
Intrinsic and extrinsic innervation of the heart in zebrafish (Danio rerio)
Upload
khangminh22Category
view
0download
0
/ . Embryo/, exp. Morph. Vol. 54, pp. 141-154, 1979Printed in Great Britain © Company of Biologists Limited 1977
Segmental innervation of the chick forelimbfollowing embryonic manipulation
By M. R. BENNETT,1 R. LINDEMAN1 AND A. G. PETT1GREW1
From the Neurobiology Laboratory, Department of Physiology,University of Sydney
SUMMARYA number of studies have shown that the segmental innervation of some muscles in the
developing limb undergoes some modification during the earliest stages of ontogeny. Theseobservations can be interpreted in support of the hypothesis that the motor axons andmuscles are matched during this period of development. As a further test of this suggestionwe have made a quantitative examination of the motor innervation of the chick forelimbunder conditions of controlled abnormal development.
Embryos were surgically manipulated at stages before the motor axons invade the limb.The operations were controlled such that forelimbs were induced with segments deleted orreduplicated or simply that a segment of the spinal cord had been deleted. In preparationswith abnormal limbs the motor innervation of the muscles present was the same as forthose muscles in the normal limb. Where a spinal segment had been deleted the limbsdeveloped normally and their innervation was completed by the remaining brachial segments.
These results suggest that any particular matching property of a developing muscle doesnot develop as a consequence of its position in the limb relative to those segments of thelimb proximal to it. Furthermore, that some muscles which are normally innervated by twospinal segments can be completely innervated by one of those spinal segments, in the absenceof the other, suggests that any matching between growing axons and developing muscle cellsis hierarchical rather than strictly all-or-nothing.
INTRODUCTION
The segmental distribution of motoneurones which project to particularmuscles in amphibian and avian limbs changes during the earliest stages ofontogeny (Lamb, 1976, 1977; McGrath & Bennett, 1979; Pettigrew, Lindeman& Bennett, 1979; see, however, Landmesser & Morris, 1975). Furthermore,Harris & Dennis (1977) and McGrath & Bennett (1979) have shown thatfunctional synapses formed by some axon terminals in a muscle regress infavour of those synapses formed by the axons which are destined to form themature innervation of the muscle. These results are not consistent with theidea that axons are selectively guided to a muscle such that the initial projectionis the same as that observed in mature animals of the same species (see Sperry,1963; Landmesser & Morris, 1975).
1 Authors' address: The Neurobiology Laboratory, Department of Physiology, Universityof Sydney, Sydney, N.S.W., Australia 2006.

142 M. R. BENNETT, R. LINDEMAN AND A. G. PETTIGREW
The observations above on developing nerve-muscle systems have beenlikened to those made on adult axolotls, where muscles can be competitivelyreinnervated by axon terminals of both the original and a foreign nerve. In suchpreparations both types of terminal compete for synaptic sites and this isfollowed by the elimination of the terminals formed by the foreign nerve (Cass &Mark, 1975; Bennett & Raftos, 1977; Dennis & Yip, 1978; Bennett, McGrath &Davey, 1979). The suggestion has been made that motor nerve terminalsare matched to particular muscles of the limb (see Bennett & Pettigrew, 1976)and that when well-matched and poorly matched terminals compete for thesame synaptic site, the poorly matched terminal will be eliminated (Bennett &Raftos, 1977; Dennis & Yip, 1978; Bennett et al. 1979). Thus, during develop-ment it is possible that the changes in the segmental innervation of a musclereflects the elimination of poorly matched terminals from the initial projectionto that muscle.
In the present work we have investigated this suggestion further by attemptingto force newly formed muscles to accept a stable innervation from nerveterminals which are normally eliminated. These experiments have involved thesurgical manipulation of embryos at very early stages of development toproduce either abnormal limbs, or embryos with abnormal spinal cords.
In an earlier study, Stirling & Summerbell (1977) made a largely histologicalanalysis of the pattern of nerves in forelimbs with truncations or deletionsalong the proximo-distal axis. These authors found that the pattern in thoseparts of the limb which were present was essentially normal, and that theinnervation of some of the major muscles was qualitatively similar to that inthe normal limb. The present study both confirms and extends the work ofStirling & Summerbell (1977) by providing a quantitative analysis of muscleinnervation in such limbs with truncations and deletions and also in limbswith reduplications of various segments or normal limbs innervated by anabnormal spinal cord.
