
Dive and beak movement patterns in leatherback turtles Dermochelys coriacea during internesting...
11
Journal of Animal Ecology 2008, 77, 236–246 doi: 10.1111/j.1365-2656.2008.01344.x © 2008 The Authors. Journal compilation © 2008 British Ecological Society Blackwell Publishing Ltd Dive and beak movement patterns in leatherback turtles Dermochelys coriacea during internesting intervals in French Guiana Sabrina Fossette 1 *, Philippe Gaspar 2 , Yves Handrich 1 , Yvon Le Maho 1 and Jean-Yves Georges 1 1 IPHC-Département Ecologie, Physiologie, Ethologie, ULP, CNRS; 23 rue Becquerel, 67087 Strasbourg, France; and 2 Collecte Localization Satellites, Space Oceanography Division, Marine Ecosystem Modelling and Monitoring by Satellites, 8–10 rue Hermès, 31520 Ramonville St Agne, France Summary 1. Investigating the foraging patterns of free-ranging species is essential to estimate energy/time budgets for assessing their real reproductive strategy. Leatherback turtles Dermochelys coriacea (Vandelli 1761), commonly considered as capital breeders, have been reported recently to prospect actively during the breeding season in French Guiana, Atlantic Ocean. In this study we investigate the possibility of this active behaviour being associated with foraging, by studying concurrently diving and beak movement patterns in gravid females equipped with IMASEN (Inter-MAndibular Angle SENsor). 2. Four turtles provided data for periods varying from 7·3 to 56·1 h while exhibiting continuous short and shallow benthic dives. Beak movement (‘b-m’) events occurred in 34% of the dives, on average 1·8 ± 1·4 times per dive. These b-m events lasted between 1·5 and 20 s and occurred as isolated or grouped (two to five consecutive beak movements) events in 96·0 ± 4·0% of the recorded cases, and to a lesser extent in series (> five consecutive beak movements). 3. Most b-m events occurred during wiggles at the bottom of U- and W-shaped dives and at the beginning and end of the bottom phase of the dives. W-shaped dives were associated most frequently with beak movements (65% of such dives) and in particular with grouped beak movements. 4. Previous studies proposed wiggles to be indicator of predatory activity, U- and W-shaped dives being putative foraging dives. Beak movements recorded in leatherbacks during the first hours of their internesting interval in French Guiana may be related to feeding attempts. 5. In French Guiana, leatherbacks show different mouth-opening patterns for different dive patterns, suggesting that they forage opportunistically on occasional prey, with up to 17% of the dives appearing to be successful feeding dives. 6. This study highlights the contrasted strategies adopted by gravid leatherbacks nesting on the Pacific coasts of Costa Rica, in the deep-water Caribbean Sea and in the French Guianan shallow continental shelf, and may be related to different local prey accessibility among sites. Our results may help to explain recently reported site-specific individual body size and population dynamics. Key-words: alternative reproductive strategy, Atlantic Ocean, foraging, hall sensor, sea turtle Introduction The resources (water, nutrients, energy) acquired by organisms in the wild are limited and cannot be allocated simultaneously to survival, growth and reproduction (Stearns 1992). Organisms are thus predicted to adopt strategies for facing trade-offs between these competing life-history traits (Stearns 1992). For instance, organisms compensate for the high energy demands of reproduction by increasing the amount of total food resources available through two main strategies: schemat- ically, capital breeders store body reserves prior to reproduction and rely exclusively on them during breeding; conversely, income breeders show limited storage capacities and adjust their food intake during reproduction (Jönsson 1997). However, in some cases where body reserves do not fulfil reproductive *Correspondence author: E-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Dive and beak movement patterns in leatherback turtles Dermochelys coriacea during internesting...

Journal of Animal Ecology
2008,
77
, 236–246 doi: 10.1111/j.1365-2656.2008.01344.x
© 2008 The Authors. Journal compilation © 2008 British Ecological Society
Blackwell Publishing Ltd
Dive and beak movement patterns in leatherback turtles
Dermochelys coriacea
during internesting intervals in French Guiana
Sabrina Fossette
1
*, Philippe Gaspar
2
, Yves Handrich
1
, Yvon Le Maho
1
and Jean-Yves Georges
1
1
IPHC-Département Ecologie, Physiologie, Ethologie, ULP, CNRS; 23 rue Becquerel, 67087 Strasbourg, France; and
2
Collecte Localization Satellites, Space Oceanography Division, Marine Ecosystem Modelling and Monitoring by Satellites,
8–10 rue Hermès, 31520 Ramonville St Agne, France
Summary
1.
Investigating the foraging patterns of free-ranging species is essential to estimate energy/timebudgets for assessing their real reproductive strategy. Leatherback turtles
Dermochelys coriacea
(Vandelli 1761), commonly considered as capital breeders, have been reported recently to prospectactively during the breeding season in French Guiana, Atlantic Ocean. In this study we investigatethe possibility of this active behaviour being associated with foraging, by studying concurrently divingand beak movement patterns in gravid females equipped with IMASEN (Inter-MAndibular AngleSENsor).
2.
Four turtles provided data for periods varying from 7·3 to 56·1 h while exhibiting continuousshort and shallow benthic dives. Beak movement (‘b-m’) events occurred in 34% of the dives, onaverage 1·8 ± 1·4 times per dive. These b-m events lasted between 1·5 and 20 s and occurred asisolated or grouped (two to five consecutive beak movements) events in 96·0 ± 4·0% of the recordedcases, and to a lesser extent in series (> five consecutive beak movements).
3.
Most b-m events occurred during wiggles at the bottom of U- and W-shaped dives and at thebeginning and end of the bottom phase of the dives. W-shaped dives were associated most frequentlywith beak movements (65% of such dives) and in particular with grouped beak movements.
4.
Previous studies proposed wiggles to be indicator of predatory activity, U- and W-shaped divesbeing putative foraging dives. Beak movements recorded in leatherbacks during the first hours oftheir internesting interval in French Guiana may be related to feeding attempts.
5.
In French Guiana, leatherbacks show different mouth-opening patterns for different dive patterns,suggesting that they forage opportunistically on occasional prey, with up to 17% of the divesappearing to be successful feeding dives.
6.
This study highlights the contrasted strategies adopted by gravid leatherbacks nesting on thePacific coasts of Costa Rica, in the deep-water Caribbean Sea and in the French Guianan shallowcontinental shelf, and may be related to different local prey accessibility among sites. Our resultsmay help to explain recently reported site-specific individual body size and population dynamics.
Key-words:
alternative reproductive strategy, Atlantic Ocean, foraging, hall sensor, sea turtle
Introduction
The resources (water, nutrients, energy) acquired by organismsin the wild are limited and cannot be allocated simultaneouslyto survival, growth and reproduction (Stearns 1992). Organismsare thus predicted to adopt strategies for facing trade-offs
between these competing life-history traits (Stearns 1992).For instance, organisms compensate for the high energydemands of reproduction by increasing the amount of totalfood resources available through two main strategies: schemat-ically, capital breeders store body reserves prior to reproductionand rely exclusively on them during breeding; conversely,income breeders show limited storage capacities and adjusttheir food intake during reproduction (Jönsson 1997). However,in some cases where body reserves do not fulfil reproductive
*Correspondence author: E-mail: [email protected]

Dive and beak movement patterns in leatherbacks
237
© 2008 The Authors. Journal compilation © 2008 British Ecological Society,
Journal of Animal Ecology
,
77
, 236–246
requirements, certain organisms considered commonly as capitalbreeders may refuel by feeding during the breeding season.Several examples of this intermediate strategy have been reportedin birds (Durant, Massemin & Handrich 2004), ungulates(Byers & Moodie 1990), pinnipeds (Boness, Bowen & Oftedal1994) and sea turtles (Hays
et al
. 2002; Myers & Hays 2006).Assessing real reproductive strategy by investigating the
foraging patterns during and outside the breeding season isessential for the reliable estimation of energy and time budgetsin order to establish ultimately how organisms ensure theirreproduction and their own survival optimally (e.g. Boyd,Lunn & Barton 1991; Challet
et al
. 1994; Hochscheid
et al
.1999; Georges & Guinet 2000). This has obvious consequencesin terms of conservation in the case of species that feed onsites overlapping with human activities (e.g. Delord
et al
.2005; Fossette
et al
. 2007b). Accordingly, there has been ageneral impetus to investigate in much greater detail theforaging behaviour of free-ranging species during their life cycle.In the case of marine species, where foraging occurs mainlyunderwater during dives, investigating foraging behaviour(i.e. diving pattern and actual feeding activity) is particularlydifficult due to the limited accessibility of marine environments(Kooyman
et al
. 1992). Consequently, foraging behaviour inmarine vertebrates has been explored in different waysbased on two-dimensional time–depth dive profiles and three-dimensional compass (e.g. Schreer & Testa 1996; Le Boeuf
et al
. 2000; Mitani
et al
. 2003), echolocation buzzes in spermwhales
Physeter macrocephallus
(e.g. Miller
et al
. 2004;Watwood
et al
. 2006), stomach/oesophageal/gastrointestinaltemperature (e.g. Ancel, Horning & Kooyman 1997; South-wood
et al
. 2005) or underwater video (e.g. Reina
et al
. 2005;Watanabe
et al
. 2006). One of the most recent techniques is therecording of dive depth in conjunction with beak-openingevents (e.g. Wilson
et al
. 2002; Wilson
et al
. 2003; Hochscheid
et al
. 2004; Myers & Hays 2006; Liebsch
et al
. 2007). Thislatest approach uses a magnet/Hall sensor combinationconnected to a data-logger that records changes in the inter-mandible angle with a high frequency [Inter-MAndibularAngle SENsor (IMASEN), see Wilson
et al
. 2002].IMASENs have been proved to be a reliable system to
investigate fine-scale ingestion and/or respiratory patterns inrelation to diving behaviour. Given the relative difficulty ofIMASEN deployment, this technique has been used mainlyin captive animals (e.g. Ropert-Coudert
et al
. 2002, 2004;Wilson
et al
. 2002, 2003; Hochscheid
et al
. 2004; Liebsch
et al
. 2007). Data on free-living individuals concern only fourdifferent species of marine animals:
Leptonychotes weddellii
(the Weddell seal,
n
= 7 individuals, Plötz
et al
. 2001; Liebsch
et al
. 2007),
Spheniscus magellanicus
(the Magellanic penguin,
n
= 19 individuals, Wilson
et al
. 2002, 2003; Simeone &Wilson 2003),
Pygoscelis antarctica
(the Chinstrap penguin,
n
= 1 individual, Takahashi
et al
. 2004) and
Dermochelyscoriacea
(the leatherback turtle,
n
= 1 individual, Myers &Hays 2006).
