
Pectoral fins of rhizodontids and the evolution of pectoral appendages in the tetrapod stem‐group
Upload
independentCategory
view
0download
0
Journal of Vertebrate Paleontology 15(3):516-531, September 1995© 1995 by the Society of Vertebrate Paleontology
THE PECTORAL GIRDLE AND FORELIMB OF CARSOSAURUS MARCHESETTI(AIGIALOSAURIDAE), WITH A PRELIMINARY PHYLOGENETIC ANALYSIS OF
MOSASAUROIDS AND VARANOIDS
MICHAEL W. CALDWELL*, ROBERT L. CARROLL, AND HINRICH KAISERtRedpath Museum and Department of Biology, McGill University,859 Sherbrooke Street West, Montreal, Quebec, Canada, H3A 2K6
ABSTRACT- The aigialosaur Carsosaurus marchesetti is represented by a nearly complete skeletonfrom the Upper Cretaceous (Cenomanian-Turonian) of Slovenia. Dermal girdle elements include portions of the clavicles and a small interclavicle with a short anterior process. Endochondral girdleelements include a small scapula and large unfenestrated coracoid. A mineralized sternum is alsopresent. The carpus is anguid-like and consists of ten ossified elements. Reduction of the procoelousnature of centrum articular surfaces is restricted to the caudal series. The type of C. marchesetti is thelargest aigialosaur specimen known (> 1.5 m) and has proportionally larger propodials than any otheraigialosaur. Phylogenetic reconstruction places Carsosaurus within a polytomous clade composed ofall known aigialosaurs. Aigialosaurs are the sister-group to Mosasauridae, forming the Mosasauroidea,and are within Anguimorpha but distinct from Varanoidea. Supposed synapomorphies of varanidsand mosasauroids are identified as either plesiomorphies ofAnguimorpha, or as misidentified homologs.Results of a principal component analysis of limb measurements suggest that aigialosaurs occupy aposition on a functional continuum reflective of terrestrial not aquatic animals. Morphometric criteriacannot validate the exclusion of Carsosaurus from the Aigialosauridae.
INTRODUCTION
The aigialosaur Carsosaurus marchesetti was described by Komhuber (1893) on the basis of a singlespecimen collected from a cave in the Trieste Limestone (Cenomanian-Turonian, U pper Cretaceous) nearKomen, Slovenia (Calligaris, 1988). The more complete specimens assigned to the Aigialosauridae (Table1) also come from the Trieste Limestone, though fromdifferent localities than C. marchesetti (Calligaris, 1988;Carroll and deBraga, 1992). The only specimen of C.marchesetti is exposed in ventral view and is missingthe skull, anterior cervicals, and a large part of the tail(Fig. 1). The remaining postcranial skeleton is nearlycomplete.
The Aigialosauridae, exclusive of C. marchesetti,contains two monotypic genera (Table 1), each represented by a single specimen, and a single unnamedspecimen referred to here as the Trieste Aigialosaur("Trieste specimen" of Carroll and deBraga, 1992). Aseries of vertebrae, referred to Aigialosaurus novaki byKramberger (1892), show no features that allow differentiation from other taxa. A skull fragment fromthe Upper Jurassic, referred to Proaigialosaurus hueniby Kuhn (1958), is now lost (Table 1).
* Current address: Department ofBiological Sciences, University of Alberta, Edmonton, Alberta T69 2E9, Canada.
t Current address: Institut fur Humangenetik, UniversitatWiirzburg, Biozentrum am Hubland, 97074 Wiirzburg, Germany.
516
Komhuber (1893) classified C. marchesetti as amember of Aigialosauridae on the basis of postcranialfeatures. However, Carroll and deBraga (1992) excluded C. marchesetti from their review of the Aigialosauridae because of limb bone ratios which theyconsidered indicative ofsignificant differences betweenC. marchesetti and the other aigialosaurs. Carroll anddeBraga (1992) also suggested that A. dalmaticus andOpetiosaurus bucchichi might be conspecific, therebymaking the Aigialosauridae monotypic. However, theirassertion that C. marchesetti differs from other aigialosaurs only means that this species might not belongto the genus or species of the conspecific forms; it doesnot merit its exclusion from the family. It is thereforeimportant to determine the conspecificity of A. dalmaticus and O. bucchichi and whether or not thesetaxa form a monophyletic group with C. marchesetti.
The postcranium of C. marchesetti is an importantsource of characters as it includes details of the shoulder girdle not preserved in other aigialosaurs. Conventional approaches to varanoid phylogeny consistently hypothesize that an aigialosaur-mosasaur cladeis nested within the Varanoidea (McDowell and Bogert, 1954; Russell, 1967; Rieppel, 1980a; Borsuk-Bialynicka, 1984; Carroll and deBraga, 1992; deBragaand Carroll, 1993). However, it is difficult to accountfor the large number of reversals which result whenpolarity is determined using fossil or extant varanoids(sensu Pregill et aI., 1986) as outgroups.
Gauthier (1982) and Pregill et aI. (1986) suggestedthat an aigialosaur-mosasaur clade might in fact lie

CALDWELL ET AL.-CARSOSAURUS PECTORAL GIRDLE 517
TABLE 1. List of taxa assigned to the family Aigialosauridae (Squamata: Mosasauroidea), with a short characterization ofeach specimen.
Species assignment
Aigialosaurus dalmaticusA. novakiCarsosaurus marchestettiOpetiosaurus bucchichiProaigialosaurus hueniTrieste Aigialosaur
Location
MiinchenTriesteTriesteViennaLostTrieste
Status
Mostly complete38 caudal vertebraeComplete except for skullMostly completeSkull fragmentComplete, associated, described
but not named
Reference
Kramberger, 1892Kramberger, 1892Kronhuber, 1893Komhuber, 1901Kuhn, 1958Carroll and deBraga, 1992
outside of Varanoidea. This is an important and interesting suggestion which has had variable support,for very different reasons, from late nineteenth andearly twentieth century squamate systematists (Fejervary, 1918 for a review). Hypotheses of a sistergrouprelationship between aigialosaurs and mosasaurs appear well supported, making aigialosaur morphologyimportant to polarizing mosasaur characters. In a general sense, the skull is almost indistinguishable fromthat of primitive mosausaurs (Carroll and deBraga,1992). However, as will be discussed below, charactersin the postcranial skeleton show only minor differencesfrom those of terrestrial anguimorphs.
We present a detailed description of the repreparedpectoral girdle and forelimb of C. marchesetti alongwith a review of the remaining postcranial skeleton.From this description we have added characters tothose used by Pregill et al. (1986) and Rieppel (1980a)in order to construct a character matrix for phylogenetic analysis of aigialosaurs, mosasaurs, and varanoids. We also present a preliminary phylogenetic hypothesis of mosasauroid-varanoid relationships usingthese characters. This analysis was motivated by theuncertain phylogenetic relationship of C. marchesettiwith other aigialosaurs (Carroll and deBraga, 1992).
To investigate further the morphometrically-basedassertion of Carroll and deBraga (1992) that C. marchesetti should not be considered an aigialosaur, wehave added measurements of limb and vertebral elements for Varanus salvator, Lanthanotus borneensis,Heloderma sp., and Clidastes propython (Table 2) totheir data set, and conducted a principle componentanalysis (PCA). In association with qualitative morphological descriptions, our PCA results are interpreted in terms of function not phylogeny.
MATERIALS AND METHODS
The shoulder girdle and forelimb of Carsosaurusmarchesetti were reprepared and drawn by RLC duringthe summer of 1991. Due to difficulties encounteredin differentiating bone from matrix it was not possibleto prepare the remainder of the skeleton. Reconstruction of C. marchesetti is based on drawings and photographs of the entire specimen. Institutional abbreviations are FMNH (Field Museum of Natural His-
tory, Chicago, Illinois, U.S.A.), RM (Redpath Museum, McGill University, Montreal, Quebec, Canada),KU (Museum of Natural History, University ofKansas, Lawrence, U.S.A.), BSP (Bayerische Staatssammlung fur Palaontologie und historische Geologie,Miinchen, Germany), MCSNT (Museo Civico di Storia Naturale, Trieste, Italy), and NMW (Naturhistorisches Museum, Wien, Austria).
Phylogenetic Analysis
Phylogenetic analysis ofnine mosasauroid-varanoidtaxa was conducted using PAUP Version 3.1.1 for theMacintosh (Swofford, 1993). A data matrix of sixtysix characters (sixty-one binary and five multistate)was constructed (Appendix I), using characters andcharacter states from Pregill et al. (1986) and Rieppel(1980a), and from characters identified in this study(Appendix II). Some characters were modified to reflectingroup character states. All multistate character transformations were unordered to avoid a priori assumptions of transformation vectors.
Outgroup polarity was determined using charactersof Gerrhonotus. Information on gerrhonotines was derived from an unnumbered skull of Gerrhonotus (=EIgaria) multicarinatus in the Redpath Museum collection, and from Meszoely (1970), Rieppel (1980a), Gauthier (1982), and Good (1987). Examination of hypotheses of anguimorph phylogeny indicate thatgerrhonotines are among the least derived anguoids(Rieppel, 1980a) or anguids (Good, 1988). Gauthier(1982) and Estes et al. (1988) reported a basal anguimorphan trichotomy between varanoids, xenosaurids,and anguids. Estes et al. (1988:fig. 5C) also present acladogram with snakes, dibamids, and amphisbaenidsdeleted, in which Anguidae is the closest sistergroupof living varanoids. A consensus of these phylogenetichypotheses would seem to indicate that an anguid taxon is a reasonable choice for outgroup polarization ofvaranoid characters.