METHODS
Experiments were performed on White Leghorn embryos, staged accordingto Hamburger & Hamilton (1951).
Operations
All operations were performed on embryos between stages 17 and 25.Truncation of the limb at various levels was achieved by removing the entireapical ectodermal ridge (AER) (Summerbell, 1974#). For trunction at the wrist,the AER was removed at stage 22 and for truncation at the elbow, the operationwas performed at stage 20. To produce limbs with mirror image reduplications(Summerbell, 19746) about the proximo-distal axis at the level of the elbow, asmall piece of tissue, which contained the zone of polarizing activity (ZPA),was taken from the posterior edge of a donor limb at stage 21 and then placed

Segmental innervation of chick forelimb 143in a similar sized niche on the anterior edge of a host limb at the same stage.The graft was held in place by a fine tungsten pin. To produce limbs reduplicatedat the wrist, a similar operation was performed at stages 23-24. To deletevarious portions of the proximo-distal axis of the forelimb, the wing tip,containing the AER and some mesenchyme, of older stage embryos, wasattached to wing stumps of younger embryos (Summerbell & Lewis, 1975).For preparations where digits were connected to the scapula, most of a stage-18limb bud was removed and replaced by the AER of a stage-23 limb bud. Forpreparations where the humerus was deleted, the donor AER was taken froma stage-21 embryo and attached to a stage-18 embryo.
Extirpation of a spinal segment was carried out on embryos at stages 18-20.Following identification of the segment to be removed, the spinal cord was cuton either side of the somite using iris forceps and the segment was removedby suction.
After each of these operations surviving embryos were incubated for abouta further week.
Estimation of muscle innervation
The pattern of innervation of various muscles in both control and operatedforelimbs was determined by recording either the electrical activity in themuscle nerves or the contraction of each muscle in response to stimulation ofspinal nerves 14, 15 and 16. All embryos were examined at 10-14 days' incu-bation. Individual embryos were removed from the egg and decapitated. Theoperated limb, together with the brachial spinal cord, was dissected free andplaced in a perspex organ bath containing a circulating modified Ringersolution (Pettigrew et al. 1979). The limb was skinned and the spinal roots wereexposed and cut close to the spinal cord. The whole preparation was thenpinned at the joints so that movement artifacts would not complicate therecording of muscle contraction. The spinal nerves were stimulated via asuction glass capillary electrode using pulses of 0-01-0-05 msec and 1-10 V.Muscle nerve activity was recorded in the intrinsic hand muscles (extensorindicis brevis, eib; abductor medius, am; flexor digiti quarti, fdq) using anothersuction glass capillary electrode placed over the point of entry of the individualnerve to each muscle. The tetanic contraction of the major limb muscles(biceps, b; triceps, t; flexor digitorum profundus,/d/>; flexor carpi ulnaris, feu;extensor metacarpi radialis, emr) was recorded using a Grass tension transducer.Individual muscles and tendons were freed from the distal insertion (theproximal insertion and point of nerve entry were left intact) and connected atrest length to the transducer using suture cotton tied securely around the tendon.The output of the transducer was monitored on a chart recorder. Repetitivestimulation (40 Hz) of each spinal nerve was halted as soon as the contractionreached its maximum amplitude and this amplitude was compared to thatrecorded with simultaneous stimulation of all three spinal nerves. The amplitude

144 M. R. BENNETT, R. LINDEMAN AND A. G. PETTIGREW
of the contraction produced by stimulation of all spinal nerves has been usedto provide a measure of the total number of muscle fibres which are innervated.The comparison of the response to stimulation of one spinal nerve with theresponse to combined stimulation provides, therefore, a measure of the pro-portion of muscle fibres in the muscle which are innervated by that particularspinal nerve. Thus in this study, the percentage innervation of a muscle by aspinal nerve has been determined by expressing the size of the tetanic con-traction with stimulation of that nerve, as a percentage of the contraction withsimultaneous stimulation of all three spinal nerves. At the end of an experiment,the limb was stained for cartilage using the Lundvall technique.
