
Ultrastructural evidence of a fungus-sponge association in the Ligurian Sea: a case study of...
8
This article was downloaded by: [University of Genova], [Giorgio Bavestrello] On: 02 October 2014, At: 07:00 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Italian Journal of Zoology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tizo20 Ultrastructural evidence of a fungus-sponge association in the Ligurian Sea: a case study of Clathrina coriacea (Porifera: Calcarea) E. Gaino a , M. Bo b , F. Betti b , M. Bertolino b , F. Scoccia a & G. Bavestrello b a Dipartimento di Chimica, Biologia e Biotecnologie, Università of Perugia, Perugia, Italy b Dipartimento di Scienze della Terra, dell’Ambiente e della Vita, Università di Genova, Genova, Italy Published online: 01 Sep 2014. To cite this article: E. Gaino, M. Bo, F. Betti, M. Bertolino, F. Scoccia & G. Bavestrello (2014): Ultrastructural evidence of a fungus-sponge association in the Ligurian Sea: a case study of Clathrina coriacea (Porifera: Calcarea), Italian Journal of Zoology, DOI: 10.1080/11250003.2014.953219 To link to this article: http://dx.doi.org/10.1080/11250003.2014.953219 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions
Transcript of Ultrastructural evidence of a fungus-sponge association in the Ligurian Sea: a case study of...

This article was downloaded by: [University of Genova], [Giorgio Bavestrello]On: 02 October 2014, At: 07:00Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Italian Journal of ZoologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tizo20
Ultrastructural evidence of a fungus-sponge associationin the Ligurian Sea: a case study of Clathrina coriacea(Porifera: Calcarea)E. Gainoa, M. Bob, F. Bettib, M. Bertolinob, F. Scocciaa & G. Bavestrellob
a Dipartimento di Chimica, Biologia e Biotecnologie, Università of Perugia, Perugia, Italyb Dipartimento di Scienze della Terra, dell’Ambiente e della Vita, Università di Genova,Genova, ItalyPublished online: 01 Sep 2014.
To cite this article: E. Gaino, M. Bo, F. Betti, M. Bertolino, F. Scoccia & G. Bavestrello (2014): Ultrastructural evidence ofa fungus-sponge association in the Ligurian Sea: a case study of Clathrina coriacea (Porifera: Calcarea), Italian Journal ofZoology, DOI: 10.1080/11250003.2014.953219
To link to this article: http://dx.doi.org/10.1080/11250003.2014.953219
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Ultrastructural evidence of a fungus-sponge association in the LigurianSea: a case study of Clathrina coriacea (Porifera: Calcarea)
E. GAINO1, M. BO2, F. BETTI2, M. BERTOLINO2, F. SCOCCIA1, & G. BAVESTRELLO2*
1Dipartimento di Chimica, Biologia e Biotecnologie, Università of Perugia, Perugia, Italy, and 2Dipartimento di Scienze dellaTerra, dell’Ambiente e della Vita, Università di Genova, Genova, Italy
(Received 7 March 2014; accepted 31 July 2014)
AbstractConsecutive underwater observations carried out in shallow waters of Pontetto (Ligurian Sea) from June to the end ofNovember 2013 revealed the presence of purple areas on the white surface of the asconoid calcareous sponge Clathinacoriacea. Histological and ultrastructural investigations performed on the purple areas of the sponge showed the occurrenceof a network of hyphae of an indeterminate fungus permeating the sponge tissues and presumably responsible for theunusual colour of the sponge. The hyphae, varying in size and morphology according to their location in the sponge body,are visible on the outermost sponge surface and, after crossing the mesohyl, penetrate inwards into the choanodermal layer,being mainly located along the apical border of the choanocytes. The occurrence of undamaged flagella and microvillarfringes, which are the organelles characterising the choanocyte cells, is consistent with a normal functionality of the spongetissues, notwithstanding the presence of the fungal hyphae. The fungus develops only in summer because it disappearsconcomitantly with the approach of autumn. This feature is coherent with the consideration that the sponge acts as a suitablesubstrate for the developing fungus, which, in turn, does not interfere with the sponge filter-feeding activity.
