
Comparative Use of Riparian Corridors and Oases by Migrating Birds in Southeast Arizona

INTRODUCTION
All muscles of the vertebrate body derive from the segmentedparaxial mesoderm of the somites (reviewed in Hauschka,1994; Christ and Ordahl, 1995). Epaxial muscles, such as thoseof the back, derive from progenitor cells in their dorsomedialquarter. These initially form the myotome, under the influenceof signal(s) from axial structures, neural tube and notochord,which induce myogenic factor expression. In contrast, hypaxialmuscles, such as those of the body wall, derive from thedorsolateral quarter of the somite and myogenesis depends oninductive signals from the dorsal ectoderm and is retarded bylateral mesoderm (see Cossu et al., 1996b for review).
The somitic contribution to limb muscles was firstdemonstrated for birds using chick/quail chimaeras (Chevallieret al., 1977; Christ et al., 1977). Morphological observationsalso pointed to cell migration from somites to the limb inmouse embryos (Milaire, 1976, 1987). Experiments in whichlabelled rodent somites were transplanted into culturedembryos showed that marked somitic cells moved into the limbbud (Beddington and Martin, 1989; Lee and Sze, 1993; Sze etal., 1995). However, it was the transplantation of somites fromdesmin-lacZ embryos which provided clear evidence for themigration of muscle progenitor cells from the somites, sincedesmin is an early muscle marker (Fontaine-Pérus et al., 1995;Sze et al., 1995). The timing of migration is well-documented
2689Development 126, 2689-2701 (1999)Printed in Great Britain © The Company of Biologists Limited 1999DEV1366
In myoblast cell cultures, the Msx1 protein is able to repressmyogenesis and maintain cells in an undifferentiated andproliferative state. However, there has been no evidencethat Msx1 is expressed in muscle or its precursors in vivo.Using mice with the nlacZ gene integrated into the Msx1locus, we show that the reporter gene is expressed in thelateral dermomyotome of brachial and thoracic somites.Cells from this region will subsequently contribute toforelimb and intercostal muscles. Using Pax3 genetranscripts as a marker of limb muscle progenitor cells asthey migrate from the somites, we have defined preciselythe somitic origin and timing of cell migration from somitesto limb buds in the mouse. Differences in the timing ofmigration between chick and mouse are discussed. Somitesthat label for Msx1nlacZ transgene expression in theforelimb region partially overlap with those that contributePax3-expressing cells to the forelimb. In order to seewhether Msx1 is expressed in this migrating population, wehave grafted somites from the forelimb level of Msx1nlacZ
mouse embryos into a chick host embryo. We show that
most cells migrating into the wing field express theMsx1nlacZ transgene, together with Pax3. In theseexperiments, Msx1 expression in the somite depends on theaxial position of the graft. Wing mesenchyme is capable ofinducing Msx1 transcription in somites that normallywould not express the gene; chick hindlimb mesenchyme,while permissive for this expression, does not induce it.In the mouse limb bud, the Msx1nlacZ transgene isdownregulated prior to the activation of the Myf5 gene, anearly marker of myogenic differentiation. Theseobservations are consistent with the proposal that Msx1 isinvolved in the repression of muscle differentiation in thelateral half of the somite and in limb muscle progenitorcells during their migration.
Key words: Mouse, Homeodomain protein, Msx genes, Homologousrecombination, Heterospecific grafts, β-galactosidase,Differentiation, In situ hybridisation, Muscle precursor cells, Pax3,Pax-3, Msx-1, Msx2, Msx-2
SUMMARY
The homeobox gene Msx1 is expressed in a subset of somites, and in muscle
progenitor cells migrating into the forelimb
Denis Houzelstein1,‡,*, Gwenola Auda-Boucher2,*, Yvonnick Chéraud2,*, Thierry Rouaud2, Isabelle Blanc1,Shahragim Tajbakhsh3, Margaret E. Buckingham3, Josiane Fontaine-Pérus2 and Benoît Robert1,§
1Laboratoire de Génétique Moléculaire de la Morphogenèse, CNRS URA 1947, Département de Biologie Moléculaire, InstitutPasteur, 25-28 rue du Docteur Roux, 75724 Paris Cedex 15, France2CNRS EP 1593, Faculté des Sciences et Techniques, 44322 Nantes Cedex 03, France3Unité de Génétique Moléculaire du Développement, CNRS URA 1947, Département de Biologie Moléculaire, Institut Pasteur, 25-28 rue du Docteur Roux, 75724 Paris Cedex 15, France*D. Houzelstein, G. Auda-Boucher and Y. Chéraud contributed equally to this work‡Present address: Division of Mammalian Development, National Institute for Medical Research, The Ridgeway, Mill Hill, London NW7 1AA, UK§Author for correspondence (e-mail: [email protected])
Accepted 8 April; published on WWW 19 May 1999

2690
in chick, where invasion of the wing bud is initiated at aboutthe 20-somite stage and is complete at the 36-somite stage(Chevallier et al., 1977; Christ et al., 1977; Solursh et al.,1987). In mammals, information is less precise. In mouse,migratory muscle cells reach the forelimb bud from about the21-somite stage (Sze et al., 1995; see also Milaire, 1987) andby about the 35-37-somite stage for the hindlimb (Tajbakhshand Buckingham, 1994).
Transcripts of the Pax3 gene have been shown to mark cellsmigrating from somites to limbs in avian embryos (Williamsand Ordahl, 1994). In splotch (Pax3−/−) mutant mice, cellsfrom the lateral dermomyotome do not migrate, limbmusculature is absent, and other hypaxial skeletal muscles areaffected (Franz et al., 1993; Bober et al., 1994; Goulding etal., 1994; Daston et al., 1996). Recent results suggest thatPax3 may act through the c-met receptor, which is alsoexpressed by this cell population and is required for migration(Bladt et al., 1995; Epstein et al., 1996; Yang et al., 1996;Maina et al., 1996). Other markers of the somitically derivedcells in the limb bud are N-cadherin, and follistatin (Brand-Saberi et al., 1996).
During early stages of limb morphogenesis, distal ectodermis induced to differentiate into a transitory epithelial structure,the apical ectodermal ridge (AER). The action of the AER ismediated by diffusible growth factors and maintains theunderlying mesenchyme, referred to as the progress zone,undifferentiated and proliferative, permitting distal limboutgrowth (see Tickle, 1996 for review). As a result of thisdistal outgrowth of the limb bud, proximal cells are removedfrom the proliferative influence of the AER and allowed todifferentiate. When they reach the limb bud, muscle progenitorcells proliferate for at least 24 hours, before they activate themyogenic factor genes Myf5 (Tajbakhsh and Buckingham,1994), MyoD and then Myogenin (Sassoon et al., 1989) priorto differentiation. Growth factors present in the progress zoneprobably contribute to the repression of muscle differentiation(Robson and Hughes, 1996).
The homeobox-containing gene Msx1 (previously Hox-7),whose expression is regulated by the AER, is stronglyexpressed in the nascent limb bud when limb muscleprogenitor cells migrate from the somites (Robert et al., 1989;Hill et al., 1989; see Davidson, 1995). Forced expression ofthis gene blocks terminal differentiation of a myogenic cellline, resulting in a decrease in levels of MyoD (Song et al.,1992); a result subsequently extended to the closely relatedMsx2 gene, also expressed in the limb bud (Takahashi et al.,1996). Furthermore, experiments in which humanchromosomes were transfected into mouse 10T1/2 fibroblastcells showed that activation of the human MyoD gene isinhibited by an additional chromosomal fragment containingthe Msx1 locus. Indeed, Msx1 binds to the MyoD enhancerand it has therefore been suggested that it may directlyregulate its transcription (Woloshin et al., 1995). However,there has been no direct demonstration that muscle progenitorcells in vivo do actually express Msx1 at any stage; the strongexpression of Msx1 throughout the early limb mesenchymecomplicates this issue. Embryos with a null mutation in theMsx1 gene show no skeletal muscle phenotype (Satokata andMaas, 1994; Houzelstein et al., 1997). However Msx2 iscoexpressed with Msx1 at many sites during embryonicdevelopment, and there may therefore be redundancy between
these closely related proteins (Catron et al., 1996; Houzelsteinet al., 1997).
We have generated mice carrying a null mutation in theMsx1 gene, by insertion of the bacterial nlacZ reporter gene,encoding the β-galactosidase protein with a nuclearlocalisation signal, into its homeobox. The pattern ofexpression of β-galactosidase from the targeted Msx1nlacZ
allele closely mimics that of Msx1 mRNA previouslydescribed (Davidson, 1995) and marks cells that normallyexpress the Msx1 gene during development (Houzelstein et al.,1997). The resolution and sensitivity conferred by the nlacZreporter have permitted us to detect expression in the lateraldermomyotome of somites in the vicinity of forelimbs and inthe interlimb region. The timing and extent of this expressionon the anteroposterior axis have been compared to that ofPax3, a gene expressed in cells migrating to the forelimb andhindlimb buds. These results show that Msx1 is expressed ina subset of somites some of which contribute cells to limbmusculature. By grafting mouse somites in chick embryos, wehave been able to demonstrate that cells in the lateraldermomyotome which have activated Msx1 are muscleprogenitor cells which migrate into the wing field. Heterotopicgrafts show that the wing, but not the leg mesenchyme, iscapable of inducing Msx1 expression in mouse somites fromdifferent rostrocaudal levels and in cells migrating from them.This may be related to the expression of Bmp4, which ishighest in limb mesenchyme at the forelimb level at theappropriate time. The nlacZ reporter gene ceases to beexpressed in the mouse limb bud prior to Myf5 activation,consistent with a role for Msx1 in the negative regulation ofmyogenic differentiation.