RESULTS
Innervation of the normal forelimb
The forelimb of the chick receives the major part of its innervation via spinalnerves 14, 15 and 16 (Roncali, 1970). Nerve 13 also innervates the limb in someembryos but in each case its contribution is small and has not been includedin the present study. The normal innervation of the major muscles of the limband the muscles in the hand is shown in the control distribution in Figs. 1 and2. The proximal part of the limb (biceps and triceps) is innervated mainlyby nerves 14 and 15; nerve 16 contributes only about 10% of the innervationof the triceps muscle. The more distal muscles in the limb are, in general,innervated by more caudal spinal segments. Nerve 14 is found only in extensormetacarpi radialis. The remaining distal forelimb muscles (flexor carpi ulnarisand flexor digitorum profundus) are innervated to some extent by nerve 15but mainly by nerve 16. The intrinsic muscles of the hand are innervated almostcompletely by nerve 16. None of the muscles listed above is innervated bymore than two of the three spinal segments studied.
In a number of cases the summed percentage innervation of a muscle by twospinal nerves exceeds 100 %. This observation has been noted previously in thedevelopment of muscle innervation in the chick wing (Pettigrew et al. 1979).The most likely explanation for this observation is that some muscle fibres aredually innervated at their synaptic site (Pettigrew et al. 1979) by separate axonsfrom each of the spinal nerves. These muscle fibres will develop a maximaltension in response to activity in either nerve terminal, and simultaneousstimulation of both terminals will have no additional effect on the tensiondeveloped. Thus, while the tension produced in response to simultaneousstimulation of spinal nerves provides an accurate measure of the total numberof muscle fibres which are innervated, it also probably reflects an underestimateof the total number of active synapses.

Segmental innervation of chick forelimb100 F
S 14
145
S15
S 16
Muscle
Fig. 1. Innervation of the major muscles of the chick forelimb. The percentageinnervation (see Methods) of forelimb muscles by segmental nerves S14, S15 andSI6 is shown for control limbs (filled circles) and for limbs with reduplicationsbelow the elbow (open circles, anterior reduplication; open triangles, posteriorreduplication). Preparations were examined at 12 days' incubation. Each pointrepresents the mean ± S.E.M. of four determinations for each muscle in each of eightpreparations. Note that the total percentage innervation of some muscles exceeds100%. This is believed to be the result of dual synaptic contact of some musclefibres by separate axons from the two spinal nerves which innervate each muscle(see text). Abbreviations are b, biceps; emr, extensor metacarpi radialis; /, triceps;feu flexor carpi ulnaris ;fdp, flexor digitorum profundus. Note that the innervationof all parts of the reduplicated limbs is substantially the same as that of the controllimb.
Innervation offorelimbs with reduplications about the proximo-distal axis
Two types of reduplicated or 'twinned' limbs were examined. Embryos wereused only when the gross anatomy of the reduplicated parts appeared quitenormal and all the muscles of interest could be easily identified.