In sea turtles, reproduction consists of successive oviposi-tions on land separated by internesting intervals of a few days,during which turtles remain at sea for egg maturation. Sea
turtles are considered commonly to be capital breeders, i.e.they ensure reproduction by relying on body reserves storedprior to reproduction during migration (Miller 1997). However,previous studies suggest that leatherbacks nesting in theAtlantic Ocean may forage during the nesting season (Eckert
et al
. 1989; Hays
et al
. 2004; Fossette
et al
. 2007b; Georges
et al
. 2007). This suggestion has been supported by Myers &Hays (2006), who reported some rhythmic beak movementsin a single leatherback equipped with an IMASEN duringthe nesting season on Grenada Island (Atlantic Ocean).These rhythmic beak movements occurred as the animal wasdescending to depth and have been interpreted as potentialattempts to detect prey by relying on gustatory cues to sensethe surrounding waters (Myers & Hays 2006). Such putativeforaging behaviour in Atlantic leatherbacks may help tocompensate for particularly high reproductive costs. How-ever, in the Pacific Ocean leatherbacks nesting in Costa Ricaappear to compensate for high reproductive costs (Wallace
et al
. 2007) by reducing their activity (Reina
et al
. 2005) andtheir metabolic rate (Wallace
et al
. 2005) during internestingintervals. These two different strategies may be linked withdifferent reproductive outputs, i.e. fewer nests and fewereggs in Pacific populations (Wallace
et al
. 2006) and/or withdifferent oceanographic conditions and local food availability(Hays
et al
. 2002; Saba
et al
. 2007).The Atlantic Ocean hosts the world’s two largest leatherback
nesting populations, found within the large estuary of theMaroni river at the border between French Guiana andSuriname (South America) and on the coasts of Gabon (westernAfrica, review in Fossette
et al
. in press). In the Guianasregion, the leatherback’s nesting season coincides with thepeak of Amazonian influence where rich and turbid watermasses cross off the continental shelf, enhancing biologicalproduction (Froidefond
et al
. 2002; Baklouti
et al
. 2007;S. Fossette, T. Bastian, F. Blanchard, C. Girard, Y. Le Maho,P. Vendeville & J.-Y. Georges, unpublished data). This isillustrated by massive stranding of jellyfish on the leatherbackrookery of Awala-Yalimapo beach (French Guiana) and bythe presence of large amounts of gelatinous plankton near theseabed in shallow waters during the leatherback nestingseason (S. Fossette, T. Bastian, F. Blanchard, C. Girard, Y. LeMaho, P. Vendeville & J.-Y. Georges, unpublished data). Asleatherbacks feed primarily on jellyfish (James & Herman2001), this suggests favourable local trophic conditions forgravid females feeding during the nesting season in FrenchGuiana. Consistently, leatherbacks have been reported todisperse widely across the Guiana continental shelf (Georges
et al
. 2007) and to dive continuously day and night, concen-trating mainly on the area close to the seabed where theyperform numerous vertical excursions of several metres ofamplitude at the bottom of the dives (hereafter called‘wiggles’; Fossette
et al
. 2007b). Wiggles are interpreted com-monly as prospecting and foraging behaviour (e.g. Schreer,Kovacs & O’Hara Hines 2001; Simeone & Wilson 2003), so ithas been suggested that leatherbacks may attempt to feed onthe Guiana continental shelf (Fossette
et al
. 2007b). Thishypothesis is supported further by the fact that leatherbacks

238
S. Fossette
et al.
© 2008 The Authors. Journal compilation © 2008 British Ecological Society,
Journal of Animal Ecology
,
77
, 236–246
nesting in French Guiana are larger and heavier compared tothose present on other nesting sites, suggesting either that theyforage during the nesting season and/or that there is a site-specific growth strategy in this species (Georges & Fossette2006). However, there is no direct evidence to date of leather-backs actually feeding in French Guiana.
Here, we test the hypothesis that in French Guiana gravidleatherbacks do forage during the nesting season by jointlyinvestigating the fine-scale diving and beak movementpatterns. We predict that if leatherbacks in French Guianaattempt to forage during the nesting season, mouth-openingsshould occur at least during the descent phase of dives fordetecting prey (as reported by Myers & Hays 2006), but alsoduring wiggles performed at the bottom of benthic dives(as suggested by Fossette
et al
. 2007b).
Materials and methods
Fieldwork was conducted during the leatherback’s nesting season inMay–June 2006 at Awala-Yalimapo beach (5·7
°
N–53·9
°
W), FrenchGuiana, South America.
INSTRUMENT
DEPLOYMENT
Twelve turtles were selected opportunistically during our nightpatrolling on the beach and equipped with an IMASEN (Terralog,JUV Elektronic; Borstel, Germany). The IMASEN consisted of athree-channel data logger (70
×
50
×
20 mm, 55 g in air, data resolution:16 bit, memory size: 128 Mbit), sampling at the same frequency(8 Hz for six turtles and 4 Hz for six other turtles) depth (range:0–200 m) and strength of the magnetic field from which jaw move-ments can be extrapolated. The logger was cast in resin and poweredby a lithium 3·6-V battery (Sonnenschein Lithium GmbH, BüdingenGermany). Theoretically, these sampling protocols allow 92 and184 h of recording for 8 and 4 Hz sampling frequency, respectively,the total amount of data stored being limited by instrument memorycapacity rather than by the battery.
The devices were attached at night during oviposition which tookapproximately 10 min. The logger was fixed on the pseudo-carapaceusing 4·5 mm wide plastic nylon ties introduced in 6 mm diameterholes drilled in a lateral dorsal ridge (similar to Fossette
et al
.2007a). The logger was connected with a 1 m cable to a Hall sensor(15
×
5
×
3 mm), highly sensitive to magnetic field, set in epoxy andglued onto the upper jaw directly onto the skin (see Myers & Hays2006). A small, rare earth magnet (10
×
10
×
2 mm) also set in epoxywas glued onto the lower jaw face to the Hall sensor, directly onto theskin using cyanoacrylite glue (see Myers & Hays 2006). Dimensionsof the Hall sensor and the magnet set in epoxy were approximately30
×
15
×
3 mm, necessitating a minimum distance of 2·5 cm betweensensor and magnet locations.
IMASENs are based on the Hall effect, where an increase inthe distance between the Hall sensor and the magnet (as the beakopened) results in a decrease in the magnetic field strength, recordedby the logger as a drop in voltage. The Hall sensor output can thusbe related theoretically to the intermandible distance by calibratingthe logger before deployment once fixed on the turtle. For each logger,an exponential relationship was obtained between the magnetic fieldstrength and the intermandible distance, with differences notedamong loggers due to different sensitivity and position of the sensorrelative to the magnet. However, for all loggers, the magnetic field
strength did not vary for intermandible distances larger than 3·5 cm.According to the narrow reading window (2·5–3·5 cm), most (> 95%)of the recorded data could not be used for quantitatively estimatingmouth-opening amplitude. In addition, the baseline of the Hallsignal was affected highly by pressure and changed parallel to thediving profile (Fig. 1). Simple mathematical high-pass filtering (butter-high and filtfilt functions in
version 6, > 6 s) was used tosmooth the voltage baseline (Fig. 1). However, due to the above-mentioned patterns, we did not investigate the amplitude changepatterns of the Hall signal. Consequently, this study is based on thepattern of beak-opening event occurrence.
DATA
ANALYSIS
IMASENs data were analysed using MT Dive software (JensenSoftware Systems 2006, Laboe, Germany). Individual dives weredefined as any depth exceeding 3 m. For each dive, the programcalculated the start and end time, the maximum depth reached duringthe dive, dive duration, the post-dive surface interval, time spent atthe bottom of the dive, the number of wiggles and the number ofdive steps. The bottom phase was defined as the period during which(a) depth was greater than 70% of maximum depth of a given dive and(b) the vertical speed was less than 0·3 ms
–1
. Wiggles were definedfor any event involving a rapid opposite change in depth > 1·2 m(equivalent to
c
. twice the maximum ventro-dorsal height of aleatherback turtle; Georges & Fossette 2006) during the bottom phaseof the dive. ‘Dive steps’ were defined as any events where verticalrate was less than 0·3 ms
–1
during the descent or ascent phase of thedive (Hochscheid
et al
. 1999). Dives were then classified into threetypes according to their profiles (see Fossette
et al
. 2007b) – ‘V-type’and ‘U-type’ dives – where bottom phase duration was < 30% and> 30% of dive duration, respectively, and ‘W-type’ (U-type dive showingat least one wiggle with an amplitude of 40% of the maximum wiggleamplitude recorded for a given turtle).