MATERIALS AND METHODS
Mouse embryosIn Msx1 nlacZ-targeted embryos, the nlacZ reporter gene has beeninserted into the region encoding the third helix of the Msx1homeodomain, such that the nlacZ-coding sequences are in phase fortranslation. This allele was maintained on a C57BL/6J background.Typing of embryos was performed by PCR as previously described(Houzelstein et al., 1997). For Myf5 nlacZ-targeted embryos, thenlacZ reporter gene has been fused into the first exon of the Myf5gene, 13 codons after the ATG codon such that the nlacZ codingsequence is in phase for translation. The Myf5 gene itself is disrupted,and a small deletion is introduced. This allele was maintained on amixed (C57BL/6-129-DBA/2) background (Tajbakhsh et al., 1996a).
In this study, the first somite to express Pax3 or Myf5 at the anteriorof the embryo was counted as somite 3 (according to Tajbakhsh et al.,1996b; Spörle and Schughart, 1997). This locates the anterior marginof the forelimb between somites 9 and 10, and the posterior marginbetween somites 14 and 15. In this way, the somite that we arecounting as 13 was referred to as somite 11 by Sze et al. (1995) andas somite 12 by Burke et al. (1995).
In situ hybridisation and staining proceduresThe Msx1 probe corresponds to the 3′ untranslated region of themouse gene as previously described (Lyons et al., 1992). The Pax3(Goulding et al., 1991) probe is derived from a 519 bp PstI-HindIIIfragment from the 3′ coding end of the mouse cDNA (kindly providedby P. Gruss). To generate a mouse-specific Pax3 probe, twooligonucleotides, 5′-GGGGTAGTTCCTCCTGGAAG-3′ and 5′-TTCATGTCTAGTCTGTGGAGGC-3′, were used to amplify the 3′
D. Houzelstein and others

2691Msx1 in muscle progenitor cells
UTR region of the Pax3 gene directly from mouse C57BL/6 DNA.The amplification product (560 nucleotides in length) was clonedinto the pnoTA plasmid using the Prime PCR Cloner System(5prime→3prime, Inc.). The Bmp4 probe (a kind gift from B. Hogan)contains the 3′ UTR and most of the coding sequence from a mousecDNA (Jones et al., 1991). Whole-mount in situ hybridisation wasperformed according to Henrique et al. (1995) with modifications(post-antibody washes were carried out for 3 days and revelation waswith BM purple AP substrate (Boehringer), 1% Tween-20 for 24hours). In situ hybridisation on sections was performed according toMyat et al. (1996). X-gal staining on whole embryos and cryostatsections was as previously described (Houzelstein et al., 1997).Cryostat sections of mouse-chick chimaeras were stained with X-gal,washed twice in PBS before counterstaining with Hoechst (bis-benzimide Hoechst No. 33342, Sigma; saturated solution of Hoechstin ethanol diluted 1/1000 in PBS). They were subsequently washedtwice in PBS and mounted in Mowiol (Calbiochem). Salmon-gal(Apollo Scientific Ltd, UK) staining was performed following thesame protocol.
All pictures have been scanned and assembled using AdobePhotoshop.
Somite and neural tube transplantation Somite grafting was performed as previously described (Fontaine-Pérus et al., 1995) except that Msx1nlacZ mice were maintained on aC57BL/6J background. For homotopic grafting, donor somites wereexcised from 18- to 20-somite stage Msx1nlacZ mouse embryos.Somites 8-15 were retained to be transplanted into the 15- to 21-somite region of the chick host, a region previously described to giverise to myogenic progenitor cells of the wing muscles (Chevallier,1979; Zhi et al., 1996). Mouse somites were replaced withoutchanging their orientation. Heterotopic and heterochronic grafts wereperformed in the same way. In all series, chick embryos were fixedbetween 16 and 24 hours following surgery.
Neural tube grafting was performed as previously described(Fontaine-Pérus et al., 1997). Donor neural tubes were excised fromthe caudalmost part of 15- to 18-somite stage Msx1nlacZ mice. Hostswere fixed between 16 and 24 hours following surgery.
RESULTS
Origin and timing of the somitic contribution to thelimb budsUsing a probe specific for the Pax3 gene, which is expressedin limb muscle progenitor cells during their migration (Boberet al., 1994; Goulding et al., 1994; Williams and Ordahl, 1994),we examined which somites contribute muscle progenitor cellsto the limb and when this migration takes place in the mouseembryo, based on the detection of delaminating Pax3-positivecells.
Lateral dermomyotomal cells were first detected todelaminate in 24-somite embryos (about embryonic day {E}9.75) from somites 9-11, adjacent to the forelimb bud (data notshown). In 25-somite embryos, delaminated Pax3-positivecells are seen adjacent to somites 9-12 (Fig. 1a; arrowhead).The contribution to the limb bud gradually extends morecaudally to include somite 14, in keeping with theanteroposterior gradient of somite maturation. Migration ismost intense in 29-somite embryos (Fig. 1b). In 32-somiteembryos, fewer cells are delaminating adjacent to the forelimbbud and somite 9 no longer contributes; by the 33-somite stageonly a few cells continue to delaminate from somite 14 (datanot shown). At the 38-somite stage forelimb premuscle masses
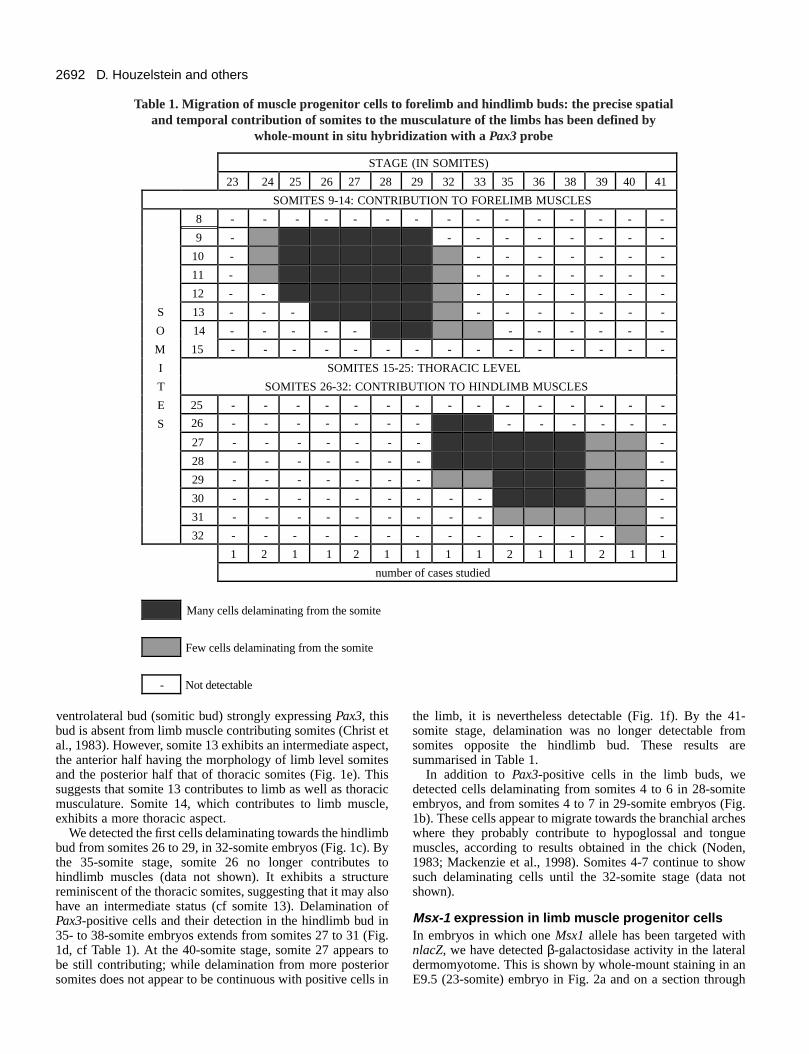
are clearly isolated from the adjacent somites (Fig. 1e).Therefore, Pax3-positive cells delaminating from somites 9 to14 are detected in the forelimb bud field in 24- to 33-somiteembryos (results summarised in Table 1).
Somites contributing to limb musculature exhibit a typicalaspect after completion of migration, when compared tosomites at the thoracic level. While thoracic somites exhibit a
Fig. 1. Migration of muscle progenitor cells into the forelimb andhindlimb buds as revealed by a Pax3 probe on whole mounts. (a) 25-somite embryo. The black arrowhead indicates muscle progenitorcells that have left somites 9-12 at the level of the forelimb bud. Atthis stage, no Pax3-expressing cells are detectable delaminating fromsomites at the level of the hindlimb field, nor more anteriorly, at thelevel of occipital somites 4-7. (b) 29-somite embryo. Pax3-expressing cells are visible within the proximal part of the forelimbbud (fl), adjacent to somites 9-14. The black arrowhead indicatescells that have left occipital somites 4-7, and which will subsequentlycontribute to tongue and hypoglossal muscles. Somites 9 and 13 areindicated. (c) 32-somite embryo. Cells have begun to delaminatefrom somites 26-29 at the level of the hindlimb bud (hl). Somites 26and 31 are indicated. (d) 36-somite embryo. Many cells havedelaminated from the adjacent somites to locate within the proximalpart of the hindlimb bud. Somite 31 is indicated. (e) 38-somiteembryo. The forelimb premuscle mass is now clearly separated fromthe adjacent somites indicating that Pax3-expressing cells haveceased to migrate into the forelimb bud (fl). In the lateral domain,somite 13 exhibits a posterior domain with the morphology ofthoracic somites, characterised by a ventral bud, and an anteriordomain typical of limb-muscle-contributing somites, which lack theventral bud. Many Pax3-positive cells can be seen immediatelyadjacent to somites 27-30 in the hindlimb bud (hl). Somites 13, 26and 31 are indicated. (f) 40-somite embryo. At the level of thehindlimb bud (hl), the premuscle mass is now also distinct from thesomites, where a few Pax3-positive cells still appear to bedelaminating into the limb field, particularly from somites 27, 31 and32. Somite 26, although contributing to the hindlimb musculature,exhibits a structure characteristic of flank somites. Somites 26 and 31are indicated. Scale bars, 500 µm.