Keywords: Porifera, fungal filaments, sponge-hyphae interaction, ultrastructure
Introduction
Porifera, one of the oldest metazoan groups, havecolonised a wide range of freshwater and marinehabitats, becoming fundamental components of thebenthic communities. In different organisationalmodels of sponges, the mesohyl has acquired a grow-ing relevance for the increasing number of theencompassed cell types and for the differentiationof a variety of skeletal elements. Additionally, themesohyl matrix has allowed the establishment ofsymbiotic associations with microorganisms, whichin some species can constitute up to 40% of thecellular component (Vacelet 1975; Hentschel et al.2006).
In fact, one of the most fascinating aspects ofsponge ecology is the transphyletic interactions thatthese animals are able to establish. A remarkablenumber of papers have been devoted to document-ing the relationships between sponges and sym-bionts, namely photosynthetic and heterotrophic
bacteria (Taylor et al. 2007). In addition, due tothe body organisation in which they are permeatedby a complex canal system where a continuous waterflow is induced by their filter-feeding activity,sponges are temporary or permanent substrata alsofor many metazoan species (Magnino & Gaino 1998;Puce et al. 2005; Calcinai et al. 2006a).Symbiosis involving sponges shows a trend from
relationships in which the two partners are indepen-dents, and live together to obtain a better chance forrefuge or food supply, to relationships in which thetwo partners are deeply interconnected to form aunique symbiotic super-organism (Calcinai et al.2006b).The association with marine microorganisms
represents the most outstanding feature because itis commonly known that their concentration withinthe sponge tissue exceeds by up to 2–4 orders ofmagnitude that of the microbes living in seawater(Hentschel et al. 2006). Sponge-associated algae
*Correspondence: G. Bavestrello, Dipartimento di Scienze della Terra, dell’Ambiente e della Vita, Università di Genova, Genova, Italy. Tel:+39 010 3538031.Fax: +39 010 353. Email: [email protected]
Italian Journal of Zoology, 2014, 1–7http://dx.doi.org/10.1080/11250003.2014.953219
© 2014 Unione Zoologica Italiana
Dow
nloa
ded
by [
Uni
vers
ity o
f G
enov
a], [
Gio
rgio
Bav
estr
ello
] at
07:
00 0
2 O
ctob
er 2
014

have also been widely explored: although symbioticzooxanthellae are mainly diffused in the species ofthe family Clionaidae (Granados et al. 2008), theseorganisms have been also sporadically recorded inother families (Scalera-Liaci et al. 1999).
The conveyance of unicellular autotrophic sym-bionts from generation to generation, by the inter-mediate of germinal cells, has been documented inChondrilla australiensis Carter, 1873 (Usher et al.2005), whereas in Tethya orphei Sarà, 1990, filamen-tous cyanobacteria were transferred from the mothertissue to the new individuals during the differentia-tion of asexually produced buds (Gaino et al. 2006).
Fungi associated with marine sponges have beenfrequently observed from temperate, tropical andpolar waters (Gao et al. 2008; Wang et al. 2008;Baker et al. 2009; Lie & Wang 2009; Liu et al.2010; Menezes et al. 2010; Paz et al. 2010; Dinget al. 2011; Wiese et al. 2011; Zhou et al. 2011;Suryanarayanan 2012; Thirunavukkarasu et al.2012; Henriquez et al. 2013). Their diversity wasmainly explored in order to assess the potential forthe production of novel active secondary metaboliteswith biological activity (Höller et al. 2000;Henriquez et al. 2013) and, consequently, the realbiological significance of the relationships remainsunclear. In fact, from the isolation of a fungus froma marine sponge, it is not possible to assume if it isoccasional or actually associated: it may grow therebecause sponges filter from the surrounding environ-ment, which contains spores of terrestrial fungi thatare facultative marine organisms (Höller et al. 2000).To date, only marine ascomycetes of the genusKoralionastes have been reported to have a uniquephysical association with encrusting sponges, devel-oping their ascomata on or within these hosts(Kohlmeyer & Volkmann-Kohlmeyer 1990).