Morphometric Analysis
Measurements of upper and lower limb charactersand average lengths of trunk vertebrae (Table 2) weretaken from Varanus salvator (RM 1969.30a), Lanthanotus borneensis (FMNH 134711), Heloderma sp.
518 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 15, NO.3, 1995
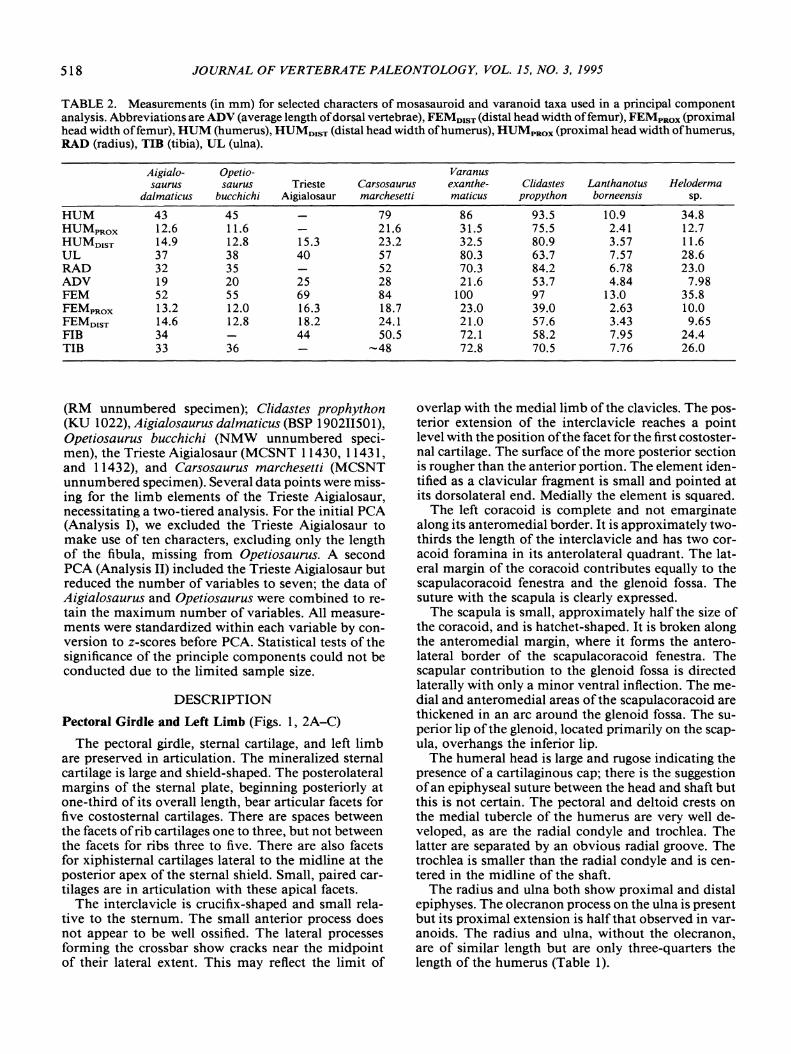
TABLE 2. Measurements (in mm) for selected characters of mosasauroid and varanoid taxa used in a principal componentanalysis. Abbreviations are ADV (average length ofdorsal vertebrae), FEMD1ST (distal head width offemur), FEMpROX (proximalhead width offemur), HUM (humerus), HUMDIsT (distal head width ofhumerus), HUMpRox (proximal head width ofhumerus,RAD (radius), TID (tibia), UL (ulna).
Aigialo- Opetio- Varanussaurus saurus Trieste Carsosaurus exanthe- Clidastes Lanthanotus Heloderma
dalmatieus bueehiehi Aigialosaur marehesetti matieus propython borneensis sp.
HUM 43 45 79 86 93.5 10.9 34.8HUMpRox 12.6 11.6 21.6 31.5 75.5 2.41 12.7HUMo1sT 14.9 12.8 15.3 23.2 32.5 80.9 3.57 11.6UL 37 38 40 57 80.3 63.7 7.57 28.6RAD 32 35 52 70.3 84.2 6.78 23.0ADV 19 20 25 28 21.6 53.7 4.84 7.98FEM 52 55 69 84 100 97 13.0 35.8FEMpRox 13.2 12.0 16.3 18.7 23.0 39.0 2.63 10.0FEM01ST 14.6 12.8 18.2 24.1 21.0 57.6 3.43 9.65FIB 34 44 50.5 72.1 58.2 7.95 24.4TIB 33 36 -48 72.8 70.5 7.76 26.0
(RM unnumbered specimen); Clidastes prophython(KU 1022), Aigialosaurus dalmaticus (BSP 190211501),Opetiosaurus bucchichi (NMW unnumbered specimen), the Trieste Aigialosaur (MCSNT 11430, 11431,and 11432), and Carsosaurus marchesetti (MCSNTunnumbered specimen). Several data points were missing for the limb elements of the Trieste Aigialosaur,necessitating a two-tiered analysis. For the initial PCA(Analysis I), we excluded the Trieste Aigialosaur tomake use of ten characters, excluding only the lengthof the fibula, missing from Opetiosaurus. A secondpeA (Analysis II) included the Trieste Aigialosaur butreduced the number of variables to seven; the data ofAigialosaurus and Opetiosaurus were combined to retain the maximum number of variables. All measurements were standardized within each variable by conversion to z-scores before PCA. Statistical tests of thesignificance of the principle components could not beconducted due to the limited sample size.
DESCRIPTION
Pectoral Girdle and Left Limb (Figs. 1, 2A-C)
The pectoral girdle, sternal cartilage, and left limbare preserved in articulation. The mineralized sternalcartilage is large and shield-shaped. The posterolateralmargins of the sternal plate, beginning posteriorly atone-third of its overall length, bear articular facets forfive costosternal cartilages. There are spaces betweenthe facets ofrib cartilages one to three, but not betweenthe facets for ribs three to five. There are also facetsfor xiphisternal cartilages lateral to the midline at theposterior apex of the sternal shield. Small, paired cartilages are in articulation with these apical facets.
The interclavicle is crucifix-shaped and small relative to the sternum. The small anterior process doesnot appear to be well ossified. The lateral processesforming the crossbar show cracks near the midpointof their lateral extent. This may reflect the limit of
overlap with the medial limb of the clavicles. The posterior extension of the interclavicle reaches a pointlevel with the position ofthe facet for the first costosternal cartilage. The surface of the more posterior sectionis rougher than the anterior portion. The element identified as a clavicular fragment is small and pointed atits dorsolateral end. Medially the element is squared.
The left coracoid is complete and not emarginatealong its anteromedial border. It is approximately twothirds the length of the interclavicle and has two coracoid foramina in its anterolateral quadrant. The lateral margin of the coracoid contributes equally to thescapulacoracoid fenestra and the glenoid fossa. Thesuture with the scapula is clearly expressed.
The scapula is small, approximately half the size ofthe coracoid, and is hatchet-shaped. It is broken alongthe anteromedial margin, where it forms the anterolateral border of the scapu1acoracoid fenestra. Thescapular contribution to the glenoid fossa is directedlaterally with only a minor ventral inflection. The medial and anteromedial areas of the scapulacoracoid arethickened in an arc around the glenoid fossa. The superior lip of the glenoid, located primarily on the scapula, overhangs the inferior lip.
The humeral head is large and rugose indicating thepresence of a cartilaginous cap; there is the suggestionofan epiphyseal suture between the head and shaft butthis is not certain. The pectoral and deltoid crests onthe medial tubercle of the humerus are very well developed, as are the radial condyle and trochlea. Thelatter are separated by an obvious radial groove. Thetrochlea is smaller than the radial condyle and is centered in the midline of the shaft.
The radius and ulna both show proximal and distalepiphyses. The olecranon process on the ulna is presentbut its proximal extension is half that observed in varanoids. The radius and ulna, without the olecranon,are of similar length but are only three-quarters thelength of the humerus (Table 1).

CALDWELL ET AL.-CARSOSAURUS PECTORAL GIRDLE 519
The carpus contains ten ossified elements. These areidentified according to the conventions ofRenous-Ucuru (1973) as the radiale, lateral centrale, intermedium, ulnare, pisiform, medial centrale, and distal carpals two through five. The radiale is an irregularlyshaped element, compressed proximodistally, and expanded into small lobes on its medial and lateral margins. The medial expansion of the element contributesto the distal margin of the antebrachial space. Thelateral centrale is an irregular hexagon and articulatesproximally with the intermedium, medially with theradiale and medial centrale, laterally with the ulnare,and distally with distal carpals two to four. The ulnareis the largest element in the carpus. It articulates withboth the lateral centrale and intermedium, as well aswith distal carpals four and five. It likely articulatedwith the pisiform but this element appears to haverotated proximally out ofarticulation, making its exactmorphology and position difficult to determine. Distalcarpal four is the largest element in the distal carpalrow. In order of largest to smallest in size, the remaining distal carpals are respectively three, two, andfive.