The first type of preparation had been reduplicated at, or just proximal to,the elbow and had two distal forelimbs and hands (Fig. 3B). We have studiedonly those preparations where the radii were completely separated. In all suchpreparations, the segmental innervation of corresponding muscles identified inboth distal forelimbs was almost identical. Furthermore, the segmental inner-vation of the muscles studied, in both the proximal and reduplicated parts ofthe abnormal limb, was essentially the same as that for those muscles in the
146 M. R. BENNETT, R. LINDEMAN AND A. G. PETTIGREW
100
16St'gmental nerve
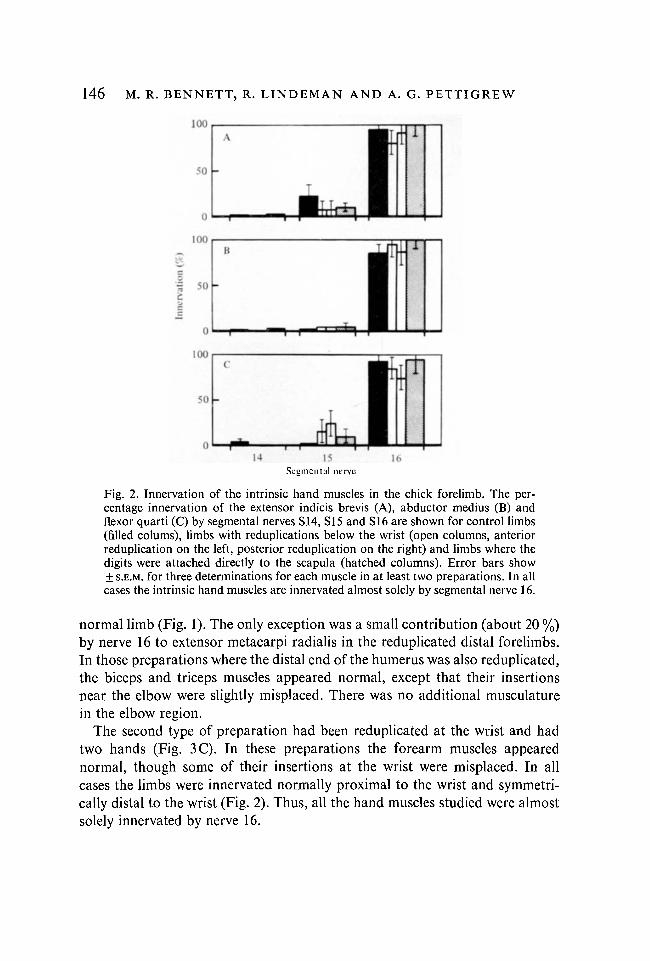
Fig. 2. Innervation of the intrinsic hand muscles in the chick forelimb. The per-centage innervation of the extensor indicis brevis (A), abductor medius (B) andflexor quarti (C) by segmental nerves S14, S15 and SI6 are shown for control limbs(filled colums), limbs with reduplications below the wrist (open columns, anteriorreduplication on the left, posterior reduplication on the right) and limbs where thedigits were attached directly to the scapula (hatched columns). Error bars show± S.E.M. for three determinations for each muscle in at least two preparations. In allcases the intrinsic hand muscles are innervated almost solely by segmental nerve 16.
normal limb (Fig. 1). The only exception was a small contribution (about 20 %)by nerve 16 to extensor metacarpi radialis in the reduplicated distal forelimbs.In those preparations where the distal end of the humerus was also reduplicated,the biceps and triceps muscles appeared normal, except that their insertionsnear the elbow were slightly misplaced. There was no additional musculaturein the elbow region.
The second type of preparation had been reduplicated at the wrist and hadtwo hands (Fig. 3C). In these preparations the forearm muscles appearednormal, though some of their insertions at the wrist were misplaced. In allcases the limbs were innervated normally proximal to the wrist and symmetri-cally distal to the wrist (Fig. 2). Thus, all the hand muscles studied were almostsolely innervated by nerve 16.

Segmental innervation of chick forelimb 147
Fig. 3. Photomicrographs of the bones of the right forelimb (dorsal view) at 11-14days' incubation. (A) Normal limb; (B) limb with mirror image reduplication distalto the elbow; (C) limb with mirror image reduplication distal to the wrist, h,humerus; r, radius; u, ulna; m, metacarpals.
148 M. R. BENNETT, R. LINDEMAN AND A. G. PETTIGREW
w^
D
Fig. 4. For legend see opposite.

Segmental innervation of chick forelimb 149
S 14
100
50
0
-
1
— i
o'
1
S 15
S 16
M usclc
Fig. 5. Percentage innervation of major limb muscles in preparations where digitswere deleted (open circles), parts distal to the elbow were deleted (open triangles)and where the humerus was deleted (open squares). The normal innervation is shownwith filled circles. Recordings were made at 14 days'incubation and each point repre-sents the mean of three determinations for each muscle from at least two prepara-tions; error bars show ±S.E.M. b, biceps; emr, extensor metacarpi radialis; /, triceps;feu, flexor carpi ulnaris; fdp, flexor digitorum profundus. Note that the segmentalinnervation of all muscles in the abnormal limbs is very similar to the innervation ofthose muscles in the normal limb.