Interesting results in understanding the ecologicalrole of fungi associated with sponges were obtained bymolecular analysis of the two Hawaiian demospongesSuberites aurantiacus (de Laubenfels, 1936) andMycalearmata Thiele, 1903, where 23 and one fungal specieswere respectively found (Gao et al. 2008). This phylo-genetic analysis indicated sponge-derived sequenceswere clustered into “marine fungus clades”.
How marine sponges and filamentous fungi estab-lish ecological relationships remains unknown (Lie &Wang 2009). In order to confirm the real associationbetween fungi and marine sponges, and to determinethe vital functions of the fungi inside the spongetissues, transmission electron microscopy (TEM)and non-cultivation-dependant diagnosis are needed(Passarini et al. 2013).
The aim of this paper was the description, at anultrastructural level (TEM and scanning electron
microscopy, SEM), of the association observed inthe shallow waters of the Ligurian Sea between thecalcarea Clathrina coriacea (Montagu, 1914), charac-terised by a simple, asconoid structure, and an uni-dentified purple fungus.
Materials and methods
Sponge collection
Specimens of the calcareous sponge Clathrina coriaceawere collected by scuba diving in Pontetto (EasternLigurian Sea) (44°22’ 34.23 N; 9°04’ 24.10 E), at adepth varying from 1 to 5 m. Diving was performedfrom June to the end of November 2013. Onceexcided, sponge specimens were fixed underwater in2.5% glutaraldehyde buffered with filtered artificialseawater (ASW) (pH adjusted to 7.5–7.8, with0.1 N sodium hydroxide, NaOH) for 1 h, and trans-ferred directly to sterilised locked bags containingASW to prevent contact of sponge tissue with air.The samples were transported to the laboratory andimmediately processed for histological and ultrastruc-tural analyses. To obtain an estimation of the numberof sponges hosting the purple symbiont, all the speci-mens of C. coriacea showing or not the purple col-ouration were counted during three sessions of 5’ ofslow swimming (about 100 m long each).
Histological and ultrastructural analyses
For histological investigations, several fixed frag-ments of the purple areas (about 1 cm2 each) wererepeatedly washed in ASW and processed accordingto the routinely used procedures; sections of 5–7 μmin thickness were then observed with a Leica micro-scope (Leica LMS Holdings GmH, Wetzlar,Germany).For TEM, small fragments of the purple areas
(about 2 mm2 each) were repeatedly washed inASW and post-fixed in 1% osmium tetroxide, inASW used as a buffer, for 1 h at 4°C; after repeatedwashes, the material was dehydrated in a series ofethanol dilutions and embedded in an Epon-Aralditemixture. Ultrathin sections were cut on a Leica EMUC6 ultracut (Leica Microsystem GmbH, Vienna,Austria), collected on formvar-coated copper grids,stained with uranyl acetate and lead citrate andexamined with a Philips EM 208 (Philips).Some fragments of the fixed material were pro-
cessed for SEM. In this regard, they were repeatedlywashed in ASW, dehydrated in a series of ethanoldilutions, critical point-dried, mounted on stubswith silver conducting paint, sputter-coated withgold-palladium in an Emitech K 550X (Emitech,
2 E. Gaino et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f G
enov
a], [
Gio
rgio
Bav
estr
ello
] at
07:
00 0
2 O
ctob
er 2
014

Ashford, UK) sputter and observed with a PhilipsXL30 (Philips, Eindhoven, the Netherlands) micro-scope at an accelerating voltage of 18 kV.
Results
Clathrina coriacea is a white sponge living in theupper fringe of the rocky infra-littoral community.This sponge had a size ranging from 2 to 10 cm andwas characterised by a net of contorted and anasto-mosed tubes of 0.5–1 mm in diameter, ending insharp conules bearing apical oscules (Figure 1a).About 10% of the observed sponges showed patchedpurple areas, particularly outstanding on the whitetissue of the sponge (Figure 1b). In some specimens,the purple areas widely invaded the sponge tissuesoccupying as much as 80% of the volume (Figure1b–c). In vivo, the sponge showed no sign of degen-eration on its purple areas.