The distal portion of metacarpals one and two areincomplete, but metacarpals three through five arecomplete. Fragments of digits two and four are foundin articulation with the respective metacarpals. Thecomplete complement of three phalanges is present fordigit five. The metacarpals all show epiphyseal platesat their proximal ends as do the phalangeal fragmentsfor digits two and four, and all the phalanges of digitfive.
Overview of Remaining Axial Skeleton (Figs. 1, 3)
The three most anterior vertebrae preserved areidentified as the fifth, sixth, and seventh cervicals. Carroll and deBraga (1992) identify seven cervicals in theTrieste Aigialosaur. The criteria used here to identifycervicals are the presence ofhypapophyseal peduncleson the fifth and sixth, and the absence of a sternal ribon the seventh; small, short, pointed ribs are associatedwith these vertebrae (Hoffstetter and Gasc, 1967, 1969).The cervical centra are subequal or slightly shorter,than the trunk centra.
There are twenty-one trunk vertebrae. The first tothe fifth are preserved with their ventral, calcified, costosternal cartilages in place and in articulation with thecalcified sternal cartilage. The ribs shorten quickly beginning at the fourteenth trunk rib. The ribs for thelast trunk vertebra are missing. Kornhuber (1893) described two sacral ribs; however, the pelvic region ispoorly preserved and this cannot be confirmed. Twelvecaudals are present. All vertebrae in this series havetransverse processes and either haemal arches or theirrespective haemal facets.
Pelvic Girdle and Limb (Figs. 1, 3)
The iliac blade is directed posteriorly. Its posteriorlimit is approximately three vertebrae posterior to the
FIGURE 1. Outline drawing ofholotype specimen of Carsosaurus marchesetti; modified from Kornhuber (1893). Thespecimen is preserved in ventral view; note the relationshipofthe interclavicle to the calcified sternum and coracoid, andthe varanoid-like nature of cervicals and presacrals as compared to the caudals. Housed in the Museo Civico di StoriaNaturale Trieste.

520 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 15, NO.3, 1995
c
·· ........ ssc
B
1cm-
A
H
Ie
FIGURE 2. Front limb and pectoral girdle of Carsosaurus marchesetti. A, Forelimb and portions of pectoral girdle basedon holotype; B, Reconstruction of forelimb and scapulacoracoid complex (with cartilages); C, Reconstruction of pectoral girdle(ventral view). Abbreviations: C, clavicle; el, lateral centrale; em, medial centrale; Cor, coracoid; IC, interclavicle; iot, intermedium; pis, pisiform; rl-5, sternal cartilages of presacral ribs 1-5; R, radius; rad, radiale; sCor, supracoracoid cartilage; sSc,suprascapular cartilage; sc, scapula; St, sternum; U, ulna; ul, ulnare; 2-5, distal carpals; i-v, metacarpals.
acetabulum. The acetabulum appears well developed,with good delimitation of the margins. The ischia andpubi are poorly preserved and cannot be adequatelydescribed. The femur is longer than the humerus, whilethe tibia and fibula are shorter than the radius and ulna(Table 2). A small patellar element is present as well.The tarsus is poorly preserved and only fragments remain from the metatarsus and phalanges. It is not possible to refute or substantiate the descriptions of theseelements as given by Komhuber (1893).
Discussion and Comparison
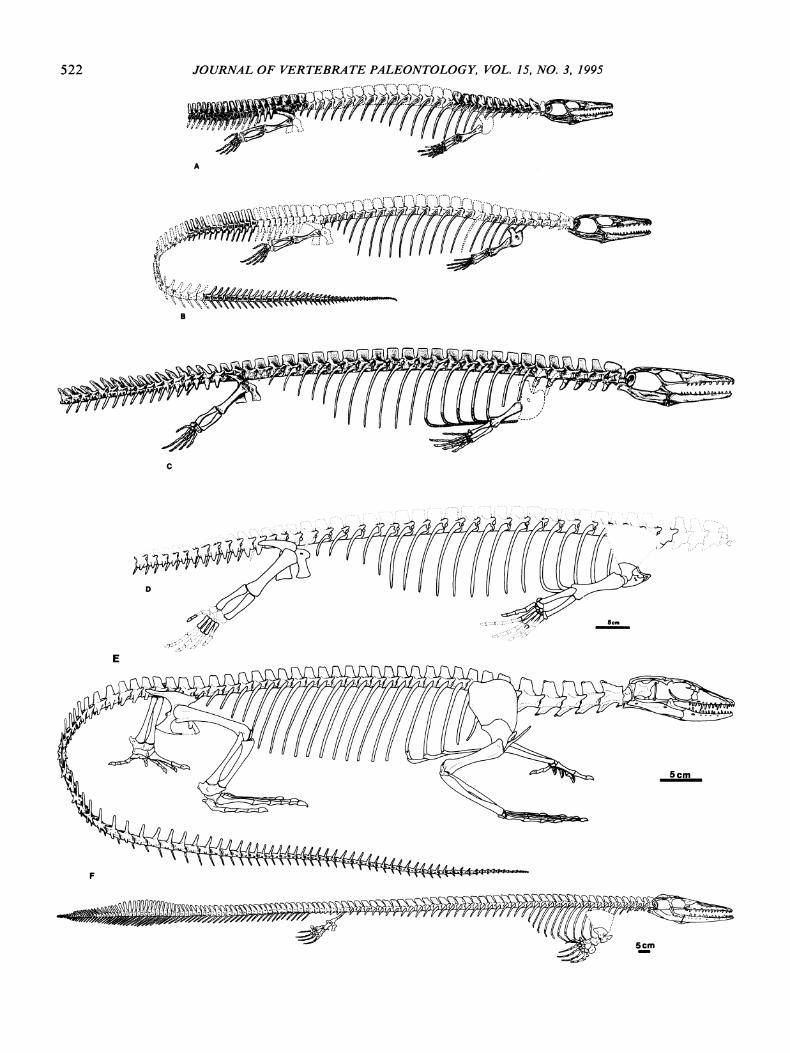
The condition of possessing short cervicals relativeto the trunk vertebrae is observed in aigialosaurs.. mosasaurs, (Fig. 3A-D, F), and most anguimorphs (Hoffstetter and Gasc, 1967, 1969; Hecht and Costelli, 1969).Extant varanids are the exception, where the cervical
centra are one-and-a-half times to almost twice thelength of trunk centra (Fig. 3E).
A lateral view of the axial skeleton of C. marchesettiis not possible. However, ventral examination of thecondyle-cotyle articulations shows that the transitionsin vertebral centrum morphology are similar to thoseof other aigialosaurs (Fig. 3A-D). The vertebral articulations show a progressive decrease in the concavityof the cotyle beginning in the caudal series (Fig. 1).The curvature of the condyle and the constriction ofthe centrum are reduced so that in ventral view thecentra appear spool-shaped or amphicoelic, not procoelic. Mosasaurs show similar changes in centrummorphology, but the effect begins in the anterior dorsals. The more vertical orientation of the articular facets has been considered an important adaptation toanguilliform locomotion in these aquatically evolvinganimals (Russell, 1967; Carroll and deBraga, 1992). It

CALDWELL ET AL.-CARSOSAURUS PECTORAL GIRDLE 521
is important to note that while the centra are approaching the mosasaur condition, the orientation ofthe zygapophyses of the cervicals and anterior trunkvertebrae in other aigialosaurs are varanoid-like (Fig.3A-C), though the presence of this feature cannot beconfirmed in C. marchesetti.
Carsosaurus marchesetti retains a high number ofelongate trunk ribs as in the Trieste Aigialosaur andO. bucchichi (Fig. 3B-D). This condition is plesiomorphic for squamates (Hoffstetter and Gasc, 1969) andsuggests that these three species are more primitivethan A. dalmaticus if Carroll and deBraga's (1992)characterization of rib length is correct.
Two other important features of C. marchesetti arethe number of sternal cartilages and the position in thetrunk series of the corresponding vertebrae; there arefive sternal cartilages articulating with trunk ribs oneto five. Assuming seven cervicals, as in other aigialosaurs (Carroll and deBraga, 1992), vertebrae eightthrough twelve articulate with the sternal ribs (the difficulty of defining the cervical-trunk transition makesit useful to compare positional relationships in bothrelative and absolute terms). In Varanus (Fig. 3F) thereare only three ribs articulating with the sternum. Thesearticulate with the third to the fifth trunk vertebrae, orvertebrae ten through twelve; trunk vertebrae one andtwo articulate with short, gravile ribs that do not reachthe sternum. Adult Lanthanotus have nine cervicalsand only two sternal ribs articulating with the first andsecond trunk vertebrae, or in absolute terms, vertebraeten and eleven (Rieppel, 1980b). A cleared and stainedjuvenile Lanthanotus shows a third cartilaginous sternal rib, originating on the sternum anterior to the ribwhich articulates with vertebra ten (Rieppel, 1992).However, the ossified rib articulating with vertebraenine, the putative terminal cervical, does not contactthis cartilaginous structure.
