
Pectoral fins of rhizodontids and the evolution of pectoral appendages in the tetrapod stem‐group
20
Biological Journal of the Linnean Society (2001), 74: 217–236. With 9 figures doi: 10.1006/bijl.2001.0572, available online at http://www.idealibrary.com on Pectoral fins of rhizodontids and the evolution of pectoral appendages in the tetrapod stem-group JONATHAN E. JEFFERY University Museum of Zoology, Downing Street, Cambridge CB2 3E Received 18 November 2000; accepted for publication 30 April 2001 The Rhizodontida are an extinct order of large, predatory, lobe-finned fishes, found world-wide in Devonian and Carboniferous freshwater deposits. They are thought to be basal members of the tetrapod lineage. In this paper the pectoral fin skeleton of Rhizodus hibberti, a derived member of the group, is described in detail for the first time. It shows that muscular processes of the humerus (the deltoid and supinator processes) may have appeared later in tetrapod evolution than previously thought. The rhizodontids share an unusual fin-ray morphology, with highly elongated basal hemisegments, overlapping extensively with the endoskeleton. This morphology raises the possibility that segmentation and endoskeletal overlap share some common upstream genetic control during lepidotrichial development. The relative size of the lepidotrichia and the complexity of the endoskeleton does not fit with a ‘clockface’ model of limb developmental evolution. 2001 The Linnean Society of London ADDITIONAL KEY WORDS: Rhizodontida – Rhizodus – Sauripterus – lepidotrichia – endoskeleton – fin-bud – embryology – development. (1972; 1985), mostly on the basis of pectoral girdle mor- INTRODUCTION phology. The Rhizodontida (Pisces: Sarcopterygii) is an order of There has been considerable interest in the pectoral freshwater predatory fishes, known world-wide from fin morphology of rhizodontids since Hall (1843) de- the Upper Devonian (e.g. Gooloogongia from the Fras- scribed the similarity of the fin endoskeleton of Sau- nian Mandagery Sandstone, Australia; Johanson & ripterus (Fig. 1F) to that of a tetrapod limb. This Ahlberg, 1998) to the Upper Carboniferous (Strepsodus was the first time that an ‘intermediate’ morphology sauroides from the Westphalian B Newsham Coal- between fins and limbs had been recognized in the shales, UK; e.g. Boyd, 1984). Recent phylogenetic ana- fossil record, and Sauripterus was used as a ‘typical’ lyses (e.g. Cloutier & Ahlberg, 1996; Zhu & Schultze, tetrapod ancestor for many years (e.g. Broom, 1913; 1997; Forey, 1998; Ahlberg & Johanson, 1998) suggest Eastman, 1917; Gregory, 1911, 1915, 1935; Gregory, they are basal members of the tetrapod ‘total-group’ Miner & Noble, 1923; Romer & Byrne, 1931; Westoll, (sensu Patterson, 1993), also known as the Tetra- 1943), even though nothing was known of the rest podomorpha Ahlberg, 1991. A large body size is ac- of its skeleton. However, much of the pectoral fin companied by a suite of unusual adaptations, including endoskeletal material of rhizodontids has been in- a highly elaborated lateralis system (Andrews, 1985; terpreted in the light of one crushed and incomplete Johanson & Ahlberg, 1998). The pectoral fin is highly specimen of Rhizodus (PMG 297; Fig. 4A–D) (Andrews derived, with an extensive endoskeleton and robust le- & Westoll, 1970b) and the extensive material of the pidotrichia (Andrews & Westoll, 1970b; Daeschler & tristichopterid osteolepiform Eusthenopteron (An- Shubin, 1998; Johanson & Ahlberg, 1998). The mon- drews & Westoll, 1970a). I describe here new material ophyly of Order Rhizodontida was originally es- of Rhizodus hibberti from Wardie Beach near Ed- tablished by Andrews & Westoll (1970b) and Andrews inburgh which challenges many details of the original interpretation of PMG 297. PREVIOUSLY DESCRIBED MATERIAL Current Address: Institute for Evolutionary and Ecological Material of the dermal pectoral girdle is well known Sciences, University of Leiden, Postbus 9516, 2300 RA Leiden, The Netherlands. E-mail: [email protected] for many rhizodontid species. In contrast material of 217 0024–4066/01/100217+20 $35.00/0 2001 The Linnean Society of London
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Pectoral fins of rhizodontids and the evolution of pectoral appendages in the tetrapod stem‐group

Biological Journal of the Linnean Society (2001), 74: 217–236. With 9 figures
doi: 10.1006/bijl.2001.0572, available online at http://www.idealibrary.com on
Pectoral fins of rhizodontids and the evolution ofpectoral appendages in the tetrapod stem-group
JONATHAN E. JEFFERY
University Museum of Zoology, Downing Street, Cambridge CB2 3E
Received 18 November 2000; accepted for publication 30 April 2001
The Rhizodontida are an extinct order of large, predatory, lobe-finned fishes, found world-wide in Devonian andCarboniferous freshwater deposits. They are thought to be basal members of the tetrapod lineage. In this paperthe pectoral fin skeleton of Rhizodus hibberti, a derived member of the group, is described in detail for the firsttime. It shows that muscular processes of the humerus (the deltoid and supinator processes) may have appearedlater in tetrapod evolution than previously thought. The rhizodontids share an unusual fin-ray morphology, withhighly elongated basal hemisegments, overlapping extensively with the endoskeleton. This morphology raises thepossibility that segmentation and endoskeletal overlap share some common upstream genetic control duringlepidotrichial development. The relative size of the lepidotrichia and the complexity of the endoskeleton does notfit with a ‘clockface’ model of limb developmental evolution. 2001 The Linnean Society of London
ADDITIONAL KEY WORDS: Rhizodontida – Rhizodus – Sauripterus – lepidotrichia – endoskeleton – fin-bud –embryology – development.
(1972; 1985), mostly on the basis of pectoral girdle mor-INTRODUCTIONphology.
The Rhizodontida (Pisces: Sarcopterygii) is an order of There has been considerable interest in the pectoralfreshwater predatory fishes, known world-wide from fin morphology of rhizodontids since Hall (1843) de-the Upper Devonian (e.g. Gooloogongia from the Fras- scribed the similarity of the fin endoskeleton of Sau-nian Mandagery Sandstone, Australia; Johanson & ripterus (Fig. 1F) to that of a tetrapod limb. ThisAhlberg, 1998) to the Upper Carboniferous (Strepsodus was the first time that an ‘intermediate’ morphologysauroides from the Westphalian B Newsham Coal- between fins and limbs had been recognized in theshales, UK; e.g. Boyd, 1984). Recent phylogenetic ana- fossil record, and Sauripterus was used as a ‘typical’lyses (e.g. Cloutier & Ahlberg, 1996; Zhu & Schultze, tetrapod ancestor for many years (e.g. Broom, 1913;1997; Forey, 1998; Ahlberg & Johanson, 1998) suggest Eastman, 1917; Gregory, 1911, 1915, 1935; Gregory,they are basal members of the tetrapod ‘total-group’ Miner & Noble, 1923; Romer & Byrne, 1931; Westoll,(sensu Patterson, 1993), also known as the Tetra- 1943), even though nothing was known of the restpodomorpha Ahlberg, 1991. A large body size is ac- of its skeleton. However, much of the pectoral fincompanied by a suite of unusual adaptations, including endoskeletal material of rhizodontids has been in-a highly elaborated lateralis system (Andrews, 1985; terpreted in the light of one crushed and incompleteJohanson & Ahlberg, 1998). The pectoral fin is highly specimen of Rhizodus (PMG 297; Fig. 4A–D) (Andrewsderived, with an extensive endoskeleton and robust le- & Westoll, 1970b) and the extensive material of thepidotrichia (Andrews & Westoll, 1970b; Daeschler & tristichopterid osteolepiform Eusthenopteron (An-Shubin, 1998; Johanson & Ahlberg, 1998). The mon- drews & Westoll, 1970a). I describe here new materialophyly of Order Rhizodontida was originally es- of Rhizodus hibberti from Wardie Beach near Ed-tablished by Andrews & Westoll (1970b) and Andrews inburgh which challenges many details of the original
interpretation of PMG 297.
PREVIOUSLY DESCRIBED MATERIALCurrent Address: Institute for Evolutionary and Ecological
Material of the dermal pectoral girdle is well knownSciences, University of Leiden, Postbus 9516, 2300 RA Leiden,The Netherlands. E-mail: [email protected] for many rhizodontid species. In contrast material of
2170024–4066/01/100217+20 $35.00/0 2001 The Linnean Society of London
218 J. E. JEFFERY
50 mm
ACD E B
over. boneent.
rad. uln. cond.
cap. hum.dors. ridgeclav.
E
Dent.
cap. hum. rad.
uln.ulne.
elong.
C ent.
uln.
rad.
ulne.
elong.
B
A
scales
dist.lepid.seg.
elong.scap. cor.
clav.
operc.
scales
scap. cor. elong.
cleith.
50 mmFG
G
F
rad.
uln.ulne.
elong.
H
dist.lepid.seg.
elong.
H
uln.
ent.
cap. hum.
cleith.elong.
rad.?ent.
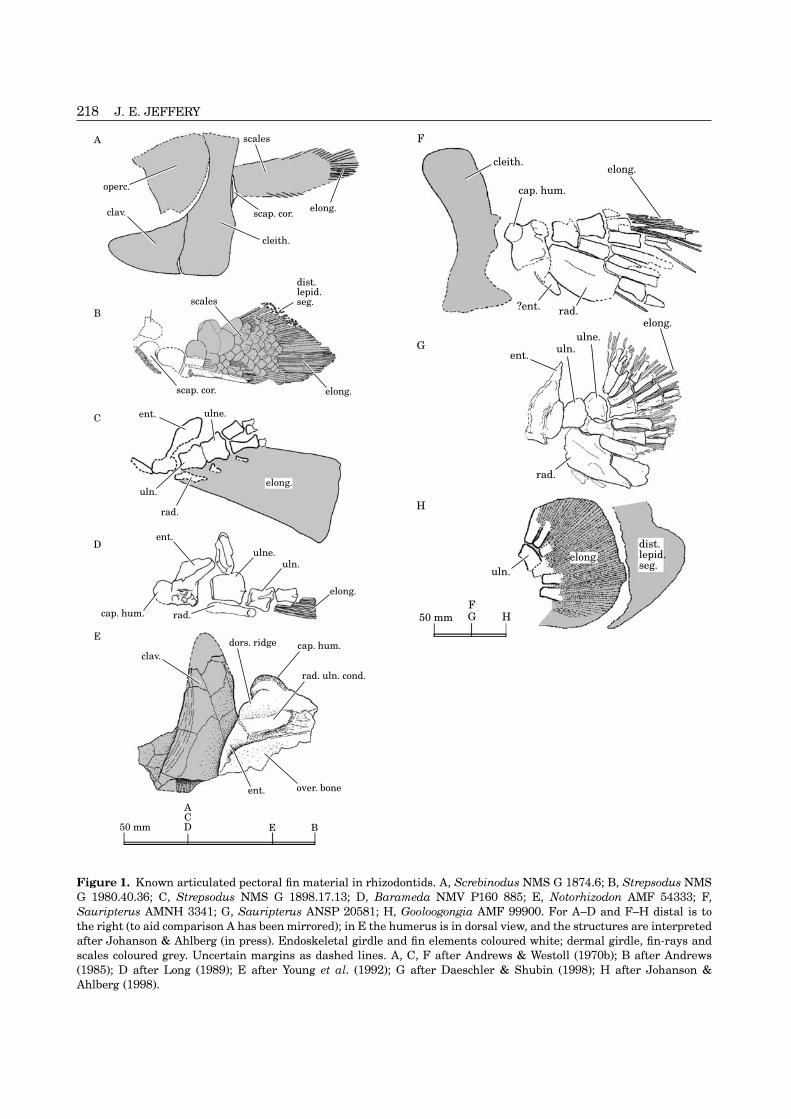
Figure 1. Known articulated pectoral fin material in rhizodontids. A, Screbinodus NMS G 1874.6; B, Strepsodus NMSG 1980.40.36; C, Strepsodus NMS G 1898.17.13; D, Barameda NMV P160 885; E, Notorhizodon AMF 54333; F,Sauripterus AMNH 3341; G, Sauripterus ANSP 20581; H, Gooloogongia AMF 99900. For A–D and F–H distal is tothe right (to aid comparison A has been mirrored); in E the humerus is in dorsal view, and the structures are interpretedafter Johanson & Ahlberg (in press). Endoskeletal girdle and fin elements coloured white; dermal girdle, fin-rays andscales coloured grey. Uncertain margins as dashed lines. A, C, F after Andrews & Westoll (1970b); B after Andrews(1985); D after Long (1989); E after Young et al. (1992); G after Daeschler & Shubin (1998); H after Johanson &Ahlberg (1998).