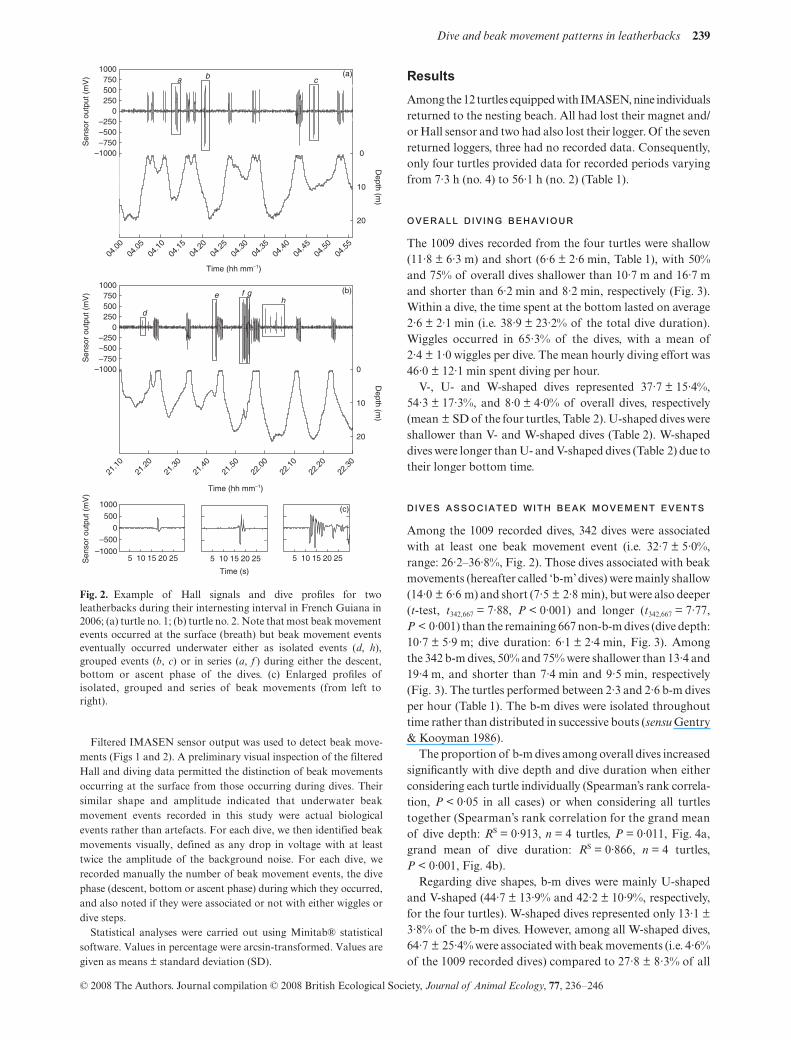
Fig. 1. Example of a record of pressure and Hall signal obtained ina leatherback turtle concurrently equipped with a time depthrecorder and an Inter-MAndibular Angle SENsor (IMASEN)during one internesting interval in French Guiana in 2006. Note thatthe raw Hall signal (bottom) was highly affected by pressure(middle), which implied to filter Hall data (top). Consequently, beakmovement patterns were investigated in terms of occurrence ratherthan quantitatively (see Methods).
Dive and beak movement patterns in leatherbacks
239
© 2008 The Authors. Journal compilation © 2008 British Ecological Society,
Journal of Animal Ecology
,
77
, 236–246
Filtered IMASEN sensor output was used to detect beak move-ments (Figs 1 and 2). A preliminary visual inspection of the filteredHall and diving data permitted the distinction of beak movementsoccurring at the surface from those occurring during dives. Theirsimilar shape and amplitude indicated that underwater beakmovement events recorded in this study were actual biologicalevents rather than artefacts. For each dive, we then identified beakmovements visually, defined as any drop in voltage with at leasttwice the amplitude of the background noise. For each dive, werecorded manually the number of beak movement events, the divephase (descent, bottom or ascent phase) during which they occurred,and also noted if they were associated or not with either wiggles ordive steps.
Statistical analyses were carried out using Minitab® statisticalsoftware. Values in percentage were arcsin-transformed. Values aregiven as means ± standard deviation (SD).
Results
Among the 12 turtles equipped with IMASEN, nine individualsreturned to the nesting beach. All had lost their magnet and/or Hall sensor and two had also lost their logger. Of the sevenreturned loggers, three had no recorded data. Consequently,only four turtles provided data for recorded periods varyingfrom 7·3 h (no. 4) to 56·1 h (no. 2) (Table 1).
OVERALL
D IV ING
BEHAVIOUR
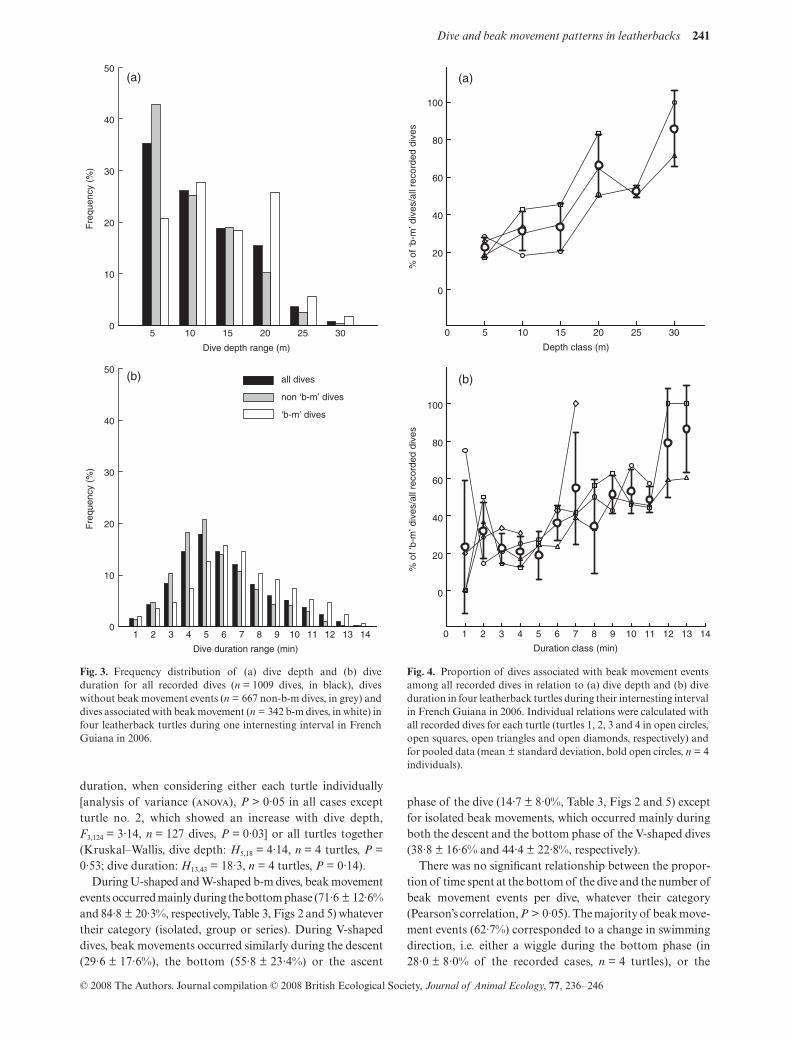
The 1009 dives recorded from the four turtles were shallow(11·8 ± 6·3 m) and short (6·6 ± 2·6 min, Table 1), with 50%and 75% of overall dives shallower than 10·7 m and 16·7 mand shorter than 6·2 min and 8·2 min, respectively (Fig. 3).Within a dive, the time spent at the bottom lasted on average2·6 ± 2·1 min (i.e. 38·9 ± 23·2% of the total dive duration).Wiggles occurred in 65·3% of the dives, with a mean of2·4 ± 1·0 wiggles per dive. The mean hourly diving effort was46·0 ± 12·1 min spent diving per hour.
V-, U- and W-shaped dives represented 37·7 ± 15·4%,54·3 ± 17·3%, and 8·0 ± 4·0% of overall dives, respectively(mean ± SD of the four turtles, Table 2). U-shaped dives wereshallower than V- and W-shaped dives (Table 2). W-shapeddives were longer than U- and V-shaped dives (Table 2) due totheir longer bottom time.
D IVES
ASSOCIATED
WITH
BEAK
MOVEMENT
EVENTS
Among the 1009 recorded dives, 342 dives were associatedwith at least one beak movement event (i.e. 32·7 ± 5·0%,range: 26·2–36·8%, Fig. 2). Those dives associated with beakmovements (hereafter called ‘b-m’ dives) were mainly shallow(14·0 ± 6·6 m) and short (7·5 ± 2·8 min), but were also deeper(
t
-test,
t
342,667
= 7·88,
P
< 0·001) and longer (
t
342,667
= 7·77,
P
< 0·001) than the remaining 667 non-b-m dives (dive depth:10·7 ± 5·9 m; dive duration: 6·1 ± 2·4 min, Fig. 3). Amongthe 342 b-m dives, 50% and 75% were shallower than 13·4 and19·4 m, and shorter than 7·4 min and 9·5 min, respectively(Fig. 3). The turtles performed between 2·3 and 2·6 b-m divesper hour (Table 1). The b-m dives were isolated throughouttime rather than distributed in successive bouts (
sensu
Gentry& Kooyman 1986).