Histological tangential sections of the sponge tubesshowed fungal hyphae permeating the mesohyl region(Figure 1d). In cross sections, the fungal hyphae wereeasily detectable as translucent elements overlappingthe sponge choanocytes (Figure 2a). The shape of thehyphal filaments varied according to the location of
these elements: from elongated in shape, whenlocated inside the atrial cavity (Figure 2b) or whencrossing the thickness of the mesohyl (Figure 2c), tofairly roundish, when closer to the choanodermallayer (Figure 2a).SEM images confirmed the occurrence of the
hyphae along the choanodermal layer facing theatrial cavity. TEM analysis showed the irregularshape of the hyphae, the diameter of which variedalong their length (Figure 2e). In some fracturedsponge tubes, observed under SEM, it becameevident that the hyphae were so large as toovergrow and partially envelop entire portions ofthe choanodermal layer (Figure 2f). This featurewas also confirmed by TEM, which showed thehyphae interposed between adjacent choanocytes(Figure 2g).In addition, TEM investigations allowed the loca-
tion of the fungal hyphae in the sponge tissue to bedetailed. In the choanodermal region, the hyphaewere almost adherent to the choanocytes that bor-dered the atrial cavity, where collar fringes encirclingthe central flagellum (Figure 3a), a characteristic ofthe choanocytes, were also evident. The hyphae pre-sented a subtle electron-dense border delimiting a
Figure 1. The white marine sponge Clathrina coriacea (a–c) in its natural environment, and (d) histological section of a body region.(a) typical anastomosed tubes ending in sharp conules bearing apical oscules (arrows); (b) patched purple areas on the sponge surface;(c) purple areas covering almost the entire sponge surface; (d) tangential histological section of a purple area of the sponge showing thenetwork of fungal hyphae (arrows).
Ultrastructure of a fungus-sponge association 3
Dow
nloa
ded
by [
Uni
vers
ity o
f G
enov
a], [
Gio
rgio
Bav
estr
ello
] at
07:
00 0
2 O
ctob
er 2
014

vacuolated central region irregularly crossed by thinsepta (Figure 3b). The hyphae permeated the choa-nodermal layer in such a way that their apical regioninfiltrated among choanocytes and the rest of the
filament protruded into the sponge atrial cavity(Figure 3c). Consequently, the continuity of thechoanodermal layer was frequently interrupted bythe presence of the hyphae reaching this tissue after
Figure 2. (a–c) histological sections, (d, f) scanning and (e, g) transmission electron microscopy of the purple areas of the sponge Clathrinacoriacea. (a) cross section of a tube showing the position of the fungal hyphae (arrows) bordering the choanodermal layer (cl); m, mesohyl;pl, pinacodermal layer; (b) a fairly tangential section showing a hyphal filament in the sponge cavity (arrow); cl, choanodermal layer; (c) ahyphal filament (arrow) crossing the mesohyl (m), cl, choanodermal layer; (d) a hyphal filament on the choandodermal layer (cl); f, flagellaof the choncocytes; (e) a hyphal filament showing its irregular pattern; (f) a fractured sponge tube showing a hyphal filament (hf)overgrowing a portion of the choanodermal layer (cl); (g) irregular shape of hyphal filaments (hf) interposed between two choanocytes (ch).