PECTORAL FINS OF RHIZODONTIDS 219
?foss. musc. fossa possibly for musclethe pectoral endoskeleton, including the scapulo-coracoid of the girdle, is rare. However, the general attachment
glen. buttr. glenoid buttressform of the pectoral fin has been known for sometime, from a small number of articulated specimens glen. for. glenoid foramen
glen. foss. glenoid fossabelonging to several species (see Fig. 1 and Table 1).Also, isolated endoskeletal elements are known for hum. humerus
hum. waist ‘waist’ of humerusStrepsodus (Andrews & Westoll, 1970b). For Rhizodusthe only previously known endoskeletal material was infra. buttr. infraglenoid buttressan articulated dermal pectoral girdle and sca- intermed. intermediumpulocoracoid (NMS G 1963.16.16) from Niddire, Ed- lepid. lepidotrichial segmentsinburgh (Namurian & Westphalian A) and an lepid. branch. branching point ofincomplete scapulocoracoid (PMG 298) and humerus lepidotrichium(PMG 297; Fig. 4A–D) from Todhill’s Pit, Dalry, Ayr- ldp. latissimus dorsi ridgeshire (?Namurian). These were described by Andrews mes. f.f. mesenchyme in fin-fold& Westoll (1970b). oblq. vent. ridge oblique ventral ridge
operc. opercleov. clav. overlap area for clavicleABBREVIATIONS USED IN FIGURESover. bone overlying bone fragmentactin. remnants of actinotrichiapect. proc. pectoralis processAER apical ectodermal ridgepect. proc. rug. rugosity on pectoralis processant. marg. rad. anterior margin of radiuspostbranch. lam. postbranchial laminaar. m. sup. supinator muscular areapr. delt. deltoid processar. sch. scapulo-humeral muscular areapr. H. process of humerusbasal campt. seg. basal camptotrichial segmentspr. sup. supinator processbasal lepid. basal lepidotrichialprox. artic. rad. proximal articular surface ofhemiseg. hemisegments
radiuscampt. camptotrichiumprox. artic. uln. proximal articular surface ofcampt. branch. branching point of
ulnacamptotrichiumprox. artic. ulne. proximal articular surface ofcap. hum. caput humeri
ulnareclav. claviclePZ progress zonecleith. cleithrumrad. radiusdep. post. flange depressed posterior flange ofrad. cond. radial condylecleithrumrad. uln. cond. radial and ulnar condylesdist. artic. uln. distal articular surfaces of ulnascap. cor. scapulocoracoiddistal campt. distal camptotrichialscales area of articulated scaleshemiseg. hemisegments;supra. buttr. supraglenoid buttressdist. endoskel. distal endoskeletal elementsuln. ulnadistal lepid. distal lepidotrichialuln. cond. ulnar condylehemiseg. hemisegmentsulne. ulnaredist. lepid. seg. distal lepidotrichial segmentsvent. lam. cleith. incomplete ventral lamina ofdist. marg. ent. distal margin of entepicondyle
cleithrumdors. lam. clieth. dorsal lamina of cleithrumdors. ridge dorsal ridge of humerusec. c. ectepicondyle
ABBREVIATIONS OF MUSEUM COLLECTIONSelong. elongate basal lepidotrichialANMH American Museum of Natural His-segments
tory, New Yorkendoskel. endoskeletal fin supportsANSP Academy of Natural Sciences, Phil-?ent. possible entepicondyle
adelphiaent. entepicondyleAMF Australian Museum, Sydneyent. foramina foramina on entepicondyleBGS British Geological Survey, Keyworthfor. ‘ect.’ ‘ectepicondylar’ foramenGLAHM Hunterian Museum, Glasgowfor. ent. entepicondylar foramenNEWHM Hancock Museum, Newcastle uponfor. vent. foramen on oblique ventral
Tyneridgefoss. ‘ect.’ fossa leading from MNHN Museum National d’Histoire Nat-
urelle, Paris‘ectepicondylar’ foramen

220 J. E. JEFFERY
NMS G National Museums of Scotland, Ed- only to the material identified as rhizodontid by Johan-son & Ahlberg (in press).inburgh
NMV National Museum of Victoria, Mel- For ease of comparison, the terminology used belowto describe the morphology of the humerus broadlybourne
PMG Paisley Museum Geological Col- follows that of Andrews & Westoll (1970a,b) and Jarvik(1980). However, Coates (1996) noted the uncertaintieslection
SME Sedgwick Museum of Geology, Cam- in interpreting the musculature of primitive limb (orfin) skeletons, and two particular structures havebridge
UCLGMZ Grant Museum of Zoology and Com- equally common alternative names; the ‘deltopectoralcrest’ (=Andrews & Westoll’s pectoralis process) andparative Anatomy, UCL, London.the ‘latissimus dorsi process’ (=Andrews & Westoll’sdeltoid process).
MATERIAL AND METHODS
COLLECTION AND PALAEONTOLOGYPALAEOENVIRONMENT
The Rhizodus fin skeleton described below (NMS G Wardie was a subtropical freshwater environment, a1972.27.434c) was collected from Wardie Beach, Ed- relatively deep part of the permanent Lake Cadell,inburgh (OS Landranger Grid NT245771). The site is which covered a large part of the Midlands of Scotlandapproximately 340 Ma (Holkerian of Visean, Lower during the Carboniferous (see Whyte, 1994). The wholeCarboniferous; (Chisholm & Castanet, 1994)) and con- area was part of a low-lying coastal alluvial plain, andtains a number of fossil-bearing horizons, exposed in subject to occasional influx of brackish or salt water.different areas along the beach. The fish fossils are Rhizodontid remains are usually restricted to bedswell preserved in ironstone nodules within shales formed in this type of palaeoenvironment.(Wood, 1975, 1992). The fin skeleton was originallycollected by Mr S. P. Wood, probably from Fish Bed 2,and is now part of the geological collections of the DESCRIPTIONNational Museum of Scotland.
GROSS MORPHOLOGYThe Strepsodus cleithrum and scapulocoracoid de-scribed below (BGS GSL 1527) is part of the British The external morphology of the fin is only well-pre-Geological Survey collection. It is preserved in coal- served in Strepsodus NMS G 1980.40.36 (Fig. 1B) andshales, but is of uncertain provenance. Screbinodus NMS G 1874.6 (Fig. 1A). In these species
Both specimens were partially prepared before the the fin is covered in deeply imbricated cycloid scalescurrent study; further preparation of NMS G and with a short, exposed, distal fringe of lepidotrichia1972.27.434c was carried out using a Chicago Pneu- (dist. lepid. seg. Fig. 1B). The fin of both species ismatic Airscribe CP9361 under a Meiji EMX stereo- rhomboid in shape. The endoskeleton and lepidotrichiamicroscope, which revealed fine detail without the are known in articulation in Sauripterus (Hall, 1843;need for a mounted needle. The specimen drawings in Daeschler & Shubin, 1998; Fig. 1F,G) and Gooloogon-Figures 2, 3 and 5 were made from the original material gia (Johanson & Ahlberg, 1998; Fig. 1H). In theseusing a camera lucida. The specimen drawings in species the fin has a broader, fan-shaped profile.Figures 1 and 6 were traced from published photo-graphs (source publications are listed in the figure
FIN ENDOSKELETONlegends). Figure 4E–D was redrawn from VorobyevaNMS G 1972.27.434c (Fig. 2) consists of an incomplete(2000), with modifications based on the original speci-right pectoral fin scattered across a large (35 cm long)men. Figure 4A–D is reproduced with the permissionnodule. Although disarticulated, the fin parts retainof the Royal Society of Edinburgh from Andrews &some of their proximodistal arrangement. SeveralWestoll (1970b). Figure 8A,B,D–F is reproduced frompoorly preserved elongate basal lepidotrichial hem-Goodrich (1904) with the permission of the Companyisegments (lepid.) are scattered over the specimen.of Biologists Ltd. Figure 8C is reproduced from Jarvik
(1959) with the permission of the Royal Swedish Acad-emy of Sciences.
HumerusJohanson & Ahlberg (in press) have redescribed thematerial attributed to Notorhizodon by Young, Long The humerus (Fig. 3) has a proximodistal length of
about 9 cm (approximately the same size as PMG 297;& Ritchie (1992). They conclude that only part of thetype series (including the pectoral fin material) belongs Fig. 4A–D). The caput humeri (cap. hum.) is semi-
spherical and lacks a periosteal surface. It is separatedto a rhizodontid. To prevent complications of no-menclatural priority, I will continue to use the name from the rest of the humerus by a distinct ‘waist’ (hum.
waist). The radial condyle (rad. cond.) is a convexNotorhizodon for this paper, but my comments refer