Innervation of forelimbs with deletions along the proximo-distal axis
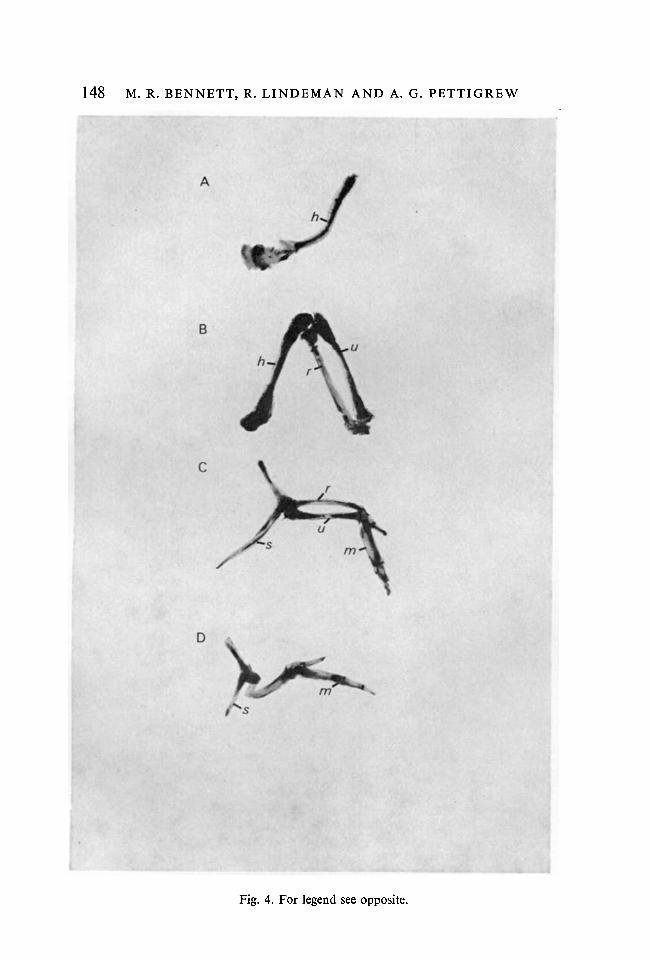
In forelimbs which had been truncated so that the digits failed to develop(Fig. 4B), the proximal and distal forelimb muscles were innervated normally(Fig. 5). In forelimbs which had failed to develop parts distal to the elbow(Fig. 4A), the biceps and triceps muscles were innervated normally by nerves14 and 15 (Fig. 5). Nerve 16 was absent from these limbs but it still normallyinnervated muscles adjacent to the limb (e.g. deltoids).
F IGURE 4
Photomicrographs of the bones of the right forelimb (dorsal view) following induceddeletion of segments along the proximo-distal axis. (A) Deletion of parts distal to theelbow; (B) deletion of parts distal to the wrist; (C) deletion of humerus; (D) deletionof the humerus, radius and ulna (hand attached to scapula), s, scapula; h, humerus;r, radius; //, ulna; m, metacarpals.

150 M. R. BENNETT, R. LINDEMAN AND A. G. PETTIGREW
Fig. 6. Photomicrographs of the right brachial plexus (ventral, rostral uppermost)in normal preparation (A) and a preparation where segment 14 had been removedat stage 20 (B). Calibration is 240/*m for (A) and 150 /*m for (B). sc, Supracora-coideus nerve. The inset in (B) shows a low power magnification of the brachialregion. Even though spinal nerve 14 is missing the basic geometry of the plexus hasbeen retained.
In preparations where the proximal ends of the radius and ulna were attachedto the scapula (humerus deleted) (Fig. 4C), the distal forelimb had developednormally and the three muscles examined received their normal pattern ofinnervation (Fig. 5). Similarly, where the hand was attached directly to thescapula (Fig. 4D), the three hand muscles examined were innervated almostsolely by nerve 16 (Fig. 3).
Thus in all the experimental situations described, where growing axons haveinvaded a grossly abnormal limb, the innervation of each of the muscles studiedat later stages was the same as for those muscles in the normal embryo.
Innervation of the forelimb following removal of a spinal cord segment
In embryos where a segment of the spinal cord was removed at stages beforethe axons have left the cord, the forelimb developed normally, with normalsize muscles, and appeared to show correct movement and coordination.Similarly, the brachial plexus in these embryos retained the basic geometry of
Segmental innervation of chick forelimb
100 F—
151
50
0 b_
100
S 16
S 50o
~ 0
S 15
100 F
50
0 -.