The condition seen in C. marchesetti may be primitive for anguimorphs, indicating that extant varanidshave lost two anterior rib articulations with the sternum. It is reasonable to assume that the five ribs ofthe Trieste Aigialosaur, as restored by Carroll anddeBraga (1992), articulated with trunk vertebrae oneto five as in C. marchesetti. However, such broad statements are complicated by other aspects of C. marchesetti (Fig. 2C). Its sternum bears paired xiphisternalcartilages found at the posterior apex of the sternalplate as well as the five sternal rib cartilages. In Varanusthe paired xiphisternals bear ribs which articulate withtrunk vertebra five. It is unfortunate that so little isknown about the sternum in mosasaurs as there appearto be a high number of sternal ribs, with a maximumof ten known in Plotosaurus (Camp, 1942; Russell,1967).
Limb Girdles and Locomotion
The limbs and girdles of C. marchesetti are fullyossified with well-defined articulating surfaces. Thepectoral girdle is varanoid-like (Fig. 2C), most closely
resembling that of Heloderma in terms of scapulocoracoid features and proportions (Lecuru, 1968a), andmost closely with Varanus regarding sternal proportions (Lecuru, 1968b). The articulations ofthe clavicle,interclavicle, coracoid, and sternal elements indicatethat the girdle functioned to increase the range of motion of the forelimb in a manner similar to that ofextant varanids (Jenkins and Goslow, 1983). In mosasaurs, the sternum and interclavicle are present butrarely preserved, while clavicles are known only fromMosasaurus (Russell, 1967). Without further materialit is very difficult to compare pectoral girdle structureand function between mosasaurs and other varanoids.
The coracoids of C. marchesetti, o. bucchichi, Heloderma (Lecuru, 1968a), and tylosaurine mosasaurs(Russell, 1967) are not fenestrated. Varanids have anterior and posterior fenestra, while the remaining mosasaurs, anguids, and Lanthanotus have only the anterior one. Gilmore (1922) described only an anteriorfenestra for the extinct varanid Saniwa ensidens. Thepolarity ofcoracoid fenestration is difficult to ascertianwhen mosasauroids are considered as members ofVaranoidea.
Though mineralized cartilages are poorly known inmosasaurs, it appears that the small scapula and largesuprascapular cartilage of varanoids, aigialosaurs, andmosasaurs (Fig. 3A-F) may be a synapomorphy ofanguimorphs. The suprascapular cartilage of the xenosaur Shinisaurus is almost twice the length of thescapula proper (Costelli and Hecht, 1971). Examination of iguanids, lacertids, and geckos (RM unnumbered specimens) shows that the suprascapular cartilage is equal in length to, or much shorter than, theossified portion of the scapula. The phylogenetic importance ofthis character is uncertain, but functionallyit affects both the origin, insertion, and relative massof shoulder and upper limb musculature.
Functional aspects of the humeral-scapulocoracoidarticulation in aigialosaurs appear similar to those oflimbed terrestrial anguimorphs. Articular surfaces arewell developed, unlike the mosasaur condition in whichthe articular surfaces are often rough and poorly finished. The superior lip of the glenoid fossa, locatedprimarily on the scapula, overhangs the inferior lip asin Varanus and Heloderma. The fossa is constructedto resist vertical dislocation of the humerus by jointloading during the impact phase of terrestrial locomotion. The glenoid in C. marchesetti is directed laterally with only a slight ventral component. A minorposterior component exists, but, as in Varanus, thisappears to result from the absence of a posterior bonycup on the glenoid. Resistance to posterior humeraldislocation at the point of maximal excursion mayhave been provided by strong cruciate ligaments at theshoulder joint as in Varanus (Haines, 1952; Jenkinsand Goslow, 1983). This idea is supported by the sizeofthe deltoid and pectoral tuberosities on the humerusof C. marchesetti (sites of insertion for the dorsal cranial and caudal, and ventral cranial and caudal, cruciate ligaments). Primitive mosasaurs possess large tu-
522 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 15, NO.3, 1995
A
~1t;{:~~{~N4'2' ~B
c
E

CALDWELL ET AL.-CARSOSAURUS PECTORAL GIRDLE 523
bercles and crests on the humeri, while more derivedforms have reduced tuberosities (Russell, 1967). Theposterior and ventral direction of the glenoid in mosasaurs suggests a very different function associatedwith aquatic locomotion, due largely to the absence ofdislocative joint loading encountered during terrestriallocomotion. This is not the case in aigialosaurs.
The potential terrestrial nature ofaigialosaurs is alsosupported by interpretations ofosteology and functionin the lower portion of the forelimb. The large olecranon process of the ulna would have stabilized theelbow during both pronation of the lower limb, andthe downward and forward thrust phases of forwardpropulsion. The olecranon is not as well developed asthat seen in extant varanoids. Due to major changesin the limbs the olecranon is often difficult to identifyin mosasaurs.
The carpi ofall aigialosaurs appear similar and showno obvious structural differences from the carpi of extant anguimorphs (Renous-Ucuru, 1973). However,differences between the carpi of aigialosaurs and mosasaurs are profound (Fig. 3A-D, F). In mosasaurs thecarpus is altered by reduction of the number of elements and the degree to which they are ossified. Reduction may have occurred by terminal deletions during development, or by chondrogenic-osteogenic fusions.
The phalangeal count of the manus in aigialosaursis plesiomorphic for squamates, though in C. marchesetti only the three phalanges of digit five are preserved. Phalangeal count is important as it indicatesthat neither reduction nor multiplication ofphalangealelements has occurred in C. marchesetti. Unlike mosasaurs, the ungual of digit five indicates that C. marchesetti had claws as did Aigialosaurus, Opetiosaurus,and other terrestrial anguimorphs (Fig. 3A-C).
The presence of two sacral vertebrae in C. marchesetti is plesiomorphic for squamates. However, theposterior limits of the iliac blade are slightly furtherposterior along the vertebral column, extending pastthe second sacral and onto the first caudal. In otheraigialosaurs the iliac blade extends only to the posteriormargin of the second sacral (Fig. 3A-D). In Varanus,Heloderma, and Lanthanotus, the iliac blade coversalmost three vertebral centra beyond the position ofthe acetabulum. The elongate posterior process is likelyrelated functionally to terrestrial locomotion in largeanguimorphs. A longer process would allow more muscle mass at the point of origin, as well as increase theeffective angle of contraction for the large flexor muscles.
PHYLOGENETIC ANALYSIS
Results
Branch-and-Bound search options found nine equally parsimonious cladograms during analysis ofthe datamatrix (Appendix I). Tree length is 107 steps, with aconsistency index (CI) of0.654. Both Strict Consensus(Fig. 4A) and 50% Majority-Rule Consensus (Fig. 4B)trees show 100% support for all major dichotomieswithin varanoids (Varanus + Heloderma + Estesia +Lanthanotus), and between varanoids and mosasauroids (Mosasauridae + Aigialosauridae). While theStrict Consensus (Fig. 4A) shows no resolution withinthe mosasauroids, the 50% Majority-Rule Consensus(Fig. 4B) supports a resolved Mosasauridae-Aigialosauridae dichotomy in 89% , or eight out of nine, ofthe most parsimonious trees. Neither consensus provides any resolution within the Aigialosauridae.
Only a single character (Character 58 [2]: elongateribs present only as far posteriorly as trunk vertebraeseven to nine) supports the tree topology responsiblefor the mosasauroid polytomy in the Strict Consensustree (Fig. 4A). Because of such tenuous support for asingle tree topology out of nine, the data set was alsotested with Character 58 excluded. This produced eightmost parsimonious trees (103 steps; CI = 0.66). Thetopology of the Strict Consensus tree for this reduceddata set was identical to the 50% Majority-Rule Consensus obtained for the full data set (Fig. 4B), therebystrengthening hypotheses of monophyly for the Mosasauridae and Aigialosauridae as independent mosasauroid clades. Overall mosasauroid (Aigialosauridae + Mosasauridae) monophyly is supported in boththe first and second (reduced data set) analyses.
Varanoidea (sensu Pregill et aI., 1986), including Estesia sinensis, is a robust clade with consistent supportat all levels of our analysis and an identical topologyto that hypothesized by Norell et al. (1992). The relationships of mosasauroids with other anguimorphs(xenosaurids and anguids) outside of Varanoidea, areuncertain at this point. As Gauthier (1982) stated indiscussing the uncertainty of xenosaurid-anguid-varanoid relationships, this analysis shows only that mosasauroids are non-varanoid anguimorphs.
The use of Gerrhonotus to polarize character transformations in this analysis serves as a tool in testingconventional hypotheses of relationship within varanoids. The trees we have presented (Fig. 4A, B) wereintentionally rooted at an internal basal polytomy tocommunicate our uncertainty ofbasal anguimorph relationships. We acknowledge the paraphyly of such an
~
FIGURE 3. Reconstructions and skeletal drawings of fossil and extant varanoids. A, holotype of Aigialosaurus dalmaticus;B, holotype of Opetiosaurus bucchichi; C, Trieste Aigialosaur; D, holotype of Carsosaurus marchesetti; E, skeleton of Varanussalvator; F, reconstruction of Clidastes sternbergi.