PECTORAL FINS OF RHIZODONTIDS 221
Table 1. Previously described articulated pectoral fin material. References given are for the original descriptions only
Genus Articulated fin material Formation and age
Barameda Long, 1989 NMV P160 885 (Long, 1989) Mansfield Formation, Victoria,Australia (?Visean)
Gooloogongia Johanson & Ahlberg, AMF 99900 (Johanson & Ahlberg, 1998) Mandagery Sandstone, Canowindra,1998 NSW, Australia (Frasnian)
Notorhizodon Young et al., 1992 AMF 54333 (Young et al., 1992) Aztec Sandstones, Southern VictoriaLand, Antarctica (?Givetian or earlyFrasnian)
Rhizodus Owen, 1840 NMS G 1972.27.434c (see main text) Wardie Shales (Visean)
Sauripterus Hall, 1843 i. AMNH 3341 (Hall, 1843) Catskill formation, Pennsylvania, USAii. ANSP 20581 (Daeschler & Shubin, (Famennian)1998)
Screbinodus Andrews, 1985 NMS G 1874.6 (Traquair, 1875) Gilmerton Ironstone, Edinburgh(Visean)
Strepsodus Huxley in Huxley & i. NMS G 1980.40.36 (Andrews, 1985) i. Foulden, Berwickshire (Tournaisian)Etheridge, 1865 ii. NMS G 1898.17.13 (Andrews & ii. Location unknown (?Westphalian)
Westoll, 1970b)
rad.40 mm
lepid.dist. endoskel.
lepid.
dist. artic. uln.
ant. marg. rad.prox. artic. rad.
pect. proc. rug.
cap. hum.
hum.
for. ent.
for. vent.ent.
intermed. dist. endoskel. lepid.dist. endoskel.
prox. artic. ulne.
prox. artic. uln.
Figure 2. Rhizodus hibberti. NMS G 1972.27.434c. Right pectoral fin skeleton. Dashed lines=cracks, broken oruncertain margins; diagonal hatching=obscured bone; grey=matrix.
region of unfinished bone, running into a similarly least 1 cm thick. It is pierced by a number of smallforamina. A canal runs proximodistally through thesized and shaped ulnar condyle (uln. cond.).
The robust entepicondyle (ent.) is obliquely offset base of the entepicondyle, producing large foramina(for. ent.) on its anterodistal and posteroproximalfrom the plane of the ulnar and radial condyles by
about 30°. Its margins are all unfinished, but are at faces. This canal is probably homologous with the

222 J. E. JEFFERY
for. ent.
A
cap. hum.
pect. proc.rug.
part of radius intermed.
rad. cond.
oblq. vent. ridgefor. vent.
dist. marg. ent.
for. ent.
hum. waist
20 mm
cap. hum.
for. 'ect'.
hum. waist.
pect. proc.
part of radiusintermed.
foss. 'ect'.
rad. cond.
uln. cond.
oblq. vent. ridge
ent.
B
dors. ridge
Figure 3. Rhizodus hibberti. NMS G 1972.27.434c. A, right humerus in ventral view. B, right humerus in dorsal view.Dashed lines=cracks, broken or uncertain margins; diagonal hatching=obscured bone; grey=matrix.
entepicondylar canal of Eusthenopteron. Jarvik (1980: back onto the dorsal surface to form a proximal dorsalridge (dors. ridge) running almost parallel with the141) suggested this carried the ulnar nerve.
The entepicondyle is not a discrete structure, but ulnar and radial condyles. The oblique ventral ridgeand the pectoral process were identified on PMG 297forms part of an extended helical ridge, running prox-
imally onto the ventral surface of the humerus to form by Andrews & Westoll (1970b) (see Fig. 4A–D), al-though the continuity with the entepicondyle was notthe oblique ventral ridge (oblq. vent. ridge), then across
the anterior edge of the bone, where it swells to form recognized. There is a large foramen (for. vent.) onthe oblique ventral ridge, close to its junction with thethe pectoralis process (pect. proc.) and then finally

PECTORAL FINS OF RHIZODONTIDS 223
for. ent.
A
cap. hum.
pect. proc.
rad. cond.
oblq. vent. ridge(hu. p. & ar. ld.)
for. vent.
for. ent.
uln. cond.
oblq. vent. ridge
ent.
dors. ridge(includes delt.)
part ent.
dors.ridge
(pr. delt.)
(pr. sup.)
(ec. c.)
ent
cap. hum.
rad. cond.
for.vent.
oblq. vent. ridge
for. ent.
ent.
uln. cond.
rad. cond.
part of ent. (pr. H.)
cap. hum.
B
C D
for. vent.(for. ent.)
ent.
20 mm A–D E–H
dors. ridge
for. ent.ent.
rad. cond.
(ar. m. sup.)
?ec. c.
E F G
H
uln. cond.(includes ?ar. u.)
cap. hum.
(ar. sch.)(ldp.)
?ec. c.
pect. proc.(includes hu. ri.)
Figure 4. Previous interpretations of rhizodontid humeri. A–D, Rhizodus hibberti. Isolated right humerus PMG 297(A, in ventral view; B, in dorsal view; C, in proximal view; D, in distal view). E–H, Strepsodus sauroides. Isolated righthumerus, part of NMS 1970.16.1 (E, in dorsal view (‘postaxial’); F, in anterior view (‘ventro-preaxial’); G, in posteriorview (‘dorsal’); H, in distal view). A–D reproduced from Andrews & Westoll (1970b: Plate IXe–h). E–H after Vorobyeva(2000: Fig. 2). Where they differ from the current study, the interpretations of the original authors are given inparentheses (NB the incorrect orientations given by Vorobyeva, 2000).
entepicondyle. This is probably homologous with thought to correspond respectively to the deltoidprocess, the supinator process and the ectepicondylethe foramina observed on the ventral ridge of
Eusthenopteron (Andrews & Westoll, 1970a). of basal tetrapods and Eusthenopteron (e.g. Andrews& Westoll, 1970a: text-fig.10C; Jarvik, 1980: fig. 104B).The tip of the pectoralis process (pect. prog. rug.)
and the peak of the dorsal ridge are of unfinished bone, On NMS G 1972.27.434c an ectepicondylar region isdiscernible at the posterior end of the ridge, separatedpresumably where connective tissues attached. The
dorsal ridge is poorly preserved in PMG 297, but from the rest of the ridge by a narrow strip of bone,apparently pierced by a small foramen (for. ‘ect.’). AAndrews & Westoll (1970b) identified three divisions
(pr. delt., pr. sup. and ec. c.; Fig. 4B). These were shallow fossa (foss. ‘ect.’) runs distally from this

224 J. E. JEFFERY
foramen to the meeting point of the ulnar and radial ectepicondyle. It bears a similar relationship to theulnar condyle as the ectepicondyle of Eusthenopteroncondyles (cf. Eusthenopteron (Andrews & Westoll,
1970a: text-fig. 10A)). However, there are no clearly (e.g. Andrews & Westoll 1970a: text-fig 10a). However,the ectepicondyle of Eusthenopteron is also connecteddifferentiated deltoid or supinator processes. There is
also no process separating the ulnar condyle from the to the supinator process via a bridge of bone, formingthe ectepicondylar foramen (cf. a similar connectionbase of the entepicondyle as there is in Eusthenopteron
(Andrews & Westoll, 1970a). The structure identified in Rhizodus, above). The ‘ectepicondyle’ of NMS G1970.16.1 is widely separated from the dorsal ridgeas such on PMG 297 by Andrews & Westoll (1970b) is
in fact a fragment of the dorsal face of the entepicondyle (contra Vorobyeva 2000), which means that its iden-tification must remain tentative. It may prove to be(part ent. and pr. H.; see Fig. 4A,C).
The entepicondyle of Strepsodus (Figs 1C, 4D, E) is convergent with the ectepicondyle of Eusthenopteronand basal tetrapods.relatively longer and more gracile than that of Rhi-
zodus. Vorobyeva (2000) figured and described a right Finally, Vorobyeva (2000) identified articular areasfor the latissimus dorsi muscle. One of these (ar.ld.;humerus from Newsham, Northumberland (part of
NMS G 1970.16.1). Although her description is largely Fig. 4G) is simply part of the oblique ventral ridge.The other two (ldp. and the associated ar. sch.) areaccurate, comparisons with the better-preserved Rhi-
zodus NMS G 1972.27.434c shows that some of her probably due to damage to the caput humeri. There areno similar structures in Rhizodus (contra Vorobyeva,interpretations are incorrect. A revised interpretation
is given in Figure 4E, F. 2000).The humerus of Screbinodus (pers. obs. of SME 4714)The entepicondyle of NMS 1970.16.1 has been
slightly crushed. This has distorted the ulnar condyle appears to have a long entepicondyle, similar in shapeto that of Strepsodus. However, little more can be said(uln. cond.) into a bi-lobed shape, which Vorobyeva
(2000) interpreted as two separate structures; the ul- about the morphology of the humerus in this species.The humerus of Barameda is known from a singlenar condyle, plus a distal part of the ectepicondyle.
The entepicondylar foramen has been correctly iden- endocast (Fig. 1D). The structures identified by Long(1989) as the deltoid and supinator processes may betified on the posteroproximal face of the entepicondyle.
However, the two ‘entepicondylar’ foramina identified part of a continuous dorsal ridge, preserved in a similarcondition to Rhizodus PMG 297.by Vorobyeva (2000) in anterior view actually pierce
the oblique ventral ridge, not the entepicondyle. As The humerus of Sauripterus is damaged on AMNH3341 (Fig. 1F), and its orientation is uncertain. Al-with Rhizodus, they are likely to be homologous with
the foramina observed on the ventral ridge of Eus- though the rest of the fin endoskeleton is undisturbed,the humerus has an unusual, large process directedthenopteron (Andrews & Westoll, 1970a). A more prob-
able entepicondylar foramen (for. ent.; Fig. 4G,H) is anteriorly (?ent.) as opposed to a posteriorly directedentepicondyle. However, a recently discovered speci-by the side of the ulnar condyle, closely aligned to the
foramen on the posteroproximal face, and in a similar men (ANSP 20581; Daeschler & Shubin, 1998) (Fig.1G), whilst very similar to AMNH 3341, shows a largeposition to that of Rhizodus.
As with Rhizodus, the entepicondyle connects to an entepicondyle but no such anterior expansion. Thus itis possible that the humerus of ANMH 3341 has beenoblique ventral ridge (oblq. vent. ridge; Fig. 4F,G)
which swells anteriorly to form pectoralis process (pect. rotated by 180° during fossilization, or that weatheringand/or preparation have damaged it in a deceptiveproc; Fig. 4F). Although slightly damaged, the pec-
toralis process seems almost complete. However, way. The ectepicondyle of ANSP 20581 does not appearto be a large, offset blade as in Rhizodus, nor is aStrepsodus NEWHM G5.05, also from Newsham,
shows a very much larger process in a similar position ventral ridge apparent. Further description of thisspecimen (and new material) is in progress (Davis,(Andrews & Westoll, 1970b: plate XIIn ‘ri.H.’). This
may represent individual variation. The pectoralis pro- Shubin & Daeschler, in press).The humerus is the only part of the fin endoskeletoncess is itself connected to the dorsal ridge (dors. ridge;
Fig. 4E,F,H) by a shallow ridge of unfinished bone. known for Notorhizodon (Fig. 1E). Young et al. (1992)identified an ectepicondyle, entepicondyle, deltoid pro-The dorsal ridge is not pierced by an ectepicondylar
foramen. It has been slightly crushed, creating (or cess and the articulation surface for the latissimusdorsi muscle. However, Johanson & Ahlberg (in press)exaggerating) a hollow between its distal face and
the radial condyle. Vorobyeva (2000) interpreted this have reinterpreted the humerus after further pre-paration. They suggest that the ridge Young et al.hollow as a region for the attachment of supinator
musculature (ar. m. sup; Fig. 4B). (1992) identified as an articulation surface for thelatissimus dorsi muscle comprises an ectepicondyleThere is a ridge of bone running along the dor-
soposterior edge of the shaft of the humerus (?ec. and indistinct deltoid and supinator regions, similarto the dorsal ridge of Rhizodus.c.; Fig. 4E,G). Vorobyeva (2000) identified this as an