The proportion of b-m dives among overall dives increasedsignificantly with dive depth and dive duration when eitherconsidering each turtle individually (Spearman’s rank correla-tion,
P
< 0·05 in all cases) or when considering all turtlestogether (Spearman’s rank correlation for the grand meanof dive depth:
R
S
= 0·913,
n
= 4 turtles,
P
= 0·011, Fig. 4a,grand mean of dive duration:
R
S
= 0·866,
n
= 4 turtles,P < 0·001, Fig. 4b).
Regarding dive shapes, b-m dives were mainly U-shapedand V-shaped (44·7 ± 13·9% and 42·2 ± 10·9%, respectively,for the four turtles). W-shaped dives represented only 13·1 ±3·8% of the b-m dives. However, among all W-shaped dives,64·7 ± 25·4% were associated with beak movements (i.e. 4·6%of the 1009 recorded dives) compared to 27·8 ± 8·3% of all
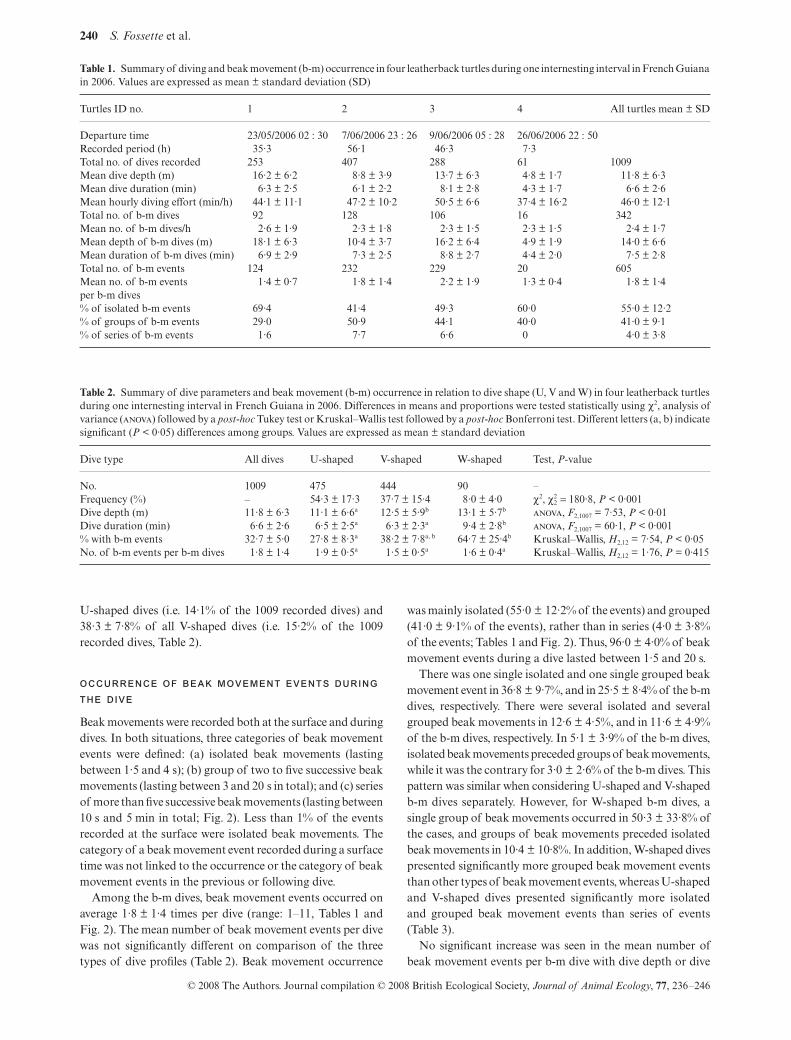
Fig. 2. Example of Hall signals and dive profiles for twoleatherbacks during their internesting interval in French Guiana in2006; (a) turtle no. 1; (b) turtle no. 2. Note that most beak movementevents occurred at the surface (breath) but beak movement eventseventually occurred underwater either as isolated events (d, h),grouped events (b, c) or in series (a, f ) during either the descent,bottom or ascent phase of the dives. (c) Enlarged profiles ofisolated, grouped and series of beak movements (from left toright).
240 S. Fossette et al.
© 2008 The Authors. Journal compilation © 2008 British Ecological Society, Journal of Animal Ecology, 77, 236–246
U-shaped dives (i.e. 14·1% of the 1009 recorded dives) and38·3 ± 7·8% of all V-shaped dives (i.e. 15·2% of the 1009recorded dives, Table 2).
OCCURRENCE OF BEAK MOVEMENT EVENTS DURING THE DIVE
Beak movements were recorded both at the surface and duringdives. In both situations, three categories of beak movementevents were defined: (a) isolated beak movements (lastingbetween 1·5 and 4 s); (b) group of two to five successive beakmovements (lasting between 3 and 20 s in total); and (c) seriesof more than five successive beak movements (lasting between10 s and 5 min in total; Fig. 2). Less than 1% of the eventsrecorded at the surface were isolated beak movements. Thecategory of a beak movement event recorded during a surfacetime was not linked to the occurrence or the category of beakmovement events in the previous or following dive.
Among the b-m dives, beak movement events occurred onaverage 1·8 ± 1·4 times per dive (range: 1–11, Tables 1 andFig. 2). The mean number of beak movement events per divewas not significantly different on comparison of the threetypes of dive profiles (Table 2). Beak movement occurrence
was mainly isolated (55·0 ± 12·2% of the events) and grouped(41·0 ± 9·1% of the events), rather than in series (4·0 ± 3·8%of the events; Tables 1 and Fig. 2). Thus, 96·0 ± 4·0% of beakmovement events during a dive lasted between 1·5 and 20 s.
There was one single isolated and one single grouped beakmovement event in 36·8 ± 9·7%, and in 25·5 ± 8·4% of the b-mdives, respectively. There were several isolated and severalgrouped beak movements in 12·6 ± 4·5%, and in 11·6 ± 4·9%of the b-m dives, respectively. In 5·1 ± 3·9% of the b-m dives,isolated beak movements preceded groups of beak movements,while it was the contrary for 3·0 ± 2·6% of the b-m dives. Thispattern was similar when considering U-shaped and V-shapedb-m dives separately. However, for W-shaped b-m dives, asingle group of beak movements occurred in 50·3 ± 33·8% ofthe cases, and groups of beak movements preceded isolatedbeak movements in 10·4 ± 10·8%. In addition, W-shaped divespresented significantly more grouped beak movement eventsthan other types of beak movement events, whereas U-shapedand V-shaped dives presented significantly more isolatedand grouped beak movement events than series of events(Table 3).
No significant increase was seen in the mean number ofbeak movement events per b-m dive with dive depth or dive
Table 1. Summary of diving and beak movement (b-m) occurrence in four leatherback turtles during one internesting interval in French Guianain 2006. Values are expressed as mean ± standard deviation (SD)
Turtles ID no. 1 2 3 4 All turtles mean ± SD
Departure time 23/05/2006 02 : 30 7/06/2006 23 : 26 9/06/2006 05 : 28 26/06/2006 22 : 50Recorded period (h) 35·3 56·1 46·3 7·3Total no. of dives recorded 253 407 288 61 1009Mean dive depth (m) 16·2 ± 6·2 8·8 ± 3·9 13·7 ± 6·3 4·8 ± 1·7 11·8 ± 6·3Mean dive duration (min) 6·3 ± 2·5 6·1 ± 2·2 8·1 ± 2·8 4·3 ± 1·7 6·6 ± 2·6Mean hourly diving effort (min/h) 44·1 ± 11·1 47·2 ± 10·2 50·5 ± 6·6 37·4 ± 16·2 46·0 ± 12·1Total no. of b-m dives 92 128 106 16 342Mean no. of b-m dives/h 2·6 ± 1·9 2·3 ± 1·8 2·3 ± 1·5 2·3 ± 1·5 2·4 ± 1·7Mean depth of b-m dives (m) 18·1 ± 6·3 10·4 ± 3·7 16·2 ± 6·4 4·9 ± 1·9 14·0 ± 6·6Mean duration of b-m dives (min) 6·9 ± 2·9 7·3 ± 2·5 8·8 ± 2·7 4·4 ± 2·0 7·5 ± 2·8Total no. of b-m events 124 232 229 20 605Mean no. of b-m events 1·4 ± 0·7 1·8 ± 1·4 2·2 ± 1·9 1·3 ± 0·4 1·8 ± 1·4per b-m dives% of isolated b-m events 69·4 41·4 49·3 60·0 55·0 ± 12·2% of groups of b-m events 29·0 50·9 44·1 40·0 41·0 ± 9·1% of series of b-m events 1·6 7·7 6·6 0 4·0 ± 3·8
Table 2. Summary of dive parameters and beak movement (b-m) occurrence in relation to dive shape (U, V and W) in four leatherback turtlesduring one internesting interval in French Guiana in 2006. Differences in means and proportions were tested statistically using χ2, analysis ofvariance () followed by a post-hoc Tukey test or Kruskal–Wallis test followed by a post-hoc Bonferroni test. Different letters (a, b) indicatesignificant (P < 0·05) differences among groups. Values are expressed as mean ± standard deviation
Dive type All dives U-shaped V-shaped W-shaped Test, P-value
No. 1009 475 444 90 –Frequency (%) – 54·3 ± 17·3 37·7 ± 15·4 8·0 ± 4·0 χ2, χ2
2 = 180·8, P < 0·001Dive depth (m) 11·8 ± 6·3 11·1 ± 6·6a 12·5 ± 5·9b 13·1 ± 5·7b , F2,1007 = 7·53, P < 0·01Dive duration (min) 6·6 ± 2·6 6·5 ± 2·5a 6·3 ± 2·3a 9·4 ± 2·8b , F2,1007 = 60·1, P < 0·001% with b-m events 32·7 ± 5·0 27·8 ± 8·3a 38·2 ± 7·8a, b 64·7 ± 25·4b Kruskal–Wallis, H2,12 = 7·54, P < 0·05No. of b-m events per b-m dives 1·8 ± 1·4 1·9 ± 0·5a 1·5 ± 0·5a 1·6 ± 0·4a Kruskal–Wallis, H2,12 = 1·76, P = 0·415
Dive and beak movement patterns in leatherbacks 241
© 2008 The Authors. Journal compilation © 2008 British Ecological Society, Journal of Animal Ecology, 77, 236–246
duration, when considering either each turtle individually[analysis of variance (), P > 0·05 in all cases exceptturtle no. 2, which showed an increase with dive depth,F3,124 = 3·14, n = 127 dives, P = 0·03] or all turtles together(Kruskal–Wallis, dive depth: H5,18 = 4·14, n = 4 turtles, P =0·53; dive duration: H13,43 = 18·3, n = 4 turtles, P = 0·14).