2692
ventrolateral bud (somitic bud) strongly expressing Pax3, thisbud is absent from limb muscle contributing somites (Christ etal., 1983). However, somite 13 exhibits an intermediate aspect,the anterior half having the morphology of limb level somitesand the posterior half that of thoracic somites (Fig. 1e). Thissuggests that somite 13 contributes to limb as well as thoracicmusculature. Somite 14, which contributes to limb muscle,exhibits a more thoracic aspect.
We detected the first cells delaminating towards the hindlimbbud from somites 26 to 29, in 32-somite embryos (Fig. 1c). Bythe 35-somite stage, somite 26 no longer contributes tohindlimb muscles (data not shown). It exhibits a structurereminiscent of the thoracic somites, suggesting that it may alsohave an intermediate status (cf somite 13). Delamination ofPax3-positive cells and their detection in the hindlimb bud in35- to 38-somite embryos extends from somites 27 to 31 (Fig.1d, cf Table 1). At the 40-somite stage, somite 27 appears tobe still contributing; while delamination from more posteriorsomites does not appear to be continuous with positive cells in
the limb, it is nevertheless detectable (Fig. 1f). By the 41-somite stage, delamination was no longer detectable fromsomites opposite the hindlimb bud. These results aresummarised in Table 1.
In addition to Pax3-positive cells in the limb buds, wedetected cells delaminating from somites 4 to 6 in 28-somiteembryos, and from somites 4 to 7 in 29-somite embryos (Fig.1b). These cells appear to migrate towards the branchial archeswhere they probably contribute to hypoglossal and tonguemuscles, according to results obtained in the chick (Noden,1983; Mackenzie et al., 1998). Somites 4-7 continue to showsuch delaminating cells until the 32-somite stage (data notshown).
Msx-1 expression in limb muscle progenitor cellsIn embryos in which one Msx1 allele has been targeted withnlacZ, we have detected β-galactosidase activity in the lateraldermomyotome. This is shown by whole-mount staining in anE9.5 (23-somite) embryo in Fig. 2a and on a section through
D. Houzelstein and others
STAGE (IN SOMITES)
23 24 25 26 27 28 29 32 33 35 36 38 39 40 41
SOMITES 9-14: CONTRIBUTION TO FORELIMB MUSCLES
8 - - - - - - - - - - - - - - -
9 - - - - - - - - -
10 - - - - - - - -
11 - - - - - - - -
12 - - - - - - - - -
S 13 - - - - - - - - - -
O 14 - - - - - - - - - - -
M 15 - - - - - - - - - - - - - - -
I SOMITES 15-25: THORACIC LEVEL
T SOMITES 26-32: CONTRIBUTION TO HINDLIMB MUSCLES
E 25 - - - - - - - - - - - - - - -
S - - - - - -
27 - - - - - - - -
28 - - - - - - - -
29 - - - - - - - -
30 - - - - - - - - - -
31 - - - - - - - - - -
32 - - - - - - - - - - - - - -
1 2 1 1 2 1 1 1 1 2 1 1 2 1 1
number of cases studied
Many cells delaminating from the somite
Few cells delaminating from the somite
- Not detectable
Table 1. Migration of muscle progenitor cells to forelimb and hindlimb buds: the precise spatialand temporal contribution of somites to the musculature of the limbs has been defined by
whole-mount in situ hybridization with a Pax3 probe
26 - - - - - - -

2693Msx1 in muscle progenitor cells
a somite in Fig. 2b, at the level of the forelimb bud. Aspreviously reported for the endogenous gene, there is stronglabelling of the neural tube, lateral mesoderm and limb buds,at this stage. This labelling of the lateral dermomyotome is firstseen in 22-somite embryos, in somites 11-15. By the 25-somitestage, labelling in heterozygous embryos is detectable insomites 13-16. In 32-somite embryos, it is no longer seen(summarised in Table 2).
In homozygous embryos, where both Msx1 alleles aretargeted with nlacZ, more extensive expression is detectablein somites along the anteroposterior axis (Fig. 2c), where it isconcentrated in cells of the lateral dermomyotome (Fig. 2d).In the dermomyotome of these somites, the expression domain
of Msx1 overlaps with the lateral domain of Pax3 expression(Fig. 2d; see also Fig. 3c). Low-level expression is seen insomites in the interlimb region from the 25-somite stage,extending to somite 26 in 30-somite embryos. By the 32-somite stage, β-galactosidase-positive cells are no longerdetectable in most somites, but persist in somites 25-26 untilthe 32-somite stage.
These results indicate that Msx1 is a marker of the lateraldermomyotome of somites, in the forelimb and interlimbregion. Msx1nlacZ does not appear to be expressed in somitesthat contribute the muscle progenitor cell population of thehindlimb, with the possible exception of somite 26, which,as previously noted, has a mixed thoracic/limb morphology;nor is Msx1nlacZ expression detectable more anteriorly insomites that will contribute migratory cells to the branchialarches. At the forelimb level, Msx1 expression is maximal insomites 12-16, while somites 9-14 are the main contributorsof Pax3-positive cells. The onset of Msx1nlacZ expressionbegins at stage 22, prior to delamination of Pax3-positivecells, and is not detectable in somite 9, which alsocontributes Pax3-positive cells to the forelimb bud (cf Tables1 and 2). In contrast, β-galactosidase activity derived fromthe Msx1nlacZ allele is no longer detectable at somite stage32 from somites opposite the forelimb in embryos with bothalleles targeted, when Pax3-positive cells are stilldelaminating.
In the avian embryo, it is well demonstrated that thedorsolateral zone of the brachial somites provides themyogenic progenitors of the forelimbs, a demonstration relyinglargely on exchange of somites between quail and chick(Reviewed in Hauschka, 1994; Christ and Ordahl, 1995). Inmammals, experiments in which labelled somites weretransplanted into cultured embryos also pointed to myogenicprogenitor migration from somites to the limb (Beddington andMartin, 1989; Lee and Sze, 1993; Sze et al., 1995). In theMsx1nlacZ mice, at the stage when limb muscle progenitor cellsmigrate from the somites, limb buds strongly express β-galactosidase from the nlacZ-targeted Msx1 allele throughoutmost if not all the mesenchyme (Fig. 2a,b). It is therefore verydifficult to assess against this background whether muscleprogenitor cells express the Msx1 gene. To address this issue,murine somites, which would normally contribute to forelimbmuscles, were grafted in place of chick somites, which wouldnormally contribute to wing muscles, prior to the onset ofmigration. Surgery was performed on 27 embryos in which thelast 4-6 somites from the brachial level of 18- to 20-somitechick hosts were replaced unilaterally with somites 8-15 fromE9 mouse embryos heterozygous or homozygous for theMsx1nlacZ allele. In all specimens examined 6 hours aftergrafting (10 cases), β-galactosidase-positive cells weredetected in the dorsolateral border of the implanted somites(Fig. 3a-c). β-galactosidase was detected only in somites 11-13 when somites 8-13 were grafted, implying that 6 hours aftergrafting the expression of Msx1nlacZ in the grafted somites stillcorresponds to that seen in the mouse. 12 hours after grafting(9 cases), labelled cells were observed located outside thesomites in the proximal wing field (Fig. 3d,e). In slightly olderchimaeric embryos (5 cases), they were all present within thewing field (not shown).
To further characterise the mouse somitic cells that expressMsx1, we investigated whether they also express Pax3. For this
STAGE (IN SOMITES)
21 22 25 27 30 32 34
10 - - - - - -11 - - -12 - - -13 - - -14 - - -15 - - -16 - - - -
S 17 - - - -O 18 - - - -M 19 - - - -I 20 - - - -T 21 - - - -E 22 - - - -S 23 - - - -
24 - - - -25 - - -26 - - - -27 - - - - - - -28 - - - - - - - 29 - - - - - - -30 - - - - - - -
1 2 4 4 2 1 2
number of cases studied
Strong expression in homo- and heterozygous embryos
Weak expression in homo- and heterozygous embryos
Strong expression in homozygous embryos only
Weak expression in homozygous embryos only
- Not detectable
Table 2. Spatial and temporal expression of the Msx1-nlacZgene in the lateral dermomyotome of the somite
during development

2694
purpose, we prepared a mouse Pax3 probe entirely derived fromthe 3′ UTR of the gene. This probe shows very little crosshybridisation with the chick Pax3 mRNA as deduced from thevery faint staining of the dorsolateral region of the chickdermomyotome (see Fig. 3b, star). Somites from the wing levelof chick hosts were replaced by brachial somites from Msx1nlacZ
heterozygous mice and the expression patterns of the mousePax3 gene and of β-galactosidase were simultaneously analysed(13 experiments). Sections of such doubly stained chimaericembryos illustrated that cells from the dorsolateral region of thedermomyotome co-accumulate β-galactosidase activity andPax3 transcripts, whereas Pax3 transcripts only are detectedmedially (Fig. 3b,c). The dorsolateral area is the source of themyogenic progenitors in the limbs, characterised by Pax3expression, and our results demonstrate that these cells alsoexpress Msx1.
Mouse somite cells were further analysed for theirproperties after they had migrated into the wing field. Mousecell nuclei can be readily distinguished from chick by bis-benzimide staining. 24 hours after grafting of mouse somites,mouse cells can be observed migrating out of the somite inthe proximal area of the wing bud (Fig. 3g). Simultaneoustreatments for β-galactosidase and nuclear type detectionrevealed that ≥90% of mouse cells leaving the somite are β-galactosidase-positive, indicating that most cells that escapefrom the grafted somites express Msx1nlacZ (not shown). Sincemost cells leaving this region of the somite are muscleprogenitors, our results suggest that Msx1 is expressed by thegreat majority of myogenic cells that will form limb muscles.This was assessed by double labelling of these cells for Pax3transcripts and β-galactosidase. For this purpose, one sectionfrom a chimaeric embryo was treated with salmon-gal toreveal β-galactosidase activity. After photography, the samesection was hybridised with the mouse-specific Pax3 probeand rephotographed in exactly the same orientation (Fig. 3h,i).Comparison of the two images demonstrates that the greatmajority of Pax3-expressing cells also express β-galactosidase(Table 3). Occasionally, cells expressing only Pax3 wereobserved (Fig. 3h,i, arrow).