PECTORAL FINS OF RHIZODONTIDS 225
Radius DERMAL FIN RAYS
The radius (rad.) of Rhizodus is preserved in ventral None of the lepidotrichia on NMS G 1972.27.434cview (Fig. 2). The anterior margin is largely convex. (lepid.; Fig. 2) is preserved in natural articulation withThe distal margin is overlain by the ulna, but what is the fin endoskeleton. However, the scattered segmentsvisible appears quite thick, and bears articular sur- strongly resemble those found in articulation in Streps-faces for at least one more distal element. odus, Screbinodus (Andrews, 1973, 1985; Andrews &
The radius of Strepsodus (Fig. 1B,C) is similar to Westoll, 1970b), Sauripterus (Andrews & Westoll,that of Rhizodus, although more elongate. It may also 1970b; Daeschler & Shubin, 1998), Baramedahave articulated with more distal elements (pers. obs. (Long, 1989) and Gooloogongia (Johanson & Ahlberg,of isolated radius NEWHM G21.28). In Sauripterus 1998). In these species, most of the length of each
lepidotrichium is formed by elongated basal hemi-(Fig. 1F,G) the radius is extremely broad, and ar-segments (elong.; Figs 1A–G, 7H). These hemi-ticulates with two distal elements. In one of the speci-segments are round in cross-section and do not branch.mens (ANSP 20581) there is a another, more distalThere is no clear evidence in any species that the basalelement (Daeschler & Shubin, 1998). Long (1989) re-lepidotrichial hemisegments were closely aligned inconstructed the partially preserved radius of Ba-dorsal and ventral pairs. The overlap of the basalrameda (Fig. 1D) as similar to that of Strepsodus.hemisegments with the endoskeleton is extensive, run-ning over several elements distal to the ulna and radius(e.g. Fig. 1B,C). The much smaller distal hemisegmentsUlna and ulnare(dist. lepid. seg.; Fig. 1B,H) are very seldom reserved.The ulna of Rhizodus (Fig. 2) is a broad, robust bone,In Gooloogongia (Fig. 1H), and probably also in Sau-which produces two distal articular surfaces, for theripterus, the most proximal segment comprises aboutulnare and intermedium (dist. artic. uln.). Although ithalf the total length of the lepidotrichia (Johanson &is still partly buried in the matrix, it appears to haveAhlberg, 1998). In Strepsodus and Screbinodus (An-a wide concave dish as its proximal articular surfacedrews, 1985; Fig. 1A–C), and probably Barameda(prox. artic. uln.) to fit the condyle on the humerus.(Long, 1989; Fig. 1D) the basal segments comprise theThe ulnare is narrower proximally than the ulna andmajority of the total lepidotrichial length. The length ofhas a fairly flat proximal articular surface (prox. artic.the incomplete basal segments on NMS G 1972.27.434culne.). It flares out distally to produce at least twosuggest that Rhizodus was similar to Strepsodus and
(possibly three) articular surfaces. The ulnare has noScrebinodus.
posterior process (cf. the ‘pisiforme’ of Eusthenopteron;e.g. Jarvik, 1980).
The ulnare of Strepsodus (ulne; Fig. 1C) probablyPECTORAL GIRDLEarticulates with three elements distally. In Sauripterus
(Fig. 1F,G) it articulates with three or four elements Endoskeletal girdledistally (Daeschler & Shubin, 1998). Strepsodus, Sau- Andrews & Westoll (1970b) described two sca-ripterus and Barameda also lack a posterior ‘pisiforme’ pulocoracoids belonging to Rhizodus (NMS Gprocess. In Rhizodus, Strepsodus and Sauripterus the 1963.16.16 & PMG 298). It is a tribasal bone, with aulna and ulnare are of approximately equal size; in deep, circular glenoid fossa. The internal face of theBarameda (Fig. 1D) the ulna appears to be much infraglenoid buttress is pierced by a canal whichbroader than the ulnare. emerges on the “surface between the glenoid and the
cleithrum” (Andrews & Westoll, 1970b: 442). There isalso a more superficial foramen, which Andrews &
Other distal bones Westoll (1970b) identified as the glenoid foramen.In Rhizodus the intermedium is a more slender bone Strepsodus BGS GSL 1527 (Fig. 5) is a previouslythan the ulna and ulnare, with a distinct ‘waist’ (in- undescribed dorsal part of a cleithrum with an attachedtermed.; Figs 2, 3). It expands distally into two articular scapulocoracoid. The scapulocoracoid is very similarsurfaces. There are at least four more distal bones to that of Rhizodus. There is a foramen (glen. for.)preserved on NMS G 1972.27.434c (dist. endoskel.; piercing the internal face of the infraglenoid buttressFig. 2) but most are badly damaged and not closely (infra. buttr.), as in Rhizodus (Fig. 6). The canal leadingassociated with any particular proximal element. Sau- from it bifurcates, emerging at two foramina on theripterus (Fig. 1G) shows that the distal elements have lateral face of the infraglenoid buttress. This is verymuch less distinct individual morphologies; they are similar to the glenoid foramen of Eusthenopteron (e.g.generally simple elongate rods, the more proximal Andrews & Westoll, 1970a: Fig. 4), and is the mostones themselves articulating with one or more distal likely identity of the canal on Strepsodus, and of the
less superficial of the two canals in Rhizodus (contraelements.

226 J. E. JEFFERY
A
40 mm
B C D
E F G Hdors. lam.cleith.
dep. post.flangesupra. buttr.
scap. cor. glen. for.
ov. clav.
infra. buttr.
vent. lam.cleith.
postbranch. lam.vent. lam.
cleith.
?foss. musc.
infra. buttr.
glen. for.
glen. foss.
dors. lam. cleith.
glen. buttr.
vent. lam.cleith.
dors. lam. cleith.
Figure 5. Strepsodus sauroides. BGS GSL 1527. A–D, Right cleithrum and scapulocoracoid (A, In anterior view; B,In mesial view; C, In posterior view; D, In lateral view). E,F, Interpretative sketches of BGS GSL 1527. Dermalcleithrum coloured grey; endoskeletal scapulocoracoid coloured white.
Andrews & Westoll, 1970b). In the place of the more to have been occluded by post-mortem compression.superficial foramen on Rhizodus (identified as the glen- Several isolated ?Strepsodus scapulocoracoids fromoid foramen by Andrews & Westoll) is a deep, peri- Newsham are in a similar condition. This suggestsosteally lined fossa, possibly for muscle attachment that much of the internal structure of the sca-(?foss. musc.). The glenoid buttress of the sca- pulocoracoid was cartilaginous, leaving only a shell ofpulocoracoid wraps around the thickened ventral parts perichondral bone after death.of the depressed posterior flange (dep. post. flange) ofthe cleithrum.
The scapulocoracoid of Screbinodus ornatus is visibleDermal girdlein situ on the holotype specimen NMS G 1874.6 (scap.The dermal girdles of Rhizodus, Strepsodus and Scre-cor.; Fig. 1A). It is mostly obscured by overlying scales,binodus were described in detail by Andrews & Westollbut what is visible appears to be similar to that of(1970b). They noted a number of synapomorphies (e.g.Strepsodus and Rhizodus. As in Strepsodus, the glenoidthe ‘reverse overlap’ of the cleithrum and clavicle, abuttress is wrapped around the ventral part of thedepressed posterior flange on the cleithrum, a semi-depressed posterior flange. Nothing is known of thecylindrical ascending spine of the clavicle). In thescapulocoracoid in other rhizodontids.cleithrum of Rhizodus the postbranchial lamina meetsIn Rhizodus and Strepsodus the scapulocoracoidsthe depressed posterior flange dorsally (Andrews &appear crushed or ‘deflated’. The supraglenoid, sub-Westoll, 1970b). It does not do so in Strepsodus, Scre-scapular and supracoracoid foramina are not as pro-
nounced as in many other sarcopterygians, and appear binodus (Andrews, 1985) or Barameda (Long, 1989).

PECTORAL FINS OF RHIZODONTIDS 227
dist.lepid.seg.
scales
ulne. uln. hum.scap. cor.
cleith.
clav.
rad.intermed.
dist. endoskel.elong.
Figure 6. Rhizodus hibberti. Reconstruction of right pectoral fin and girdle. Endoskeleton based on NMS G 1972.27.434c;scales, distal lepidotrichial web, and overall shape of fin based on Strepsodus NMS G 1980.40.36 (Andrews, 1985);pectoral girdle based on reconstruction in Andrews & Westoll (1970b: text-fig. 10). Dermal bone and scale ornamentomitted for clarity.
Gooloogongia lacks a depressed posterior flange al- a large number of lateralis pores scattered across thetogether (Johanson & Ahlberg, 1998) and the flange dorsal-most part of the dorsal lamina of the cleithrum,is poorly developed in Notorhizodon (Johanson & nestled amongst the fine dermal ornament.Ahlberg, in press). Strepsodus (NEWHM G19.41) has 5–6 parallel lines
Interclavicles are not known in any rhizodontid spe- of closely spaced lateralis pores running ante-cies. I question the interpretation of Andrews & Westoll roventrally on the dorsal part of the dorsal lamina of(1970b: 433) that depressions on some clavicles are the cleithrum. At either end of the lines the poresarticulation surfaces for interclavicles. These de- merge into the parallel grooves of the dermal ornament.pressions are irregular and are present on only a few Janvier et al. (1984) described two cleithra from thespecimens (for example Screbinodus NMS G Upper Devonian (‘TN1a’) of Turkey as “Strepsodus sp.”1890.104.8 shows a depression whilst the similarly (MNHN ZAP 1 & 2). These do not show any definitesized NMS G 1964.28.10 does not). Where present, sensory structures. However, their morphology differsthe depression could equally well be interpreted as from that of other Strepsodus material in several re-taphonomic distortion of the specimen. spects (e.g. extremely narrow dorsal lamina), and they
Andrews & Westoll (1970b) showed that the dermal may represent a new species.ornament on the cleithrum is autapomorphic in Rhi- Screbinodus ornatus (specimens NMS G 1874.6 andzodus and in Screbinodus. The cleithral ornament of NMS G 1964.28.1) have no lateralis pores, but possessStrepsodus is also distinctive (pers. obs. of NEWHM vertically oriented pit-lines on the dorsal lamina of theG19.41, NEWHM G18.70, GLAHM V8127 and BGS cleithrum, just posterior to the postbranchial lamina,GSL 1527). However the ornament is only distinctive at the level of the attachment of the scapulocoracoid.on the dorsal lamina of the cleithrum; on the ventrallamina the ornament of all species becomes a similarnetwork of ridges and craters. This latter ornament is DISCUSSIONalso seen on the clavicles of all three species (e.g.
EVOLUTION AND PHYLOGENYRhizodus NMS G 1876.8; Screbinodus NMS G 1874.6;Strepsodus NEWHM G18.72). Internal relationships
There have been four attempts to elucidate the in-Sensory canals and pits terrelationships of the rhizodontids; Young et al. (1992),
Johanson & Ahlberg (1998), Jeffery (1999) and Johan-Johanson & Ahlberg (in press) observed a large fieldson & Ahlberg (in press). Young et al. (1992) had aof lateralis pores on the cleithrum of Gooloogongia.limited data set, and Johanson & Ahlberg (1998) wereThis prompted a re-examination of the cleithra of theunable to resolve the interrelationships to any degree.European taxa.
Rhizodus (SME 4715a and NMS G 1963.16.18) has Both Jeffery (1999) and Johanson & Ahlberg (in press)