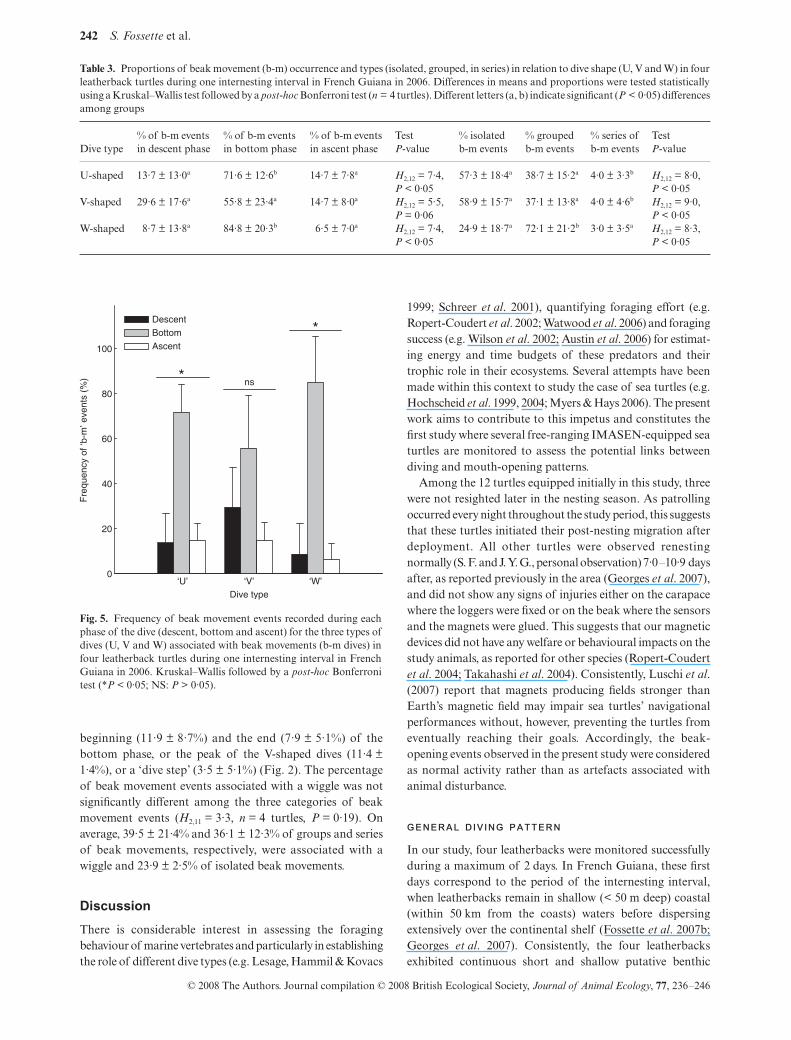
During U-shaped and W-shaped b-m dives, beak movementevents occurred mainly during the bottom phase (71·6 ± 12·6%and 84·8 ± 20·3%, respectively, Table 3, Figs 2 and 5) whatevertheir category (isolated, group or series). During V-shapeddives, beak movements occurred similarly during the descent(29·6 ± 17·6%), the bottom (55·8 ± 23·4%) or the ascent
phase of the dive (14·7 ± 8·0%, Table 3, Figs 2 and 5) exceptfor isolated beak movements, which occurred mainly duringboth the descent and the bottom phase of the V-shaped dives(38·8 ± 16·6% and 44·4 ± 22·8%, respectively).
There was no significant relationship between the propor-tion of time spent at the bottom of the dive and the number ofbeak movement events per dive, whatever their category(Pearson’s correlation, P > 0·05). The majority of beak move-ment events (62·7%) corresponded to a change in swimmingdirection, i.e. either a wiggle during the bottom phase (in28·0 ± 8·0% of the recorded cases, n = 4 turtles), or the
Fig. 3. Frequency distribution of (a) dive depth and (b) diveduration for all recorded dives (n = 1009 dives, in black), diveswithout beak movement events (n = 667 non-b-m dives, in grey) anddives associated with beak movement (n = 342 b-m dives, in white) infour leatherback turtles during one internesting interval in FrenchGuiana in 2006.
Fig. 4. Proportion of dives associated with beak movement eventsamong all recorded dives in relation to (a) dive depth and (b) diveduration in four leatherback turtles during their internesting intervalin French Guiana in 2006. Individual relations were calculated withall recorded dives for each turtle (turtles 1, 2, 3 and 4 in open circles,open squares, open triangles and open diamonds, respectively) andfor pooled data (mean ± standard deviation, bold open circles, n = 4individuals).
242 S. Fossette et al.
© 2008 The Authors. Journal compilation © 2008 British Ecological Society, Journal of Animal Ecology, 77, 236–246
beginning (11·9 ± 8·7%) and the end (7·9 ± 5·1%) of thebottom phase, or the peak of the V-shaped dives (11·4 ±1·4%), or a ‘dive step’ (3·5 ± 5·1%) (Fig. 2). The percentageof beak movement events associated with a wiggle was notsignificantly different among the three categories of beakmovement events (H2,11 = 3·3, n = 4 turtles, P = 0·19). Onaverage, 39·5 ± 21·4% and 36·1 ± 12·3% of groups and seriesof beak movements, respectively, were associated with awiggle and 23·9 ± 2·5% of isolated beak movements.
Discussion
There is considerable interest in assessing the foragingbehaviour of marine vertebrates and particularly in establishingthe role of different dive types (e.g. Lesage, Hammil & Kovacs
1999; Schreer et al. 2001), quantifying foraging effort (e.g.Ropert-Coudert et al. 2002; Watwood et al. 2006) and foragingsuccess (e.g. Wilson et al. 2002; Austin et al. 2006) for estimat-ing energy and time budgets of these predators and theirtrophic role in their ecosystems. Several attempts have beenmade within this context to study the case of sea turtles (e.g.Hochscheid et al. 1999, 2004; Myers & Hays 2006). The presentwork aims to contribute to this impetus and constitutes thefirst study where several free-ranging IMASEN-equipped seaturtles are monitored to assess the potential links betweendiving and mouth-opening patterns.
Among the 12 turtles equipped initially in this study, threewere not resighted later in the nesting season. As patrollingoccurred every night throughout the study period, this suggeststhat these turtles initiated their post-nesting migration afterdeployment. All other turtles were observed renestingnormally (S. F. and J. Y. G., personal observation) 7·0–10·9 daysafter, as reported previously in the area (Georges et al. 2007),and did not show any signs of injuries either on the carapacewhere the loggers were fixed or on the beak where the sensorsand the magnets were glued. This suggests that our magneticdevices did not have any welfare or behavioural impacts on thestudy animals, as reported for other species (Ropert-Coudertet al. 2004; Takahashi et al. 2004). Consistently, Luschi et al.(2007) report that magnets producing fields stronger thanEarth’s magnetic field may impair sea turtles’ navigationalperformances without, however, preventing the turtles fromeventually reaching their goals. Accordingly, the beak-opening events observed in the present study were consideredas normal activity rather than as artefacts associated withanimal disturbance.
GENERAL DIV ING PATTERN
In our study, four leatherbacks were monitored successfullyduring a maximum of 2 days. In French Guiana, these firstdays correspond to the period of the internesting interval,when leatherbacks remain in shallow (< 50 m deep) coastal(within 50 km from the coasts) waters before dispersingextensively over the continental shelf (Fossette et al. 2007b;Georges et al. 2007). Consistently, the four leatherbacksexhibited continuous short and shallow putative benthic
Table 3. Proportions of beak movement (b-m) occurrence and types (isolated, grouped, in series) in relation to dive shape (U, V and W) in fourleatherback turtles during one internesting interval in French Guiana in 2006. Differences in means and proportions were tested statisticallyusing a Kruskal–Wallis test followed by a post-hoc Bonferroni test (n = 4 turtles). Different letters (a, b) indicate significant (P < 0·05) differencesamong groups
Dive type% of b-m eventsin descent phase
% of b-m eventsin bottom phase
% of b-m eventsin ascent phase
Test P-value
% isolated b-m events
% groupedb-m events
% series ofb-m events
Test P-value
U-shaped 13·7 ± 13·0a 71·6 ± 12·6b 14·7 ± 7·8a H2,12 = 7·4, 57·3 ± 18·4a 38·7 ± 15·2a 4·0 ± 3·3b H2,12 = 8·0, P < 0·05 P < 0·05
V-shaped 29·6 ± 17·6a 55·8 ± 23·4a 14·7 ± 8·0a H2,12 = 5·5, 58·9 ± 15·7a 37·1 ± 13·8a 4·0 ± 4·6b H2,12 = 9·0, P = 0·06 P < 0·05
W-shaped 8·7 ± 13·8a 84·8 ± 20·3b 6·5 ± 7·0a H2,12 = 7·4, 24·9 ± 18·7a 72·1 ± 21·2b 3·0 ± 3·5a H2,12 = 8·3, P < 0·05 P < 0·05
Fig. 5. Frequency of beak movement events recorded during eachphase of the dive (descent, bottom and ascent) for the three types ofdives (U, V and W) associated with beak movements (b-m dives) infour leatherback turtles during one internesting interval in FrenchGuiana in 2006. Kruskal–Wallis followed by a post-hoc Bonferronitest (*P < 0·05; NS: P > 0·05).

