
Orientation of migrating leatherback turtles in relation to ocean currents
11
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Orientation of migrating leatherback turtles in relation to ocean currents

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Orientation of migrating leatherback turtles in relation to ocean currents
S. Galli a, P. Gaspar b, S. Fossette c, B. Calmettes b, G. C. Hays c, J. R. E. Lutjeharms d,þ, P. Luschi a,*aDepartment of Biology, University of Pisa, Pisa, ItalybCollecte Localisation Satellites, Direction Océanographie Spatiale, Ramonville, FrancecDepartment of Biosciences, College of Science, Swansea University, Swansea, U.K.dDepartment of Oceanography, University of Cape Town, Rondebosch, South Africa
a r t i c l e i n f o
Article history:Received 15 March 2012Initial acceptance 19 April 2012Final acceptance 10 September 2012Available online 22 October 2012MS. number: 12-00214R
Keywords:Dermochelys coriaceadriftleatherback sea turtlemigrationnavigationswimming speed
During their offshore movements, leatherback turtles, Dermochelys coriacea, associate frequently withocean currents and mesoscale oceanographic features such as eddies, and their movements are often inaccordance with the current flow. To investigate how individual turtles oriented their ground- andwater-related movements in relation to the currents encountered on their journeys, we used oceano-graphic techniques to estimate the direction and intensity of ocean currents along the course of15 leatherbacks tracked by satellite during their long-distance movements in the Indian and AtlanticOceans. For all individuals a non-negligible component of active swimming was evident throughout thejourneys, even when their routes closely followed the currents, but overall the turtle water-relatedorientation was random with respect to current directions. For turtles in the North Atlantic, theground-related movements largely derived from the turtles’ active swimming, while in the Indian Oceancurrents contributed substantially to the observed movements. The same pattern was shown whendistinct parts of the routes corresponding to foraging bouts and travelling segments were consideredseparately. These findings substantiate previous qualitative observations of leatherback movements, byrevealing that turtles were not simply drifting passively but rather swam actively during most of theirjourneys, although with a random orientation with respect to currents. Our analysis did not provide anyindication that leatherbacks were able to detect the current drift they were exposed to, further high-lighting the navigational challenges they face in their oceanic wanderings.� 2012 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.
For animals living in the open sea, ocean currents representa key environmental factor that greatly affects their movementsand overall behaviour (Chapman et al. 2011). Currents can influencethe distribution and movements of pelagic animals in a variety ofways, for instance determining the distribution of food resources(Olson et al. 1994; Bost et al. 2009; Kai et al. 2009) or physicallyinfluencing their displacement (Girard et al. 2006, 2009; Cotté et al.2007). Indeed, the observed velocity of marine animals is the vectorsum of their swimming velocity and the water velocity (i.e. thecurrent velocity) so that the resulting ground-based path maydiffer, sometimes substantially, from the animal’s water-relatedroute (Green & Alerstam 2002; Gaspar et al. 2006; Girard et al.2006). Such a current drift is expected to play a strong role in thelong-distance oceanic movements of many marine animals,possibly leading to special navigational performances (Sale &Luschi 2009; Chapman et al. 2011).
Leatherback sea turtles, Dermochelys coriacea, are ideal subjectsfor studying the role that oceanographic features play in themovements of marine animals. Leatherbacks spend a lot of theirtime in the open sea (Eckert et al. 2012), foraging for gelatinousplankton associated with ocean currents (Olson et al. 1994;Lutcavage 1996). In recent years, satellite telemetry has provideda wealth of data documenting the migratory movements of adultsea turtles (Godley et al. 2008), and studies that have integratedtracking datawith oceanographic information have shown how theturtles’ movements are often influenced by the oceanic circulationof the areas frequented (Luschi et al. 2003a). Leatherbacks, inparticular, have been found to associate frequently with mesoscalefeatures such as eddies, following circuitous courses in accordancewith the rotation of the water masses (Luschi et al. 2003b; Doyleet al. 2008; Lambardi et al. 2008). Movements in general agree-ment with the current flow have also been revealed outside eddies,for instance in the presence of stable, strong currents (Luschi et al.2003b; Lambardi et al. 2008; Shillinger et al. 2008). Quantitativeassessments of the currents along the leatherback routes havefurther revealed that the geographical displacements of ocean-moving turtles are largely determined by currents (Gaspar et al.
* Correspondence: P. Luschi, Department of Biology, University of Pisa, Via A.Volta 6, I-56126 Pisa, Italy.
E-mail address: [email protected] (P. Luschi).þ Deceased
Contents lists available at SciVerse ScienceDirect
Animal Behaviour
journal homepage: www.elsevier .com/locate/anbehav
0003-3472/$38.00 � 2012 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.anbehav.2012.09.022
Animal Behaviour 84 (2012) 1491e1500
Author's personal copy
2006; Shillinger et al. 2008; Fossette et al. 2010a) and that thewater-related motor paths of individuals often differ from theground-related courses as recorded by satellite (Gaspar et al. 2006).
However, no experimental data are currently available onhow individual leatherbacks actually deal with the currents theyencounter, and to what degree they take them into account in theirlong-distance journeys. For instance, it is unclear whether the oftenobserved pattern of leatherback movements that are in accord withprevailing currents derives from a simple drift of the turtle withlittle or no active swimming in the horizontal plane, or whetherindividuals contribute actively to the observedmovements in thesecases. This issue can be profitably addressed by relying on existingmethods to evaluate the direction and intensity of the currents ata given time in oceanic locations. These methods are based uponestimations of the currents’ geostrophic component deduced fromsatellite altimetry data and of the wind-driven component derivedfrom satellite scatterometer measurements and have proven toprovide reliable quantitative estimates of the currents encounteredby satellite-tracked turtles at various points along their paths(Gaspar et al. 2006; Girard et al. 2006; Shillinger et al. 2008;Fossette et al. 2010a, 2012). In the present study, we applied thisapproach to the extended and prolonged routes of 15 leatherbackstracked in the Indian and Atlantic Oceans (1) to study how indi-vidual turtles oriented their movements in relation to the currentsencountered during their open-sea journeys, (2) to quantify towhatextent the currents actually contributed to the reconstructed turtlemovements, and (3) to establish whether the leatherbacks’responses to currents were indicative of their detection of thecurrent flow or of their being unaware of the current drift theyweresubjected to. While previous studies on green sea turtles, Cheloniamydas, employing thesemethods have provided no indications thatthese turtles were able to detect currents during offshore move-ments (Girard et al. 2006; Luschi et al. 2007), it is not possible toextend these findings readily to leatherbacks, which belong toa different family and broadly differ from green turtles in theirecology and spatial behaviour. For instance, because of theirmarked pelagic habits, leatherbacks may have evolved special skillsto detect currents that are not available to the green turtles, whichspend far less time in the oceanic environment.
METHODS
Analysed Movement Data
We have analysed a large set of tracking data on leatherbackstracked in the Atlantic (N¼ 11) and the Indian (N¼ 4)Oceans. Detailsof the duration and length of tracks can be found in Table 1 and inpreviously published papers (Hughes et al. 1998; Luschi et al. 2003b;Hays et al. 2004a, 2006; James et al. 2006;Doyle et al. 2008; Lambardiet al. 2008; Fossette et al. 2010a). In all cases, turtles were tracked bymeans of satellite-linked radiotransmitters localized by the ArgosSystem, which provided locations unevenly spaced in time and spaceand differing in spatial accuracy (see www.argos-system.org fordetails). In 13 turtles, transmitters were attached using custom-fittedharnesses, while in turtles A6 and A7 the units were directly attachedto the carapace (Fossette et al. 2008). The turtleswere tracked for longperiods (mean: 6.3 months; range 2e11 months, Table 1) whilemoving in the oceanic environment (see Figs S1, S2 in the Supple-mentary Material) during different periods of their migratory cycle,including the postnesting migrations towards high-latitude foragingareas, the successive migrations towards tropical and subtropicalwaters, as well as the residence periods in relatively limited areas forfeeding purposes (Table 1). During their journeys, they encountereda variety of oceanographic conditions, such as strong currents like theGulf Stream or the Agulhas Current, eddies and gyres (Luschi et al.
2003b; Hays et al. 2006; Doyle et al. 2008; Lambardi et al. 2008).The reconstructed routes displayed a large degree of variation,probably deriving from the different migratory strategies adopted bythe turtles (Fossette et al. 2010a). Most journeys extended over hugedistances (2023e8994 km; Table 1) and led the turtles to visit well-known leatherback foraging areas such as the North Americancontinental shelf (James et al. 2005a, b; Eckert et al. 2006), theoffshore convergence zones of North and central Atlantic (Ferraroliet al. 2004; Eckert 2006) and the southwestern Indian Ocean(Luschi et al. 2006).Most routes included both straight and circuitoussegments, but some only consisted of straight or convoluted parts(Figs S1, S2 in the Supplementary Material).