228 J. E. JEFFERY
identified Gooloogongia as the most primitive rhi- of [osteolepiforms+[elpistostegids+tetrapods]]. Thereis not yet a consensus about which of the remainingzodontid. Johanson & Ahlberg (in press) found two
more derived clades; one comprised the remaining sarcopterygian orders is closest to the tetrapod total-group (e.g. Cloutier & Ahlberg, 1996; Zhu & Schultze,Devonian rhizodontids (Notorhizodon and Sau-
ripterus), and the other comprised the Carboniferous 1997).Ahlberg & Johanson (1998) revised relationshipsrhizodontids. They recovered four most parsimonious
trees, which differed only in the internal relationships within the tetrapod stem-group (see Fig. 7). They foundone genus (Kenichthys) to the most primitive memberof the Carboniferous clade. Jeffery (1999) recovered a
single most parsimonious tree, in which the re- of the stem-group. They also found that the osteo-lepiforms s.l. were paraphyletic relative tolationships of the four Carboniferous rhizodontids (Ba-
rameda, Strepsodus, Screbinodus and Rhizodus) were [elpistostegids+tetrapods]. Thus many patterns andcharacters previously seen as osteolepiform syn-fully resolved, but Sauripterus and Notorhizodon were
not included in the analysis. However, the single most apomorphies must now be seen as plesiomorphic forelpistostegids and digited tetrapods (i.e. the crown-parsimonious tree of Jeffery (1999) is congruent with
one of the most parsimonious trees of Johanson & group tetrapods plus the most derived members of thestem-group). Patterns of evolution within the osteo-Ahlberg (in press). Therefore, to examine the evolu-
tionary patterns of the pectoral fin and girdle within lepiforms s.l. may also be seen in the context of atransformational sequence leading to crown-group tet-the group, the tree topologies will be combined, with
Johanson & Ahlberg (in press) being used for the rapods.Devonian taxa and Jeffery (1999) being used for theCarboniferous taxa (see Fig. 7D–H).
Endoskeletal patternThe basal position of Gooloogongia suggests thatrelatively short basal lepidotrichial segments in the The evolution of the fin endoskeleton in sar-pectoral fin (>50% of total lepidotrichial length) are copterygians has been discussed in detail by Ahlbergprimitive for the group. This is supported by the fact (1989) and Forey (1998). Recent sarcopterygians sharethat the lepidotrichia in the pelvic fin of Gooloogongia features of the pectoral fin (or limb) endoskeleton whichhave basal segments which resemble the primitive are probably plesiomorphic for the group. A number ofcondition for crown-group osteichthyians (Johanson endoskeletal mesomeres form a mid-line proximodistal& Ahlberg, in press; see below). The well-developed axis (the so-called ‘metapterygial axis’). The first threedepressed posterior flange on the cleithrum is a derived mesomeres correspond to the humerus, ulna and ul-character of the Carboniferous taxa; a weakly de- nare of tetrapods. In Recent lungfish genera and theveloped flange is present on Notorhizodon (Johanson coelacanth Latimeria the mesomeric axis forms both& Ahlberg, in press), and the flange is entirely absent proximal and distal parts of the endoskeleton. In lung-from Gooloogongia (Johanson & Ahlberg, 1998). fish the axis is long (at least 18 mesomeres in Neo-
The shape of the fin appears to move from the broad, ceratodus; Fig. 7C) and, where known, fossil lungfish‘fan’ of Gooloogongia and Sauripterus (Fig. 1F–H), to also possess long axes (Ahlberg, 1989). The poro-the rhomboid ‘paddle’ of Strepsodus and Screbinodus lepiform Glyptolepis is the only other taxon with a(Fig. 1A–C). This correlates with a reduction in the well-preserved endoskeleton known to possess a longrelative size of the radius, which is much larger in axis (possibly up to 20 mesomeres; Ahlberg, 1989).Sauripterus than in the more derived rhizodontids However, two recent cladistic analyses (Cloutier &(Barameda, Strepsodus and Rhizodus). There is an Ahlberg, 1996; Forey, 1998) placed porolepiformsapparent reduction in the complexity of the branching within the lungfish stem-group, on the basis of a num-of the distal endoskeleton in Carboniferous rhi- ber of morphological features besides the fin. In La-zodontids when compared to Sauripterus. This may be timeria the axis is short (4–5 mesomeres; Fig. 7B).artefactual; the fins are better preserved in Sauripterus Laugia is the only fossil coelacanth to preserve thethan the other members of the group. pectoral fin skeleton in any detail. It has 4–5
mesomeres, and is generally very similar to that ofLatimeria (Forey, 1998). Recent tetrapods show a
Relationship to other sarcopterygians unique morphological pattern in the distal parts of theendoskeleton (the autopodium, including digits). ThisThere is a general consensus about the phylogenetic
position of the rhizodontid clade within the sar- morphology is first seen in stem-tetrapods more de-rived than Panderichthys (e.g. Acanthostega; Coates,copterygians. All recent analyses (e.g. Cloutier &
Ahlberg, 1996; Zhu & Schultze, 1997; Ahlberg & Johan- 1996). It may be a neomorphic structure (e.g. seeNelson et al., 1996; Coates & Ruta, 2000).son, 1998; Forey, 1998; Jeffery, 1999; Zhu, Yu & Jan-
vier, 1999) have found them to be basal members of In Recent sarcopterygians secondary radials ar-ticulate with the mesomeres. In lungfish, the secondarythe tetrapod total-group, specifically as the sister group

PECTORAL FINS OF RHIZODONTIDS 229
Polyodon
Latimeria
Neoceratodus
Kenichthys
Sauripteris
Sauripteris
Gooloogongia
Notorhizodon
Barameda
Actinopterygii
Sarcopterygii
Tetrapodtotal-group
Rhizodontida
Gogonasus
Osteolepis
Other Megalichthyids
Megalichthys
Medoevia
Canowindrids
Gyroptychius
Other Tristichopterids
Eusthenopteron
Panderichthys
TetrapodsK
J
I
H
G
FE
D
C
B
A
Strepsodus
Screbinodus
Rhizodus
Figure 7. Phylogeny of the tetrapod total-group. Rhizodontid clade shown in bold lines. Outgroups represented bythe Recent basal actinopterygian Polyodon, and two Recent sarcopterygians (the coelacanth Latimeria and the lungfishNeoceratodus). Main phylogeny after Ahlberg & Johanson (1998); rhizodontid phylogeny after Jeffery (1999) andJohanson & Ahlberg (in press). Diagrammatic articulation patterns of pectoral fin endoskeleton (A–K) shown whereknown. In fin patterns, grey=uncertain or poorly preserved elements (if missing, the presence of some distal elementsmay be inferred by the articular surfaces on proximal elements). Proximal towards left of page, anterior towards thebottom of the page. A, based on Grande & Bemis (1991); B, based on Forey (1998); C, based on Schultze (1987); upperD based on Daeschler & Shubin, (1998); E, based on Long (1989); F,G, based on Andrews & Westoll (1970b); lower Dand H based on pers. obs.; I,J, based on Andrews & Westoll (1970b); K, based on Worobyeva (1975).
radials are segmented and unbranching. Those ar- mesomeres. It should be noted here that during on-togeny the pectoral fin skeleton of lungfishes rotatesticulating anteriorly show a 1:1 correspondence with
the axial mesomeres; those articulating posteriorly to bring the anterior margin into a dorsal position.The adult morphology of Latimeria suggests that itsshow no clear relationship to the axial mesomeres. In
Latimeria the anterior secondary radials are much fin is also rotated, although there are no ontogeneticdata available to confirm this (see Rosen et al., 1981;reduced, and only the proximal two (radius and in-
termedium) show a clear correspondence with the axial Ahlberg, 1989; Forey, 1998 for a discussion). For sim-plicity, the terms ‘anterior’, ‘posterior’, etc., will bemesomeres (Forey, 1998). The distal secondary radials
(anterior and posterior) form a mosaic surrounding applied as if the fins had not rotated. In digited tet-rapods only the first and second anterior radials (thethe tip of the fin, with no clear relationship to the