4 E. Gaino et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f G
enov
a], [
Gio
rgio
Bav
estr
ello
] at
07:
00 0
2 O
ctob
er 2
014
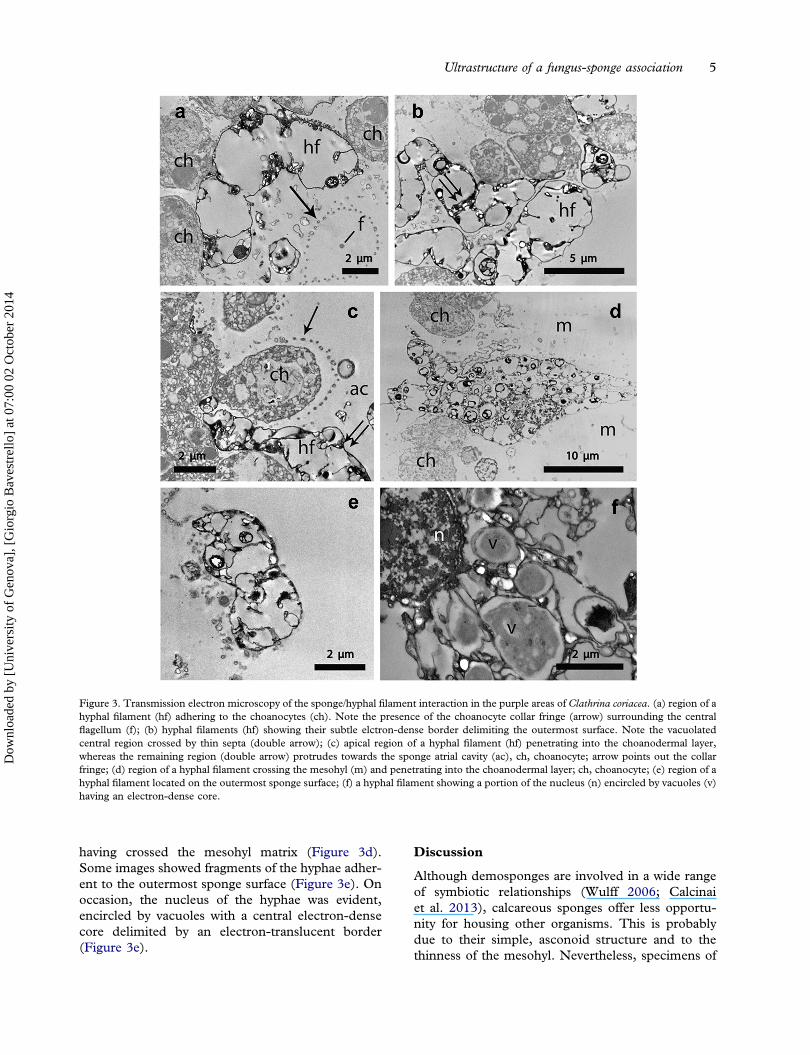
having crossed the mesohyl matrix (Figure 3d).Some images showed fragments of the hyphae adher-ent to the outermost sponge surface (Figure 3e). Onoccasion, the nucleus of the hyphae was evident,encircled by vacuoles with a central electron-densecore delimited by an electron-translucent border(Figure 3e).
Discussion
Although demosponges are involved in a wide rangeof symbiotic relationships (Wulff 2006; Calcinaiet al. 2013), calcareous sponges offer less opportu-nity for housing other organisms. This is probablydue to their simple, asconoid structure and to thethinness of the mesohyl. Nevertheless, specimens of
Figure 3. Transmission electron microscopy of the sponge/hyphal filament interaction in the purple areas of Clathrina coriacea. (a) region of ahyphal filament (hf) adhering to the choanocytes (ch). Note the presence of the choanocyte collar fringe (arrow) surrounding the centralflagellum (f); (b) hyphal filaments (hf) showing their subtle elctron-dense border delimiting the outermost surface. Note the vacuolatedcentral region crossed by thin septa (double arrow); (c) apical region of a hyphal filament (hf) penetrating into the choanodermal layer,whereas the remaining region (double arrow) protrudes towards the sponge atrial cavity (ac), ch, choanocyte; arrow points out the collarfringe; (d) region of a hyphal filament crossing the mesohyl (m) and penetrating into the choanodermal layer; ch, choanocyte; (e) region of ahyphal filament located on the outermost sponge surface; (f) a hyphal filament showing a portion of the nucleus (n) encircled by vacuoles (v)having an electron-dense core.
Ultrastructure of a fungus-sponge association 5
Dow
nloa
ded
by [
Uni
vers
ity o
f G
enov
a], [
Gio
rgio
Bav
estr
ello
] at
07:
00 0
2 O
ctob
er 2
014

Clathrina luteoculcitella Wörheide & Hooper, 1999from South China showed a great culturable fungaldiversity represented by 12 genera among which thespecies Nigrospora oryzae (Berk & Br.) strain PF18,having a strong and broad spectrum of antimicrobialactivities, could have potential for antimicrobialcompound production (Ding et al. 2011).