S 14
feuMuscle
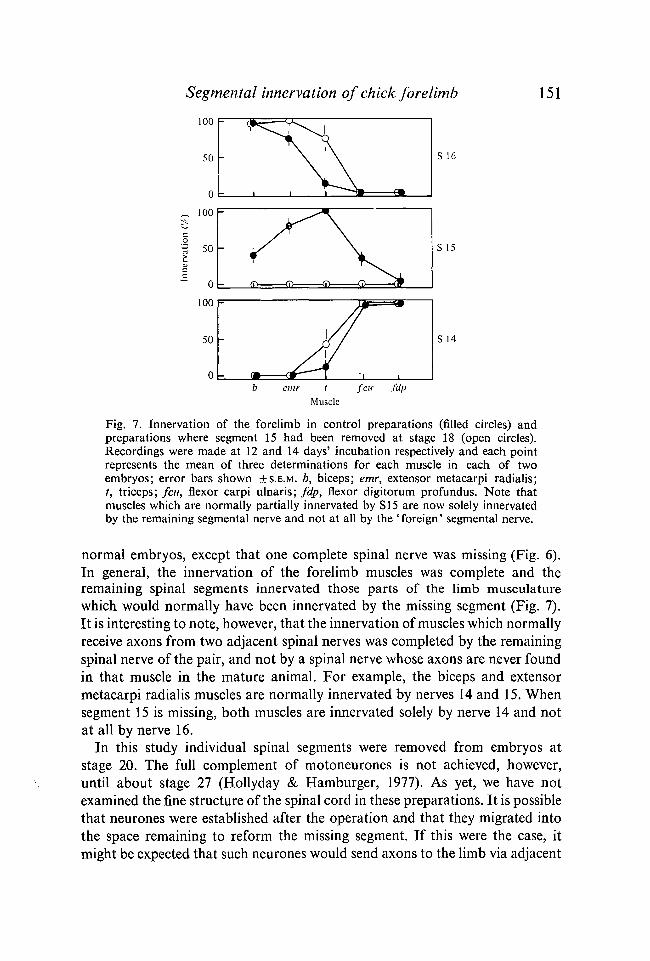
Fig. 7. Innervation of the forelimb in control preparations (filled circles) andpreparations where segment 15 had been removed at stage 18 (open circles).Recordings were made at 12 and 14 days' incubation respectively and each pointrepresents the mean of three determinations for each muscle in each of twoembryos; error bars shown ±S.E.M. b, biceps; emr, extensor metacarpi radialis;t, triceps; feu, flexor carpi ulnaris; fdp, flexor digitorum profundus. Note thatmuscles which are normally partially innervated by SI 5 are now solely innervatedby the remaining segmental nerve and not at all by the 'foreign' segmental nerve.
normal embryos, except that one complete spinal nerve was missing (Fig. 6).In general, the innervation of the forelimb muscles was complete and theremaining spinal segments innervated those parts of the limb musculaturewhich would normally have been innervated by the missing segment (Fig. 7).It is interesting to note, however, that the innervation of muscles which normallyreceive axons from two adjacent spinal nerves was completed by the remainingspinal nerve of the pair, and not by a spinal nerve whose axons are never foundin that muscle in the mature animal. For example, the biceps and extensormetacarpi radialis muscles are normally innervated by nerves 14 and 15. Whensegment 15 is missing, both muscles are innervated solely by nerve 14 and notat all by nerve 16.
In this study individual spinal segments were removed from embryos atstage 20. The full complement of motoneurones is not achieved, however,until about stage 27 (Hollyday & Hamburger, 1977). As yet, we have notexamined the fine structure of the spinal cord in these preparations. It is possiblethat neurones were established after the operation and that they migrated intothe space remaining to reform the missing segment. If this were the case, itmight be expected that such neurones would send axons to the limb via adjacent

152 M. R. BENNETT, R. LINDEMAN AND A. G. PETTIGREW
spinal nerves, because of the abnormal bone structure in this region. Thus, inpreparations where segment 15 had been removed we would have expected tofind the biceps muscle innervated in part by spinal nerve 16. In all such pre-parations, however, the biceps was innervated solely by nerve 14.
DISCUSSIONInnervation of abnormal limbs
The initial operations on the embryos used in this study were performed atstages before the axons which are to innervate the limb have reached theshoulder region. Nevertheless, in the series of experiments where the morphologyof the limb was abnormal, the motor innervation of all but one muscle examinedat 10-14 days' incubation was the same as that found in the control forelimb.This observation was made in limbs with either deletions along the proximo-distal axis or with reduplications about this axis. Thus, wherever a muscle wasplaced in the limb in relation to the parts proximal to it, it was almost alwaysinnervated normally. These results both confirm and extend the work ofStirling & Summerbell (1977).