230 J. E. JEFFERY
radius and intermedium respectively) can be identified existence of a metapterygial axis within the auto-podium, which has yet to be unambiguously dem-with any certainty (see below).onstrated (e.g. see Coates, 1995; Nelson et al., 1996;Outgroup comparisons can also provide informationCoates & Ruta, 2000).on the primitive conditions for sarcopterygians. It is
In rhizodontids the branching is so complex that noclear that a fin endoskeleton formed solely from aclear mesomeric ‘axis’ is discernible distally. There aremetapterygial axis and its associated secondary radialsno posterior secondary radials proximally (articulatingis a derived feature of the sarcopterygians, as arewith the humerus, ulna or ulnare) but it is impossible tosecondary radials articulating with the posterior mar-say whether there are any posterior secondary radialsgin of the metapterygium. Primitively in gnathostomesdistally. At the very least, the endoskeleton (meso-the metapterygium is the posterior-most of a numbermeres and/or secondary radials) are segmented andof radials articulating with the pectoral girdle and hascontinue to branch distally. In view of the retainedsecondary radials articulating only its anterior marginprimitive morphology of the more derived stem-tetra-(Fig. 7A; illustrated by the actinopterygian Polyodon).pods, the complex distal endoskeleton of rhizodontidsFor reviews of primitive fin patterns see Rosen et al.is probably a synapomorphy of that group. It may(1981), Ahlberg (1989), Coates (1994), Forey (1998)even be a neomorphic structure, convergent with theand references therein. Outgroup comparison does notrepatterning of the distal endoskeleton seen in tetra-provide unambiguous information on the primitivepods (cf. Daeschler & Shubin, 1998). In any case,length of the axis in sarcopterygians. In chon-rhizodontid fins appear to retain the primitive ‘short’drichthyians, the metapterygium primitively has anaxis; in the best preserved fin (Sauripterus) there areelongated proximal segment with a variable numberno more than 5–7 segments proximodistally, much less(1–13) of small segments distally (Rosen et al., 1981;than in lungfish.Forey, 1998). In actinopterygians (widely accepted as
the sister group of the sarcopterygians), the me-tapterygium is primitively unsegmented, or at least Entepicondylebears only a very few distal segments (Ahlberg, 1989;
Where data are available, the humerus of coelacanths,Gardiner & Schaeffer, 1989; Coates, 1994; Forey, 1998).lungfish and basal tetrapods all bear a ventroposteriorHowever, Ahlberg (1989: 149) noted that, althoughprocess, the entepicondyle. This may be a syn-unsegmented, the primitive actinopterygian meta-apomorphy of the sarcopterygians (see Ahlberg, 1989).pterygium is a short structure bearing only a few
The entepicondyle of Eusthenopteron is hook-shapedsecondary radials. He therefore suggested that a shortand projects more-or-less in line with the ulnar and
metapterygium is primitive for sarcopterygians, whichradial condyles. The base of the entepicondyle is
in turn implied that the long axis of lungfish (includingpierced by a canal running proximodistally, resulting
porolepiforms) is a derived feature of that group. in proximal and distal entepicondylar foramina. ThisUnfortunately, no data are available for the fin also appears to be the case with the osteolepiforms
pattern of the most basal stem-tetrapod, Kenichthys. s.l. Medoevia (Lebedev, 1995) and Beelarongia (Long,However, the osteolepiforms s.l. Sterropterygion, Eu- 1987) and in Panderichthys (Vorobyeva, 2000).sthenopteron and Megalichthys have a short axis of no The large subrectangular entepicondyle of earlymore than five mesomeres (Rackoff, 1980; Andrews & tetrapods also projects more-or-less in line with theWestoll, 1970b; Fig. 7I,J), suggesting that the primitive ulnar and radial condyles, but has broad dorsal andshort axis is retained in stem-tetrapods. An even more ventral ‘faces’ (Coates, 1996). The canal connecting theabbreviated endoskeleton is seen in Panderichthys, entepicondylar foramina runs proximodistally, roughlywith only three mesomeres, including an expanded in the plane of the faces.‘ulnare plate’ (Worobyeva, 1975; Fig. 7K). These taxa In Rhizodus (Fig. 3) and Strepsodus (Fig. 4E–H) thehave no secondary radials on the posterior margin of entepicondyle is very broad dorsoventrally. The dorsalthe axis, which may also be a retention of the primitive margin projects in line with the ulnar and radialsarcopterygian condition. Some of the axial mesomeres condyles, and the ventral margin is confluent with theof Eusthenopteron bear posterior processes, which Wes- oblique ventral ridge on the body of the humerus. Thetoll (1943) and Jarvik (1980) proposed were ho- broad ‘faces’ are anterodistal and posteroproximal. Themologous with posterior secondary radials. This has canal connecting the entepicondylar foramina runsbeen rejected by subsequent authors (e.g. Ahlberg proximodistally through the base of the entepicondyle,1989; Forey, 1998) as they are never seen as separate and therefore runs from the anterodistal to the post-ossifications. The digits of tetrapods have also been eroproximal face. This is quite unlike the condition inequated with secondary radials (see Shubin & Alberch, tetrapods, and may be a synapomorphy of the rhi-1986; Ahlberg, 1989; Forey, 1998 for reviews). However, zodontids. However, the superficial similarity with the
broad entepicondyle of tetrapods seems to have causedthese arguments are based on assumptions of the

PECTORAL FINS OF RHIZODONTIDS 231
Vorobyeva (2000: Fig. 2) to interpret the orientation of gradually decreases distally (e.g. Grandel & Merker,the humerus of Strepsodus NMS G 1970.16.1 in- 1998). The hemisegments have a ‘U’-shaped cross-correctly (see Fig. 4E–H). section (Fig. 8E,F), with the dorsal and ventral hemi-
segments aligning to enclose a central space containingconnective tissue, nerves and blood vessels (LaforestSupinator and deltoid processeset al., 1998). At the distal tip of each lepidotrichium
The humeri of basal crown-group tetrapods bear three the remains of an actinotrichium (‘actin’) protrudes,processes on the dorsal surface; the deltoid and su- providing a short, flexible end to the fin ray. In two orpinator processes and the ectepicondyle (e.g. Coates, three places along the lepidotrichia the hemisegments1996). The homologues of these processes have been will bifurcate (lepid. branch; Fig. 8C). The points ofidentified in the stem-tetrapods Acanthostega (Coates, bifurcation are co-ordinated across the fin, and are1996), Panderichthys (Vorobyeva, 2000), Eus- similar in every lepidotrichium (Laforest et al., 1998).thenopteron (Andrews & Westoll, 1970a) and Medoevia The most basal hemisegments of each ray are usually(Lebedev, 1995). In Acanthostega and Panderichthys slightly longer than the other segments, and are roundthe processes appear to be distinct structures, as they in cross-section (e.g. Goodrich, 1904; Jarvik, 1959) (Fig.are in crown-group tetrapods. However, in Eus- 8D). These segments overlap (and may be fused with)thenopteron and Medoevia they appear consist of swell- the fin endoskeleton (basal lepid. hemiseg.; Fig. 8C,ings on a more-or-less continuous dorsal ridge. G). The degree of overlap usually encompasses only theRhizodus and Strepsodus (and possibly Barameda and most distal parts of the endoskeleton (e.g. LatimeriaNotorhizodon) also show a single ridge. Rhizodus has (Forey, 1998), Polypterus (pers. obs. of specimensome limited subdivision (via the ‘ectepicondylar fo- UCLGMZ V416), Lepisosteus (Goodrich, 1958), Amiaramen’), and Strepsodus may possess and distinct ec- (Grande & Bemis, 1998), the zebrafish Brachydaniotepicondyle. (Grandel & Merker, 1998); see Fig. 8G). The slightly
Ahlberg (1989) suggested that the three processes greater overlap seen in Recent acipenseriforms, whereof tetrapods were homologous with the single dorsal the lepidotrichia can entirely cover some distal radialsprocess seen on the humerus of Latimeria, Glyptolepis (e.g. Acipenser (Goodrich, 1904), Polyodon (Grande &and lungfish. He was unable to determine whether Bemis, 1991) and Psephurus (Grande & Bemis, 1991)either condition was primitive with respect to the other appears to be a synapomorphy of that group.(i.e. whether the three processes were derived from a The pattern seen in rhizodontids (highly elongatedsingle process by subdivision and/or neomorphy, or basal hemisegments with extensive overlap with thewhether the single process was derived by loss and/or endoskeleton; Fig. 8H) is probably a synapomorphy offusion of the three processes). However, the separation that group, and not the primitive condition for stem-into three distinct structures appears to be restricted group tetrapods. More derived stem-tetrapod taxa re-to a clade including Panderichthys and more derived
tain the plesiomorphic crown-group osteichthyian mor-stem-tetrapods plus the crown-group. More basal stem-
phology (except digited tetrapods, where there is notetrapods show an ‘intermediate’ morphology, with the
trace of dermal fin rays).partial subdivision of a single dorsal ridge. It would
Amongst the Recent genera of non-tetrapod sar-therefore be most parsimonious to assume that the
copterygians, the coelacanth Latimeria, as describedsingle ridge is the primitive condition for sar-
above, retains the primitive fin ray morphology, al-copterygians, with a gradual specialization occurringthough there is some superficial secondary ossificationwithin the tetrapod stem-lineage.across the proximal joints (Castanet et al., 1975). Thefin rays of Recent lungfish (Fig. 8A) show several
Dermal fin rays convergent features with those of rhizodontids. Each‘camptotrichium’ (campt.; the equivalent of a hemiray)Amongst Recent sarcopterygians, only the coelacanthhas a round cross-section and is not aligned with itsLatimeria bears fully ossified dermal fin rays (Castanetantimere. They are branched (campt. branch.), butet al., 1975). However, the morphology of these fin raysthere are only slight traces of segmentation. In theis similar to that seen in actinopterygians (e.g. theDevonian fossil lungfish Scaumenacia (Fig. 8B) thezebrafish Brachydanio; Grandel & Merker, 1998). It israys are segmented, with the basal hemisegmentsalso similar to what is known of the fin rays of stem-comprising about half the total length of the ray (‘basalcoelacanths, and the extinct porolepiforms and osteo-campt. seg.’), similar to the lepidotrichia of Gooloogon-lepiforms s.l. (e.g. see Goodrich, 1904 and Jarvik, 1959).gia. The fin rays of Recent lungfish also have someIt is thus likely that this morphology is primitive forunique characters, such as a division of each campto-crown-group osteichthyians.trichium into distinct superficial and deep regions (ofEach lepidotrichium consists of dorsal and ventralbone and collagen respectively (Geraudie & Meunier,hemirays (Kemp & Park, 1970). Each hemiray consists
of a number of hemisegments, the length of which 1984)). In Scaumenacia the rays are more fully ossified,

232 J. E. JEFFERY
endoskel. basal lepid. hemiseg. actin.distal lepid. hemiseg.
basal lepid. hemiseg.lepid. branch.
distal campt. hemiseg.distal lepid. hemiseg.
campt.
basal campt. seg.
A
B
C
DE F
G
H
campt. branch.
Figure 8. Proximodistal morphology of fin rays. A, unsegmented camptotrichia of Neoceratodus. B, segmentedcamptotrichia of Scaumenacia (this example from the dorsal fin). Note elongated basal segments. C, lepidotrichia ofEusthenopteron, showing the primitive sarcopterygian morphology. D–F, cross-sections of lepidotrichia in Eu-sthenopteron (D, paired dorsal and ventral hemisegments showing basal morphology (round in cross-section); E, moredistal morphology – three paired dorsal and ventral hemisegments showing beginnings of a ‘U’-shaped cross-section;F, distal morphology – four paired dorsal and ventral hemisegments showing complete ‘U’-shaped cross-section). G,diagrammatic lateral longitudinal cross-section of a primitive sarcopterygian lepidotrichium; dorsal and ventralhemirays divided into paired hemisegments. H, Diagrammatic lateral longitudinal cross-section of a lepidotrichium ofStrepsodus; the basal hemisegments of the lepidotrichium are highly elongated, overlapping several endoskeletalelements. The basal morphology (unbranched and round in cross section) is retained. A,B and D–F after Goodrich(1904); C, after Jarvik (1959).
similar to primitive osteichthyian pattern. Thus the or even how it is achieved in vivo; whether by thepositive allometric growth of the endoskeleton relativeloss of segmentation seen in later lungfish is convergent
with rhizodontids, and may reflect some similar to the lepidotrichia, or by proximad growth of thelepidotrichia. However, the co-incidence of the ex-changes in the developmental patterning of the lep-
idotrichia. tensive endoskeletal overlap and elongation of thebasal segment of rhizodontid lepidotrichia raises aThe developmental patterning of lepidotrichia has
only been studied in the actinopterygian zebrafish possibility that they may share some common up-stream genetic controls.(Laforest et al., 1998), although the mechanisms de-
scribed may be primitive for crown-group os-teichthyians. The overlying ectoderm interacts with
DEVELOPMENTAL MODELSscleroblasts laying down the lepidotrichia to co-or-dinate branching and segmentation (Laforest et al., The paired appendage bud is a ‘model system’ in both
evolutionary and developmental biology (e.g. Coates,1998). However, little is known of the up-stream mech-anisms inducing the proximodistal pattern. Also, there 1996; Johnson & Tabin, 1997). Only a few non-tetrapod
species have been studied in any detail, all of themare no published data to show how the limit of overlapof the endoskeleton with the dermal rays is patterned, teleost actinopterygians (e.g. the trout Oncorhynchus