Since neural crest cells traverse the somite, they may becarried over with the graft and contribute migrating cells tothe chick host. Previous work has shown that activeemigration of neural crest cells from mouse neural tubeoccurs after in ovo grafting (Fontaine-Pérus et al., 1997).Under these conditions, neural crest cells from the implantcontribute to the development of the host peripheral nervoussystem and to the melanoblast population. These cells mayalso express Pax3 at certain levels along the rostrocaudal axis(Goulding et al., 1991; Natoli et al., 1997). The major routesfollowed by neural crest cells do not traverse the wing bud(Fontaine-Pérus et al., 1997). However, to rule out thepossibility that the cells expressing Msx1nlacZ in the somiteand proximal wing field are related to the neural crestpopulation, a fragment of mouse neural tube was grafted inplace of part of the chick neural tube, adjacent to a levelcaudal in the segmental plate to the youngest newly formedsomite in 15 to 18-somite-stage chick embryos. A total of 10cases were examined between 16 and 24 hours followingsurgery. At 16 hours post surgery, neural crest cells had leftthe mouse neural tube. At this time, Msx1nlacZ transgeneexpression was restricted to the dorsal area of the graftedmouse neural tube. 24 hours post surgery, the mouse crestcells compacted and constituted the early spinal ganglionicprimordia; at this stage enzyme activity was still restricted tothe neural tube area. In one chimaera, in which in situhybridisation with the Pax3 probe and β-galactosidasestaining were carried out simultaneously, co-expression wasseen in the dorsal aspect of the grafted neural tube (Fig. 3f).No staining was observed in the wing bud, demonstrating thatmouse neural crest cells do not contribute to the Msx1-expressing population detected in the proximal wing bud aftersomite grafting.
Mouse somites can be induced to express Msx1 byectopic graftingChick/quail transplantation experiments have demonstratedthat unsegmented paraxial mesoderm or somites, not derivedfrom the forelimb region of the mouse embryo, are
D. Houzelstein and others
Fig. 2. Expression of the Msx1nlacZ gene in the lateraldermomyotome of somites. (a) An E9.5 (23 somites)Msx1nlacZ/+ embryo, with β-galactosidase activity revealedon whole mount. The position of somite 13 is indicated. TheMsx1nlacZ gene is expressed in the lateral dermomyotome ofsomites 12-15, and more weakly in somite 11. (b) An E9.5Msx1nlacZ/+ embryo. This embryo has been whole-mountstained for β-galactosidase, embedded in paraffin andsectioned. A section at forelimb (fl) level is shown, theMsx1nlacZ gene is expressed in the lateral dermomyotome.The forelimb bud is strongly labelled. (c) An E9.75 (30somites) Msx1nlacZ/nlacZ embryo. This embryo has beenwhole-mount stained for β-galactosidase. The position ofsomite 13 is indicated. β-galactosidase activity is detectedwith variable intensities in the lateral domain of somites 10-26. (d) E9.75 Msx1nlacZ/nlacZ embryo cryostat sectioned atthe level of the forelimb (fl) and stained for β-galactosidaseactivity. Strong staining in the forelimb and also in the lateraldermomyotome is seen. The inset shows an adjacent sectionhybridised with the Pax3 probe. Pax3 and Msx1nlacZ
expression domains are superimposed in the lateral region ofthe dermomyotome. Scale bar (a,c) 250 µm; (b,d) 100 µm.

2695Msx1 in muscle progenitor cells
reprogrammed to contribute migratory limb cells when placedopposite the chick wing field (Chevallier, 1979). To assesswhether such reprogramming also takes place for Msx1expression, heterotopic and heterochronic grafts wereperformed. In the mouse, the anterior-most somites neverexpress Msx1 (see Table 2). When such somites (somite 1-5level) from E8.5 Msx1nlacZ heterozygous mice were implantedinto the chick at the brachial level, β-galactosidase expressionwas observed laterally to the grafted somites (Fig. 4a) (4 casesanalysed).
Somites from the wing level of 15- to 18-somite chick hostswere unilaterally replaced by the unsegmented paraxialmesoderm from 18- to 20-somite Msx1nlacZ heterozygousmouse embryos (18 cases). The unsegmented paraxialmesoderm does not express Msx1 in the mouse. In theseexperiments, it was derived from an area that would later formthoracic level somites, which do not express the Msx1 gene ata high level. Histological analysis showed that the murinesegmental plate was able to become segmented in ovo (notshown). After 18 hours of incubation, β-galactosidase-positivecells in the newly formed murine somites were clearly
detectable, although migration had not yet been initiated insuch less advanced embryos (Fig. 4b).
Conversely, somites from the forelimb region of a mouseembryo were grafted slightly anterior to the chick wing field,such that only the posteriormost somite of the graft replaced achick somite that would contribute muscle progenitor cells tothe wing bud (Fig. 4c). Cells migrating from this somiteexpressed the Msx1nlacZ gene, whereas the more anterior mousesomites did not provide β-galactosidase-positive cells.Noticeably, somites adjacent to the anterior area of the limbfield in the mouse do not express Msx1 in the lateral part of thedermomyotome (see Table 2), although it has not beendemonstrated whether muscle precursors migrating from thesesomites express the gene. Our results show that signals fromthe wing somatopleure (Hayashi and Ozawa, 1995) are able toinduce strong Msx1nlacZ expression in the lateral margin of thedermomyotome and in cells migrating from it, but suggest thatthe distribution of the inducing activity may be different inchick and mouse.
To better define the inductive properties of the limbmesenchyme, mouse somites from the hindlimb level, which
Fig. 3. Expression of the Msx1nlacZ gene in limb muscleprecursors from mouse somites grafted into chick hosts. (a) A20-somite chick embryo examined 6 hours after grafting ofsomites 8-13 from an E9 Msx1nlacZ mouse, to replace chicksomites at the wing level. Cells from the lateral border ofgrafted somites 11-13 express β-galactosidase. The limits ofthe graft are indicated (white bars). (b) Transverse section of achick embryo into which somites 9-13 of an E9 Msx1nlacZ
mouse have been implanted at the 18-somite stage at thebrachial level. 6 hours post surgery, Msx1nlacZ expression wasrevealed on the operated side by salmon-gal (red) staining andmouse Pax3 expression by in situ hybridisation (bluestaining). Note the faint cross-hybridisation of the mouse-specific Pax3 probe with chick transcripts in the chickdermomyotome (*). (c) Higher magnification of the operatedside in b. The dorsolateral zone of the mouse dermomyotomeco-expresses the Pax3 and Msx1nlacZ genes (blue and redlabelling are superimposed – arrowhead) whereas themediodorsal region only accumulates Pax3 transcripts (bluelabelling only – arrow). (d) Expression of the Msx1nlacZ genein a chick host embryo examined 12 hours after implantationof somites 8-13 from an E9 mutant mouse at the wing level.A mass of β-galactosidase-positive cells identified by X-galblue staining is detected outside the grafted somite.(e) Cryostat section from an embryo similar to that shown ind. The β-galactosidase-positive cells revealed by salmon-gal(red staining) appear just outside the grafted somites in theproximal wing (w) field. (f) Section at the wing bud levelfrom a mouse-chick chimaera where part of the neural tubewas replaced by a fragment of neural tube from an Msx1nlacZ
heterozygous mouse at the 18-somite stage, adjacent to thesegmental plate. Pax3 expression was revealed by in situhybridisation with a mouse-specific probe (pink staining) andMsx1nlacZ by X-gal (blue) staining of β-galactosidase activityon the day following surgery. X-gal staining is restricted to the upper part of the grafted neural tube (nt), which also expresses Pax3 in a broaderdomain. No β-galactosidase-positive (blue) cells can be detected in the wing field (w). The myotome (m) is revealed by in situ hybridisation with achick MyoD probe (also as pink staining). nc, notochord. (g) Transverse section of a chick embryo implanted at the 18-somite stage at the brachiallevel with somites 9-13 of an E9 Msx1nlacZ mouse, stained with bis-benzimide 24 hours post-surgery. Mouse cells in the somite (s) and in the wingfield (w) appear brighter, as do the compacted cells of the chick neural tube (nt). (h,i) Adjacent section to that in g, corresponding to the boxedregion. This section was first treated with salmon-gal to reveal β-galactosidase activity and, after photography (h), the same section was hybridisedwith the mouse Pax3 probe and re-photographed (i). Comparison of h and i clearly shows that most mouse cells are co-labelled. Occasional cellsexpress only Pax3 (arrow). Scale bars, (a,d) 500 µm, (f,g) 100 µm, (b,c,e) 50 µm, (h,i) 25 µm.

2696
never express Msx1, were grafted either at the forelimb(wing) or at the hindlimb (leg) level in a chick host. This wasthe first attempt to graft mouse somites at a posterior level inchick embryos older than the 20-somite stage; all grafts weresuccessful. In accordance with experiments described above,mouse somites from the hindlimb level, when grafted to thewing level in the chick, were reprogrammed to expressMsx1nlacZ and provide β-galactosidase-positive cells to thewing mesenchyme (Fig. 4d) (7 cases). In contrast, whengrafted at the leg level (somite level 26-32), these somites andthe cells migrating from them never expressed Msx1 (4 casesanalysed) (data not shown). This shows that, in the chick, theinductive capacity resides in forelimb, not hindlimb territory,in accordance with the endogenous pattern of expressionobserved for Msx1 in the mouse somites. However, whenmouse somites from the brachial level were transplanted,before detectable Msx1nlacZ expression, to the leg level of achick host, β-galactosidase-positive cells were observed inthe lateral margin of these somites (10/11 cases) andmigrating from them (4/11 cases) (Fig. 4e). Therefore,induction of a potential for Msx1 expression must take placebefore expression itself is detectable and the leg mesenchymethen constitutes a permissive region for Msx1 expression, asopposed to the neck mesenchyme (Fig. 4c). Cells migratingto the hindlimb were confirmed to be muscle precursors byhybridisation with the mouse-specific Pax3 probe (notshown).