Dive and beak movement patterns in leatherbacks 243
© 2008 The Authors. Journal compilation © 2008 British Ecological Society, Journal of Animal Ecology, 77, 236–246
dives and dived for more than 45 min per hour, as reportedpreviously in the area (Fossette et al. 2007b) and in other sites,both in the Atlantic and in the Pacific oceans (e.g. Eckert et al.1996; Southwood et al. 1999). Our four study leatherbacksperformed mainly U- or W-shaped dives (60% of all dives),spending about 30% of their time at the bottom of the divewhere they performed numerous wiggles. U- and W-shapeddives are commonly considered as foraging dives duringwhich animals spend time in a food patch, but they may alsobe associated with resting, as reported in the green turtleChelonia mydas (Hochscheid et al. 1999). Conversely, V-shapeddives are considered as travelling or exploration (e.g. Hindell,Slip & Burton 1991; Le Boeuf et al. 1992, 2000; Schreer & Testa1996; Lesage et al. 1999; Hays et al. 2002). The continuousbenthic diving associated with extended movements overthe continental shelf indicate that in French Guiana gravidleatherbacks are active rather than quiescent, contrary to thebehaviour reported in gravid leatherbacks on the Pacificcoasts of Costa Rica (Reina et al. 2005).
GENERAL BEAK MOVEMENT PATTERN
In our study all monitored leatherbacks showed beak-openingactivity, both at the surface and during dives, during the firsthours of their internesting interval. One-third of the dives wereassociated with beak movements that occurred irregularlyduring dives rather than rhythmically or in long seriesthrough time. In marine vertebrates, mouth-opening activitymay be involved in several kinds of behaviour: communication(e.g. Wilson et al. 2002), courtship (e.g. Booth & Peters 1972;Reina et al. 2005), drinking (e.g. Lutz 1997; Myers & Hays2006), foraging (e.g. Wilson et al. 2002; Hochscheid et al.2004; Takahashi et al. 2004; Myers & Hays 2006; Liebschet al. 2007) and breathing when beak movements occur at thesurface (e.g. Wilson et al. 2003; Reina et al. 2005).
For communication and courtship, social and sexualinteractions are initiated commonly by adult males in marinevertebrates (reviewed in Miller 1997 and Scott et al. 2005).Females are not generally observed acting aggressivelytowards adults of either sex (e.g. bottlenose dolphins Tursiopstruncatus, Scott et al. 2005; leatherbacks, Reina et al. 2005),with the exception of Caretta caretta (the loggerhead seaturtle), where female–female aggressive interactions may occureventually, yet in low frequency, on resting sites rather thanduring basking or swimming (Schofield et al. 2007). In FrenchGuiana, where leatherback turtles clearly do not rest duringtheir internesting intervals, the relatively high frequency of beakmovement events reported in the present study suggests thatthey may not be associated with sexual or social interactions.
Regarding drinking, adult sea turtles have been reportedto normally avoid drinking sea water directly (Lutz 1997;Southwood et al. 2005). This suggests that beak-openingevents observed in our study may not be associated withdrinking. However, if beak movements were associated withdrinking, their pattern should have been similar to that ofleatherbacks from different areas. The irregular pattern ofbeak movements observed in our four leatherbacks in French
Guiana contrasts, however, with the continuous rhythmicbeak movements reported in an IMASEN-equipped leather-back performing an internesting interval in the CaribbeanSea (Myers & Hays 2006). The beak movement patternsrecorded in our study may not, therefore, be associatedexclusively with drinking.
The last function of beak movement events may be associatedwith foraging. In our study, beak movement events occurredirregularly, primarily (62·3 ± 18·0%) as one isolated beakmovement or one group of beak movements during the dive,and secondly (24·3 ± 9·4%) as successive isolated beakmovements or groups of beak movements (see Fig. 2). Similarirregular patterns of mouth-opening events have been reportedin several species of marine vertebrates, where the number,duration, frequency and amplitude of mouth movementsdepends on whether the predator attempts then succeeds ornot to catch its prey, then manipulates, swallows or ingests it(Wilson et al. 2002; Hochscheid et al. 2004; Ropert-Coudertet al. 2004; Liebsch et al. 2007). In our study, beak-openingevents lasted mainly between 1·5 and 20 s. This is similar toresults obtained in captive loggerhead turtles feeding on crabs,where isolated beak movements last 1–5 s and a completefeeding cycle lasts from 10 to 50 s until prey ingestion,depending on prey type (Hochscheid et al. 2004). Similardurations of feeding events have been reported in free-livingpenguins (0·1–11 s; Wilson et al. 2002; Takahashi et al. 2004)and in free-ranging Weddell seals (0·6–30·7 s; Liebsch et al.2007), the interspecies differences in feeding duration probablybeing related to differences in prey size among species. Inall cases, captured prey is transported intra-orally thanks torepeated mouth-opening and -closing events. Consistently,the groups and series of successive beak movements observedin our study may correspond to successful feeding events andfood processing. Conversely, isolated beak movements maybe interpreted as unsuccessful capture attempts. Nevertheless,unsuccessful attacks are considered unlikely in our study, asleatherbacks feed mainly on slowly moving gelatinousplankton. Because isolated beak movements represent 55% ofthe beak movement events recorded in our study and mainlyprecede putative foraging grouped beak movements, isolatedbeak movements may, rather, permit the turtle to sense thesurrounding waters for detecting potential prey olfactorysignatures, as suggested by Myers & Hays (2006).
In our study, beak movements were also recorded at thesurface. They were mainly in groups and series and wereinterpreted classically as breathing events. However, if ananimal feeds at the surface this might be difficult to separatefrom the breathing signal. Leatherbacks have been observedfeeding on large jellyfish at the surface (James & Herman2001). Such observations have never been conducted in FrenchGuiana, probably because of the smaller size of jellyfish(unpublished data). In our study, there was no correlationbetween the type of beak-opening events at the surface andthe occurrence of beak movements during the preceding/following dive. This suggests that surface beak movementsreported in leatherbacks are probably not associated withfood processing. Nevertheless, feeding on subsurface prey

244 S. Fossette et al.
© 2008 The Authors. Journal compilation © 2008 British Ecological Society, Journal of Animal Ecology, 77, 236–246
cannot be excluded, even if this phenomenon has never beenobserved in French Guiana.
DIVE TYPE AND FORAGING EVENTS
The combined analysis of dive types and mouth-openingpatterns provides an additional means of determining under-water activity in free-ranging animals (e.g. Plötz et al. 2001;Liebsch et al. 2007). In our study, most of the beak movementevents, whatever their category, occurred during the bottomphase of U- and W-shaped dives. In penguins and elephantseals (Mirounga longirostris), beak-opening events associatedwith prey ingestion have been reported to occur mainly duringthe bottom phase of U- and W-shaped dives, rather thanduring the descending and ascending phases (Le Boeuf et al.1992; Wilson et al. 2002; Takahashi et al. 2004), suggestingthat animals swim directly to and from a depth where food islocated. This suggests that in our study, beak-opening eventsoccurring during U- and W-shaped dives may correspond toforaging events, as suggested by Fossette et al. (2007b).Additionally, the present study proposes the idea that, inFrench Guiana, leatherbacks may attempt to feed during theentire bottom phase of the dives, probably as long as they areclose to the seabed where their prey is concentrated (Artigaset al. 2003; S. Fossette, T. Bastian, F. Blanchard, C. Girard, Y.Le Maho, P. Vendeville & J.-Y. Georges, unpublished data).Indeed, dives associated with beak movements were slightlybut significantly deeper and longer than other dives, and theirproportion increased as turtles dived deeper and for longer.This suggests that in French Guiana, the probability for aleatherback to find prey increases as it dives for a longertime at deeper depth, at least during the first 2 days of theinternesting interval.
In French Guiana, beak movement events, whatever theircategory, occurred mainly when turtles were changing theirvertical angle, either during wiggles or at the beginning andthe end of the bottom phase. Additionally, W-shaped dives(i.e. dives with one or more ample wiggles), although lessfrequent than other dive types, were those associated mostfrequently with beak movements (65% of such dives) and inparticular with grouped beak movements. Previous studieshave shown that marine vertebrates capture prey mainly duringwiggles at the bottom phase (Schreer et al. 2001; Simeone &Wilson 2003; Takahashi et al. 2004) and that undulations inthe depth profile over time are associated with prey pursuit(Simeone & Wilson 2003; Takahashi et al. 2004). If this holdsfor leatherbacks, this suggests that the groups and series ofbeak movements associated with wiggles reported in ourstudy may be potential indicators of foraging success. Thissupports our recent hypothesis that W-shaped dives have aforaging function in leatherback turtles (Fossette et al. 2007b).Conversely, V-shaped dives were associated mainly withisolated beak movements during the descent and the bottomphases. In marine vertebrates, V-shaped dives are interpretedcommonly as exploration dives, a significant portion of thedescent being devoted to prey searching (e.g. Hindell et al.1991; Le Boeuf et al. 1992, 2000; Schreer & Testa 1996; Myers
& Hays 2006; Watwood et al. 2006). If this theory is alsoapplicable to leatherbacks, this would support the hypothesisthat leatherbacks taste the surrounding waters for detectingpotential prey as they descend to depths (see Myers & Hays2006), rather than actually feeding.