Data Processing
Each track was processed by using the same protocol appliedpreviously (Gaspar et al. 2006) to reconstruct the turtles’ routes.Argos locations of any accuracy were used, but those implyinga ground speed above 10 km/h (278 cm/s) were discarded, and theresulting track was resampled with a fixed sampling periodDt ¼ 8 h. Locations for which a proper interpolation was notpossible because of large temporal gaps in the original satellite fixeswere discarded. As a result, some segments of the turtles’ rou-tes were not considered for the analysis (Figs S1, S2 in theSupplementary Material). When successive resampled positions½XðtÞ; Xðt þ DtÞ� were available, a track vector (T) was systemati-cally computed:
TðtÞ ¼ ½Xðt þ DtÞ � XðtÞ�=Dt (1)
This vector is the actual velocity on the ground of the trackedanimal averaged over the whole time interval ½t; t þ Dt�.
Table 1Main statistics of the analysed turtle tracks
TurtleID
Startinglocation
Trackingduration(months)
Tracklength(km)
Type of movement
A1 Grenada 11.4 2831 Postnesting migration(equatorial)
A2 Grenada 7.5 7564 Postnesting migration(round trip)
A3 Grenada 6.7 4608 Postnesting migration(northbound)
A4 Grenada 7.0 8994 Postnesting migration(northbound)
A5 Grenada 6.0 6913 Postnesting migration(northbound)
A6 French Guiana 3.3 5271 Postnesting migration(northbound)
A7 Ireland 5.3 8276 Southbound migrationA8 Nova Scotia 4.3 8923 Feeding grounds
residenceþsouthboundmigration
A9 Nova Scotia 2.2 2023 Feeding groundsresidence
A10 Ireland 9.3 7479 Southbound migrationA11 Grenada 7.5 8494 Postnesting migration
(round trip)I1 South Africa 7.4 7683 Postnesting migration
(southbound)I2 South Africa 5.6 5766 Postnesting migration
(round trip)I3 South Africa 4.1 5416 Postnesting migration
(southbound)I4 South Africa 7.4 8806 Postnesting migration
(southbound)
All tracked turtles were females except turtle A7. Values of tracking duration andlength refer to the parts of the routes considered for the analysis (i.e. excluding theparts for which current estimation or a proper interpolation was not possible).
S. Galli et al. / Animal Behaviour 84 (2012) 1491e15001492

Author's personal copy
Following Gaspar et al. (2006), a surface current vector C wasthen computed, at each resampled position X(t). This vector wasobtained as the sum of the geostrophic current componentdeduced from satellite altimetry and the ageostrophic, wind-drivencomponent estimated using the Rio & Hernandez (2003) model.The geostrophic current was obtained as the sum of the meangeostrophic current deduced from the mean sea level of Rio &Hernandez (2004) and the deviation from the mean deducedfrom altimeter-derived weekly sea level anomalymaps provided byAVISO (http://www.aviso.oceanobs.com) with a spatial resolutionof 1/3�. The Ekman component of the current was computed fromdaily wind stresses derived from QuickSCAT scatterometermeasurements and obtained from CERSAT (http://www.ifremer.fr/cersat) on a 25 � 25 km grid. This technique of surface currentestimation is now widely used in physical oceanography, and itsaccuracy has been assessed by Pascual et al. (2006) and Sudre &Morrow (2008). Information on surface currents provides a reli-able description of the currents experienced by migrating leather-backs, as these turtles are known to dive mostly in the upper layersof thewater column (Lutcavage & Lutz 1997). This was the case herewith most individuals remaining at depths shallower than 200 m,with only a few exceptional dives to hundreds of metres (Hays et al.2004b, 2006; McMahon & Hays 2006; Sale et al. 2006).
Basic fluid mechanics dictate that the instantaneous velocity onthe ground (Vg) of any marine animal actively swimming withinoceanic currents is simply the vector sum of the current vector (C)plus the swimming velocity vector, also called heading vector (H)(e.g. Gaspar et al. 2006; see inset in Fig. 1):
VgðtÞ ¼ CðtÞ þ HðtÞ (2)
Unfortunately, while current velocities were estimated here atdiscrete times t, t þ Dt ., instantaneous velocities on the groundwere not available, as we could only compute the T vector (equation1) representing the mean value of Vg over the time interval½t; t þ Dt�. Still, the equation relating T(t) to the instantaneouscurrent and heading vectors is readily obtained by integrating (2)between t and t þ Dt, and dividing by Dt. Using (1), this yields:
TðtÞ ¼ 1Dt
264
ZtþDt
t
CðtÞdtþZtþDt
t
HðtÞdt
375 (3)
Making the simple, and accurate, assumption that the currentvelocity varies linearly during the (short) period Dt, the mean valueof the current vector during the time interval ½t; t þ Dt� is easilycomputed as:
1Dt
ZtþDt
t
CðtÞdt ¼ 12½CðtÞ þ Cðt þ DtÞ� (4)
so that (3) finally yields:
1Dt
ZtþDt
t
HðtÞdt ¼ TðtÞ � 12½CðtÞ þ Cðt þ DtÞ� (5)
This relation shows that, under the only and simple assumptionthat currents vary linearly over the resampling period, the exactmean swimming velocity (or heading) vector computed over thetime interval ½t; t þ Dt� is simply the vector difference between T(t)and ½CðtÞ þ Cðt þ DtÞ�=2.
For the purpose of simplifying notations in the rest of the paper,T(t) will be denoted T, ½CðtÞ þ Cðt þ DtÞ�=2 will be denoted C andaccordingly H ¼ TeC. One should keep inmind that these T, H and C
vectors actually represent our best estimates of the turtle velocityon the ground (track vector), of its swimming velocity (headingvector) and of the current, averaged over a sampling period of 8 h.These vectors are represented by a direction (in degrees, clockwisefrom north) and a norm (speed in cm/s). Note that the distancestravelled by the turtles during these 8 h periods were around 10e20 km on average (individual means between 13.9 and 19.9 kmfor Indian Ocean turtles; between 4.0 and 14.6 km for Atlanticturtles).
Data Analysis
To study how the turtles oriented their water-related move-ments in relation to the currents, we calculated the angles betweenthe H and the C vector (HeC angles) and studied their distributionin the individual turtles. This provided us a way to evaluatewhether turtles detected currents during their offshore move-ments: a nonrandom distribution of HeC angles would indicatethat turtles tended to orient their movements at some angle withrespect to the current (having probably detected it), while if turtlesmoved while having no information on the direction of the currentthey were in, HeC angles would be distributed randomly. Similarly,we calculated the angle between the T and the C vector (TeC angle),which provides a measure of how the turtles’ ground-relatedmovements (expressed by the T vectors) were oriented withrespect to the currents. For instance, TeC angles close to 0� willindicate that the turtles’ resulting movements were in accordancewith the currents, while angles around 180� will indicate that theresulting turtle movements were oriented away from the currents,because of an intense swimming activity overcoming currentaction.
These analyses were first done by taking the whole routes oftracked turtles into account, and then by considering separatelydifferent route segments. Indeed, applying the procedure to thewhole route may somewhat mask any variations in turtle behav-iour during different parts of their journeys. As mentioned above,the prolonged movements of the tracked leatherback turtlesencompassed different phases, such as travelling and foragingsegments, differentiated by the turtles’ differing behaviouralstrategies and needs (e.g. James et al. 2005a, b; Jonsen et al. 2007;Bailey et al. 2008; Fossette et al. 2010a). It is likely that the turtleswill not react in the same way to the currents in these differentconditions, which were combined in this analysis. To circumventthis problem, a subsample of eight routes was broken up intodifferent phases, separating convoluted segments, where turtleswere more likely to be foraging, from linear ones where otheractivities such as travelling were predominant. Making suchdistinctions based on telemetry data only is not a straightforwardtask (Fossette et al. 2010a), and so we focused only on those turtlesin which this distinction was most evident. These included thesegments for which other evidence was available (concomitantpresence of mesoscale oceanographic features or periods in knownproductive foraging areas) showing that these were probableforaging bouts (Doyle et al. 2008; Fossette et al. 2010a, b; Luschiet al. 2003b; Lambardi et al. 2008). For comparison, one leg foreach turtle was chosen that was straight and covered at high speedand that was considered to be a travelling segment. For the otherroutes, no such clear subdivision was possible and so we excludedthem to avoid arbitrary distinctions.