The interpretation of our data relies on the observation that the motor axonsof each spinal nerve at the brachial level have had prior access to muscles ofthe forelimb. All the spinal nerves which form the brachial plexus duringnormal development are known to supply axon terminals to muscles of theforelimb (Pettigrew et al. 1979), although it has been claimed that the spinalnerves which form the lumbar plexus only supply axon terminals to the musclesof the hindlimb identical to those found in the mature muscles (Landmesser &Morris, 1975; for commentary on this paper see the Discussion in Pettigrewet al. 1979). In the present work operations which produced abnormal limbswere performed at stages before the motor axons have invaded the limbs. It isreasonable to assume, therefore, that at least some of the muscles in abnormallimbs receive an initial widespread segmental innervation, just as they wouldduring normal development (Pettigrew et al. 1979). That these muscles havethe same more restricted segmental innervation as in the normal limb by laterstages of development, suggests that the processes which result in the loss ofpart of the early innervation of a muscle are operating normally, even thoughthe muscle is in an abnormal position.
The development of limb innervation
The observation by Lamb (1976, 1977), McGrath & Bennett (1979) andPettigrew et al. (1979) that the segmental projection to at least some majorlimb muscles undergoes considerable modification at early stages is more easilyexplained in terms of matching between growing axons and muscle cells (seeHughes, 1968; McGrath & Bennett, 1979; Pettigrew et al 1979), rather thanin terms of strict fibre guidance (see Weiss, 1934; Sperry, 1963; Landmesser &

Segmental innervation of chick forelimb 153Morris, 1975). From the present experiments on abnormal limbs, it seemslikely that any selective matching properties of embryonic muscle cells may notbe determined by the position of the muscle in the limb relative to the partsproximal to it.
As a further test of the possible matching between embryonic nerve andmuscle cells, it was of interest to determine to what extent a normal limb wouldbe innervated by a spinal cord which was deficient in a group of motoneuronesof a particular spinal segment. In such preparations the limbs and musculaturewere fully developed and the motor innervation of each limb was completed bythe segments which remained. Previous studies, using only anatomical tech-niques, have described the pattern of nerves in developing chick limbs followingearly removal of two or three of the brachial (Castro, 1963) or lumbar spinalsegments (Kieny & Fouvet, 1974). While Castro describes hyperplasia in theone or two remaining nerves, it was nevertheless insufficient to reconstitute thenormal size of some of the limb nerves. The results of the present physiologicalstudy imply, however, that if only one spinal segment is removed, rather thantwo or three, hyperplasia in the remaining segmental nerves is sufficient tocompensate for the missing segmental nerve. The observations of Kieny andFouvet, in which the anatomical pattern of nerves in the hindlimb was deter-mined after removal of part of the lumbar spinal cord and associated somites,are difficult to interpret. In this study destruction of the somites produced grossabnormalities in hindlimb structure.
It is likely that early during the development of preparations with a singlesegment deleted, the remaining segments initially supplied axons to most musclesof the limb, as they do during normal ontogeny and that some axons weresubsequently lost from individual muscles during later development (Pettigrewet al. 1979). In some cases, however, muscles were left completely innervatedby axons which only partially innervate that muscle in the normal embryo. Thisobservation suggests that any matching between growing axons and musclesmay be hierarchical rather than strictly all-or-nothing, as is the matchingbetween axons and muscles in the axolotl (Bennett et al. 1979). Furthermore,such a hierarchy may be related to the general rostro-caudal spinal gradientalong the proximo-distal and antero-posterior gradients of innervation in thenormal limb. For example, the biceps muscle (anterior) is normally innervatedby nerves 14 and 15. When segment 15 has been removed the biceps is innervatedsolely by nerve 14 (rostral) and not at all by nerve 16 (caudal). In this context,it will be of interest to examine preparations where the spinal segment whichhas been removed is normally solely responsible for a muscle's innervation;for example, examination of the intrinsic hand muscles after removal of segment16.
We are very grateful to Ms J. Stratford for her excellent technical assistance. This workwas supported by the Australian Research Grants Committee.

154 M. R. BENNETT, R. LINDEMAN AND A. G. PETTIGREW
REFERENCES
BENNETT, M. R., MCGRATH, P. A. & DAVEY, D. F. (1979). The regression of synapses formedby a foreign nerve in a mature axolotl striated muscle. Brain Research. (In the Press.)