Conclusion
Our study shows that in French Guiana, gravid leatherbackturtles perform different patterns of mouth-opening associatedwith different dive patterns within the first days of theirinternesting intervals (this study), during which they disperseextensively over the continental shelf where they then performbenthic dives continuously (Fossette et al. 2007b; Georgeset al. 2007). This indicates that, in French Guiana, gravidleatherbacks do indeed feed, or at least attempt to do so, duringthe nesting season. Dives associated with beak movementevents were isolated throughout time, rather than distributedin successive dives inside a bout (sensu Gentry & Kooyman1986), suggesting that leatherbacks forage opportunisticallyon dispersed prey, with up to 17% of the dives appearing to besuccessful feeding dives. The actual function of the differentbeak-opening patterns is, however, more difficult to identify,and further validation is required either by in-captivity studiesor by video recording. Contrasted diving and foraging patternsamong leatherback populations from the Pacific coasts ofCosta Rica, deep-water Caribbean Sea and French Guianashallow continental shelf suggest that leatherbacks adoptdifferent strategies during the nesting season, most probablyin response to local food occurrence and distribution. InFrench Guiana, trophic conditions may permit leatherbacksto compensate for reproductive costs by feeding between twoconsecutive nesting events. This may help to explain therelative large body size of French Guianan leatherbacks(Georges & Fossette 2006), as well as the relatively highreproductive effort and the demography of this populationcompared to others sites (e.g. Girondot & Fretey 1996; Girondotet al. 2007; Fossette et al., in press). However, as leatherbacksstore most of their body reserves prior to reproduction, furtherstudies and developments are required to assess individualforaging success during their pan-oceanic migrations (Myers,Lovell & Hays 2006) using more integrated techniques involv-ing IMASEN-like technologies similar to those used success-fully in the present study.
Acknowledgements
We are grateful to the inhabitants of Awala-Yalimapo for their hospitality andto all those involved in the sea turtle monitoring programmes through theDirection Régionale de l’Environnement-Guyane, Kulalasi NGO, the RéserveNaturelle de l’Amana and WWF for their logistical contribution in the field. Wethank A. Avril, M. Boutrif, L. Gagnon, N. Gauter, M. Hamel, N. Hanuise, J.Marin-Pache, S. Martini, M. Laur and V. Plot for their assistance in the field.Financial support for S. F. was provided in the form of a studentship from theFrench Ministry of Research. Funding was provided by grants to J.-Y. G. fromthe European Union FEDER Program, the Contrat Plan Etat Région CPER/DocUp 2000–06, the ‘Programme Amazonie’ held by Centre National de laRecherche Scientifique. This study was carried out under CNRS-DEPEinstitutional licence (B67 482 18) with an individual licence attributed to toJ.-Y. G. (67–220 and 04–199) and S. F. (67–256). The study adhered strictly

Dive and beak movement patterns in leatherbacks 245
© 2008 The Authors. Journal compilation © 2008 British Ecological Society, Journal of Animal Ecology, 77, 236–246
to the legal requirements of the country in which the work was carried out andto all institutional guidelines.
References
Ancel, A., Horning, M. & Kooyman, G.L. (1997) Prey ingestion revealed byoesophagus and stomach temperature recordings in cormorants. Journal ofExperimental Biology, 200, 149–154.
Artigas, L.F., Vendeville, P., Leopold, M., Guiral, D. & Ternon, J.-F. (2003)Marine biodiversity in French Guiana: estuarine, coastal, and shelf ecosystemsunder the influence of Amazonian waters. Gayana, 67, 302–326.
Austin, D., Bowen, W.D., Mcmillan, J.I. & Boness, D.J. (2006) Stomachtemperature telemetry reveals temporal patterns of foraging success in afree-ranging marine mammal. Journal of Animal Ecology, 75, 408–420.
Baklouti, M., Devenon, J.-L., Bourret, A., Froidefond, J.-M., Ternon, J.-F. &Fuda, J.-L. (2007) New insights in the French Guiana continental shelfcirculation and its relation to the North Brazil Current retroflection. Journalof Geophysical Research, 112, C02023, doi: 10.1029/2006JC003520.
Boness, D.J., Bowen, W.D. & Oftedall, O.T. (1994) Evidence of a maternalforaging cycle resembling that of otariid seals in a small phocid, the harborseal. Behavioral Ecology and Sociobiology, 34, 95–104.
Booth, J. & Peters, J.A. (1972) Behavioural studies on the green turtle (Cheloniamydas) in the sea. Animal Behaviour, 20, 808–812.
Boyd, I.L., Lunn, N.J. & Barton, T. (1991) Time budgets and foragingcharacteristics of lactating antarctic fur seals. Journal of Animal Ecology, 60,577–592.
Byers, J.A. & Moodie, J.D. (1990) Sex-specific maternal investment in pronghornand the question of a limit on differential provisioning in ungulates.Behavioral Ecology and Sociobiology, 26, 157–164.
Challet, E., Bost, C.-A., Handrich, Y., Gendner, J.-P. & Le Maho, Y. (1994)Behavioural time budget of breeding king penguins (Aptenodytes patagonica).Journal of Zoology, 233, 669–681.
Delord, K., Gasco, N., Weimerskirch, H., Barbraud, C. & Micol, T. (2005) Sea-bird mortality in the Patagonian toothfish longline fishery around Crozetand Kerguelen Islands, 2001–03. CCAMLR Science, 12, 53–80.
Durant, J.M., Massemin, S. & Handrich, Y. (2004) More eggs the better: eggformation in captive barn owls (Tyto alba). Auk, 121, 103–109.
Eckert, S.A., Eckert, K.L., Ponganis, P. & Kooyman, G.L. (1989) Diving andforaging behavior of leatherback sea turtles (Dermochelys coriacea). Cana-dian Journal of Zoology, 67, 2834–2840.
Eckert, S.A., Liew, H.-C., Eckert, K.L. & Chan, E.-H. (1996) Shallow waterdiving by leatherback turtles in the South China Sea. Chelonian Conserva-tion and Biology, 2, 237–243.
Fossette, S., Corbel, H., Gaspar, P., Le Maho, Y. & Georges, J.-Y. (2007a) Analternative technique for the long-term satellite tracking of LeatherbackTurtles. Endangered Species Research 3.
Fossette, S., Ferraroli, S., Tanaka, T., Ropert-Coudert, Y., Arai, N., Sato, K.,Naito, Y., Le Maho, Y. & Georges, J.-Y. (2007b) Dispersal and dive patternsin gravid leatherback turtles during the nesting season in French Guiana.Marine Ecology Progress Series, 338, 233–247.
Fossette, S., de Thoisy, B., Kelle, L., Verhage, B., Fretey, J. & Georges, J.-Y. (inpress) The world’s largest leatherback rookeries: conservation and researchin French Guiana, Surinam and Gabon. Journal of Experimental MarineBiology and Ecology, in press.
Froidefond, J.M., Gardel, L., Guiral, D., Parra, M. & Ternon, J.F. (2002)Spectral remote sensing reflectances of coastal waters in French Guianaunder the Amazon influence. Remote Sensing of Environment, 80, 225–232.
Gentry, R.L. & Kooyman, G.L. (1986) Fur Seals: Maternal Strategies on Landand at Sea. Princeton University Press, Princeton.
Georges, J.-Y. & Fossette, S. (2006) Estimating body mass in leatherback turtlesDermochelys coriacea. Marine Ecology Progress Series, 318, 255–262.
Georges, J.-Y., Fossette, S., Billes, A., Ferraroli, S., Fretey, J., Grémillet, D., LeMaho, Y., Myers, A.E., Tanaka, H. & Hays, G.C. (2007) Meta-analysis ofmovements in Atlantic leatherback turtles during the nesting season:conservation implications. Marine Ecology Progress Series, 338, 225–232.
Georges, J.-Y. & Guinet, C. (2000) Maternal care in the subantarctic fur seals onAmsterdam island. Ecology, 81, 295–308.
Girondot, M. & Fretey, J. (1996) Leatherback turtles, Dermochelys coriacea,nesting in French Guiana, 1978–1995. Chelonian Conservation Biology, 2,204–208.
Girondot, M., Godfrey, M.H., Ponge, L. & Rivalan, P. (2007) Modelingapproaches to quantify leatherback nesting trends in French Guiana andSuriname. Chelonian Conservation and Biology, 6, 37–46.
Hays, G.C., Glen, F., Broderick, A.C., Godley, B.J. & Metcalfe, J.D. (2002)Behavioural plasticity in a large marine herbivore: contrasting patterns of
depth utilisation between two green turtle (Chelonia mydas) populations.Marine Biology, 141, 985–990.
Hays, G.C., Isaacs, C., King, R.S., Lloyd, C. & Lovell, P. (2004) First records ofoceanic dive profiles for leatherback turtles, Dermochelys coriacea, indicatebehavioural plasticity associated with long-distance migration. AnimalBehaviour, 67, 733–743.