524 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 15, NO.3, 1995
B I-<
Majority rule ~0
eu CI)CI) ca
CI) CI) cd 2 2 '6'0 CI)
:::3 cd :::3 :z~
20 8 0 ~
:::3 :.(~Q Q CI)
cd 0 ~
~0 eu cd :::3 ·m tI) ca 0 0
-E "0 ...c:: Q cd '£0 Q e B tI) '6"0 eu
~"i) 0 0.. E5d cd cd~ ~ :.(::I: .....:l ;> 0
Discussion
Our goal with this analysis was not to review orrevise phylogenetic hypotheses regarding the Anguimorpha, but rather to assess the relationships of aigialosaurs. We do not therefore, intend to discuss at
analysis through exclusion of some fossil taxa (necrosaurids, Saniwa, Cherminotus), as well as the possible polyphyly resulting from the absence of xenosaurids and other anguids. However, this last point isderived from a posteriori recognition of clade statusfor mosasauroids apart from varanoids. As our goalwas a preliminary analysis of the conventional taxonVaranoidea, new phylogenetic hypotheses, based onrecognition of our results here, can now be tested.
length the character transformation series implied byour results for all characters. These will be consideredin forthcoming studies of the Anguimorpha. However,there are a number of characters and character states(Characters 3-6, 11-13, 16-17, concerned with narialretraction) which can be discussed confidently in termsof homology and transformation between varanoidsand mosasauroids.
Nares and Narial Retraction (Fig. 5A-D): The degreeof narial retraction has been cited as a synapomorphyofvaranids and mosasauroids without attention to theactual homologous structures involved. It cannot bestressed strongly enough that an open space (fenestra),created by the differential spatial interaction of numerous developmental units (bones, soft tissues), cannot be treated as one independent character. Each element involved in such a structure must be considerednot in terms of the fenestra itself, but in terms of itstopological relations with other developmental units.This erroneous notion ofhomolog characterization hasmisconstrued notions of synapomorphy and relationships within the conventional taxon Varanoidea (for areview Fejervary, 1918; McDowell and Bogert, 1954).
In Varanus (Fig. 5A) (Bahl, 1938; Rieppel, 1980a)perceived narial retraction results from the following:elongation of the narial and vomerine processes of thepremaxilla; anterior elongation and narrowing of thenasal bone and medial retraction of the nasal from theprefrontal and maxilla; elongation of the anterolateralprocesses of the frontal around the posterior processesof the nasal; shortening of the prefrontal; exposure ofthe septomaxilla; elongation of the vomer anterior tothe vomerine aperture; and anterior elongation of themaxilla, with medial expansion and elongation anteriorly.
In aigialosaurs, and in less derived mosasaurs (Fig.5D) such as Halisaurus and some mosasaurines (Russell, 1967; deBraga and Carroll, 1993), narial retractionresults from quite different structural relationships: extreme elongation of the narial process of the premaxilla; elongation of the maxilla and development of ananterodorsal maxillary process; extreme elongation andnarrowing of the septomaxilla (Camp, 1942); loss orfusion of the nasal bone with the anterior process ofthe frontal with no evidence of nasal elongation contributing to the internarial bar as in Varanus (in somemosasaurs there is evidence of nasals, but these aresmall and lie at or near the junction of the narial barand frontal [Russell, 1967]); extreme elongation of thefrontals anterior to the anterior orbital margin (exceptPlotosaurus bennisoni; Camp [1942]); no size decreasein the anterior portion of the prefrontals (in Ectenosaurus clear contact is maintained between the maxilla,prefrontal and frontal, preventing apparent narialemargination to a point superior to the maxillary-lacrimal suture as in Varanus).
In short, the narial opening has not retracted at all.The frame of reference for elongation should not befrom the snout backwards, but rather from the orbit-
I-<
~eu CI)
0
B CI) cd e CI) caCI)
cd B ] ~ '6'0~ ~
~
0 8 0 ~ :.( ~Q Q CI) a ~0 eu cd ~ ·m
0 0~
~
-E '1:l oS ~ ~ ca '£ §0"i) Q ~ B 0 '6"0 eu
eu cd~ ::E :.( ~ E5 cd
d ::r: .....:l ;> U
A
FIGURE 4. Consensus cladograms of mosasauroid-varanoid interrelationships based on nine most parsimonius trees(107 steps; CI = 0.654) derived from the data matrix inAppendix I. A, Strict Consensus tree ofrelationships. B, 50%Majority-Rule Consensus tree. It is important to note thatin 89% (8 out of 9) of the most parsimonious trees that theMosasauridae is resolved relative to the aigialosaur polytomy. Only one character in one topology, Character 58 (0),places the Mosasauridae in a clade with Aigialosaurus dalmaticus.
Strict

CALDWELL ET AL.-CARSOSAURUS PECTORAL GIRDLE 525
B 0
c
FIGURE 5. Dorsal view ofnarial regions in three anguimorph diapsids. A, Varanus salvator (RM 1969.30a). B, Gerrhonotusmulticarinatus (unnumbered RM specimen). C, dorsolateral view of Gerrhonotus multicarinatus to illustrate the size and shapeofthe septomaxilla. D, Plotosaurus bennisoni (UCMP 32778), narial bar ofpremaxilla has been cut away (redrawn from Camp,1942). Abbreviations: f, frontal; lac, lacrimal; mx, maxilla; 0, nasals; pi, palatine; pm, premaxilla; prf, prefrontal; sm, septomaxilla; v, vomer.
braincase region forward. In relative proportions, thisregion is equivalent in the skulls of Varanus (Fig. 5A),Heloderma, and Gerrhonotus (Fig. 5B, C) we examined(see also the illustrations provided by Rieppel [1980a]),and in aigialosaurs and mosasaurs (Fig. 5D). Changesin the topological relation of the nasals with the maxillae and prefrontals appear to be the major effector ofnarial enlargement (Heloderma) compared to the plesiomorphic condition (Gerrhonotus). The elongate narialbar is a result of snout elongation and should not bethought of as the effector of that change. The elongatefrontals of aigialosaurs and mosasaurs may be a resultof ontogenetic fusion with the nasals. The elongatefrontals of mosasauroids extend past the orbits abovethe point where the orbital process of the maxilla expands dorsally; this is equivalent to the position of thenasal-premaxillary bar contact in Varanus (Fig. 5A).The only mosasauroid exception is Plotosaurus bennisoni (Fig. 5D) where the frontal is short, the nasalsare present and small, and the prefrontals are excludedfrom the narial margin. In terms of sutural relationships, the insertion of the narial bar into the frontal inmosasauroids is identical to the insertion of the barinto the nasals in Varanus. Together, these pointsstrongly suggest that ontogenetically, mosasauroidspossessed nasals which fused to the frontals, but maintained the plesiomorphic topology with the prefrontals,and variably, the maxillae. This condition is in directcontrast to Varanus. Snout elongation is therefore accomplished convergently in mosasauroids and varanoids.
MORPHOMETRIC ANALYSIS
Results
In both morphometric analyses, principal component (PC) 1 and PC2 explained over 98% of the totalvariance in the data set (Table 3). The percentage oftotal variance explained by PC3 and PC4 combinedwas negligible and these components were thus notconsidered further. Loadings for PCl in both caseswere all positive, identifying this component as an indicator of size variation between taxa, while PC2 loadings included both positive and negative values, characterizing it as a shape component (Table 3).
Analysis I-Loadings of PC1 are all nearly equal inmagnitude. Conversely, loadings of PC2 emphasizerelative lengths of ulna, femur, and the heads of thehumerus (Table 3). Graphic representaiton ofPC1 andpe2 (Fig. 6A) placed Carsosaurus in a position nearlyequidistant to either Varanus or the aigialosaurs Opetiosaurus and Aigialosaurus, along both the size (PCl)and shape (PC2) components. The distance from thesetaxa to the mosasaur Clidastes is greater along bothsize and shape axes. While Clidastes and Varanus occupy positions of maximum size and shape, respectively, Opetiosaurus and Aigialosaurus cannot be differentiated confidently along either axis (Fig. 6A). Alltaxa except the aquatic Clidastes fit well (r2 = 0.955)along a line indicating proportionality (slope = 1.11)of size and shape. Morphometric change for both sizeand shape occurs gradually from Lanthanotus at oneextreme to Varanus at the other, with the three aigi-

526 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 15, NO.3, 1995
TABLE 3. Prinicipal component loadings (PCL) for a principal component analysis of measurements from varanoidsand mosasauroids. Analyses were conducted in a stepwisefashion to circumvent data limitations. The first analysis (I)included ten variables and omitted the Trieste aigialosaur.The second analysis (II) included seven variable and combined data for Aigialosaurus and Opetiosaurus. Characterswith the greatest contribution to each PCL (assigned arbitrarily) are indicated by asterisk (*). Abbreviations of measurements are as listed in Table 2.