BENNETT, M. R. & PETTIGREW, A. G. (1976). The formation of neuromuscular synapses.Cold Spring Harb. Symp. quant. Biol. 40, 409-424.
BENNETT, M. R. & RAFTOS, J. (1977). The formation and regression of synapses duringreinnervation of axolotl striated muscles. / . Physiol. 265, 261-295.
CASS, D. T. & MARK, R. F. (1975). Reinnervation of axolotl limbs. I. motor nerves. Proc.R. Soc. B 190, 45-58.
CASTRO, G. (1963). Effects of reduction of nerve centers on the development of residualganglia and on nerve patterns in the wing of the chick embryo. / . exp. Zool. 152, 279-295.
DENNIS, M. J. & YIP, J. W. (1978). Formation and elimination of foreign synapses on adultsalamander muscle. / . Physiol. 274, 299-310.
HAMBURGER, V. & HAMILTON, H. L. (1951). A series of normal stages in the development ofthe chick embryo. / . Morph. 88, 49-92.
HARRIS, A. J. & DENNIS, M. J. (1977). Deletion of 'mistakes' in nerve-musle connectivityduring development of rat embryos. Soc. for Neurosciences, 7th Meeting, Anaheim,California. Abstracts, vol. in, p. 107.
HOLLYDAY, M. & HAMBURGER, V. (1977). An autoradiographic study of the formation ofthe lateral motor column in the chick embryo. Brain Res. 132, 197-208.
HUGHES, A. (1968). Development of limb innervation. In Growth of the Nervous System,pp. 110-117. London: Churchill.
KIENY, M. & FOUVET, B. (1974). Innervation et morphogenese de la patte chez l'embryonde poulet. Archs Anat. microsc. Morph. exp. 63, 281-298.
LAMB, A. H. (1976). The projection patterns of the ventral horn to the hind limb duringdevelopment. Devi Biol. 54, 82-99.
LAMB, A. H. (1977). Neuronal death in the development of the somatotopic projections ofthe ventral horn in Xenopus. Brain Res. 134, 145-150.
LANDMESSER, L. & MORRIS, D. G. (1975). The development of functional innervation in thehind limb of the chick embryo. J. Physiol. 249, 301-326.
MCGRATH, P. & BENNETT, M. R. (1979). The development of synaptic connections betweendifferent segmental motoneurones and and striated muscles in an axolotl limb. Devi Biol.69, 133-145.
PETTIGREW, A. G., LINDEMAN, R. & BENNETT, M. R. (1979). Development of the segmentalinnervation of the chick forelimb. J. Embryol. exp. Morph. 49, 115-137.
RONCALI, L. (1970). The brachial plexus and the wing nerve pattern during early develop-mental phases in chick embryo. Monitore Zool. Ital. 4, 81-98.
SAUNDERS, J. W. & GASSELING, M. T. (1968). Ectodermal-mesenchymal interactions in theorigin of limb symmetry. In Epithelial-Mesenchymal Interactions (ed. R. Fleishmajer &R. E. Billingham), pp. 78-97. Baltimore: Williams & Wilkins.
SPERRY, R. W. (1963). Chemoafnnity in the orderly growth of nerve fibre patterns andconnections. Proc. natn. Acad. Sci., U.S.A. 50, 703-710.
STIRLING, R. V. & SUMMERBELL, D. (1977). The development of functional innervation inthe chick wing-bud following truncations and deletions of the proximo-distal axis. J.Embryol. exp. Morph. 41, 189-207.
SUMMERBELL, D. (1974a). Interaction between the proximo-distal and antero-posterior co-ordinates of positional value during the specification of positional information in theearly development of the chick limb-bud. J. Embryol. exp. Morph. 32, 227-237.
SUMMERBELL, D. (19746). A quantitative analysis of the effect of excision of the AER fromthe chick limb-bud. / . Embryol. exp. Morph. 32, 651-660.
SUMMERBELL, D. & LEWIS, J. H. (1975). Time, place and positional value in the chick limb-bud. / . Embryol. exp. Morph. 33, 621-643.
WEISS, P. (1934). In vitro experiments on the factors determining the course of the outgrowingnerve fibre. / . exp. Zool. 68, 393-448.
(Received 5 February 1979, revised 8 April 1979)
Copyright © 2022 FDOKUMEN




