Hindell, M.A., Slip, D.J. & Burton, H.R. (1991) The diving behaviour of adultmale and female southern elephant seals, Mirounga leonina (Pinnipedia:Phocidae). Australian Journal of Zoology, 39, 595–619.
Hochscheid, S., Godley, B.J., Broderick, A.C. & Wilson, R.P. (1999) Reptiliandiving: highly variable dive patterns in the green turtle Chelonia mydas.Marine Ecology Progress Series, 185, 101–112.
Hochscheid, S., Maffucci, F., Bentivegna, F. & Wilson, R.P. (2004) Gulps,wheezes, and sniffs: how measurement of beak movement in sea turtles canelucidate their behaviour and ecology. Journal of Experimental MarineBiology and Ecology, 316, 45–53.
James, M.C. & Herman, T.B. (2001) Feeding of Dermochelys coriacea onMedusae in the Northwest Atlantic. Chelonian Conservation Biology, 4,202–205.
Jönsson, K.I. (1997) Capital and income breeding as alternative tactics ofresource use in reproduction. Oikos, 78, 57–66.
Kooyman, G.L., Ponganis, P.J., Castellini, M.A., Ponganis, E.P., Ponganis,K.V., Thorson, P.H., Eckert, S.A. & LeMaho, Y. (1992) Heart rates and swimspeeds of emperor penguins diving under sea ice. Journal of ExperimentalBiology, 165, 161–180.
Le Boeuf, B.J., Crocker, D.E., Costa, D.P., Blackwell, S.B., Webb, P.M. &Houser, D.S., (2000) Foraging ecology of northern elephant seals. EcologicalMonographs, 70, 353–382.
Le Boeuf, B.J., Naito, Y., Asaga, T., Crocker, D. & Costa, D.P. (1992) Swimspeed in a female northern elephant seal: metabolic and foraging implications.Canadian Journal of Zoology, 70, 786–795.
Lesage, V., Hammill, M.O. & Kovacs, K.M. (1999) Functional classification ofharbor seal (Phoca vitulina) dives using depth profiles, swimming velocity,and an index of foraging success. Canadian Journal of Zoology, 77, 74–87.
Liebsch, N.S., Wilson, R.P., Bornemann, H., Adelung, D. & Plötz, J. (2007)Mouthing off about fish capture: jaw movements in pinnipeds reveal the realsecrets of ingestion. Deep-Sea Research II, 54, 256–269.
Luschi, P., Benhamou, S., Girard, C., Ciccione, S., Roos, D., Sudre, J. &Benvenuti, S. (2007) Marine turtles use geomagnetic cues during open-seahoming. Current Biology, 17, 126–133.
Lutz, P.L. (1997) Salt, water, and pH balance in sea turtles. The Biology ofSea Turtles (eds P.L. Lutz & J.A. Musick), pp. 343–362. CRC Press, NewYork.
Miller, J.D. (1997) Reproduction in sea turtles. The Biology of Sea Turtles (edsP.L. Lutz & J.A. Musick), pp. 51–73. CRC Press, New York.
Miller, P.J.O., Johnson, M.P., Tyack, P.L. & Terray, E.A. (2004) Swimming gaits,passive drag and buoyancy of diving sperm whales Physeter macrocephalus.Journal of Experimental Biology, 207, 1953–1967.
Mitani, Y., Sato, K., Ito, S., Cameron, M.F., Siniff, D.B. & Naito, Y. (2003) Amethod for reconstructing three-dimensional dive profiles of marinemammals using geomagnetic intensity data: results from two lactatingWeddell seals. Polar Biology, 26, 311–317.
Myers, A.E. & Hays, G.C. (2006) Do leatherback turtles Dermochelys coriaceaforage during the breeding season? A combination of novel and traditionaldata-logging devices provide next insights. Marine Ecology Progress Series,322, 259–267.
Myers, A.E., Lovell, P. & Hays, G.C. (2006) Tools for studying animal behaviour:validation of dive profiles relayed via the Argos satellite system. AnimalBehaviour, 71, 989–993.
Plötz, J., Bornemann, H., Knust, R., Schröder, A. & Bester, M. (2001) Foragingbehaviour of Weddell seals, and its ecological implications. Polar Biology,24, 901–909.
Reina, R.D., Abernathy, K.J., Marshall, G.J. & Spotila, J.R. (2005) Respiratoryfrequency, dive behaviour and social interactions of leatherback turtles,Dermochelys coriacea, during the inter-nesting interval. Journal of ExperimentalMarine Biology and Ecology, 316, 1–16.
Ropert-Coudert, Y., Kato, A., Liebsch, N., Wilson, R.P., Müller, G. & Baubet, E.(2004) Monitoring jaw movements: a cue to feeding activity. Game andWildlife Science, 20, 1–19.
Ropert-Coudert, Y., Liebsch, Y., Kato, A., Bedford, G., Leroy, M. & Wilson, R.P.(2002) Mouth opening in dolphins, as revealed by magnetic sensors. Isana,36, 72–74.
Saba, V.S., Santidrian-Tomillo, P., Reina, R.D., Spotila, J.R., Musick, J.A.,Evans, D.A. & Paladino, F.V. (2007) The effect of the El Ninõ SouthernOscillation on the reproductive frequency of eastern Pacific leatherback.Journal of Applied Ecology, 44, 395–404.

246 S. Fossette et al.
© 2008 The Authors. Journal compilation © 2008 British Ecological Society, Journal of Animal Ecology, 77, 236–246
Schofield, G., Katselidis, K.A., Pantis, J.D., Dimopoulos, P. & Hays, G.C.(2007) Female–female aggression: structure of interaction and outcome inloggerhead sea turtles. Marine Ecology Progress Series, 336, 267–274.
Schreer, J.F., Kovacs, K.M. & O’Hara Hines, R.J. (2001) Comparative divingpatterns of pinnipeds and seabirds. Ecological Monographs, 71, 137–162.
Schreer, J.F. & Testa, J.W. (1996) Classification of Weddell seal diving behaviour.Marine Mammal Science, 12, 227–250.
Scott, E.M., Mann, J., Watson-Capps, J.J., Sargeant, B.L. & Connor, R.C.(2005) Aggression in bottlenose dolphins: evidence for sexual coercion,male-male competition, and female tolerance through analysis of tooth-rakemarks and behaviour. Behaviour, 142, 21–44.
Simeone, A. & Wilson, R.P. (2003) In-depth studies of magellanic penguin(Spheniscus magellanicus) foraging: can we estimate prey consumption byperturbations in the dive profile? Marine Biology, 143, 825–831.
Southwood, A.L., Andrews, D.D., Lutcavage, M.E., Paladino, F., West, N.H.,George, R.H. & Jones, D.R. (1999) Heart rates and diving behavior ofleatherback sea turtles in the eastern Pacific Ocean. Journal of ExperimentalBiology, 202, 1115–1125.
Southwood, A.L., Andrews, R.D., Paladino, F.V. & Jones, D.R. (2005)Effects of diving and swimming behaviour on body temperatures of Pacificleatherback turtles in tropical seas. Physical and Biochemical Zoology, 78,285–297.
Stearns, S.C. (1992) The Evolution of Life Histories. Oxford University Press,Oxford.
Takahashi, A., Dunn, M.J., Trathan, P.N., Croxall, J.P., Wilson, R.P., Sato, K.& Naito, Y. (2004) Krill-feeding behaviour in a chinstrap penguin Pygoscelisantarctica compared with fish-eating in magellanic penguins Speniscusmagellanicus: a pilot study. Marine Ornithology, 32, 47–54.
Wallace, B.P., Cassondra, L.W., Paladino, F.V., Morreale, S.J., Lindstrom, R.T.& Spotila, J.R. (2005) Bioenergetics and diving activity of internestingleatherback turtles Dermochelys coriacea at Parque Nacional Marino LasBaulas, Costa Rica. Journal of Experimental Biology, 208, 3873–3884.
Wallace, B.P., Seminoff, J.A., Kilham, S.S., Spotila, J.R. & Dutton, P.H. (2006)Leatherback turtles as oceanographic indicators: stable isotope analysesreveal a trophic dichotomy between ocean basins. Marine Biology, 149,1432–1793.
Wallace, B.P., Sotherlan, P.R., Tomillo, P.S., Reina, R.D., Spotila, J.R. &Paladino, F.V. (2007) Maternal investment in reproduction and its con-sequences in leatherback turtles. Oecologia, 152, 37–47.
Watanabe, Y., Bornemann, H., Liebsch, N., Plötz, J., Sato, K., Naito, Y. &Miyazaki, N. (2006) Seal-mounted cameras detect invertebrate fauna onunderside of Antarctic ice shelf. Marine Ecology Progress Series, 309, 297–300.
Watwood, S.L., Miller, P.J.O., Johnson, M., Madsen, P.T. & Tyack, P.L. (2006)Deep-diving foraging behaviour of sperm whales (Physeter macrocephalus).Journal of Animal Ecology, 75, 814–825.
Wilson, R.P., Simeone, A., Luna-Jorquera, G., Steinfurth, A., Jackson, S. &Fahlkman, A. (2003) Patterns of respiration in diving penguins: is the lastgasp an inspired tactic? Journal of Experimental Biology, 206, 1751–1763.
Wilson, R.P., Steinfurth, A., Ropert-Coudret, Y., Kato, A. & Kurita, M. (2002)Lip-reading in remote subjects: an attempt to quantify and separateingestion, breathing and vocalisation in free-living animals using penguinsas a model. Marine Biology, 140, 17–27.
Received 23 July 2007; accepted 16 October 2007Handling Editor: Graeme Hayes