Finally, we also quantified the contribution of the currents andof the turtles’ active swimming to the ground-related movements.For this, we computed the projection of the C vector on the T one(PCT), calculated as PCT ¼ C � cos(AceAt), where C is the speed ofthe C vector, Ac is the direction of the C vector and At is thedirection of the T vector; and the projection of the H vector on
S. Galli et al. / Animal Behaviour 84 (2012) 1491e1500 1493
Author's personal copy
T vector (PHT), calculated using the formula above with theappropriate changes.
Circular data (geographical direction of currents and differencesbetween angles) were elaborated following standard methods ofcircular statistics (Batschelet 1981). For the distribution of angles ofeach turtle, a mean vector was calculated, described by a meanangle and a mean vector length (r) which ranged from 0 (uniformlyscattered distribution) to 1 (fully concentrated distribution). Sincethe original angular data were not independent (being obtained byrepeated measures from the same individuals), the distributions ofthe mean vectors of the individual turtles were tested forrandomness by using second-order statistical analysis (Hotellingtest, Batschelet 1981). Because of such lack of independence, water-related speeds were compared between Atlantic and Indian Oceanturtles using a generalized linear mixed model (GLMM) witha normal error distribution and correlation structure R1.Geographical area was the fixed factor, and turtle individual wasthe random factor. Calculations were made with the NRLMEpackage in R 2.15 (The R Project for Statistical Computing, http://www.r-project.org/).
RESULTS
The currents experienced by tracked turtles along their routeswere generally non-negligible (Fig. 1), especially in the IndianOcean where they often reached values of 40e50 cm/s. They werevery variable in direction, as shown by the circular distributions oftheir geographical directions which mostly have a small meanvector length (<0.5 in 13 cases out of 15; Table 2). Only the Atlanticturtles A1 and A9 encountered currents that were rather uniform in
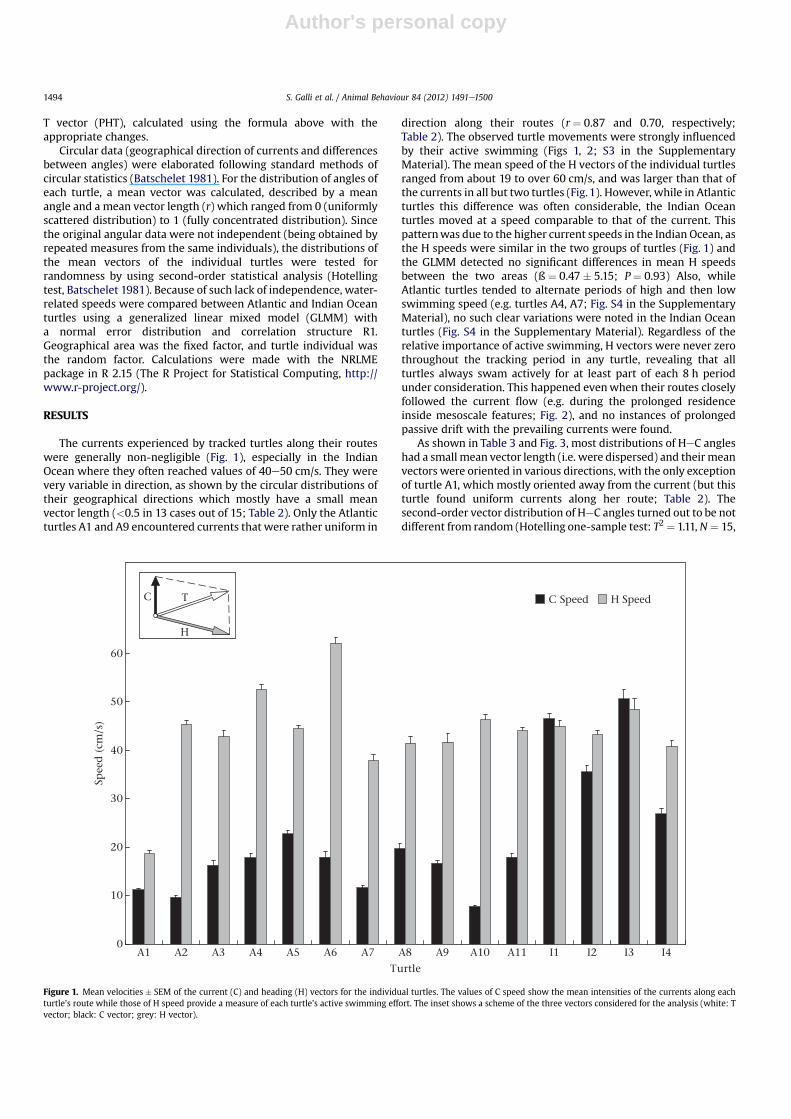
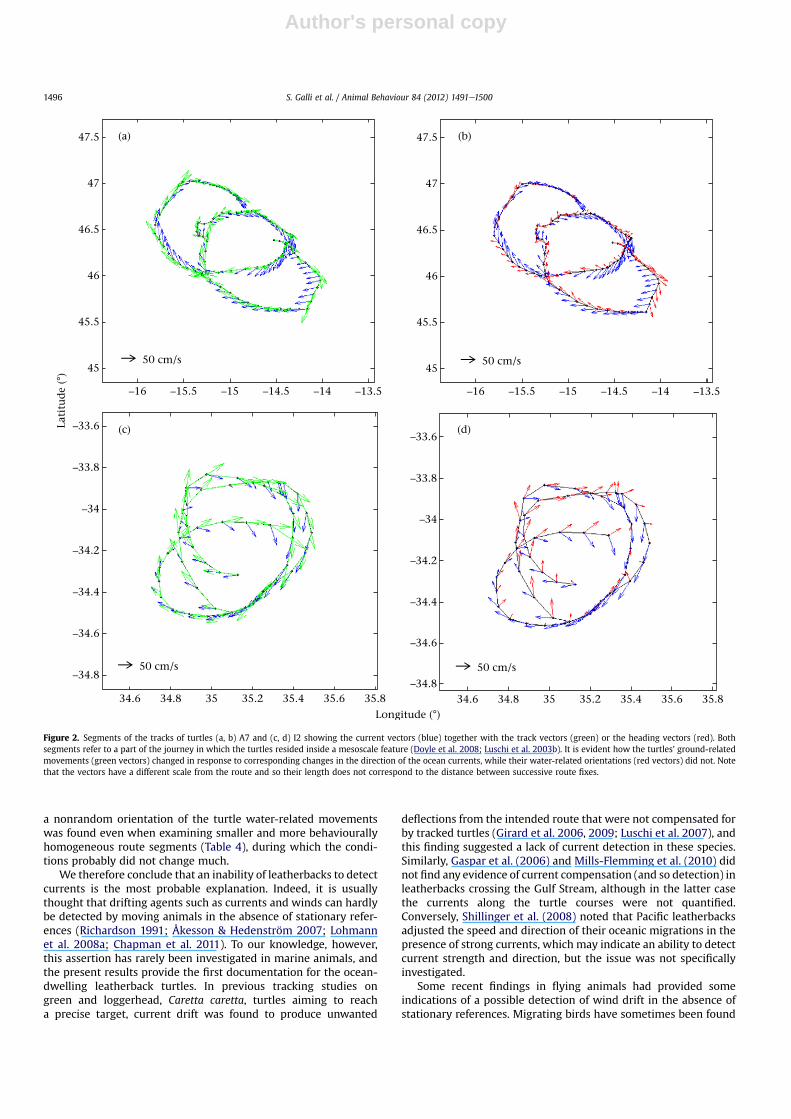
direction along their routes (r ¼ 0.87 and 0.70, respectively;Table 2). The observed turtle movements were strongly influencedby their active swimming (Figs 1, 2; S3 in the SupplementaryMaterial). The mean speed of the H vectors of the individual turtlesranged from about 19 to over 60 cm/s, and was larger than that ofthe currents in all but two turtles (Fig.1). However, while in Atlanticturtles this difference was often considerable, the Indian Oceanturtles moved at a speed comparable to that of the current. Thispatternwas due to the higher current speeds in the Indian Ocean, asthe H speeds were similar in the two groups of turtles (Fig. 1) andthe GLMM detected no significant differences in mean H speedsbetween the two areas (ß ¼ 0.47 � 5.15; P ¼ 0.93) Also, whileAtlantic turtles tended to alternate periods of high and then lowswimming speed (e.g. turtles A4, A7; Fig. S4 in the SupplementaryMaterial), no such clear variations were noted in the Indian Oceanturtles (Fig. S4 in the Supplementary Material). Regardless of therelative importance of active swimming, H vectors were never zerothroughout the tracking period in any turtle, revealing that allturtles always swam actively for at least part of each 8 h periodunder consideration. This happened evenwhen their routes closelyfollowed the current flow (e.g. during the prolonged residenceinside mesoscale features; Fig. 2), and no instances of prolongedpassive drift with the prevailing currents were found.