PECTORAL FINS OF RHIZODONTIDS 233
In non-tetrapods the AER enlarges and develops aninternal cavity, becoming a ‘fin-fold’ (Fig. 9B). Althoughinitially excluded, a distal population of finbud me-senchyme (mes. f.f.) soon enters the fold, producingelastoidin fin rays (actinotrichia) (Grandel & Merker,1998). Later, scleroblasts move between the ectodermand actinotrichia and start to lay down the bony finrays (lepidotrichia) (Laforest et al., 1998). In tetrapods,the transformation of AER to fin-fold fails to occur,and progress zone proliferation becomes asymmetrical,with greater mitosis posteriorly (Haack & Gruss, 1993;Vargesson et al., 1997). This is coincident with a modi-fication of the expression patterns of hoxa and hoxd,by a combination of cell movements and changes incell expression (Nelson et al., 1996; Vargesson et al.,1997). There are also changes in the expression of
BA
C
AER
PZ
fin-fold
mes. f.f.
tetrapod
lungfish
teleost
AER transformsto fin fold
Late
Early
members of the FGF family and of the growth factorFigure 9. Pectoral fin development. A,B, diagrammatic sonic hedgehog (shh). This alteration in gene ex-representations of the development of the zebrafish pec- pression domains represents the initial stages of thetoral fin. A, early bud. The AER maintains rapid cell patterning and formation of carpals and digits. Tho-proliferation in the PZ (triangular arrows) During this rogood (1991) proposed a model of paired appendagephase the endoskeletal anlage are laid down. B, later bud.
evolution based on a shift in timing of the trans-The AER has transformed into the fin-fold. Mesenchymeformation of the AER to fin-fold (the ‘clockface’ model;has invaded the fold and started to lay down the fin rays.Fig. 9C). Actinopterygians typically possess a smallProliferation in the PZ has almost ceased. C, ‘clockface’number of endoskeletal radials supporting lengthymodel proposed by Thorogood (1991). Teleosts transformdermal lepidotrichia. The endoskeleton is not heavilytheir AER early, and therefore have a small endoskeletalbranched, although the most posterior radial oftencontribution to the fin. Tetrapods do not transform theirsupports a small number of anterior radials. RecentAER and thus have no fin rays. Lungfish have a more
robust endoskeleton and less significant fin ray skeleton sarcopterygian fishes (lungfish and the coelacanth)than do teleosts. This may be due to a later transformation possess highly segmented endoskeleton, and a rel-of the AER to a fin-fold. A,B after Grandel & Merker atively smaller lepidotrichial skeleton. Studies of ac-(1998) and Sordino et al. (1995). C, after Sordino et al. tinopterygian fin development have shown that when(1995). the AER transforms into the fin-fold, proliferation in
the PZ is drastically reduced. It has been suggestedthat this is caused by a cessation or mechanical in-
(Geraudie & Francois, 1973), the killifish Aphyosemionterruption of FGF signalling (Grandel & Merker, 1998;
(Bouvet, 1974; Geraudie, 1978; Geraudie & Landis,Sordino & Duboule, 1996; Sordino, van der Hoeven &1982; Wood, 1982) and the zebrafish (Grandel &Duboule, 1995). Thorogood (1991) proposed that theMerker, 1998; Kimmel et al., 1995 and referencesadult fin morphology was a consequence of the timingtherein)). Much attention has been given to a fun-of the transformation of the AER to the fold (Fig. 9C).damental difference between the development of finIn actinopterygians the greater contribution of theand limb buds. In both there is a thickened ridge oflepidotrichial skeleton is a reflection of an early trans-ectoderm along the distal tip of the bud, the apicalformation of the AER to a fin-fold. In contrast, theectodermal ridge (‘AER’; Fig. 9A). This produces se-greater contribution of the endoskeleton in sar-creted factors (members of the fibroblast growth factorcopterygians reflects a later transition of the AER.(FGF) family) which keeps a subjacent population ofThe complete dominance of the elaborately patternedrapidly dividing mesenchyme, the progress zone (PZ)endoskeleton in tetrapods reflects the failure of thein an undifferentiated state. The PZ is the major regionAER to transform at all. Although not explicitly stated,of growth in the bud, and is involved in thethis model implies that endoskeletal and lepidotrichialproximodistal patterning of the appendage throughsystems are each given a portion of a fixed ‘time budget’members of the hoxa and hoxd gene families. Theof overall bud development.AER is also involved (via a positive feedback loop of
The morphology of rhizodontid fins does not fit withmaintaining signals) with the mechanisms for es-this model, as there is an equally robust contributiontablishing the anteroposterior and dorsoventral axesfrom both endoskeletal and lepidotrichial elements.of the appendage (see Johnson & Tabin (1997) for a
recent review). Furthermore, the endoskeleton displays an extremely

234 J. E. JEFFERY
complex pattern of segmentation. Thus it is not neces- Sauripterus. Sarah Finney guided my fossil pre-paration, and prepared NEWHM G19.41. I am gratefulsary per se to suppress the transformation of the AER
(thus reducing or losing the lepidotrichia) in order to to the Royal Society of Edinburgh for permission toreproduce Andrews & Westoll (1970b: plate IXe–g),evolve an increased complexity of the endoskeleton.
Rather, the loss of the fin-fold may relate to the modi- to the Company of Biologists Ltd., for permission toreproduce Goodrich (1904: figs 4, 36 and 45) and tofication of gene expression, and asymmetric growth in
the progress zone, both of which are unique to tetrapods the Royal Swedish Academy of Sciences for permissionto reproduce Jarvik (1959: fig. 10B).(Nelson et al., 1996; Sordino et al., 1995). This would
be a contingency of tetrapod evolution, rather than anecessary step in the evolution of a robust and complex
REFERENCESendoskeleton.Rhizodontid fin morphology suggests two alternative Ahlberg PE. 1989. Paired fin skeletons and relationships of
models for the developmental relationship between the the fossil group Porolepiformes (Osteichthyes: Sar-endoskeleton and the lepidotrichia: copterygii). Zoological Journal of the Linnean Society 96:
119–166.(1) Rhizodontids differed from actinopterygians in that
Ahlberg PE. 1991. A re-examination of sarcopterygian in-AER signalling to the PZ did not cease with its terrelationships, with special reference to the Porolepi-transformation to a fin-fold, and endoskeletal pat- formes. Zoological Journal of the Linnean Society 103:terning could therefore continue in unison with 241–287.lepidotrichial growth. Ahlberg PE, Johanson Z. 1998. Osteolepiforms and the
(2) Rhizodontids were similar to actinopterygians, and ancestry of tetrapods. Nature (London) 395: 792–794.had to form a fin-fold early in order to develop the Andrews SM. 1972. The shoulder girdle of ‘Eogyrinus’. In:large lepidotrichia. However, this must have been Joysey KA, Kemp TS, eds. Studies in Vertebrate Evolution.preceded by an extremely rapid patterning of very Edinburgh: Oliver & Boyd, 35–48.small endoskeletal primordia. Later positive al- Andrews SM. 1973. Interrelationships of crossopterygians.lometric growth could result in the robust and In: Greenwood PH, Miles RS, Patterson C, eds. Inter-
relationships of Fishes. London: Academic Press, 138–177.complex endoskeleton.Andrews SM. 1985. Rhizodont crossopterygian fish from theAlso, a third possibility exists:
Dinatian of Foulden, Berwickshire, Scotland, with a re-(3) The ‘time budget’ is more flexible than supposed,evaluation of this group. Transactions of the Royal Societyallowing extended development of both endo-of Edinburgh: Earth Sciences 76: 67–95.skeleton and lepidotrichia in turn.
Andrews SM, Westoll TS. 1970a. The postcranial skeletonThis third model is not mutually exclusive with the of Eusthenopteron foodi Whiteaves. Transactions of the
Royal Society of Edinburgh 68: 207–329.alternatives (1) and (2); it is possible that some degreeAndrews SM, Westoll TS. 1970b. The postcranial skeletonof manipulation of the ‘time budget’ could occur in
of rhipidistian fishes excluding Eusthenopteron. Trans-concert with either alternative. However, if the ‘timeactions of the Royal Society of Edinburgh 68: 391–489.budget’ is flexible, the abbreviated patterning and
Bouvet J. 1974. Differenciation et ultrastructure du sque-small size of the endoskeleton of teleost ac-lette de la nageoire pectorale chez la truite indigene (Salmotinopterygians [e.g. the zebrafish (Sordino & Duboule,trutta fario L.) I. Differenciation et ultrastructure des1996; Sordino et al., 1995)] might be due to activeactinotriches. Archives d’Anatomie Microscopique 63: 79–selection on that morphology, rather than being con-96.strained by the demands of lepidotrichial development.
Boyd MJ. 1984. The Upper Carboniferous tetrapod as-semblage from Newsham, Northumberland. Palaeontology27: 367–392.
ACKNOWLEDGEMENTS Broom R. 1913. On the origin of the Cheiropterygium.Bulletin of the American Museum of Natural History 22:Much of this work was carried out as part of my Ph.D.,459–464.
funded by BBSRC grant 9430 6007; I am indebted toCastanet J, Meunier F, Bergot C, Francois Y. 1975.
my supervisor, Dr Jenny Clack, for her help and advice Donnees preliminaires sur les structures histologiques duduring my research. I would also like to thank Dr Mike squelette de Latimeria chalumnae. 1. Dent, ecailles, rayonsCoates and Dr Mike Richardson for discussion and de nageoires. In: Colloque international du Centre nationalcomments during the preparation of the manuscript. de la Recherche Scientifique, 218. Problemes actuels deDr P. E. Ahlberg made perceptive comments on the final paleontologie: evolution des vertebres (Paris, 4–9 juin 1973).manuscript and, along with Dr Z. Johanson shared Paris: Editions du C.N.R.S., 159–168.valuable information about Gooloogongia and No- Chisholm JI, Castanet J. 1994. Revision of the late Din-torhizodon. Dr E. B. Daeschler, Mr M. C. Davis and antian sequence in Edinburgh and West Lothian. Scottish
Journal of Geology 30: 97–104.Professor N. Shubin were similarly forthcoming about