2
N
U 0~
-I
A
lanthanotus
Varanus 0
II
PCL 1 PCL2 PCL 1 PCL2
HUM 0.958 0.255HUMpRox 0.934 -0.331 *HUMD1sT 0.931 -0.348* 0.913 0.363UL 0.877 0.478* 0.879 -0.464*RAD 0.991 * 0.127ADV 0.931 -0.278 0.930 0.305FEM 0.935 0.345* 0.937 -0.335FEMpRox 0.980* -0.188 0.978* 0.201FEMD1ST 0.946 -0.321 0.940 0.339FIB 0.886 -0.460*TIB 0.949 0.2910/0 variation 89.1 9.6 85.3 13.1
Clidastes 0
-2-2 -I 0 2
PC 1
2
B 0Clidastes
N
U 0~
OCarsosaurus
-I
Varanus 0
2oPC 1
-I-2~--~-_---L. ..I..-__-J
-2
FIGURE 6. Principle component analyses (PCA) using datafor Aigalosaurus dalmaticus, Opetiosaurus bucchichi, TriesteAigialosaur, Carsosaurus marchesetti, Varanus salvator,Lanthanotus borneensis, Heloderma sp., and Clidastes propython (Table 2). A, PCA (Analysis I), exclusion ofthe TriesteAigialosaur and the variable "length of the fibula." Slope ofregression line for all taxa except Clidastes is 1.11 (r2 =0.955). B, PCA (Analysis II) including the Trieste Aigialosaurbut with a reduction in the number of variables to seven,combining the data ofAigialosaurus and Opetiosaurus. Slopeof regression line for all taxa except Clidastes is - 1.02 (r2 =0.885).
alosaurs and Heloderma occupying intermediate positions (Fig. 6A).
Analysis II-Loadings for PC1 (size axis) in this caseare influenced nearly equally by the measurements,while the shape axis is greatly influenced by the fibularand ulnar lengths (Table 3). Graphically, Clidastes liesat the high extreme for both size and shape axes (Fig.6B). Both Clidastes and Varanus hold positions nearlyequidistant from an "aigialosaur morphospace," as defined by the presence ofAigialosaurus (=Opetiosaurus),Carsosaurus, and the Trieste Aigialosaur. The latter isin a position intermediate to Carsosaurus and Aigialosaurus (=Opetiosaurus). As in the previous analysis,all taxa, except Clidastes, lie closely along a straightline (r2 = 0.885; slope = -1.02) with Lanthanotus andVaranus at the upper and lower extremes, respectively.
Discussion
The most important conclusions to be drawn fromthis approach reflect on functional aspects of limb proportions relative to body size. Since the position ofCarsosaurus is equivocal when compared to Lanthanotus, Varanus, and Clidastes (near the centroid of anequilateral triangle), consideration of the relative influence of variables and their component loadings becomes important. The selective, though necessary, exclusion of variables in these two analyses results in astepwise operation of sorts, which assists in determining some functional relationships between variables.While size is generally of great influence on structure,the dual analysis conducted here shifts the importanceof the simple length measurements from size in thelarger data set, to shape in the more exclusive data set
(Table 3). Thus, in defining limb and body characteristics, aspects of size variability and functionality, asinferred from shape, can be incorporated.
Conventional varanoid taxonomy can easily be derived from the groupings shown here (Fig. 6A, B), suggesting that past hypotheses have been based on measures ofoverall similarity. Such studies have attemptedto represent a phylogenetic continuum from morphometric information we interpret as a functional continuum (McDowell and Bogert, 1954; Borsuk-Bialynicka, 1984; Carroll and deBraga, 1992).
We have interpreted our PCA results to indicate twofunctionally disparate groups. The first is representedby one unique and disparate taxon, the aquatic mosasaur Clidastes. The second is a series of taxa fromLanthanotus to Varanus, arranged linearly along afunctional continuum, indicating adaptive trends from

CALDWELL ET AL.-CARSOSAURUS PECTORAL GIRDLE 527
fossoriality to active locomotor pursuit. This suggeststhat aigialosaurs were more like Heloderma in termsof their locomotor capabilities and limb function. Further, aigialosaurs cannot be assumed to have beenaquatic reptiles on the basis of limb morphometrics.Their limbs do not show any size or shape changesindicative ofaquatic adaptation anymore than do Heloderma and Lanthanotus. The lateral compression ofthe tail and changes in caudal centrum size and articular surfaces support hypotheses ofaquatic habits, butcannot prove them. There is no evidence that havinga mosasaurian skull is an aquatic adaptation. In thecontext of the phylogenetic hypotheses presented (Fig.4A, B) and the results of PCA (Fig. 6A, B), it is betterstated that mosasaurs possess aigialosaurian cranialand caudal characteristics.
The placement of Carsosaurus in "aigialosaur morphospace," as defined by the two most important principal components (Fig. 6A, B), and despite limitationsof the data set, also provides some additional supportfor our preliminary phylogenetic hypotheses (Fig. 4A,B). Morphometrically, the intermediate position ofCarsosaurus in both analyses (Fig. 6A, B) confirmsgross morphological impressions that it is clearly closerin size and shape to other aigialosaurs than it is to nonaigialosaurs. The distance of Carsosaurus from Aigialosaurus (=Opetiosaurus) and the Trieste Aigialosauralong the size axis (PC1) may be explained by its largersize, a factor not removed by PCA. This analysis showsthat Carsosaurus cannot be excluded from the Aigialosauridae on the basis of minor size and shape ratios(Carroll and deBraga, 1992).
CONCLUSIONS
Carsosaurus marchesetti is an important source ofinformation for structure, function, and evolution ofthe postcranial skeleton during phylogenesis of earlymosasauroid anguimorphs. Phylogenetic analysis indicates that C. marchesetti is an aigialosaur, and thatmosasauroids (mosasaurs + aigialosaurs) are a monophyletic assemblage distinct from Varanoidea. The exact position of mosasauroids within Anguimorpha isuncertain but it is tentatively accepted here that membership is within that clade.
Osteological information indicates that the skeletonof Carsosaurus is very similar to that of some anguids,lanthanotids, and helodermatids (in carpus structure,large number of sternal ribs, absence of coracoid fenestra, and relative limb to body proportions). Thisalone suggests a terrestrial habit and terrestrial locomotor capabilities. The position of aigialosaurs, between Lanthanotus and Varanus, in a PCA comparingthe proportions and shape of limbs to the body (Fig.6A, B), can be easily interpreted in terms of function.Aigialosaurs are clearly intermediate in a linear seriesthat describes active terrestrial locomotion to fossoriallocomotor habits, further supporting the conceptualization of aigialosaurs as terrestrial animals.
This information, in association with lithofacies
analysis and interpretations of depositional environments from aigialosaur fossil localities (Calligaris,1988), suggests that it is reasonable to conclude thataigialosaurs lived along or near coastal margins. Theymay in fact have had aquatic habits, but there is littleosteological evidence to support speculations that aigialosaurs were obligatorily aquatic. Postcranially, theyare plesiomorphic compared to their aquatic sistergroup (Mosasauridae). However, by virtue oftheir discovery in marine deposits, and cranial synapomorphies shared with mosasaurs, it has been concludedthat these animals were at least facultatively aquatic.From the observations presented here, the only osteological feature which supports such an interpretationis the most posterior portion of the tail. The anteriorcaudals show the same vertical aspect of the neuralspines and haemal arches as do terrestrial varanids.The relative proportions of the transverse processesare also similar to both varanids and helodermatids.Only the posterior caudals show both an absence oftransverse processes and laterally compressed centra.Cranial characters, such as the purported aquatic adaptation of a calcified tympanum, a synapomorphy ofmosasaurs and aigialosaurs, is symplesiomorphic withtheir unknown terrestrial mosasauroid common ancestor. From this perspective, mosasaurian innovations for successful adaptation to aquatic environments were entirely postcranial.
The primitive anguoid-like postcranium of Carsosaurus is significant in terms of limb evolution. In aglobal sense, the morphology of the terrestrial lacertilian limb has changed very little from that of earlydiapsids (Carroll and Currie, 1991; Caldwell, 1994).Only flying and limb-assisted swimming require structural modifications which significantly alter the bonypatterns and proportions of the limb (limb loss is notconsidered in this statement). Swimming reptiles, suchas mosasaurs and ichthyosaurs, reduced their limb proportions and evolved locomotor styles where propulsive forces were generated by muscles supported by theaxial skeleton, not the limb skeleton (Carroll, 1985).This sort of shift in the limbs of Carsosaurus cannotbe derived from either the qualitative descriptions wehave given, nor from the results of quantitative analysis (PCA).
The available postcranial material of other aigialosaurs shows no major differences in anatomy whichwould suggest family-level distinction for C. marchesetti. Phylogenetic relationships indicate that C. marchesetti is allied with other aigialosaurs and not withVaranoidea or Mosasauridae. Our analysis qualitatively (phylogenetic) and quantitatively (PCA) supports Carroll and deBraga's (1992) suggestion that Opetiosaurus and Aigialosaurus may belong to the samegenus; the posterior shortening of the ribs is the onlyequivocal character. If both are members of the samegenus, then Aigialosaurus Kramberger, 1892 wouldhave priority. Carsosaurus and the Trieste Aigialosaurcannot be differentiated qualitatively from Opetiosaurus on the basis of postcranial material, but we retain

528 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 15, NO.3, 1995
C. marchesetti as a valid taxon pending discovery ofassociated cranial and postcranial material. We conclude with the following systematic treatment of theaigialosaur taxa examined in this study
Infraorder ANGUIMORPHA Fiirbringer, 1900Superfamily MOSASAUROIDEA Camp, 1923
Family AIGIALOSAURIDAE Kramberger, 1892
Genus AIGIALOSAURUS Kramberger, 1892
Type Species-Aigialosaurus dalmaticus Kramber-ger, 1892, PI. 3.