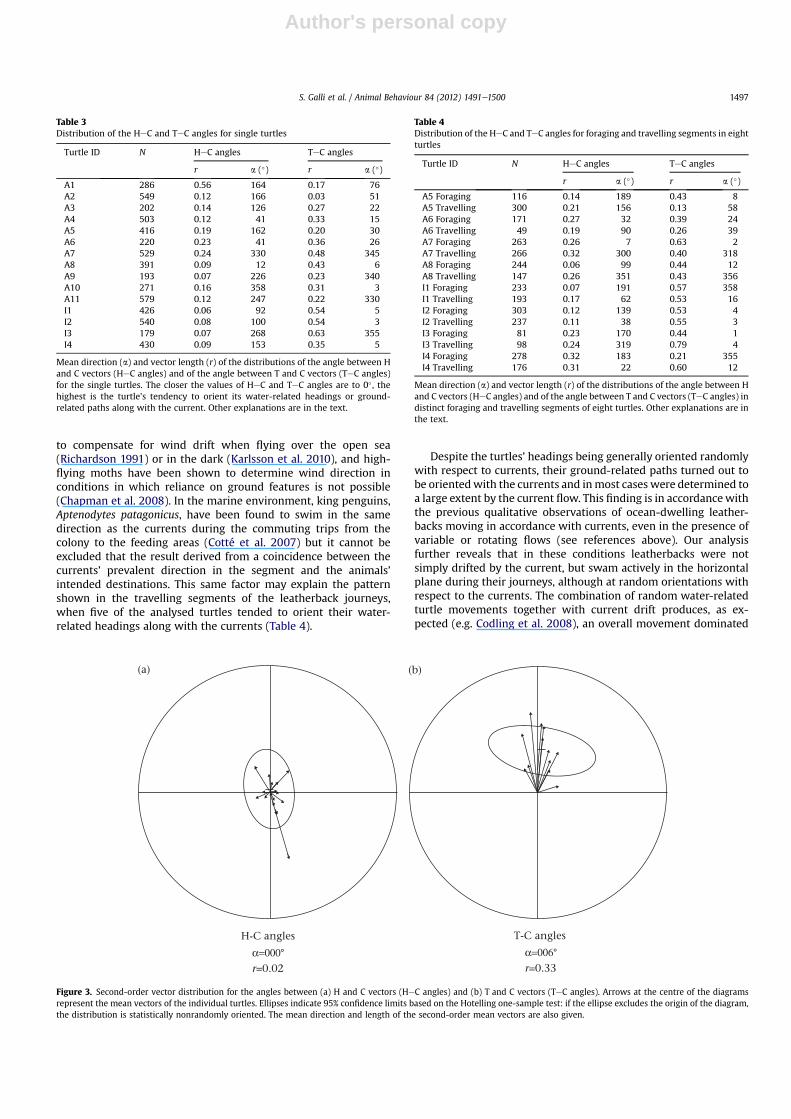
As shown in Table 3 and Fig. 3, most distributions of HeC angleshad a small mean vector length (i.e. were dispersed) and their meanvectors were oriented in various directions, with the only exceptionof turtle A1, which mostly oriented away from the current (but thisturtle found uniform currents along her route; Table 2). Thesecond-order vector distribution of HeC angles turned out to be notdifferent from random (Hotelling one-sample test: T2 ¼ 1.11,N ¼ 15,
0
10
20
30
40
50
60
A1 A2 A3 A4 A5 A6 A7 A8 A9 A10 A11 I1 I2 I3 I4
Turtle
Spee
d (
cm/s
)
C Speed H SpeedC T
H
Figure 1. Mean velocities � SEM of the current (C) and heading (H) vectors for the individual turtles. The values of C speed show the mean intensities of the currents along eachturtle’s route while those of H speed provide a measure of each turtle’s active swimming effort. The inset shows a scheme of the three vectors considered for the analysis (white: Tvector; black: C vector; grey: H vector).
S. Galli et al. / Animal Behaviour 84 (2012) 1491e15001494

Author's personal copy
P ¼ 0.69; Fig. 3). This result shows that turtles oriented at randomwith respect to the currents and provides no indication that theturtles adjusted their headings in relation to the (varying) currents(see below for a detailed discussion of this finding).
A largely different pattern emerges from the analysis of thedistributions of TeC angles, which were usually oriented close to0� (Table 3, Fig. 3) and often showed a small variability (asexpressed by the relatively large vector lengths, Table 3). Thesecond-order vector distribution was indeed significantly differentfrom random (Hotelling one-sample test: T2 ¼ 71.19, N ¼ 15,P < 0.001; Fig. 3) and oriented close to 0�. In other words, theturtles’ ground-relatedmovements were on average oriented in thesame direction as the current, as can also be qualitatively appre-ciated by examining small portions of the turtles’ route such asthose shown in Fig. 2.
The analysis of the turtle orientation in different segmentsconfirmed the trend shown for the whole route: in both foragingand travelling segments the distributions of HeC angles were verydispersed (r values <0.40), while TeC distributions were moreconcentrated and usually oriented close to 0� (Table 4). In six out ofeight turtles, HeC distributions were oriented closer to 0� duringtravelling segments than during foraging ones, which indicates thatwhen performing straight movements, these turtles tended toorient their water-related headings with the prevailing currents,even though they always displayed a large variability (Table 4).
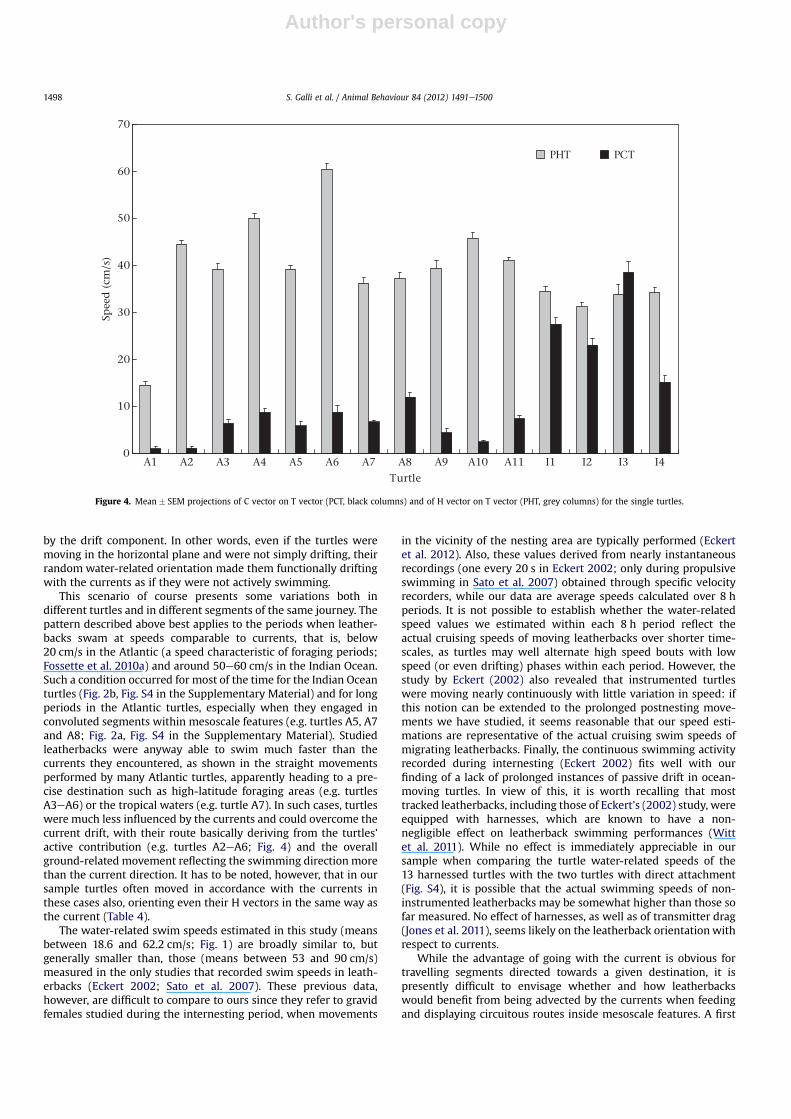
As shown in Fig. 4, in all turtles the mean values of PHT weregenerally large, although PCT too was positive in all turtles, andespecially large in the Indian Ocean ones. This shows how theturtles’ active movements (represented by PHT) provided thehighest contribution to the observed movement, especially inthe Atlantic turtles. In the Indian Ocean turtles, conversely, theground-related course of the turtles depended on the turtles’ activeswimming and on the currents to a comparable extent, withcurrents often contributing substantially to the determination ofthe turtles’ overall movement.
