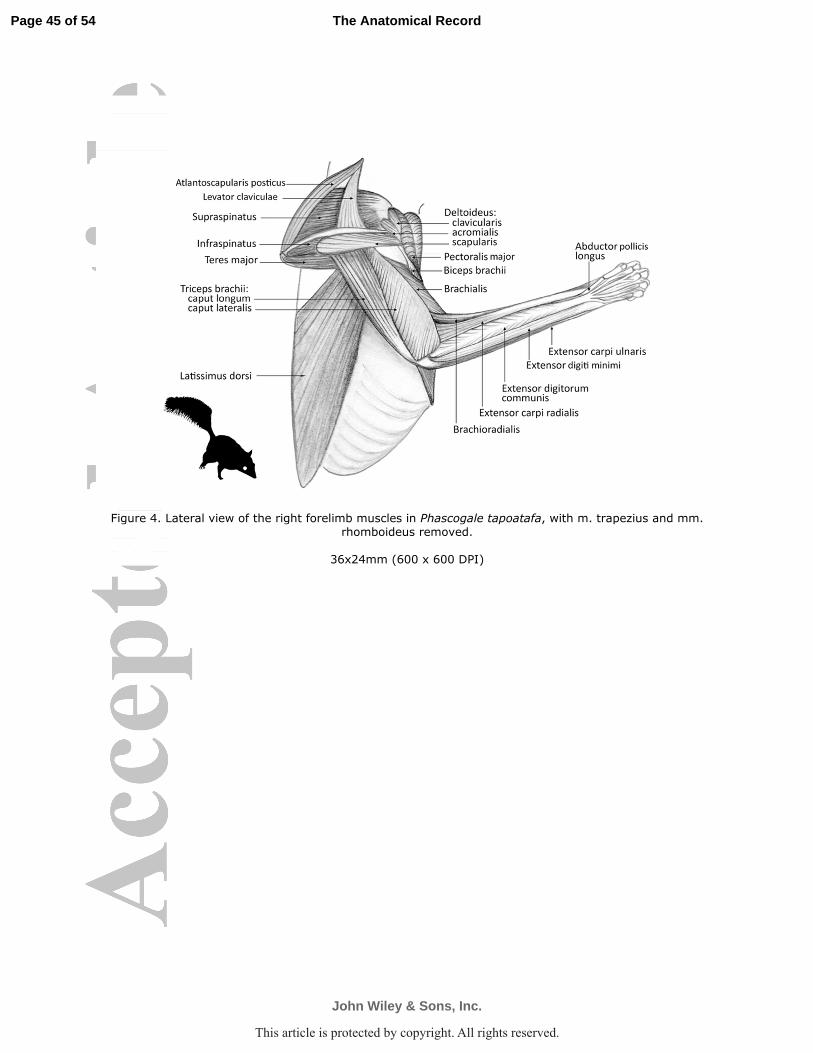
Figure 4. Lateral view of the right forelimb muscles in Phascogale tapoatafa, with m. trapezius and mm. rhomboideus removed.
36x24mm (600 x 600 DPI)
Page 45 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.