At present, among the species of the genusClathrina living the Liguria Sea, namely C. cerebrum(Haeckel, 1872), C. clathrus (Schmidt, 1864), C. con-torta (Bowerbank, 1866) and C. rubra Sarà, 1958,C. coriacea represents the most diffused sponge(Longo & Pronzato 2011). In fact, C. clathrus,C. contorta and C. rubra are becoming rare represen-tatives of the sponge community, and C. cerebrum,which in the past survived the summer heat crisis inminute forms (Gaino et al. 1996), has recently beenstrongly reduced (unpublished data).
In C. cerebrum, elongated bacteria, tentativelyattributed to the genus Cytophaga, were recordedadherent to the choanodermal region and, morerarely, penetrating into the mesohyl (Burlandoet al. 1988). These authors suggested this associationwas probably based on some species-specificmechanisms because such a relationship was lackingin other congeneric species.
In Sycon sp., characterised by a syconoid struc-ture, two species of fungi (Emericellopsis minimaStolk and Scopulariopsis candida Vuill.) were isolated(Höller et al. 2000), but no information about thereal nature of the association between the fungalstrains and the sponge was provided.
This paper gives the first ultrastructural evidenceof the relationship between a fungus and a marinesponge, allowing us to demonstrate the location ofthe fungal hyphae within both the mesohyl matrixand the choanodermal layer.
The occurrence of the fungal infection does notapparently affect the sponge organisation, as evi-denced at the ultrastructural level by the integrity ofboth flagella and the collar fringes of the choano-cytes. This evidence strongly suggests that the choa-nocyte functionality also remains unchanged.
The presence of the fungus does not constitute astable relationship with the sponge tissues becauserepeated underwater observations, carried out for 6months between summer and autumn, allowed us toverify the disappearance of the pigmented colonies ofC. coriacea after the middle of October. This featureproves that the fungus develops inside the spongebody only during the summer season. The thin andtypically not pigmented sponge tubes facilitated arapid detection of the presence/absence of the fungalhyphae, without the need to apply the complex tech-niques usually utilised for isolating the fungal
communities (Höller et al. 2000; Gao et al. 2008;Henriquez et al. 2013; Passarini et al. 2013).Additional surveys, carried out in early and late win-ter, confirmed the presence of the population ofC. coriacea characterized by the typical white colour.This feature proves that the sponge does not have aseasonality development, being present all yearround in the sampling site.The sporadic presence of the fungus did not allow
its characterisation to be performed. As the sponge/fungus association does not interfere with the survi-val of the sponge population, we can speculate thatthe fungus exploited the sponge body as a suitablesubstrate for its further development, without affect-ing sponge functionality. Nevertheless, the total lackof records concerning the presence of the fungus onthe surrounding substratum or associated with otherbenthic organisms induces us to suppose a certaindegree of specificity in this association.
References
Baker PW, Kennedy J, Dobson ADW, Marchesi JR. 2009.Phylogenetic diversity and antimicrobial activities of fungiassociated with Haliclona simulans isolated from Irish coastalwaters. Marine Biotechnology 11:540–547. doi:10.1007/s10126-008-9169-7.
Burlando B, Sabatini MA, Gaino E. 1988. Association betweencalcareous Clathrina cerebrum (Haeckel) and bacteria: Electronmicroscope study. Journal of Experimental Marine Biologyand Ecology 116:35–42. doi:10.1016/0022-0981(88)90244-4.
Calcinai B, Bavestrello G, Bertolino M, Pica D, Wagner D,Cerrano C. 2013. Sponges associated with octocorals in theIndo-Pacific, with the description of four new species. Zootaxa3617:1–61. doi:10.11646/zootaxa.3617.1.1.
Calcinai B, Cerrano C, Bavestrello G. 2006a. Sponges as hotspotof biodiversity. Biologia Marina Mediterranea 12:63–68.