DISCUSSION
A number of papers have shown that leatherback turtle oceanicmovements are often made in accordance with the main currentflow, either in the presence of stable currents (Luschi et al. 2003b;Lambardi et al. 2008; Shillinger et al. 2008) or within mesoscalefeatures (Luschi et al. 2003b; Doyle et al. 2008; Lambardi et al.2008). The present analysis sheds further light on the behaviourof migrating leatherbacks with respect to the currents, showing
how turtles oriented their water-related movements and providinga quantification of the contribution of currents to the observedroute.
In the analysed data set, no indication has been found thatturtles oriented their water-related movements (H vectors) inrelation to the currents, both when the whole routes were takeninto account and when smaller track portions were considered.This in turn indicates that the turtles were probably unable todetect the currents they experienced along their routes: if a turtlemoved while having no information on the direction of the currentit was in, HeC angles will tend to be distributed randomly, as wasthe case.
The soundness of this finding depends on the reliability of thecurrent estimations (Fossette et al. 2012) as well as on the spatialuncertainty of the original Argos locations used to reconstruct theturtle routes (Robel et al. 2011). Both factors, however, are unlikelyto have introduced any systematic bias into our analyses, whoseoutcomes were very consistent among different turtles and indifferent geographical areas. The method we used to assesscurrents has been validated with tracking data from surface drifters(Fossette et al. 2012) and has been found to provide reliable esti-mates of mesoscale current velocities, except for coastal locationsand for the equatorial band (Pascual et al. 2006; Sudre & Morrow2008). The inaccuracies in Argos locations for sea turtle applica-tions are estimated to be around 10e15 km in the worst cases(Hazel 2009; Witt et al. 2010) and so are of a smaller magnitudethan the spatial resolution of current estimations, for which thegeostrophic component is computed with 1/3� resolution (i.e.>30 km). Also, in the present study the effects of these inaccuracieshave been reduced, at least in part, by the filtering and interpola-tion processes that we have applied. Futurework on the subject willtake advantage of the improvements that are being made to thespatial and temporal resolution of the turtle tracking data andocean current estimates. For example, Fastloc GPS tags are nowproviding a more detailed view of turtle oceanic movements (Hazel2009;Witt et al. 2010) and higher resolution oceanographic modelsare starting to allow better current estimates for some areas.
Our key finding that there was no indication that turtlesoriented their water-related movements (H vectors) in relation tothe currents may therefore derive from two different situations.
(1) The turtle water-related movements are variable (as isthought to happen in foraging bouts; Fossette et al. 2010a) andrandomly oriented relative to currents. These can be either variableor stable (like in the A9 turtle case), as in both cases the H andC vectors will remain uncorrelated.
(2) The turtle movements are directed (as is typically the caseduring travelling segments), but the currents are variable along thefollowed trajectory (owing, for example, to mesoscale variability).In that case again, H and C vectors will exhibit little or nocorrelation.
In both cases, the behaviour of studied turtles is that of ananimal moving at random with respect to the currents, mostprobably because of a lack of knowledge of the currents encoun-tered during the journey and also given that there were no meansfor turtles to anticipate the direction of the currents in advance asthey mostly moved in regions with variable currents (Table 2). Thepresent data do not totally rule out the possibility that leatherbackscan detect offshore currents, since from our analysis we cannotexclude that they may be aware of the currents but did not accountfor them in orienting their movements. It may also be that theturtles’ swimming patterns relative to the currents can changewithlocal environmental and behavioural conditions (e.g. current speedand direction, presence of a destination), and that our evaluationsbased on data pooled over long periods may have missed suchsubtle variations in turtle behaviour. However, no indication of
Table 2Variability of the currents along each turtle’s route
Turtle ID N r a (�)
A1 286 0.87 279A2 549 0.23 242A3 202 0.22 308A4 503 0.19 59A5 416 0.28 269A6 220 0.21 234A7 529 0.23 265A8 391 0.33 108A9 193 0.70 76A10 271 0.17 185A11 579 0.19 299I1 426 0.33 220I2 540 0.13 228I3 179 0.49 202I4 430 0.42 289
Data shown are the mean directions (a, 0� ¼ North) and vector lengths (r, seeMethods section) of the current vectors used for the analysis.
S. Galli et al. / Animal Behaviour 84 (2012) 1491e1500 1495
Author's personal copy
a nonrandom orientation of the turtle water-related movementswas found even when examining smaller and more behaviourallyhomogeneous route segments (Table 4), during which the condi-tions probably did not change much.
We therefore conclude that an inability of leatherbacks to detectcurrents is the most probable explanation. Indeed, it is usuallythought that drifting agents such as currents and winds can hardlybe detected by moving animals in the absence of stationary refer-ences (Richardson 1991; Åkesson & Hedenström 2007; Lohmannet al. 2008a; Chapman et al. 2011). To our knowledge, however,this assertion has rarely been investigated in marine animals, andthe present results provide the first documentation for the ocean-dwelling leatherback turtles. In previous tracking studies ongreen and loggerhead, Caretta caretta, turtles aiming to reacha precise target, current drift was found to produce unwanted
deflections from the intended route that were not compensated forby tracked turtles (Girard et al. 2006, 2009; Luschi et al. 2007), andthis finding suggested a lack of current detection in these species.Similarly, Gaspar et al. (2006) and Mills-Flemming et al. (2010) didnot find any evidence of current compensation (and so detection) inleatherbacks crossing the Gulf Stream, although in the latter casethe currents along the turtle courses were not quantified.Conversely, Shillinger et al. (2008) noted that Pacific leatherbacksadjusted the speed and direction of their oceanic migrations in thepresence of strong currents, which may indicate an ability to detectcurrent strength and direction, but the issue was not specificallyinvestigated.
Some recent findings in flying animals had provided someindications of a possible detection of wind drift in the absence ofstationary references. Migrating birds have sometimes been found
47.5 (a)
(c) (d)
(b)
47
46.5
46
45.5
45
–33.6
–33.8
–34
–34.2
–34.4
–34.6
–34.8
34.6 34.8 35 35.2 35.4 35.6 35.8
–16 –15.5 –15 –14.5 –14 –13.5 –16 –15.5 –15 –14.5 –14 –13.5
47.5
47
46.5
46
45.5
45
–33.6
–33.8
–34
–34.2
–34.4
–34.6
–34.834.6 34.8 35 35.2 35.4 35.6 35.8
Lati
tud
e (°
)
Longitude (°)
50 cm/s 50 cm/s
50 cm/s 50 cm/s
Figure 2. Segments of the tracks of turtles (a, b) A7 and (c, d) I2 showing the current vectors (blue) together with the track vectors (green) or the heading vectors (red). Bothsegments refer to a part of the journey in which the turtles resided inside a mesoscale feature (Doyle et al. 2008; Luschi et al. 2003b). It is evident how the turtles’ ground-relatedmovements (green vectors) changed in response to corresponding changes in the direction of the ocean currents, while their water-related orientations (red vectors) did not. Notethat the vectors have a different scale from the route and so their length does not correspond to the distance between successive route fixes.
S. Galli et al. / Animal Behaviour 84 (2012) 1491e15001496
Author's personal copy
to compensate for wind drift when flying over the open sea(Richardson 1991) or in the dark (Karlsson et al. 2010), and high-flying moths have been shown to determine wind direction inconditions in which reliance on ground features is not possible(Chapman et al. 2008). In the marine environment, king penguins,Aptenodytes patagonicus, have been found to swim in the samedirection as the currents during the commuting trips from thecolony to the feeding areas (Cotté et al. 2007) but it cannot beexcluded that the result derived from a coincidence between thecurrents’ prevalent direction in the segment and the animals’intended destinations. This same factor may explain the patternshown in the travelling segments of the leatherback journeys,when five of the analysed turtles tended to orient their water-related headings along with the currents (Table 4).