Referred Genus - Opetiosaurus Komhuber, 1901.Holotype-BSP 190211501.Diagnosis-as in Kramberger (1892).Assigned Species-A. bucchichi (Komhuber, 1901),
(Komhuber, 1901, PI. 1-2).Holotype-NMW unnumbered specimen.Diagnosis-as in Komhuber (1901).
Genus CARSOSAURUS Komhuber, 1893
Type Species - Carsosaurus marchesetti Komhuber,1893, PI. 1.
Assigned Materials-Trieste specimen (MCSNT,11430,11431, and 11432 [three parts]); Carroll anddeBraga (1992:figs. 4, 8).
Holotype-MCSNT specimen figured by Komhuber(1893, PI. 1).
Syntype-MCSNT, 11430, 11431,and 11432(threeparts), figured by Carroll and deBraga (1992:figs. 4,8).
Diagnosis-as in Komhuber (1893).
ACKNOWLEDGMENTS
We thank P. Gaskill for illustrating five of the reconstructions shown in Figure 3. Data on mosasaurswas obtained by the first author while visiting the Museum of Paleontology, University of Kansas. The assistance of L. Martin and J. Chom is greatly appreciated. R. L. Carroll gratefully acknowledges the assistance of Dr. R. Calligaris and the staff at the MuseoCivico di Storia Naturale, Trieste, Italy, while preparing, drawing, and measuring the specimen of Carsosaurus marchesetti. O. Rieppel and S. Evans are gratefully acknowledged for making useful comments onthe manuscript. This research was supported by a Natural Sciences and Engineering Research Council ofCanada (NSERC) Post Graduate Scholarship to M.W.Caldwell, and by NSERC operating grants to R. L.Carroll.
LITERATURE CITED
Bahl, K. N. 1938. Skull of Varanus monitor (Linn.). Records ofthe Indian Museum (Journal ofIndian Zoology)39: 133-174.
Borsuk-Bialynicka, M. 1984. Anguimorphans and relatedlizards from the Late Cretaceous of the Gobi Desert,Mongolia. Palaeontologica Polonica 46:5-105.
Caldwell, M. W. 1994. Developmental constraints and limb
evolution in Permian and modern Lepidosauromorphdiapsids. Journal of Vertebrate Paleontology 14:459471.
Calligaris, R. 1988. I rettili fossili degli "Strati calcarieiittilitici di comeno" e dell'isola di lesina. Atti del MuseoCivico di Storia naturale Trieste 41 :85-125.
Camp, C. L. 1942. California Mosasaurs. University ofCalifornia Memoirs 13: 1-68.
Carroll, R. L. 1985. Evolutionary constraints in aquaticdiapsid reptiles. Special Papers in Palaeontology 33: 145155.
--- and P. J. Currie. 1991. The early radiation ofdiapsid reptiles; pp. 354-424 in H.-P. Schultze and L. Trueb(eds.), Origins ofthe Higher Groups ofTetrapods. Comstock Publishing Associates, Ithaca.
--- and M. deBraga. 1992. Aigialosaurs: Mid-Cretaceous varanoid lizards. Journal of Vertebrate Paleontology 12:66-86.
Costelli, J., Jr., and M. K. Hecht. 1971. The postcranialosteology of the lizard Shinisaurus: the appendicularskeleton. Herpetologica 27:87-98.
deBraga, M., and R. L. Carroll. 1993. The origin of mosasaurs as a model of macroevolutionary patterns andprocesses. Evolutionary Biology 27:245-322.
Estes, R., K. De Quieroz, and J. Gauthier. 1988. Phylogenetic relationships within Squamata; pp. 119-281 inR. Estes and G. Pregill (eds.), Phylogenetic Relationshipsof the Lizard Families. Stanford University Press, Stanford.
Fejervary, G. J. de. 1918. Contributions to a monographyon fossil Varanidae and Megalanidae. Budapesch, Annales Historico-Naturales Musei Nationalis Hungarici16:341-467.
Gauthier, J. A. 1982. Fossil xenosaurid and anguid lizardsfrom the early Eocene Wasatch Formation, southeastWyoming, and a revision of Anguoidea. Contributionsto Geology, University of Wyoming 21:7-54.
Gilmore, C. W. 1922. A new description ofSaniwa ensidensLeidy, an extinct varanid lizard from Wyoming. Proceedings of the United States Museum 60: 1-28.
Good, D. A. 1987. A phylogenetic analysis of cranial osteology in the gerrhonotine lizards (Squamata: Anguidae). Journal of Herpetology 21:283-295.
--- 1988. The phylogenetic position of fossils assignedto the Gerrhonotinae (Squamata: Anguidae). Journal ofVertebrate Paleontology 8: 188-195.
Haines, R. W. 1952. The shoulder joint of lizards and theprimitive reptilian shoulder mechanism. Journal ofAnatomy 86:412-422.
Hecht, M. K., and J. Costelli, Jr. 1969. The postcranialosteology of the lizard Shinisaurus 1. The vertebral column. American Museum Novitates 2378:1-21.
Hoffstetter, R., and J.-P. Gasc. 1967. Observations sur Iesquelette cervical et specialement sur les hypapophysesdes sauriens Varanoides actuels et fossiles. Bulletin duMuseum D'Histoire Naturelle, Deuxieme Serie 39:10281043.
--- and --- 1969. Chapter 5. Vertebrae and ribs ofmodern reptiles; pp. 201-310 in C. Gans (ed.), Biologyof the Reptilia, Vol. 1. Academic Press, New York.
Jenkins, F. A., and G. E Goslow. 1983. The functionalanatomy ofthe shoulder ofthe Savannah monitor lizard(Varanus exanthematicus). Journal of Morphology 175:195-216.
Kornhuber, A. G. 1893. Carsosaurus marchesetti, ein neuerfossiler Lacertilier aus den Kreideschichten des Karstes
CALDWELL ET AL.-CARSOSAURUS PECTORAL GIRDLE 529
bei Komen. Abhandlungen der geologischen Reichsanstalt Wien 17: 1-15.
--- 1901. Opetiosaurus bucchichi, eine neue fossile Eidechse aus der unteren Kreide von Lesina in Dalamtien.Abhnadlungen der geologischen Reichsanstalt Wien 17:1-24.
Kramberger, K. G. 1892. Aigialosaurus, eine neue Eidechseaus den Kreideschiefern der Insel Lesina mit Riicksichtaufdie bereits beschriebenen Lacertiden von Comen undLesina. Glasnik huvatskoga naravolosovnoga derstva(Societas historico-matulis croatica) u Zagrebu 7:74-106.
Kuhn, O. 1958. Ein neuer Lacertilier aus dem frankischenLithographieschiefer. Neues Jahrbuch fur Geologie undPalaontologie, Monatshefte 1958:380-382.
Lecuru, S. 1968a. Remarques sur Ie scapulo-coracoldes desLacertiliens. Annales des Sciences Naturelles, Zoologie,Paris, Douzieme Serie, 10:475-510.
--- 1968b. Etude des variations morphologiques dusternum, des clavicules et de l'interclavicule des Lacertiliens. Annales des Sciences Naturelles, Zoologie, Paris, Douzieme Serie, 10:511-544.
McDowell, S. B., Jr., and C. M. Bogert. 1954. The systematic position of Lanthanotus and the affinities of theanguimorph lizards. Bulletin of the American Museumof Natural History 105:1-142.
Meszoely, C. A. M. 1970. North American fossil anguidlizards. Bulletin ofthe Museum ofComparative Zoology139:87-150.
Norell, M. A., M. C. McKenna, and M. J. Novacek. 1992.
Estesia mongoliensis, a new fossil varanoid from theLate Cretaceous ofthe Barun Goyot Formation ofMongolia. American Museum Novitates 3045: 1-24.
Pregill, G. K., J. A. Gauthier, and H. W Greene. 1986. Theevolution of helodermatid squamates, with descriptionof a new taxon and an overview of Varanoidea. Transactions of the San Diego Society of Natural History 21:167-202.
Renous-Lecuru, S. 1973. Morphologie comparee du carpechez les Lepidosauriens actuels (Rhyncocephales, Lacertiliens, Amphisbeniens). Gegenbaurs morphologischesJahrbuch, Leipzig 119:727-766.
Rieppel, O. 1980a. The Phylogeny of Anguinomorph Lizards. Birkhauser Verlag, Basel, 86 pp.
--- 1980b. The postcranial skeleton of Lanthanotusborneensis (Reptilia, Lacertilia). Amphibia-Reptilia 1:95112.
--- 1992. The skeleton ofajuvenile Lanthanotus (Varanoidea). Amphibia-Reptilia 13:27-34.
Russell, D. A. 1967. Systematics and morphology ofAmerican mosasaurs. Peabody Museum of Natural History,Yale University Bulletin 23:1-241.
Swofford, D. L. 1993. PAUP: Phylogenetic Analysis UsingParsimony, Version 3.1.1 Computer program distributed by the Illinois Natural History Survey, Champaign,Illinois.
Received 9 September 1993; accepted 14 July 1994.
APPENDIX I. Data Matrix
5 10 15 20 25 30
Gerrhonotus 00000 00000 00000 00000 00000 00000Heloderma 01010 00011 01111 11001 00100 10100Lanthanotus 01001 10010 11101 11001 11101 11110Varanus 01101 11110 11010 01010 11011 01010Estesia 01011 10011 01101 11010 OlIO? 00010Mosasauridae 10?01 10100 00000 01100 0101? 01000Aigialosaurus ????? 1??00 ?OOO? 0010? ?OO?? 0100?Opetiosaurus 10??? ?1??? ?OO?? OO??? ?OO?? O?OOOTrieste Aigialosaur ????? ????? ????? ????? ????? ?????Carsosaurus ????? ????? ????? ????? ????? ?????