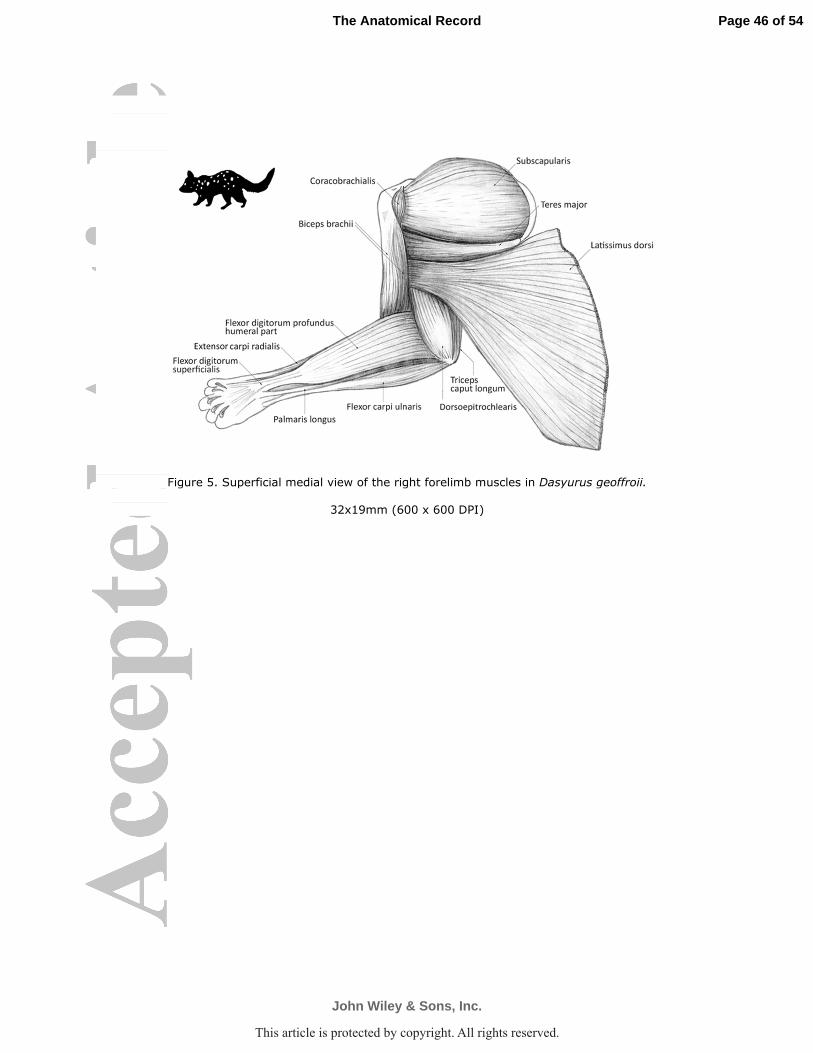
Figure 5. Superficial medial view of the right forelimb muscles in Dasyurus geoffroii.
32x19mm (600 x 600 DPI)
Page 46 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.
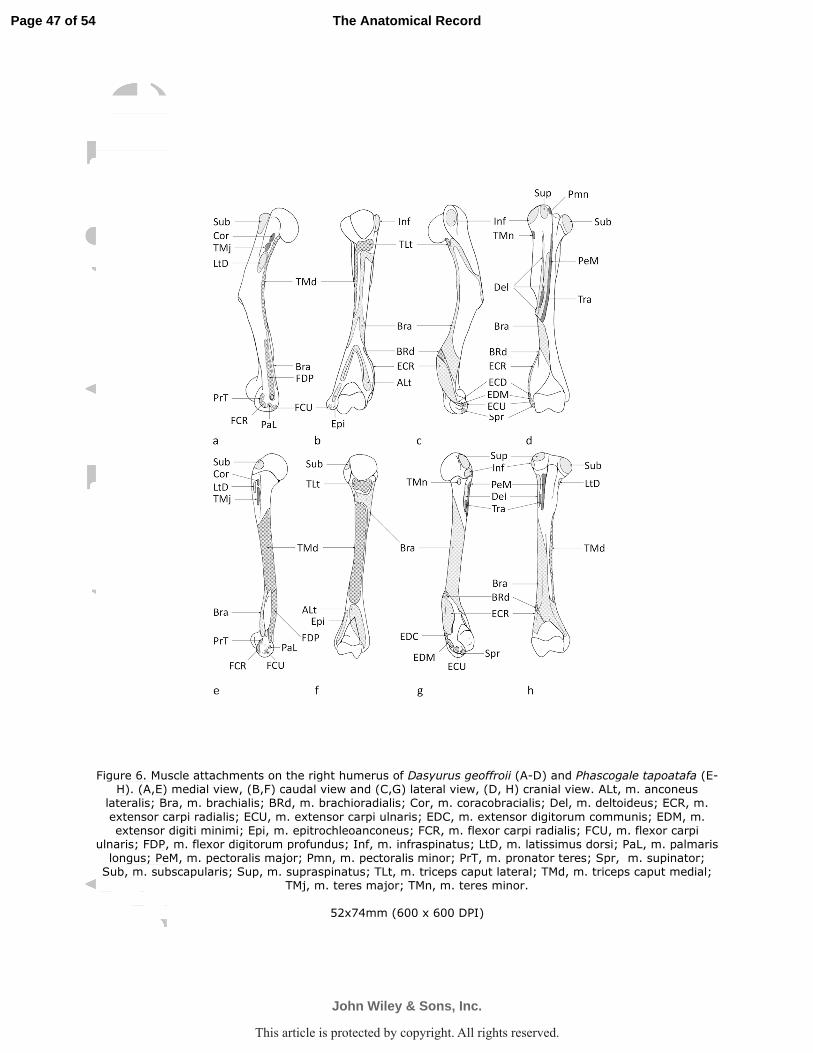
Figure 6. Muscle attachments on the right humerus of Dasyurus geoffroii (A-D) and Phascogale tapoatafa (E-H). (A,E) medial view, (B,F) caudal view and (C,G) lateral view, (D, H) cranial view. ALt, m. anconeus
lateralis; Bra, m. brachialis; BRd, m. brachioradialis; Cor, m. coracobracialis; Del, m. deltoideus; ECR, m.