Calcinai B, Cerrano C, Totti C, Romagnoli T, Bavestrello G.2006b. Symbiosis of Mycale (Mycale) vansoesti sp. nov.(Porifera, Demospongiae) with a coralline alga from NorthSulawesi (Indonesia). Invertebrate Biology 125:195–204.doi:10.1111/j.1744-7410.2006.00052.x.
Ding B, Yin Y, Zhang F, Li Z. 2011. Recovery and phylogeneticdiversity of culturable fungi associated with marine spongesClathrina luteoculcitella and Holoxea sp. in the South ChinaSea. Marine Biotechnology 13:713–721. doi:10.1007/s10126-010-9333-8.
Gaino E, Bavestrello G, Cerrano C, Sarà M. 1996. Survival of thecalcareous sponge Clathrina cerebrum (Haeckel, 1872) on avertical cliff during the summer crisis. Italian Journal ofZoology 63:41–46. doi:10.1080/11250009609356105.
Gaino E, Sciscioli M, Lepore E, Rebora M, Corriero G. 2006.Association of the sponge Tethya orphei (Porifera,Demospongiae) with filamentous cyanobacteria. InvertebrateBiology 125:281–287. doi:10.1111/j.1744-7410.2006.00061.x.
Gao Z, Li B, Zheng C, Wang G. 2008. Molecular detection offungal communities in the Hawaiian marine sponges Suberiteszeteki and Mycale armata. Applied and EnvironmentalMicrobiology 74:6091–6101. doi:10.1128/AEM.01315-08.
Granados C, Camargo C, Zea S, Sánchez JA. 2008. Phylogeneticrelationships among zooxanthellae (Symbiodinium) associated
6 E. Gaino et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f G
enov
a], [
Gio
rgio
Bav
estr
ello
] at
07:
00 0
2 O
ctob
er 2
014

to excavating sponges (Cliona spp.) reveal an unexpected line-age in the Caribbean. Molecular Phylogenetics and Evolution49:554–560. doi:10.1016/j.ympev.2008.07.023.
Henriquez M, Vergara K, Norambuena J, Beiza A, Maza F,Ubilla P, Araya I, Chàvez R, San-Martin A, Darias J,Darias MJ, Yaca I. 2013. Diversity of cultivable fungi asso-ciated with Antarctic marine sponges and screening for theantimicrobial, antitumoral and antioxidant potential. WorldJournal Microbiology Biotechnology doi:10.1007/s11274-013-1418–x.
Hentschel U, Usher KM, Taylor MV. 2006. Marine sponges asmicrobial fermenters. FEMS Microbiology and Ecology55:167–177. doi:10.1111/j.1574-6941.2005.00046.x.
Höller U, Wright AD, Matthee GF, Konig GM, Draeger S, AustH-J, Schulz B. 2000. Fungi from marine sponges: Diversity,biological activity and secondary metabolites. MycologicalResearch 104:1354–1365. doi:10.1017/S0953756200003117.
Kohlmeyer J, Volkmann-Kohlmeyer B. 1990. New species ofKoralionastes (Ascomycotina) from the Caribbean andAustralia. Canadian Journal of Botany 68:1554–1559.doi:10.1139/b90-199.
Lie Q, Wang G. 2009. Diversity of fungal isolates from threeHawaiian marine sponges. Microbiological Research164:233–241. doi:10.1016/j.micres.2007.07.002.
Liu WC, Li CQ, Zhu P, Yang JL, Cheng KD. 2010. Phylogeneticdiversity of culturable fungi associated with two marine sponges:Haliclona simulans and Gelliodes carnosa, collected from theHainan Island coastal waters of the South China Sea. FungalDiversity 42:1–15. doi:10.1007/s13225-010-0022-8.
Longo C, Pronzato R. 2011. Class Calcarea. In: Pansini M,Manconi R, Pronzato R, editors. Fauna d’Italia, Porifera I.Milano: Calderini. pp. 117–244.
Magnino G, Gaino E. 1998. Haplosyllis spongicola GrubePolychaeta, Syllidae associated with two species of spongesfrom East Africa Tanzania, Indian Ocean. Marine Ecology19:77–87. doi:10.1111/j.1439-0485.1998.tb00455.x.