These results show that signals emanating from the wingsomatopleure are able to induce Msx1nlacZ expression in thelateral margin of the dermomyotome of grafted somites and incells migrating from them into the wing field, which would nothave expressed the gene in the mouse embryo. BMP4 has beenshown to induce Msx1 expression in mouse dentalmesenchyme (Vainio et al., 1993), and is expressed in the limbbud and lateral plate mesenchyme (Pourquié et al., 1996). Toinvestigate whether BMP4 might be the Msx1 inducer in thelateral margin of the dermomyotome as well, we performed insitu hybridisation with a Bmp4 probe on mouse embryos at the20- to 25-somite stages (Fig. 4f). This shows that Bmp4expression is highest in the limb mesenchyme facing somites11-15, which correspond to those somites in which Msx1expression is maximal by the 22-somite stage (Table 2). Thesame pattern has been observed at slightly earlier stages byHogan et al. (1994). This is therefore consistent with theproposition that BMP4 induces Msx1 expression in the lateraldermomyotome of these somites.
Msx-1 exclusion from differentiated musclesExpression of the myogenic factor gene Myf5 serves as an
early marker of myogenic cells (Ott et al., 1991; Tajbakhshand Buckingham, 1994). We have used Myf5nlacZ-targetedmice (Tajbakhsh et al., 1996a) to detect the onset ofmyogenesis in the limb. At the 32-somite stage, when most ofthe muscle progenitor cells are already present in the proximalforelimb (see Table 1), the whole bud expresses the Msx1nlacZ
gene (data not shown). We have generated mice heterozygousfor both Msx1nlacZ and Myf5nlacZ. In such a doubleheterozygous embryo shown in Fig. 5a (35 somites), the β-galactosidase-positive cells of the progress zone, expressingthe Msx1nlacZ allele, are already restricted to the distal domainof the forelimb bud. Cells in the proximal part of the limb,located more than 300 µm away from the AER, no longerexpress the Msx1nlacZ gene. It is in this proximal area thatmyogenesis will be initiated as shown by the activation of theMyf5nlacZ allele from the 37-somite stage (Tajbakhsh andBuckingham, 1994). At the same stage (39 somites) shown inFig. 5d, Msx1 is not expressed in this proximal area (Fig. 5c).The Msx1nlacZ gene is downregulated proximally prior to anysign of Myf5nlacZ activation in the forelimb bud (Fig. 5a,b).The delay between Msx1nlacZ downregulation and thedetection of Myf5nlacZ expression is about 5 hours in theC57BL/6 mouse strain. These observations point to exclusionbetween the expression of Msx1 and that of the earlymyogenic factor gene Myf5.
DISCUSSION
The results reported here document the expression of Msx1 inthe lateral dermomyotome of a subset of somites, whichinclude most of those that will contribute migratory musclecells to the forelimb, as defined here by the presence of Pax3transcripts. Orthotopic grafting experiments show that somitesfrom Msx1nlacZ mice contribute β-galactosidase-positive cellsto the chick wing field. The fact that most of the migratorymouse cells are labelled strongly suggests that these includemuscle progenitor cells, rather than representing a smallpopulation of endothelial progenitors that are also somite-derived, or of mouse neural crest cells co-transplanted with thesomite (Beddington and Martin, 1989; Wilting et al., 1995).Double labelling for Msx1nlacZ and Pax3 expression confirmsthat the great majority of these cells are muscle precursors.Direct comparison of Msx1nlacZ expression with that ofMyf5nlacZ demonstrates that the former is downregulated in thelimb bud before expression of this first myogenic factor gene.These results are consistent with a role for the Msx1 gene inthe initial repression of myogenesis in muscle progenitor cellsin the limb.
Origin and timing of limb muscle progenitor cellmigration in the mouse; comparison with chickWe have used the expression of Pax3 as a marker of cells thatmigrate from the lateral dermomyotome into the limb buds, inorder to define which somites contribute this cell populationand when migration occurs. Pax3−/− (splotch) mutant mice lacklimb musculature, clearly indicating that expression of thisgene marks muscle progenitor cells, which indeed make up thebulk of the migratory population (see Christ and Ordahl, 1995).It is not clear whether Pax3 is also expressed by other somite-derived migratory cells. However, the apparent absence of an
D. Houzelstein and others
Table 3. The proportion of Msx1-expressing cells amonglimb muscle precursors, scored as Pax3 positive, for a
representative sample of limb bud sections fromchick/mouse chimeras with grafted Msx1nlacZ mouse
somites (see text)β-galactosidase-
Sections Pax3-expressing cells positive cells
A 52 48 B 51 49C 26 25D 24 23

2697Msx1 in muscle progenitor cells
endothelial phenotype in the limbs of splotch mice suggeststhat Pax3 may not be a marker of this cell type. In the regionof the forelimb bud, we detect Pax3-positive cells delaminatingfrom somites 9-14 between somitic stages 24-33(approximately 12-15 hours in the C57BL/6J background). Theonset of migration is similar to that deduced previously (21-somite stage which corresponds to our 23-somite stage – seeMaterial and Methods) (Sze et al., 1995; see also Milaire,1987). Interestingly, we detect the onset of delamination ofcells from more anterior somites (4-7) in the field of thebranchial arches rather later, in 28- to 29-somite embryos,continuing to the 32-somite stage. In quail/chick chimaeras,Mackenzie et al. (1998) have shown that myogenic cellsmigrating from anterior somites to colonise the hypoglossalmusculature do express Pax3. In splotch mutants, muscleprogenitor cells which contribute to the hypoglossal chord andto some throat and tongue muscles are absent (Tajbakhsh et al.,1997). Thus, we conclude that Pax3-positive cells delaminatingfrom the anterior somites correspond in part to this population.On the basis of experiments with hindlimb bud explants wherethe appearance of Myf5-expressing myogenic cells wasmonitored, we had previously deduced that migratory cells hadleft the somite by the 35- to 37-somite stage (Tajbakhsh andBuckingham, 1994). Consistent with this, we show here thatsomites 26-32 contribute Pax3-expressing cells to thehindlimbs in 32- to 40-somite embryos.
Somites that contribute migratory cells to the limbs have adistinct lateral morphology; they lack the ventral somitic bud,clearly labelled by the Pax3 probe, which is typical of interlimbsomites. Interestingly, somites 13 and 26 have an intermediatemorphology suggesting differences in specification betweenrostral and caudal halves of the lateral dermomyotome. Suchrostral/caudal differences have been reported for somitic neuralcrest colonisation (reviewed in Christ and Ordahl, 1995) ormotor nerve innervation (Keynes and Stern, 1984).Furthermore each hemivertebra is composed of two adjacenthemisclerotomes, the caudal half of the rostral one and therostral half of the caudal one (Aoyama and Asamoto, 1988).Our results suggest that the anterior half of somite 13contributes to muscles of the forelimb bud, while muscleprogenitor cells from the posterior half of this somite formintercostal and body wall muscles. This resembles observationson chick/quail chimaeras where it has been shown that somite20 contributes both to limb and intercostal muscles (Zhi et al.,1996).
In comparing the data reported here on the somitecontribution to limb muscle with those for chicken, it isnecessary to take into account differences in axial formulae.While mice have 7 cervical and 13 thoracic vertebrae, chickenhave 14 cervical and 7 thoracic vertebrae, with theconsequence that wing buds in chick form at the 15- to 20-somite level (Burke et al., 1995). In both species, the first limbmuscle progenitor cells leave the somites at about the samestage (22-24 somites in chick) (Christ et al., 1977; Jacob etal., 1978; Solursh et al., 1987). However, due to their differentposition along the anterior-posterior axis, the chick somitesare less mature. If the most recently formed somite is countedas I (see Christ and Ordahl, 1995), this means that wingmuscle progenitor cells begin to migrate from stage VIIsomites in chick, compared to stage XIV-XVI somites for themouse forelimb buds. Chicken and mouse hindlimbs are at
approximately the same somite level. In both cases, migrationtakes place from relatively less mature somites; in mouse fromstage IV-VII somites and in chicken, even more strikingly,from the most recently formed somites (I-III), even before adistinct dermomyotome is formed (Jacob et al., 1979). Thesedifferences in timing indicate that the initiation ofdelamination and onset of cell migration are not an intrinsicproperty of somites, but may be induced at different stages oftheir maturation, by external factors. Indeed in chick, it hasbeen shown that the migration of muscle progenitor cells isinduced by contacts between the medial region of the limbfield and the lateral margin of somites at the wing level(Hayashi and Ozawa, 1995). The delay in migration of mouselimb muscle precursors towards the limb bud, compared tochick, may be due to a difference in the timing of the lateralrotation of the embryo, which brings limb presumptivemesoderm into contact with the lateral margin of the somiteas shown in chick (Solursh et al., 1987). As a result, in birds,the wing field is still immature when migration begins and theapical ectodermal ridge (AER), which is important indetermining the direction of migration, is not present (seeHauschka, 1994), whereas in mice the forelimb bud alreadyhas a well-developed AER.