extensor carpi radialis; ECU, m. extensor carpi ulnaris; EDC, m. extensor digitorum communis; EDM, m. extensor digiti minimi; Epi, m. epitrochleoanconeus; FCR, m. flexor carpi radialis; FCU, m. flexor carpi
ulnaris; FDP, m. flexor digitorum profundus; Inf, m. infraspinatus; LtD, m. latissimus dorsi; PaL, m. palmaris longus; PeM, m. pectoralis major; Pmn, m. pectoralis minor; PrT, m. pronator teres; Spr, m. supinator; Sub, m. subscapularis; Sup, m. supraspinatus; TLt, m. triceps caput lateral; TMd, m. triceps caput medial;
TMj, m. teres major; TMn, m. teres minor.
52x74mm (600 x 600 DPI)
Page 47 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.
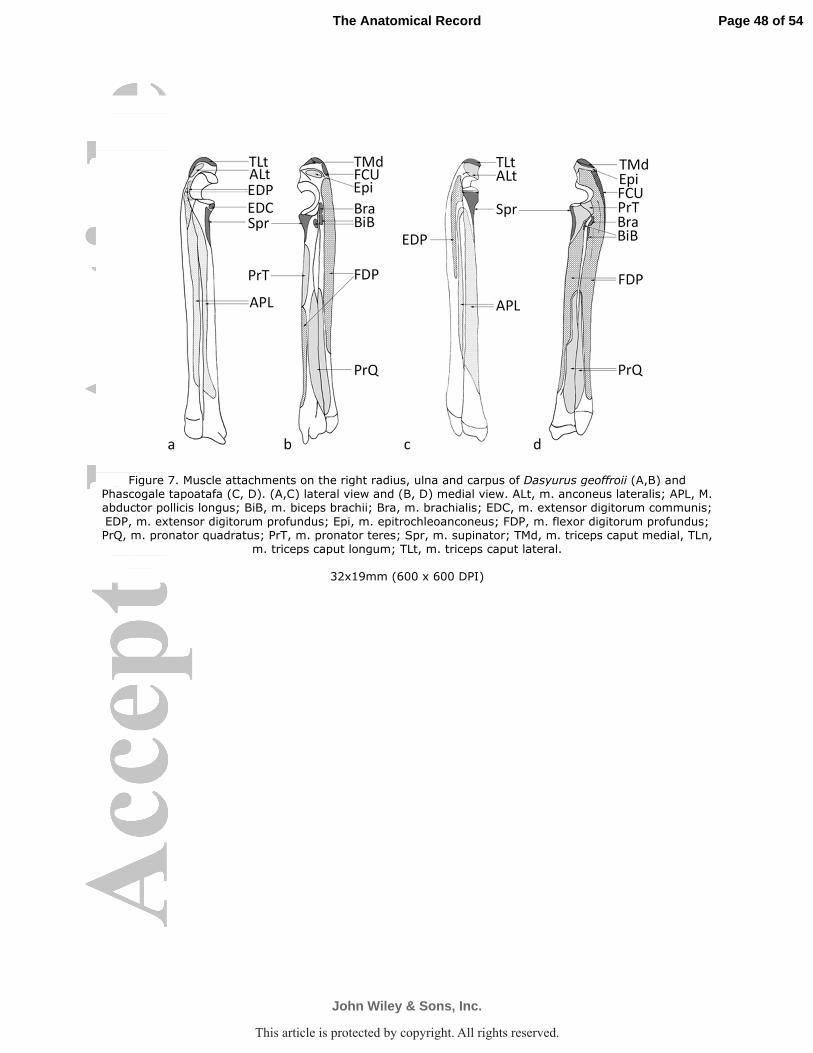
Figure 7. Muscle attachments on the right radius, ulna and carpus of Dasyurus geoffroii (A,B) and Phascogale tapoatafa (C, D). (A,C) lateral view and (B, D) medial view. ALt, m. anconeus lateralis; APL, M. abductor pollicis longus; BiB, m. biceps brachii; Bra, m. brachialis; EDC, m. extensor digitorum communis;
EDP, m. extensor digitorum profundus; Epi, m. epitrochleoanconeus; FDP, m. flexor digitorum profundus; PrQ, m. pronator quadratus; PrT, m. pronator teres; Spr, m. supinator; TMd, m. triceps caput medial, TLn,
m. triceps caput longum; TLt, m. triceps caput lateral.