35 40 45 50 55 60 65
Gerrhonotus 00000 00000 00000 00000 00000 00000 00000 0Heloderma 10001 11110 01101 10000 10100 00101 10000 0Lanthanotus 01101 00010 11111 11110 11001 10111 00010 0Varanus 01110 10111 11100 11110 11121 11011 01011 1Estesia ?OOOO 1111? 111?0 ?1??0 10??? ????? ????? ?Mosasauridae 00000 10100 00010 00011 01002 00202 00210 1Aigialosaurus 100?? ????? ????? ???11 01002 012?0 ??1?0 0Opetiosaurus 100?? ????? ????? ???11 010?? 010?? 101?? 0Trieste Aigialosaur 1000? ????? ????? ????1 01012 01000 ????? ?Carsosaurus ????? ????? ????? ????? ???1? 00000 10110 0
Character state abbreviations: ? = not preserved/unknown.
530 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 15, NO.3, 1995
APPENDIX II
SPECIMENS AND CHARACTERDESCRIPTIONS
Descriptions ofcharacters and character states identified in the varanoids and mosasauroids examined.Numbered characters followed by parentheses containing a number and an 'R' are found in Rieppel(1980a); those with a 'P' are found in Pregill et al.(1986). In several cases characters were recoded to reflect the character states of taxa considered for thisanalysis. Examination of the following specimens andpublished descriptions were used in association withPregill et al. (1986) and Rieppel (1980a): Varanus salvator RM 1969.30a, and Bahl (1938), McDowell andBogert (1954), Norell et al. (1992); Lanthanotus, FMNH134711, and McDowell and Bogert (1954), Rieppel(1980b; 1992), Norell et al. (1992); Heloderma sp., RMspecimen, and McDowell and Bogert (1954), Gauthier(1982), Norell et al. (1992); Estesia, Norell et al. (1992);Clidastes prophython, KU 1022 and Russell (1967);Aigialosaurus dalmaticus, BSP 190211501, and Kramberger (1892), Carroll and deBraga (1992); Opetiosaurus buccichi, NMW specimen, and Kornhuber (1901),Carroll and deBraga (1992); the Trieste Aigialosaur,MCSNT, 11430, 11431, and 11432 (three parts), andCarroll and deBraga, (1992); and Carsosaurus marchesetti, MCSNT unnumbered specimen, and Kornhuber (1893).
Cranium
1(IR)
2 (23P)
3 (2R)
4 (2P)
5 (3--4P)
6 (5P)
7 (3R)
8 (6R)
9 (28-29P)
10 (30P)
11 (lOR)12 (13R)13 (14R)
Number ofpremaxillary teeth: more than6 (0); less than 6 (1).Premaxillary teeth, size: same size orlarger than maxillary teeth (0); abruptlysmaller than maxillary teeth (1).Nasal process ofpremaxilla elongate entering deeply between nasal bones: no(0); yes (1).Nasal and prefrontal bones: little or nocontact (0); broad contact (1).Nasal and maxillaries: contact (0); nocontact (1).Nasal process of maxilla: arises frommiddle ofmaxilla (0); arises on posterioraspect of maxilla (1).Premaxillary foramen: absent (0); present (1).Vomerine process of premaxilla: shortand blunt (0); elongate (1).Maxillary tooth number: greater thantwelve (0); less than twelve (1).Venom groove on maxillary teeth: absent (0); present (1).Nasal bones: paired (0); fused (1).Frontal bones: fused (0); paired (1).Contribution offrontal to dorsal marginof orbit: enter the dorsal margin (0); excluded from margin (1).
14 (16R)
15 (17R)
16 (lOP)
17 (20R)
18 (22R)
19 (23R)
20 (24R)
21 (25R)
22 (27R)
23 (30R)24 (32R)
25 (33R)
26 (35R)
27 (14P)
28 (13P)
29 (18P)
30 (37R)
31 (38R)
32 (39R)
33 (20P)
34 (41R)
Palate
35 (42R)
36 (44R)
Descensus frontalis development: poorlydeveloped with no sutural contact belowolfactory tract (0); well developed andcontacts in a suture below olfactory tract(1).Descensus frontalis separation of orbitae: does not separate orbits (0); separates orbits (1).Prefrontal contact with postfrontal: nocontact (0); contact or closely approaches(1).Postfrontal and postorbital: separate (0);fused (1).Lacrimal bone lateral exposure: extensive (0); narrow (1).Lacrimal spine on caudal edge lateral tolacrimal foramen: no spine (0); spinepresent (1).Lacrimal separation of prefrontal fromjugal: yes (0); no (1).Foramen pro ductus lacrimalis: single (0);double (1).Jugal has distinct triangular projectionon caudo-ventral edge: yes (0); no (1).Pineal foramen: present (0); absent (1).Supratemporal process of parietal contact: contacts distal end of paroccipitalprocess (0); separate from paroccipitalprocess (1).Contact between supraoccipital and parietal: narrow (0); broad contact (1).Upper temporal arch: complete (0); interrupted (1).Temporal musculature insertion on parietal table: ventral (0); dorsal (1).Squamosal size and contact with postorbital process: large and contacts postorbital (0); small and reduced (1).Supratemporal process ofparietal in dorsal aspect: broad (0); narrow (1).Squamosal contact with supratemporalprocess ofparietal: no contact (0); in frontof supratemporal (1).Posterior tip of squamosal articulationwith quadrate: in a socket on dorsal surface of quadrate (0); notch on laterodorsal edge of quadrate (1).Anterodorsal edge of quadrate: broadtransversely (0); narrow and pointed (1).Quadrate outer conch: large (0); reduced(1).Mesial crest of quadrate: distinct (0);weakly developed (1).
Palate: neochoanate (0); palaeochoanate(1).Length ofchoanal groove: more than half

CALDWELL ET AL.-CARSOSAURUS PECTORAL GIRDLE 531
Axial and Appendicular Skeleton
54 Length ofcervical centra relative to trunkcentra: twice as long (0); centra equal (1);centra subequal or shorter (2).
55 (49P) Number of cervical vertebrae: eight (0);nine (1); seven (2).
56 (53P) Peduncles on cervical and caudal vertebrae: short (0); long (1).
57 (51 P) Number of presacral vertebrae: greaterthan 30 (0); less than 30 (1).
58 Elongate trunk ribs posterior on vertebral column: to trunk vertebrae 14-15(0); to vertebrae 16-25 (1); to trunk vertebrae 7-9 (2).
59 (61P) Number of rib attachments on sternum:more than three pairs (0); three or fewerpairs (1).
60 Posterior iliac process: extends the lengthof two vertebrae (0); extends the lengthof three vertebrae (1); no posterior extension (2).
61 Anterior coracoid fenestra: present (0);absent (1).
62 Posterior coracoid fenestra: absent (0);present (1).
63 Proximal portion of olecranon: largeprocess (0); moderate sized process (1);process absent (2).
64 (59P) Size ofinterclavicle anterior process: long(0); short or absent (1).
65 Intermedium ossification of carpus:present (0); absent (1).
66 Ulna length: approximately % length ofhumerus (0); approximately the samelength (1).
the length of body of palatine (0); lessthan half the length (1).
37 (46R) Length of mesial palatine process articulating with dorsal surface of pterygoid:elongate (0); short (1).
38 (48R) Palatal shelves of vomers: present (0);absent (1).
39 (33P) Palatines size: longer than wide (0); widthroughly equal to length (1).
40 (35P) Pterygoid teeth: present (0); absent (1).
Brain Case
41 (49R) Lateral head on distal end of ectopterygoid: concealed behind jugal (0); exposedlaterally (1).
42 (36P) Ectopterygoid contact with palatine anteriorly: no contact (0); contact (1).
43 (50R) Length and articulation ofproximal headof ectopterygoid: short and lies alongdorsal edge of pterygoid (0); long andcontacts body of pterygoid (1).
44 (52R) Spine on mesial tip of medial pterygoidridge which articulates basipterygoidprocess: absent (0); present (1).
45 (55R) Crista prootica development: well developed (0); weakly developed (1).
46 (57R) Basisphenoid participation in sphenooccipital tubercle: excluded (0); included(1).
47 (59R) Foramina for facial nerve; single (0);double (1).
48 (16P) Hypoglossal foramen size: not enlarged(0); enlarged (1).
Mandible
49 (38N) Coronoid and surangular processes ofdentary: well developed (0); reduced (1).
50 (63R) Suture between angular and splenial: angular, interdigitating (0); vertical andstraight (1).
51 (64R) Medial exposure ofangular compared tolateral exposure: lateral greater (0); medial greater (1).
52 (65R)
53 (68R)
Anterior extension of surangular beyondthe anterior process of coronoid: does(0); does not (1).Splenial contact with the dentary: contact in front of inferior alveolar foramento tip of splenial (0); no contact andMeckels groove open anteriorly (1).
Copyright © 2022 FDOKUMEN




