Restriction of Msx1 expression to a subgroup ofsomites along the anteroposterior axisWe detect expression of the targeted Msx1nlacZ allele in thelateral part of the dermomyotome, where it coincides withPax3 expression. Recently, somite expression has beenreported for Msx1-nlacZ transgenes, and then detected by insitu hybridisation for the endogenous gene (Mackenzie et al.,1997; Pereira et al., 1999). A striking feature of theexpression of Msx1, which we only detect in the lateral partof the dermomyotome, is that it is not uniform in all somiteson the rostrocaudal axis, in contrast to Pax3. β-galactosidaselabelling from the targeted Msx1nlacZ allele is particularlystrong in somites 11-14 which contribute Pax3-positive cellsto the forelimb. Labelling in somites 9 and 10, which alsocontribute migratory muscle progenitor cells, is very weak(10) or not detectable (9), indicating that Msx1 expressionmay mark a subpopulation of these cells. Within the limb,different somites will contribute to more or less anteriormuscle masses (see Hauschka, 1994), indicating that Msx1-positive muscle progenitor cells may preferentially colonisea subset of muscles. With the exception of somite 26, whereweak β-galactosidase labelling is detectable, Msx1 is notexpressed in the lateral dermomyotome at the level of thehindlimb. Differences in gene expression between forelimband hindlimb have been reported notably for the Tbx4 andTbx5 genes; Tbx4 is predominantly expressed in theembryonic hindlimb, and Tbx5 in the forelimb. However,these genes are not normally expressed in the interlimbregion (Chapman et al., 1996; Gibson-Brown et al., 1996).In contrast, β-galactosidase is detectable, when both allelesof Msx1 are targeted with nlacZ, in the lateraldermomyotome of interlimb somites, which will contributeto intercostal and body wall muscles (see Christ and Ordahl,1995), and is particularly strong in somites 15 and 16.Differences in gene expression on the anterior-posterior axisare classically identified for the Hox genes (see Krumlauf,1994). Although somites show flexibility in terms of their

2698
adaptation to changes in axial position, they do registerpositional information. This is demonstrated by micecarrying the MLC1F-CAT myosin transgene, which show anunexpected caudal-rostral gradient of expression as musclesform (Donoghue et al., 1991). Differences in the extent ofMsx1 expression in somites along the axis may well be aresponse to anterior-posterior positional information,perhaps via Hox genes.
Msx1 in muscle progenitor cells of the limbIn keeping with the observation that somites can bereprogrammed to produce limb muscle progenitor cells whentransplanted ectopically (Chevallier, 1979), mouse somitescan be reprogrammed to express Msx1 when transplantedfrom a non-expressing level to the wing level in the chick.Expression can be induced in somites grafted at this levelwhether they come from the most rostral region (somites 1-5) or from the hindlimb level (somites 26-32), and even insomites that form after grafting of the segmental plate. Thissuggests that the mesenchyme from the brachial level has thecapacity to induce Msx1 expression. In accordance withMsx1 expression in the mouse, chick leg mesenchyme doesnot induce expression of Msx1 in somites grafted at thisposterior level (somites 26-31). However, mouse somites
from the forelimb level, at a stage prior to detectable Msx1expression, when grafted at the hindlimb level in the chickdo express Msx1, while they do not when grafted rostrally tothe forelimb level. This suggests that two different signalslead to Msx1 expression in the somite: one that is instructive,for the induction of Msx1, present in the forelimb (andthoracic) mesenchyme, and another that is permissive,present also at the hindlimb level but not in neckmesenchyme. The forelimb, hindlimb and interlimbmesenchyme, in contrast to that at more rostral or caudalpositions on the axis, is also distinguished by its capacity torespond to FGFs and form limb buds (Cohn et al., 1995). Itis probable that the initial activation of Msx1 dependson BMP4. This has been demonstrated during toothmorphogenesis (Vainio et al., 1993), and for themesenchymal cells that are recruited to form the dorsal partof the vertebrae (Monsoro-Burq et al., 1996). In the case ofthe somite, expression of Bmp4, which has a dynamic patternduring mouse embryogenesis (Hogan et al., 1994), is highestin lateral plate mesoderm adjacent to the forelimb/thoracicsomites at the time when Msx1 is activated in the lateraldermomyotome at this level. Furthermore, BMP4 has beenshown to lateralise the somite, thus preventing or delayingthe onset of myogenesis, as evidenced by the inhibition of
D. Houzelstein and others
Fig. 4. Mouse somitic cells are reprogrammed toexpress Msx1 by grafting mouse somites in thechick at the brachial level. (a) A mouse-chickchimaera produced by implantation of theanteriormost (1-5) somites from an E8.5 Msx1nlacZ
mouse at the brachial level of an 18-somite chickembryo. In toto salmon-gal treatment shows redstaining in cells migrating towards the wing field(arrow), lateral to the grafted somites. (b) A mouse-chick chimaeric embryo, 18 hours after grafting.Unsegmented paraxial mesoderm from an Msx1nlacZ
mouse embryo at the thoracic level has replacedchick somite 16 and part of the segmental plate in a16-somite chick embryo. The unsegmented paraxialmesoderm has started to segment and β-galactosidase-positive cells are detected in thelateral margin of the newly formed mouse somitesopposite the wing field (arrow). (c) A mouse-chickchimaeric embryo, 24 hours after grafting. Fivesomites (11-15) from an Msx1nlacZ mouse embryohave been grafted to replace chick somites justanterior to the wing level in a 19-somite chickembryo. Only cells migrating from the mostposterior grafted somite, which is adjacent to thewing field, express the Msx1nlacZ gene and aredetected by β-galactosidase staining (arrow). Thedorsalmost aspect of the chick neural tube is lightlylabelled blue by the β-galactosidase reagents, as issometimes observed for the most externalstructures. (d) Lateral view of a mouse-chickchimaera, 20 hours after grafting. Mouse somites26-32 (hindlimb level) were grafted to an 18-somitechick embryo at the brachial level. Msx1nlacZ
expression is induced in the mouse somites and incells migrating from them, as viewed through the apical region of the chick wing bud (w). (e) Dorsal view of a mouse-chick chimaera, 19 hoursafter grafting. Somites 8-13 from an E9 mouse were grafted to replace presumptive somites 26-32 (hindlimb level) in a 25-somite chickembryo. β-galactosidase-positive cells are observed in the grafted somites and in cells migrating from them in the leg mesenchyme (l). (f) Insitu hybridisation of a 25-somite mouse embryo with a Bmp4 probe. Expression of Bmp4 is maximum in the forelimb mesenchyme facingsomites 11-15 (arrows), which are those where expression of Msx1 is first detected. Somites 9 and 14 are numbered. Scale bars, 500 µm.

2699Msx1 in muscle progenitor cells
MyoD expression in the chick (Pourquié et al., 1996). Anegative effect of lateral mesoderm on myogenesis is alsoseen in mouse explants from the interlimb level (Cossu et al.,1996a). We suggest that, in part at least, this may be due toBMP4 induction of Msx1.
Based on ex vivo observations (Song et al., 1992; Woloshinet al., 1995), which show that Msx1 represses the onset ofmyogenesis, and the demonstration here that Msx1 isdownregulated in the proximal region of the limb bud beforeactivation of Myf5, it is possible that Msx1 acts as an inhibitorof myogenesis in most, if not all muscle progenitor cells inthe forelimb. At other levels on the axis, other homeoboxgenes (see below) may perform a similar function. It alsopotentially fulfils this role in the lateral dermomyotome insomites of the forelimb and thoracic region. It has beenproposed that the distal limb environment is able to repressMyoD expression through an FGF-mediated pathway(Robson and Hughes, 1996). Furthermore, it has been shownthat FGF2 and FGF4 are able to maintain Msx1 expression inmurine limb buds grown in culture (Vogel et al., 1995; Wangand Sassoon, 1995). It is tempting to propose that theinhibitory action of FGFs in the limb may be mediated byMsx1 in vivo. Although it has been shown that Msx1 can bindto sequences in the MyoD enhancer (Woloshin et al., 1995),no information is available for Myf5. The lag of 3-4 somites(about 5 hours) before activation of Myf5nlacZ, and afterdownregulation of Msx1nlacZ, might suggest that this is not adirect effect of derepression, particularly since activation ofMyoD in the limb occurs slightly later than that of Myf5 (Ottet al., 1991). Indeed the large number of sites where Msx1 isexpressed in the embryo suggests that it may act as a generalrepressor of terminal differentiation, while maintainingproliferation, in various cell types.It may also be playing a role inapoptosis; BMP4 induction ofMsx2 has been associated withapoptosis of neural crest cells, forexample (Graham et al., 1994).
Limb muscle progenitor cellsmigrate from the somites for ashort period of time (10 hours inchick; 13-15 hours in mice). Inchick, 30-100 cells migrate fromeach dermomyotome (reviewed inHauschka, 1994). Since 6-7somites contribute, only about 700cells are progenitors of all the limbmusculature. Thus, amplificationof this population prior to the onsetof differentiation and formationof primary fibres, must beconsiderable. Msx1 may be one ofthe factors involved in the initialphase of cell division. Othergenes expressed in the lateraldermomyotome are also potentiallyinvolved in migration/proliferation/repression of differentiation of themuscle progenitor cells. Theseinclude Pax3, probably acting viathe c-met receptor, which is clearly
essential. Lbx1, homologue of the Drosophila ladybirdhomeobox gene, shows expression in the lateral margin ofsomites at the level of the forelimb bud in the mouse, and atlater stages, in the dorsal and ventral pre-muscle masses ofthe forelimb and hindlimb (Jagla et al., 1995). In Pax3-deficient (splotch) mice, Lbx1 transcripts are not detectablein somites (Mennerich et al., 1998). In the chick, it has beenshown recently to mark cells in the lateral lip ofdermomyotomes from occipital, cervical, forelimb andhindlimb, but not interlimb, somites, and in cells migratingfrom these (Dietrich et al., 1998). This expression patterntherefore differs from that of Msx1, which is detectable onlyin forelimb and, more weakly, interlimb somites on the axis.Another homeobox gene, Mox2, is also expressed in limbmuscle progenitor cells and mutation of this gene gives apartial limb muscle phenotype (Mankoo et al., 1999).Although migration and proliferation of muscle progenitorcells are most marked in the limb buds, these phenomena alsoapply to the interlimb somites where the onset of myogenesisis initially delayed in the lateral somitic bud (see Tajbakhshand Buckingham, 1999). The relative roles of these differentproteins, and that of the bHLH protein Sim1 also expressedin the lateral dermomyotome, remains to be determined.Msx2, the sister gene to Msx1, has also been reported toinhibit myogenesis (Takahashi et al., 1996). Transcripts ofMsx2 were not detectable in somites by in situ hybridisation(data not shown), but it should be kept in mind that Msx1transcripts were not detected either in the dermomyotomeprior to introduction of an nlacZ reporter gene. Targeting ofMsx2 with a reporter gene is underway in our laboratory, andthis should, as in the case of Msx1, facilitate detection of itsexpression. The Msx1 null mouse (Satokata and Maas, 1994;
Fig. 5. Msx1nlacZ downregulation prior to Myf5nlacZ activation and the onset of muscle differentiation.(a) An E10 double heterozygote Msx1nlacZ/+/Myf5nlacZ/+ embryo (35 somites), visualised by wholemount. The β-galactosidase signal in the myotomes is due to expression from the Myf5nlacZ allele(compare with the slightly older embryo shown in b) and at other sites (including expression in theprogress zone of the limb bud, dorsal neural tube and head, compare with the older embryo in c), toexpression from the Msx1nlacZ allele. The latter has been downregulated in the proximal triangle ofthe forelimb bud (inset) where myogenesis will be initiated only 5 hours later (2-3 additionalsomites). (b) An E10 Myf5nlacZ/+ embryo (36 somites). β-galactosidase activity is detected on wholemount at every site where Myf5 is normally expressed (myotomes, and also in the head in thetemporalis muscle and mesencephalon). No expression is detected in the forelimb bud, indicatingthat the onset of myogenesis has not yet occurred (inset). (c) An E10 Msx1nlacZ/+ embryo (39somites). At this stage, the whole proximal mesenchyme of the forelimb bud no longer expressesMsx1 (inset). (d) An E10 Myf5nlacZ/+ embryo (39 somites). A few Myf5nlacZ-expressing cells are nowdetected by β-galactosidase staining on whole mounts within the proximal region of the forelimbbud (inset), where the Msx1 gene has been downregulated (compare with c). Scale bars, 500 µm.