32x19mm (600 x 600 DPI)
Page 48 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.
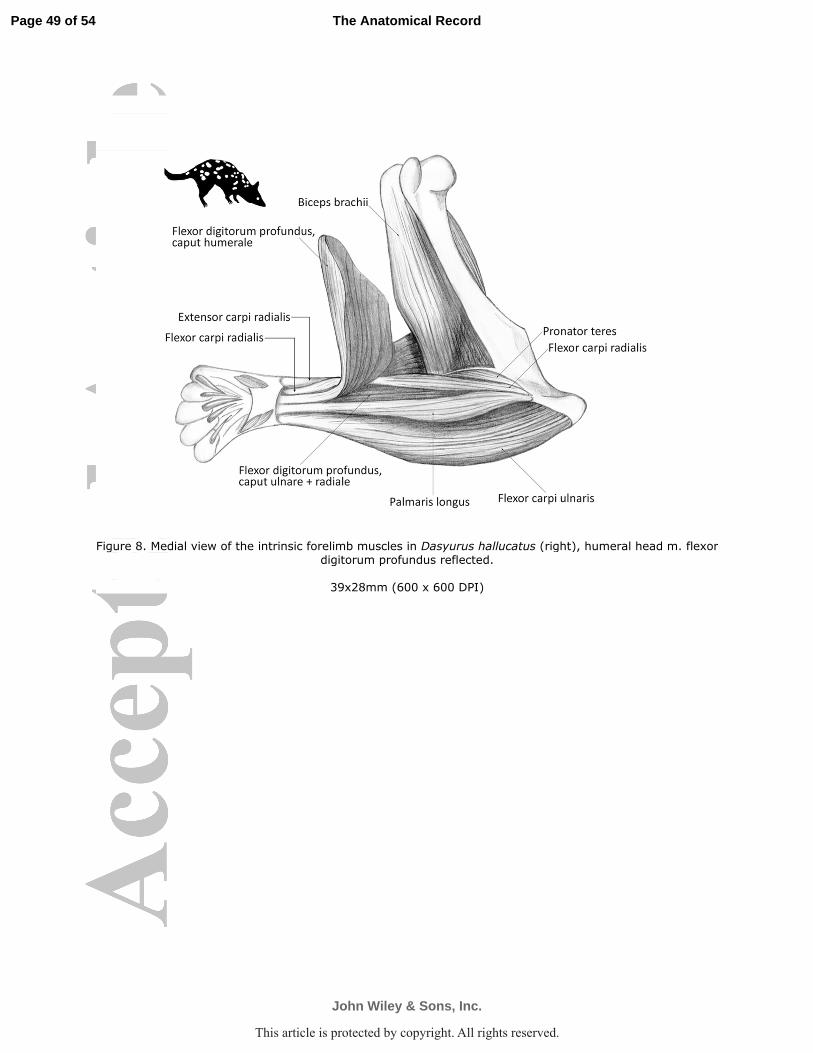
Figure 8. Medial view of the intrinsic forelimb muscles in Dasyurus hallucatus (right), humeral head m. flexor digitorum profundus reflected.
39x28mm (600 x 600 DPI)
Page 49 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.