Despite the turtles’ headings being generally oriented randomlywith respect to currents, their ground-related paths turned out tobe orientedwith the currents and inmost cases were determined toa large extent by the current flow. This finding is in accordancewiththe previous qualitative observations of ocean-dwelling leather-backs moving in accordance with currents, even in the presence ofvariable or rotating flows (see references above). Our analysisfurther reveals that in these conditions leatherbacks were notsimply drifted by the current, but swam actively in the horizontalplane during their journeys, although at random orientations withrespect to the currents. The combination of random water-relatedturtle movements together with current drift produces, as ex-pected (e.g. Codling et al. 2008), an overall movement dominated
Table 3Distribution of the HeC and TeC angles for single turtles
Turtle ID N HeC angles TeC angles
r a (�) r a (�)
A1 286 0.56 164 0.17 76A2 549 0.12 166 0.03 51A3 202 0.14 126 0.27 22A4 503 0.12 41 0.33 15A5 416 0.19 162 0.20 30A6 220 0.23 41 0.36 26A7 529 0.24 330 0.48 345A8 391 0.09 12 0.43 6A9 193 0.07 226 0.23 340A10 271 0.16 358 0.31 3A11 579 0.12 247 0.22 330I1 426 0.06 92 0.54 5I2 540 0.08 100 0.54 3I3 179 0.07 268 0.63 355I4 430 0.09 153 0.35 5
Mean direction (a) and vector length (r) of the distributions of the angle between Hand C vectors (HeC angles) and of the angle between T and C vectors (TeC angles)for the single turtles. The closer the values of HeC and TeC angles are to 0� , thehighest is the turtle’s tendency to orient its water-related headings or ground-related paths along with the current. Other explanations are in the text.
H-C angles
(a) (b)
r=0.02α=000
T-C angles
r=0.33α=006
Figure 3. Second-order vector distribution for the angles between (a) H and C vectors (HeC angles) and (b) T and C vectors (TeC angles). Arrows at the centre of the diagramsrepresent the mean vectors of the individual turtles. Ellipses indicate 95% confidence limits based on the Hotelling one-sample test: if the ellipse excludes the origin of the diagram,the distribution is statistically nonrandomly oriented. The mean direction and length of the second-order mean vectors are also given.
Table 4Distribution of the HeC and TeC angles for foraging and travelling segments in eightturtles
Turtle ID N HeC angles TeC angles
r a (�) r a (�)
A5 Foraging 116 0.14 189 0.43 8A5 Travelling 300 0.21 156 0.13 58A6 Foraging 171 0.27 32 0.39 24A6 Travelling 49 0.19 90 0.26 39A7 Foraging 263 0.26 7 0.63 2A7 Travelling 266 0.32 300 0.40 318A8 Foraging 244 0.06 99 0.44 12A8 Travelling 147 0.26 351 0.43 356I1 Foraging 233 0.07 191 0.57 358I1 Travelling 193 0.17 62 0.53 16I2 Foraging 303 0.12 139 0.53 4I2 Travelling 237 0.11 38 0.55 3I3 Foraging 81 0.23 170 0.44 1I3 Travelling 98 0.24 319 0.79 4I4 Foraging 278 0.32 183 0.21 355I4 Travelling 176 0.31 22 0.60 12
Mean direction (a) and vector length (r) of the distributions of the angle between Hand C vectors (HeC angles) and of the angle between T and C vectors (TeC angles) indistinct foraging and travelling segments of eight turtles. Other explanations are inthe text.
S. Galli et al. / Animal Behaviour 84 (2012) 1491e1500 1497
Author's personal copy
by the drift component. In other words, even if the turtles weremoving in the horizontal plane and were not simply drifting, theirrandom water-related orientation made them functionally driftingwith the currents as if they were not actively swimming.
This scenario of course presents some variations both indifferent turtles and in different segments of the same journey. Thepattern described above best applies to the periods when leather-backs swam at speeds comparable to currents, that is, below20 cm/s in the Atlantic (a speed characteristic of foraging periods;Fossette et al. 2010a) and around 50e60 cm/s in the Indian Ocean.Such a condition occurred for most of the time for the Indian Oceanturtles (Fig. 2b, Fig. S4 in the Supplementary Material) and for longperiods in the Atlantic turtles, especially when they engaged inconvoluted segments within mesoscale features (e.g. turtles A5, A7and A8; Fig. 2a, Fig. S4 in the Supplementary Material). Studiedleatherbacks were anyway able to swim much faster than thecurrents they encountered, as shown in the straight movementsperformed by many Atlantic turtles, apparently heading to a pre-cise destination such as high-latitude foraging areas (e.g. turtlesA3eA6) or the tropical waters (e.g. turtle A7). In such cases, turtleswere much less influenced by the currents and could overcome thecurrent drift, with their route basically deriving from the turtles’active contribution (e.g. turtles A2eA6; Fig. 4) and the overallground-related movement reflecting the swimming direction morethan the current direction. It has to be noted, however, that in oursample turtles often moved in accordance with the currents inthese cases also, orienting even their H vectors in the same way asthe current (Table 4).
The water-related swim speeds estimated in this study (meansbetween 18.6 and 62.2 cm/s; Fig. 1) are broadly similar to, butgenerally smaller than, those (means between 53 and 90 cm/s)measured in the only studies that recorded swim speeds in leath-erbacks (Eckert 2002; Sato et al. 2007). These previous data,however, are difficult to compare to ours since they refer to gravidfemales studied during the internesting period, when movements
in the vicinity of the nesting area are typically performed (Eckertet al. 2012). Also, these values derived from nearly instantaneousrecordings (one every 20 s in Eckert 2002; only during propulsiveswimming in Sato et al. 2007) obtained through specific velocityrecorders, while our data are average speeds calculated over 8 hperiods. It is not possible to establish whether the water-relatedspeed values we estimated within each 8 h period reflect theactual cruising speeds of moving leatherbacks over shorter time-scales, as turtles may well alternate high speed bouts with lowspeed (or even drifting) phases within each period. However, thestudy by Eckert (2002) also revealed that instrumented turtleswere moving nearly continuously with little variation in speed: ifthis notion can be extended to the prolonged postnesting move-ments we have studied, it seems reasonable that our speed esti-mations are representative of the actual cruising swim speeds ofmigrating leatherbacks. Finally, the continuous swimming activityrecorded during internesting (Eckert 2002) fits well with ourfinding of a lack of prolonged instances of passive drift in ocean-moving turtles. In view of this, it is worth recalling that mosttracked leatherbacks, including those of Eckert’s (2002) study, wereequipped with harnesses, which are known to have a non-negligible effect on leatherback swimming performances (Wittet al. 2011). While no effect is immediately appreciable in oursample when comparing the turtle water-related speeds of the13 harnessed turtles with the two turtles with direct attachment(Fig. S4), it is possible that the actual swimming speeds of non-instrumented leatherbacks may be somewhat higher than those sofar measured. No effect of harnesses, as well as of transmitter drag(Jones et al. 2011), seems likely on the leatherback orientation withrespect to currents.
While the advantage of going with the current is obvious fortravelling segments directed towards a given destination, it ispresently difficult to envisage whether and how leatherbackswould benefit from being advected by the currents when feedingand displaying circuitous routes inside mesoscale features. A first
0
10
20
30
40
50
60
70
A1 A2 A3 A4 A5 A6 A7 A8 A9 A10 A11 I1 I2 I3 I4
Turtle
Spee
d (
cm/s
)
PHT PCT
Figure 4. Mean � SEM projections of C vector on T vector (PCT, black columns) and of H vector on T vector (PHT, grey columns) for the single turtles.
S. Galli et al. / Animal Behaviour 84 (2012) 1491e15001498

Author's personal copy
immediate consequence is that in this way they took advantage ofthe current drift and would increase their speed and distancecovered, ultimately hitchhiking the currents (as shown e.g. forhigh-flying insects; Chapman et al. 2010). The actual advantage ofsuch an enhanced speed for a foraging turtle is not clear: mesoscalefeatures are thought to represent foraging hotspots for planktivo-rous species (Lévy et al. 1999; Lima et al. 2002) and so leatherbacksshould tend to remain associated with them as long as possible. It ispossible that the turtles’ overall drift with the current is notnecessarily advantageous to them, at least for their spatial behav-iour: theymostly dive in the water column to find their prey, whichdrift with the currents, and the resulting effect of current drift ontheir horizontal movement is merely a side-effect of this feeding/diving behaviour (which may rather benefit from the turtle asso-ciationwith currents). In other words, the overall orientation of theturtle courses along with the current is just an outcome of theturtles’ unawareness (or disregard) of the currents and of theirconsequent random orientation with respect to them. Given thatthe gelatinous plankton prey of leatherbacks are advected bycurrents as well, the turtles’ movements would in the end mirrorthose of their prey.