2700
Houzelstein et al., 1997) does not have a limb phenotype.Double mutant analyses should clarify the potential role ofthe Msx genes, and also the involvement of the other potentialregulatory factors present in the lateral dermomyotome andthe muscle progenitor cells which migrate from it.
We are very grateful to Drs B. Mankoo, V. Pachnis, S. Dietrich andA. Lumsden for sharing data prior to publication. We would like tothank Drs M. Goulding and P. Gruss for the gift of the mouse Pax3probe, and Dr B. Hogan for the Bmp4 probe. This work was supportedby the Institut Pasteur and the Centre National de la RechercheScientifique (CNRS) and by grants from the Association pour laRecherche contre le Cancer (ARC), the Association Française contreles Myopathies (AFM), and the Ministère de l’Éducation Nationale,de l’Enseignement Supérieur et de la Recherche (MENESR). DenisHouzelstein was funded by the MENESR and subsequently by theARC, and then by the AFM.
REFERENCES
Aoyama, H. and Asamoto, K. (1988). Determination of somite cells:independence of cell differentiation and morphogenesis. Development 104,15-28.
Beddington, R. S. and Martin, P. (1989). An in situ transgenic enzymemarker to monitor migration of cells in the mid-gestation mouseembryo. Somite contribution to the early forelimb bud. Mol. Biol. Cell 6,263-274.
Bladt, F., Riethmacher, D., Isenmann, S., Aguzzi, A. and Birchmeier, C.(1995). Essential role for the c-met receptor in the migration of myogenicprecursor cells into the limb bud. Nature 376, 768-771.
Bober, E., Franz, T., Arnold, H. H., Gruss, P. and Tremblay, P. (1994). Pax-3 is required for the development of limb muscles: a possible role for themigration of dermomyotomal muscle progenitor cells. Development 120,603-612.
Brand-Saberi, B., Wilting, J., Ebensperger, C. and Christ, B. (1996). Theformation of somite compartments in the avian embryo. Int. J. Dev. Biol.40, 411-420.
Burke, A. C., Nelson, C. E., Morgan, B. A. and Tabin, C. (1995). Hox genesand the evolution of vertebrate axial morphology. Development 121, 333-346.
Catron, K. M., Wang, H. Y., Hu, G. H., Shen, M. M. and Abate-Shen, C.(1996). Comparison of Msx-1 and Msx-2 suggests a molecular basis forfunctional redundancy. Mech. Dev. 55, 185-199.
Chapman, D. L., Garvey, N., Hancock, S., Alexiou, M., Agulnik, S. I.,Gibson-Brown, J. J., Cebra-Thomas, J., Bollag, R. J., Silver, L. M., andPapaioannou, V. E. (1996). Expression of the T-box family genes, Tbx1-Tbx5, during early mouse development. Develop. Dyn. 206, 379-390.
Chevallier, A. (1979). Role of the somitic mesoderm in the development ofthe thorax in bird embryos. II. Origin of thoracic and appendicularmusculature. J. Embryol. Exp. Morph. 49, 73-88.
Chevallier, A., Kieny, M. and Mauger, A. (1977). Limb-somite relationship:origin of the limb musculature. J. Embryol. Exp. Morph. 41, 245-258.
Christ, B., Jacob, H. J. and Jacob, M. (1977). Experimental analysis of theorigin of the wing musculature in avian embryos. Anat. Embryol. 150, 171-186.
Christ, B., Jacob, M. and Jacob, H. J. (1983). On the origin and developmentof the ventrolateral abdominal muscles in the avian embryo. Anexperimental and ultrastructural study. Anat. Embryol. 166, 87-101.
Christ, B. and Ordahl, C. P. (1995). Early stages of chick somitedevelopment. Anat. Embryol. 191, 381-396.
Cohn, M. J., Izpisua-Belmonte, J. C., Abud, H., Heath, J. K., and Tickle,C. (1995). Fibroblast growth factors induce additional limb developmentfrom the flank of chick embryos. Cell 80, 739-46.
Cossu, G., Kelly, R., Tajbakhsh, S., Didonna, S., Vivarelli, E. andBuckingham, M. (1996a). Activation of different myogenic pathways –Myf-5 is induced by the neural tube and MyoD by the dorsal ectoderm inmouse paraxial mesoderm. Development 122, 429-437.
Cossu, G., Tajbakhsh, S. and Buckingham, M. (1996b). How is myogenesisinitiated in the embryo? Trends Genet. 12, 218-223.
Daston, G., Lamar, E., Olivier, M. and Goulding, M. (1996). Pax-3 is
necessary for migration but not differentiation of limb muscle precursors inthe mouse. Development 122, 1017-1027.
Davidson, D. (1995). The function and evolution of MSX genes- pointers andparadoxes. Trends Genet. 11, 405-411.
Dietrich, S., Schubert, F. R., Healy, C., Sharpe, P. T. and Lumsden, A.(1998). Specification of the hypaxial musculature. Development 125, 2235-2249.
Donoghue, M. J., Merlie, J. P., Rosenthal, N. and Sanes, J. R. (1991).Rostrocaudal gradient of transgene expression in adult skeletal muscle.Proc. Natl. Acad. Sci. USA 88, 5847-5851.
Epstein, J. A., Shapiro, D. N., Cheng, J., Lam, P. Y. and Maas, R. L. (1996).Pax3 modulates expression of the c-Met receptor during limb muscledevelopment. Proc. Natl. Acad. Sci. USA 93, 4213-4218.
Fontaine-Pérus, J., Jarno, V., Fournier le Ray, C., Li, Z. and Paulin, D.(1995). Mouse chick chimera: a new model to study the in ovodevelopmental potentialities of mammalian somites. Development 121,1705-1718.
Fontaine-Pérus, J., Halgand, P., Chéraud, Y., Rouaud, T., Velasco, M. E.,Diaz, C. C., and Rieger, F. (1997). Mouse-chick chimera: a developmentalmodel of murine neurogenic cells. Development 124, 3025-3036.
Franz, T., Kothary, R., Surani, M. A., Halata, Z. and Grim, M. (1993). TheSplotch mutation interferes with muscle development in the limbs. Anat.Embryol. 187, 153-160.
Gibson-Brown, J. J., Agulnik, S. I., Chapman, D. L., Alexiou, M., Garvey,N., Silver, L. M., and Papaioannou, V. E. (1996). Evidence of a role forT-box genes in the evolution of limb morphogenesis and the specificationof forelimb/hindlimb identity. Mech. Dev. 56, 93-101.
Goulding, M. D., Chalepakis, G., Deutsch, U., Erselius, J. R. and Gruss,P. (1991). Pax-3, a novel murine DNA binding protein expressed duringearly neurogenesis. EMBO J. 10, 1135-1147.
Goulding, M., Lumsden, A. and Paquette, A. J. (1994). Regulation of Pax-3 expression in the dermomyotome and its role in muscle development.Development 120, 957-971.
Graham, A., Francis-West, P., Brickell, P., and Lumsden, A. (1994). Thesignalling molecule BMP4 mediates apoptosis in the rhombencephalicneural crest. Nature 372, 684-686.
Hauschka, S. D. (1994). The embryonic origin of muscle. In Myology, basicand clinical (ed. A. G. Engel and C. Franzini-Armstrong), Vol. 1, pp. 3-73.McGraw-Hill Inc., New York.
Hayashi, K. and Ozawa, E. (1995). Myogenic cell migration from somites isinduced by tissue contact with medial region of the presumptive limbmesoderm in chick embryos. Development 121, 661-669.
Henrique, D., Adam, J., Myat, A., Chitnis, A., Lewis, J. and Ish-Horowicz,D. (1995). Expression of a delta homologue in prospective neurons in thechick. Nature 375, 787-790.
Hill, R. E., Jones, P. F., Rees, A. R., Sime, C. M., Justice, M. J., Copeland,N. G., Jenkins, N. A., Graham, E. and Davidson, D. R. (1989). A newfamily of mouse homeo box-containing genes: molecular structure,chromosomal location, and developmental expression of Hox-7.1. GenesDev. 3, 26-37.
Hogan, B., Blessing, M., Winnier, G. E., Suzuki, N., and Jones, C. M.(1994). Growth factors in development – the role of TGF-β relatedpolypeptide signalling molecules in embryogenesis. Development 1997Supplement, 53-60.
Houzelstein, D., Cohen, A., Buckingham, M. E. and Robert, B. (1997).Insertional mutation of the mouse Msx1 homeobox gene by an nlacZreporter gene. Mech. Dev. 65, 123-133.