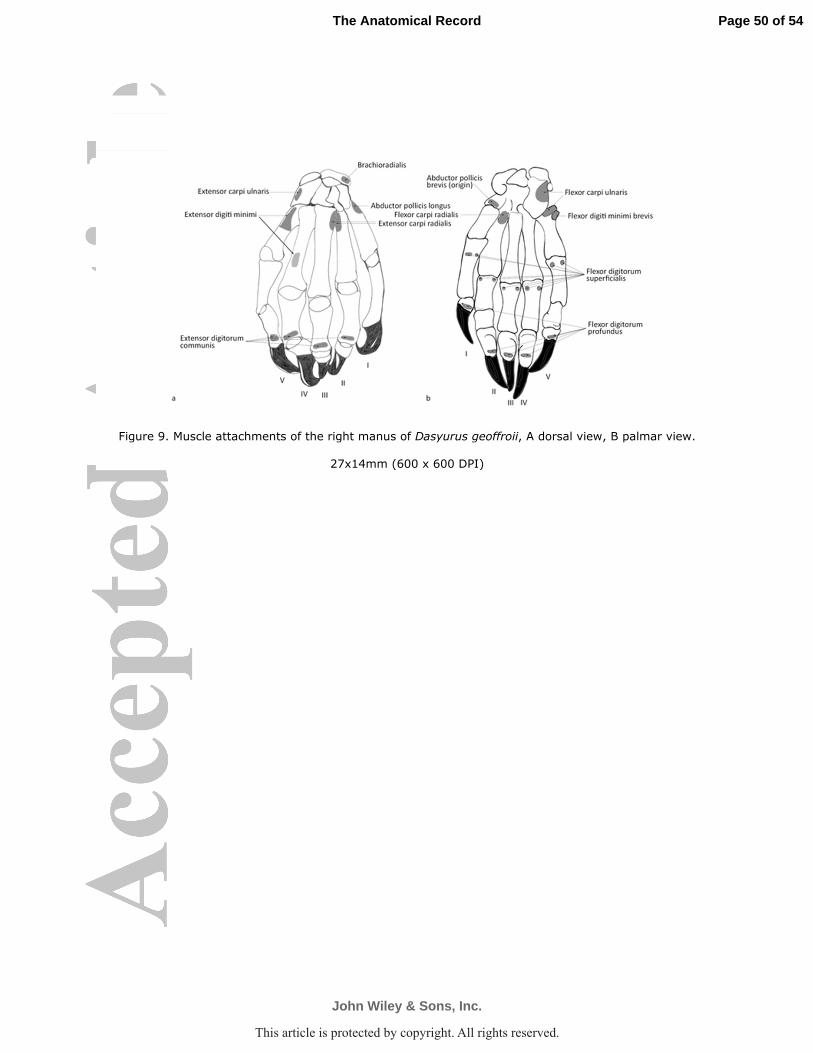
Figure 9. Muscle attachments of the right manus of Dasyurus geoffroii, A dorsal view, B palmar view.
27x14mm (600 x 600 DPI)
Page 50 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.
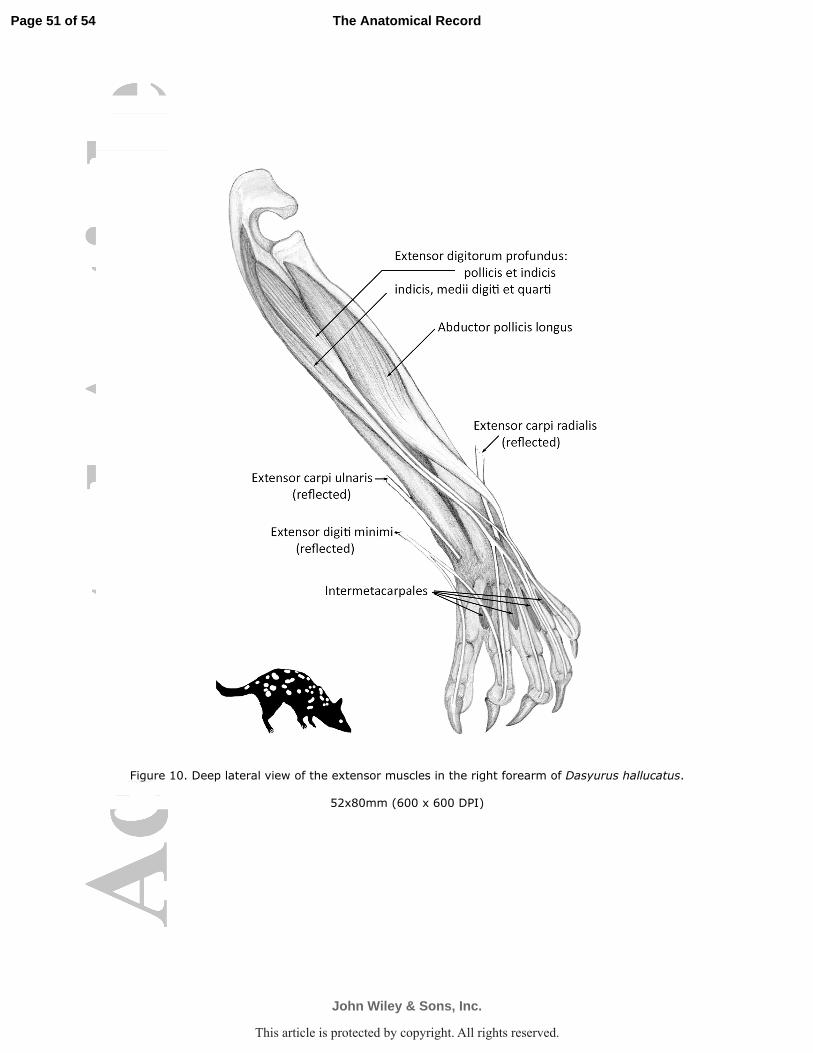