Menezes CBA, Bonugli-Santos RC, Miqueletto PB, PassariniMRZ, Silva CHD, Justo MR, Leal RR, Fantinatti-Garboggini F, Oliveira VM, Berlinck RGS, Sette LD. 2010.Microbial diversity associated with algae, ascidians andsponges from the north coast of São Paulo state, Brazil.Microbiological Research 165:466–482. doi:10.1016/j.micres.2009.09.005.
Passarini MRZ, Santos C, Lima N, Berlinck RGS, Sette LD.2013. Filamentous fungi from the Atlantic marine spongeDragmacidon reticulatum. Archives of Microbiology 195:99–111. doi:10.1007/s00203-012-0854-6.
Paz Z, Komon-Zelazowska M, Druzhinina IS, Aveskamp MM,Shnaiderman A, Aluma Y, Carmeli S, Ilan M, Yarden, O.2010. Diversity and potential antifungal properties of fungiassociated with a Mediterranean sponge. Fungal Diversity42:17–26. doi:10.1007/s13225-010-0020-x.
Puce S, Calcinai B, Bavestrello G, Cerrano C, Gravili C, Boero F.2005. Hydrozoa (Cnidaria) symbiotic with Porifera: A review.Marine Ecology 26:73–81.
Scalera-Liaci L, Sciscioli M, Lepore E, Gaino E. 1999. Symbioticzooxanthellae in Cynachira tarentina a non-boring demos-ponge. Endocytobiosis & Cell Research 13:105–114.
Suryanarayanan TS. 2012. The diversity and importance of fungiassociated with marine sponges. Botanica Marina 55:553–564.doi:10.1515/bot-2011-0086.
Taylor MW, Radax R, Steger D, Wagner M. 2007. Sponge-asso-ciated microorganisms: Evolution, ecology, and biotechnologi-cal potential. Microbiology and Molecular Biology Reviews71:295–347. doi:10.1128/MMBR.00040-06.
Thirunavukkarasu N, Suryanarayanan TS, Girivasan KP,Venkatachalam A, Geetha V, Ravishankar JP, Doble M.2012. Fungal symbionts of marine sponges fromRameswaram, southern India: Species composition and bioac-tive metabolites. Fungal Diversity 55:37–46. doi:10.1007/s13225-011-0137-6.
Usher KM, Sutton DC, Toze S, Kuo J, Fromont J. 2005. Inter-generational transmission of microbial symbionts in the marinesponge Chondrilla australiensis (Demospongiae). Marine andFreshwater Research 56:125–131. doi:10.1071/MF04304.
Vacelet J. 1975. Étude en Microscopie Électronique del’Association entre Bactéries et Spongiaires du GenreVerongia (Dictyoceratida). Journal de Microscopie et deBiologie Cellulaire 23:271–288.
Wang G, Li Q, Zhu P. 2008. Phylogenetic diversity of culturablefungi associated with the Hawaiian sponges Suberites zeteki andGelliodes fibrosa. Antonie van Leeuwenhoek 93:163–174.doi:10.1007/s10482-007-9190-2.
Wiese J, Ohlendorf B, Blümel M, Schmaljohann R, Imhoff JF. 2011.Phylogenetic identification of fungi isolated from the marinesponge Tethya aurantium and identification of their secondarymetabolites. Marine Drugs 9:561–585. doi:10.3390/md9040561.
Wulff JL. 2006. Ecological interactions of marine sponges.Canadian Journal of Zoology 84:146–166. doi:10.1139/z06-019.
Zhou K, Zhang X, Zhang F, Li Z. 2011. Phylogenetically diversecultivable fungal community and polyketide synthase (PKS),non-ribosomal peptide synthase (NRPS) genes associated withthe South China Sea sponges. Microbiology Ecology 62:644–654. doi:10.1007/s00248-011-9859-y.
Ultrastructure of a fungus-sponge association 7
Dow
nloa
ded
by [
Uni
vers
ity o
f G
enov
a], [
Gio
rgio
Bav
estr
ello
] at
07:
00 0
2 O
ctob
er 2
014




