Jacob, M., Christ, B. and Jacob, H. J. (1978). On the migration of myogenicstem cells into the prospective wing region of chick embryos. A scanningand transmission electron microscope study. Anat. Embryol. 153, 179-193.
Jacob, M., Christ, B. and Jacob, H. J. (1979). The migration of myogeniccells from the somites into the leg region of avian embryos. Anultrastructural study. Anat. Embryol. 157, 291-309.
Jagla, K., Dolle, P., Mattei, M. G., Jagla, T., Schuhbaur, B., Dretzen, G.,Bellard, F. and Bellard, M. (1995). Mouse Lbx1 and human LBX1 definea novel mammalian homeobox gene family related to the Drosophila ladybird genes. Mech. Dev. 53, 345-356.
Jones, C. M., Lyons, K. M., and Hogan, B. L. M. (1991). Involvement ofBone Morphogenetic Protein-4 (BMP-4) and Vgr-1 in morphogenesis andneurogenesis in the mouse. Development 111, 531-542.
Keynes, R. J. and Stern, C. D. (1984). Segmentation in the vertebrate nervoussystem. Nature 310, 786-789.
Krumlauf, R. (1994). Hox genes in vertebrate development. Cell 78, 191-201.Lee, K. K. and Sze, L. Y. (1993). Role of the brachial somites in the
D. Houzelstein and others

2701Msx1 in muscle progenitor cells
development of the appendicular musculature in rat embryos. Dev. Dyn. 198,86-96.
Lyons, G. E., Houzelstein, D., Sassoon, D., Robert, B. and Buckingham,M. E. (1992). Multiple sites of Hox-7 expression during mouseembryogenesis: comparison with retinoic acid receptor mRNA localization.Mol. Reprod. Dev. 32, 303-314.
Mackenzie, A., Purdie, L., Davidson, D., Collinson, M. and Hill, R. E.(1997). Two enhancer domains control early aspects of the complexexpression pattern of Msx1. Mech. Dev. 62, 29-40.
Mackenzie, S., Walsh, F. S., and Graham, A. (1998). Migration ofhypoglossal myoblast precursors. Develop. Dyn. 213, 349-358.
Maina, F., Casagranda, F., Audero, E., Simeone, A., Comoglio, P. M.,Klein, R. and Ponzetto, C. (1996). Uncoupling of Grb2 from the Metreceptor in vivo reveals complex roles in muscle development. Cell 87, 531-542.
Mankoo, B. S., Collins, N. S., Ashby, P., Grigorieva, E., Pevny, L. H.,Candia, A., Wright, C. V., Rigby, P. W. and Pachnis, V. (1999). Mox2, acritical component of the genetic hierarchy controlling limb muscledevelopment. Nature (in press).
Mennerich, D., Schafer, K., and Braun, T. (1998). Pax-3 is necessary butnot sufficient for lbx1 expression in myogenic precursor cells of the limb.Mech. Dev. 73, 147-158.
Milaire, J. (1976). Contribution cellulaire des somites à la genèse desbourgeons de membre postérieurs chez la souris. Arch. Biol. 87, 315-343.
Milaire, J. (1987). Early structural changes associated with the onset offorelimb development in the mouse. Arch. Biol. 98, 67-98.
Monsoro-Burq, A. H., Duprez, D., Watanabe, Y., Bontoux, M., Vincent,C., Brickell, P. and Le Douarin, N. (1996). The role of bonemorphogenetic proteins in vertebral development. Development 122, 3607-3616.
Myat, A., Henrique, D., Ish-Horowicz, D., and Lewis, J. (1996). A chickhomologue of Serrate and its relationship with Notch and Delta homologuesduring central neurogenesis. Dev. Biol. 174, 233-247.
Natoli, T. A., Ellsworth, M. K., Wu, C., Gross, K. W., and Pruitt, S. C.(1997). Positive and negative DNA sequence elements are required toestablish the pattern of Pax3 expression. Development 124, 617-26.
Noden, D. M. (1983). The embryonic origins of avian cephalic and cervicalmuscles and associated connective tissues. Am. J. Anat. 168, 257-276.
Ott, M. O., Bober, E., Lyons, G., Arnold, H. and Buckingham, M. (1991).Early expression of the myogenic regulatory gene, myf-5, in precursorcells of skeletal muscle in the mouse embryo. Development 111, 1097-1107.
Pereira, M., Houzelstein, D., Vinci, G., Cohen, A., Abdelhay, E., andRobert, B. (1999). Analysis of the mouse Msx1 gene promoter usingembryonic stem cell-mediated transgenesis. Transgenics In press.
Pourquié, O., Fan, C. M., Coltey, M., Hirsinger, E., Watanabe, Y., Breant,C., Francis-West, P., Brickell, P., Tessier-Lavigne, M. and Le Douarin,N. M. (1996). Lateral and axial signals involved in avian somite patterning:a role for BMP4. Cell 84, 461-471.
Robert, B., Sassoon, D., Jacq, B., Gehring, W. and Buckingham, M. (1989).Hox-7, a mouse homeobox gene with a novel pattern of expression duringembryogenesis. EMBO J. 8, 91-100.
Robson, L. G. and Hughes, S. M. (1996). The distal limb environmentregulates Myod accumulation and muscle differentiation in mouse-chickchimaeric limbs. Development 122, 3899-3910.
Sassoon, D., Lyons, G., Wright, W. E., Lin, V., Lassar, A., Weintraub, H.,and Buckingham, M. (1989). Expression of two myogenic regulatoryfactors myogenin and MyoD1 during mouse embryogenesis. Nature 341,303-307.
Satokata, I. and Maas, R. (1994). Msx1 deficient mice exhibit cleft palate
and abnormalities of craniofacial and tooth development. Nat. Genet. 6, 348-356.
Solursh, M., Drake, C. and Meier, S. (1987). The migration of myogeniccells from the somites at the wing level in avian embryos. Dev. Biol. 121,389-396.
Song, K., Wang, Y. and Sassoon, D. (1992). Expression of Hox-7.1 inmyoblasts inhibits terminal differentiation and induces cell transformation.Nature 360, 477-481.
Spörle, R., and Schughart, K. (1997). System to identify individual somitesand their derivatives in the developing mouse embryo. Develop. Dyn. 210,216-226.
Sze, L. Y., Lee, K., Webb, S. E., Li, Z. L. and Paulin, D. (1995). Migrationof myogenic cells from the somites to the fore-limb buds of developingmouse embryos. Dev. Dyn. 203, 324-336.
Tajbakhsh, S. and Buckingham, M. E. (1994). Mouse limb muscle isdetermined in the absence of the earliest myogenic factor Myf-5. Proc. Natl.Acad. Sci. USA 91, 747-751.
Tajbakhsh, S., Bober, E., Babinet, C., Pournin, S., Arnold, H. andBuckingham, M. (1996a). Gene targeting the Myf-5 locus with nlaczreveals expression of this myogenic factor in mature skeletal muscle fibresas well as early embryonic muscle. Dev. Dyn. 206, 291-300.
Tajbakhsh, S., Rocancourt, D. and Buckingham, M. (1996b). Muscleprogenitor cells failing to respond to positional cues adopt non-myogenicfates in Myf-5 null mice. Nature 384, 266-270.
Tajbakhsh, S., Rocancourt, D., Cossu, G. and Buckingham, M. (1997).Redefining the genetic hierarchies controlling skeletal myogenesis – Pax-3and Myf-5 act upstream of Myod. Cell 89, 127-138.
Tajbakhsh, S., and Buckingham, M. (1999). The birth of muscle progenitorcells in the mouse: spatiotemporal considerations. In Somitogenesis (ed. C.P. Ordhal), San Diego: Academic Press. (In press).
Takahashi, C., Akiyama, N., Matsuzaki, T., Takai, S., Kitayama, H. andNoda, M. (1996). Characterization of a human Msx-2 cDNA and itsfragment isolated as a transformation suppressor gene against v-ki-rasoncogene. Oncogene 12, 2137-2146.
Tickle, C. (1996). Vertebrate limb development. Semin. Cell Dev. Biol. 7, 137-143.
Vainio, S., Karavanova, I., Jowett, A. and Thesleff, I. (1993). Identificationof BMP-4 as a Signal mediating secondary induction between epithelial andmesenchymal tissues during early tooth development. Cell 75, 45-58.
Vogel, A., Roberts-Clarke, D. and Niswander, L. (1995). Effect of FGF ongene expression in chick limb bud cells in vivo and in vitro. Dev. Biol. 171,507-520.
Wang, Y. and Sassoon, D. (1995). Ectoderm-mesenchyme and mesenchyme-mesenchyme interactions regulate Msx-1 expression and cellulardifferentiation in the murine limb bud. Dev. Biol. 168, 374-382.
Williams, B. A. and Ordahl, C. P. (1994). Pax-3 expression in segmentalmesoderm marks early stages in myogenic cell specification. Development120, 785-796.
Wilting, J., Brand-Saberi, B., Huang, R., Zhi, Q., Kontges, G., Ordahl, C.P. and Christ, B. (1995). Angiogenic potential of the avian somite. Dev.Dyn. 202, 165-171.
Woloshin, P., Song, K. N., Degnin, C., Killary, A. M., Goldhamer, D. J.,Sassoon, D. and Thayer, M. J. (1995). Msx1 inhibits Myod expression infibroblast × 10t1/2 cell hybrids. Cell 82, 611-620.
Yang, X. M., Vogan, K., Gros, P. and Park, M. (1996). Expression of themet receptor tyrosine kinase in muscle progenitor cells in somites and limbsis absent in splotch mice. Development 122, 2163-2171.
Zhi, Q. X., Huang, R. J., Christ, B. and Brand-Saberi, B. (1996).Participation of individual brachial somites in skeletal muscles of the aviandistal wing. Anat. Embryol. 194, 327-339.
Copyright © 2022 FDOKUMEN



