Figure 10. Deep lateral view of the extensor muscles in the right forearm of Dasyurus hallucatus.
52x80mm (600 x 600 DPI)
Page 51 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.
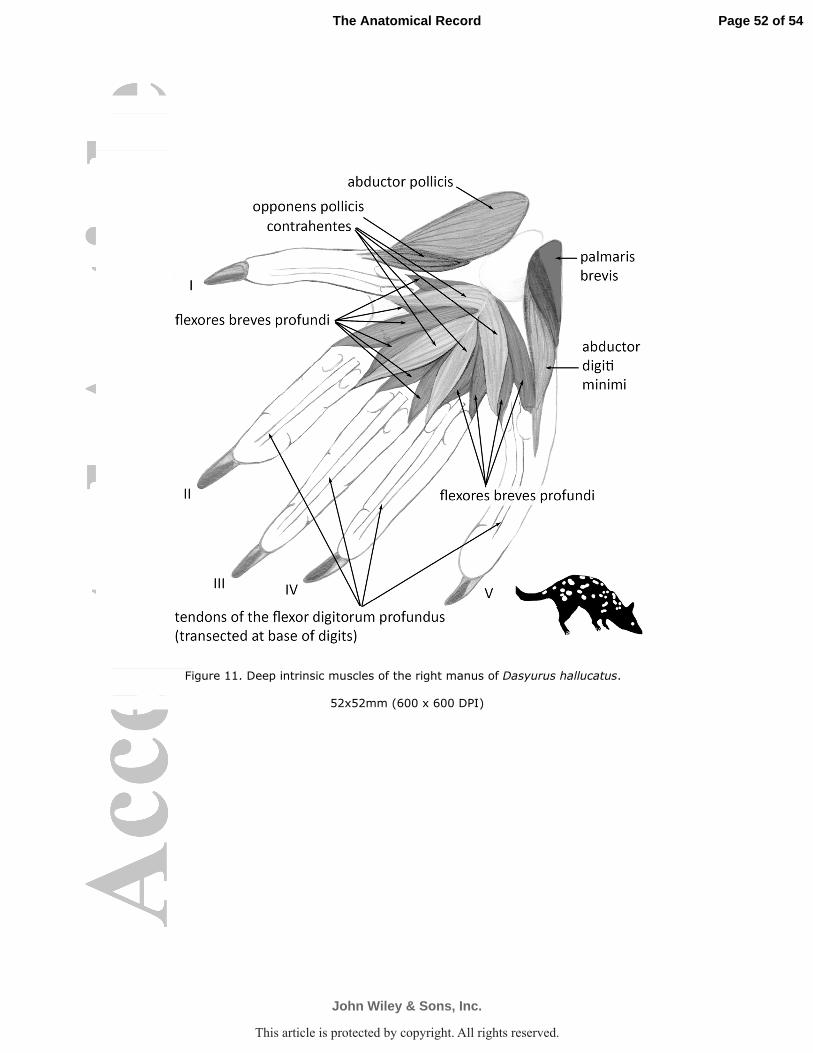
Figure 11. Deep intrinsic muscles of the right manus of Dasyurus hallucatus.