In any case, the present findings have a number of importantconsequences for the leatherback navigational performances andabilities. Female leatherbacks are known to display nesting sitefidelity and this means that after 2 or 3 years spent foraging in theopen ocean, in each breeding season they have to navigate back totheir specific nesting area (Lohmann et al. 2008b; Sale & Luschi2009). The present analysis shows that the offshore migrations ofleatherback turtles are largely governed by oceanic currents andthat leatherback swimming directions are not directly linked to thedirection of the current in which they are swimming. Returning toa fixed site after years spent wandering in these conditions there-fore represents a most challenging task that renders the leather-back oceanic migration one of the most astonishing feats ofnavigation known in marine animals.
Acknowledgments
The authors wish to dedicate the paper to the memory ofProfessor Johann Lutjeharms, who died during the final stages ofmanuscript preparation. S.F. was supported by the Esmee FairbairnFoundation. Funding for the original research was provided bygrants to P.L. from Italian MIUR and the University of Pisa and toG.C.H. from the Natural Environment Research Council of the U.K.(NERC). We thank J-Y Georges for sharing his data set. We aregrateful to George Hughes, Andy Myers, Jon Houghton, Tom Doyleand Mike James for logistical help attaching tags in the field; toDimitri Giunchi for help with circular statistics; to Floriano Papi,Silvano Benvenuti, Jessi Brown and Resi Mencacci for comments onthe manuscript. The paper benefited from the useful suggestions ofthree anonymous referees. This study adhered to the legalrequirements of the countries in which the work was carried outand to all institutional guidelines.
Supplementary Material
Supplementarymaterial is available, in the online version of thisarticle, at http://dx.doi.org/10.1016/j.anbehav.2012.09.022.
References
Åkesson, S. & Hedenström, A. 2007. How migrants get there: migratory perfor-mance and orientation. BioScience, 57, 123e133.
Bailey, H., Shillinger, G., Palacios, D., Bograd, S., Spotila, J. R., Paladino, F. V. &Block, A. B. 2008. Identifying and comparing phases of movement by
leatherback turtles using state-space models. Journal of Experimental MarineBiology and Ecology, 356, 128e135.
Batschelet, E. 1981. Circular Statistics in Biology. New York: Academic Press.Bost, C. A., Cotté, C., Bailleul, F., Cherel, Y., Charrassin, J.-B., Guinet, C.,
Ainley, D. G. & Weimerskirch, H. 2009. The importance of oceanographicfronts to marine birds and mammals of the southern oceans. Journal of MarineSystems, 78, 363e376.
Chapman, J. W., Reynolds, D. R., Mouritsen, H., Hill, J. K., Riley, J. R., Sivell, D.,Smith, A. D. & Woiwod, I. P. 2008. Wind selection and drift compensationoptimize migratory pathways in a high-flying moth. Current Biology, 18, 514e518.
Chapman, J. W., Nesbit, R. L., Burgin, L. E., Reynolds, D. R., Smith, A. D.,Middleton, D. R. & Hill, J. K. 2010. Flight orientation behaviours promoteoptimal migration trajectories in high-flying insects. Science, 327, 682e685.
Chapman, J. W., Klaassen, R. H., Drake, V. A., Fossette, S., Hays, G. C.,Metcalfe, J. D., Reynolds, A. M., Reynolds, D. R. & Alerstam, T. 2011. Animalorientation strategies for movement in flows. Current Biology, 21, R861eR870.
Codling, E. A., Planck, M. J. & Benhamou, S. 2008. Randomwalks model in biology.Journal of the Royal Society Interface, 5, 813e834.
Cotté, C., Park, Y. H., Guinet, C. & Bost, C. A. 2007. Movements of foraging kingpenguins through marine mesoscale eddies. Proceedings of the Royal Society B,274, 2385e2391.
Doyle, T. K., Houghton, J. D. R., O’Suilleabhain, P. F., Hobson, V. J., Marnell, F.,Davenport, J. & Hays, G. C. 2008. Leatherback turtles satellite-tagged in Euro-pean waters. Endangered Species Research, 4, 23e31.
Eckert, K. L., Wallace, B. P., Frazier, J. G., Eckert, S. A. & Pritchard, P. C. H. 2012.Synopsis of the Biological Data on the Leatherback Sea Turtle (Dermochelys cor-iacea). Washington, D.C.: U.S. Department of Interior, Fish and Wildlife Service
Eckert, S. A. 2002. Swim speed and movement patterns of gravid leatherback seaturtles (Dermochelys coriacea) at St Croix, US Virginian Islands. Journal ofExperimental Biology, 205, 3689e3697.
Eckert, S. A. 2006. High-use oceanic areas for Atlantic leatherback sea turtles(Dermochelys coriacea) as identified using satellite telemetered location anddive information. Marine Biology, 149, 1257e1267.
Eckert, S. A., Bagley, D. A., Kubis, S., Ehrhart, L. M., Johnson, C., Stewart, K. &DeFreese, D. 2006. Internesting and postnesting movements and foraginghabitats of leatherback sea turtles (Dermochelys coriacea) nesting in Florida.Chelonian Conservation and Biology, 5, 239e248.
Ferraroli, S., Georges, J. Y., Gaspar, P. & Le Maho, Y. 2004. Where leatherbackturtles meet fisheries. Nature, 429, 521e522.
Fossette, S., Corbel, H., Gaspar, P., Le Maho, Y. & Georges, J. Y. 2008. An alternativetechnique for the long-term satellite tracking of leatherback turtles. EndangeredSpecies Research, 4, 33e41.
Fossette, S., Hobson, V. J., Girard, C., Calmettes, B., Gaspar, P., Georges, J. Y. &Hays, G. C. 2010a. Spatio-temporal foraging patterns of a giant zooplanktivore,the leatherback turtle. Journal of Marine Systems, 81, 225e234.
Fossette, S., Girard, C., López-Mendilaharsu, M., Miller, P., Domingo, A., Evans, D.,Kelle, L., Plot, V., Prosdocimi, L. & Verhage, S. 2010b. Atlantic leatherbackmigratory paths and temporary residence areas. PLoS ONE, 5, e13908.
Fossette, S., Putman, N. F., Lohmann, K. J., Marsh, R. & Hays, G. C. 2012.A biologist’s guide to assessing ocean currents: a review. Marine Ecology Prog-ress Series, 457, 285e301.
Gaspar, P., George, J. C., Fossette, S., Lenoble, A., Ferraroli, S. & Le Maho, Y. 2006.Marine animal behaviour: neglecting ocean currents can lead us up the wrongtrack. Proceedings of the Royal Society B, 273, 2697e2702.
Girard, C., Sudre, J., Benhamou, S., Roos, D. & Luschi, P. 2006. Homing in greenturtles Chelonia mydas: oceanic currents act as a constraint rather than as aninformation source. Marine Ecology Progress Series, 322, 281e289.
Girard, C., Tucker, A. D. & Calmettes, B. 2009. Post-nesting migrations of logger-head sea turtles in the Gulf of Mexico: dispersal in highly dynamic conditions.Marine Biology, 156, 1827e1839.
Godley, B. J., Blumenthal, J. M., Broderick, A. C., Coyne, M., Godfrey, M. H.,Hawkes, L. A. & Witt, M. J. 2008. Satellite tracking of sea turtles: where havewe been and where do we go next? Endangered Species Research, 4, 3e22.
Green, M. & Alerstam, T. 2002. The problem of estimating wind drift in migratingbirds. Journal of Theoretical Biology, 218, 485e496.
Hays, G. C., Houghton, J. D. R. & Myers, A. E. 2004a. Pan-Atlantic leatherback turtlemovements. Nature, 429, 522.
Hays, G. C., Houghton, J. D. R., Isaacs, C., King, R. S., Lloyd, C. & Lovell, P. 2004b.First records of oceanic dive profiles for leatherback turtles, Dermochelys cor-iacea, indicate behavioural plasticity associated with long-distance migration.Animal Behaviour, 67, 733e743.
Hays, G. C., Hobson, V. J., Metcalfe, J. D., Righton, D. & Sims, D. W. 2006. Flexibleforaging movements of leatherback turtles across the North Atlantic Ocean.Ecology, 87, 2647e2656.
Hazel, J. 2009. Evaluation of fast-acquisition GPS in stationary tests and fine-scaletracking of green turtles. Journal of Experimental Biology, 374, 58e68.
Hughes, G. R., Luschi, P., Mencacci, R. & Papi, F. 1998. The 7000-km oceanicjourney of a leatherback turtle tracked by satellite. Journal of ExperimentalMarine Biology and Ecology, 229, 209e217.