PECTORAL FINS OF RHIZODONTIDS 235
Cloutier R, Ahlberg PE. 1996. Morphology, characters, and origin of the Tetrapoda, with special reference to the skulland paired limbs. Annals of the New York Academy ofthe interrelationships of basal sarcopterygians. In: Stiassny
MLJ, Parenti LR, Johnson GD, eds. Interrelationships of Science 26: 317–383.Gregory WK. 1935. Further observations on the pectoralFishes. London: Academic Press, 445–479.
Coates MI. 1994. The origin of vertebrate limbs. Development girdle and fin of Sauripterus taylori Hall, a crossopterygianfish from the Upper Devonian of Pennsylvania, with specialSupplement: 169–180.
Coates MI. 1995. Fish fins or tetrapod limbs – a simple reference to the origin of the pentadactylate extremitiesof Tetrapoda. Proceedings of the American Philosophicaltwist of fate? Current Biology 5: 844–848.
Coates MI. 1996. The Devonian tetrapod Acanthostega Society 75: 673–690.Gregory WK, Miner RW, Noble GK. 1923. The carpus ofgunnari Jarvik: postcranial anatomy, basal tetrapod inter-
relationships and patterns of skeletal evolution. Trans- Eryops and the structure of the primitive chiropterygium.Bulletin of the American Museum of Natural History 48:actions of the Royal Society of Edinburgh: Earth Sciences
87: 363–421. 279–288.Haack H, Gruss P. 1993. The establishment of murineCoates MI, Ruta M. 2000. Early tetrapod evolution. Trends
in Ecology and Evolution 15: 327–328. Hox-1 expression domains during patterning of the limb.Developmental Biology 157: 410–422.Daeschler EB, Shubin N. 1998. Fish with fingers? Nature
(London) 391: 133. Hall J. 1843. Natural History of New York; Geology of New-York, Part IV. comprising the Survey of the Fourth Geo-Davis MC, Daeschler EB, Shubin N (in press). Immature
rhizodontids from the Devonian of North America. Bulletin logical District. Albany, USA: Carroll & Cook.Huxley TH, Etheridge R. 1865. A Catalogue of the Col-of the Museum of Comparative Zoology, Harvard.
Eastman CR. 1917. Fossil fishes in the collections of the lection of Fossils in the Museum of Practical Geology, withan explanatory introduction. London: G.E. Eyre & W. Spot-United States National Museum. Proceedings of the United
States National Museum 52: 235–304. tiswoode.Janvier P, Lethier F, Monod O, Balkas O. 1984. DiscoveryForey PL. 1998. History of the coelacanth fishes. London:
NHM/Chapman & Hall. of a vertebrate fauna at the Devonian–Carboniferousboundary in SE Turkey (Hakkari Province). Journal ofGardiner BG, Schaeffer B. 1989. Interrelationships of
lower actinopterygian fishes. Zoological Journal of the Lin- Petroleum Geology 7: 147–168.Jarvik E. 1959. Dermal fin-rays and Holmgren’s principlenean Society 97: 135–187.
Geraudie J. 1978. Scanning electron microscope study of of delamination. Kungliga Svenska VetenskapsakademiensHandlinger 6 (4th Series): 3–51.the developing trout pelvic fin bud. Anatomical Records
191: 391–396. Jarvik E. 1980. Basic Structure and Evolution of Vertebrates.London: Academic Press.Geraudie J, Francois Y. 1973. Les premiers stades de la
formation de l’ebauche de nageoire pelvienne de Truite Jeffery JE. 1999. The morphology and phylogeny of theEuropean members of Order Rhizodontida (Pisces: Sar-(Salmo fario et Salmo gairdneri ). I. Etude anatomique.
Journal of Embryology and Experimental Morphology 29: copterygii). Unpublished Ph.D. Thesis, University of Cam-bridge Museum of Zoology.221–237.
Geraudie J, Landis WJ. 1982. The fine structure of the Johanson Z, Ahlberg PE. 1998. A complete primitive rhi-zodont from Australia. Nature (London) 394: 569–573.developing pelvic fin dermal skeleton in the trout Salmo
gairdneri. American Journal of Anatomy 163: 141–156. Johanson Z, Ahlberg PE (in press). Devonian rhizodontids(Sarcopterygii; Tetrapodomorpha) from East Gondwana.Geraudie J, Meunier FJ. 1984. Structure and comparative
morphology of camptotrichia of lungfish fins. Tissue and Transactions of the Royal Society of Edinburgh: EarthScience.Cell 16: 217–236.
Goodrich ES. 1904. On the dermal fin-rays of fishes – living Johnson RL, Tabin CJ. 1997. Molecular models for ver-tebrate limb development. Cell 90: 979–990.and extinct. Quarterly Journal of Microscopical Science 47
(New Series): 466–522. Kemp NE, Park JH. 1970. Regeneration of lepidotrichiaand actinotrichia in the tailfin of the teleost Tilapia mos-Goodrich ES. 1958. Studies on the Structure and De-
velopment of Vertebrates. New York: Dover. sambica. Developmental Biology 22: 321–342.Kimmel CB, Ballard WW, Kimmel SR, Ullmann B, Schil-Grandel H, Merker SS. 1998. The development of the
paired fins in the Zebrafish (Danio rerio). Mechanisms of ling TF. 1995. Stages of embryonic development of thezebrafish. Developmental Dynamics 203: 253–310.Development 79: 99–120.
Grande L, Bemis WE. 1991. Osteology and phylogenetic Laforest L, Brown CW, Poleo G, Geraudie J, Tada M,Ekker M, Akimenko MA. 1998. Involvement of the Sonicrelationships of fossil and Recent paddlefishes (Poly-
odontidae) with comments on the interrelationships of Hedgehog, patched 1 and bmp2 genes in patterning ofzebrafish dermal rays. Development 125: 4175–4184.Acipenseriformes. Journal of Vertebrate Paleontology 11
(Suppl. 1): 1–121. Lebedev O. 1995. Morphology of a new osteolepid fish fromRussia. Bulletin du Musee National d’Histoire Naturelle,Gregory WK. 1911. The limbs of Eryops and the origin of
paired limbs from fins. Science (New Series) 33: 508–509. Paris, 4e Series 17: 287–341.Long JA. 1987. An unusual osteolepiform fish from theGregory WK. 1915. Present status of the problem of the

236 J. E. JEFFERY
Late Devonian of Victoria, Australia. Palaeontology 30: evolution. In: Hinchliffe JR, Hurle JM, Summerbell D, eds.839–852. Developmental patterning of the vertebrate limb. New York:
Long JA. 1989. A new rhizodontiform fish from the Early Plenum Press, 347–354.Carboniferous of Victoria, Australia, with remarks on the Traquair RH. 1875. On some Fossil Fishes from the Neigh-phylogenetic position of the group. Journal of Vertebrate bourhood of Edinburgh. Annals and Magazine of NaturalPaleontology 9: 1–17. History 15: 258–268.
Nelson CE, Morgan BA, Burke AC, Laufer E, DiMambro Vargesson N, Clarke JDW, Vincent K, Coles C, WolpertE, Murtaugh LC, Gonzales E, Tessarollo L, Parada L, Tickle C. 1997. Cell fate in the chick limb bud andLF, Tabin C. 1996. Analysis of Hox gene expression in the relationship to gene expression. Development 124: 1909–chick limb bud. Development 122: 1449–1466. 1918.
Owen R. 1840. Odontography, or, a treatise on the com- Vorobyeva EI. 2000. Morphology of the humerus in theparative anatomy of the teeth; their physiological relations, rhipidistian Crossopterygii and the origin of tetrapods.mode of development, and microscopic structure in the Paleontological Journal 34: 632–641.vertebrate animals. London: Hippolyte Bailliere. Westoll TS. 1943. The origin of the primitive tetrapod
Patterson C. 1993. Naming names. Nature (London) 366: limb. Proceedings of the Royal Society London 131B:518. 373–393.
Rackoff JS. 1980. The origin of the tetrapod limb and the Whyte MA. 1994. Scottish fresh-water limestones in theirancestry of tetrapods. In: Panchen AL, ed. The Terrestrial regional setting. Transactions of the Royal Society of Ed-Environment and the Origin of Land Vertebrates. London:
inburgh: Earth Sciences 84: 239–248.Academic Press, 255–292.
Wood A. 1982. Early pectoral fin development and mor-Romer AS, Byrne F. 1931. The pes of Diadectes: Notes on
phogenesis of the apical ectodermal ridge in the killifish,the primitive tetrapod limb. Palaeobiologica 4: 24–48.
Aphyosemion scheeli. Anatomical Records 204: 349–356.Rosen DE, Forey PL, Gardiner BG, Patterson C. 1981.
Wood SP. 1975. Recent discoveries of Carboniferous fishesLungfishes, tetrapods, palaeontology, and plesiomorphy.in Edinburgh. Scottish Journal of Geology 11: 251–258.Bulletin of the American Museum of Natural History 167:
Wood SP. 1992. A Challenging Edinburgh Fossil Site. Ed-163–275.inburgh: Mr Wood’s Fossils.Schultze HP. 1987. Dipnoans as sarcopterygians. Journal
Worobyeva [Vorobyeva] EI. 1975. Bemerkungen zu Pan-of Morphology (Supplement) 1: 39–74.derichthys rhombolepis (Gross) aus Lode in LettlandShubin NH, Alberch P. 1986. A morphogenetic approach(Gauja-Schichten, Oberdevon). Neues Jahrbuch fur Ge-to the origin and basic organisation of the tetrapod limb.ologie und Palaontologie, Monatshafte 5: 315–320.In: Hecht MK, Wallace B, Prance GI, eds. Evolutionary
Young GC, Long JA, Ritchie A. 1992. CrossopterygianBiology. New York: Plenum Press, 319–387.fishes from the Devonian of Antarctica: Systematics, re-Sordino P, Duboule D. 1996. A molecular approach tolationships and biogeographic significance. Records of thethe evolution of vertebrate paired appendages. Tree 11:Australian Museum, Supplement 14: 1–77.114–119.
Zhu M, Schultze HP. 1997. The oldest sarcopterygian fish.Sordino P, van der Hoeven F, Duboule D. 1995. Hox geneLethaia 30: 293–304.expression in teleost fins and the origin of vertebrate digits.
Zhu M, Yu XB, Janvier P. 1999. A primitive fossil fishNature (London) 375: 678–681.sheds light on the origin of bony fishes. Nature (London)Thorogood P. 1991. The development of the teleost fin
and implications for our understanding of tetrapod limb 397: 607–610.


