52x52mm (600 x 600 DPI)
Page 52 of 54
John Wiley & Sons, Inc.
The Anatomical Record
This article is protected by copyright. All rights reserved.

Table 1 - Biological characteristics of dasyuromorphian species investigated in this study (modified from (Warburton, 2014).
Species Body mass Ecology References
Antechinus flavipes
Yellow-footed antechinus,
mardo
26-79 g males;
21-52 g females
Primarily terrestrial; insectivorous; inhabits varied
habitats from forests to swamps and dry mulga
country.
(Crowther, 2008)
Dasycercus cristicauda
Crest-tailed mulgara,
ampurta
110-185 g males;
65-120 g females
Terrestrial; diet of arthropods and small
vertebrates; constructs burrows in desert sand
dunes with sparse cover.
(Masters, 2008)
Dasyurus geoffroii
Western quoll, chuditch
1310 g males (mean);
890 g females (mean)
Arboreal and terrestrial; mixed carnivorous diet,
mostly small to medium sized vertebrates;
sclerophyll forest, dry woodland or shrubland of
the south-west Australia.
(Serena and Soderquist,
2008)
Dasyurus hallucatus
Northern quoll
340-1120 g males; 240-
690 g females
Arboreal and terrestrial; diet mostly small
vertebrates; most commonly inhabits rocky
escarpments; also found in woodlands.
(Oakwood, 2008)
Phascogale tapoatafa
Brush-tailed phascogale,
common wambenger
175-311 g males;
106-212 g females
Arboreal specialist; primarily arthropod diet; open
forest with sparse groundcover.
(Soderquist and Rhind,
2008)
Sarcophilus harrisii
Tasmanian devil
9 kg males (mean);
7 kg females (mean)
Terrestrial; hypercarnivore, efficient scavenger;
inhabits varied habitats across Tasmania (formerly
Australian mainland)
(Jones, 1995)
Thylacinus cynocephalus
Tasmanian tiger
15-35 kg (species
mean)
Cursorial apex predator; varied habitats, most
abundant in open forest and woodland. Extinct.
(Rounsevell and Mooney,
1995)
Page 53 of 54
John Wiley & Sons, Inc.
The Anatomical Record
5051525354555657585960
This article is protected by copyright. All rights reserved.
Table 2 – Sources of Data for the Order Marsupialia (taxonomy after (Aplin and Archer, 1987))
Didelphimorphia Dasyuromorphia Diprotodontia Peramelemorphia Notoryctemorphia
Caluromys Dasycercus Dendrolagus Chaeropus Notoryctes
(Argot, 2001) (Jones, 1949) (Carlsson, 1914) (Parsons, 1903) (Wilson, 1894)
(Warburton et al., 2011) (Warburton, 2006)
Didelphis Dasyurus Echymypera
(Coues, 1871) (MacCormick, 1886a) Macropus (Filan, 1990)
(Jenkins and Weijs, 1979) (Boardman, 1941)
(Cheng, 1955) Sarcophilus (Hopwood, 1974) Isoodon
(MacAlister, 1870) (Harvey and Warburton, 2010) (Filan, 1990)
(Macalister, 1872) (Horiguchi, 1981)
Petrogale (Warburton et al., 2014)
Thylacinus (Parsons, 1896)
(Cunningham, 1878b) Macrotis
(Cunningham, 1878a) Phalanger (Filan, 1990)
(Sonntag, 1922) (Warburton et al., 2014)
Phascolactos
(Macalister, 1872)
(Young, 1882)
(Sonntag, 1922)
Phascolomys
(MacAlister, 1870)
(Sonntag, 1922)
Trichosurus
(Barbour, 1963)
Page 54 of 54
John Wiley & Sons, Inc.
The Anatomical Record
5051525354555657585960
This article is protected by copyright. All rights reserved.




