James, M. C., Eckert, S. A. & Myers, R. A. 2005a. Migratory and reproductivemovements of male leatherback turtles (Dermochelys coriacea). Marine Biology,147, 845e853.
James, M. C., Myers, R. A. & Ottensmeyer, C. A. 2005b. Behaviour of leatherback seaturtles, Dermochelys coriacea, during the migratory cycle. Proceedings of theRoyal Society B, 272, 1547e1555.
S. Galli et al. / Animal Behaviour 84 (2012) 1491e1500 1499

Author's personal copy
James, M. C., Davenport, J. & Hays, G. C. 2006. Expanded thermal niche for a divingvertebrate: a leatherback turtle diving into near-freezing water. Journal ofExperimental Marine Biology and Ecology, 335, 221e226.
Jones, T. T., Bostrom, B., Carey, M., Imlach, B., Mikkelsen, J., Ostafichuk, P.,Eckert, S., Opay, P., Swimmer, Y., Seminoff, J. A., et al. 2011. DeterminingTransmitter Drag and Best-practice Attachment Procedures for Sea Turtle Biote-lemetry Studies. Washington, D.C.: U.S. Department of Commerce
Jonsen, I. D., Myers, R. A. & James, M. C. 2007. Identifying leatherback turtleforaging behaviour from satellite telemetry using a switching state-spacemodel. Marine Ecology Progress Series, 337, 255e264.
Kai, E. T., Rossi, V., Sudre, J., Weimerskirch, H., Lopez, C., Hernandez-Garcia, E.,Marsac, F. & Garçon, V. 2009. Top marine predators track Lagrangian coherentstructures. Proceedings of the National Academy of Sciences, U.S.A., 106,8245e8250.
Karlsson, H., Henningsson, P., Bäckman, J., Hedenström, A. & Alerstam, T. 2010.Compensation for wind drift by migrating swifts. Animal Behaviour, 80,399e404.
Lambardi, P., Lutjeharms, J. R. E., Mencacci, R., Hays, G. C. & Luschi, P. 2008.Influence of ocean currents on long-distance movement of leatherback seaturtles in the Southwest Indian Ocean. Marine Ecology Progress Series, 353,289e301.
Lévy, M., Memery, L. & Madec, C. 1999. The onset of the spring bloom in theMEDOC area: mesoscale spatial variability. Deep Sea Research, 46, 1137e1160.
Lima, I. D., Olson, D. B. & Doney, S. C. 2002. Biological response to frontal dynamicsand mesoscale variability in oligotrophic environments: biological productionand community structure. Journal of Geophysical Research, 107, C8, http://dx.doi.org/10.1029/2000JC000393.
Lohmann, K. J., Lohmann, C. M. F. & Endres, C. S. 2008a. The sensory ecology ofocean navigation. Journal of Experimental Biology, 211, 1719e1728.
Lohmann, K. J., Luschi, P. & Hays, G. C. 2008b. Goal navigation and island-findingin sea turtles. Journal of Experimental Marine Biology and Ecology, 356, 83e95.
Luschi, P., Hays, G. C. & Papi, F. 2003a. A review of long-distance movements bymarine turtles, and the possible role of ocean currents. Oikos, 103, 293e302.
Luschi, P., Sale, A., Mencacci, R., Hughes, G. R., Lutjeharms, J. R. E. & Papi, F.2003b. Current transport in leatherback sea turtles (Dermochelys coriacea) inthe ocean. Proceedings of the Royal Society B Supplement, 270, 129e132.
Luschi, P., Lutjeharms, J. R. E., Lambardi, P., Mencacci, R., Hughes, G. R. &Hays, G. C. 2006. A review of migratory behaviour of sea turtles off southernAfrica. South African Journal of Science, 102, 51e58.
Luschi, P., Benhamou, S., Girard, C., Ciccione, S., Roos, D., Sudre, J. & Benvenuti, S.2007. Marine turtles use geomagnetic cues during open-sea homing. CurrentBiology, 17, 126e133.
Lutcavage, M. E. 1996. Planning your next meal: leatherback travel routes andocean fronts. In: Proceedings of the 15th Annual Symposium on Sea Turtle Biologyand Conservation (Ed. by J. A. Keinath, D. E. Barnard, J. A. Musick & B. A. Bell), pp.174e178. Miami: National Oceanic and Atmospheric Administration.
Lutcavage, M. E. & Lutz, P. L. 1997. Diving behaviour. In: The Biology of Sea Turtles(Ed. by P. L. Lutz & J. A. Musick), pp. 277e296. Boca Raton: CRC Press.
McMahon, C. R. & Hays, G. C. 2006. Thermal niche, large-scale movements andimplications of climate change for a critically endangered marine vertebrate.Global Change Biology, 12, 1330e1338.
Mills-Flemming, J., Field, C. A., James, M. C., Jonsen, I. D. & Myers, R. A. 2010.Hierarchical state-space estimation of leatherback turtle navigation ability. PLoSONE, 5, e14245.
Olson, D. B., Hitchcock, G. L., Mariano, A. J., Ashjian, C., Peng, G., Nero, R. W. &Podestà, G. 1994. Life on the edge: marine life and fronts. Oceanography, 7,52e60.
Pascual, A., Faugere, Y., Larnicol, G. & Le Traon, P.-Y. 2006. Improved descriptionof the ocean mesoscale variability by combining four satellite altimeters.Geophysical Research Letters, 33, 13e16, http://dx.doi.org/10.1029/2005GL024633.
Richardson, W. J. 1991. Wind and orientation of migrating birds: a review. In:Orientation in Birds (Ed. by P. Berthold), pp. 226e249. Basel: Birkhauser Verlag.
Rio, M.-H. & Hernandez, F. 2003. High-frequency response of wind-driven currentsmeasured by drifting buoys and altimetry over the world ocean. Journal ofGeophysical Research, 108, C8, 3283, http://dx.doi.org/10.1029/2002JC001655.
Rio, M.-H. & Hernandez, F. 2004. A mean dynamic topography computed over theworld ocean from altimetry, in situ measurements, and a geoid model. Journalof Geophysical Research, 109, C12032, http://dx.doi.org/10.1029/2003JC002226.
Robel, A. A., Lozier, M. S., Gary, S. F., Shillinger, G. L., Bailey, H. & Bograd, S. J. 2011.Projecting uncertainty onto marine megafauna trajectories. Deep Sea Research,58, 915e921.
Sale, A. & Luschi, P. 2009. Navigational challenges in the oceanic migrations ofleatherback sea turtles. Proceedings of the Royal Society B, 276, 3737e3745.
Sale, A., Luschi, P., Mencacci, R., Lambardi, P., Hughes, G. R. & Hays, G. C. 2006.Long-term monitoring of leatherback turtle diving behaviour during oceanicmovements. Journal of Experimental Marine Biology and Ecology, 328, 197e201.
Sato, K., Watanuki, Y., Takahashi, A., Miller, P. J. O., Tanaka, H., Kawabe, R.,Ponganis, P. J., Handrich, Y., Akamatsu, T., Watanabe, Y., et al. 2007. Strokefrequency, but not swimming speed, is related to body size in free-rangingseabirds, pinnipeds and cetaceans. Proceedings of the Royal Society B, 274,471e477.
Shillinger, G. L., Palacios, D. M., Bailey, H., Bograd, S. J., Swithenbank, A. M.,Gaspar, P., Wallace, B. P., Spotila, J. R., Paladino, F. V., Piedra, R., et al. 2008.Persistent leatherback turtle migrations present opportunities for conservation.PLoS Biology, 6, e171.
Sudre, J. & Morrow, R. A. 2008. Global surface currents: a high-resolution productfor investigating ocean dynamics. Ocean Dynamics, 58, 101e118.
Witt,M. J., Åkesson, S., Broderick, A. C., Coyne,M. S., Ellick, J., Formia, A., Hays, G. C.,Luschi, P., Stroud, S. & Godley, B. J. 2010. Assessing accuracy and utility ofsatellite-trackingdatausingArgos-linkedFastloc-GPS.AnimalBehaviour,80,1e11.
Witt, M. J., Bonguno, E. A., Broderick, A. C., Coyne, M. S., Formia, A., Gibudi, A.,Mounguengui, G. A. M., Moussounda, C., Nsafou, M., Nougessono, S., et al.2011. Tracking leatherback turtles from the world’s largest rookery: assessingthreats across the South Atlantic. Proceedings of the Royal Society B, 278,2338e2347.
S. Galli et al. / Animal Behaviour 84 (2012) 1491e15001500




















